Агенты, специфически связывающие ангиопоэтин-2





























Формула / Реферат
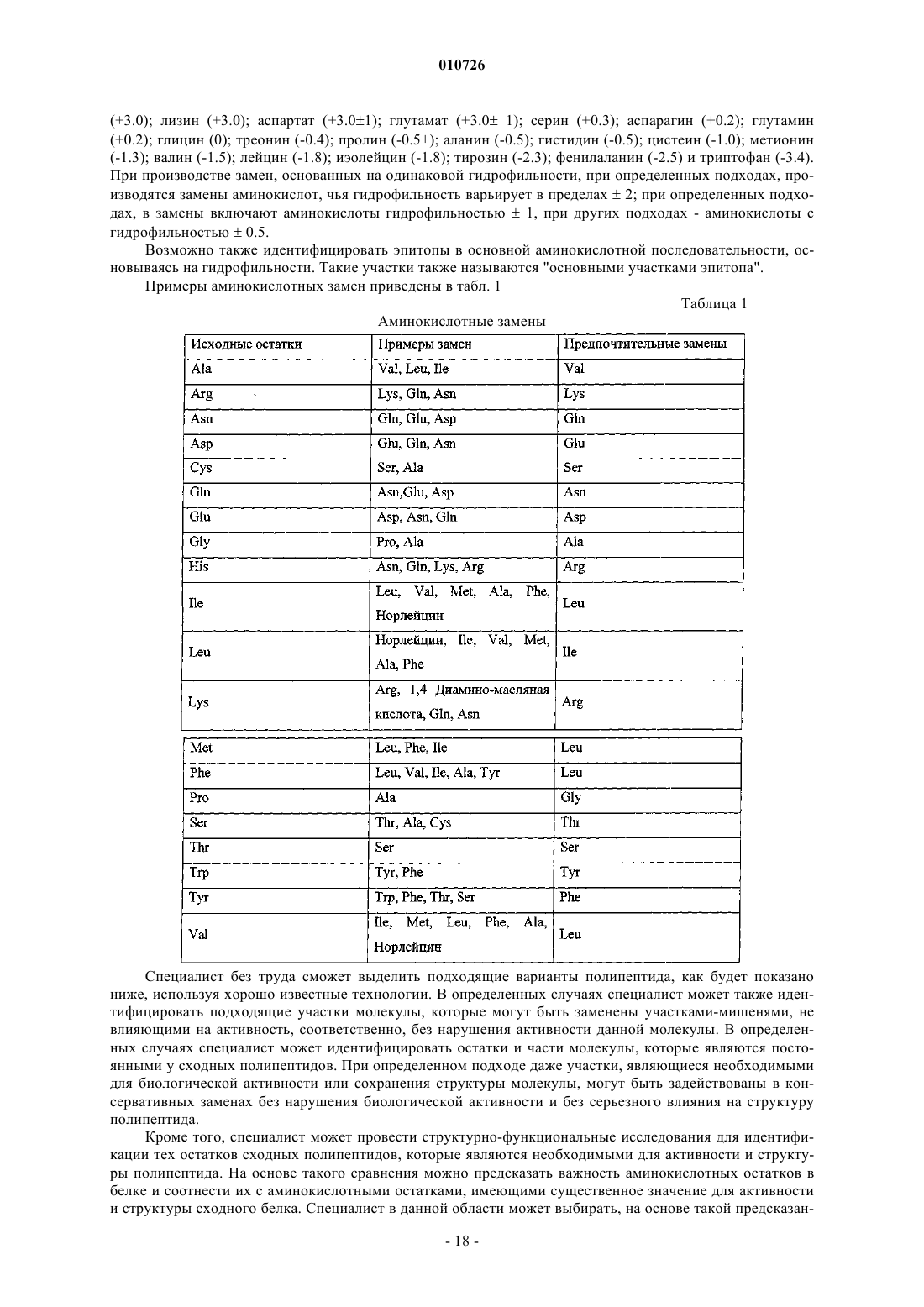
1. Агент, специфически связывающий ангиопоэтин-2, содержащий по крайней мере один пептид, с аминокислотной последовательностью, выбранной из группы, включающей
![]()

![]()
и их фрагменты.
