Новые иммуноадъювантные соединения на основе флагеллина и их применение




























Формула / Реферат
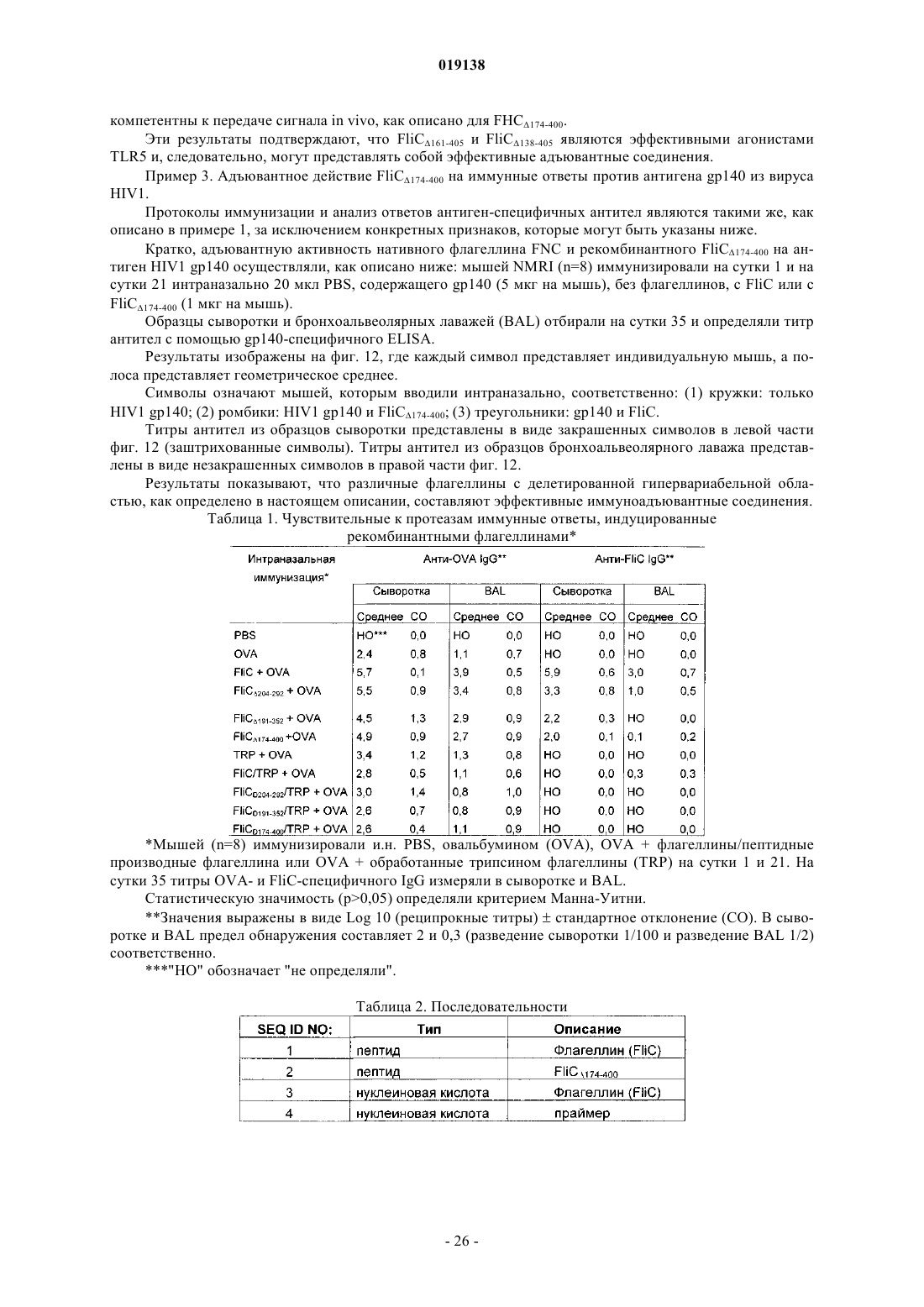
1. Иммуноадъювантное соединение, содержащее:
а) N-концевой пептид, обладающий по меньшей мере 90% аминокислотной идентичностью с аминокислотной последовательностью, начинающейся с аминокислотного остатка, локализованного в положении 1 SEQ ID NO:1, и оканчивающейся аминокислотным остатком, выбранным из группы, состоящей из любого из аминокислотных остатков, локализованных в положениях 99-173 SEQ ID NO:1; и
б) С-концевой пептид, обладающий по меньшей мере 90% аминокислотной идентичностью с аминокислотной последовательностью, начинающейся с аминокислотного остатка, выбранного из группы, состоящей из любого из аминокислотных остатков, локализованных в положениях 401-406 SEQ ID NO:1, и оканчивающейся аминокислотным остатком, локализованным в положении 494 SEQ ID NO:1; где
указанный N-концевой пептид непосредственно связан с указанным С-концевым пептидом или
указанный N-концевой пептид и указанный С-концевой пептид связаны друг с другом опосредованно через спейсерную цепь, где спейсерная цепь содержит от 1 до 20 аминокислот, связанных пептидными связями, где аминокислоты выбраны из 20 природных аминокислот.
2. Иммуноадъювантное соединение по п.1, где указанный N-концевой пептид выбран из группы, состоящей из аминокислотных последовательностей 1-99, 1-137, 1-160 и 1-173 SEQ ID NO:1.
3. Иммуноадъювантное соединение по любому из пп.1 или 2, где указанный С-концевой пептид выбран из группы, состоящей из аминокислотных последовательностей 401-494 и 406-494 SEQ ID NO:1.
4. Иммуноадъювантное соединение по любому из пп.1-3, где указанные N-концевой и С-концевой пептиды состоят из аминокислотных последовательностей 1-173 и 401-494 SEQ ID NO:1 соответственно.
5. Иммуноадъювантное соединение по любому из пп.1-3, где указанные N-концевой и С-концевой пептиды состоят из аминокислотных последовательностей 1-160 и 406-494 SEQ ID NO:1 соответственно.
6. Иммуноадъювантное соединение по любому из пп.1-3, где указанные N-концевой и С-концевой пептиды состоят из аминокислотных последовательностей 1-137 и 406-494 SEQ ID NO:1 соответственно.
7. Иммуноадъювантное соединение по любому из пп.1-6, где указанный N-концевой пептид и указанный С-концевой пептид связаны друг с другом опосредованно через промежуточную спейсерную цепь, состоящую из пептидной последовательности NH2-Gly-Ala-Ala-Gly-COOH.
8. Иммуноадъювантное соединение по любому из пп.1-7, где аминокислотный остаток аспарагина, локализованный в положении 488 SEQ ID NO:1, заменен серином.
9. Иммуноадъювантное соединение по любому из пп.1-7, где указанное соединение содержит дополнительный остаток метионина на N-конце.
10. Фармацевтическая композиция, содержащая иммуноадъювантное соединение по любому из пп.1-9 вместе с одним или более чем одним фармацевтически приемлемым эксципиентом.
11. Иммуногенная композиция, содержащая иммуноадъювантное соединение по любому из пп.1-9 вместе с одним или более чем одним антигеном.
12. Вакцинная композиция, содержащая иммуноадъювантное соединение по любому из пп.1-9 вместе с одним или более чем одним антигеном.
13. Иммуногенная композиция по п.11 или вакцинная композиция по п.12, где указанное иммуноадъювантное соединение нековалентно связано с одним или более чем одним указанным антигеном.
14. Применение иммуноадъювантного соединения по любому из пп.1-9 в качестве лекарственного средства.
15. Применение иммуноадъювантного соединения по любому из пп.1-9 для изготовления фармацевтической композиции.
16. Нуклеиновая кислота, кодирующая иммуноадъювантное соединение по любому из пп.1-9.
17. Рекомбинантный вектор, содержащий встроенную в него нуклеиновую кислоту по п.16.
18. Клетка-хозяин, трансфицированная или трансформированная нуклеиновой кислотой по п.16 или рекомбинантным вектором по п.17.
Текст
НОВЫЕ ИММУНОАДЪЮВАНТНЫЕ СОЕДИНЕНИЯ НА ОСНОВЕ ФЛАГЕЛЛИНА И ИХ ПРИМЕНЕНИЕ Изобретение относится к новым пептидным соединениям, являющимся производными флагеллина, имеющего происхождение из Salmonelle enterica, которые проявляют иммуноадъювантную активность in vivo.(71)(73) Заявитель и патентовладелец: ИНСЭРМ (ИНСТИТЮТ НАСИОНАЛЬ ДЕ ЛЯ САНТЭ Э ДЕ ЛЯ РЕШЕРШ МЕДИКАЛЬ); ИНСТИТЮТ ПАСТЕР ДЕ ЛИЛЛЬ Область изобретения Изобретение относится к индукции и/или стимуляции иммунного ответа у индивидуума или животного. Оно касается, в частности, новых иммуноадъювантных соединений, полезных для иммуногенных и вакцинных композиций. Предшествующий уровень техники Разработка безопасных и эффективных вакцин остается основной целью в мировом здравоохранении. В частности, появились вакцины, называемые "мукозальными", в качестве привлекательной потенциальной альтернативы инъекционным вакцинам. Мукозальное введение обладает многими потенциально желательными свойствами. Возможно,наиболее обоснованным доводом для разработки методов доставки мукозальных вакцин является развитие первой линии иммунологической защиты путем создания местного иммунитета в сайте внедрения многих инвазивных агентов в слизистую оболочку. Кроме того, некоторые исследователи сообщили, что существует общая мукозальная иммунная система, где мукозальный иммунитет, индуцированный в одном сайте, может привести к иммунитету в дистальном мукозальном сайте (McGhee, J. R. et al. The mucosal immune system: from fundamental conceptsto vaccine development. Vaccine 1992, 10:75-88). Кроме того, доставка антигена через мукозальный сайт также обладает потенциалом генерирования системного иммунного ответа. Это позволяет предположить, что значительные преимущества могут быть достигнуты благодаря доставке вакцин неинвазивным путем, например интраназальным или другим мукозальным путем, чтобы вызвать иммунитет к широкому ряду патогенов, которые могут внедряться в различные сайты слизистой оболочки. Большинство современных вакцин (мукозальных или других вакцин) состоит из двух основных компонентов: (1) антигена-мишени, представляющего терапевтический интерес, и (2) иммуноадъювантного соединения(й), которое стимулирует и/или индуцирует иммуногенность против указанного антигена. Природа известных иммуноадъювантов широко варьирует, но включает, в частности, минеральные масла, бактериальные экстракты, живые и аттенуированные организмы и суспензии алюмогидроксидов металлов. Даже если адъюванты обеспечивают усиленные иммунные ответы, их применение может также вызвать вредные побочные эффекты, зависящие исключительно от пути их введения. Поэтому число адъювантов, которые одобрены и эффективны у людей, остается относительно ограниченным. Успехи в области наследственного иммунитета привели к лучшему пониманию как клеточных, так и молекулярных механизмов, управляющих регуляцией иммунного ответа хозяина. Это лучшее знание иммунной системы дало возможность исследования и разработки новых потенциально полезных иммуноадъювантов. В частности, toll-подобные рецепторы (TLR) являются инструментами в координированной индукции наследственного и адаптивного иммунитета у млекопитающих. Поскольку TLR экспрессируются широким рядом типов клеток, они способны запускать иммунитет во всем организме. После инфицирования патогенными микроорганизмами TLR распознают консервативные мотивы,называемые микроорганизм-ассоциированными молекулярными паттернами (МАМР). ВовлеченностьTLR индуцирует программу генной экспрессии, направленной как на наследственно-обусловленный клиренс патогенных микроорганизмов, так и на приобретенный иммунитет к ним. Например, TLR индуцируют продуцирование хемокинов, которые, в свою очередь, специфично привлекают полиморфноядерные нейтрофилы (ПМН), непосредственно вовлеченные в наследственно-обусловленный клиренс микроорганизмов. Кроме того, TLR стимулируют секрецию плейотропных иммунных медиаторов (таких как TNF) и функциональное созревание дендритных клеток (ДК), которые специализированы на презентацию антигенов лимфоцитам. Следовательно, агонисты TLR не только стимулируют провоспалительные иммунные ответы "широкой специфичности", но также усиливают адаптивный иммунный ответ на определенные антигены, и поэтому их считают иммуноадъювантами. Несмотря на эти потенциально полезные эффекты, системная токсичность МАМР вызвала попытки разработать производные, которые сдвигают активность МАМР в направлении адъювантной способности. Действительно, конструирование молекул с уникальными свойствами является основным направлением в манипуляции иммунными ответами. Бактериальные флагеллины (основные компоненты жгутиков во многих бактериальных патогенах) являются специфичными уникальными агонистами для активации TLR5. Флагеллин FNC из Salmonella enterica серовар Typhimurium (S. Typhimurium) является парадигмой для исследований по структуре и функции флагеллинов, иммунитету и передаче сигнала TLR5. Он представляет собой 494-аминокислотный белок с двумя отдельными доменами. Амино- и карбоксиконцевые "консервативные" области образуют домен, который существенен для активации TLR5.-1 019138 Средний домен флагеллина FNC содержит аминокислоты, не обязательные для передачи сигналаTLR5. Он предназначен в качестве "гипервариабельной" области, поскольку первичные последовательности значительно варьируют по составу и размеру от одного вида бактерий к другому. Напротив, известно, что гипервариабельная область существенна для антигенности флагеллина. Показано, что внутривенная (в.в.) инъекция флагеллинов стимулирует системный ответ, характеризующийся продуцированием провоспалительных медиаторов (таких как TNF или IL-6) и активацией ДК. Кроме того, флагеллины запускают специфичные для слизистой оболочки наследственнообусловленные и адаптивные защитные механизмы. Например, эпителиальные клеточные линии и слизистая оболочка легкого осуществляют позитивную регуляцию продуцирования хемокинов, таких какCXCL8 (IL-8) и CCL20, которые, в свою очередь, рекрутируют ПМН и ДК слизистой оболочки соответственно. Разные авторы также сообщали, что флагеллины являются сильными системными и мукозальными иммуноадъювантами, которые вызывают (1) ответы сывороточных и/или секреторных антител и (2) клеточные ответы Th1 и Th2 как на сами флагеллины, так и на совместно введенные антигены. Вследствие своих сильных системных и мукозальных иммуноадъювантных активностей флагеллины могут представлять особый интерес для разработки вакцины и, в частности, вакцины мукозального типа. Однако большинство указанных адъювантов флагеллинового типа не полностью подходит для применения в таких вакцинах, и, в частности, для стратегии указанных мукозальных вакцин. Действительно, известные флагеллиновые адъюванты проявляют значительные побочные эффекты,и, в частности, собственную антигенную активность и системные провоспалительные свойства при введении in vivo. Кроме того, большинство известных флагеллиновых иммуноадъювантов необходимо физически сшивать с антигеном-мишенью, чтобы вызвать сильный иммунный ответ при введении in vivo. Это требование обязывает к дополнительным сложным манипуляциям для получения подходящей связи флагеллин-антиген и конечного полезного иммуногенного вещества. Таким образом, существует необходимость в новых соединениях, которые можно было бы использовать в качестве иммунологических адъювантов, в частности, для индукции и/или для усиления мукозального иммунного ответа против антигена, исключительно без запуска какого-либо значительного системного воспалительного побочного эффекта. Эти новые соединения также предпочтительно должны обладать способностью к запуску иммунного ответа посредством простого смешения с антигеном-мишенью. В настоящем изобретении, таким образом, предложены новые иммуноадъювантные соединения,которые удовлетворяют эту потребность, и которые могут быть особенно полезны для получения иммуногенных композиций и вакцины (в частности, мукозального типа). Краткое изложение сущности изобретения Согласно данному изобретению обнаружены новые пептидные соединения, являющиеся производными флагеллина, имеющего происхождение из Salmonella enterica серовар Typhimurium, типа SEQ IDNO:1, которые проявляют иммуноадъювантную активность in vivo, как проиллюстрировано в примерах,приведенных в данной заявке. Согласно настоящему изобретению также показано, что эти новые адъювантные соединения проявляют, в частности, свойства мукозальных адъювантов, не оказывающих значительных системных провоспалительных эффектов. Указанные новые соединения по изобретению, являющиеся производными флагеллина, следовательно, особенно полезны в качестве иммуноадъювантных веществ, преимущественно для индукции и/или для усиления мукозального иммунного ответа. Таким образом, настоящее изобретение относится к иммуноадъювантному соединению, включающему: а) N-концевой пептид, обладающий по меньшей мере 90% аминокислотной идентичностью с аминокислотной последовательностью, начинающейся с аминокислотного остатка, локализованного в положении 1 SEQ ID NO:1, и оканчивающейся аминокислотным остатком, выбранным из группы, состоящей из любого из аминокислотных остатков, локализованных в положениях 99-173 SEQ ID NO:1; и б) С-концевой пептид, обладающий по меньшей мере 90% аминокислотной идентичностью с аминокислотной последовательностью, начинающейся с аминокислотного остатка, выбранного из группы,состоящей из любого из аминокислотных остатков, локализованных в положениях 401-406 SEQ ID NO:1,и оканчивающейся аминокислотным остатком, локализованным в положении 494 SEQ ID NO:1,где указанные N-концевой и С-концевой пептиды непосредственно связаны друг с другом или указанные N-концевой и С-концевой пептиды связаны друг с другом опосредованно через спейсерную цепь. Предпочтительные воплощения иммуноадъювантного соединения по изобретению определены в описании ниже. Изобретение также относится к фармацевтической композиции, содержащей иммуноадъювантное-2 019138 соединение, как оно определено выше (или в последующем описании), вместе с одним или более чем одним фармацевтически приемлемым эксципиентом. Фармацевтическая композиция по изобретению содержит иммуноадъювантное соединение, как оно определено выше, вместе с одним или более чем одним антигеном. Указанная фармацевтическая композиция, таким образом, преимущественно является иммуногенной композицией (то есть композицией, которая направлена на индукцию иммунного ответа против антигена, например, для выработки антител) или вакцинной композицией (то есть композицией, которая направлена на индукцию иммунного ответа у субъекта или животного с целью лечения или предупреждения заболевания). В соответствии с предпочтительным воплощением указанная иммуногенная композиция или указанная вакцина преимущественно содержит указанное иммуноадъювантное соединение по изобретению,которое нековалентно связано с одним или более чем одним указанным антигеном. Настоящее изобретение также относится к иммуноадъювантному соединению, как оно определено выше, для применения в качестве лекарственного средства (в частности, для индукции и/или для усиления мукозальной адъювантной активности). Данное изобретение также относится к применению иммуноадъювантного соединения в соответствии с изобретением для изготовления фармацевтической композиции, в частности, для индукции и/или для усиления иммунного ответа против одного или более чем одного антигена, отличного от белка флагеллина (в частности, в мукозальном компартменте после введения мукозальным путем). Данное изобретение также относится к (1) нуклеиновой кислоте, кодирующей иммуноадъювантное пептидное соединение, как раскрыто выше, (2) рекомбинантному вектору, содержащему указанную нуклеиновую кислоту, встроенную в него, (3) клетке-хозяину, трансфицированной или трансформированной указанной нуклеиновой кислотой или указанным рекомбинантным вектором. Краткое описание графических материалов Фиг. 1. Характеристики и перекрестная реактивность флагеллинов с делетированной гипервариабельной областью.(А) Схематическое 3D изображение рекомбинантных флагеллинов. Структура флагеллина FliC дикого типа представлена на левой панели с использованием Pymol(http://www.pymol.org). В мономере концевые области (1-170 и 400-494) плотно уложены в виде осспиралей и образуют структурный домен, вовлеченный в функцию жгутика. Мотив 89-96 (показан черным цветом) является существенным для передачи сигнала TLR5. "Гипервариабельный" домен FliC в основном состоит из -структур и -витков. С использованием программы Swiss-Model (http://www.expasy.org/spdbv/) общая структура была предсказана для FliC204-292 и PliC174-400, показывая частичную и полную делецию гипервариабельной области соответственно. Для FliC191-352 положения аминокислот,очерчивающих делецию, показаны на левой панели. FliC174-400 и FliC191-352 содержат линкеры GAAG иLELE на стыке делеции соответственно.(В, С) Перекрестная реактивность FliC-специфичных сывороток. Гипериммунные сыворотки были получены после подкожного (п.к.) введения флагеллина, включенного в препарат с CFA для примирования с последующими бустер-инъекциями IFA. Сыворотку титровали в ELISA (твердофазном иммуноферментном анализе) на FliC, FliC204-292, FliC191-352 и FliC174-400. Результаты являются репрезентативными для 2 экспериментов. (В) Перекрестная реактивность сыворотки анти-FliC. (С) Перекрестная реактивность сыворотки анти-FliC174-400. Статистическая значимость (р 0,05 в критерии Манна-Уитни) указана звездочкой. Фиг. 2. Эпителиальная и мукозальная провоспалительная активность флагеллинов с делетированной гипервариабельной областью.