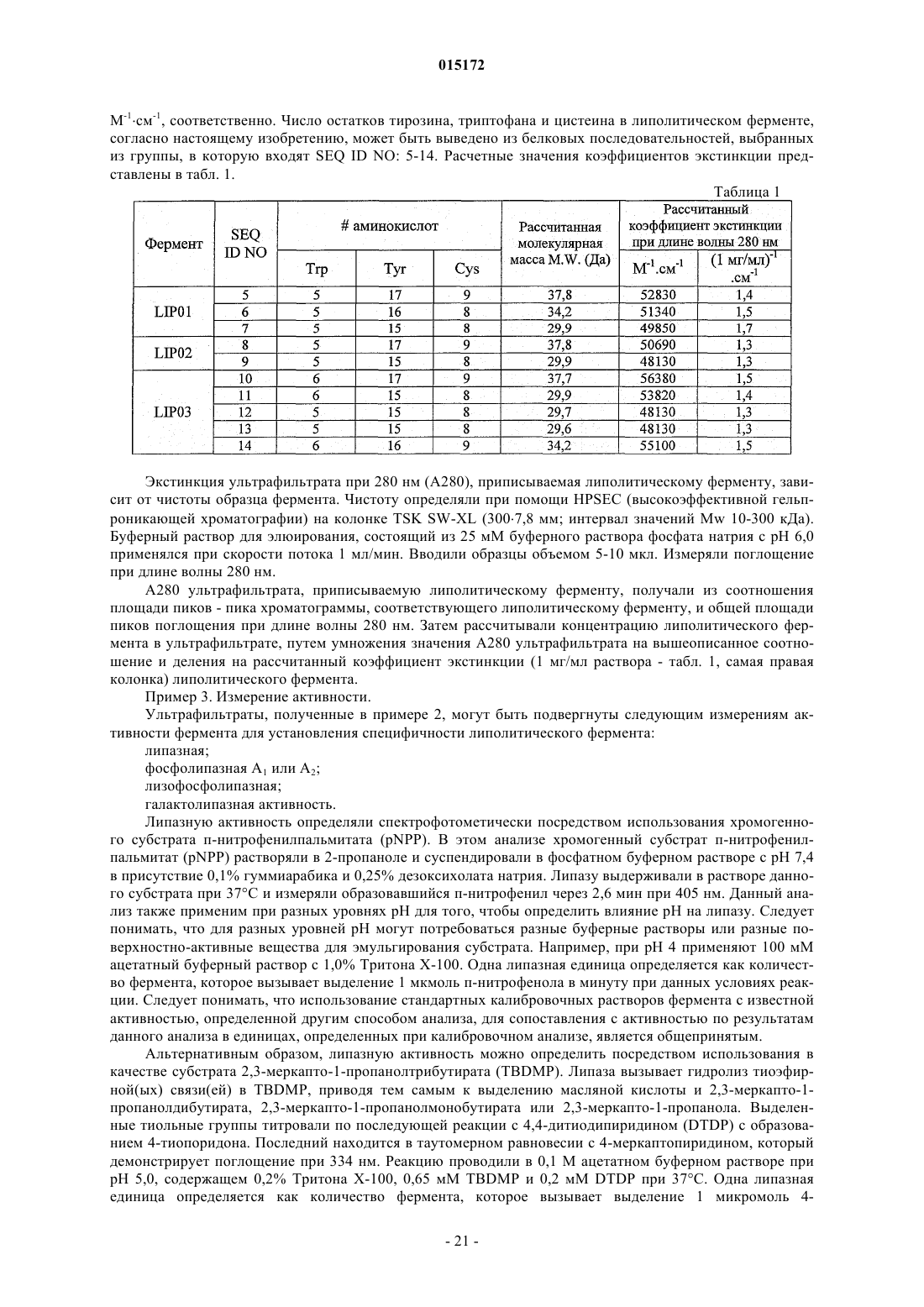
Новые липазы и их применение
Номер патента: 15172
Опубликовано: 30.06.2011
Авторы: Шонефельд-Бергманс Маргот Элизабет Франсуаз, Лаан Ван Дер Ян Метске


























Формула / Реферат
1. Выделенный полинуклеотид, кодирующий липолитический фермент, выбранный из группы, состоящей из:
a) выделенного полинуклеотида, кодирующего полипептид, содержащий последовательность аминокислот, соответствующую любой из SEQ ID NO: 5-7 или функциональному эквиваленту любой из них, по меньшей мере на 70% гомологичному аминокислотной последовательности SEQ ID NO: 5-7;
b) выделенного полинуклеотида, содержащего последовательность нуклеотидов, соответствующую любой из SEQ ID NO: 2-4 или их функциональным эквивалентам, по меньшей мере на 85% гомологичным нуклеотидной последовательности SEQ ID NO: 2;
c) выделенного полинуклеотида, кодирующего полипептид, содержащий последовательность аминокислот, соответствующую любой из SEQ ID NO: 8-9 или SEQ ID NO:10-14;
d) выделенного полинуклеотида, соответствующего любой из SEQ ID NO: 3-4.
2. Вектор, содержащий последовательность полинуклеотидов по п.1.
3. Вектор по п.2, где упомянутая последовательность полинуклеотидов по п.1 функционально связана с регуляторными последовательностями, пригодными для экспрессии указанной последовательности полинуклеотидов в подходящей клетке-хозяине.
4. Вектор по п.3, где указанная подходящая клетка-хозяин представляет собой мицелиальный гриб, предпочтительно вида Aspergillus, более предпочтительно Aspergillius niger.
5. Способ получения полинуклеотида по п.1 или вектора по пп.2-4, включающий этапы культивирования клетки-хозяина, трансформированной указанным полинуклеотидом или указанным вектором, и выделения указанного полинуклеотида или указанного вектора из указанной клетки-хозяина.
6. Выделенный полипептид, выбранный из группы, состоящей из:
a) выделенного полипептида, соответствующего любой из SEQ ID NO: 5-14 или функциональным эквивалентам любой из них, по меньшей мере на 85% гомологичным аминокислотной последовательности SEQ ID NO: 5-7;
b) выделенного полипептида, который может быть получен экспрессией полинуклеотида по п.1 или вектора по пп.2-4 в соответствующей клетке-хозяине, например Aspergillius niger;
c) выделенного полипептида, соответствующего любой из SEQ ID NO: 8-9 или SEQ ID NO: 10-14.
7. Способ получения полипептида по п.6, включающий этапы трансформации подходящей клетки-хозяина выделенным полинуклеотидом по п.1 или вектором по пп.2-4, культивирования указанной клетки в условиях, предоставляющих возможность для экспрессии указанного полинуклеотида и при необходимости очистки кодируемого полпептида из указанной клетки или культуральной среды.
8. Рекомбинантная клетка-хозяин, содержащая полинуклеотид по п.1 или вектор по пп.2-4.
9. Рекомбинантная клетка-хозяин, экспрессирующая полипептид по п.6.
10. Применение выделенного полипептида по п.6 при получении теста.
11. Способ получения теста, включающий этапы добавления полипептида по п.6 по меньшей мере к одному из ингредиентов теста.
12. Тесто, содержащее полипептид по п.6.
13. Тесто по п.12, имеющее повышенную стабильность.
14. Тесто по любому из пп.12-13, имеющее по меньшей мере одно из улучшенных свойств, выбранных из группы, состоящей из повышенной прочности, повышенной эластичности, повышенной стабильности, пониженной липкости и/или повышенной растяжимости теста.
15. Способ получения печеного изделия, включающий этап выпекания теста по любому из пп.12-14.
16. Печеное изделие, которое можно получить способом по п.15.
17. Печеное изделие по п.16, являющееся хлебом.
18. Печеное изделие по любому из пп.16-17, имеющее повышенный объем буханки.
19. Печеное изделие по любому из пп.16-18, имеющее по меньшей мере одно улучшенное свойство, выбранное из группы, состоящей из повышенного объема, улучшенного аромата, улучшенной структуры мякиша, улучшенной мягкости мякиша, уменьшенного образования пузырей и/или улучшенной способности не черстветь.
20. Применение выделенного полипептида по п.6 в одном из способов промышленного производства, выбранных из группы, состоящей из:
a) получения печеного изделия,
b) производства дигалактозилмоноглицерида из источника, содержащего дигалактозилдиглицерид,
c) производства сиропов глюкозы из клейковины пшеницы,
d) рафинирования гидратацией растительных масел или
е) модификации фосфолипидного эмульгатора.
Текст
Лаан Ван Дер Ян Метске, ШонефельдБергманс Маргот Элизабет Франсуаз (NL) Изобретение относится к вновь идентифицированным последовательностям полинуклеотидов,содержащим гены, кодирующие новые липолитические ферменты LIP01-LIP03. Фермент LIP01 может быть выделен из Magnaporthe grisae, LIP02 и LIP03 могут быть получены посредством мутации последовательности полинуклеотидов, кодирующей LIP01. Изобретение включает следующие признаки: полноразмерная кодирующая последовательность нового гена, который подходит для экспрессии в подходящей клетке-хозяине, а также последовательность аминокислот полноразмерного функционального белка и функциональные эквиваленты гена или последовательности аминокислот. Настоящее изобретение также относится к способам применения данных белков в промышленном производстве, например, в хлебопекарной промышленности, рафинировании растительного масла гидратацией, производстве или модификации пищевых эмульгаторов и производстве глюкозы из клейковины пшеницы.(71)(73) Заявитель и патентовладелец: ДСМ АйПи АССЕТС Б.В. (NL) 015172 Область техники, к которой относится изобретение Изобретение относится к вновь идентифицированным последовательностям полинуклеотидов, содержащим гены, кодирующие новый липолитический фермент. Ферменты могут быть выделены из Magnaporthe grisae. Изобретение включает следующие признаки: полиоразмерную кодирующую последовательность нового гена, а также последовательность аминокислот полиоразмерного функционального белка и функциональные эквиваленты гена или последовательности аминокислот. Настоящее изобретение также относится к способам применения данных белков в промышленном производстве, например в хлебопекарной промышленности. В настоящее изобретение также включены клетки, трансформированные полинуклеотидом согласно настоящему изобретению, пригодные для производства данных белков, а также клетки, в которых белок согласно настоящему изобретению генетически модифицирован с целью повышения или уменьшения его активности и/или уровня экспрессии. Сведения о предшествующем уровне техники Для улучшения пригодности теста для обработки и/или свойств готовых печеных изделий предпринимаются постоянные усилия по разработке технологических добавок с улучшенными свойствами. Технологические добавки определяются здесь как соединения, улучшающие пригодность теста для обработки и/или свойства готовых печеных изделий. Свойства теста, которые могут быть улучшены, включают стабильность, пригодность для механической переработки, газоудерживающая способность, уменьшенное образование пузырей, пониженную липкость, эластичность, растяжимость, пластичность и тому подобные. Свойства печеных изделий, которые могут быть улучшены, включают объем буханки, хрустящие свойства корки, текстуру мякиша, структуру мякиша, мягкость мякиша, аромат, относительную черствость и срок хранения. Данные технологические добавки для улучшения теста и/или печеных изделий можно подразделить на две группы: химические добавки и ферменты (также называемые пекарными ферментами). Химические добавки с улучшенными свойствами включают окислительные средства, такие как аскорбиновая кислота, бромат и азодикарбонат, восстановительные средства, такие как L-цистеин, и глутатион, эмульгаторы, действующие как улучшители теста, такие как сложные эфиры диацетилвинной кислоты и моно/диглицеридов (DATEM), стеароиллактилат натрия (SSL) или стеароиллактилат кальция(CSL), или действующие как смягчители мякиша, такие как моностеарат глицерина (GMS), и тому подобные, жировые материалы, такие как триглицериды (жир) или лецитин, и другие. Благодаря определяемой покупателем необходимости заменять химические добавки более натуральными продуктами были разработаны несколько пекарных ферментов, обладающих улучшающими тесто и/или печеные изделия свойствами, и которые применяются во всех возможных сочетаниях, в зависимости от конкретных условий применения в хлебопечении. Эмульгаторы, применяемые в хлебопекарной промышленности, грубо могут быть разделены на средства, смягчающие мякиш, и на средства, повышающие прочность теста. Для смягчения мякиша главным образом применяются дистиллированные моноглицериды. Комплексообразование моноглицеридов с крахмалом препятствует полной рекристаллизации крахмала, приводящей к начальной мягкости мякиша и/или уменьшению скорости затвердевания мякиша во время хранения печеных изделий. Для повышения прочности теста применяются несколько синтетических аналогов полярных липидов, таких как DATEM, CSL и SSL. Их действие в хлебопечении заключается в повышении стабильности теста,увеличении объема буханки и в том, что они дают тонкую и однообразную структуру мякиша. Относительно этого последнего аспекта следует заметить, что при применении данных эмульгаторов происходит также смягчение мякиша. Кроме того, могут быть достигнуты пониженная липкость теста, улучшенная пригодность теста для механической переработки, повышенный объем буханки печеного изделия,улучшенная структура мякиша, повышенная мягкость мякиша, улучшенные хрустящие свойства корки. Благодаря полярным и неполярным заместителям эмульгаторы могут концентрироваться на границах раздела фаз масло-вода и газ-вода. В хлебопечении пузырьки газа изначально заключены в клейковиново-крахмальную массу, но во время ферментации пузырьки газа увеличиваются в объеме и поверхности раздела между пузырьками газа содержат только жидкую пленку поверхностно-активного материала. В данных жидких пленках присутствуют эндогенные полярные липиды пшеничной муки, а также добавленные эмульгаторы. Известно, что полярные диацилглицериды, такие как лецитины или DATEM,полученные из диацилглицеридов, оказывают при хлебопечении по сравнению с их аналогамимоноацилглицеридами лишь ограниченное действие. Из уровня техники известно, что некоторые липолитические ферменты могут применяться в качестве заменителей DATEM, так, как, например, описано у L. Chirstiansen и сотр. в Proceedings of the ThirdSymposium on Enzymes in Grain Processing, 25-25 September 2002, стр. 269-274. Липолитические ферменты представляют собой ферменты, катализирующие гидролиз сложноэфирных связей в липидных субстратах. Липидные ферменты могут действовать на несколько типов липидов, начиная с глицеридов (например, триглицериды), фосфолипидов, сфинголипидов или гликолипидов, таких как галактолипиды. Глицериды являются сложными эфирами глицерина и жирных кислот. Триглицериды (известные-1 015172 также как триацилглицерин или триацилглицериды) присутствуют главным образом в растительных маслах и животном жире. Липазы (ЕС 3.1.1.3) определяют здесь как ферменты, вызывающие путем гидролиза образование одной или более жирных кислот из липидов, более конкретно, они гидролизуют сложноэфирную связь между жирной кислотой и гидроксильными группами глицерина. Галактолипиды состоят из глицериновой главной цепи, содержащей две связанные сложноэфирными связями жирные кислоты в крайнем (sn-1) и среднем (sn-2) положении, при этом третья гидроксильная группа связана с сахарными заместителями, такими как галактоза, например моногалактозилдиглицерид или дигалактозилдиглицерид. Галактолипаза (ЕС 3.1.1.26) катализирует гидролиз одной или обеих жирных ацильных групп в sn-1- и sn-2-положениях галактозилдиглицерида, соответственно. Фосфолипиды состоят из глицериновой главной цепи, содержащей две связанные сложноэфирными связями жирные кислоты в крайнем (sn-1) и среднем (sn-2) положении, при этом третья гидроксильная группа глицерина этерифицирована фосфорной кислотой. Фосфорная кислота, в свою очередь, может быть этерифицирована, например, до аминоспирта, такого как этаноламин (фосфатидилэтаноламин), холин (фосфатидилхолин). Фосфолипазы определяют здесь как ферменты, участвующие в гидролизе одной или более связей в фосфолипидах. Липолитические ферменты включают, например, липазы, галактолипазы и фосфолипазы, такие как,например, фосфолипазы A1, A2 и лизофосфолипазу, в зависимости от субстрата, на который они действуют. Существует постоянная потребность в улучшенных липолитических ферментах, которые могут применяться в качестве заменителей химических эмульгаторов, таких как DATEM, CSL и SSL, при производстве хлеба. Задача данного изобретения Задачей настоящего изобретения является предложить новый липолитический фермент, пригодный для применения в качестве ферментного заменителя химических эмульгаторов. Кроме того, задачей настоящего изобретения является предложить новые полинуклеотиды, кодирующие новый липолитический фермент. Дополнительной задачей является предложить липолитический фермент, получаемый натуральным или рекомбинантным способом, а также рекомбинантные штаммы, производящие его. Также частью настоящего изобретения являются слитые полипептиды и способы получения и применения полинуклеотидов и полипептидов согласно настоящему изобретению. Сущность изобретения В настоящем изобретении предложен новый липолитический фермент, пригодный для применения в качестве ферментного заменителя химических эмульгаторов. Неожиданно, новый липолитический фермент является чрезвычайно подходящим для применения в качестве заменителя химических эмульгаторов, поскольку данный фермент обладает по меньшей мере одним из следующих свойств in situ при использовании в тесте: относительно низкая активность по отношению к неполярным липидам,относительно высокая активность по отношению к полярным диациллипидам, по меньшей мере, по отношению к диацилгалактолипидам,относительно низкая активность по отношению к полярным моноацил-производным. Например, фермент согласно настоящему изобретению может демонстрировать in situ относительно низкую лизофосфолипазную активность и относительно низкую липазную активность. Все эти неожиданные свойства, как обнаружено, являются предпочтительными при применении в качестве заместителя химических эмульгаторов в тесте. Новый липолитический фермент дает одно или более улучшенных свойств теста и/или печеного изделия при данном использовании, выбранных из группы, в которую входят повышенная прочность теста,повышенная эластичность теста, повышенная стабильность теста, пониженная липкость теста, повышенная растяжимость теста, улучшенная пригодность теста для механической переработки, повышенный объем печеного изделия, улучшенная структура мякиша печеного изделия, уменьшенное образование пузырей в печеном изделии, повышенная мягкость печеного изделия, улучшенная способность печеного изделия не черстветь, улучшенная корка печеного изделия, или тех, которые имеют широкую специфичность к субстрату. Кроме того, в настоящем изобретении предложены новые полинуклеотиды, кодирующие новый липолитический фермент. В частности, в изобретении предложены полинуклеотиды, имеющие последовательность нуклеотидов, которая гибридизуется предпочтительно в очень строгих условиях с последовательностью, соответствующей любой из SEQ ID NO: 2, SEQ ID NO: 3 или SEQ ID NO: 4 (в дальнейшем именуемой "SEQ IDNO: 2-4"). Следовательно, в изобретении предложены нуклеиновые кислоты, которые по меньшей мере на 85%, предпочтительно по меньшей мере на 88%, более предпочтительно по меньшей мере на 90%,еще более предпочтительно по меньшей мере на 95, 96, 97, 98 или на 99% гомологичны последовательности, соответствующей SEQ ID NO: 2, SEQ ID NO: 3 или SEQ ID NO: 4. В одном из вариантов реализации настоящего изобретения для этого предложен выделенный полинуклеотид, который можно получить из мицелиального гриба, в частности, Magnaporthe является пред-2 015172 почтительным, и еще более предпочтительным является Magnaporthe grisae. В дополнительном варианте реализации настоящего изобретения такой выделенный полинуклеотид может быть получен синтетическим путем посредством способов, известных специалистам в данной области техники. В еще одном варианте реализации настоящего изобретения предложен выделенный полинуклеотид,содержащий последовательность нуклеиновых кислот, кодирующую полипептид, содержащий последовательность аминокислот, представленную в SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8,SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14 (в дальнейшем именуемых "SEQ ID NO: 5-14"), или функциональные эквиваленты любой из них. В дополнительном варианте реализации настоящего изобретения предложен выделенный полинуклеотид, кодирующий по меньшей мере один функциональный домен полипептида, соответствующего любой из SEQ ID NO: 5-14, или его функциональные эквиваленты. В другом варианте реализации настоящего изобретения предложен ген липолитического фермента,соответствующий любой из SEQ ID NO: 2-4, или его варианты или фрагменты, еще способные кодировать активный фермент. Настоящее изобретение относится также к векторам, содержащим последовательность полинуклеотидов согласно настоящему изобретению и праймеры, зонды и фрагменты, которые могут применяться для амплификации или обнаружения ДНК, согласно настоящему изобретению. В дополнительном предпочтительном варианте реализации настоящего изобретения предложен вектор, где последовательность полинуклеотидов согласно настоящему изобретению