2. Агент по п.1, являющийся антителом.
3. Агент по п.2, являющийся поликлональным, моноклональным, химерным, гуманизированным или полноразмерным антителом человека.
4. Агент по п.3, являющийся одноцепочечным антителом.
5. Гибридома, продуцирующая моноклональное антитело по п.3.
6. Конъюгат, содержащий агент по п.1.
7. Конъюгат, содержащий агент по любому из пп.2, 3 или 4.
8. Молекула нуклеиновой кислоты, кодирующая агент по п.1.
9. Молекула нуклеиновой кислоты, кодирующая агент по любому из пп.2, 3 или 4.
10. Вектор, содержащий молекулу нуклеиновой кислоты по любому из пп.8 или 9.
11. Клетка-хозяин, содержащая вектор по п.10.
12. Способ получения агента, специфически связывающего ангиопоэтин-2, включающий:
(a) трансформацию клетки-хозяина по крайней мере одной молекулой нуклеиновой кислоты, кодирующей агент по п.1;
(b) обеспечение экспрессии молекулы нуклеиновой кислоты в указанной клетке-хозяине; и
(c) выделение указанного агента.
13. Способ получения антитела, специфически связывающего ангиопоэтин-2, включающий:
(a) трансформацию клетки-хозяина по крайней мере одной молекулой нуклеиновой кислоты, кодирующей агент по любому из пп.2, 3 или 4;
(b) обеспечение экспрессии молекулы нуклеиновой кислоты в указанной клетке-хозяине; и
(c) выделение указанного антитела.
14. Способ ингибирования нежелательного ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по п.1.
15. Способ лечения раковых заболеваний у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по п.1.
16. Способ ингибирования нежелательного ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по любому из пп.2, 3 или 4.
17. Способ лечения раковых заболеваний у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по любому из пп.2, 3 или 4.
18. Фармацевтическая композиция, содержащая специфический связывающий агент по п.1 и дополнительный агент в фармакологически доступной форме.
19. Фармацевтическая композиция, содержащая агент по любому из пп.2, 3 или 4 и дополнительный агент в фармакологически доступной форме.
20. Способ модулирования или ингибирования активности ангиопоэтина-2, заключающийся во введении агента по п.1.
21. Способ модулирования или ингибирования активности ангиопоэтина-2, заключающийся во введении агента по любому из пп.2, 3 или 4.
22. Способ модулирования сосудистой проницаемости или утечки плазмы в организме млекопитающих, заключающийся во введении терапевтически эффективного количества агента по п.1.
23. Способ лечения глазного неоваскулярного заболевания, ожирения, гемангиобластомы, гемангиомы, артериосклероза, воспалительных заболеваний, воспалительных нарушений, атеросклероза, эндометриоза, неопластических заболеваний, заболеваний костной системы и псориаза у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по п.1.
24. Способ модулирования сосудистой проницаемости или утечки плазмы в организме млекопитающих, заключающийся во введении терапевтически эффективного количества агента по любому из пп.2, 3 или 4.
25. Способ лечения глазного неоваскулярного заболевания, ожирения, гемангиобластомы, гемангиомы, артериосклероза, воспалительных заболеваний, воспалительных нарушений, атеросклероза, эндометриоза, неопластических заболеваний, заболеваний костной системы и псориаза у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по любому из пп.2, 3 или 4.
26. Способ лечения раковых заболеваний у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по п.1 и химиотерапевтического агента.
27. Способ по п.26, заключающийся в неодновременном введении связывающего агента и химиотерапевтического агента.
28. Способ лечения раковых заболеваний у млекопитающих, заключающийся во введении терапевтически эффективного количества агента по любому из пп.2, 3 или 4 и химиотерапевтического агента.
29. Способ по п.26, заключающийся в неодновременном введении связывающего агента и химиотерапевтического агента.
30. Агент, специфически связывающий ангиопоэтин-2, содержащий CDR1, выбранную из
![]()


![]()
причем локализация CDR1 соответствует представленной в табл. 7.