(А, В) Активация эпителиальных клеток рекомбинантными флагеллинами. Эпителиальные клетки человека активировали флагеллинами FliC, FliC204-292, FliC191-352, FliC174-400 или FliC174-400/89-96 в указанных концентрациях. Клетки Caco-Rumbo, несущие репортерный слитый генCCL20-luc, активировали в течение 6 ч и активность люциферазы нормализовали по максимальной активности, измеренной с насыщающими уровнями FliC (А). Бронхиальные эпителиальные клетки BEAS2B стимулировали в течение 16 ч, после чего измеряли уровни IL-8 в надосадочной жидкости. Результаты являются репрезентативными для 1 из 2 независимых экспериментов (В).(C, D) Стимуляция мукозального наследственно-обусловленного ответа делетированными флагеллинами. Рекомбинантные флагеллины или препараты, обработанные трипсином (1 мкг-эквивалент), вводили интраназально (и.н.) анестезированным мышам (n=3-5). Через 2 ч определяли уровни CCL20 специфичной мРНК в целых легких, используя количественную полимеразную цепную реакцию с обратной транскриптазой (кОТ-ПЦР) в реальном времени (С). Через шесть часов после инстилляции брали образцы бронхоальвеолярных лаважей (BAL) (черные столбики) и легких (незакрашенные столбики) для измерения концентрации CCL20 (D). Статистическую значимость (р 0,05) определяли по критерию Манна-Уитни.-3 019138 Фиг. 3. Адъювантный эффект флагеллинов с делецией гипервариабельной области. Мышей (n=8) иммунизировали и.н. овальбумином (OVA)флагеллины или холерный токсин (СТ) на сутки 1 и 21. На сутки 35 измеряли титры OVA-специфичного IgG в сыворотке (А) и BAL (В). Определяли концентрацию OVA-специфичного IgA в BAL (С). Результаты являются репрезентативными для 1 из 2 независимых экспериментов. Статистическую значимость (р 0,05) определяли по критерию Манна-Уитни. Фиг. 4. Собственные антигенные свойства флагеллинов без гипервариабельной области. Мышей (n=8) иммунизировали и.н. овальбумином (OVA)флагеллины или холерный токсин (СТ) или липополисахаридами (LPS) на сутки 1 и 21. На сутки 35 измеряли титры FliC-специфичного IgG в сыворотке (А) и BAL (В). Результаты являются репрезентативными для 1 из 2 независимых экспериментов. Статистическую значимость (р 0,05) определяли по критерию Манна-Уитни. Фиг. 5. Нейтрализация передачи сигнала TLR5 флагеллин-специфичными антителами. Мышей NMRI иммунизировали п.к. в первую неделю 1 мкг флагеллина FliC и CFA с последующими бустер-инъекциями FNC и IFA в недели 3, 5, 7. В контрольных условиях животных подобным образом обрабатывали овальбумином и адъювантами или одними адъювантами. Эксперименты проводили в неделю 9.(А) TLR5-нейтрализующая активность флагеллин-специфичной иммунной сыворотки in vitro. Эпителиальные клетки Caco-Rumbo, несущие репортерную конструкцию CCL20-luc, активировали в течение 6 ч флагеллином FliC, инкубировали с 50% об./об. FliC гипериммунными (незакрашенные кружки) или контрольными (черные кружки). Активность люциферазы определяли и нормализовали по активности, полученной с 100 нг/мл FliC. Результаты являются репрезентативными для 1 из 3 независимых экспериментов.(В,С) TLR5-нейтрализующая активность флагеллин-специфичной иммунной сыворотки in vivo. Иммунизированным животным (n=3) инъецировали в.в. забуференный фосфатом солевой раствор(PBS) (черные столбики) либо 0,1 мкг (серые столбики) или 1 мкг флагеллина FliC (незакрашенные столбики). Сыворотки собирали через 2 ч и определяли концентрации CCL20 (В) и CXCL2 (С) путем ELISA.(D) Нейтрализующая активность иммунной сыворотки. Животным (n=3 на дозу) пассивно переносили в.в. различные количества флагеллин-специфичной или контрольной сыворотки и через 1 ч их обрабатывали в.в. рекомбинантными флагеллинами, как указано. Продуцирование хемокинов в сыворотке через 2 ч после провокации измеряли путем ELISA. Статистическую значимость (р 0,05) определяли, используя критерий Манна-Уитни. Фиг. 6. Интраназальная активность доза-ответ флагеллинов FliC и FliC174-400. Мышам (n=3-5) вливали и.н. различные количества флагеллинов FliC (черные квадраты) илиBAL, используя ELISA. Статистическую значимость (р 0,05) определяли по U-критерию Манна-Уитни. Фиг. 7. Изменение способности к системной активации флагеллина FliC174-400 с делегированной гипервариабельной областью. Различные количества флагеллина FliC (черные квадраты) или FliC174-400 (незакрашенные квадраты) вводили в.в. Концентрации CCL20 (А) и CXCL2 (В) определяли через 2 ч в сыворотке, используяELISA. Статистическую значимость (р 0,05) определяли по критерию Манна-Уитни. Фиг. 8. Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (ДСНПААГ) на различных рекомбинантных флагеллинах с делетированной гипервариабельной областью. Фиг. 8 состоит из фотографии электрофореза в ДСН-ПААГ продуцируемых рекомбинантным путемFliC174-400, FliC161-405, FliC138-405 и FliC100-405 после окрашивания Кумасси синим. Фиг. 9. Иммуноблоттинг различных рекомбинантных флагеллинов с делетированной гипервариабельной областью. Фиг. 9 состоит из фотографии Вестерн-блот электрофореза продуцируемых рекомбинантным путемFliC174-400, FliC161-405, FliC138-405 и FliC100-405 после окрашивания антителами анти-FliC. Фиг. 10. Индукция продуцирования хемокина CCL20 различными рекомбинантными флагеллинами с делетированной гипервариабельной областью. Стимуляция системного наследственно-обусловленного ответа делетированными флагеллинами. Рекомбинантные флагеллины или препараты, обработанные трипсином (10 мкг-эквивалентов), вводили внутрибрюшинно (в.б.) мышам (n=2). Через два часа после инъекции брали образцы сыворотки для измерения концентрации CCL20. Фиг. 11. Индукция продуцирования хемокина CXCL2 различными рекомбинантными флагеллинами с делегированной гипервариабельной областью. Стимуляция системного наследственно-обусловленного ответа делетированными флагеллинами. Рекомбинантные флагеллины или препараты, обработанные трипсином (10 мкг-эквивалентов), вво-4 019138 дили в.б. мышам (n=2). Через два часа после инъекции брали образцы сыворотки для измерения концентрации CXCL2. Фиг. 12. Адъювантный эффект рекомбинантного FliC174-400 для иммунизации против антигена(незакрашенные символы). Результаты являются репрезентативными для 1 из 2 независимых экспериментов. Фиг. 13. Хроматографическая характеристика при 280 нм цикла очистки FliC на иммуноаффинном субстрате, на котором иммобилизованы мышиные моноклональные антитела анти-FliC174-400. Фиг. 14. Электрофоретический анализ различных хроматографических фракций, собранных во время цикла очистки FliC на иммуноаффинном субстрате, на котором иммобилизованы мышиные моноклональные антитела анти-FliC174-400. Фиг. 14 состоит из фотографии электрофореза в ДСН-ПААГ фракций, собранных, как изображено на фиг. 13, после окрашивания Кумасси синим. Подробное описание изобретения Согласно настоящему изобретению показано, что новые соединения индуцируют in vivo мукозальную иммуноадъювантную активность, дающую возможность индукции иммунного ответа против антигена-мишени, когда указанные новые соединения вводят с подходящим соответствующим антигеном(ами). Конкретно в данной заявке показано, что новые адъювантные соединения по изобретению проявляют свои иммуноадъювантные свойства также после интраназального введения мышам. Указанные иммуноадъювантные соединения по изобретению, таким образом, способны потенцировать системный и мукозальный иммунный ответ. Также продемонстрировано, что указанное иммуноадъювантное соединение по изобретению, являющееся производным флагеллина, обладает TLR5-опосредованными свойствами мукозального адъюванта с провоспалительным эффектом in vivo в слизистой оболочке, но не проявляет никакого значительного системного провоспалительного побочного эффекта после системной инъекции. Кроме того, результаты, содержащиеся в приведенных в данной заявке примерах, показывают, что указанное иммуноадъювантное соединение, являющееся производным флагеллина, не проявляет значительного собственного антигенного эффекта, то есть интересующая молекула предотвращает или ослабляет эффективность запуска флагеллин-специфичных антител, а именно в сыворотке или бронхоальвеолярном лаваже (BAL) при введении интраназальным путем. Вышеописанные результаты показывают, что указанное иммуноадъювантное соединение по изобретению, являющееся производным флагеллина, можно применять в качестве эффективного адъюванта иммунного ответа, в частности, для индукции мукозальных иммунных ответов. Указанное пептидное соединение, таким образом, может быть полезно, в частности, когда оно содержится в (1) мукозальных вакцинных композициях для предупреждения или для лечения заболеваний посредством индукции мукозального иммунного ответа в организме субъекта или (2) иммуногенной композиции для усиления или запуска иммунного ответа против желаемого антигена. В частности, как показано в приведенных здесь примерах, авторы изобретения неожиданно обнаружили, что передача сигнала TLR5 компартментализирована, поскольку новый конкретный флагеллинSEQ ID NO:1 флагеллина FliC Salmonella enterica серовар. Typhimurium ATCC14028 делегирована от положения 174 до положения 400) стимулирует иммунитет в слизистой оболочке, но лишен какого-либо значительного системного провоспалительного эффекта. Авторы изобретения также установили, что флагеллин FliC174-400 обладает выраженными полезными свойствами вследствие его слабой способности генерировать нейтрализующие FliC-специфичные антитела. Кроме того, в данном изобретении обнаружено, что флагеллин FliC174-400 сильно аттенуирован для системной передачи сигнала по сравнению с флагеллином дикого типа, тогда как мукозальная активность остается неизменной. В данном изобретении также показано, что другие флагеллины с делетированной гипервариабельной областью, включая FliC161-405 и FliC138-405, наделены иммуноадъювантными свойствами. Иммуноадъювантные пептиды по изобретению Эти обнаружения позволили авторам изобретения сконструировать семейство пептидов, которые должны обладать такими же свойствами и преимуществами, что и флагеллины FliC174-400, FliC161-405 иFliC138-405. Указанное семейство пептидов определено, исходя из флагеллинов FliC174-400, FliC161-405 И FliC138, исследованных в приведенных здесь примерах, и основываясь на пептидной последовательности 405 флагеллина SEQ ID NO:1 и на кристаллографической структуре пептида, чтобы предсказать укорочен-5 019138 ные варианты, которые могут обладать TLR5-стимулирующей активностью. Настоящее изобретение, таким образом, преимущественно относится к иммуноадъювантному соединению, включающему: а) N-концевой пептид, имеющий аминокислотную последовательность, начинающуюся с аминокислотного остатка, локализованного в положении 1 SEQ ID NO:1, и оканчивающуюся аминокислотным остатком, выбранным из группы, состоящей из любого из аминокислотных остатков, локализованных в положениях 99-173 SEQ ID NO:1; и б) С-концевой пептид, имеющий аминокислотную последовательность, начинающуюся с аминокислотного остатка, выбранного из группы, состоящей из любого из аминокислотных остатков, локализованных в положениях 401-406 SEQ ID NO:1, и оканчивающуюся аминокислотным остатком, локализованным в положении 494 SEQ ID NO:1, где указанный С-концевой пептид непосредственно связан с N-концевым пептидом или указанные N-концевой пептид и С-концевой пептид связаны друг с другом опосредованно через промежуточную спейсерную цепь."Включает/включающий" и их грамматические варианты при использовании в данном описании следует считать указывающими на присутствие установленных признаков, целых чисел, стадий или компонентов или их групп, но не исключающим присутствие или добавление одного или более чем одного другого признака, целого числа, стадии или компонента или их групп. Соединение по изобретению может быть взаимозаменяемо названо здесь "иммуноадъювантным соединением" или "пептидным производным флагеллина". Под "иммуноадъювантным соединением" понимают, что пептидное производное флагеллина по изобретению может индуцировать и/или усиливать иммунный ответ против антигена при введении субъекту или животному. Под ним также подразумевают вещество, которое действует в целом на ускорение, пролонгирование или усиление качества специфичных иммунных ответов на специфичный антиген. Как описано здесь, указанное иммуноадъювантное соединение можно применять в вакцинной или иммуногенной композиции вместе с одним или более чем одним антигеном и фармацевтически приемлемым эксципиентом. Вышеупомянутая пептидная последовательность SEQ ID NO:1 имеет происхождение из Salmonellaenterica серовар Typhimurium ATCC14028 флагеллин FliC (номер доступа AAL20871). Нумерация полипептида начинается с первой аминокислоты после возможного N-концевого метионина (не показанного в SEQ ID NO:1), который типично отсекается метионинаминопептидазой в бактериальных клетках-хозяевах, как упомянуто ниже.N-концевой и С-концевой пептиды указанного пептидного производного флагеллина по изобретению предпочтительно обладают аминокислотной идентичностью по меньшей мере 90% и даже более с соответствующим участком аминокислотной последовательности SEQ ID NO:1. Описание идентичности и того, как это можно определить, хорошо известно специалистам в данной области техники. Как подразумевается здесь, данная представляющая интерес аминокислотная последовательность обладает идентичностью 90% или более с референсной аминокислотной последовательностью, когда указанная представляющая интерес аминокислотная последовательность обладает аминокислотной идентичностью по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 99,5% с указанной референсной аминокислотной последовательностью. Для определения процента идентичности между двумя аминокислотными последовательностями в целях оптимального сравнения аминокислотные последовательности выравнивают. Например, для оптимального выравнивания бреши можно вводить в одну или в обе из первой и второй аминокислотных последовательностей, и в целях сравнения негомологичные последовательности можно не рассматривать. В целях оптимального сравнения процент идентичности двух аминокислотных последовательностей может быть получен с помощью программы CLUSTAL W (версия 1.82) с приведенными ниже параметрами: (1) CPU MODE = ClustalW mp; (2) ALIGNMENT = "full"; (3) OUTPUT FORMAT = "alnGAP OPEN = "default"; (14) END GAPS = "default"; (15) GAP EXTENSION = "default"; (16) GAP DISTANCES = "default"; (17) TREE TYPE = "cladogram" и (18) TREE GRAP DISTANCES = "hide". В частности, понятно, что могут быть проведены минорные модификации без нарушения преимуществ и активности пептидного производного флагеллина по изобретению. Такие модификации включены в значение терминов "иммуноадъювантное соединение" или "пептидное производное флагеллина" по изобретению до той степени, пока сохранена конкретная иммунная активность, в частности TLR6-опосредованные свойства мукозального адъюванта без какого-либо значительного системного провоспалительного ответа. Кроме того, различные молекулы могут быть ковалентно или нековалентно присоединены к пеп-6 019138 тидному производному флагеллина по изобретению, включая, например, другие полипептиды, углеводы,нуклеиновые кислоты или липиды. Эти присоединенные молекулы возможно состоят из антигена, против которого подразумевается данный иммунный ответ. Такие модификации включены в определение изобретения. Минорные модификации могут также касаться, например, консервативных замен встречающихся в природе аминокислот, а также структурные изменения, которые включают неприродные аминокислоты,аминокислотные аналоги и функциональные миметики. Например, аминокислотный остаток лизина считают консервативной заменой для аминокислотного остатка аргинина. Таким образом, как подразумевают здесь, первый полипептид, обладающий по меньшей мере 90% аминокислотной идентичностью со вторым референсным полипептидом, охватывает первые полипептиды, содержащие одно или более чем одно аминокислотное различие по сравнению со вторым референсным полипептидом, и где указанные аминокислотные различия выбраны из группы, состоящей из (1) замены одной или более чем одной аминокислоты, (2) делеции одной или более чем одной аминокислоты и (3) добавления одной или более чем одной аминокислоты, или любой комбинации (1), (2) и (3). В целом изобретение, таким образом, охватывает варианты полипептидов, имеющих одну или более чем одну аминокислотную замену, делецию или добавление по сравнению с референсным полипептидом, предпочтительно замену 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот и/или делецию 1, 2, 3, 4, 5, 6, 7,8, 9 или 10 аминокислот и/или добавление 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот по сравнению с референсным полипептидом. Специалисты в данной области техники знают или могут определить, какая структура составляет функционально эквивалентные аминокислотные аналоги и аминокислотные миметики. Как упомянуто выше, С-концевой и N-концевой пептиды пептидного производного флагеллина по изобретению могут быть связаны непосредственно, предпочтительно ковалентно, пептидной