является функционально связанной по меньшей мере с одной регуляторной последовательностью, подходящей для экспрессии кодированной последовательности аминокислот в подходящей клетке-хозяине, такой как Aspergillus, более конкретно, Aspergillus niger, oryzae или nidulans. Клетка-хозяин предпочтительно представляет собой Aspergillus niger. Кроме того, в изобретении предложены способы получения полинуклеотидов и векторов согласно настоящему изобретению. Настоящее изобретение относится также к полученным рекомбинантным способом клеткамхозяевам, содержащим гетерологичные или гомологичные полинуклеотиды, согласно настоящему изобретению. В другом варианте реализации настоящего изобретения предложены полученные рекомбинантным способом клетки-хозяева, где экспрессия липолитического фермента, согласно настоящему изобретению,значительно повышена, или где повышена активность липолитического фермента. В другом варианте реализации настоящего изобретения предложены полученные рекомбинантным способом клетки-хозяева, содержащие гетерологичные или гомологичные ДНК, согласно настоящему изобретению, и где клетки способны производить функциональный липолитический фермент, согласно настоящему изобретению, предпочтительно клетки, способные к сверхэкспрессии липолитического фермента, согласно настоящему изобретению, например штамм Aspergillus, содержащий увеличенное число копий гена, согласно настоящему изобретению. В еще одном варианте реализации настоящего изобретения предложен очищенный полипептид. Полипептиды, согласно настоящему изобретению, включают полипептиды, кодируемые полинуклеотидами, согласно настоящему изобретению. Особенно предпочтительным является полипептид, соответствующий любой из SEQ ID NO: 5-14 или функциональным эквивалентам любой из них. Кроме того, в рамки настоящего изобретения попадают слитые белки, содержащие полинуклеотид,согласно настоящему изобретению. В настоящем изобретении предложены также способы получения полипептидов, согласно настоящему изобретению. Настоящее изобретение также относится к применению липолитического фермента, согласно настоящему изобретению, в любом способе изготовления, как описано здесь. Сведения, подтверждающие возможность осуществления изобретения Полинуклеотиды В одном из вариантов реализации настоящего изобретения предложены полинуклеотиды, кодирующие липолитические ферменты, условно названные LIP01, имеющие последовательность аминокислот, соответствующую любой из SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7 (в дальнейшем именуемым"SEQ ID NO: 5-7") или функциональным эквивалентам любой из них. В другом варианте реализации настоящего изобретения предложены полинуклеотиды, кодирующие липолитические ферменты, условно названные LIP02, имеющие последовательность аминокислот, соответствующую любой из SEQ ID NO: 8, SEQ ID NO: 9 (в дальнейшем именуемым "SEQ ID NO: 8-9") или функциональным эквивалентам любой из них. В дополнительном варианте реализации настоящего изобретения предложены полинуклеотиды, кодирующие липолитические ферменты, условно названные LIP03, имеющие последовательность аминокислот, соответствующую любой из SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12 (в дальнейшем именуемым "SEQ ID NO: 10-12") или функциональным эквивалентам любой из них. Последовательность гена, кодирующего LIP01, была определена путем секвенирования геномного клона, полученного из Magnaporthe grisae согласно SEQ ID NO: 1. Последовательности генов, кодирующих LIP02 и LIP03, были получены путем мутации геномного клона, полученного из Magnaporthe grisae-3 015172 согласно SEQ ID NO: 1. LIP02 образуется точечной мутацией относительно LIP01. В настоящем изобретении предложены последовательности полинуклеотидов, содержащие ген, кодирующий липолитические ферменты LIP01 - LIP03, а также кодирующие их последовательности. Соответственно, изобретение относится к выделенному полинуклеотиду, содержащему последовательность нуклеотидов, соответствующую любой из SEQ ID NO: 2-4 или его функциональным эквивалентам. В частности, изобретение относится к выделенному полинуклеотиду, гибридизируемому в строгих условиях, предпочтительно в очень строгих условиях, с полинуклеотидом согласно SEQ ID NO: 2-4. Предпочтительно такой выделенный полинуклеотид может быть получен из мицелиального гриба,в частности из Magnaporthaseae, таких как Magnaporthe, например grisae, oryzae, poae, rhizophila, salvinii,предпочтительно из Magnaporthe grisae. Более конкретно, изобретение относится к выделенному полинуклеотиду, содержащему последовательность нуклеотидов согласно SEQ ID NO: 2. В другом варианте реализации настоящего изобретения изобретение относится к выделенному полинуклеотиду, гибридизируемому в строгих условиях, предпочтительно в очень строгих условиях, с полинуклеотидом согласно SEQ ID NO: 3 или SEQ ID NO: 4. Такой выделенный полинуклеотид может быть получен синтетически при помощи способов, известных специалистам в данной области техники. Даже еще более предпочтительно, такой выделенный полинуклеотид может быть получен путем мутации полинуклеотида, полученного из мицелиального гриба, в частности из Magnaporthaseae, таких какMagnaporthe, например grisae, oryzae, poae, rhizophila, salvinii, предпочтительно из Magnaporthe grisae, и,наиболее предпочтительно, полинуклеотид содержит последовательность нуклеотидов согласно SEQ IDNO: 1 или SEQ ID NO: 2. Изобретение относится также к выделенному полинуклеотиду, кодирующему по меньшей мере один функциональный домен полипептида согласно любой из SEQ ID NO: 5-14 или функциональным эквивалентам любой из них. Используемые здесь термины "ген" и "рекомбинантный ген" относятся к молекулам нуклеиновых кислот, которые могут быть выделены из хромосомной ДНК, которая содержит открытую рамку считывания, кодирующую белок, например липолитический фермент Magnaporthe grisae. Ген может содержать кодирующие последовательности, некодирующие последовательности, интроны и регуляторные последовательности. Кроме того, ген относится к выделенной молекуле нуклеиновой кислоты, согласно данному здесь определению. Молекула нуклеиновой кислоты, согласно настоящему изобретению, такая как молекула нуклеиновой кислоты, содержащая последовательность нуклеотидов согласно любой из SEQ ID NO: 2-4, или ее функциональный эквивалент, может быть выделена при помощи стандартных микробиологических технологий и информации о последовательности, обеспеченной здесь. Например, посредством использования всей или части последовательности нуклеиновых кислот любой из SEQ ID NO: 2-4 в качестве гибридизационного зонда, молекулы нуклеиновых кислот, согласно настоящему изобретению, могут быть выделены при помощи стандартных технологий гибридизации и клонирования (например, как описано уLaboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Кроме того, молекула нуклеиновой кислоты, включающая все или часть любой из SEQ ID NO: 2-4,может быть выделена посредством полимеразной цепной реакции (ПНР) при помощи синтетических олигонуклеотидных праймеров, сконструированных на основе информации, содержащейся в любой из последовательностей SEQ ID NO: 2-4. Нуклеиновая кислота согласно настоящему изобретению может быть амплифицирована при помощи использования кДНК, иРНК или, альтернативно, используя геномную ДНК, в качестве матрицы, и соответствующих олигонуклеотидньгх праймеров, согласно стандартным технологиям амплификации ПЦР. Таким образом амплифицированная нуклеиновая кислота может быть клонирована в соответствующий вектор и определена при помощи секвенирования ДНК. Кроме того, олигонуклеотиды, соответствующие последовательностям нуклеотидов, согласно настоящему изобретению, или гибридизируемые с последовательностями нуклеотидов, согласно настоящему изобретению, могут быть получены при помощи стандартных технологий синтеза, например при помощи автоматического ДНК-синтезатора. В предпочтительном варианте реализации настоящего изобретения выделенная молекула нуклеиновой кислоты согласно настоящему изобретению содержит последовательность нуклеотидов, представленную в любой из SEQ ID NO: 2-4. Последовательность SEQ ID NO: 2 соответствует кодирующей области кДНК липолитического фермента, на основе геномной ДНК согласно SEQ ID NO: 1. Данная кДНК содержит последовательности, кодирующие LIP01 Magnaporthe grisae согласно любой из SEQ ID NO: 5-7. В другом предпочтительном варианте реализации выделенная молекула нуклеиновой кислоты согласно настоящему изобретению содержит молекулу нуклеиновой кислоты, комплементарную по отношению к последовательности нуклеотидов, представленной в SEQ ID NO: 2-4, или функциональному эквиваленту любой из них. Молекула нуклеиновой кислоты, комплементарная другой последовательности нуклеотидов, представляет собой молекулу нуклеиновой кислоты, в значительной степени комплементарную другой по-4 015172 следовательности нуклеотидов, таким образом, что она может гибридизироваться с другой последовательностью нуклеотидов, образуя таким образом стабильный дуплекс. Один аспект настоящего изобретения относится к выделенным молекулам нуклеиновой кислоты,которые кодируют полипептид, согласно настоящему изобретению, или его функциональный эквивалент, такой как биологически активный фрагмент или домен, а также к молекулам нуклеиновой кислоты,достаточным для использования в качестве гибридизационных зондов для идентификации молекул нуклеиновой кислоты, кодирующих полипептид, согласно настоящему изобретению, и к фрагментам таких молекул нуклеиновой кислоты, пригодным для применения в качестве ПЦР-праймеров для амплификации или мутации молекул нуклеиновой кислоты."Выделенный полинуклеотид" или "выделенная нуклеиновая кислота" представляют собой ДНК или РНК, которые не являются непосредственно смежными с обеими кодирующими последовательностями, с которыми они являются непосредственно смежными (один на 5'-конце и один на 3'-конце) в природном геноме организма, из которого они получены. Таким образом, в одном из вариантов реализации выделенная нуклеиновая кислота содержит несколько или все 5'-некодирующие (например, промотор) последовательности, которые являются непосредственно смежными с кодирующей последовательностью. Следовательно, термин включает, например, рекомбинантную ДНК, введенную в вектор, в автономно реплицирующуюся плазмиду или вирус, или в геномную ДНК прокариота или эукариота, или который существует в виде отдельной молекулы (например, кДНК или фрагмент геномной ДНК, полученный посредством ПЦР или посредством обработки эндонуклеазой рестрикции), независимой от других последовательностей. Он также включает рекомбинантную ДНК, являющуюся частью гибридного гена,кодирующего добавочный полипептид, который является в значительной степени свободным от клеточного материала, вирусного материала, или от культуральной среды (когда он получен посредством технологий рекомбинантнои ДНК), или от химических предшественников или других химикатов (когда он синтезирован химическим путем). Кроме того, "фрагмент выделенной нуклеиновой кислоты" представляет собой фрагмент нуклеиновой кислоты, который не встречается в природе в виде фрагмента и, предположительно, не встречается в естественном состоянии. Используемые здесь термины "полинуклеотид" или "молекула нуклеиновой кислоты" включает молекулы ДНК (например, кДНК или геномной ДНК) и молекулы РНК (например, иРНК), а также аналоги ДНК и РНК, полученные с использованием аналогов нуклеотидов. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, однако, предпочтительно, она является двухцепочечной ДНК. Нуклеиновая кислота может быть синтезирована с использованием аналогов олигонуклеотидов или производных (например, нуклеотидов инозина или фосфоротиоата). Такие олигонуклеотиды могут использоваться, например, для получения нуклеиновых кислот с измененной способностью к спариванию оснований или повышенной устойчивостью к нуклеазам. В другом варианте реализации настоящего изобретения предложена выделенная молекула нуклеиновой кислоты, которая является антисмысловой к молекулам нуклеиновой кислоты LIP01-LIP03, например кодирующей цепи молекулы нуклеиновой кислоты LIP01-LIP03. Также в рамки настоящего изобретения попадают комплементарные цепи молекул нуклеиновой кислоты, описанных здесь. Ошибки секвестрования Информация, содержащаяся в последовательности, как предложено здесь, не должна быть истолкована настолько узко, чтобы требовать включения ошибочно идентифицированных оснований. Конкретные последовательности, описанные здесь, могут легко использоваться для выделения полного гена из мицелиальных грибов, в частности Magnaporthe grisae, который, в свою очередь, может быть легко подвергнут дополнительному секвенированию для идентификации ошибок. Если не оговорено противное, все последовательности нуклеотидов, определенные посредством секвенирования молекулы ДНК, были здесь определены при помощи автоматического секвенатора ДНК, и все последовательности аминокислот полипептидов, кодируемые молекулами ДНК, определенными здесь, были предсказаны путем трансляции последовательности ДНК, определенной как указано выше. Следовательно, как известно из уровня техники для любой последовательности ДНК, определенной при помощи данного автоматического способа, любая последовательность нуклеотидов, определенная здесь,может содержать некоторое количество ошибок. Последовательности нуклеотидов, определенные автоматически, типично идентичны по меньшей мере на примерно 90%, более типично по меньшей мере на примерно 95% по отношению к идентичной по меньшей мере на примерно 99,9% фактической последовательности нуклеотидов секвенированной молекуле ДНК. Фактическая последовательность может быть более точно определена при помощи других способов, включая способы ручного секвенирования ДНК,хорошо известные из уровня техники. Как также известно из уровня техники, единичная инсерция или делеция в определенной последовательности нуклеотидов по сравнению с фактической последовательностью будет вызывать сдвиг рамки при трансляции последовательности нуклеотидов, так что предсказанная последовательность аминокислот, кодируемая определенной последовательностью нуклеотидов,будет совершенно отличной от последовательности аминокислот, фактически кодируемой секвенированной молекулой ДНК, начиная с точки такой инсерции или делеции. Специалист в данной области техники способен идентифицировать такие ошибочно определенные-5 015172 основания и знает, как исправлять такие ошибки. Фрагменты нуклеиновой кислоты, зонды и праймеры Молекула нуклеиновой кислоты, согласно настоящему изобретению, может содержать только участок или фрагмент последовательности нуклеиновых кислот, согласно любой из SEQ ID NO: 2-4, например фрагмент, который может быть использован в качестве зонда или праймера, или в качестве фрагмента, кодирующего участок белка LIP01-LIP03. Последовательность нуклеотидов, определенная путем клонирования гена LIP01-LIP03 и кДНК дает возможность для получения зондов и праймеров, созданных для применения при идентификации и/или клонировании других представителей семейства LIP01-LIP03, а также гомологов LIP01-LIP03 других видов. Зонд/праймер обычно содержит в значительной степени очищенный олигонуклеотид, который обычно содержит участок последовательности нуклеотидов, который гибридизируется предпочтительно в строгих условиях по меньшей мере с примерно 12 или 15, предпочтительно примерно 18 или 20, предпочтительно примерно 22 или 25, более предпочтительно примерно 30, 35, 40, 45, 50, 55, 60, 65 или 75 или более последовательными нуклеотидами последовательности нуклеотидов, соответствующей любой из SEQ ID NO: 2-4 или ее функциональному эквиваленту. Зонды на основе последовательностей нуклеотидов LIP01-LIP03 могут применяться для обнаружения транскриптов или геномных последовательностей LIP01-LIP03, кодирующих их или гомологичные белки, например, в других организмах. В предпочтительных вариантах реализации зонд дополнительно содержит присоединенную к нему метку, например, метка может представлять собой радиоизотоп, флуоресцентное соединение, фермент или кофермент. Такие зонды могут также применяться как часть диагностического набора для обнаружения клеток, экспрессирующих белок LIP01-LIP03. Идентичность и гомология Термины "гомология" или "процент идентичности" применяются здесь взаимозаменяемо. Для цели настоящего изобретения здесь определено, что для того чтобы определить процент идентичности двух последовательностей аминокислот или двух последовательностей нуклеиновых кислот, последовательности выравнивают для оптимального сравнения (например, в первую последовательность аминокислот или последовательность нуклеиновых кислот для оптимального выравнивания со второй последовательностью аминокислот или последовательностью нуклеиновых кислот могут быть введены бреши). Затем сравнивают аминокислотные остатки или нуклеотиды в соответствующих позициях аминокислот или положениях нуклеотидов. Когда позиция в первой последовательности занята таким же аминокислотным остатком или нуклеотидом в соответствующей позиции во второй последовательности, это означает, что молекулы идентичны в этой позиции. Процент идентичности двух последовательностей является функцией количества идентичных положений у данных последовательностей (например, % идентичности = количество идентичных положений/общее количество положений (т.