31. Агент, специфически связывающий ангиопоэтин-2, содержащий CDR2, выбранную из
![]()

![]()
причем локализация CDR2 соответствует представленной в табл. 7.
32. Агент, специфически связывающий ангиопоэтин-2, содержащий CDR3, выбранную из
![]()

![]()
причем локализация CDR3 соответствует представленной в табл. 7.
33. Молекула нуклеиновой кислоты, кодирующая агент по любому из пп.30, 31 или 32.
34. Способ определения уровня ангиопоэтина-2 в биологическом образце, включающий:
(a) обеспечение взаимодействия агента по п.1 с биологическим образцом;
(b) определение продолжительности связывания агента с указанным образцом, и
(c) корреляцию продолжительности связывания агента с образцом с уровнем ангиопоэтина-2 в образце.
35. Способ определения уровня ангиопоэтина-2 в биологическом образце, включающий:
(a) обеспечение взаимодействия агента по п.2 с биологическим образцом;
(b) определение продолжительности связывания антитела с указанным образцом, и
(c) корреляцию продолжительности связывания агента с образцом с уровнем ангиопоэтина-2 в образце.
36. Антитело, содержащее тяжелую цепь и легкую цепь, причем
тяжелая цепь включает вариабельный участок, выбранный из
![]()

![]()
и связывающих ангиопоэтин-2 фрагментов данного участка;
и легкая цепь содержит вариабельный участок, выбранный из


![]()
и связывающих ангиопоэтин-2 фрагментов данного участка.
37. Молекула нуклеиновой кислоты, кодирующая антитело по п.36.
38. Агент по п.32, содержащий CDR3, выбранную из 536 НС (SEQ ID NO. 11) или 536 каппа (SEQ ID NO. 12), и дополнительно содержащий по меньшей мере одну из CDR1 или CDR2 536 каппа (SEQ ID NO. 12) или 536 НС (SEQ ID NO. 11).
39. Агент по п.32, содержащий CDR3 последовательности SEQ ID NO. 11 или CDR3 последовательности SEQ ID NO. 12.
40. Изолированное антитело, специфически связывающее ангиопоэтин-2, содержащее тяжелую цепь и легкую цепь, причем тяжелая цепь включает вариабельный участок, выбранный из CDR1, CDR2 и CDR3 последовательности SEQ ID NO. 11, а легкая цепь включает вариабельный участок, выбранный из CDR1, CDR2 и CDR3 последовательности SEQ ID NO. 12.
41. Антитело по п.40, у которого вариабельный участок тяжелой цепи включает последовательность SEQ ID NO. 11, а вариабельный участок легкой цепи включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO. 12.
42. Антитело по п.40, у которого вариабельный участок тяжелой цепи включает CDR1, CDR2 и CDR3 последовательности SEQ ID NO. 11, а вариабельный участок легкой цепи включает последовательность SEQ ID NO. 12.
43. Антитело по п.40, у которого вариабельный участок тяжелой цепи включает последовательность SEQ ID NO. 11, а вариабельный участок легкой цепи включает последовательность SEQ ID NO. 12.
44. Антитело по п.40, которое представляет собой одноцепочечное Fv антитело.
45. Антитело по п.40, которое представляет собой Fab фрагмент антитела.
46. Антитело по п.40, которое представляет собой Fab' фрагмент антитела.
47. Антитело по п.40, которое представляет собой (Fab')2 фрагмент антитела.
48. Изолированное антитело, специфически связывающее ангиопоэтин-2, содержащее тяжелую цепь и легкую цепь, причем тяжелая цепь включает вариабельный участок, содержащий аминокислотную последовательность SEQ ID NO. 11.
49. Изолированное антитело, специфически связывающее ангиопоэтин-2, содержащее тяжелую цепь и легкую цепь, причем легкая цепь включает вариабельный участок, содержащий аминокислотную последовательность SEQ ID NO. 12.