связью. В альтернативном воплощении указанные N-концевой и С-концевой пептиды пептидного производного флагеллина по изобретению связаны друг с другом опосредованно через спейсерную цепь. Спейсерная цепь должна быть выбрана так, чтобы не препятствовать биологической активности конечного соединения, а также так, чтобы иммуногенность конечного соединения значительно не повышалась. Спейсерная цепь предпочтительно состоит из аминокислот, связанных вместе пептидными связями,и присоединена ковалентно между N-концевой и С-концевой последовательностью пептидного производного флагеллина по изобретению. Таким образом, в предпочтительных воплощениях спейсерная цепь содержит от 1 до 20 аминокислот, связанных пептидными связями, где аминокислоты выбраны из 20 природных аминокислот. В более предпочтительном воплощении указанные от 1 до 20 аминокислот выбраны из Gly, Ala, Pro, Asn, Gln, Cys, Lys. Даже более предпочтительно спейсерная цепь состоит из последовательности NH2-Gly-Ala-Ala-Gly-COOH. Непептидные линкеры также возможны: например, алкильные линкеры. Эти алкильные линкеры могут быть дополнительно замещены любой стерически не затрудненной группой, низшим ацилом, галогеном, CN, NH2, фенилом и так далее. Другим типом непептидного линкера является полиэтиленгликолевая группа. Специалисту в данной области техники хорошо известны эти спейсерные цепи, и он может выбрать подходящую спейсерную цепь, конкретно зависящую от последовательностей N-концевого пептида и Сконцевого пептида, которые ему нужно связать друг с другом. Кроме того, аспарагиновый аминокислотный остаток С-концевой последовательности, локализованный в положении аминокислоты 488 SEQ ID NO:1, предпочтительно заменен сериновым остатком. Эта замена введена для специфичного мечения пептидного производного флагеллина по изобретению. Такая замена встречается в природе в флагеллинах других видов бактерий, таких как Legionnelapneumophila, без изменения TLR5-стимулирующей активности. Другие замены могут быть введены в положениях, которые не изменяют адъювантную TLR5-стимулирующую активность, для дополнительного мечения пептидного производного флагеллина по изобретению. Предпочтительные воплощения пептидного производного флагеллина по изобретению В соответствии с предпочтительными воплощениями, в свете пептидной последовательности флагеллина SEQ ID NO:1 и кристаллографической структуры, N-концевой пептид иммуноадъювантного соединения по изобретению предпочтительно выбран из группы, состоящей из аминокислотных последовательностей 1-99, 1-137, 1-160 и 1-173 SEQ ID NO:1. В частности, трехмерная (3D) структура флагеллина FliC показывает, что N-концевой домен организован в виде 3 альфа-спиралей, разделенных бета-витками, за которыми следуют бета-складчатые слои и бета-витки. Остальная(ые) часть(и) этих вторичных структур на N-конце может(гут) быть достаточной(ыми) для сохранения TLR5-стимулирующей активности (и, в частности, мукозальной TLR5 стимулирующей активности), то есть аминокислотные последовательности 1-99 SEQ ID NO:1 содержат первые 2 альфа-спирали, аминокислотные последовательности 1-137 SEQ ID NO:1 содержат первые 3 альфа-спирали, и аминокислотные последовательности 1-173 SEQ ID NO:1 содержат N-концевые структуры, обнаруженные в флагеллине FliC174-400.-7 019138 В следующих предпочтительных воплощениях указанный С-концевой пептид иммуноадъювантного соединения выбран из группы, состоящей из аминокислотных последовательностей 401-494 и 406-494SEQ ID NO:1. В частности, 3D структура флагеллина FliC показывает, что С-концевой домен организован в виде 2 альфа-спиралей, разделенных бета-витками. Остальная(ые) часть(и) этих вторичных структур на N-конце может(гут) быть достаточной(ыми) для сохранения TLR5-стимулирующей активности (и, в частности,мукозальной TLR5-стимулирующей активности): аминокислотные последовательности 401-494 SEQ IDNO:1 представляют собой последовательности, обнаруженные в флагеллине FliC174-400, тогда как аминокислотные последовательности 406-494 SEQ ID NO:1 содержат только две вторичные С-концевые альфаспирали. В некоторых предпочтительных воплощениях N-концевой пептид иммуноадъювантного соединения по изобретению состоит из аминокислотной последовательности, начинающейся с остатка аланина,локализованного в положении 1 SEQ ID NO:1, и оканчивающейся аминокислотным остатком, локализованным в положении SEQ ID NO:1, выбранным из группы, состоящей из аминокислотных остатков, локализованных в положениях 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152,153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172 и 173. В некоторых предпочтительных воплощениях С-концевой пептид иммуноадъювантного соединения по изобретению состоит из аминокислотной последовательности, начинающейся с аминокислотного остатка, локализованного в положении SEQ ID NO:1, выбранном из группы, состоящей из аминокислотных остатков, локализованных в положениях 401, 402, 403, 404, 405 и 406, и оканчивающейся остатком аргинина, локализованным в положении 494 SEQ ID NO:1. В конкретном аспекте этих предпочтительных воплощений указанный N-концевой пептид и указанный С-концевой пептид иммуноадъювантного соединения по изобретению предпочтительно связаны друг с другом посредством вышеупомянутой спейсерной цепи NH2-Gly-Ala-Ala-Gly-COOH (то есть при замене делетированной последовательности 174-400); аспарагиновый аминокислотный остаток, локализованный в положении 488 SEQ ID NO:1, также предпочтительно заменен сериновым остатком. Иллюстративные воплощения таких иммуноадъювантных соединений, описанных выше, охватывают FliC174-400, FliC161-405 и FliC138-405, которые показаны в приведенных здесь примерах; и которые также более подробно описаны ниже. Еще в одном следующем воплощении указанные N-концевой и С-концевой пептиды интересующего иммуноадъювантного соединения состоят из аминокислотных последовательностей 1-173 и 401-494SEQ ID NO:1 соответственно. В еще одном воплощении указанные N-концевой и С-концевой пептиды интересующего иммуноадъювантного соединения состоят из аминокислотных последовательностей 1-160 и 406-494 SEQ IDNO:1 соответственно. В еще одном воплощении указанные N-концевой и С-концевой пептиды интересующего иммуноадъювантного соединения состоят из аминокислотных последовательностей 1-137 и 406-494 SEQ IDNO:1 соответственно. В некоторых воплощениях иммуноадъювантные соединения в соответствии с изобретением содержат дополнительный остаток метионина на их N-конце, в частности, когда эти соединения продуцируют в виде рекомбинантных белков в бактериальных клетках. В воплощении, где указанные N-концевой и С-концевой пептиды интересующего иммуноадъювантного соединения состоят из аминокислотных последовательностей 1-173 и 401-494 SEQ ID NO:1,пептидное производное флагеллина по изобретению состоит из аминокислотной последовательностиSEQ ID NO:1, из которой делетирована аминокислотная последовательность, продолжающаяся от положения аминокислоты 174 до положения аминокислоты 400. Эту пептидную последовательность флагеллина по изобретению также называют в настоящем описании "FliC174-400" или "флагеллин FliC174-400". В соответствии с предпочтительным воплощением указанный N-концевой пептид и указанный Сконцевой пептид иммуноадъювантного соединения по изобретению предпочтительно связаны друг с другом посредством вышеупомянутой спейсерной цепи NH2-Gly-Ala-Ala-Gly-COOH (то есть при замене делетированной последовательности 174-400); аспарагиновый аминокислотный остаток, локализованный в положении 488 SEQ ID NO:1, также предпочтительно заменен сериновым остатком. Пептидное производное флагеллина по изобретению, полученное таким образом, представляет собой последовательность из 271 аминокислот, где пептидная последовательность состоит из SEQ ID NO:2. Нумерация полипептида начинается с первой аминокислоты после возможного N-концевого метионина (не показанного в SEQ ID NO:2), который типично отсекается метионинаминопептидазой в бактериальных клетках-хозяевах, как раскрыто ниже. В воплощении, где указанные N-концевой и С-концевой пептиды интересующего иммуноадъювантного соединения состоят из аминокислотных последовательностей 1-160 и 406-494 SEQ ID NO 1,пептидное производное флагеллина по изобретению состоит из аминокислотной последовательностиSEQ ID NO 1, из которой делетирована аминокислотная последовательность, продолжающаяся от поло-8 019138 жения аминокислоты 161 до положения аминокислоты 405. Эту пептидную последовательность флагеллина по изобретению также называют в настоящем описании "FliC161-405" или "флагеллин FliC161-405". В соответствии с предпочтительным воплощением указанный N-концевой пептид и указанный Сконцевой пептид иммуноадъювантного соединения по изобретению предпочтительно связаны друг с другом посредством вышеупомянутой спейсерной цепи NH2-Gly-Ala-Ala-Gly-COOH (то есть при замене делетированной последовательности 161-405), аспарагиновый аминокислотный остаток, локализованный в положении 488 SEQ ID NO 1, также предпочтительно заменен сериновым остатком. Пептидное производное флагеллина по изобретению, полученное таким образом, представляет собой последовательность из 253 аминокислот, где пептидная последовательность состоит из SEQ ID NO 25. Нумерация полипептида начинается с первой аминокислоты после возможного N-концевого метионина (не показанного в SEQ ID NO 25), который типично отсекается метионинаминопептидазой в бактериальных клетках-хозяевах, как раскрыто ниже. В воплощении, где указанные N-концевой и С-концевой пептиды интересующего иммуноадъювантного соединения состоят из аминокислотных последовательностей 1-137 и 406-494 SEQ ID NO 1,пептидное производное флагеллина по изобретению состоит из аминокислотной последовательностиSEQ ID NO 1, из которой делетирована аминокислотная последовательность, продолжающаяся от положения аминокислоты 138 до положения аминокислоты 405. Эту пептидную последовательность флагеллина по изобретению также называют в настоящем описании "FliC138-405" или "флагеллин FliC138-405". В соответствии с предпочтительным воплощением указанный N-концевой пептид и указанный Сконцевой пептид иммуноадъювантного соединения по изобретению предпочтительно связаны друг с другом посредством вышеупомянутой спейсерной цепи NH2-Gly-Ala-Ala-Gly-COOH (то есть при замене делетированной последовательности 138-405); аспарагиновый аминокислотный остаток, локализованный в положении 488 SEQ NO:1, также предпочтительно заменен сериновым остатком. Пептидное производное флагеллина по изобретению, полученное таким образом, представляет собой последовательность из 230 аминокислот, где пептидная последовательность состоит из SEQ IDNO:26. Нумерация полипептида начинается с первой аминокислоты после возможного N-концевого метионина (не показанного в SEQ ID NO:26), который типично отсекается метионинаминопептидазой в бактериальных клетках-хозяевах, как раскрыто ниже. В воплощении, где указанные N-концевой и С-концевой пептиды интересующего иммуноадъювантного соединения состоят из аминокислотных последовательностей 1-99 и 406-494 SEQ ID NO:1, пептидное производное флагеллина по изобретению состоит из аминокислотной последовательности SEQID NO:1, из которой делетирована аминокислотная последовательность, продолжающаяся от положения аминокислоты 100 до положения аминокислоты 405. Эту пептидную последовательность флагеллина по изобретению также называют в настоящем описании FliC100-405" или "флагеллин FliC100-405". В соответствии с предпочтительным воплощением указанный N-концевой пептид и указанный Сконцевой пептид иммуноадъювантного соединения по изобретению предпочтительно связаны друг с другом посредством вышеупомянутой спейсерной цепи NH2-Gly-Ala-Ala-Gly-COOH (то есть при замене делетированной последовательности 100-405); аспарагиновый аминокислотный остаток, локализованный в положении 488 SEQ ID NO:1, также предпочтительно заменен сериновым остатком. Пептидное производное флагеллина по изобретению, полученное таким образом, представляет собой последовательность из 192 аминокислот, где пептидная последовательность состоит из SEQ IDNO:27. Нумерация полипептида начинается с первой аминокислоты после возможного N-концевого метионина (не показанного в SEQ ID NO:27), который типично отсекается метионинаминопептидазой в бактериальных клетках-хозяевах, как раскрыто ниже. Синтез иммуноадъювантного пептида по изобретению Пептидное производное флагеллина по изобретению может быть синтезировано рекомбинантными клетками, полученными с помощью генной инженерии, или любым из способов химического или ферментативного пептидного синтеза, которые хорошо известны специалистам в данной области техники. 1. Синтез рекомбинантными клетками. Пептидное производное флагеллина согласно изобретению можно продуцировать рекомбинантным путем посредством рекомбинантных клеток, которые трансфицированы нуклеиновой кислотой, которая кодирует его аминокислотную последовательность и дает возможность его эффективного продуцирования внутри трансфицированных клеток. Последовательность нуклеиновой кислоты, кодирующая пептидное производное флагеллина по изобретению Модификации указанной пептидной последовательности флагеллина могут быть получены с использованием методик мутагенеза рекомбинантной ДНК. Различные способы конструирования и модификации последовательности ДНК известны специали-9 019138 стам в данной области техники, и выбор указанных рекомбинантных методов известен специалистам в данной области техники. Методики "рекомбинантного мутагенеза" включают, например, сайт-направленный мутагенез и ПЦР-мутагенез (см., в частности, Current Protocols in Molecular Biology, 2007 by John Wiley and Sons, Inc.,Chapter 8 and 15). Указанная полимеразная цепная реакция (ПЦР) особенно полезна для широкого ряда мутационных методик и применений. Методики ПЦР-мутагенеза дают возможность модифицировать и конструировать любую целевую ДНК легко и эффективно. Они включают, например, введение точечных мутаций,делеций или инсерций. Эти методы применяют, например, на гене дикого типа fliC SEQ ID NO:3, выделенном из штаммовS. Typhimurium ATCC14028, который кодирует пептид флагеллин, идентифицированный в SEQ ID NO:1. В предпочтительном воплощении вышеупомянутый ген fliC делетируют по центральному участку его длины путем ПЦР-мутагенеза (см., в частности, Current Protocols in Molecular Biology, 2007 by JohnWiley and Sons, Inc., Chapter 8 and 15), используя подходящие пары праймеров, выбранных в зависимости от желаемых N-концевой и С-концевой последовательностей, которые найдены для пептида по изобретению. Например, на основе плазмиды, имеющей происхождение от pBR322, несущей указанный ген fliC дикого типа SEQ ID NO:3 под контролем его собственного промотора, в методике ПЦР-мутагенеза можно использовать приведенные ниже пары праймеров:SEQ ID NO:4 и SEQ ID NO:5 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-99 и 401-494 SEQ ID NO:1 соответственно;SEQ ID NO:4 и SEQ ID NO:6 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-99 и 406-494 SEQ ID NO:1 соответственно;SEQ ID NO:7 и SEQ ID NO:5 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-137 и 401-494 SEQ ID NO:1 соответственно;SEQ ID NO:7 и SEQ ID NO:6 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-137 и 406-494 SEQ ID NO:1 соответственно;SEQ ID NO:8 и SEQ ID NO:5 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-160 и 401-494 SEQ ID NO:1 соответственно;SEQ ID NO:8 и SEQ ID NO:6 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-160 и 406-494 SEQ ID NO:1 соответственно;SEQ ID NO:9 и SEQ ID NO:5 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-173 и 401-494 SEQ ID NO:1 соответственно;SEQ ID NO:9 и SEQ ID NO:6 для N-концевого и С-концевого пептидов, состоящих из аминокислотных последовательностей 1-173 и 406-494 SEQ ID NO:1 соответственно. Чтобы заменить аспарагин в положении 488 SEQ ID NO:1 на серин, например, можно использовать сайт-направленный мутагенез с приведенными ниже праймерами SEQ ID NO:10 и SEQ ID NO:11. Чтобы ввести линкер NH2-Gly-Ala-Ala-Gly-COOH в точке соединения 1-99, 1-137, 1-160 или 1-173 с 401-494 или 406-494 рекомбинантных пептидов флагеллина, следующую последовательность ДНК:SEQ ID NO:12 и F-линкер-406" последовательности SEQ ID NO:13. Последовательностью ДНК, подходящей для продуцирования пептидного производного флагеллина по изобретению FliC174-400, является, например, последовательность SEQ ID NO:14. Нуклеиновая кислота, подходящая для продуцирования пептидного производного флагеллина по изобретению FliC161-405, имеет, например, последовательность SEQ ID NO:28. Нуклеиновая кислота, подходящая для продуцирования пептидного производного флагеллина по изобретению FliC138-405, имеет, например, последовательность SEQ ID NO:29. Нуклеиновая кислота, подходящая для продуцирования пептидного производного флагеллина по изобретению FliC100-405, имеет, например, последовательность SEQ ID NO:30. Выбор и использование реплицируемого вектора Последовательность нуклеиновой кислоты, раскрытую в данной заявке, кодирующую интересующее пептидное производное флагеллина, можно встраивать в реплицируемый вектор для клонирования(амплификации ДНК) или для экспрессии. Различные векторы широко доступны. Вектор может