е. перекрьщающихся положений)100). Предпочтительно, две последовательности имеют одинаковую длину. Специалисту в данной области известен тот факт, что существует некоторое различие в компьютерных программах, применяемых для определения гомологичности двух последовательностей. Например, сравнение последовательностей и определение процента идентичности двух последовательностей может быть выполнено при помощи математического алгоритма. В предпочтительном варианте реализации процент идентичности двух последовательностей аминокислот определяют при помощи алгоритмаNeedleman и Wunsch (J. Mol. Biol. (48): 444-453 (1970, который встроен в программу GAP пакета программного обеспечения Accelrus GCG (доступного на http://www.accelrus.com/products/gcl/), использующего или матрицу Blossom 62, или матрицу РАМ 250 и веса бреши 16, 14, 12, 10, 8, 6 или 4 и веса длины 1, 2, 3, 4, 5 или 6. Специалист в данной области оценит, что данные различные параметры будут давать слабо различающиеся результаты, но общее значение процента идентичности двух последовательностей при использовании других алгоритмов в значительной степени не изменится. В еще одном варианте реализации процент идентичности между двумя последовательностями нуклеотидов определяют при помощи программы GAP пакета программного обеспечения Accelrus GCG(доступного на http://www.accelrus.com/products/gcl/), использующего матрицу NWSgapdna.CMP и веса бреши 40, 50, 60, 70 или 80 и веса длины 1, 2, 3, 4, 5 или 6. В другом варианте реализации процент идентичности двух аминокислот или последовательностей нуклеотидов определяют при помощи алгоритма Е. Meyers и W. Miller (CABIOS, 4:11-17 (1989, встроенного в программу ALIGN (версия 2.0) (доступную у ALIGN Query, использующем данные последовательностей с сервера Genestream IGH Monpellier,Франция, http://vega.igh.cnrs.fr/bin/align-guess.cgi) при помощи таблицы весов остатков РАМ 120, штраф за длину бреши 12 и штраф за брешь 4. Последовательности нуклеиновых кислот и белков согласно настоящему изобретению могут дополнительно применяться в качестве "искомой последовательности" для осуществления поиска в общедоступных базах данных для идентификации других представителей семейства или родственных последовательностей. Подобный поиск может быть осуществлен при помощи программ NBLAST и XBLAST(версия 2.0) Altschul и сотр. (1990) J. Mol. Biol. 215:403-10. Поиск нуклеотидов BLAST может быть осуществлен при помощи программы NBLAST, показатель = 100, длина кодового слова = 12, для того чтобы получить последовательности нуклеотидов, гомологичные молекулам нуклеиновой кислоты LIP01-6 015172LIP03, согласно настоящему изобретению. Поиск белков BLAST может быть осуществлен при помощи программы XBLAST, показатель = 50, длина кодового слова = 3, для того чтобы получить последовательности аминокислот, гомологичные молекулам белков LIP01-LIP03, согласно настоящему изобретению. Для того чтобы получить выравнивания, содержащие бреши, для целей сравнения, можно использовать Gapped BLAST, как описано у Altschul и сотр., (1997) Nucleic Acids Res. 25 (17): 3389-3402. При использовании программ BLAST и Gapped BLAST, могут быть применены параметры по умолчанию для соответствующих программ (например, XBLAST и NBLAST). См. домашнюю страницу National Centerfor Biotechnology Information на http://www.ncbi.nlm.nih.gov/. Гибридизация Используемый здесь термин "гибридизация" описывает условия гибридизации и промывания, при которых последовательности нуклеотидов, гомологичные друг другу по меньшей мере на примерно 60%,по меньшей мере на примерно 70%, по меньшей мере на примерно 80%, более предпочтительно по меньшей мере на примерно 85%, еще более предпочтительно по меньшей мере на примерно 90%, наиболее предпочтительно по меньшей мере на примерно 95%, типично остаются гибридизованными друг с другом. Предпочтительным примером (но не ограничиваясь им) таких условий гибридизации является гибридизация в 6 Х хлорид натрия/цитрат натрия (SSC) при примерно 45 С, с одним или более последующим промыванием в 1X SSC, 0,1% SDS при 50 С, предпочтительно при 55 С, предпочтительно при 60 С и более предпочтительно при 65 С. Очень строгие условия включают, например, гибридизацию при 68 С в 5 х SSC/5x раствор Денхардта/ 1,0% SDS и промывание в 0,2x SSC/0,1% SDS при комнатной температуре. Альтернативным образом,промывание может быть осуществлено при 42 С. Опытный исполнитель в данной области знает, какие условия применять в качестве строгих условий гибридизации и очень строгих условий гибридизации. Дополнительное руководство относительно таких условий легко доступно из уровня техники, например, у Sambrook и сотр., 1989, Molecular Cloning,A Laboratory Manual, Cold Spring Harbor Press, N.Y.; и Ausubel и сотр. (eds.), 1995, Current Protocols inMolecular Biology, (John WhileySons, N.Y.). Конечно, полинуклеотиды, которые гибридизируются только с поли (А) последовательностью (такие, как 3'-концевой поли(А) тракт иРНК) или с комплементарным участком Т (или U) остатков, не могут быть включены в полинуклеотид, согласно настоящему изобретению, использованный для специфической гибридизации с участком нуклеиновой кислоты, согласно настоящему изобретению, поскольку такой нуклеотид будет гибридизироваться с любой молекулой нуклеиновой кислоты, содержащей поли(А) участок или его комплемент (например, практически любой двухцепочечный клон кДНК). Получение полноразмерной ДНК из других организмов В типичном способе библиотеки кДНК, сконструированные из других организмов, например мицелиальных грибов, в частности из вида Magnaporthe, могут быть подвергнуты скринингу. Например, штаммы Magnaporthe могут быть подвергнуты скринингу на гомологи полинуклеотидовLIP01-LIP03 при помощи нозерн-блоттинга. При обнаружении транскриптов, гомологичных полинуклеотидам, согласно настоящему изобретению, библиотеки кДНК могут быть сконструированы из РНК,выделенных из соответствующего штамма при помощи стандартных технологий, хорошо известных специалистам в данной области техники. Альтернативным образом полная библиотека геномной ДНК может быть подвергнута скринингу при помощи зонда, гибридизующегося с полинуклеотидом LIP01LIP03, согласно настоящему изобретению. Гомологичные последовательности генов могут быть выделены, например, при помощи ПЦР с использованием двух пулов вырожденных олигонуклеотидных праймеров, созданных на основе последовательностей нуклеотидов, как описано здесь. Матрицей для реакции может быть кДНК, полученная путем обратной транскрипции иРНК, полученной из штаммов, про которые известно или предполагается, что они экспрессируют полинуклеотид,согласно настоящему изобретению. Продукт ПЦР может быть субклонирован и секвенирован для того,чтобы убедиться, что амплифицированные последовательности представляют последовательности новойLIP01-LIP03 последовательности нуклеиновых кислот, или ее функциональный эквивалент. ПЦР-фрагмент может затем быть использован для выделения клона первичной кДНК при помощи разнообразных известных способов. Например, амплифицированный фрагмент может быть помечен и использован для обнаружения библиотеки кДНК бактериофага или космиды. Альтернативным образом,помеченный фрагмент может быть использован для обнаружения геномной библиотеки. Технология ПЦР может также применяться для выделения полноразмерных последовательностей кДНК из других организмов. Например, посредством стандартных процедур может быть выделена РНК из соответствующего клеточного или тканевого источника. Реакция обратной транскрипции РНК может быть проведена при помощи олигонуклеотидного праймера, специфичного к большей части 5'-конца амплифицированного фрагмента для прайминга синтеза первой цепи. Полученный гибрид РНК/ДНК может затем быть "наращен" (например, гуанинами) при помощи стандартной реакции терминальной трансферазы, гибрид может быть обработан рибонуклеазой Н, после-7 015172 чего может быть примирован синтез второй цепи (например, при помощи поли-С праймера). Таким образом, последовательности кДНК, находящиеся против направления транскрипции от амплифицированного фрагмента, могут быть легко выделены. Обзор применимых стратегий клонирования см., например,в Sambrook и сотр. (упомянуто ранее) и Ausubel и сотр. (упомянуто ранее). Векторы Другой аспект настоящего изобретения относится к векторам, предпочтительно экспрессирующим векторам, содержащим нуклеиновую кислоту, кодирующую белок LIP01-LIP03 или его функциональный эквивалент. Используемый здесь термин "вектор" относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой он связан. Один тип вектора представляет собой "плазмиду", которая относится к циклическому петлевому фрагменту двухцепочечной ДНК,внутри которой могут находиться связанные дополнительные участки ДНК. Другой тип вектора представляет собой вирусный вектор, где дополнительные участки ДНК могут быть связаны с геномом вируса. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены(например, бактериальные векторы, имеющие бактериальное происхождение репликации или эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) встроены в геном клетки-хозяина при введении в клетку-хозяина и поэтому реплицируются вместе с геномом носителя. Кроме того, определенные векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы названы здесь как "векторы экспрессии". В общем случае, векторы экспрессии, применяемые в методе рекомбинантных ДНК, часто имеют форму плазмид. Термины "плазмида" и "вектор" могут здесь применяться взаимозаменяемо, поскольку плазмида представляет собой наиболее часто используемую форму вектора. Однако настоящее изобретение включает другие формы векторов экспрессии, такие как вирусные векторы (например, ретровирусы, дефектные по репликации, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции. Рекомбинантные векторы экспрессии, согласно настоящему изобретению, содержат нуклеиновую кислоту, согласно настоящему изобретению, в форме, пригодной для экспрессии нуклеиновой кислоты в клетке-хозяине, что означает, что рекомбинантный вектор экспрессии содержит одну или более регуляторную последовательность, выбранную на основе клеток-хозяев, используемых для экспрессии, которая функционально связана с последовательностью нуклеиновых кислот, которую нужно экспрессировать. В рамках рекомбинантного вектора экспрессии "функционально связанный" означает, что представляющая интерес последовательность нуклеотидов связана с регуляторной последовательностью (последовательностями) способом, разрешающим экспрессию последовательности нуклеотидов (например, в системе транскрипция/трансляция in vitro или в клетке-хозяине, когда вектор введен в клетку-хозяина). Термин"регуляторная последовательность" означает включающий промоторы, энхансеры и другие регулирующие экспрессию элементы (например, сигнал полиаденилирования). Такие регуляторные последовательности описаны, например, в Goeddel; Gene Expression Technology: Methods in Enzymology 185, AcademicPress, San Diego, CA (1990). Регуляторные последовательности включают последовательности, управляющие конститутивной экспрессией последовательности нуклеотидов в клетках-хозяевах многих типов,и последовательности, которые управляют экспрессией последовательности нуклеотидов только в определенных типах клеток-хозяев (например, тканеспецифичные регуляторные последовательности). Специалисты в данной области техники оценят тот факт, что конструкция вектора экспрессии может зависеть от таких факторов, как выбор клетки-хозяина, которую нужно трансформировать, уровень экспрессии требуемого белка, и т.д. Векторы экспрессии, согласно настоящему изобретению, могут быть введены в клетки-хозяина для получения, таким образом, белков или пептидов, кодируемых нуклеиновыми кислотами, согласно описанному здесь (например, белки LIP01-LIP03, мутантные формы белков LIP01LIP03, фрагменты, варианты их функциональных эквивалентов, слитые белки и т.д.). Рекомбинантные векторы экспрессии, согласно настоящему изобретению, могут быть сконструированы для экспрессии белков LIP01-LIP03 в прокариотных или эукариотных клетках. Например, белкиLIP01-LIP03 могут быть экспрессированы в клетках бактерий, таких как Е. coli, клетках насекомых (при помощи векторов экспрессии бакуловируса), дрожжевых клетках или клетках млекопитающих. Подходящие клетки хозяева обсуждены дополнительно в Goeddel, Gene Expression Technology: Methoda in Enzymology 185, Academic Press, San Diego, CA (1990). Альтернативным образом, рекомбинантный вектор экспрессии может быть транскрибирован и транслирован in vitro, например, при помощи Т 7-промотора регуляторных последовательностей промотора и Т 7 полимеразы. Векторы экспрессии, применяемые в настоящем изобретении, включают векторы хромосомного,эписомального и вирусного происхождения, например векторы, полученные из бактериальных плазмид,бактериофагов, эписом дрожжей, элементов хромосом дрожжей, вирусов, таких как бакуловирусы, паповавирусы, вирусы коровьей оспы, аденовирусы, птичьи поксвирусы, вирусы псевдобешенства и ретровирусы, а также векторы, полученные из их комбинаций, такие как векторы, полученные из плазмид и генетических элементов бактериофагов, таких как космиды и фагемиды. ДНК-вставка должна быть функционально связана с подходящим промотором, таким как PL промотор фага лямбда, lac, trp и tac промоторы Е. coli, ранние и поздние промоторы SV 40 и промоторы рет-8 015172 ровирусных LTR, если назвать несколько. Другие подходящие промоторы известны специалистам в данной области техники. В специальном варианте предпочтительны промоторы, способные управлять высоким уровнем экспрессии липолитического фермента в мицелиальных грибах. Такие промоторы известны из уровня техники. Экспрессирующие конструкции могут содержать участки для инициации и терминации транскрипции, а также в транскрибированной области связывающий рибосомы участок для трансляции. Кодирующий участок в зрелых транскриптах, экспрессированных конструктами, будет включать кодон инициации AUG в начале и терминирующий кодон, соответственно расположенный на конце полипептида, который должен быть транслирован. Векторная ДНК может быть введена в прокариотные или эукариотные клетки посредством стандартных технологий трансформации или трансфекции. Используемые здесь термины "трансформация" и"трансфекция" относятся к разнообразным принятым в данной области техники технологиям введения инородной нуклеиновой кислоты (например, ДНК) в клетку-хозяина, включая совместное осаждение фосфатом кальция и хлоридом кальция, опосредованную декстраном трансфекцию, трансдукцию, инфекцию, липофекцию, опосредованную катионоактивным липидом трансфекцию или электропорацию. Способы, применимые для трансформации или трансфекции клеток-хозяев, можно найти в Sambrook и сотр. (Molecular Cloning: A Laboratory Manual, 2nd, ed. Cold Spring Harbor Laboratory, Cold Sprind HarborPress, Cold Spring Harbor, NY, 1989), Davis и сотр., Basic Methods in Molecular Biology (1986) и в других лабораторных справочниках. Относительно стабильной трансфекции клеток млекопитающих известно, что, в зависимости от используемых вектора экспрессии и способа трансфекции, лишь малая часть клеток может интегрировать инородную ДНК в свой геном. Для того чтобы идентифицировать и селектировать эти вставки, в клеткихозяева, наряду с представляющим интерес геном, обычно вводят ген, кодирующий селектируемый маркер (например, резистентность к антибиотикам). Предпочтительные селектируемые маркеры включают селектируемые маркеры, которые дают резистентность к лекарствам, такие как G418, гидромицин и метатрексат. Нуклеиновая кислота, кодирующая селектируемый маркер, может быть введена в клеткухозяина при помощи того же вектора, что и вектор, кодирующий белок LIP01-LIP03, или при помощи отдельного вектора. Клетки, стабильно трансфицированные введенной нуклеиновой кислотой, могут быть идентифицированы посредством лекарственного отбора (например, клетки, которые инкорпорировали ген селектируемого маркера, выживут, тогда как другие клетки погибнут). Экспрессия белков, в прокариотах часто проводится в Е. coli, при помощи векторов, содержащих конститутивные или индуцибельные промоторы, управляющие экспрессией как слитых, так и не слитых белков. Слитые векторы добавляют ряд аминокислот к кодируемому ими белку, например амино-конец рекомбинантного белка. Такие слитые векторы обычно служат трем целям: 1) повышают экспрессию рекомбинантного белка; 2) повышают растворимость рекомбинантного белка и 3) помогают очистке рекомбинантного белка, действуя как лиганд при аффинной очистке. Часто в слитые векторы экспрессии вводят участок протеолитического расщепления в место соединения участка слияния и рекомбинантного белка для того, чтобы сделать возможным отделение рекомбинантного белка от участка слияния после очистки слитого белка. Как указано, векторы экспрессии предпочтительно содержат селектируемые маркеры. Такие маркеры включают дигидрофолатредуктазу или резистентность к неомицину для культуры эукариотных клеток и устойчивость к тетрациклину или ампициллину для культивирования в Е. coli и других бактериях. Репрезентативные примеры подходящих носителей включают клетки бактерий, таких как Е. coli, Streptomyces и Salmonella typhimurium; клетки грибов, таких как дрожжи; клетки насекомых, такие как клеткиDrosophila S2 и Spodoptera Sf9; клетки животных, такие как СНО, COS и меланома Bowes; и клетки растений, известные из уровня техники. Векторы, предпочтительные для применения в бактериях, раскрыты, например, в WO-A12004/074468, содержание данной ссылки включено здесь в качестве ссылки. Другие подходящие векторы очевидны для опытных специалистов в данной области техники. Известные бактериальные промоторы, пригодные для использования в настоящем изобретении,включают промоторы, раскрытые в WO-A1-2004/074468, содержание данной ссылки включено здесь путем отсылки. Транскрипция ДНК, кодирующей полипептиды согласно настоящему изобретению высшими эукариотами, может быть увеличена посредством инсерции в вектор энхансерной последовательности. Энхансеры являются действующими в цис-положении элементами ДНК, обычно приблизительно от 10 до 300 п.