50. Изолированное антитело, специфически связывающее ангиопоэтин-2, содержащее вариабельный участок тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO. 11, или его антигенсвязывающий фрагмент, причем антигенсвязывающий фрагмент содержит, по меньшей мере, CDR1, CDR2 и CDR3 последовательности SEQ ID NO. 11, и вариабельный участок легкой цепи, содержащий аминокислотную последовательность SEQ ID NO. 12, или его антигенсвязывающий фрагмент, причем антигенсвязывающий фрагмент содержит, по меньшей мере, CDR1, CDR2 и CDR3 последовательности SEQ ID NO. 12
51. Изолированное антитело, специфически связывающее ангиопоэтин-2, содержащее:
a) каркасный участок тяжелой цепи, CDR1 участок тяжелой цепи, содержащий CDR1 последовательности SEQ ID NO. 11, CDR2 участок тяжелой цепи, содержащий CDR2 последовательности SEQ ID NO. 11, и CDR3 участок тяжелой цепи, содержащий CDR3 последовательности SEQ ID NO. 11, и
b) каркасный участок легкой цепи, CDR1 участок легкой цепи, содержащий CDR1 последовательности SEQ ID NO. 12, CDR2 участок легкой цепи, содержащий CDR2 последовательности SEQ ID NO. 12, и CDR3 участок легкой цепи, содержащий CDR3 последовательности SEQ ID NO. 12.
Текст
010726 Область техники Данное изобретение относится к специфическим связывающим агентам, которые распознают и связывают ангиопоэтин-2 (Ang-2). Более детально, изобретение относится к производству, диагностическому использованию и терапевтическому применению моноклональных и поликлональных антител и их фрагментов, которые специфически связывают Ang-2. Предпосылки создания изобретения Ангиогенез, формирование новых кровеносных сосудов из уже существующих, является обязательным для многих физиологических и патологических процессов. В норме ангиогенез строго регулируется с помощью про- и антиангиогенических факторов, но в случае таких заболеваний, как рак, глазные неоваскулярные заболевания, артриты и псориаз, этот процесс может пойти неправильно. Folkman, J., NatMed., 1:27-31 (1995). Существует целый ряд заболеваний, для которых известна их связь с нерегулируемым и нежелательным ангиогенезом. Эти патологии включают, но не ограничиваются указанными, глазные неоваскулярные патологии, такие как ретинопатии (включая диабетическую ретинопатию), старческая атрофия макулы; псориаз, гемиангиобластома, гемангиома, артериосклероз, воспалительные заболевания, такие как ревматоидные или ревматические воспалительные заболевания, особенно артриты (включая ревматоидный артрит), или другие хронические воспалительные заболевания, такие как бронхиальная астма,артериальный или посттрансплантационный атеросклероз, эндометриоз и неопластические болезни, например, так называемые солидные опухоли и гемопоэтические опухоли (такие как лейкемии и лимфомы). Другие заболевания, ассоциированные с нежелательным ангиогенезом, хорошо известны специалистам в данной области. Хотя многие сигнальные трансдуктивные системы задействованы в регуляции ангиогенеза, наиболее хорошо охарактеризованная и обладающая наилучшей избирательностью к клеткам эндотелия является система, включающая Tie-2 рецептор тирозинкиназы (известный как "Tie-2" или "Tie-2R"(также обозначаемый "ORK"); (мышиный Tie-2 также обозначается "tek") и его лиганды, ангиопоэтины (Gele,N.W. and Yancopoulos, G. D., Genes Dev, 13:1055-1066[1999]). Известно 4 ангиопоэтина; от ангиопоэтина 1 ("Ang-1") до ангиопоэтина-4 ("Ang-4"). Эти ангиопоэтины также обозначаются как "Tie-2 лиганды".Academy of Sciences of the USA, 96:1904-1909 [1999]; Witzenbichler, В., et al., J Biol Chem, 273:1851418521 [1998]). В то время как Ang-1, связываясь с Tie-2, стимулирует фосфорилирование рецептора в культуре эндотелиальных клеток, Ang-2, как было замечено, является агонистом и антагонистом фосфорилирования Tie-2 рецептора (Davis, S., et al., [1996], supra; Maisonpierre, P.C., et al., [1997], supra; Kim, I.,J.H. Kim, et al., Oncogene 19(39):4549-4552 (2000); Teichert-Kuliszewska, К., Р.