находиться, например, в форме плазмиды,космиды, вирусной частицы или фага. Соответствующую последовательность нуклеиновой кислоты можно встраивать в вектор с помощью ряда методик. Как правило, ДНК встраивают в соответствующий сайт(ы) рестрикционной эндонуклеазы, используя методики, известные в данной области техники. Компоненты вектора обычно включают, но не ограничены ими, одно или более чем одно из следующего: сигнальная последовательность, если последовательность нужно секретировать, точка начала репликации, один или более чем один маркерный ген, энхансерный элемент, промотор и последователь- 10019138 ность терминации транскрипции. При конструировании подходящих векторов, содержащих один или более чем один из этих компонентов, используют стандартные методики лигирования, которые известны специалистам в данной области техники. Интересующее пептидное производное флагеллина можно продуцировать рекомбинантным путем не только непосредственно, но также в виде слитого полипептида с гетерологичным полипептидом, который может представлять собой сигнальную последовательность или другой полипептид, имеющий специфичный сайт расщепления на N-конце зрелого белка или пептида. Как правило, сигнальная последовательность может быть компонентом вектора, либо она может составлять часть ДНК, кодирующей интересующий полипептид, который встроен в вектор. Сигнальная последовательность может представлять собой прокариотическую сигнальную последовательность, выбранную, например, из группы лидерных последовательностей щелочной фосфатазы, пенициллиназы, Ipp или термостабильного энтеротоксина II. Для дрожжевой секреции сигнальная последовательность может представлять собой, например,лидерную последовательность инвертазы дрожжей, лидерную последовательность альфа-фактора (включая лидерные последовательности альфа-фактора Saccharomyces и Kluyveromyces; последняя описана в патенте США 5010182) или лидерную последовательность кислой фосфатазы, лидерную последовательность глюкоамилазы С. albicans (ЕР 362179, опубликованный 4 апреля 1990) или сигнальную последовательность, раскрытую в WO 90/13646, опубликованной 15 ноября 1990). При экспрессии в клетках млекопитающих можно использовать сигнальные последовательности млекопитающих, чтобы направлять секрецию белка, такие как сигнальные последовательности из секретируемых полипептидов того же или родственного вида, а также вирусные секреторные лидерные последовательности. Как экспрессионные, так и клонирующие векторы содержат последовательность нуклеиновой кислоты, которая дает возможность вектору реплицироваться в одной или более чем одной из выбранных клеток-хозяев. Такие последовательности хорошо известны для ряда бактерий, дрожжей и вирусов. Точка начала репликации из плазмиды pBR322 подходит для большинства грамотрицательных бактерий,точка начала репликации 2-микронной плазмиды подходит для дрожжей, а различные вирусные точки начала репликации (SV40, полиомы, аденовируса, VSV или BPV) полезны для клонирующих векторов в клетках млекопитающих. Экспрессионные и клонирующие векторы типично содержат ген селекции, также называемый селективным маркером. Типичные гены селекции кодируют белки, которые (а) придают устойчивость к антибиотикам или другим токсинам, например к ампициллину, неомицину, метотрексату или тетрациклину, (б) восполняют ауксотрофные дефициты или (в) обеспечивают важные питательные вещества, недоступные из комплексных сред, например ген, кодирующий D-аланинрацемазу для Bacilli. Примерами подходящих селективных маркеров для клеток млекопитающих являются те, которые обеспечивают идентификацию клеток, компетентных к поглощению нуклеиновой кислоты, кодирующей интересующее пептидное производное флагеллина, такие как дигидрофолатредуктаза (DHFR) или тимидинкиназа. Подходящей клеткой-хозяином при использовании DHFR дикого типа является клеточная линия СНО с дефицитом по активности DHFR, которую получают и размножают как описано Urlaub etal., Proc. Natl. Acad. Sci. USA, 77: 4216 (1980). Подходящим селективным геном для использования в дрожжах является ген trp 1, присутствующий в дрожжевой плазмиде YRp7. Stinchcomb et al., Nature, 282: 39 (1979); Kingsman et al., Gene, 7: 141 (1979); Tschemper et al, Gene, 10: 157 (1980). Ген trp1 обеспечивает селективный маркер для мутантного штамма дрожжей в отсутствие способности роста при триптофане,например, АТСС No. 44076 или РЕР 4-1. Jones, Genetics, 85: 12 (1977). Экспрессионные и клонирующие векторы обычно содержат промотор, функционально связанный с последовательностью нуклеиновой кислоты, кодирующей пептидное производное флагеллина, для направления синтеза мРНК. Промоторы, распознаваемые рядом потенциальных клеток-хозяев, хорошо известны. Промоторы, подходящие для использования с прокариотическими хозяевами, включают промоторные системы бета-лактамазы и лактозы (Chang et al., Nature, 275: 615 (1978); Goeddel et al., Nature,281: 544 (1979, щелочной фосфатазы, промоторную систему триптофана (trp) (Goeddel, Nucleic AcidsRes., 8:4057 (1980); EP 36776) и гибридные промоторы, такие как промотор tac (deBoer et al., Proc. Natl.Acad. Sci. USA, 80: 21-25 (1983. Промоторы для использования в бактериальных системах также содержат последовательность Шайна-Далгарно (S.D.), функционально связанную с ДНК, кодирующей интересующее пептидное производное флагеллина. Примеры подходящих промоторных последовательностей для использования с дрожжевыми хозяевами включают промоторы 3-фосфоглицераткиназы (Hitzeman et al., J. Biol. Chem., 255: 2073 (1980 или других гликолитических ферментов (Hess et al., J. Adv. Enzyme Reg., 7: 149 (1968); Holland, Biochemistry,17: 4900 (1978, таких как енолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа,триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа. Другие дрожжевые промоторы, которые являются индуцибельными промоторами, обладающими дополнительным преимуществом транскрипции, контролируемой условиями роста, представляют собой промоторные области алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, ферментов расщеп- 11019138 ления, связанных с метаболизмом азота, металлотионеина, глицеральдегид-3-фосфатдегидрогеназы и ферментов, ответственных за утилизацию мальтозы и галактозы. Подходящие векторы и промоторы для использования при дрожжевой экспрессии дополнительно описаны в ЕР 73657. Транскрипцию интересующей нуклеиновой кислоты в клетках-хозяевах млекопитающих контролируют, например, промоторы, полученные из геномов вирусов, таких как вирус полиомы, вирус оспы кур(UK 2211504, опубликован 5 июля 1989), аденовирус (такой как аденовирус 2), вирус бычьей папилломы,вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и вакуолизирующий вирус обезьян 40 (SV40); гетерологичные промоторы млекопитающих, например промотор актина или промотор иммуноглобулина; и промоторы теплового шока, при условии, что такие промоторы совместимы с системами клетки-хозяина. Транскрипцию ДНК, кодирующей интересующее пептидное производное флагеллина, высшими эукариотами можно повысить путем инсерции энхансерной последовательности в вектор. Энхансеры представляют собой цис-активные элементы ДНК, обычно примерно от 10 до 300 пар оснований (п.о.),которые действуют на промотор с повышением его транскрипции. В настоящее время известны многие энхансерные последовательности из генов млекопитающих (глобина, эластазы, альбумина, альфафетопротеина и инсулина). Однако типично используют энхансер из вируса эукариотической клетки. Примеры включают энхансер SV40 на поздней стороне точки начала репликации (п.о. 100-270), энхансер раннего промотора цитомегаловируса, энхансер вируса полиомы на поздней стороне точки начала репликации и аденовирусные энхансеры. Энхансер может быть сплайсирован в векторе в положении 5' или 3' к последовательности, кодирующей интересующие полипептиды, но предпочтительно локализован с 5'-стороны от промотора. Экспрессионные векторы, используемые в эукариотических клетках-хозяевах (дрожжей, грибов,насекомых, растений, животных, человека или нуклеированных клетках из других многоклеточных организмов), также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно доступны из 5'- и иногда 3'-нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты,транскрибируемые в виде полиаденилированных фрагментов в нетранслируемом участке мРНК, кодирующей интересующее пептидное производное флагеллина. Другие способы, векторы и клетки-хозяева, подходящие для адаптации к синтезу интересующего пептидного производного флагеллина в культуре рекомбинантных клеток позвоночных, описаны вGething et al., Nature, 293: 620-625 (1981); Mantei et al., Nature, 281: 40-46 (1979); EP 117060 и ЕР 117058. Выбор и трансформация клеток-хозяев Клетки-хозяева трансфицируют или трансформируют экспрессионными или клонирующими векторами, описанными в данной заявке, для продуцирования пептидного производного флагеллина и культивирования в традиционной питательной среде, соответствующим образом модифицированной для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих желаемые последовательности. Условия культивирования, такие как среды, температура, рН и тому подобное, могут быть выбраны специалистом в данной области техники без излишнего экспериментирования. В целом принципы, протоколы и практические методики для максимизации продуктивности клеточных культур можно найти вMammalian Cell Biotechnology: A Practical Approach, M. Butler, ed. (IRL Press, 1991). Способы трансфекции известны обычным специалистам в данной области техники, например обработка СаРО 4 и электропорация. В зависимости от используемой клетки-хозяина трансформацию осуществляют, используя стандартные методики, пригодные для таких клеток. Кальциевую обработку с использованием хлорида кальция, как описано в Sambrook et al., см. выше, или электропорацию используют, как правило, для прокариот или других клеток, которые содержат существенные барьеры клеточной стенки. Инфицирование Agrobacterium tumefaciens используют для трансформации определенных растительных клеток, как описано Shaw et al., Gene 23: 315 (1983) и WO 89/05859, опубликованной 29 июня 1989 года. Для клеток млекопитающих без клеточных стенок можно использовать метод кальцийфосфатной преципитации Graham and van der Eb, Virology, 52:456-457 (1978). Общие аспекты трансформации системы клеток-хозяев млекопитающих описаны в патенте США 4399216. Трансформации в дрожжи типично осуществляют в соответствии со способом Van Solingen et al., J. Bact., 130: 946 (1977) иHsiao et al., Proc. Natl. Acad. Sci. (USA), 76: 3829 (1979). Однако можно также использовать другие способы введения ДНК в клетки, как, например, путем микроинъекции в ядро, электропорации, слияния бактериальных протопластов с интактными клетками или поликатионами, например, полибреном или полиорнитином. Для различных методик трансформации клеток млекопитающих см. Keown et al., Methods in Enzymology, 185: 527-537 (1990) и Mansouret al., Nature, 336: 348-352 (1988). Подходящие клетки-хозяева для клонирования или экспрессии ДНК в векторах в данной заявке включают клетки прокариот, дрожжей или высших эукариот. Подходящие прокариоты включают, но не ограничены ими, эубактерии, такие как грамотрицательные или грамположительные организмы, например Enterobacteriaceae, такие как Е. coli. Различные штаммы Е. coli находятся в открытом доступе, такие как штамм Е. coli K12 ММ 294 (АТСС 31446); Е. coli- 12019138 Х 1776 (АТСС 31537); штамм Е. coli W3110 (АТСС 27325); и К 5772 (АТСС 53635). Другие подходящие прокариотические клетки-хозяева включают Enterobacteriaceae, такие как Escherichia, например Е. coli,Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, напримерSerratia marcescans, и Shigella, а также Bacilli, такие как В. subtilis и В. licheniformis (например В. licheniformis 41P, раскрытый в DD 266710, опубликованной 12 апреля 1989), Pseudomonas, такие как P. aeruginosa, и Streptomyces. Эти примеры скорее являются иллюстративными, чем ограничивающими. Штамм SIN41 Salmonella typhimurium (fliC fljB) представляет особый интерес для продуцирования пептидного производного флагеллина, поскольку эти прокариотические клетки-хозяева не секретируют никаких флагеллинов (Proc Natl Acad Sci USA. 2001;98:13722-7). Однако флагеллины секретируются посредством специализированной системы секреции: так называемой "системы секреции типа III". Интересно, что штамм SIN41 продуцирует все компоненты системы секреции типа III, необходимой для оптимальной секреции флагеллина. Клонирование последовательности, кодирующей новые пептиды флагеллина под промотором fliC, обеспечивает секрецию в больших количествах интересующих пептидных производных флагеллина в штамме SIN41. Штамм W3110 также представляет интерес, поскольку он является распространенным штаммомхозяином для промышленных ферментации рекомбинантных ДНК. Предпочтительно клетка-хозяин секретирует минимальные количества протеолитических ферментов. Например, штамм W3110 можно модифицировать путем осуществления генетической мутации в генах, кодирующих белки, эндогенные для хозяина, где примеры таких хозяев включают штамм Е. coli W3110 1 А 2, который имеет полный генотипtonA; штамм Е. coli W3110 9 Е 4, который имеет полный генотип tonA ptr3; штамм Е. coli W3110 27 С 7(АТСС 55244), который имеет полный генотип tonA ptr3 phoA E15 (argF-lac)169 degP ompT kan.sup.r; штамм E. coli W3110 37D6, который имеет полный генотип tona ptr3 phoA E15 (argF-lac)169 degP ompTrbs7 ilvG kan.sup.r; штамм E. coli W3110 40B4, который представляет собой штамм 37D6 с неустойчивой к канамицину делеционной мутацией degP; и штамм Е. coli, имеющий мутантную периплазматическую протеазу, раскрытый в патенте США 4946783, опубликованном 7 августа 1990 года. Штаммы Е. coliMG1655, MG1655 fimA-H или MKS12, fliD- и -fimA-H-делетированный штамм MG1655 также являются интересными кандидатами для продуцирования рекомбинантных флагеллинов в виде секретируемых белков (Nat Biotechnol. 2005; (4):475-81). Альтернативно подходят способы клонирования in vitro, например ПЦР или другие полимеразные реакции нуклеиновых кислот. Кроме прокариот, эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи,являются подходящими хозяевами для клонирования или экспрессии векторов, кодирующих пептидное производное флагеллина.Saccharomyces cerevisiae являются общепринято используемым низшим эукариотическим микроорганизмом-хозяином. Другие включают Schizosaccharomyces pombe (Beach and Nurse, Nature, 290: 140pastons (EP 183070, Sreekrishna et al, J Basic Microbiol , 28 265-278 [1988]), Candida, Tnchoderma reesia (EP 244234), Neurospora crassa (Case et al, Proc Natl Acad Sci USA, 76 5259-5263 [1979]), Schwanniomyces, такие как Schwanniomyces occidentalis (EP 394538, опубликованный 31 октября 1990 года), и мицелиальные грибы, такие как, например, хозяева Neurospora, Penicillium, Tolypocladium (WO 91/00357, опубликованная 10 января 1991 года) и Aspergillus, такие как A nidulans (Ballance et al., Biochem Biophys Res Commun,112 284-289 [1983], Tilburn et al., Gene, 26 205-221 [1983], Yelton et al., Proc Natl Acad Sci USA, 81 14701474 [1984]) и A niger (Kelly and Hynes, EMBO J, 4 475-479 [1985]) Метилотрофные дрожжи являются подходящими в данном изобретении и включают, но не ограничены ими, дрожжи, способные к росту на метаноле, выбранные из родов, состоящих из Hansenula, Candida, Kloeckera, Pichia, Saccharomyces, Torulopsis и Rhodotorula Перечень конкретных видов, которые являются примерами данного класса дрожжей,можно найти в С Anthony, The Biochemistry of Methylotrophs, 269 (1982). Подходящие клетки-хозяева для экспрессии нуклеиновой кислоты, кодирующей интересующее пептидное производное флагеллина, имеют происхождение от многоклеточных организмов Примеры клеток беспозвоночных включают клетки насекомых, такие как Drosophila S2 и Spodoptera Sf9, а также клетки растений. Примеры полезных линий клеток-хозяев млекопитающих включают клетки яичника китайского хомячка (СНО) и клетки африканской зеленой мартышки (COS) Более конкретные примеры включают линию почек обезьяны CV1, трансформированную SV40 (COS-7, АТСС CRL 1651), линию эмбриональных почек человека (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Grahain et al., J Gen Virol, 36 59 (1977, клетки яичника китайского хомячка - DHFR (CHO,Urlaub and Chasm, Proc Natl Acad Sci USA, 77 4216 (1980, клетки Сертоли мыши (ТМ 4, Mather, Biol Reprod , 23 243-251 (1980, клетки легких человека (W138, АТСС CCL 75), клетки печени человека (HepG2, НВ 8065), и опухолевые клетки молочной железы мыши (ММТ 060562, АТСС CCL51) Выбор подходящей клетки-хозяина находится в пределах компетенции специалистов в данной области техники. Общие способы очистки интересующего пептидного производного флагеллина Формы интересующего пептидного производного флагеллина могут быть выделены из культуральной среды или из лизатов клеток-хозяев. Если эти производные связаны с мембраной, их можно освободить от мембраны, используя подходящий раствор детергента (например, ТРИТОН-Х.