н., которые действуют, увеличивая транскрипционную активность промотора для данного типа клеток-хозяев. Примеры энхансеров включают энхансер SV40, который находится на поздней стороне точки начала репликации при п.н. 100-270, ранний энхансер промотора цитомегаловируса, энхансер полиомы на поздней стороне точки начала репликации, и энхансеры аденовируса. Для секреции транслированного белка в просвет эндоплазматической сети, внутрь периплазматического пространства или во внеклеточную среду, в экспрессируемый полипептид может быть инкорпорирован соответствующий сигнал к секреции. Сигналы могут быть эндогенными к полипептиду или могут являться гетерологичными сигналами.-9 015172 Полипептид LIP01-LIP03 может быть экспрессирован в модифицированной форме, такой, как слитый белок, и может включать не только сигналы секреции, но также и дополнительные гетерологичные функциональные области. Так, например, к N-концу полипептида может быть добавлена область дополнительных аминокислот, особенно имеющих заряд аминокислот, для улучшения стабильности и персистенции в клетке-хозяине, во время очистки или при последующем использовании и хранении. Кроме того, в полипептид могут быть введены пептидные заместители, способствующие очистке. Полипептиды согласно настоящему изобретению В настоящем изобретении предложен выделенный полипептид, содержащий последовательность аминокислот, соответствующую любой из SEQ ID NO:5-14, и последовательность аминокислот, которая может быть получена экспрессией полинуклеотида согласно любой из SEQ ID NO: 2-4 в приемлемом хозяине. Также настоящее изобретение включает пептид или полипептид, содержащий функциональный эквивалент вышеупомянутых полипептидов. Как известно специалистам в данной области техники, возможно, что из-за ошибок процессинга во время созревания N-концы SEQ ID NO: 5-14 должны быть гетерологичными, также как и С-конец SEQID NO: 5-14. В частности, такие ошибки процессинга должны иметь место при сверхэкспрессии полипептида. Дополнительно, экзопротеазная активность может приводить к росту гетерогенности. Степень,в которой проявляется данная гетерогенность, зависит также от хозяина и применяемых протоколов ферментации. Такие артефакты С-терминального процессинга должны приводить к образованию более коротких или более длинных полипептидов, чем представлено в SEQ ID NO: 5-14. В результате таких ошибок N-конец также может быть гетерологичным. В дополнительном варианте реализации изобретения предложен выделенный полинуклеотид, кодирующий по меньшей мере один функциональный домен полипептида, согласно любой из SEQ ID NO: 514, содержащий дополнительные остатки и точку начала трансляции в позиции -1, или -2, или -3 и т.д. Альтернативным образом, в нем могут отсутствовать некоторые остатки и как следствие - точка начала трансляции в позиции 2, или 3, или 4 и т.д. Более конкретно, для LIP01, в одном из вариантов реализации настоящего изобретения SEQ ID NO: 5 раскрывает белок, как непосредственно транслированный с кДНК, так и представленный в SEQ ID NO: 2. Обычно такой белок будет процессироваться перед образованием зрелого белка и, например, будет утрачивать сигнальную последовательность, предпочтительно давая тем самым SEQ ID NO: 6 или 7. Для последовательности аминокислот, представленной в SEQ ID NO: 6 и SEQ ID NO:7, N-конец, в том случае, если он содержит дополнительные остатки, должен содержать следующие дополнительные последовательности аминокислот R, GR или EGR, соответствующие инициации N-конца в позициях -1, -2 или -3,соответственно. Аналогично, артефакты С-концевого процессинга должны приводить к образованию более коротких или более длинных полипептидов. В специальном случае SEQ ID NO: 7, С-конец в том случае, если он содержит дополнительные остатки, предпочтительно содержит следующие дополнительные последовательности аминокислот R, RR или RRD, в соответствии с С-концом, продолженном в позициях 310 +1, +2 или +3, соответственно. Более конкретно, для LIP02 в еще одном варианте реализации, в настоящем изобретении предложен выделенный полинуклеотид, содержащий последовательность нуклеиновых кислот, кодирующую полипептид, содержащий последовательность аминокислот, представленную в SEQ ID NO: 9, или функциональные эквиваленты любой из них. SEQ ID NO: 8 раскрывает белок, как непосредственно транслированный с кДНК, так и представленный в SEQ ID NO: 3. Обычно такой белок будет процессироваться перед образованием зрелого белка и, например, будет утрачивать сигнальную последовательность, предпочтительно давая тем самым SEQ ID NO: 9. При этом С- и N-концы полученного белка должны быть гетерологичными, например, из-за артефактов процессинга. Более конкретно, для LIP03, в еще одном варианте реализации, SEQ ID NO: 10 раскрывает белок,как непосредственно транслированный с кДНК, так и представленный в SEQ ID NO: 4. Обычно такой белок будет процессироваться перед образованием зрелого белка и, например, будет утрачивать сигнальную последовательность, предпочтительно давая тем самым SEQ ID NO: 11, 12, 13 или 14. Вышеупомянутые полипептиды собирательно обозначены термином "полипептиды согласно настоящему изобретению". Предполагается, что термины "пептид" и "олигопептид" являются синонимами (поскольку они являются общепринятыми), и каждый термин может использоваться взаимозаменяемо, поскольку в контексте требуется обозначать цепь, состоящую по меньшей мере из двух аминокислот, связанных пептидильными связями. Слово "полипептид" применяется здесь для цепей, содержащих более семи аминокислотных остатков. Все формулы олигопептидов и полипептидов или последовательности в настоящем изобретении написаны слева направо и в направлении от амино-конца к карбоксильному концу. Однобуквенный код аминокислот, используемый здесь, является общеизвестным в данной области техники, и его можно найти в Sambrook и сотр. (Molecular Cloning: A Laboratory Manual, 2nd, ed. Cold Spring HarborLaboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Под "выделенным" полипептидом или белком понимается полипептид или белок, удаленный из своего нативного окружения. Например, полученные рекомбинантным способом полипептиды и белки,- 10015172 экспрессированные в клетках-хозяевах, рассматриваются как выделенные в целях настоящего изобретения, поскольку являются нативными и рекомбинантными полипептидами, которые в значительной степени очищены посредством любого подходящего способа, такого как, например, одношаговый способ очистки, раскрытый в Smith и Johnson, Gene 67:31-40 (1988). Липолитический фермент LIP01-LIP03, согласно настоящему изобретению, может быть извлечен и очищен от культур рекомбинантных клеток способами, известными из уровня техники (Protein Purification Protocols, Methods in Molecular Biology series by Paul Cutler, Humana Press, 2004). Полипептиды, согласно настоящему изобретению, включают естественным образом очищенные продукты, продукты химического синтеза, а также продукты, произведенные рекомбинантными способами из прокариотного и эукариотного носителя, включающего, например, клетки бактерий, дрожжей,высших растений, насекомых и млекопитающих. В зависимости от носителя, используемого в рекомбинантном способе получения, полипептиды, согласно настоящему изобретению, могут быть гликозилированными или не гликозилированными. Дополнительно, полипептиды, согласно настоящему изобретению, могут содержать модифицированный первичный метиониновый остаток, в некоторых случаях являющийся результатом опосредованных носителем процессов. Фрагменты белков В настоящем изобретении предложены также биологически активные фрагменты полипептидов, согласно настоящему изобретению. Биологически активные фрагменты полипептида, согласно настоящему изобретению, включают полипептиды, содержащие последовательности аминокислот, в значительной степени идентичные или полученные и последовательности аминокислот белка LIP01-LIP03 (например, последовательности SEQID NO: 5-14, которая включает меньше аминокислот, чем первичный белок, но демонстрирует по меньшей мере один вид биологической активности соответствующего первичного белка). Обычно биологически активные фрагменты содержат домен или мотив, обладающей по меньшей мере одним типом биологической активности белка LIP01-LIP03. Биологически активный фрагмент белка, согласно настоящему изобретению, может представлять собой полипептид, который имеет длину, например, 10, 25, 50, 100 или более аминокислот. Кроме того, рекомбинантными способами могут быть получены другие биологически активные участки, в которых удалены другие области белка, и произведена их оценка по одному или более видам биологической активности, проявляемой нативной формой полипептида, согласно настоящему изобретению. Также особенностью настоящего изобретения являются фрагменты нуклеиновых кислот, кодирующие вышеупомянутые биологически активные фрагменты белка LIP01-LIP03. Слитые белки Белки, согласно настоящему изобретению, или их функциональные эквиваленты, например их биологически активные участки, могут быть функционально связаны с полипептидом, отличным от LIP01LIP03 (например, гетерологичными последовательностями аминокислот), с образованием слитых белков."Отличный от LIP01-LIP03 полипептид" означает полипептид, содержащий последовательность аминокислот, соответствующую белку, не являющемуся в значительной степени гомологичным белку LIIP01LIP03. Такой "отличный от LIP01-LIP03 полипептид" может быть получен из того же или из другого организма. В рамках слитого белка LIP01-LIP03, полипептид может соответствовать полностью или биологически активному фрагменту белка LIP01-LIP03. В предпочтительном варианте реализации слитый белок LIP01-LIP03 содержит по меньшей мере два биологически активных участка белка LIP01-LIP03. В рамках слитого белка, термин "функционально связанный" означает, что полипептид LIP01-LIP03 и полипептид, отличный от LIP01-LIP03, слиты в рамке считывания друг с другом. Полипептид, отличный отLIP01-LIP03, может быть слит с N-концом или С-концом полицептида LIP01-LIP03. Например, в одном из вариантов реализации, слитый белок представляет собой слитый белок GSTLIP01-LIP03, в котором последовательности LIP01-LIP03 слиты с С-концом последовательностей GST. Такие слитые белки могут способствовать очистке рекомбинантных LIP01-LIP03. В другом варианте реализации, слитый белок представляет собой белок LIP01-LIP03, содержащий гетерологичную сигнальную последовательность на N-конце. В некоторых клетках-хозяевах (например, клетках-хозяевах млекопитающих и дрожжей) экспрессия и/или секреция LIP01-LIP03 может быть повышена посредством использования гетерологичной сигнальной последовательности. В другом варианте примере секреторная последовательность gp67 белка оболочки бакуловируса может быть использована в качестве гетерологичной сигнальной последовательности (Current Protocolsin Molecular Biology, Ausubel и сотр., John WileySons, 1992). Другие примеры эукариотных гетерологичных сигнальных последовательностей включают секреторные последовательности меллитина и щелочной фосфатазы человека (Stratagene; La Jolla, California). В еще одном примере, применимые прокариотные гетерологичные сигнальные последовательности включают секреторный сигнал phoA (Sambrook и сотр. (упомянуто ранее) и секреторный сигнал белка А (Pharmacia Biotech; Piscataway, NewJersey). Сигнальная последовательность может использоваться для облегчения секреции и выделения белка и полипептида, согласно настоящему изобретению. Сигнальные последовательности обычно характери- 11015172 зуются ядром гидрофобных аминокислот, которые, в общем случае, отщеплены от зрелого белка в процессе секреции посредством одного или более отщеплений. Такие сигнальные пептиды содержат центры процессинга, которые дают возможность отщепления сигнальной последовательности от зрелых белков при прохождении ими секреторного пути. Сигнальная последовательность управляет секрецией белка,например, из эукариотного носителя, в котором трансформирован вектор экспрессии, и сигнальная последовательность отщепляется, после или одновременно. Затем белок может быть легко очищен от внеклеточной среды посредством известных способов. Альтернативным образом, сигнальная последовательность может быть связана с представляющим интерес белком при помощи последовательности, способствующей очистке, такой как GST домен. Таким образом, например, последовательность, кодирующая полипептид, может быть слита с маркерной последовательностью, такой как последовательность,кодирующая пептид, способствующий очистке слитого полипептида. В некоторых вариантах реализации данного аспекта настоящего изобретения маркерная последовательность представляет собой гексагистидиновый пептид, такой как метка, обеспеченная в векторе pQE (Quagen, Inc.), среди других, многие из которых имеются в продаже. Как описано в Gentz и сотр., Proc. Natl. Acad. Sci. USA 86:821-824 (1989),например, гексагистидин предложен для удобной очистки слитого белка. Метка HA представляет собой другой пептид, применимый для очистки, который соответствует эпитопу, полученному из белка гемагглютинина гриппа, который описан, например, у Wilson и сотр., Cell 37:767 (1984). Предпочтительно слитый белок LIP01-LIP03 согласно настоящему изобретению получают при помощи стандартных методов рекомбинантных ДНК. Например, фрагменты ДНК, кодирующие различные последовательности полипептидов, лигированы друг с другом в рамке считывания, в соответствии с общепринятыми способами, например, посредством "тупых" концов или ступенчатых концов для лигирования, обработка рестриктазой для получения подходящих концов, заполнение "липких" концов подходящим способом, обработка щелочной фосфатазой для того, чтобы избежать нежелательных соединений,и ферментативное лигирование. В другом варианте реализации, ген слияния может быть синтезирован общепринятыми способами, включая использование автоматических ДНК-синтезаторов. Альтернативным образом, ПЦР-амплификация фрагментов генов может быть осуществлена с использованием якорных праймеров, которые дают комплементарные "липкие" участки между двумя последовательными фрагментами генов, которые могут затем быть соединены и реамплифицированы с образованием последовательности гибридного гена (см., например, Current Protocols in Molecular Biology, eds. Ausubel и сотр.John WileySons: 1992). Кроме того, имеются в продаже многие векторы экспрессии, уже кодирующие участки слияния (например, полипептид GST). Нуклеиновая кислота, кодирующая LIIP01-LIP03, может быть клонирована в такой вектор экспрессии таким образом, что участок слияния лигирован в рамке считывания с белком LIP01-LIP03. Функциональные эквиваленты Термины "функциональные эквиваленты" и "функциональные варианты" применяются здесь взаимозаменяемо. Функциональные эквиваленты ДНК LIP01-LIP03 представляют собой фрагменты выделенной ДНК, которые кодируют полипептид, демонстрирующий особую функцию липолитического фермента LIP01-LIP03, согласно данному здесь определению. Функциональный эквивалент полипептидаLIP01-LIP03, согласно настоящему изобретению, представляет собой полипептид, демонстрирующий по меньшей мере одну функцию липолитического фермента Magnaporthe grisae, согласно данному здесь определению. Функциональные эквиваленты, следовательно, охватывают биологически активные фрагменты. Функциональные эквиваленты белка или полипептида могут содержать только консервативные замены одной или более аминокислот последовательностей SEQ ID NO: 5-14 или замены, инсерции или делеции заменимых аминокислот. Соответственно, заменимая аминокислота в остатке может быть изменена в SEQ ID NO: 5-14 без значительного изменения биологической функции. Например, аминокислотные остатки, которые консервативны в белках LIP01-LIP03, согласно настоящему изобретению, как предсказано, являются особенно не подлежащими изменению. Кроме того, аминокислоты, которые консервативны в белках LIP01-LIP03, согласно настоящему изобретению, и другом липолитическом ферменте, вероятно, не являются подлежащими изменению. Термин "консервативная замена" означает замену, при которой аминокислотный остаток заменяется аминокислотным остатком, имеющим похожую боковую цепь. Данные семейства известны из уровня техники и включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, и гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагины, глутамин, серии, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин,фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин,изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Функциональные эквиваленты нуклеиновых кислот типично могут содержать молчащие мутации или мутации, которые не изменяют биологической функции кодируемого полипептида. Соответственно,в настоящем изобретении предложены молекулы нуклеиновой кислоты, кодирующие белки LIP01-LIP03,- 12015172 содержащие изменения в аминокислотных остатках, которые не являются существенными для конкретной