С. Maisonpierre, et al., Cardiovascular Research 49(3): 659-70(2001. Фенотипы Ang-1 и Tie-2 нокаутированных мышей сходны и позволяют предположить, что стимулированное Ang-1 фосфорилирование Tie-2 опосредует ремоделирование и стабилизацию развивающихся сосудов в матке путем поддержания клеточной эндотелиальной адгезии. (Dumont, D. J., et al., GenesDevelopment, 8:1897-1909 [1994]; Sato, T. N., et al., Nature, 376:70-74 [1995]; Suri, C., et al., [1996], supra). Считается, что способность Ang-1 стабилизировать сосуды сохраняется у взрослых людей, у которыхAng-1 наиболее широко и конститутивно экспрессирован (Hanahan, D., Science, 277:48-50 [1997]; Zagzag,D., et al., Experimental Neurology, 159-391-400 [1999]). В противоположность этому, экспрессия Ang-2, в первую очередь, ограничена местами сосудистой ремодуляции, где, предполагается, он блокирует функционирование Ang-1, таким образом индуцируя состояние сосудистой пластичности, сопровождающей ангиогенез (Hanahan, D., [1997], supra; Holash, J., et al., Science, 284: 1994-1998 [1999]; Maisonpierre, P. C.,et al., [1997], supra). Многие опубликованные исследования наглядно продемонстрировали селективную сосудистую экспрессию Ang-2 при патологических состояниях, ассоциированных с ангиогенезом. Эти патологические состояния включают псориаз, атрофию макулы и рак. (Bunone, G., et al., American Journal of Pathology, 155:1967-1976 [1999]; Etoh, Т., et al., Cancer Research, 61:2145-2153 [2001]; Hangai, M., et al., Investigative OphtalmologyVisial Science, 42:1617-1625 [2001]; Holash, J., et al., [1999] supra; Kuroda, K., et al.,Journal of Investigative Dermatology, 116:713-720 [2001]; Otani, A., et al., Investigative Ophtalmologyal., [1999] supra). Большинство из этих исследований, в основном, посвящены изучению злокачественных опухолей, многие из которых, по-видимому, обнаруживают экспрессию сосудистого Ang-2. В противоположность тому, что экспрессия Ang-2 связана с патологическим ангиогенезом, его экспрессия в нор-1 010726 мальных тканях существенно ограничена (Maisonpierre, P.С., et al., [1997] supra; Mezquita, J., et al., Biochemical and Biophysical Research Communications, 260:492-498 [1999]). У здорового взрослого человека существует три основных органа, где происходит ангиогенез: яичники, плацента и матка; указанные органы являются основными у здоровых (т.е. не больных раком) людей, в тканях которых обнаружена матричная РНК (mRNA) Ang-2. Результаты определенных функциональных исследований позволяют предположить, что Ang-2 может участвовать в ангиогенезе, происходящем в злокачественных опухолях. Ahmad et al. (Cancer Res.,61:1255-1259 [2001]) описал сверхэкспрессию Ang-2 на модели ксенотрансплантации у мышей и показал,что она, скорее всего, ассоциируется с увеличением роста опухоли. См. также Etoh et al., supra и Tanaka etal., supra, которые в своих работах показали возможную связь сверхэкспрессии Ang-2 с опухолевой гиперваскуляризацией. Однако, в противоположность этому, Yu et al. (Am.J. Path, 158:563-570 [2001]) опубликовали данные, согласно которым сверхэкспрессия Ang-2 в клетках легочной карциномы Левиса и ТА 3 карциномы молочной железы, предположительно, продлевает продолжительность жизни мышей,которым были трансплантированы клетки вышеуказанных опухолей. В последние пять лет различные исследования предполагали, что Ang-1, Ang-2 и/или Tie-2 могут являться возможными мишенями противораковой терапии. Например, патенты США 6166185, 5650490,и 5814464 описывают анти-Tie-2 лигандные антитела и рецепторные антитела. Lin et al. (Proc. Natl. Acad.Sci USA, 95:8829-8834 [1998]) вводили аденовирус, экспрессирующий растворимый Tie-2, мышам; растворимый Tie-2, предположительно, уменьшал число и размер опухолей, развивающихся у этих мышей. В аналогичном исследовании, Lin et al. (J. Clin. Invest., 100: 2072-2078 [1997]) вводили растворимую форму Tie-2 крысам; это, предположительно, приводило к уменьшению размера опухолей у крыс. Siemeisteret al. (Cancer Res., 59:3185-3189 [1999]) получили клеточные линии меланомы человека, экспрессирующие внеклеточный домен Tie-2, инъецируя эти клеточные линии "голым" мышам, и