ТМ. 100), или путем ферментативного расщепления. Клетки, используемые в экспрессии нуклеиновой кислоты, кодирующей интересующее пептидное производное флагеллина, можно разрушить различными физическими или химическими способами, такими как цикл замораживания-оттаивания, обработка ультразвуком, механическое разрушение или агенты, лизирующие клетки. Может быть желательным очистить интересующий полипептид от рекомбинантных клеточных белков или полипептидов. Приведенные ниже методики являются примерами подходящих методик очистки: фракционирование на ионообменной колонке; осаждение этанолом; ВЭЖХ с обращенной фазой; хроматография на силикагеле или на катионообменной смоле, такой как диэтиламиноэтилцеллюлоза(DEAE); хроматографическое фокусирование; электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН-ПААГ); осаждение сульфатом аммония; гель-фильтрация с использованием, например, Sephadex G-75; колонки сефарозы с белком А для удаления загрязнителей, таких какIgG; и колонки, хелатирующие металлы, для связывания форм интересующего пептидного производного флагеллина с эпитопной меткой. Можно использовать различные способы очистки белка, и такие способы известны в данной области техники и описаны, например, в Deutscher, Methods in Enzymology, 182 (1990); Scopes, Protein Purification: Principles and Practice (Springer-Verlag: New York, 1982). Выбранная стадия(и) очистки зависит, например, от природы используемого способа продуцирования и от конкретного продуцируемого пептидного производного флагеллина. В предпочтительном воплощении пептидное производное флагеллина очищают из надосадочной жидкости рекомбинантного S. Typhimurium SIN41 (fliC fljB), как раскрыто в примерах. В частности, Salmonella выращивали в среде Лурия-Бертани (LB) в течение 6-18 ч при 37 С при встряхивании. Надосадочную жидкость фильтровали и насыщали 60% сульфатом аммония (Sigma Aldrich, USA). Осажденные вещества выделяли центрифугированием, солюбилизацией в 20 мМ Трис/HCl рН 7,5, а затем диализом. Белки дополнительно очищали посредством последовательных раундов хроматографии на гидроксиапатите, анионообменной и эксклюзионной хроматографии (Bio-Rad Laboratories,USA; GE Healthcare, Sweden). Наконец, белки истощали по липополисахариду (LPS), используя колонку с полимиксином В (Pierce, USA). Используя ЛАЛ-тест (анализ с использованием лизатов амебоцитовLimulus) (Associates of Cape Cod Inc., USA), определяли остаточную концентрацию LPS, так чтобы она была менее чем 30 пг LPS на мкг рекомбинантного флагеллина. Очистка интересующего пептидного производного флагеллина с использованием иммуноаффинной хроматографии В дополнительных воплощениях пептидное производное флагеллина согласно изобретению можно очищать путем разделения на иммуноаффинном хроматографическом субстрате. Указанный иммуноаффинный хроматографический субстрат содержит антитела антифлагеллин, которые иммобилизованы на нем. Под антителами "антифлагеллин" в данном описании подразумевают антитела, которые связаны либо с нативным флагеллином, либо с флагеллином с делетированной гипервариабельной областью, включая те, которые охватываются настоящим изобретением. Предпочтительно антитела анти-флагеллин состоят из моноклональных антител, включая мышиные антитела анти-флагеллин. В соответствии с изобретением показано, что антитела анти-флагеллин, которые получены способом, включающим стадию иммунизации мышей флагеллином FliC174-400 с делетированной гипервариабельной областью, который раскрыт в данном описании в других частях, распознают как нативный флагеллин, так и любой из флагеллинов с делетированной гипервариабельной областью, который раскрыт в данном описании. Таким образом, в некоторых предпочтительных воплощениях иммуноаффинного хроматографического субстрата указанный субстрат содержит мышиные моноклональные антитела, направленные против FliC174-400. Указанный предпочтительный иммуноаффинный хроматографический субстрат может быть получен, как описано ниже: асцит мыши, содержащий моноклональные антитела анти-FliC174-400, очищали на колонках с белком А Econo-Рас ( 732-2022 Affi-gel; Bio-Rad); полученные в результате очищенные моноклональные антитела анти-FliC174-400 (которые могут быть также названы "В 23 С 5") ковалентно связывали через первичные аминогруппы с активированной Nгидроксисукцинимидом высокоэффективной колонкой Sepharose ( 17-0716-01 Hitrap, NHS активированная HP от фирмы GE Healthcare), с образованием аффинной колонки, специфичной к флагеллину. Выход связывания составлял 98%.- 14019138 Как показано в примерах, приведенных в данном описании, указанная выше аффинная колонка,специфичная к флагеллину, дает возможность высокоспецифичного отделения нативного флагеллина и,следовательно, также любого из флагеллинов с делетированной гипервариабельной областью, которые раскрыты в данном описании, от других белковых компонентов или небелковых компонентов, содержащихся в исходном образце. Способ очистки флагеллина или любого из флагеллинов с делетированной гипервариабельной областью, которые раскрыты здесь, описан ниже: надосадочные жидкости, содержащие флагеллин, из культуры рекомбинантного штамма S. Typhimurium SIN41 или Е. coli, центрифугировали, фильтровали через 0,22 мкм мембрану, разбавляли один к одному связывающим буфером (75 мМ Трис-HCl с рН 8) и наносили на аффинную колонку, специфичную к флагеллину, описанную выше; затем эту аффинную колонку, специфичную к флагеллину, промывали 15-20 CV (объем колонки) связывающего буфера; далее, белки элюировали 3 CV буфера элюции (100 мМ глицин-HCl, 0,5 М NaCl, рН 2,7) и фракции немедленно нейтрализовали 500 мкл Трис 1,5 М рН 8,9 во избежание длительного воздействия кислого рН; затем колонку регенерировали 10 CV связывающего буфера и хранили при 4 С с 0,02% азидом натрия. Типичный хроматографический профиль представлен на фиг. 13, где изображены (1) кривая поглощения (OD) при 280 нм (линия с закрашенными квадратами), а также (2) кривая электропроводности. Номера со стрелками на фиг. 13 соответствуют периодам времени, когда фракции жидкости, вытекающей из колонки, успешно собраны с целью дальнейшего их анализа на содержание флагеллина (см. фиг. 14 и приведенный ниже параграф). Пронумерованные собранные фракции соответственно состоят из:1 - 5 мкл образца перед нанесением (3 мкг) равно суммарному внесенному количеству 900 мкг;2 - 20 мкл из нанесенного образца после прохождения колонки;3 - 20 мкл из промывки колонки;4 - 20 мкл из промывки колонки;5, 6 и 7 - 20 мкл из каждой из соответствующих фракций после буфера элюирования: суммарное количество примерно 900 мкг; 8 - 20 мкл после повторного уравновешивания колонки. На фиг. 14 изображена фотография Вестерн-блот-анализа, который проведен с использованием фракций 1-8, относящихся к фиг. 13, в качестве соответствующего исходного вещества. 2. Химический синтез. В некоторых воплощениях пептид по изобретению можно синтезировать посредством общепринятых методов химического пептидного синтеза. Например, последовательность интересующего производного пептида флагеллина может быть получена путем прямого пептидного синтеза, используя твердофазные методы, подобные описанным Stewart et al., Solid-Phase Peptide Synthesis (W.H. Freeman Co.: San Francisco, Calif., (1969); Merrifield, J. Am.Chem. Soc, 85: 2149-2154 (1963); Fields G.B., Noble R.L.; 1990; Int. J. Pept. Protein Res.; Vol. 35 : 161-214). Синтез белка in vitro можно осуществить, используя ручные способы или с помощью автоматизации. Автоматизированный синтез можно выполнять, например, с помощью пептидного синтезатора фирмы Applied Biosystems (Foster City, Calif.), используя инструкции изготовителя. Различные участки интересующего пептида могут быть химически синтезированы отдельно и объединены с использованием химических или ферментативных способов с получением полноразмерного интересующего пептида. Композиции, содержащие пептидное производное флагеллина по изобретению Следующий объект изобретения составляет композиция, в частности фармацевтическая композиция, содержащая адъювантное соединение, как оно определено в настоящем описании, в частности, в сочетании с одним или более чем одним фармацевтически приемлемым эксципиентом. Настоящее изобретение также относится к иммуногенной композиции, содержащей иммуноадъювантное соединение, как оно определено в настоящем описании, вместе с одним или более чем одним антигеном."Иммуногенная композиция", как только она введена субъекту или животному, вызывает защитный иммунный ответ против одного или более чем одного указанного антигена, который она содержит. Настоящее изобретение также относится к вакцинной композиции, содержащей иммуноадъювантное соединение, как оно определено в настоящем описании, вместе с одним или более чем одним антигеном. Как используют здесь, "вакцинная композиция", как только она введена субъекту или животному,вызывает защитный иммунный ответ против, например, микроорганизма, или чтобы эффективно защищать субъекта или животное против инфекции. Вакцинная композиция полезна для предупреждения или ослабления патологического состояния,которое даст благоприятный ответ на модулирование иммунного ответа. Иммуноадъювант Как упомянуто выше, термин "иммуноадъювант" при использовании со ссылкой на иммуногенную композицию или вакцину предназначен для обозначения вещества, которое в целом действует на ускорение, пролонгирование или улучшение качества специфичных иммунных ответов на антиген. Иммуноадъювант может предпочтительно также уменьшать число иммунизации или количество антигена, необходимое для защитной иммунизации. Антиген Ряд веществ можно использовать в качестве антигенов в композиции или препарате иммуногенного или вакцинного типа. Например, аттенуированные и инактивированные вирусные и бактериальные патогены, очищенные макромолекулы, полисахариды, анатоксины, рекомбинантные антигены, организмы,содержащие чужеродный ген из патогена, синтетические пептиды, полинуклеиновые кислоты, антитела и опухолевые клетки можно использовать для получения (1) иммуногенной композиции, полезной для индукции иммунного ответа у индивидуума, или (2) вакцины, полезной для лечения патологического состояния. Следовательно, пептидное производное флагеллина по изобретению можно объединять с широким рядом антигенов для получения иммуногенной композиции или вакцины, полезной для индукции иммунного ответа у индивидуума. Специалисты в данной области техники способны выбрать антиген, подходящий для лечения конкретного патологического состояния, и знают, как определить, необработанный или выделенный антиген является лучшим в конкретном вакцинном препарате. Специалисты в данной области техники также способны определить, связывать ли ковалентно или связывать нековалентно иммуноадъювант по изобретению с одним или более чем одним из указанных антигенов. В настоящих тестах in vivo продемонстрировано, что для мукозальной адъювантной активности не требуется какой-либо связи между интересующим пептидным производным флагеллина и антигеноммишенью, когда их вводят совместно мукозальным путем и, в частности, интраназальным путем. Выделенный антиген можно получить, используя ряд способов, хорошо известных в данной области техники. Ген, кодирующий любой иммуногенный полипептид, можно выделить и клонировать, например, в клетках бактерий, дрожжей, насекомых, рептилий или млекопитающих, используя рекомбинантные способы, хорошо известные в данной области техники и описанные, например, в Sambrook et al.,Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York (1992) и в Ansubel et al.,Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, MD (1998). Ряд генов, кодирующих поверхностные антигены из патогенов вирусов, бактерий и простейших, были успешно клонированы,экспрессированы и использованы в качестве антигенов для разработки вакцин. Например, основной поверхностный антиген вируса гепатита В, HbsAg, субъединица Р холерного токсина, энтеротоксин Е. coli,белок циркумспорозоита малярийного паразита и гликопротеиновый мембранный антиген из вируса Эпштейна-Барр, а также антигены опухолевых клеток экспрессированы в различных хорошо известных системах вектор/хозяин, очищены и использованы в вакцинах. Пептидное производное флагеллина по изобретению индуцирует наследственно-обусловленный иммунный ответ посредством TLR5-опосредованной мукозальной системы, которая может полезно усиливать иммунный ответ на рекомбинантный антиген. Патологически аберрантная клетка, которую нужно использовать в вакцине, может быть получена из любого источника, такого как один или более чем один индивидуум, страдающий патологическим состоянием, или клетки, культивируемые ex vivo или in vitro, полученные от одного или более чем одного такого индивидуума, включая конкретного индивидуума, подлежащего лечению полученной в результате вакциной. Иммуномодуляторные молекулы Ряд иммуномодуляторных молекул можно также использовать в комбинации с пептидным производным флагеллина по изобретению для изменения иммунного ответа у индивидуума. Тип желаемого изменения будет определять тип иммуномодуляторной молекулы, выбранной для комбинирования с указанным пептидным производным флагеллина по изобретению. Например, для усиления наследственно-обусловленного иммунного ответа пептидное производное флагеллина по изобретению можно объединять с другой иммуномодуляторной молекулой, которая стимулирует наследственно-обусловленный иммунный ответ, такой как другие РАМР или консервативная область, известная или предполагаемая в отношении индукции наследственно-обусловленного иммунного ответа. Известно, что ряд РАМР стимулирует активности различных членов toll-подобного семейства рецепторов. Такие РАМР можно объединять для стимулирования конкретной комбинации toll-подобных рецепторов, которые индуцируют благоприятный профиль цитокинов. Например, РАМР можно объединять для стимулирования профиля цитокинов, который индуцирует иммунный ответ Th1 или Th2. Другие типы иммуномодуляторных молекул, которые стимулируют гуморальный или клеточноопосредованный иммунные ответы, можно объединять с пептидным производным флагеллина по изо- 16019138 бретению. Например, можно вводить цитокины для изменения баланса иммунных ответов Th1 и Тh2. Специалистам в данной области техники известно, как определить подходящие цитокины, полезные для получения благоприятного изменения в иммунном ответе на конкретное патологическое состояние. Введение пептидного производного флагеллина по изобретению Пептидное производное флагеллина по изобретению вводят в "иммуногенном количестве" с одной или более чем одной молекулой, которое предназначено для обозначения количества, необходимого для запуска иммунного ответа, например, антигеном или другой иммуномодуляторной молекулой. Дозировка пептидного производного флагеллина по изобретению, независимо или в комбинации с одной или более чем одной молекулой, зависит, например, от патологического состояния, подлежащего лечению, от массы и состояния индивидуума и от предшествующих или сопутствующих терапий. Подходящее количество, которое считают иммуногенной дозой для конкретного применения способа, может быть определено специалистами в данной области техники. Специалистам в данной области техники понятно, что необходимо проводить мониторинг состояния пациента на протяжении всего курса терапии и что количество композиции, которое вводят, можно регулировать в соответствии с ответом пациента на терапию. В качестве вакцинного иммуноадъюванта пептидные производные флагеллина по изобретению могут вносить вклад в эффективность вакцины посредством, например, усиления иммуногенности более слабых антигенов, таких как высокоочищенные или рекомбинантные антигены, уменьшения количества антигена, требующегося для иммунного ответа, снижения частоты иммунизации, требующейся для обеспечения защитного иммунитета, улучшают эффективность вакцин у индивидуумов со сниженными или ослабленными иммунными ответами, таких как новорожденные, пожилые индивидуумы и индивидуумы с иммунной недостаточностью, и усиливают иммунитет в ткани-мишени, такой как мукозальный иммунитет, или стимулируют клеточно-опосредованный или гуморальный иммунитет, вызывая конкретный профиль цитокинов. Пептидное производное флагеллина по изобретению индуцирует наследственно-обусловленный иммунный ответ посредством активации системы TLR5, в частности, здесь TLR5-опосредованный мукозальный ответ при введении мукозальным путем. В частности, в тестах in vivo показано, что пептидное производное флагеллина по изобретению проявляет мукозальную адъювантную активность, которая способна потенцировать системные и мукозальные ответы против антигена-мишени. Наследственно-обусловленный иммунный ответ повышает иммунный ответ на антиген посредством стимуляции адаптивного иммунного ответа. Поэтому комбинация пептидного производного флагеллина по изобретению с одним или более чем одним антигеном обеспечивает эффективную иммуногенную композицию или вакцину для индукции иммунного ответа у индивидуума. Комбинацию антигена и/или иммуномодуляторной молекулы с пептидным производным флагеллина по изобретению можно тестировать в ряде доклинических токсикологических исследований и исследований на безопасность, хорошо известных в данной области техники. Например, такую комбинацию можно оценивать на животной модели, для которой обнаружено, что антиген является иммуногенным, и которую можно воспроизводимо иммунизировать тем же путем, который предложен для клинического испытания у