биологической активности. Такие белки LIP01-LIP03 отличаются по последовательности аминокислот от последовательностей SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6,однако еще сохраняют по меньшей мере один вид их биологической активности. В одном из вариантов реализации, выделенная молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую белок, где белок содержит в значительной степени гомологичную последовательность аминокислот, гомологичную последовательности аминокислот, представленной в SEQ ID NO: 5-14, по меньшей мере на 60%, предпочтительно на 65%, более предпочтительно на 70%, еще более предпочтительно на 75, 80, 85, 90, 95, 96, 97, 98, 99% или более. Например, руководство, касающееся того, как получить фенотипически молчащие последовательности аминокислот, предложено в Bowie, J.U. и сотр., Science 247:1306-1310 (1990) и в ссылках, цитируемых там. Как утверждают авторы, данные исследования показали, что белки, к удивлению авторов,являются толерантными к замещениям аминокислот. Далее, авторы указывают, какие изменения, вероятно, являются пермиссивными в определенных позициях белка. Выделенная молекула нуклеиновой кислоты, кодирующая белок LIP01-LIP03, гомологичный белку,согласно любой из SEQ ID NO: 5-7, SEQ ID NO: 8-9, SEQ ID NO: 10-14, соответственно, может быть получена посредством введения одного или более замещений нуклеотидов, добавлений или делеций в кодирующие последовательности нуклеотидов, согласно любой из соответствующих SEQ ID NO: 2-4, так,чтобы одно или более замещений, делеций или инсерций аминокислот было введено в кодируемый белок. Такие мутации могут быть введены посредством стандартных способов, таких как сайтспецифический мутагенез и опосредованный ПЦР мутагенез. Термин "функциональные эквиваленты" также охватывает ортологи белка LIP01-LIP03. Ортологии белка LIP01-LIP03 представляют собой белки, которые могут быть выделены из других штаммов или видов, и обладают похожей или одинаковой биологической активностью. Такие ортологи могут быть легко идентифицированы как содержащие последовательности аминокислот, в значительной степени гомологичные SEQ ID NO: 5-14. Согласно данному здесь определению, термин "в значительной степени гомологичные" относится к первой аминокислоте или последовательности нуклеотидов, которая содержит достаточное или минимальное количество идентичных или эквивалентных (например, с похожими боковыми цепями) аминокислот или нуклеотидов, по отношению ко второй аминокислоте или последовательности нуклеотидов,так, что первая и вторая аминокислоты или последовательности нуклеотидов имеют общий домен. Например, аминокислота или последовательности нуклеотидов, которые содержат общий домен, имеющий приблизительно 60%, предпочтительно 65%, более предпочтительно 70%, еще более предпочтительно 75, 80, 85, 90, 95, 96, 97, 98, 99% или более идентичности, определяются здесь как в значительной степени идентичные. Также, нуклеиновые кислоты, кодирующие другие представители семейства LIP01-LIP03, которые,следовательно, имеют последовательность нуклеотидов, отличающуюся от SEQ ID NO: 2-4, тоже находятся в рамках настоящего изобретения. Кроме того, нуклеиновые кислоты, кодирующие белки LIP01LIP03 из других видов, которые могут иметь последовательность нуклеозидов, отличающуюся от SEQ IDNO: 2-4, тоже находятся в рамках настоящего изобретения. Молекулы нуклеиновой кислоты, соответствующие вариантам (например, природные аллельные варианты) и гомологи ДНК LIP01-LIP03, согласно настоящему изобретению, могут быть выделены, на основе своей гомологичности по отношению к раскрытым здесь нуклеиновым кислотам LIP01-LIP03,при помощи использования раскрытых здесь кДНК или их подходящего фрагмента в качестве гибридизационного зонда, согласно стандартным способам гибридизации, предпочтительно, в очень строгих условиях. Дополнительно к встречающимся в природе аллельным вариантам последовательности LIP01LIP03, специалистам в данной области будет понятно, что посредством мутации в последовательности нуклеотидов LIP01-LIP03 могут быть внесены изменения, приводящие тем самым к изменениям в последовательности аминокислот белка LIP01-LIP03 без значительного изменения функции белка LIP01LIP03. В другом аспекте настоящего изобретения предложены улучшенные белки LIP01-LIP03. Улучшенные белки LIP01-LIP03 представляют собой белки, в которых улучшен по меньшей мере один вид биологической активности. Такие белки могут быть получены посредством статистического введения мутаций во всей кодирующей последовательности LIP01-LIP03 или в ее части, такого как насыщающий мутагенез, и полученные в результате мутации могут быть рекомбинантным способом экспрессированы, и исследованы на предмет их биологической активности. Например, в уровне техники предложены стандартные способы анализа для измерения ферментной активности липолитического фермента, и, следовательно, улучшенные белки легко могут быть выбраны. В предпочтительном варианте реализации белок LIP01-LIP03 имеет последовательность аминокислот, соответствующую SEQ ID NO: 5-7, SEQ ID NO: 8-9, SEQ ID NO: 10-14, соответственно. В другом варианте реализации, полипептид LIP01-LIP03 является в значительной степени гомологичным последо- 13015172 вательности аминокислот, соответствующей SEQ ID NO: 5-14, и сохраняет по меньшей мере один вид биологической активности полипептида, соответствующего SEQ ID NO:5-7, при этом отличаясь по последовательности аминокислот по причине природной изменчивости или по причине описанного выше мутагенеза. В другом предпочтительном варианте реализации, белок LIP01-LIP03 имеет последовательность аминокислот, кодируемую выделенным фрагментом нуклеиновой кислоты, способным к гибридизации с нуклеиновой кислотой, соответствующей, соответственно, одной из SEQ ID NO: 2-4, предпочтительно, в очень строгих условиях. Соответственно, белок LIP01-LIP03 предпочтительно представляет собой белок, содержащий последовательность аминокислот по меньшей мере на примерно 60, 65, 70, 75, 80, 85, 90, 95, 96, 97, 98, 99% или более гомологичную последовательности аминокислот, представленной в SEQ ID NO: 5-14, и еще сохраняющий по меньшей мере один вид биологической активности полипептида, соответствующегоSEQ ID NO: 5-14. Функциональные эквиваленты белка согласно настоящему изобретению также могут быть идентифицированы, например, посредством скрининга комбинаторных библиотек мутантов, например усеченных мутантов, белка согласно настоящему изобретению на предмет липолитической ферментной активности. В одном из вариантов реализации смешанную библиотеку вариантов создают посредством комбинаторного мутагенеза на уровне нуклеиновой кислоты. Смешанная библиотека вариантов может быть получена, например, путем ферментного лигирования смеси синтетических олигонуклеотиидов в генные последовательности таким образом, чтобы набор вырожденных потенциальных последовательностей белков мог быть экспрессируемым как в виде индивидуальных полипептидов, так и, альтернативным образом, в виде набора более больших слитых белков (например, фаговый дисплей). Существуют разнообразные способы, которые могут применяться для получения библиотек потенциальных вариантов полипептидов, согласно настоящему изобретению, из вырожденной последовательности олиигонуклеотидов. Способы получения вырожденных олигонуклеотидов известны из уровня техники (см., например,Narang (1983) Tetrahedron 39:3; Itakura и сотр. (1984) Annu. Rev. Biochem. 53:323; Itakura и сотр. (1984)Science 198:1056; Ike и сотр. (1983) Nucleic Acid Rees. 11:477. Дополнительно, библиотеки фрагментов кодирующей последовательности полипептида согласно настоящему изобретению могут быть использованы для генерации смешанной популяции полипептидов для скрининга последующего выбора вариантов. Например, библиотека фрагментов кодирующей последовательности может быть получена путем обработки двухцепочечного ПЦР-фрагмента представляющей интерес кодирующей последовательности нуклеазой в условиях, при которых одноцепочечный разрыв имеет место только примерно один раз на молекулу, денатурации двухцепочечной ДНК, ренатурации ДНК с образование двухцепочечной ДНК, которая может включать смысловые/антисмысловые пары различных продуктов одноцепочечного разрыва, удаления одноцепочечных участков из преобразованных дуплексов путем обработки S1 нуклеазой, и лигированиия полученного фрагмента библиотеки в вектор экспрессии. Посредством данного способа может быть получена библиотека экспрессии, кодирующая имеющие различные размеры N-концевые и внутренние фрагменты представляющего интерес белка. Из уровня техники известно несколько способов скрининга генных продуктов комбинаторных библиотек, полученных посредством точечных мутаций усечения, а также для скрининга библиотек кДНК,обладающих выбранным свойством. Наиболее широко применяемые способы скрининга больших библиотек генов, поддающихся анализу с высокой пропускной способностью, обычно включают клонирование библиотеки генов в способные к репликации векторы, трансформации подходящих клеток посредством полученной библиотеки векторов и экспрессии комбинаторных генов в условиях, при которых обнаружение требуемой активности способствует выделению вектора, кодирующего ген, продукт которого обнаружен. Ступенчатый согласованный мутагенез (recursive ensemble mutagenesis (REM, способ, повышающий частоту функциональных мутаций в библиотеках, может применяться в сочетании со скрининг-анализом для идентификации вариантов белка, согласно настоящему изобретению (Arkin и Yourvan(1992) Proc. Natl. Acad. Sci. USA 89:7811-7815; Delgrave и сотр. (1993) Protein Engineering 6(3): 327-331). Дополнительно к последовательности генов LIP01-LIP03, представленной в SEQ ID NO: 2-4, соответственно, для специалиста в данной области будет очевидно, что полиморфизмы последовательности ДНК могут существовать внутри данной популяции, что может приводить к изменениям в последовательности аминокислот белка LIP01-LIP03. Такие генетические полиморфизмы могут существовать в клетках из разных популяций или внутри популяции, что обусловлено естественной аллельной изменчивостью. Аллельные варианты могут также включать функциональные эквиваленты. Фрагменты полинуклеотида согласно настоящему изобретению могут также содержать полинуклеотиды, не кодирующие функциональные полипептиды. Такие полинуклеотиды могут исполнять функцию зондов или праймеров для ПЦР. Нуклеиновые кислоты согласно настоящему изобретению вне зависимости от того, кодируют они функциональные или нефункциональные полипептиды, могут быть использованы в качестве гибридизационных зондов или праймеров полимеразной цепной реакции (ПЦР). Способы применения молекул- 14015172 нуклеиновых кислот согласно настоящему изобретению, не кодирующих полипептид, обладающий активностью LIP01-LIP03, включают среди других (1) выделение гена, кодирующего белок LIP01-LIP03,или его аллельных вариантов, из библиотеки кДНК, например, из организмов, отличных от Magnaporthegrisae; (2) гибридизация in situ (например, FISH) с распластанными метафазными хромосомами для установления точного местоположения гена LIP01-LIP03 в хромосоме, как описано в Verma и сотр., HumanChromosomes: a Manual of Basic Techniques, Pergamon Press, New York (1988); (3) нозерн-блоттинг для установления экспрессии иРНК LIP01-LIP03 в специфических тканях и/или клетках и (4) зонды и праймеры, которые могут быть использованы в качестве средства диагностики при анализе присутствия нуклеиновой кислоты, способной гибридизоваться с зондом LIP01-LIP03 в данном биологическом (например, в ткани) образце. Кроме того, настоящее изобретение охватывает способ получения функционального эквивалента гена LIP01-LIP03. Такой способ влечет за собой получение меченого зонда, который включает выделенную нуклеиновую кислоту, кодирующую всю белковую последовательность, согласно SEQ ID NO: 5-14 или варианту любой из них, или ее участок; скрининг библиотеки фрагментов нуклеиновых кислот при помощи меченого зонда в условиях, дающих возможность для гибридизации зонда с фрагментами нуклеиновых кислот библиотеки с образованием дуплексов нуклеиновых кислот, и получение первичной последовательности генов из фрагментов нуклеиновых кислот любого меченого дуплекса для получения гена, относящегося к генам LIP01-LIP03. В одном из вариантов реализации, нуклеиновая кислота LIP 01-LIP03, согласно настоящему изобретению, приблизительно на 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99% или более гомологична последовательности нуклеиновых кислот, представленной в любой из SEQ ID NO: 2-4, или ее комплементу. Клетки-хозяева В другом варианте реализации настоящего изобретения предложены клетки, например трансформированные клетки-хозяева или рекомбинантные клетки-хозяева, содержащие нуклеиновую кислоту,охватываемую настоящим изобретением. "Трансформированная клетка" или "рекомбинантная клетка" представляют собой клетку, внутрь которой (или внутрь предка которой) посредством метода рекомбинантных ДНК была введена нуклеиновая кислота согласно настоящему изобретению. Настоящее изобретение включает как прокариотные, так и эукариотные клетки, например бактерии, грибы, дрожжи и т.п.,особенно предпочтительными являются клетки мицелиальных грибов, в частности Magnaporthe grisae или видов Aspergillus, таких как Aspergillus niger или oryzae. Могут быть выбраны такие клетки-хозяева, которые модулируют экспрессию введенных последовательностей, или модифицирует и процессирует генный продукт специфическим, требуемым образом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут способствовать оптимальному функционированию белка. Различные клетки-хозяева обладают характерными и специфическими механизмами посттрансляционного процессинга и модификации белков и генных продуктов. Подходящие линии клеток или системы носителей, хорошо известные специалистам в данной области молекулярной биологии и/или микробиологии, могут быть выбраны таким образом, чтобы иметь уверенность в требуемой и правильной модификации и процессинге экспрессированного чужеродного белка. С этой целью могут применяться эукариотные клетки-хозяева, обладающие клеточными механизмами для должного процессинга первичного транскрипта, гликозилирования и фосфорилирования генного продукта. Такие клетки-хозяева известны из уровня техники. Клетки-хозяева также включают, но не ограничиваются ими, линии клеток млекопитающих, такие как СНО, VERO, BHK, HeLa, COS, MDCK, 293, 3 Т 3, WI38 и линии клеток хороидного сплетения. Если необходимо, линии стабильно трансфицированных клеток могут производить полипептиды,согласно настоящему изобретению. Доступен ряд векторов, пригодных для стабильной трансфекции клеток млекопитающих, способы конструирования таких линий клеток также общеизвестны, например, вAusbel и сотр. (упомянуто ранее). Применение липолитического фермента LIP01-LIP03 в промышленном производстве Неожиданно действие липолитического фермента согласно настоящему изобретению не ограничивается гидролизом лишь одного специфического субстрата, но он может проявлять разные типы липолитической активности, представляющие собой фосфолипазную, липазную и галактолипазную активность. Липолитический фермент согласно настоящему изобретению может проявлять эти типы активности одновременно или может обладать узкой специфичностью с одной единственной активностью при малой или полностью отсутствующей другой активности, либо он может обладать более широкой специфичностью с одной доминирующей активностью и меньшими другими активностями, в зависимости от состава теста, времени реакции, рН, температуры, содержания воды. Благодаря данному разнообразию липолитический фермент согласно настоящему изобретению может применяться для множества промышленных применений, включая производство дигалактозилмоноглицерида из содержащего дигалактозилдиглицерид источника, или модификацию фосфолипидных эмульгаторов. Примером фосфолипидного эмульгатора является лецитин, представляющий собой смесь как полярных, так и нейтральных липидов, в которой содержание полярных липидов составляет по- 15015172 меньшей мере 60%. Фосфолипидные эмульгаторы имеют много пищевых и непищевых применений, например, яичный лецитин применяется в качестве эмульгатора, например, в молочных продуктах, особенно в майонезе, заправках, выпечке и т.д., соевый лецитин, например, применяется в качестве эмульгатора в (низкокалорийных) соусах, хлебе, маргарине, косметических средствах и т.