пришли к выводу,что растворимый Tie-2, предположительно, приводит к "значительному ингибированию" опухолевого роста и опухолевого ангиогенеза. Обобщая эту информацию и данные о том, что и Ang-1, и Ang-2 связываются с Tie-2, из этих исследований не совсем ясно, каким образом Ang-1, Ang-2 или Tie-2 могут являться привлекательными мишенями для противораковой терапии. Как показано многими исследователями, слияние определенных белков или их фрагментов со стабильным белком плазмы, таким как константный участок Ig, увеличивает продолжительность полужизни этих белков [см., например, заявку WO 00/24782, опубл. 4 мая 2000 г.; патент США 5480981; Zheng et al.,J. Immunol., 154:5590-5600, (1995); Fisher et al., N. Engl. J. Med., 334:1697-1702, (1996); Van Zee, K. et al.,J. Immunol., 156:2221-2230, (1996); патент США 5808029 от 15 сентября 1998; Capon et al., Nature,337:525-531, (1989); Harvil et al., Immunotech., 1:95-105, (1995); WO 97/23614, опубл. 3 июля 1997;PCT/US 97/23183 от 11 декабря 1997; Linsley, J. Exp. Med., 174: 561-569, (1991); WO 95/21258, опубл. 10 августа 1995]. Эффективная анти-Ang-2-терапия должна помогать огромному количеству больных раком людей,поскольку многие солидные раки нуждаются в новообразовании сосудов от 1 до 2 мм в диаметре. Такая терапия должна находить широкое применение также в лечении других ассоциированных с ангиогенезом заболеваний, таких как ретинопатии, артриты и псориаз. Существует еще неразработанная возможность идентифицировать новые агенты, которые специфически распознают и связывают Ang-2. Такие агенты будут полезны для скриннинговых обследований и терапевтического внедрения у больных, страдающих заболеваниями, связанными с активностью Ang-2. Соответственно, предметом данного изобретения является способ получения специфических связывающих Ang-2 агентов, модулирующих его активность. Сущность изобретения Данное изобретение относится к антителам, содержащим тяжелую цепь и легкую цепь, причем указанная тяжелая цепь включает вариабельный участок, выбранный из группы, включающей а также его антигенсвязывающие фрагменты; легкая цепь включает вариабельный участок, выбранный из группы, включающей а также его антигенсвязывающие фрагменты. Изобретение также относится к специфическим связывающим агентам, содержащим по крайней мере один белок, выбранный из группы, включающей а также фрагменты данного белка. Специфическими связывающими агентами могут быть, например, антитела, поликлональные, моноклональные, химерные, гуманизированные или полные антитела человека. Антитела могут состоять и из одной цепи. Изобретение также относится к гибридоме, продуцирующей моноклональные антитела,соответствующие изобретению. Изобретение также относится к конъюгатам, раскрытым в описании. Этими соединениями могут быть, например, специфические связывающие агенты (скажем, антитела), заявленные в соответствии с настоящим изобретением. Изобретение также относится к молекулам нуклеиновой кислоты, кодирующим специфические связывающие агенты (например, антитела), заявленные в соответствии с настоящим изобретением, а также к вектору, содержащему такие молекулы нуклеиновой кислоты, и к клеткам-хозяевам, содержащим указанный вектор. Кроме того, изобретение относится к способу получения специфических связывающих агентов,включающему: (а) трансформацию клеток-хозяев, содержащих по крайней мере одну молекулу нуклеиновой кислоты, кодирующей специфический связывающий агент по п. 1; (б) экспрессию нуклеиновой кислоты в указанных клетках-хозяевах; и (с) выделение указанного специфического связывающего агента. Изобретение также относится к способу получения антител, включающему: (а) трансформацию клеток-хозяев, содержащих по крайней мере одну молекулу нуклеиновой кислоты, кодирующую антитело,согласно настоящему изобретению; (б) экспрессию нуклеиновой кислоты в указанных клетках-хозяевах; и (с) выделение указанного специфического связывающего агента. Помимо этого, изобретение относится к способу ингибирования нежелательного ангиогенеза у млекопитающих путем