людей. Комбинацию антигена и/или иммуномодуляторной молекулы с пептидным производным флагеллина по изобретению можно тестировать, например, методом, описанным Center for Biologies Evaluationand Research/Food and Drug Administration and National Institute of Allergy and Infectious Diseases (Goldenthal, K.L. et al. AID Res Hum Retroviruses, 9: S45-9 (1993. Специалистам в данной области техники известно, как определить конкретную комбинацию антигена и/или иммуномодуляторной молекулы с пептидным производным флагеллина по изобретению, подходящую полезную нагрузку антигена, путь введения, объем дозы, чистоту антигена и режим вакцинации, полезный для лечения конкретного патологического состояния у конкретного вида животного. Иммуногенную композицию или вакцину по изобретению для индукции иммунного ответа можно вводить в виде раствора или суспензии вместе с фармацевтически приемлемой средой. Такой фармацевтически приемлемой средой может быть, например, вода, фосфатно-солевой буферный раствор, нормальный физиологический раствор или другой физиологически забуференный солевой раствор, либо другой растворитель или носитель, такой как гликоль, глицерин и масло, такое как оливковое масло или инъекционный органический эфир. Фармацевтически приемлемая среда может также содержать липосомы или мицеллы и может содержать иммуностимулирующие комплексы, полученные путем смешивания полипептидных или пептидных антигенов с детергентом и гликозидом, таким как Quil A. Дополнительные способы получения и введения пептидного производного флагеллина по изобретению в фармацевтически приемлемой среде представлены ниже, со ссылкой на соединения, которые индуцируют TLR5-опосредованный мукозальный ответ. Иммуногенную композицию или вакцину по изобретению можно вводить разными путями для стимуляции иммунного ответа. Например, эти иммуномодуляторные молекулы могут быть доставлены- 17019138 подкожно, внутрикожно, внутрилимфатически, внутримышечно, внутрь опухоли, интравезикально,внутрибрюшинно и интрацеребрально. Специалистам в данной области техники известно, как выбрать соответствующие пути доставки для конкретных препаратов пептидных производных флагеллина по изобретению. В предпочтительном воплощении изобретения способы вакцинации для лечения или предупреждения инфекции у млекопитающего включают применение вакцины по изобретению, которую вводят, в частности, на поверхность слизистой оболочки (например, глазной, интраназальной, ротовой, желудочной, легочной, кишечной, ректальной, вагинальной или мочевых путей). Назальные пути доставки могут быть полезны для индукции как мукозального, так и системного иммунного ответа. Возможен ряд устройств для удобной и эффективной доставки препаратов в носовую полость и легочные ткани. Назальный путь доставки в данной заявке может представлять особый интерес, поскольку пептидное производное флагеллина по изобретению проявляет значительную адъювантную активность в компартменте слизистой оболочки, не обладая каким-либо значительным системным провоспалительным побочным эффектом. В протоколе вакцинации вакцину можно предпочтительно вводить мукозальным путем в виде единственной дозы или предпочтительно несколько раз, например два, три или четыре раза, через недельные или месячные интервалы в соответствии с режимом первичной/бустерной вакцинации. Подходящая дозировка зависит от различных параметров. Протокол вакцинации может представлять собой строго мукозальный протокол или смешанный протокол, при котором первичную дозу вакцины вводят мукозальным, например интраназальным путем,в бустерную дозу(ы) вводят парентерально, или наоборот. Препарат Способы получения фармацевтических препаратов или композиций включают стадию приведения активного ингредиента(ов) в контакт с носителем и, возможно, одним или более чем одним вспомогательным ингредиентом. Как правило, препараты готовят путем однородного и тесного приведения активного ингредиента(ов) в контакт с жидкими носителями, либо с тонко измельченными твердыми носителями, либо с теми и с другими, а затем при необходимости формования продукта. Жидкие лекарственные формы для перорального введения активных ингредиентов включают фармацевтически приемлемые эмульсии, микроэмульсии, растворы, суспензии, сиропы и эликсиры. В дополнение к активному ингредиенту(ам) жидкие лекарственные формы могут содержать инертные разбавители, обычно используемые в данной области техники, такие как, например, вода или другие растворители, солюбилизирующие агенты и эмульгаторы, такие как этиловый спирт, изопропиловый спирт,этилкарбонат, этилацетат, бензиловый спирт, бензилбензоат, пропиленгликоль, 1,3-бутиленгликоль, масла (в частности, хлопковое, арахисовое, кукурузное масло, масло зародышей пшеницы, оливковое, касторовое и кунжутное масло), глицерин, тетрагидрофуриловый спирт, полиэтиленгликоли и эфиры жирных кислот и сорбитана, а также их смеси. Кроме инертных разбавителей, пероральные композиции могут также включать адъюванты, такие как увлажняющие агенты, эмульгирующие и суспендирующие агенты, подсластители, корригенты, красители, ароматизаторы и консерванты. Суспензии, в дополнение к активному ингредиенту(ам), могут содержать суспендирующие агенты,такие как, например, этоксилированные изостеариловые спирты, полиоксиэтиленсорбит и эфиры сорбитана, микрокристаллическая целлюлоза, метагидроксид алюминия, бентонит, агар-агар и трагакант, а также их смеси. Препараты фармацевтических композиций по изобретению для ректального или вагинального введения могут быть представлены в виде суппозитория, который можно готовить путем смешивания активного ингредиента(ов) с одним или более чем одним подходящим не раздражающим эксципиентом или носителем, включающим, например, масло какао, полиэтиленгликоль, воск для суппозиториев или салицилат, и который является твердым при комнатной температуре, но жидким при температуре тела, и,следовательно, плавится в прямой кишке или в полости влагалища и высвобождает активный ингредиент(ы). Препараты по настоящему изобретению, которые подходят для вагинального введения, также включают препараты в виде пессариев, тампонов, кремов, гелей, паст, пенок или спреев, содержащие такие носители, которые известны в данной области техники как подходящие. Фармацевтические композиции по данному изобретению, подходящие для парентерального введения, содержат активный ингредиент(ы) в комбинации с одним или более чем одним фармацевтически приемлемым стерильным изотоническим водным или неводным раствором, дисперсией, суспензией или эмульсией, либо стерильными порошками, которые можно восстанавливать в стерильных инъекционных растворах или дисперсии непосредственно перед применением, которые могут содержать антиоксиданты, буферы, растворенные вещества, которые делают препарат изотоническим с кровью предназначенного реципиента, либо суспендирующие или загущающие агенты. Примеры подходящих водных и неводных носителей, которые можно использовать в фармацевти- 18019138 ческих композициях по изобретению, включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и тому подобное) и их подходящие смеси, растительные масла, такие как оливковое масло, и инъекционные органические эфиры, такие как этилолеат. Соответствующую текучесть можно поддерживать, например, путем использования материалов покрытия, таких как лецитин,путем поддержания требуемого размера частиц в случае дисперсий и путем использовании поверхностно-активных веществ. Эти композиции могут также содержать адъюванты, такие как увлажняющие агенты, эмульгирующие агенты и диспергирующие агенты. Может быть также желательно включать в композиции изотонические агенты, такие как сахара, хлорид натрия и тому подобное. Кроме того, пролонгированное всасывание инъекционной фармацевтической формы можно вызвать за счет включения агентов, которые замедляют всасывание, таких как моностеарат алюминия и желатин. Инъекционные депо-препараты готовят путем формования микроинкапсулирующих матриц активного(ых) ингредиента(ов) в биоразлагаемых полимерах, таких как полилактид-полигликолид. В зависимости от отношения активного(ых) ингредиента(ов) к полимеру и от природы конкретного используемого полимера можно регулировать скорость высвобождения активного(ых) ингредиента(ов). Примеры других биоразлагаемых полимеров включают поли(ортоэфиры) и поли(ангидриды). Инъекционные депо-препараты также готовят путем включения активного(ых) ингредиента(ов) в липосомы или микроэмульсии, которые совместимы с тканью организма. Инъекционные материалы можно стерилизовать, например, фильтрованием через фильтр, задерживающий бактерии. Препараты могут быть представлены в герметичных контейнерах, содержащих однократную дозу или многократные дозы, например, в ампулах и флаконах, и их можно хранить в лиофилизированном состоянии, требующем только добавления стерильного жидкого носителя, например воды для инъекций,непосредственно перед применением. Экстемпоральные инъекционные растворы и суспензии можно готовить из стерильных порошков, гранул и таблеток вышеописанного типа. Количество антигена и иммуноадъювантного соединения в вакцинной композиции в соответствии с изобретением, вводимые дозы определяют методами, хорошо известными специалистам в области фармацевтики, с учетом таких факторов, как конкретный антиген, возраст, пол, масса, вид и состояние конкретного животного или пациента, а также путь введения. В предпочтительном воплощении вакцинная композиция в соответствии с изобретением дополнительно содержит один или более чем один компонент, выбранный из группы, состоящей из поверхностно-активных веществ, стимуляторов всасывания, полимеров, поглощающих воду, веществ, которые ингибируют ферментативное расщепление, спиртов, органических растворителей, масел, агентов, регулирующих рН, консервантов, агентов, регулирующих осмотическое давление, пропеллентов, воды и их смесей. Вакцинная композиция в соответствии с изобретением может дополнительно содержать фармацевтически приемлемый носитель. Количество носителя будет зависеть от количеств, выбранных для других ингредиентов, желаемой концентрации антигена, выбора пути введения, перорального или парентерального, и т.д. Носитель можно добавлять к вакцине в любое удобное время. В случае лиофилизированной вакцины носитель можно, например, добавлять непосредственно перед применением. Альтернативно,конечный продукт можно готовить с носителем. Примеры подходящих носителей включают, но не ограничены ими, стерильную воду, физиологический раствор, буферы, фосфатно-солевой буферный раствор, забуференный хлорид натрия, растительные масла, минимальную эссенциальную среду (MEM), MEM с буфером HEPES и т.д. Возможно, вакцинная композиция по изобретению может содержать традиционные вторичные адъюванты в варьирующих количествах в зависимости от адъюванта и от желаемого результата. Общепринятое количество находится в интервале от примерно 0,02 до примерно 20% мас./мас., в зависимости от других ингредиентов и от желаемого эффекта. Примеры подходящих вторичных адъювантов включают, но не ограничены ими, стабилизаторы; эмульгаторы; гидроксид алюминия; фосфат алюминия; регуляторы рН, такие как гидроксид натрия, соляная кислота и т.д.; поверхностно-активные вещества, такие как Tween.RTM.80 (полисорбат 80, имеющийся в продаже от фирмы Sigma Chemical Co., St. Louis, Mo.); липосомы; иском-адъювант; синтетические гликопептиды, такие как мурамилдипептиды; разбавители, такие как декстран или комбинации декстрана, например, с фосфатом алюминия; карбоксиполиметилен; бактериальные клеточные стенки, такие как экстракт клеточной стенки микобактерий; их производные, такие как Corynebacterium parvum; Propionibacterium acne; Mycobacterium bovis, например, коровья бацилла Кальметта-Герена (БЦЖ); белки вируса осповакцины или поксвирусов животных; адъюванты субвирусных частиц, такие как орбивирус; холерный токсин; N,N-диоктадецил-N',N'-бис(2-гидроксиэтил)-пропандиамин (пиридин); монофосфориллипид А; диметилдиоктадециламмония бромид (DDA, имеющийся в продаже от фирмы Kodak, Rochester, N.Y.); синтетические адъюванты и их смеси. Желательно гидроксид алюминия смешивают с другими вторичными адъювантами или с иммуноадъювантом, таким как Quil A. Примеры подходящих стабилизаторов включают, но не ограничены ими, сахарозу, желатин, пептон, переваренные белковые экстракты, такие как NZ-амин или NZ-амин AS. Примеры эмульгаторов- 19019138 включают, но не ограничены ими, минеральное масло, растительное масло, арахисовое масло и другие стандартные метаболизируемые нетоксичные масла, полезные для инъекционных или интраназальных вакцинных композиций. Для целей данного изобретения эти адъюванты идентифицированы в данной заявке как "вторичные", исключительно в противоположность вышеописанному иммуноадъювантному соединению, состоящему из активатора ГТФ-азы семейства Rho, который является существенным ингредиентом в вакцинной композиции для его эффекта в комбинации с антигенным веществом, чтобы значительно повысить гуморальный иммунный ответ на антигенное вещество. Вторичные адъюванты, в основном, включают в препарат вакцины в качестве технологических добавок, хотя некоторые адъюванты действительно обладают до некоторой степени иммунологическими усиливающими свойствами и имеют двойное назначение. Традиционные консерванты можно добавлять в вакцинную композицию в эффективных количествах, находящихся в интервале от примерно 0,0001 до примерно 0,1% мас./мас. В зависимости от консерванта, используемого в препарате, могут быть полезны количества ниже или выше данного интервала. Типичные консерванты включают, например, сорбат калия, метабисульфит натрия, фенол, метилпарабен,пропилпарабен, тимеросал и т.д. Выбор инактивированного, модифицированного или другого типа вакцинной композиции и способа получения усовершенствованного препарата вакцинной композиции по настоящему изобретению известен или может быть легко определен обычными специалистами в данной области техники. Фармакологически эффективное количество иммуноадъювантного соединения в соответствии с изобретением можно давать, например, перорально, парентерально или иным путем, и предпочтительно мукозальным путем, одновременно, последовательно или через короткое время после введения антигенного вещества, чтобы потенцировать, ускорить или продлить иммуногенность антигена. Хотя дозировка вакцинной композиции конкретно зависит от антигена, вида хозяина, вакцинированного или подлежащего вакцинации и т.д., дозировка фармакологически эффективного количества вакцинной композиции обычно будет находиться в диапазоне от примерно 0,01 до примерно 500 мкг (и,в частности, от 50 до примерно 500 мкг) иммуноадъювантного соединения по изобретению на дозу (в частности, на основании результатов, представленных на фиг. 6). Хотя количество конкретного антигенного вещества в комбинации влияет на количество иммуноадъювантного соединения в соответствии с изобретением, необходимое для улучшения иммунного ответа, подразумевается, что практикующий врач сможет легко регулировать количество эффективной дозы иммуноадъювантного соединения посредством рутинных тестов для соответствия конкретным обстоятельствам. Как общее правило, вакцинную композицию по настоящему изобретению удобно вводить перорально, парентерально (подкожно, внутримышечно, внутривенно, внутрикожно или внутрибрюшинно),интрабуккально, интраназально или чрескожно. Путь введения, рассмотренный настоящим изобретением, будет зависеть от антигенного вещества и сопутствующих веществ препарата. Например, если вакцинная композиция содержит сапонины, хотя они нетоксичны перорально или интраназально, необходимо заботиться о том, чтобы не инъецировать сапогениновые гликозиды в кровоток, поскольку они действуют как сильные гемолитики. Также многие антигены не будут эффективны при пероральном приеме. Предпочтительно вакцинную композицию вводят подкожно, внутримышечно или интраназально. Дозировка вакцинной композиции будет зависеть конкретно от выбранного антигена, от пути введения, от вида и от других стандартных факторов. Подразумевается, что обычный специалист в данной области техники может легко и быстро титровать соответствующую дозу для иммуногенного ответа на каждый антиген для достижения эффективного иммунизирующего количества и способа введения. Как уже указано в настоящем описании в других частях, еще одним объектом изобретения является вакцинная композиция по изобретению для введения на поверхность слизистой оболочки. Данный путь введения представляет огромный интерес. Действительно, слизистые оболочки содержат многочисленные дендритные клетки и клетки Лангерганса, которые являются превосходными антиген-обнаруживающими и антиген-представляющими клетками. Слизистые оболочки также связаны с лимфоидными органами, называемыми лимфоидной тканью, ассоциированной со слизистой, которые способы направлять иммунный ответ в другие области слизистой оболочки. Примером такого эпителия является носовая эпителиальная оболочка, которая состоит практически из одного слоя эпителиальных клеток (многорядного эпителия), и слизистая оболочка в верхних дыхательных путях соединена с двумя лимфоидными тканями, аденоидами и миндалинами. Обширная сеть кровеносных капилляров под слизистой оболочкой носа с высокой