д., другие лецитины применяются, например, в шоколаде, питании для телят. Модификация фосфолипидных эмульгаторов посредством липолитического фермента согласно настоящему изобретению может вызывать повышенное эмульгирование смеси масло/вода. Модификация фосфолипидных эмульгаторов посредством липолитического фермента, согласно настоящему изобретению, может увеличивать стабильность эмульсий в более широком диапазоне рН и/или температур, из-за добавления модифицированных фосфолипидных эмульгаторов. Модификация фосфолипидных эмульгаторов посредством липолитического фермента,согласно настоящему изобретению, может увеличивать стабильность эмульсий в присутствии ионов Са 2+ или Mg2+, из-за добавления модифицированных фосфолипидных эмульгаторов. Другим примером промышленного применения липолитического фермента, согласно настоящему изобретению, является то, что он может использоваться для рафинирования гидратацией растительных масел при переработки данных масел. В типичном способе рафинирования масла гидратацией из растительных масел, например из соевого масла, рапсового масла (масла канолы), льняного масла, подсолнечного масла, удаляют лецитины для повышения помимо прочего стабильности растительного масла, путем промывания масляной фазы водой, при этом смешивание воды и масла в условиях высоких усилий сдвига переводит основную массу лецитинов в водную фазу, которую затем удаляют в сепараторе. С этой так называемой водной рафинирующей фазой легко удаляются только быстро гидратируемые фосфолипиды, например, фосфатидилхолин, фосфатидилинозитол и фосфатидилэтаноламин. Негидратируемые фосфолипиды/фосфатиды, главным образом, фосфолипиды, содержащие до 50% солей магния и/или кальция, не могут быть легко удалены на этапе водного рафинирования гидратацией. Воздействие на негидратируемые фосфолипиды/фосфатиды липолитического фермента, согласно настоящему изобретению, делает данные фосфолипиды более растворимыми в воде и, следовательно, более легко экстрагируемыми в водную рафинирующую фазу. Другим примером промышленного применения липолитического фермента, согласно настоящему изобретению, является удаление осадка, который появляется при осахаривании (при помощи -амилазы и гликоамилазы) клейковины пшеницы или пшеничного крахмала для получения сиропов глюкозы. Удаление осадка значительно ускоряет последующую фильтрацию получающихся сиропов глюкозы. Еще одним примером промышленного применения липолитического фермента, согласно настоящему изобретению, в пищевой промышленности, является его использование в хлебопекарной промышленности для улучшения качества теста или печеных изделий. Удивительно, что липаза, согласно настоящему изобретению, при использовании в тесте (а также в других упомянутых промышленных процессах) демонстрирует in situ, по меньшей мере одно из следующих свойств: относительно низкая активность по отношению к неполярным липидам,относительно высокая активность по отношению к полярным диациллипидам, по меньшей мере, по отношению к диацилгалактолипидам,относительно низкая активность по отношению к полярным моноацил-производным. Например, фермент согласно настоящему изобретению может демонстрировать in situ относительно низкую лизофосфолипазную активность и относительно низкую липазную активность. Все эти неожиданные свойства, как обнаружено, являются чрезвычайно предпочтительными при применении в качестве заместителя химических эмульгаторов в тесте. Можно выделить несколько типов фосфолипазной активности при гидролизе сложноэфирной(ных) связи(ей), связывающих жирные ацильные группы с глицериновой главной цепью. Фосфолипаза А 1 (ЕС 3.1.1.32) и А 2 (ЕС 3.1.1.4) катализируют деацилирование жирной ацильной группы в sn-1 и sn-2 положениях, соответственно, от диацилглицерофосфолипида с образованием лизофосфолипида. Это желательная активность для заменителя эмульгатора. Лизофосфолипаза (ЕС 3.1.1.5 - называемая также фосфолипазой В согласно Комитету по номенклатуре международного союза биохимии и молекулярной биологии (Nomenclature Committee of the International Union of Biochemistry and Molecular Biology) (Enzyme Nomenclature, Academic Press, New York,1992 катализирует гидролиз оставшейся жирной ацильной группы лизофосфолипида. О фосфолипазе В, полученной из Penicillium notatum, сообщалось (Saito и сотр., 1991, Methods in Enzymology 197:446456), что она катализирует деацилирование обеих жирных кислот из диацилглицерофосфолипида и обладает присущей ей лизофосфолипазной активностью. Для заменителя эмульгатора лизофосфолипазная активность менее желательна, поскольку она будет приводить к разрушению сочетания полярной головы и неполярного хвоста, делая получающийся продукт неспособным влиять на поверхностные свойства. Неожиданно было показано, что липаза согласно настоящему изобретению проявляет низкую лизофосфолипазную активность в тесте. Пшеничная мука содержит приблизительно 2,2-2,9% липидов. Липиды муки можно разделить на крахмальные липиды (0,8-0,9%) и некрахмальные липиды (1,4-2,0%). В то время как крахмальные липи- 16015172 ды состоят, главным образом, из полярных лизофосфолипидов, некрахмальные липиды состоят из примерно 40% неполярных триглицеридов и 40% полярных фосфо- и гликолипидов. Для оптимизации фракции липидов муки липаза, согласно настоящему изобретению, способна к гидролизу полярных липидов, являющихся фосфолипидами и гликолипидами, более специально, галактолпидами, in situ в тесте при добавлении в него липазы, согласно настоящему изобретению. В WO 04/104193 раскрывается применение фосфолипазы С из Magnaporthe grisae в хлебопекарных целях. Однако активность фосфолипазы не является желательной для фермента, применяемого в качестве заменителя химических эмульгаторов, поскольку он не дает образования достаточно поверхностноактивных соединений. Кроме того, фосфолипаза С, раскрытая в WO 04/104193, не гомологична SEQ IDNO: 3, 4 или 5. В WO 98/45453 раскрывается обладающий липазной активностью полипептид, который можно получить из Aspergillius tubigensis и который также проявляет гидролитическую активность по отношению к дигалактозилдиглицериду. Данный фермент, однако, страдает относительно низкой специфической активностью по отношению к галактозиздиглицеридам и относительно высокой активностью по отношению к триглицеридам in situ в хлебе (пример 10), что делает данный фермент непригодным для применения в качестве полного заменителя химических эмульгаторов. Пекарные ферменты могут применяться для производства печеных изделий. Термин "печеные изделия" включает здесь хлебные изделия, такие как формовой хлеб, хлеб в виде батонов, французские булки, а также булочки, пирожные, пироги, кексы, дрожжевое тесто, пончики и т.п. Липолитический фермент согласно настоящему изобретению можно, например, применять в печеных изделиях. Такие печеные изделия, как хлеб, изготавливают из теста, которое обычно делают из муки(пшеничной), воды и, необязательно, соли в качестве основных ингредиентов. В зависимости от печеных изделий другие добавленные ингредиенты могут представлять собой сахара, ароматизаторы и т.д. Для дрожжевых изделий главным образом применяются хлебопекарные дрожжи наряду с системами химических разрыхлителей, такими как комбинация кислоты (генерирующее соединение) и бикарбонат. Дрожжи, ферменты и химические добавки обычно добавляют в тесто по отдельности. Ферменты могут быть добавлены в сухом, например, гранулированном, виде или в жидком виде. Химические добавки в большинстве случаев добавляют в виде порошка. Кроме того, композиции технологических добавок, специально созданные для хлебопекарного применения, могут состоять из специальной смеси химических добавок и фермента. Получение теста из ингредиентов и описанных выше технологических добавок хорошо известно из уровня техники и включает смешивание указанных ингредиентов и технологических добавок, а также один или несколько этапов формования и ферментации. Получение печеных изделий из такого теста также хорошо известно из уровня техники и может включать формование и придание формы и дополнительную ферментацию теста с последующим выпеканием при требуемых температурах и времени выпекания. Настоящее изобретение направлено по меньшей мере на одну, если не на все вышеупомянутые задачи. Настоящее изобретение также относится к применению липолитического фермента, согласно настоящему изобретению, в ряде промышленных способов производства. Несмотря на большой опыт,имеющийся в таких производствах, липолитический фермент, согласно настоящему изобретению, имеет ряд значительных преимуществ над применяемыми в настоящее время ферментами. В зависимости от конкретного применения, эти преимущества могут включать такие аспекты, как более низкие издержки производства, более высокую специфичность к субстрату, меньшую аллергенность, менее нежелательное побочное действие, более высокий выход при производстве подходящими микроорганизмами, более подходящие интервалы значений рН и температуры, лучший вкус готового изделия, а также качестве пищи и аспекты, связанные с кошерностью. Настоящее изобретение также относится к способам получения теста или печеного изделия, включающим введение в тесто эффективного количества липолитического фермента, согласно настоящему изобретению, который улучшает одно или более свойств теста или печеного изделия, полученного из теста, по сравнению с тестом или печеным изделием, в которые не добавлен данный полипептид. Фраза "введение в тесто" определяется здесь как добавление липолитического фермента, согласно настоящему изобретению, в тесто, в любой ингредиент, из которого делают данное тесто, и/или в любую смесь ингредиентов теста, из которых делают данное тесто. Другими словами, липолитический фермент согласно настоящему изобретению может быть добавлен на любом этапе получения теста и может быть добавлен за один, два или более этапов. Липолитический фермент согласно настоящему изобретению добавляют в ингредиенты теста, которое замешивают и выпекают для получения печеного изделия посредством способов, хорошо известных из уровня техники. См., например, патент США 4567046, ЕРА-426211, JP-A-60-7829, JP-A-62-111629 и JP-A-63-258528. Термин "эффективное количество" определяется здесь как количество липолитического фермента,согласно настоящему изобретению, достаточное для оказания измеримого действия по меньшей мере на одно представляющее интерес свойство теста и/или печеного изделия.- 17015172 Термин "улучшенное свойство" определяется здесь как любое свойство теста и/или полученного из него изделия, особенно печеного изделия, которое улучшено действием липолитического фермента, согласно настоящему изобретению, по сравнению с тестом или изделием, в которые не добавлен липолитический фермент, согласно настоящему изобретению. Улучшенное свойство может включать, но не ограничиваясь им, повышенную прочность теста, повышенную эластичность теста, повышенную стабильность теста, пониженную липкость теста, улучшенную пригодность теста для механической переработки, повышенный объем буханки печеного изделия, улучшенный аромат печеного изделия, улучшенную структуру мякиша печеного изделия, повышенную мягкость мякиша печеного изделия и/или улучшенную способность печеного изделия не черстветь. Улучшенное свойство может быть определено путем сравнения теста и/или печеного изделия, полученного с и без добавления полипептида, согласно настоящему изобретению, в соответствии со способами, согласно настоящему изобретению, описанными ниже в примерах. Органолептические свойства могут быть оценены при помощи процедур, признанных в пекарной промышленности, и могут включать,например, привлечение группы обученных дегустаторов. Термин "повышенная прочность теста" определяется здесь как свойство теста, которое обладает большей эластичностью и/или требует больше труда для помещения его в форму и формования. Термин "повышенная эластичность теста" определяется здесь как свойство теста, которое имеет более высокую склонность к восстановлению своей первоначальной формы после того, как его подвергли определенной физической деформации. Термин "пониженная липкость теста" определен здесь как свойство теста, которое имеет меньшую склонность прилипать к поверхностям, например, механического оборудования для получения теста, и которое либо оценивают эмпирически при помощи специалиста по пробной выпечке, либо измеряют при помощи анализатора текстуры (например, ТАХ 2) способом, известным из уровня техники. Термин "повышенная растяжимость теста" определяется здесь как свойство теста, которое может быть подвергнуто более большей деформации или растяжению без разрыва. Термин "улучшенная пригодность теста для механической переработки" определяется здесь как свойство теста, которое является в целом менее липким, и/или более прочным, и/или более эластичным. Термин "повышенный объем буханки печеного изделия" определяется здесь как объем данной буханки хлеба, определенный автоматическим анализатором объема хлеба (например, BVM-3, Тех Vol Instruments АВ, Викен, Швеция) при помощи ультразвукового или лазерного детектирования, согласно способам, известным из уровня техники. Термин "уменьшенное образование пузырей в печеном изделии" определяется здесь как определенное визуально уменьшение образования пузырей на корке печеного изделия. Термин "улучшенная структура мякиша печеного изделия" определяется здесь как свойство печеного изделия, имеющего более мелкие пустоты и/или более тонкие стенки пустот в мякише, и/или более равномерное и/или более гомогенное распределение пустот в мякише, и его оценивают или визуально пекарь, или определяют при помощи цифрового анализа изображения, способами, известными из уровня техники (например, C-cell, Calibre Control International Ltd, Аплтон, Уоррингтон, Великобритания). Термин "улучшенная мягкость печеного изделия" определяется здесь как свойство печеного изделия, которое более легко сжимать, и его оценивают либо эмпирически при помощи специалиста по пробной выпечке, либо измеряют при помощи анализатора текстуры (например, ТАХ 2) способом, известным из уровня техники. Термин "улучшенный аромат печеного изделия" оценивают при помощи группы обученных дегустаторов. Термин "улучшенная способность печеного изделия не черстветь" определен здесь как свойство печеного изделия, которое имен уменьшенную скорость ухудшения параметров качества, например мягкости и/или эластичности, при хранении. Термин "тесто" определен здесь как смесь муки и других ингредиентов, достаточно прочную для замешивания или раскатывания. Тесто может быть свежим, замороженным, подготовленным или предварительно выпеченным. Получение замороженного теста описана у Kulp и Lorenz в Refrigerated Doughsand Batters. Термин "печеное изделие" определен здесь как любое изделие, полученное из теста, или мягкое,или хрустящее. Примерами печеных изделий, или белых, светлых, или черных, которые могут быть предпочтительно получены при помощи настоящего изобретения, являются хлеб (в частности, белый, из непросеянной муки или ржаной хлеб), обычно в виде буханок или батонов, хлеб типа французского багета, макароны, лапша (вареная или (при постоянном помешивании) обжаренная), лаваш, тортилья, тако,пирожные, блины, сухое печенье, домашнее печенье, пончики, рогалики, сухари, паровой хлеб, а также сушки и т.п. Липолитический фермент согласно настоящему изобретению и/или дополнительные ферменты, которые используются в способах согласно настоящему изобретению могут быть в любом виде подходящем для данного применения, например в виде сухого порошка, агломерированного порошка или гранулята, в частности непылящего гранулята, жидкости, в частности стабилизированной жидкости, или за- 18015172 щищенного фермента, как описано в WO 01/11974 и WO 02/26044. Грануляты и агломерированные порошки могут быть получены стандартными способами, например распылением липолитического фермента, согласно настоящему изобретению на носитель в грануляторе с псевдоожиженным слоем. Носитель может состоять из основы в виде частиц, имеющих подходящие размеры. Носитель может быть растворимым или нерастворимым, например солью (такой как NaCl или сульфат натрия), сахаром (таким как сахароза или лактоза), сахарным спиртом (таким как сорбит), крахмалом, рисом, кукурузной крупой или соей. Липолитический фермент согласно настоящему изобретению и/или дополнительные ферменты могут содержаться в составах медленного высвобождения. Способы получения составов медленного высвобождения хорошо известны из уровня техники. Добавление приемлемых пищевых стабилизаторов,таких как сахар, сахарный спирт или другой полиол, и/или молочная кислота или другая органическая кислота, согласно общепризнанным способам может, например, стабилизировать жидкие ферментные составы. Липолитический фермент согласно настоящему изобретению может также быть введен в содержащие дрожжи композиции, такие как те, что раскрыты в ЕР-А-0619947, ЕР-А-0659344 и WO 02/49441. Для включения в мучные премиксы предпочтительно, чтобы полипептид, согласно настоящему изобретению, был в виде сухого продукта, например непылящего гранулята, тогда как для включения вместе с жидкостью он преимущественно находится в жидком виде. В тесто могут быть также введены один или более дополнительных ферментов. Дополнительный фермент может иметь любое происхождение, включая млекопитающих и растения, предпочтительно микробное (бактериальное, дрожжевое или грибное) происхождение, и может быть получен при помощи технологий, общепринятых в уровне техники. В предпочтительном варианте реализации дополнительный фермент может представлять собой амилазу - такую как альфа-амилаза (применимую для того, чтобы сделать сахара ферментируемыми дрожжами и для замедления черствения), бета-амилаза, мальтогенная амилаза или немальтогенная амилаза - циклодекстринглюканотрансферазу, пептидазу, в частности эксопептидазу (применимую для улучшения аромата), трансглутаминазу, липазу (применимую для модификации присутствующих в тесте липидов, или компонентов теста для смягчения теста), галактолипазу, фосфолипазу, целлюлазу, хемицеллюлазу, в частности пентозаназу, такую как ксиланаза (применимая для частичного гидролиза пентозанов, более конкретно, арабиноксилана, что повышает растяжимость теста), протеазу (применимую для размягчения клейковины, в частности, при использовании муки из твердых сортов пшеницы), протеин дисульфид изомеразу, как раскрыто в WO 95/00636, гликозилтрансферазу, пероксидазу (применимую для улучшения консистенции теста), лакказу или оксидазу, например, глюкозооксидазу, гексозооксидазу,альдозооксидазу, пиранозооксидазу, липоксигеназу или оксидазу L-аминокислот (применимую для улучшения консистенции теста). При добавлении одного или более активных дополнительных ферментов в способы, согласно настоящему изобретению, эти активные вещества могут быть добавлены отдельно или вместе с полипептидом, согласно настоящему изобретению, необязательно, в виде компонента(ов) состава - улучшителя хлеба или теста. Другие ферментные активные вещества могут представлять собой любые ферменты,описанные выше, и могут быть дозированы в соответствии с общепринятыми пекарными технологиями. Настоящее изобретение также относится к способам получения печеного изделия, содержащего тесто, полученное способом, согласно настоящему изобретению, для получения печеного изделия. Выпекание теста для получения печеного изделия может быть осуществлено при помощи способов, хорошо известных из уровня техники. Настоящее изобретение относится также к тестам и печеным изделиям, соответственно, полученным при помощи способов, согласно настоящему изобретению. Настоящее изобретение относится также к премиксу, например, в виде мучной композиции, для теста и/или печеных изделий, полученных из теста, в котором премикс содержит полипептид, согласно настоящему изобретению. Термин "премикс" определяется здесь так, чтобы его понимали в обычном значении, т.е. как смесь пекарных средств, обычно включающую муку, которая может применяться не только на промышленных хлебозаводах/оборудовании, но также и в булочных-пекарнях. Премикс может быть получен путем смешивания полпептида или композиции - улучшителя хлеба и/или теста, согласно настоящему изобретению, содержащего полипептид с подходящим носителем, таким как мука, крахмал,сахар или соль. Премикс может содержать другие добавки - улучшители теста и/или хлеба, например любые добавки, включающие упомянутые выше ферменты. Настоящее изобретение дополнительно относится к пекарным добавкам в виде гранулята или агломерированного порошка, который содержит полипептид, согласно настоящему изобретению. Пекарная добавка предпочтительно имеет узкое распределение частиц по размерам, где более 95 мас.% частиц попадают в интервал от 25 до 500 мкм. При изготовлении теста и хлеба настоящее изобретение может применяться в сочетании с технологическими добавками, согласно данному выше определению, такими как химические технологические добавки типа окислителей (например, аскорбиновая кислота), восстановительных средств (например, Lцистеин) и/или эмульгаторов (например, DATEM, SSL и CSL) и/или ферментных технологических доба- 19015172 вок, таких как оксидоредуктазы (например, глюкозооксидаза), ферменты, модифицирующие полисахариды (например, -амилаза, хемицеллюлаза, разветвляющие ферменты и т.д.). Вышеупомянутые способы промышленного применения липолитического фермента, согласно настоящему изобретению, включают лишь несколько примеров, и этот список не является ограничительным. Липолитический фермент LIP01-LIP03 обычно может быть произведен микроорганизмами. В вышеупомянутых способах преимущественным является применение липолитического фермента, полученного методами рекомбинантных ДНК. Такие рекомбинантные ферменты имеют ряд преимуществ над их очищенными традиционными способами аналогами. Рекомбинантные ферменты могут производиться при низких затратах, с высоким выходом, не содержат загрязнений, таких как бактерии или вирусы, а также не содержат токсинов бактерий и не загрязнены другими ферментными активными веществами. Ниже изобретение проиллюстрировано следующими не ограничивающими его примерами. Примеры Пример 1. Ферментация Aspergillius niger. Липолитические ферменты, кодируемые последовательностями нуклеотидов SEQ ID NO: 2, SEQ IDNO: 3 и SEQ ID NO: 4, как предложено здесь, были получены путем создания экспрессирующих плазмид, содержащих последовательность ДНК, трансформирования штамма A. niger посредством такой плазмиды и выращивания штаммов A. niger следующим путем. Свежие споры (106-107 ) штаммов A. niger были инокулированы в 20 мл CSL-среды (колба на 100 мл, с отбойниками) и их выращивали в течение 20-24 ч при 34 С и скорости перемешивания 174 об/мин. После инокуляции 5-10 мл прекультуры CSL в 100 мл среды CSM (колба на 500 мл, с отбойниками),штаммы ферментировали при 34 С и скорости перемешивания 170 об/мин в течение 3-5 дней. Не содержащие клеток надосадочные жидкости получили центрифугированием в 50 мл пробиркахGreiner (30 мин, 5000 об/мин). Надосадочные жидкости были префильтрованы через микроволоконный стеклянный фильтр GF/A Whatman (150 мм AE) для удаления крупных частиц, довели рН до 5 при помощи 4 N КОН (если необходимо) и подвергли стерилизующему фильтрованию на вакуум-фильтре 0,2 мкм (установленного в горловину флакона) для удаления грибного материала. Надосадочную жидкость хранили при 4 С (или -20 С). Среда CSL состояла из (даны количества на литр): 100 г сухого остатка кукурузного экстракта(Roquette), 1 г NaH2PO4 Н 2 О, 0,5 г MgSO47H2O, 10 г (глюкоза)H2O и 0,25 г базильдона (антивспениватель). Ингредиенты растворили в деминерализованной воде и довели рН до 5,8 при помощи NaOH илиH2SO4; в колбы на 100 мл с отбойной перегородкой и противопенным поплавком загрузили по 20 мл ферментационного бульона и стерилизовали в течение 20 мин при 120 С, после чего в каждую колбу,после охлаждения до комнатной температуры, загрузили по 200 мкл раствора, содержащего 5000 МЕ/мл пенициллина и 5 мг/мл стрептомицина. Среда CSM состояла из (даны количества на литр): 150 г (мальтоза)H2O, 60 г Soyton (пептон), 1 гNaH2PO4H2O, 0,5 г MgSO47H2O, 0,08 г Твин-80, 0,02 г базильдона (антивспениватель), 20 г MES, 1 г Lаргинина. Ингредиенты растворили в деминерализованной воде и довели рН до 6,2 при помощи NaOH или H2SO4; в колбы на 500 мл с отбойной перегородкой и противопенным поплавком загрузили по 100 мл ферментационного бульона и стерилизовали в течение 20 мин при 120 С, после чего в каждую колбу,после охлаждения до комнатной температуры, загрузили по 1 мл раствора, содержащего 5000 МЕ/мл пенициллина и 5 мг/мл стрептомицина. Пример 2. Очистка липолитического фермента согласно настоящему изобретению. Этап 1 - получение ультрафильтратов. Надосадочные жидкости культур, полученные согласно примеру 1, подвергли ультрафильтрации для удаления низкомолекулярных примесей, способных мешать определению ферментативной активности и пробной выпечке. Ультрафильтрация 30 мл надосадочной жидкости проводили на системе Millipore Labscale TFF, снабженной фильтром с пределом пропускания 10 кДа. В зависимости от цвета образцы промывали 3-5 раз объемами по 40 мл холодного фосфатного буфера (100 мМ) с рН 6,0, содержащего 0,5 мМ CaCl2. Конечный объем раствора фермента составлял 30 мл,и далее он обозначен здесь термином "ультрафильтрат". Общее содержание белка в образцах определяли методом Брэдфорда (Protein Protocols Handbook,2nd edition, Edited by J.M. Walker, Humana Press, Totowa 2002, стр. 15-21). Этап 2. Определение концентрации липолитического фермента посредством A280 u HPSEC. Концентрацию липолитического фермента в ультрафильтрате рассчитывали по экстинкции при длине волны 280 нм (А 280), приписываемой липолитичекому ферменту, и рассчитывали молекулярный коэффициент экстинкции липолитического фермента. Измерения А 280 проводили при помощи спектрофотометра Uvikon XL Secomam (Beun de Ronde, Абкауде, Нидерланды). Молекулярный коэффициент экстинкции фермента может быть рассчитан по числу остатков тирозина, триптофана и цистеина на молекулу фермента (S.C. Gill и Р.Н. von Hippel, Anal. Biochem. 182, 319326 (1989. Молекулярные коэффициенты экстинкции данных аминокислот составляют 1280, 5690 и 120- 20015172 М-1 см-1, соответственно. Число остатков тирозина, триптофана и цистеина в липолитическом ферменте,согласно настоящему изобретению, может быть выведено из белковых последовательностей, выбранных из группы, в которую входят SEQ ID NO: 5-14. Расчетные значения коэффициентов экстинкции представлены в табл. 1. Таблица 1 Экстинкция ультрафильтрата при 280 нм (А 280), приписываемая липолитическому ферменту, зависит от чистоты образца фермента. Чистоту определяли при помощи HPSEC (высокоэффективной гельпроникающей хроматографии) на колонке TSK SW-XL (3007,8 мм; интервал значений Mw 10-300 кДа). Буферный раствор для элюирования, состоящий из 25 мМ буферного раствора фосфата натрия с рН 6,0 применялся при скорости потока 1 мл/мин. Вводили образцы объемом 5-10 мкл. Измеряли поглощение при длине волны 280 нм. А 280 ультрафильтрата, приписываемую липолитическому ферменту, получали из соотношения площади пиков - пика хроматограммы, соответствующего липолитическому ферменту, и общей площади пиков поглощения при длине волны 280 нм. Затем рассчитывали концентрацию липолитического фермента в ультрафильтрате, путем умножения значения А 280 ультрафильтрата на вышеописанное соотношение и деления на рассчитанный коэффициент экстинкции (1 мг/мл раствора - табл. 1, самая правая колонка) липолитического фермента. Пример 3. Измерение активности. Ультрафильтраты, полученные в примере 2, могут быть подвергнуты следующим измерениям активности фермента для установления специфичности липолитического фермента: липазная; фосфолипазная A1 или А 2; лизофосфолипазная; галактолипазная активность. Липазную активность определяли спектрофотометически посредством использования хромогенного субстрата п-нитрофенилпальмитата (pNPP). В этом анализе хромогенный субстрат п-нитрофенилпальмитат (pNPP) растворяли в 2-пропаноле и суспендировали в фосфатном буферном растворе с рН 7,4 в присутствие 0,1% гуммиарабика и 0,25% дезоксихолата натрия. Липазу выдерживали в растворе данного субстрата при 37 С и измеряли образовавшийся п-нитрофенил через 2,6 мин при 405 нм. Данный анализ также применим при разных уровнях рН для того, чтобы определить влияние рН на липазу. Следует понимать, что для разных уровней рН могут потребоваться разные буферные растворы или разные поверхностно-активные вещества для эмульгирования субстрата. Например, при рН 4 применяют 100 мМ ацетатный буферный раствор с 1,0% Тритона Х-100. Одна липазная единица определяется как количество фермента, которое вызывает выделение 1 мкмоль п-нитрофенола в минуту при данных условиях реакции. Следует понимать, что использование стандартных калибровочных растворов фермента с известной активностью, определенной другим способом анализа, для сопоставления с активностью по результатам данного анализа в единицах, определенных при калибровочном анализе, является общепринятым. Альтернативным образом, липазную активность можно определить посредством использования в качестве субстрата 2,3-меркапто-1-пропанолтрибутирата (TBDMP). Липаза вызывает гидролиз тиоэфирной(ых) связи(ей) в TBDMP, приводя тем самым к выделению масляной кислоты и 2,3-меркапто-1 пропанолдибутирата, 2,3-меркапто-1-пропанолмонобутирата или 2,3-меркапто-1-пропанола. Выделенные тиольные группы титровали по последующей реакции с 4,4-дитиодипиридином (DTDP) с образованием 4-тиопоридона. Последний находится в таутомерном равновесии с 4-меркаптопиридином, который демонстрирует поглощение при 334 нм. Реакцию проводили в 0,1 М ацетатном буферном растворе при рН 5,0, содержащем 0,2% Тритона Х-100, 0,65 мМ TBDMP и 0,2 мМ DTDP при 37 С. Одна липазная единица определяется как количество фермента, которое вызывает выделение 1 микромоль 4- 21015172 тиопиридона в минуту при в данных условиях реакции. Фосфолипазную А активность определяли спектрофотометрически при использовании в качестве субстрата 1,2-димеркаптодиоктаноилфосфатидилхолина. Фосфолипаза А вызывает гидролиз тиоэфирной связи в положении 1 (PLA1) или в положении 2 (PLA2), что приводит к выделению октановой кислоты и 1,2-димеркаптомонооктаноилфосфатидилхолина или 1,2-димеркаптофосфатидилхолина. Выделенные тиольные группы титровали по последующей реакции с 4,4'-дитиопиридином с образованием 4 тиопиридона. Последний находится в таутомерном равновесии с 4-меркаптопиридином, который демонстрирует поглощение при длине волны 334 нм. Реакцию проводили в 0,1 М ацетатном буферном растворе при рН 4,0, содержащем 0,2% Тритона Х-100, при 37 С. Одна фосфолипазная А единица (APLU) определяется как количество фермента, которое вызывает выделение 1 мкмоль 4-тиопиридона в минуту при данных условиях реакции. Лизофосфолипазная активность может быть определена посредством 31 Р-ЯМР спектроскопии при использовании в качестве субстрата лизофосфатидилхолина. Лизофосфолипаза вызывает гидролиз сложноэфирной связи, приводя к выделению жирной кислоты из глицеринового соединения. Количество таким образом образовавшегося глицерофосфохолина определяли при помощи ЯМР. Реакцию проводили в 50 мМ ацетатном буферном растворе с рН 4,5, дополнительно содержавшем 1 мг/мл лизофосфатидилхолина и 5 мМ CaCl2, в течение 30 мин при 55 С. Одна лизофосфолипазная единица (LPC) определяется как количество фермента, приводящее к образованию 1 мкмоль глицеринфосфохолина в минуту при данных условиях реакции. Галактолипазную активность определяли при помощи Н-ЯМР спектроскопии при использовании в качестве субстрата дигалактозилдиглицерида, согласно способу, описанному у Hirayama и Matsuda(1972) Agric. Biol. Chem. 36, 1831. Галактолипаза вызывает гидролиз сложноэфирной связи между жирными кислотами и глицериновой главной цепью, что приводит к выделению одной ил обеих жирных кислот. Реакцию проводил в 50 мМ ацетатном буферном растворе с рН 4,5, дополнительно содержавшем 4 мМ CaCl2, 0,2% Тритона Х-100 и 1 мг/мл дигалактозилдиглицерида (Lipid Products), в течение 30 мин при 30 С. Одна галактолипазная единица определяется как количество фермента, вызывающее выделение 1 мкмоль жирной кислоты в минуту при данных условиях реакции. Дополнительно к спектрофотометрическим измерениям липазная активность может также быть определена при помощи титриметрических измерений. Например, эстеразная активность липолитического фермента может быть измерена при использовании трибутирина в качестве субстрата, согласно FoodChemical Codex, Forth Edition, National Academy Press, 1996, стр. 803. Липазную активность предпочтительно определяют с использованием в качестве субстратов триацилглицеридов с более длинноцепочечными жирными кислотами, например пальмитиновой кислотой, стеариновой кислотой, олеиновой кислотой, линолевой кислотой, линоленовой кислотой. Часто в таких анализах используют оливковое масло. Фосфолипаза, лизофосфолипаза и галактолипаза, в принципе, также могут быть подвергнуты анализу при помощи титриметрических способов. Дополнительно к упомянутым типам липолитической активности, образцы могут проявлять побочные виды нелиполитической активности, например альфа-амилазная активность. Активность альфаамилазы грибного происхождения может быть измерена при помощи амилазных тестовых таблетокPhadebas (Pharmacia). Таблетки Phadebas содержат не растворимый в воде крахмальный субстрат и синий краситель, связанный с субстратом при помощи сшивок. Субстрат гидролизуется амилазой грибного происхождения, что приводит к высвобождению окрашенных растворимых мальтодекстринов, которые переходят в раствор. Калибровочная кривая была получена при помощи раствора, содержавшего сравнительную активность грибной альфа-амилазы. Из сравнительного и исследуемых образцов были получены соответствующие разбавления в 50 м буферном растворе малеиновой кислоты с рН 5,5. Образцы по 5 мл выдержали при 30 С в течение 5 мин,добавили таблетки Phadebas и через 15 мин, реакцию остановили добавлением 1,0 мл 0,5 М раствора гидроксида натрия. Смесям дали охладиться до комнатной температуры в течение 5 мин, после чего добавили по 4,0 мл воды, встряхнули в руках и, через 15 мин, образцы подвергали центрифугированию в течение 10 мин при 4700 об/мин. Экстинкцию верхнего слоя измеряли при 620 нм. OD при 620 нм является мерой активности альфа-амилазы грибного происхождения. Одна единица альфа-амилазной (грибного происхождения) активности (FAU) определяется здесь как количество фермента, которое конвертирует 1 г крахмала (100% сухого вещества) в час в продукт,имеющий пропускание при 620 нм после реакции с раствором йода известной концентрации в данных условиях реакции. Дополнительно к упомянутым видам активности, также присутствовали незначительные гликоамилазная, протеазная и ксиланазная активности, однако, при таких малых количествах, что данные ферменты не мешали экспериментам по выпечке, описанным в нижеприведенных примерах. Не содержащее клеток ультрафильтраты, полученные в примере 1, были подвернуты липазному, фосфолипазному и галактолипазному анализам, согласно собранным в табл. 2 данным.