плотностью В- и Т-клеток особенно подходит для обеспечения быстрого распознавания антигена и обеспечивает быстрый иммунологический ответ. Предпочтительно поверхность слизистой оболочки выбрана из группы, состоящей из поверхностей слизистой оболочки носа, легких, рта, глаза, уха, желудочно-кишечного тракта, половых путей, влагалища, прямой кишки и кожи. Примеры Пример 1. Иммуноадъювантный эффект флагеллинов с делегированной гипервариабельной областью. Материалы и методы Получение рекомбинантных флагеллинов. Рекомбинантные флагеллины имели происхождение от флагеллина FliC Salmonella enterica SerovarTyphimurium ATCC14028 (номер доступа AAL20871). Флагеллины FliC и FliC205-293 либо выделяли из штаммов S. typhimurium SIN22 (fljB) и SJW46, как описано ранее (Yoshioka et al, 1995. Flagellar filament structure and cell motility of Salmonella typhimuriumBiochemicals (Switzerland). Конструкции, кодирующие FliC174-400 и FliC191-352, получали с помощью ПЦР на pBR322 производной плазмиде, несущей ген дикого типа fliC SEQ ID NO:3 под контролем его собственного промотора, и используя приведенные ниже пары праймеров: AGCACCattcagcgtatccagacc (SEQ ID NO:15) /GCTGGTgctacaaccaccgaaaac (SEQ ID NO:16) и TCGAGatatcctgtaacagttgcagcc (SEQ ID NO:17) / ACTCGAGgacggtacatccaaaactgcac (SEQ ID NO:18) (основания, кодирующие линкер, выделены курсивом). Сайт-направленный мутагенез также проводили на плазмиде, несущей FliC174-400, С целью замещения остатков 89-96 (QRVRELAV), вовлеченных в обнаружение TLR5, соответствующими последовательностями из не сигнального флагеллина (DTVKVKAT); полученный в результате белок, таким образом, представлял собой FliC174-400/89-96. В FliC174-400, FliC191-352 и FliC174-400/89-96 аспарагин, локализованный за 6 остатков от конца, был заменен на серин. Укороченные флагеллины очищали из надосадочной жидкости рекомбинантного S. typhimuriumSIN41 (fliC fljB), как описано ниже. Сальмонеллу выращивали в среде Лурия-Бертани (LB) в течение 18 ч при 37 С при встряхивании. Надосадочную жидкость фильтровали и насыщали 60% сульфатом аммония(Sigma Aldrich, USA). Осажденные вещества выделяли центрифугированием, солюбилизацией в 20 мМ Трис/HCl рН 7,5, а затем диализом. Белки дополнительно очищали путем последовательных раундов хроматографии на гидроксиапатите и анионообменной хроматографии (Bio-Rad Laboratories, USA). Наконец, белки истощали по липополисахариду (LPS), используя колонку с полимиксином В (Pierce, USA). Используя анализ Limulus (Associates of Cape Cod Inc., USA), концентрацию остаточного LPS определили как менее чем 30 пг LPS на мкг рекомбинантного флагеллина. Когда указано, флагеллины обрабатывали в течение 1 ч при 37 С 0,017% трипсином-ЭДТА (Invitrogen, USA) для полного гидролиза белков с последующим нагреванием при 70 С в течение 1 ч для инактивации трипсина. Белки анализировали, используя стандартный электрофорез ДСН-ПААГ и иммуноблоттинг с FliC-специфичными поликлональными антителами. Эксперименты на животных Самок мышей NMRI (6-8-недельного возраста) приобретали у Charles River Laboratories (France) и поддерживали в специальном виварии, свободном от патогенов, в зарегистрированной организации(А 59107; Institut Pasteur de Lille). Все эксперименты выполняли с соблюдением существующих национальных и ведомственных норм и руководств по этике. Для гипериммунизации животных инъецировали подкожно (п.к.) флагеллином FliC (1 мкг на инъекцию), эмульгированным в 200 мкл полного адъюванта Фрейнда (CFA)/PBS на сутки 1 и неполного адъюванта Фрейнда (IFA)/PBS на сутки 21, 35 и 49. На сутки 63 мышам давали 200 мкл флагеллина/PBS внутривенно (в.в.) и умерщвляли их через 2 ч путем внутрибрюшинной (в.б.) инъекции 5 мг пентобарбитала натрия (CEVA Sant Animale, France) для отбора образцов и анализа сыворотки и тканей. Для характеристики мукозального наследственно-обусловленного ответа и адъювантных свойств 20 мкл PBS с белками или без белков вводили интраназально (и.н.) мышам, анестезированным в.б. 1,5 мг кетамина (Merial, France) и 0,3 мг ксилазина (Bayer, Germany) на животное массой 25 г. Для исследования провоспалительных ответов отбор образцов у мышей проводили либо через 2 ч(для анализов РНК и генной экспрессии), либо через 6 ч (для тестирования продуцирования цитокинов). Для анализов иммунизации мышам вводили и.н. PBS с овальбумином (OVA), истощенным по LPS,или без него (20 мкг, Sigma, сорт VII, USA) с флагеллинами (1 мкг) или без флагеллинов на сутки 1 и 21. Образцы бронхоальвеолярных лаважей (BAL) и сыворотки отбирали на сутки 35. Для оценки нейтрализации иммунную и контрольную сыворотки нагревали в течение 30 мин при 56 С, чтобы инактивировать комплемент. Серийные разведения сыворотки (в 200 мкл PBS) пассивно переносили животным в.в. путем за 1 ч до системной активации флагеллинами. В некоторых экспериментах сыворотки смешивали с флагеллинами, разведенными в PBS, и вводили и.н. для тестирования мукозальной нейтрализации.BAL собирали после интратрахеальной инъекции 1 мл PBS с полной смесью ингибиторов протеаз(Roche, Switzerland) и осветляли центрифугированием. Образцы крови собирали и подвергали свертыванию при комнатной температуре, после чего отделяли сыворотку путем центрифугирования. Белковые экстракты легких получали путем гомогенизации ткани с 2 мл реагента для экстракции белков из тканей T-PER (Pierce, USA) с добавлением ингибиторов протеаз. Все образцы хранили при-80 С до анализа. Анализ антиген-специфичных ответов антител Уровни OVA- или флагеллин-специфичных антител в образцах сыворотки и BAL оценивали, используя твердофазные иммуноферментные анализы (ELISA). Кратко, OVA (20 мкг на лунку в фосфатном буфере 0,2 М рН 6,5) и флагеллин FliC (100 нг на лунку в PBS) наносили на микропланшеты MaxiSorp (Nalge Nunc Int., USA) в течение ночи при 4 С. Все микропланшеты промывали смесью PBS/Твин 20 0,05%, а затем блокировали смесью PBS/cyxoe молоко 1% в течение 1 ч при комнатной температуре. Серийные разведения образцов инкубировали в течение 1 ч при комнатной температуре до проявления. Биотинилированные антитела IgG или IgA против мыши (Southern Biotechnology Associates, USA),HRP-конъюгированный стрептавидин (GE Healthcare, USA) и 3,3',5,5'-тетраметилбензидин (Becton Dickinson Bioscience, USA) использовали в качестве реагентов для проявления. Реакцию останавливали добавлением H2SO4 и определяли OD при 450 нм. Титр IgG определяли как соответствующий самому высокому разведению образца, дающему значение поглощения 0,15 OD для OVA и 0,5 OD для FliC, и систематически сравнивали с референсной сывороткой. Титры приведены в виде геометрических средних титров от индивидуальных мышей. Уровни суммарного IgA и OVA-специфичного IgA в BAL измеряли и нормализовали, используя калибровочную кривую с имеющимся в продаже IgA мыши (Sigma). Удельное отношение IgA (выраженное в нг OVA-специфичного IgA на мкг суммарного IgA) определяли для каждой мыши. Цитокин-специфичный ELISA и генная экспрессия Уровни CXCL2 и CCL20 мыши и IL-8 (CXCL8) человека измеряли в сыворотке, BAL, суммарной надосадочной жидкости легкого и/или культуры клеток, используя коммерческие наборы ELISA (RDSystems, USA). Суммарную РНК из легких мыши экстрагировали, используя набор Nucleospin RNA II (MachereyNagel, Germany), и подвергали обратной транскрипции с помощью набора High-Capacity cDNA Archive(Applied Biosystems, USA). Полученную в результате кДНК амплифицировали, используя ПЦР в реальном времени SYBR Green-based real-time PCR (Applied Biosystems). Специфичными праймерами являются CGTCATCCATGGCGAACTG (SEQ ID NO:19) /GGACAC (SEQ ID NO:21) / TGCAGGTGAAGCCTTCAACC (SEQ ID NO:22) (CCL20) и CCCTCAACGGAAGAACCAAA (SEQ ID NO:23) / CACATCAGGTACGATCCAGGC (SEQ ID NO:24) (CXCL2). Относительные уровни мРНК (2-Ct) определяли путем сравнения (а) порогов числа циклов ПЦР (Ct) для интересующего гена и АСТВ (Ct) и (б) значений Ct для обработанных и контрольных групп (Ct), как описано ранее (Sierro et al, 2001, Flagellin stimulation of intestinal epithelial cells triggers CCL20-mediatedmigration of dendritic cells. Proc. Natl. Acad. Sci. USA 98:13722-13727). Анализы на клеточной основе Линия клеток аденокарциномы ободочной кишки человека Сасо-2 была стабильно трансфицирована плазмидой, несущей ген люциферазы под контролем промотора CCL20 человека (Rumbo et al, 2004,Lymphotoxin beta receptor signaling induces the chemokine CCL20 in intestinal epithelium. Gastroenterol. 127:213-223) с получением линии Сасо-Rumbo. Эти эпителиальные клетки кишечника выращивали в модифицированной Дульбекко среде Игла с добавлением 10% фетальной сыворотки теленка, 10 мМ HEPES, заменимых аминокислот 1 Х, пенициллина (100 ед./мл) и стрептомицина (100 ед./мл) и (для селекции трансгена) 0,7 мг/мл G418 (Invitrogen). Линию бронхиальных эпителиальных клеток человека BEAS-2B культивировали в питательной среде Kaigh's F12 с добавками, как для среды Caco-Rumbo плюс 1 мМ пируват натрия и смесь инсулина,трансферрина и селена (Invitrogen). Клетки стимулировали рекомбинантными флагеллинами в течение 6 ч для анализов люциферазы или в течение 16 ч перед сбором надосадочной жидкости для ELISA. Активность люциферазы в клеточных экстрактах измеряли, используя набор Bright Glo LuciferaseAssay (Promega, USA). Относительную люминесценцию (RLU) нормализовали в виде процента максимальной активности с флагеллином дикого типа для теста на активацию рекомбинантными флагеллинами. Для теста нейтрализации in vitro RLU нормализовали в виде процента максимальной активности для каждого белка: [(RLU обработанный/RLU необработанный)/(RLU макс/RLU необработанный)]100. Статистический анализ Статистические различия анализировали, используя критерий Манна-Уитни, и считали значимыми- 22019138 для значений р 0,05. Если не указано иное, результаты выражали в виде арифметических среднихстандартное отклонение. Результаты Делеция гипервариабельной области флагеллина придает антигенность. но не модифицирует TLR5 стимулирующую активность Две новых молекулы флагеллина (FliC191-352 и FliC174-400, состоящие соответственно из 336 и 271 аминокислот) были сконструированы путем внутренней делеции (фиг. 1 А). В качестве контроля авторы изобретения использовали охарактеризованный ранее вариант FliC204292, который имел частичную делецию в антигенном домене (Yoshioka et al, 1995, Flagellar filament structure and cell motility of Salmonella typhimurium mutants lacking part of the outer domain of flagellin. J. Bacteriol. 177:1090-1093) (фиг. 1 А). В качестве отрицательного контроля для экспериментов in vitro и in vivo мутации, которые придают передачу сигнала TLR5, вводили в FliC174-400 с получением рекомбинантного белка FliC174-400/89-96. Предсказанные структуры соответствующих флагеллинов показали, что мотив 89-96 и общая структура консервативных областей не были изменены (фиг. 1 А). За исключением FliC204-292, эти варианты были неспособны комплементировать подвижность бактерий с дефицитом по флагеллину и секретировались в культуральную среду. Затем авторы изобретения оценивали собственную антигенность рекомбинантных флагеллинов. С этой целью насыщающие концентрации флагеллинов наносили на микропланшеты и тестировали с помощью ELISA, используя гипериммунную сыворотку, специфичную к FliC или FliC174-400. Как проиллюстрировано на фиг. 1 В, авторы наблюдали титры антител в 3-10-раз ниже, когда сыворотку анти-FliC титровали против вариантов FNC, чем против FliC дикого типа. Напротив, реактивность гипериммунной сыворотки, специфичной к FliC174-400, была сходной независимо от флагеллина-мишени (фиг. 1 С). Эти результаты подтверждают, что центральная гипервариабельная область является основной мишенью для антител анти-флагеллин. Наконец, авторы изобретения решили установить, сохраняют ли рекомбинантные молекулы какуюлибо TLR5-стимулирующую активность. Анализ доза-ответ проводили, используя репортерные клетки Сасо-Rumbo и клеточную линию легочного эпителия BEAS-2B. Активацию оценивали путем измерения активности люциферазы в клеткахCaco-Rumbo и секреции IL-8 клетками BEAS-2B (клеточные линии кишечного эпителия человека являются уникальными репортерами флагеллин/TLR5-стимуляторной активности на основании экспрессии хемокина CCL20, также известного как "хемокины, активируемые и регулируемые печенью" или LARC,и IL-8). Как показано на фиг. 2 А-В, FliC204-292, FliC191-352 и FliC174-400 все были сильными клеточными активаторами. Соответствующие значения ЕС 50 флагеллинов слегка варьировали в зависимости от клеточного типа, но попадали в пределы описанного ранее интервала в нг/мл (Smith et al, 2003, Toll-like receptor 5 recognizes a conserved site on flagellin protofilament formation and bacterial motility. Nat. Immunol. 4:1247-1253). Было обнаружено, что активность рекомбинантных флагеллинов полностью зависит от TLR5, поскольку FliC174-400/89-96 был неспособен активировать эпителиальные клетки. Потребность в передаче сигнала TLR5 была дополнительно подтверждена путем использования макрофагов костного мозга, выделенных из TLR5-дефицитных мышей; клетки не синтезировали никакого IL-12 р 40 при стимуляции рекомбинантными флагеллинами (данные не показаны). Делетированные флагеллины стимулируют TLR5-зависимые мукозальные наследственно-обусловленные ответы Стимуляция TLR5 рекомбинантными флагеллинами была затем исследована in vivo мукозальным путем. С этой целью экспрессию CCL20 и CXCL2 в легких мышей, обработанных и.н. флагеллинами, количественно определяли, используя кОТ-ПЦР (фиг. 2 С). В течение 2 ч уровни мРНК CCL20 в легких были примерно в 30 раз выше у животных, обработанных флагеллином дикого типа или рекомбинантными флагеллинами, чем у мышей, обработанных контролем. Кроме того, продуцирование хемокина CCL20 обнаруживали через 6 ч после инстилляции, и в гомогенатах легких, и в BAL (фиг. 2D). В контрольных экспериментах FliC174-400/89-96 и флагеллины, расщепленные трипсином, не индуцировали данный тип эффекта. Подобные открытия наблюдали дляCXCL2 (данные не показаны). Эти результаты подтвердили, что провоспалительный ответ in vivo исключительно был следствием рекомбинантных флагеллинов. В целом флагеллины с делециями в гипервариабельной области проявляли мукозальные провоспалительные свойства, эквивалентные свойствам соответствующего FNC дикого типа. Рекомбинантные флагеллины проявляют мукозальную адъювантную активность С целью характеристики адъювантных свойств рекомбинантных молекул, полученных авторами изобретения, исследовали ответы антител в сыворотке и секрециях после и.н. иммунизации. В качестве модельного антигена использовали овальбумин (OVA), включенный в препарат с различными флагеллинами или без флагеллинов или с холерным токсином (СТ) в качестве эталонного мукозального адъюванта. Совместное введение FliC с OVA значительно повышало ответ OVA-специфичного IgG (как в сыворотке, так и в BAL, примерно в 300 и в 100 раз, соответственно) по сравнению с животными, иммунизированными только OVA (фиг. 3 А-В). Кроме того, ответ OVA-специфичного IgA был усилен в BAL, подтверждая, что FliC стимулирует прототипичный ответ секреторных антител мукозального адъюванта (фиг. 3 С). Интересно, что эффектFliC был сходным с таковым для СТ. Подобно FliC, рекомбинантные флагеллины FliC204-292, FliC191-352 И FliC174-400 были, таким образом, способны потенцировать системный и мукозальный ответы. Напротив, у FliC174-400/89-96 и флагеллинов, обработанных трипсином, отсутствовала эффективность(фиг. 3 и табл. 1). Следовательно, делеция гипервариабельной области флагеллинов не оказывает значительного влияния на TLR5-опосредованные мукозальные адъювантные свойства. Данные авторов изобретения также показали, что соответствующие эффекты рекомбинантных молекул на наследственнообусловленный и адаптивный иммунитет коррелируют. Делеция гипервариабельной области придает способность продуцировать антитела антифлагеллин Ожидают, что делеция антигенного домена снижает флагеллин-специфичный иммунный ответ и,следовательно, какую-либо нейтрализацию TLR5-опосредованного иммунитета, в частности, при повторном введении. Поэтому авторы изобретения решили оценить эффективность и.н. иммунизации в отношении индукции FliC-специфичных антител. Как показано, FliC вызывал сильный ответ IgG в сыворотке и BAL (табл. 1 и фиг. 4). Напротив,FliC204-292 запускал в 10 раз более низкие уровни антител в обеих жидкостях, чем FliC, и более выраженный эффект наблюдали после иммунизации FliC191-352 и FliC174-400. В заключение, антигенный и иммуностимулирующий домены флагеллинов функционально разобщены. Поэтому FliC191-352 и FliC174-400 являются молекулами, представляющими интерес для предупреждения или аттенуации образования флагеллин-специфичных антител с нейтрализующей активностью. Флагеллин-специфичные антитела нейтрализуют TLR5-опосредованную передачу сигнала Известно, что бактериальные флагеллины вызывают сильные ответы антител, которые в основном направлены против гипервариабельной области. Авторы изобретения выдвинули гипотезу, что антитела анти-флагеллин должны нейтрализовать TLR5-стимулирующую активность флагеллинов. Следовательно, мышей иммунизировали п.к. флагеллином FliC или контрольным препаратом (одним PBS или нерелевантным антигеном овальбумином (OVA), включенным в CFA) с последующими бустер-иммунизациями IFA. Анализы ELISA выявили, что сыворотки анти-FNC проявляли специфичные титры IgG более 106, тогда как титры контрольных сывороток были ниже порога обнаружения анализа(102). Как упомянуто выше, клеточные линии кишечного эпителия человека полезны в качестве уникальных репортеров флагеллин/TLR5-стимулирующей активности, основанных на экспрессии хемокинаCCL20 (также известного как "активируемый и регулируемый печенью хемокин", LARC). Таким образом, используя клетки Caco-Rumbo, несущие ген люциферазы под контролем промотораCCL20, здесь продемонстрировано, что сыворотка анти-FliC способна полностью нейтрализовать агонистическую активность FNC в отношении TLR5 (фиг. 5 А). Нейтрализующий эффект FliC-специфичных антител на передачу сигнала TLR5, таким образом,непосредственно оценивали у иммунизированных животных. С этой целью исследовали системные провоспалительные ответы у мышей (продуцирование хемокинов CCL20 и CXCL2) после в.в. инъекции FliC(фиг. 5 В-С). У животных, иммунизированных контролем, провокация FliC запускала значительное повышение сывороточных уровней CCL20 и CXCL2 по сравнению с провокацией PBS. Напротив, продуцирование хемокинов у FliC-иммунизированных животных не было усилено ни одной из провокаций. Используя пассивный перенос сыворотки у интактных животных, обнаружили близкую корреляцию между количеством инъецированного антитела и системным наследственнообусловленным ответом, как показано на фиг. 5D. В заключение, заранее существующий иммунитет к флагеллину может нейтрализовать последующую TLR5-стимулирующую активность как in vitro, так и in vivo. Это не имеет места в случае FliC174-400, у которого сильно нарушена способность к стимуляции- 24019138 продуцирования флагеллин-специфичных антител, включая нейтрализующие антитела, как раскрыто выше в соответствии с фиг. 4. Были определены эффективные дозы, необходимые для инициации TLR5-опосредованных наследственно-обусловленных ответов и.н. путем. FliC и FliC174-400 проявляли сходные кривые доза-ответ, и доза 0,1 мкг была выбрана для последующих анализов нейтрализации (фиг. 6). С этой целью животных подвергали гипериммунизации и.н. FNC, чтобы вызвать сильные ответыFliC-специфичных IgG (средний титр примерно 45000), а затем подвергали провокации и.н. 0,1 мкг флагеллинов FNC или FliC174-400. Проводили мониторинг продуцирования провоспалительных хемокинов вBAL. Провокация FliC или FliC174-400 приводила к продуцированию CCL20 (4,281,98 по сравнению с 1,080,54 нг/мл и 2,481,22 по сравнению с 0,930,48 нг/мл у животных, иммунизированных контролем или FliC, соответственно), что наблюдали у интактных животных. Мукозальный и системный TLR5-зависимые ответы в различной степени зависят от гипервариабельной области флагеллина Авторы изобретения также хотели исследовать нейтрализацию флагеллин-специфичными антителами TLR5-зависимых ответов, индуцированных после в.в. инъекции рекомбинантных флагеллинов. Для анализа системной активации наследственно-обусловленного иммунитета продуцирование в кровообращении провоспалительных хемокинов CCL20 и CXCL2 измеряли с помощью ELISA в сыворотке (фиг. 7). Неожиданно авторы изобретения обнаружили, что у FliC174-400 была примерно в 100 раз нарушена его способность к запуску системных провоспалительных иммунных ответов по сравнению с FliC дикого типа. Если 10 мкг FliC174-400 стимулировали небольшое продуцирование хемокинов, у варианта, мутированного внутри мотива TLR5 FliC174-400/89-96, отсутствовала активность (0,850,27 по сравнению с 0,020,00 нг/мл для CCL20). Это было противоположным FliC204-292 и FliC191-352, которые оба были сильными активаторами,подобными FliC. Следовательно, определенные молекулярные детерминанты на гипервариабельной области (или зависимые от последней) требуются для системной стимуляции TLR5, но не мукозальной стимуляцииTLR5. Взятые вместе, результаты авторов изобретения показывают, что активация TLR5 в мукозальном и системном компартментах регулируется разными механизмами. Пример 2. Биологическая активность флагеллинов с делетированной гипервариабельной областью,выбранных из группы, состоящей из FliC174-400, FliC161-405 и FliC138-405. Получение рекомбинантных флагеллинов с делетированной гипервариабельной областью Различные флагеллины с делетированной гипервариабельной областью продуцировали рекомбинантным путем, осуществляя тот же способ, который раскрыт выше в примере 1, а именно FliC174-400,FliC161-405 и FliC138-405 и FliC100-405. На фиг. 8 и 9 изображены анализы указанных белков, полученных рекомбинантным путем. На фиг. 8 показан результат электрофореза ДСН-ПААГ, который проведен на рекомбинантных белках, собранных из надосадочной жидкости культуры из соответствующих рекомбинантных бактериальных клеток S. typhimurium SIN41 после стадии осаждения белка трихлоруксусной кислотой (ТХУ). На фиг. 9 показан результат Вестерн-блот-анализа с использованием поликлональных антител антиFliC, который проведен на надосадочной жидкости культуры из соответствующих рекомбинантных бактериальных клеток S. typhimurium SIN41 после стадии осаждения белка ТХУ. Биологическая активность флагеллинов с делетированной гипервариабельной областью FliC174-400, FliC161-405 и FliC138-405 Эффект FliC174-400, FliC161-405 и FliC138-405 на индукцию CCL20 и CXCL2 анализировали путем проведения цитокин-специфичного анализа ELISA, который описан в примере 1. Кратко, С 3 Н/HeJ (TLR4-дефицитные) инъецировали внутрибрюшинно 10 мкг различных рекомбинантных флагеллинов, делетированных от положений 174-400, 161-405 и 138-405. Через 2 ч отбирали образцы сыворотки и обрабатывали для цитокин-специфичного ELISA (CCL20 и CXCL2). Препарат флагеллина был получен из надосадочной жидкости рекомбинантных Salmonella, которую предварительно осаждали сульфатом аммония и подвергали диализу (фиг. 8 и 9). Поскольку этот неочищенный препарат может быть загрязнен эндотоксином, авторы изобретения использовали мышей с дефицитом по передаче сигнала TR4, так как LPS может быть основным загрязнителем в этом неочищенном препарате. Кроме того, авторы изобретения использовали обработку трипсином, чтобы продемонстрировать, что биологическая активность присутствует в белковой фракции неочищенных препаратов. Результаты изображены на фиг. 10 (индукция CCL20) и 11 (индукция CXCL2). Результаты фиг. 10 и 11 подтверждают, что рекомбинантные флагеллины FliC161-405 и FliC138-405- 25019138 компетентны к передаче сигнала in vivo, как описано для FHC174-400. Эти результаты подтверждают, что FliC161-405 и FliC138-405 являются эффективными агонистамиTLR5 и, следовательно, могут представлять собой эффективные адъювантные соединения. Пример 3. Адъювантное действие FliC174-400 на иммунные ответы против антигена gp140 из вирусаHIV1. Протоколы иммунизации и анализ ответов антиген-специфичных антител являются такими же, как описано в примере 1, за исключением конкретных признаков, которые могут быть указаны ниже. Кратко, адъювантную активность нативного флагеллина FNC и рекомбинантного FliC174-400 на антиген HIV1 gp140 осуществляли, как описано ниже: мышей NMRI (n=8) иммунизировали на сутки 1 и на сутки 21 интраназально 20 мкл PBS, содержащего gp140 (5 мкг на мышь), без флагеллинов, с FliC или сFliC174-400 (1 мкг на мышь). Образцы сыворотки и бронхоальвеолярных лаважей (BAL) отбирали на сутки 35 и определяли титр антител с помощью gp140-специфичного ELISA. Результаты изображены на фиг. 12, где каждый символ представляет индивидуальную мышь, а полоса представляет геометрическое среднее. Символы означают мышей, которым вводили интраназально, соответственно: (1) кружки: толькоHIV1 gp140; (2) ромбики: HIV1 gp140 и FliC174-400; (3) треугольники: gp140 и FliC. Титры антител из образцов сыворотки представлены в виде закрашенных символов в левой части фиг. 12 (заштрихованные символы). Титры антител из образцов бронхоальвеолярного лаважа представлены в виде незакрашенных символов в правой части фиг. 12. Результаты показывают, что различные флагеллины с делетированной гипервариабельной областью, как определено в настоящем описании, составляют эффективные иммуноадъювантные соединения. Таблица 1. Чувствительные к протеазам иммунные ответы, индуцированные рекомбинантными флагеллинами Мышей (n=8) иммунизировали и.н. PBS, овальбумином (OVA), OVA + флагеллины/пептидные производные флагеллина или OVA + обработанные трипсином флагеллины (TRP) на сутки 1 и 21. На сутки 35 титры OVA- и FliC-специфичного IgG измеряли в сыворотке и BAL. Статистическую значимость (р 0,05) определяли критерием Манна-Уитни. Значения выражены в виде Log 10 (реципрокные титры)стандартное отклонение (СО). В сыворотке и BAL предел обнаружения составляет 2 и 0,3 (разведение сыворотки 1/100 и разведение BAL 1/2) соответственно. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Иммуноадъювантное соединение, содержащее: а) N-концевой пептид, обладающий по меньшей мере 90% аминокислотной идентичностью с аминокислотной последовательностью, начинающейся с аминокислотного остатка, локализованного в положении 1 SEQ ID NO:1, и оканчивающейся аминокислотным остатком, выбранным из группы, состоящей из любого из аминокислотных остатков, локализованных в положениях 99-173 SEQ ID NO:1; и б) С-концевой пептид, обладающий по меньшей мере 90% аминокислотной идентичностью с аминокислотной последовательностью, начинающейся с аминокислотного остатка, выбранного из группы,состоящей из любого из аминокислотных остатков, локализованных в положениях 401-406 SEQ ID NO:1,и оканчивающейся аминокислотным остатком, локализованным в положении 494 SEQ ID NO:1; где указанный N-концевой пептид непосредственно связан с указанным С-концевым пептидом или указанный N-концевой пептид и указанный С-концевой пептид связаны друг с другом опосредованно через спейсерную цепь, где спейсерная цепь содержит от 1 до 20 аминокислот, связанных пептидными связями, где аминокислоты выбраны из 20 природных аминокислот. 2. Иммуноадъювантное соединение по п.1, где указанный N-концевой пептид выбран из группы,состоящей из аминокислотных последовательностей 1-99, 1-137, 1-160 и 1-173 SEQ ID NO:1. 3. Иммуноадъювантное соединение по любому из пп.1 или 2, где указанный С-концевой пептид выбран из группы, состоящей из аминокислотных последовательностей 401-494 и 406-494 SEQ ID NO:1. 4. Иммуноадъювантное соединение по любому из пп.1-3, где указанные N-концевой и С-концевой пептиды состоят из аминокислотных последовательностей 1-173 и 401-494 SEQ ID NO:1 соответственно. 5. Иммуноадъювантное соединение по любому из пп.1-3, где указанные N-концевой и С-концевой пептиды состоят из аминокислотных последовательностей 1-160 и 406-494 SEQ ID NO:1 соответственно. 6. Иммуноадъювантное соединение по любому из пп.1-3, где указанные N-концевой и С-концевой пептиды состоят из аминокислотных последовательностей 1-137 и 406-494 SEQ ID NO:1 соответственно. 7. Иммуноадъювантное соединение по любому из пп.1-6, где указанный N-концевой пептид и указанный С-концевой пептид связаны друг с другом опосредованно через промежуточную спейсерную цепь, состоящую из пептидной последовательности NH2-Gly-Ala-Ala-Gly-COOH. 8. Иммуноадъювантное соединение по любому из пп.1-7, где аминокислотный остаток аспарагина,локализованный в положении 488 SEQ ID NO:1, заменен серином. 9. Иммуноадъювантное соединение по любому из пп.1-7, где указанное соединение содержит до- 27019138 полнительный остаток метионина на N-конце. 10. Фармацевтическая композиция, содержащая иммуноадъювантное соединение по любому из пп.1-9 вместе с одним или более чем одним фармацевтически приемлемым эксципиентом. 11. Иммуногенная композиция, содержащая иммуноадъювантное соединение по любому из пп.1-9 вместе с одним или более чем одним антигеном. 12. Вакцинная композиция, содержащая иммуноадъювантное соединение по любому из пп.1-9 вместе с одним или более чем одним антигеном. 13. Иммуногенная композиция по п.11 или вакцинная композиция по п.12, где указанное иммуноадъювантное соединение нековалентно связано с одним или более чем одним указанным антигеном. 14. Применение иммуноадъювантного соединения по любому из пп.1-9 в качестве лекарственного средства. 15. Применение иммуноадъювантного соединения по любому из пп.1-9 для изготовления фармацевтической композиции. 16. Нуклеиновая кислота, кодирующая иммуноадъювантное соединение по любому из пп.1-9. 17. Рекомбинантный вектор, содержащий встроенную в него нуклеиновую кислоту по п.16. 18. Клетка-хозяин, трансфицированная или трансформированная нуклеиновой кислотой по п.16 или рекомбинантным вектором по п.17.
МПК / Метки
МПК: C12N 15/11, C07K 14/255, A61K 39/39
Метки: новые, применение, флагеллина, соединения, основе, иммуноадъювантные
Код ссылки
<a href="https://eas.patents.su/30-19138-novye-immunoadyuvantnye-soedineniya-na-osnove-flagellina-i-ih-primenenie.html" rel="bookmark" title="База патентов Евразийского Союза">Новые иммуноадъювантные соединения на основе флагеллина и их применение</a>
Диароматические соединения пропинила или диенила, промежуточные соединения, фармацевтические и косметические композиции на основе указанных соединений и применение косметических композиций.

Номер патента: 1060
Опубликовано: 30.10.2000
Автор: Бернардон Жан-Мишель
МПК: C07C 63/66, A61P 17/00, A61K 31/192...
Метки: применение, соединений, диенила, косметические, композиции, промежуточные, диароматические, композиций, основе, указанных, соединения, пропинила, косметических, фармацевтические
Формула / Реферат:
1. Диароматические соединения пропинила или диенила общей формулы (I) где R1 является (I) радикалом -СН3, (II) радикалом -СН2-O-R6, (III) радикалом -О-R6, (IV) радикалом -CO-R7, где R6 и R7 имеют значения, приведенные ниже, Аr является радикалом, отвечающим следующей формуле (а): где R5 имеет значения, приведенные ниже, Х является радикалом формулы или или где R8 и R9 имеют значения, приведенные ниже, R2 и R3,...
Новые полициклические соединения и их применение

Номер патента: 7868
Опубликовано: 27.02.2007
Авторы: Данн Дерек, Чаттерджи Санкар, Биховски Рон, Хадкинс Роберт Л., Атор Марк А.
МПК: A61K 31/407, A61K 31/4035, A61P 25/00...
Метки: полициклические, соединения, новые, применение
Формула / Реферат:
1. Соединение формулы IIIa где каждый А и В обозначает, независимо, С(=O), СН(ОН) или CH2; Е и F вместе с атомами углерода, к которым они присоединены, образуют незамещенную С4-С7-циклоалкильную группу; R1 обозначает водород, низший алкил, низший алкил, имеющий по меньшей мере один заместитель J; формил; ацетил, низший алканоил, низший алканоил, имеющий по меньшей мере один заместитель J, низший алкилсульфонил, низший арилсульфонил, остаток...
Новые соединения, их применение и получение

Номер патента: 8148
Опубликовано: 27.04.2007
Авторы: Йенссон Маттиас, Нильссон Бьерн, Шеберг Биргер, Рингберг Эрик
МПК: A61K 31/497, A61P 25/00, A61K 31/444...
Метки: применение, новые, получение, соединения
Формула / Реферат:
1. Соединение формулы (I) где nх равно 2-4, предпочтительно 2; каждый из R0 и R1 независимо означает Н или СН3; R2 означает Н, C1-C4-алкил, 2-гидроксиэтил, 2-цианоэтил или тетрагидропиран-2-ил, C1-C4-ацил или C1-C4-алкоксикарбонил; каждый из R3-R5 независимо означает Н, галоген, метил или метокси, при условии, что по меньшей мере один из R3-R5 означает водород; каждый из X, Y и z независимо означает СН или N; A1 означает О, СН или СН2; A2...
Новые модифицированные полиэтиленгликолем соединения и их применение

Номер патента: 10016
Опубликовано: 30.06.2008
Авторы: Иин Кан, Хольмез Кристофер П., Тумелти Дэвид
МПК: C07K 14/505, A61K 47/48
Метки: полиэтиленгликолем, новые, применение, модифицированные, соединения
Формула / Реферат:
1. Соединение, содержащее: (а) пептидный фрагмент и поли(этиленгликолевый) фрагмент, который содержит не более чем примерно 50 аминокислот; и (б) поли(этиленгликолевый) фрагмент, ковалентно присоединенный к указанному пептидному фрагменту, причем указанный поли(этиленгликолевый) фрагмент является линейным, непрерывным и неразветвленным и имеет молекулярную массу, превышающую 20 кДа. 2. Соединение по п.1, в котором поли(этиленгликолевый) фрагмент...
4-аминотетрагидробензизоксазоловые или -изотиазоловые соединения, их применение и фармацевтическая композиция на их основе

Номер патента: 380
Опубликовано: 24.06.1999
Авторы: Мольтсен Ленс Сибюлле, Перрегор Енс Кристьян, Фрёлунн Бенте, Скоусбо Арне, Крогсгор-Ларсен Повл, Фальк Эрик
МПК: C07D 261/20
Метки: соединения, 4-аминотетрагидробензизоксазоловые, изотиазоловые, композиция, основе, фармацевтическая, применение
Формула / Реферат:
1. 4-Аминотетрагидробензизоксазоловые или -изотиазоловые соединения, имеющие общую формулу I: где R1 и R2, каждый, независимо выбран из группы, состоящей из А) водорода, С3-7 циклоалкила, фенила или группы где R, R8 и R9, каждый, независимо выбран из группы, состоящей из водорода, C1-4 алкила, C2-4 алкенила, С2-4 алкинила, С1-4 алкокси- C1-4 алкила, С3-7 циклоалкила, С3-7 циклоалкил-С1-4 алкила, фенила, фенил-C1-4 алкила, фенокси-C1-4...
Предыдущий патент: Устройство для внутренней остеодистракции
Следующий патент: Реактор синтеза углеводородов типа барботажной колонны и способ определения уровня суспензии
Случайный патент: Система экспрессии гетерологичных и гомологичных целлюлаз