- 22015172 Таблица 2. Типы активности липолитического фермента в свободных от клеток осветленных фильтратах, полученных согласно примеру 1(липазную активность определяли при рН 5). Следует заметить, что в различных анализах присутствует только один субстрат, и величины активностей не могут предсказывать действительные активности в смесях различных липоидных субстратов или при промышленном применении, таком как применение в тесте. В таких случаях аффинность или специфичность по отношению к субстратам становится важной. Энзиматическая характеристика Определение молекулярной массы методом ДДС Na-ПААГ для образцов ультрафильтратов проводили на NuPage 4-12% MES Simply Blue Safe Stain. Для LIP01 получили Mw = 33 кДа. Для LIP02 получили Mw = 33 кДа. Для LIP03 наблюдались две главные полосы, соответствующие Mw = 33 кДа и Mw = 41 кДа. Эксперимент по изоэлектрическому фокусированию (IEF) Расчетное pI для зрелого белка LIP01 из 275 аминокислот: 5. Расчетное pI для транслированного гена LIP01: 5,4. Расчетное pI для транслированного гена LIP02: 5,4. Расчетное pI для зрелого белка LIP02 из 276 аминокислот: 5,5. pI для LIP02 определяли экспериментально при помощи гель-электрофореза в интервале амфолитов 3-10. IEF показало множественные полосы в интервале 4-5 с главной полосой при значении pI = 4,7 и 4,3. Расчетное pI для зрелого белка LIP03 из 276 аминокислот: 5,1 (с использованиемSEQ1). Расчетное pI для транслированного гена LIP03 из 348 аминокислот: 5,1 (с использованием SEQ10). Расчетное pI для транслированного гена LIP03 из 314 аминокислот: 4,8 (с использованием SEQ14). Изоэлектрическое фокусирование для липазы LIP03, произведенной A. niger, показало мультиплетные полосы в интервале от pI = 4 до pI = 5,0 с главной полосой при pI = 4,7 и pI = 4,4. Определение гликозилирования Гликозилирование может влиять на молекулярную массу, наблюдаемую на гелях ДДС Na-ПААГ. Обычно молекулярная масса является завышенной. Для того чтобы проверить, является ли белок LIP0103 гликозилированным, а также для того, чтобы эффективно определить молекулярную массу белка, образец белка обработали PNGASE-F гликозидазой, чтобы дегликозилировать белок. Затем, как обработанный, так и необработанный образцы подвергали гель-электрофорезу. В зрелом белке LIP01 из 275 аминокислот присутствуют два потенциальных центра N-гликозилирования: 126 NLTF и 264 NYTF. В зрелом белке LIP02 из 276 аминокислот присутствует один потенциальный центр гликозилирования: 264NYTF. Необработанный LIP02 демонстрирует полосу около 33 кДа, тогда как после дегликозилирования наблюдается полоса вблизи 30 кДа. В зрелом белке из 276 аминокислот присутствует один потенциальный центр N-гликозилирования: 264 NYTF. Необработанный LIP03 демонстрирует две полосы, одну около 33 кДа, и одну около 41 кДа. После дегликозилирования снова наблюдаются две полосы, одна около 30 кДа, и одна около 38 кДа. Эти результаты предполагают, что существует две формы LIP03, обе гликозилированные в одинаковой степени. Характеристика и обработка гликопротеинов широко описана в The Protein Protocols Handbook, 2ndedition, Edited by J.M. walker, Humana Press Inc., Totowa 2002, глава VI. Интактная масса полученного липолитического фермента может быть определена посредством применения ЖХ/МС, согласно следующему протоколу: Образцы белков были обессолены путем фильтрации на 10 кДа центробежном фильтре (Pall) центрифугированием при 13000 об/мин в течение 15 мин при 4 С. Дегликозилирование осуществили путем- 23015172 ферментативного дегликозилирования пептид-N-гликозидазой F (PNGase, Roche Diagnostics GmbH, Мангейм, Германия). Фильтрат LIP01-LIP03 растворили в 100 мМ растворе гидрокарбоната аммония и денатурировали путем выдерживания при 95 С в течение десяти минут. К образцам добавили PNGASE-F (20 единиц) и провели дегликозилирование путем выдерживания при 37 С всю ночь. После дегликозлирования образцы снова профильтровали на 10 кДа центробежном фильтре (Pall) центрифугированием при 13000 об/мин в течение 15 мин для отделения от сахаров. Фильтраты после обессоливания и фильтраты после дегликозилирования растворили в 50/50/5 AcN/MQ/FA для получения конечной концентрации приблизительно 1 мг/мл. Образцы вводили в масс-спектрометр QTOFII прямым введением и рассчитали полные массы при помощи логарифма MaxEnt1 программного обеспечения Masslynx (версия 4 sp,Waters). Для LIP02 значение молекулярной массы 32265 было рассчитано по деконволюции масс-спектра интактного образца LIP02. Для дегликозилированного LIP02 была рассчитана интактная масса, составляющая 29905 кДа, что соответствует остаткам 35-310 теоретической последовательности аминокислот(SEQ NO 2). Различие в интактной массе, наблюдаемой до и после дегликозилирования, соответствует,вероятно, 12 маннозным группам и 2 группам GlcNAC, присоединенным к белку. Для дегликозилированного LIP03 наблюдалось более одной интактной массы. Были рассчитаны обе интактные массы LIP03 с и без С-концевого пептида, MW = 29835 (SEQ3, 35-310) и MW=34100 (SEQ6,35-348), соответственно. Дополнительно, С-конец фрагмента с молекулярной массой MW=29835 (SEQ3) был неровным, поскольку также были обнаружены массы остатков 35-309 (SEQ4) и 35-308 (SEQ5), где в особенности остаток 35-309 является полностью превалирующим по сравнению с остатком 35-310. Это показывает, что С-концевое отщепление не является для LIP03 полностью специфическим. Оптимальный уровень рН Зависимость оптимального уровня рН для липолитического фермента может быть определена посредством проведения анализа, при котором измеряют определенный тип липолитической активности при различных уровнях рН. рН, при котором наблюдается максимальная активность, представляет собой оптимальный уровень рН для конкретного фермента. Оптимальный уровень рН может зависеть от типа используемого субстрата или когда условия проведения анализа сильно меняются. Оптимальная температура Оптимальную температуру для липолитического фермента определяли посредством проведения данного анализа при разных температурах. Путем построения графика активности как функции от температуры можно определить оптимальную температуру для фермента. Термическая стабильность Термическую стабильность липолитического фермента можно определить посредством дифференциальной сканирующей калориметрии (DSC). Альтернативным образом, температурная стабильность может быть изучена путем установления Т 50. Т 50 определяется здесь как температура, при которой теряется 50% активности при нагревании липолтического фермента в течение 20 мин при данных условиях. Стабильность при хранении можно определить посредством хранения липолитического фермента при определенных условиях при определенной температуре. После истечения разных промежутков времени небольшие части образцов отбирали и определяли остаточную активность этих образцов при стандартных условиях анализа. Пример 4. Экспериментальная выпечка - мини-багет. Мини-багеты были выпечены из кусочков по 150 г теста, которое было получено путем смешивания 200 г муки (Kolibri), 4,6 г прессованных дрожжей, 4 г соли, 68 м.д. аскорбиновой кислоты, 1 м.д.Bakenzyme Р 500 (альфа-амилаза грибного происхождения), 5 м.д. Bakenzyme HSP6000 (хемицеллюлаза грибного происхождения) и 114 мл воды. После смешивания в течение 6 мин и 15 с в лопастном смесителе тесто раздели на два куска по 150 г, придали им круглую форму и выдержали 25 минут при комнатной температуре и относительной влажности 90%. Затем куски теста поместили в форму, сформовали и выдержали 100 мин при 32 С и относительной влажности 85%. Полностью выдержанные куски теста надрезали и выпекали в печи при 240 С в течение 20 мин с первичным добавлением пара. Различные эффекты липолитических ферментов LIP01-LIP03 в различных количествах сравнивали с пустым опытом, буханкой, не содержавшей дополнительных добавок, и контрольной буханкой, содержавшей 0,3% DATEM (Panodan 80CP). Объем буханки определяли при помощи автоматического анализатора объема хлеба (BVM-3, TexVol Instruments AB, Викен, Швеция) после остывания буханок. Другие эффекты оценивал пекарь согласно шкалам, представленным в табл. 3. Таблица 4. Рабочие характеристики при выпечке для липолитического фермента LIP01 при различных количествах (мг белка на кг муки Таблица 5. Рабочие характеристики при выпечке для липолитического фермента LIP02 при различных количествах (мг белка на кг муки Пример 5. Экспериментальная выпечка - полноразмерный багет. Рабочие характеристики при выпечке для липолитических ферментов LIP01-LIP03 были изучены также на полноразмерном багете. 3000 г муки (Kolibri), 70 г прессованных дрожжей, 60 г соли, 50 м.д. аскорбиновой кислоты, 2 м.д. Bakenzyme P500 (альфа-амилаза грибного происхождения), 15 м.д.Bakenzyme HSP6000 (хемицеллюлаза грибного происхождения) и 1740 мл воды смешивали в Doisnamixer в течение 2 мин на скорости 1 и 100 Вт/ч на скорости 2, температура конечного теста 27 С. Тесто разделил на 6 кусков по 350 г, придали круглую форму и выдержали 20 мин при 32 С и относительной влажности 90%. Затем куски теста поместили в форму, сформовали и выдержали 100 мин при 34 С и относительной влажности 90%. Полностью выдержанные куски теста надрезали и выпекали в печи при 240 С в течение 30 мин при первичном добавлении пара. Различные эффекты липолитического фермента в различных количествах, как на тесто, так и на готовое печеное изделие, сравнивали с пустым опытом, буханкой, не содержавшей дополнительных добавок, и контрольной буханкой, содержавшей 0,3% DATEM (Panodan 80CP). После остывания буханок определяли их объем при помощи автоматического анализатора объема хлеба (BVM-3, TexVol Instruments AB, Викен, Швеция). Другие эффекты ручным способом и визуально оценивал опытный пекарь согласно шкалам, представленным в таблице 6. Результаты представлены в табл. 7 и 8.- 25015172 Таблица 6. Оценки эффектов, наблюдавшихся для полноразмерного багета. Таблица 7. Рабочие характеристики при выпечке для липолитического фермента LIP01 при различных количествах (мг белка на кг муки Таблица 8. Рабочие характеристики при выпечке для липолитического фермента LIP02 при различных количествах (мг белка на кг муки Пример 6. Определение конверсии липидов в тесте мини-багета. Полярные липиды Липиды экстрагировали путем сильного взбалтывания лиофилизированного и размолотого полностью выдержанного теста (см. пример 3) с водонасыщенном бутанолом. После центрифугирования прозрачную надосадочную жидкость анализировали посредством ВЭЖХ на LiChrospher 100 DIOL 5 мкм(2504,0 мм), липоидные компоненты определяли при помощи испарительного детектора светорассеяния (Alltech ELSD 2000ES), в токе азота 1,5 л/мин, при температуре 80 С, с включенным импактором. Элюирование осуществляли с применением двух подвижных фаз по градиентной программе, при потоке 1 мл/мин: А: гептан/изопропанол/бутанол/тетрагидрофуран/изооктан/вода (64,5/17,5/7/5/5/1)- 26015172 В: изопропанол/бутанол/тетрагидрофуран/изооктан/вода (73/7/5/5/10). К обоим элюирующим растворам добавляли по 77 мкл аммиачного раствора и 77 мкл трифторуксусной кислоты на литр. Градиентная программа: линейный от 100% А до 100% В в течение 25 мин, затем 100% В в течение 5 мин, затем линейный градиент от 100% В до 100% А в течение 0,5 мин и в конце 100% А в течение 5 мин при вводимом объеме 20 мкл при температуре колонки 80 С. Для того чтобы указать порядок элюирования различных компонентов и рассчитать для них коэффициенты чувствительности детектора и количества, присутствующие в тесте, использовали эталонные галактолипиды, фосфолипиды, три-, ди- и моноглицериды, например, моногалактозилдиглицерид, моногалактозилмоноглицерид, дигалактозилдиглицерид, дигалактозилмоноглицерид, фосфатидилхолин и лизофосфатидилхолин. В табл. 9 и 10 представлены количества основных полярных липидов в полностью выдержанном тесте, содержащем различные количества LIP01-LIP03. Из этих результатов ясно, что LIP01-LIP03 эффективно превращает галактозилдиглицериды в галактозилмоноглцериды при относительно низком содержании, с преимуществом для дигалактозилдглицерида по сравнению с моногалактозилдиглицеридом,а также по сравнению с фосфатидилхолином. Кроме того, ясно, что оптимальный результат выпечки примера 4 при количестве в 2,4 м.д. (белок по методу Брэдфорда) совпадает с наивысшим уровнем дигалактозилмоноглицерида. Таблица 9. Полярные липиды в полностью расслоенном тесте(выражено в г на кг лиофилизированного теста), содержащем различные количества LIP01 (выражено в мг белка по методу Брэдфорда на кг муки).(выражено в г на кг лиофилизированного теста), содержащем различные количества LIP02 (выражено в мг белка по методу Брэдфорда на кг муки).MGDG = моногалактозилдиглицерид, MGMG = моногалактозилмоноглицерид, DGMG = дигалактозилмоноглицерид, DGMG =дигалактозилдиглицерид, PC = фосфатдилхолин, LPC = лизофосфатидилхолин. Неполярные липиды Неполярные липиды экстрагировали путем сильного взбалтывания лиофилизированного и размолотого полностью выдержанного теста (см. пример 1) с гептаном, содержащим 1% уксусной кислоты. После центрифугирования прозрачную надосадочную жидкость анализировали посредством ВЭЖХ наSpherisorb S3CN (Phenomenex OOD-0097-EO; 1004,6 мм), липоидные компоненты определяли при помощи испарительного детектора светорассеяния (Alltech ELSD 2000ES), в токе азота 1,5 л/мин, при температуре 40 С, с выключенным импактором. Элюирование осуществляли с применением двух подвижных фаз (А: гептан и В: трет-бутилметиловый простой эфир, содержащий 1% уксусной кислоты) по следующей градиентной программе, при потоке 1,0 мл/мин, вводимый объем 20 мкл и при температуре колонки, равной температуре окружающей среды: Для того чтобы указать порядок элюирования различных компонентов и рассчитать для них коэффициенты чувствительности детектора и количества, присутствующие в тесте, использовали эталонные три-, ди-, моноглицериды и жирную кислоту.
МПК / Метки
МПК: C07K 14/37
Метки: липазы, применение, новые
Код ссылки
<a href="https://eas.patents.su/30-15172-novye-lipazy-i-ih-primenenie.html" rel="bookmark" title="База патентов Евразийского Союза">Новые липазы и их применение</a>
Новые липазы и их применение

Номер патента: 8007
Опубликовано: 27.02.2007
Авторы: Ильгенфритц Хильмар, Фритц Андреас, Спреафико Фабио, Вагнер Кристиан, Мейма Рульф Бернхард, Хайнрих Оливер, Фолькерс Ульрике, Майер Дитер, Альбанг Рихард, Бур Де Лекс, Герхард Беатрикс
МПК: C12N 15/52, C07K 16/14, C12N 15/62...
Метки: липазы, новые, применение
Формула / Реферат:
1. Выделенный полинуклеотид, гибридизуемый с полинуклеотидом, выбранным из группы, состоящей из SEQ ID NO:10, 11. 2. Выделенный полинуклеотид по п.1, гибридизуемый в жестких условиях с полинуклеотидом, выбранным из группы, состоящей из SEQ ID NO:10, 11. 3. Выделенный полинуклеотид по п.1 или 2, получаемый из мицельного гриба. 4. Выделенный полинуклеотид по п.3, получаемый из Aspergillus niger. 5. Выделенный полинуклеотид, кодирующий полипептид,...
Соединения-ингибиторы липазы поджелудочной железы, их синтез и применение

Номер патента: 9368
Опубликовано: 28.12.2007
Авторы: Уиттер Дэвид, Кастелано Арлиндо Л.
МПК: A61K 31/5365, A61P 3/04, A61K 31/542...
Метки: поджелудочной, синтез, липазы, соединения-ингибиторы, применение, железы
Формула / Реферат:
1. Соединение, имеющее следующую структуру: где X представляет собой О, S или СН2; Y представляет собой О; R1 представляет собой Н, замещенный C1-C15алкил, незамещенный С2-С15алкил С1-С8алкиларил, -C(O)OR4, -C(O)NR4R5, -CR6R6'OR4, -CR6R6'OC(O)R4, -CR6R6'OC(O)NHR7, -С(О)NR8R9NR8R9, -N(R5)С(О)NHR5 или CH2R4; R2 представляет собой замещенный или незамещенный неразветвленный C1-C30алкил или разветвленный С3-С30алкил, арил, алкиларил, арилалкил,...
Новые порфирины и их применение

Номер патента: 2910
Опубликовано: 31.10.2002
Авторы: Шульга Александр, Шастак Станислав, Берр Фридер, Видеманн Петер
МПК: A61K 31/40, A61P 35/00, C07D 487/22...
Метки: порфирины, новые, применение
Формула / Реферат:
1. Соединения общей формулы где R1 обозначает C1-С6-алкил или арил, R2 - R5 независимо друг от друга обозначают Н, ОН, C1-С6-алкил, C1-С6-алкилен или OR6, причем R6 обозначает C1-С6-алкил или арил, Аn- обозначает анион и n равно 1 или 2. 2. Соединения по п.1, отличающиеся тем, что R1 обозначает метил, этил, пропил, бутил, пентил или гексил, в особенности метил. 3. Соединения по п.1 или 2, отличающиеся тем, что R2 - R5 обозначают Н. 4....
Новые производные бензимидазолов и бензотиазолов, способ их получения, их применение в качестве лекарственных средств, фармацевтические композиции и новое применение, в частности, в качестве ингибиторов cmet

Номер патента: 14315
Опубликовано: 29.10.2010
Авторы: Клерк Франсуа, Немесек Консепсьон
МПК: C07D 235/30, C07D 277/82, C07D 235/32...
Метки: бензотиазолов, фармацевтические, ингибиторов, бензимидазолов, композиции, частности, производные, новые, качестве, лекарственных, новое, применение, получения, средств, способ
Формула / Реферат:
1. Соединения формулы (I)в которой А обозначает NH или S;R1 и R2, одинаковые или разные, выбирают из атомов водорода, радикала NH2, атомов галогена и алкильных радикалов, необязательно замещенных одним или несколькими атомами галогена,и R3 обозначает атом водорода или выбран из значений R1 и R2, причем по меньшей мере один из R1, R2 и R3 не является водородом,R обозначает циклоалкил или алкил, необязательно замещенный фенилом, гетероарилом,...
Новые соединения, их применение и получение

Номер патента: 8148
Опубликовано: 27.04.2007
Авторы: Йенссон Маттиас, Нильссон Бьерн, Рингберг Эрик, Шеберг Биргер
МПК: A61K 31/497, A61K 31/444, A61P 25/00...
Метки: применение, новые, соединения, получение
Формула / Реферат:
1. Соединение формулы (I) где nх равно 2-4, предпочтительно 2; каждый из R0 и R1 независимо означает Н или СН3; R2 означает Н, C1-C4-алкил, 2-гидроксиэтил, 2-цианоэтил или тетрагидропиран-2-ил, C1-C4-ацил или C1-C4-алкоксикарбонил; каждый из R3-R5 независимо означает Н, галоген, метил или метокси, при условии, что по меньшей мере один из R3-R5 означает водород; каждый из X, Y и z независимо означает СН или N; A1 означает О, СН или СН2; A2...
Предыдущий патент: Лечение субъектов с хроническим заболеванием почек с применением соединений лантана
Следующий патент: Комбинированная терапия с использованием антител к egfr и her2