введения терапевтически эффективного количества специфических связывающих агентов, заявленных в соответствии с настоящим изобретением. Изобретение также относится к способу лечения рака у млекопитающих, заключающемуся во введении терапевтически эффективного количества специфических связывающих агентов, заявленных в соответствии с настоящим изобретением. Кроме того, изобретение относится к способу ингибирования нежелательного ангиогенеза у млекопитающих, заключающемуся во введении терапевтически эффективного количества антител, заявленных в соответствии с настоящим изобретением. Изобретение также относится к способу лечения рака у млекопитающих, заключающемуся во введении терапевтически эффективного количества антител, заявлен-3 010726 ных в соответствии с настоящим изобретением. Также изобретение относится к фармацевтическим композициям, содержащим специфические связывающие агенты, заявленные согласно настоящему изобретению, и дополнительный агент в фармакологически доступной форме. Фармацевтическая композиция может содержать антитела, заявленные в соответствии с настоящим изобретением, и дополнительный агент в фармакологически доступной форме. Изобретение относится к способу модулирования или ингибирования активности ангиопоэтина-2 путем введения одного или более специфического связывающего агента, заявленного в соответствии с настоящим изобретением. Изобретение также относится к способумодулирования или ингибирования активности ангиопоэтина-2 путем введения антител, заявленных в соответствии с настоящим изобретением. Изобретение также относится к способу модулирования сосудистой проницаемости или утечки плазмы в организме млекопитающих, заключающемуся во введении терапевтически эффективного количества специфических связывающих агентов, заявленных в соответствии с настоящим изобретением. Изобретение также относится к способу лечения глазных неоваскулярных заболеваний, ожирения, гемоангиобластомы, гемангиомы, артериосклероза, воспалительных заболеваний, воспалительных нарушений, атеросклероза, эндометриоза, неопластических заболеваний, заболеваний костной системы у млекопитающих, заключающемуся во введении терапевтически эффективного количества специфических связывающих агентов, заявленных в соответствии с настоящим изобретением. Изобретение также относится к способу модулирования сосудистой проницаемости или утечки плазмы в организме млекопитающих, заключающемуся во введении терапевтически эффективного количества антител, заявленных в соответствии с настоящим изобретением. Изобретение также относится к способу лечения глазных неоваскулярных заболеваний, ожирения, гемоангиобластомы, гемангиомы, артериосклероза, воспалительных заболеваний, воспалительных нарушений, атеросклероза, эндометриоза,неопластических заболеваний, заболеваний костной системы у млекопитающих, заключающемуся во введении терапевтически эффективного количества антител, заявленных в соответствии с настоящим изобретением. Кроме того, изобретение также относится к способу лечения рака у млекопитающих путем введения терапевтически эффективного количества специфических связывающих агентов, заявленных в соответствии с настоящим изобретением, и химиотерапевтического агента. Специалистам в данной области должно быть понятно, что специфический связывающий агент и химиотерапевтический агент не обязательно должны вводиться одновременно. Изобретение также относится к способу лечения рака у млекопитающих, заключающемуся во введении терапевтически эффективного количества антител, заявленных в соответствии с настоящим изобретением, и химиотерапевтического агента. Антитела и химиотерапевтический агент не обязательно должны вводиться одновременно. Изобретение также относится к специфическому связывающему агенту, содержащему область 1,определяющую комплементарность (CDR 1), выбранную из группы, включающей Изобретение также относится к специфическому связывающему агенту, содержащему область 2,определяющую комплементарность (CDR 2), выбранную из группы, включающей Изобретение также относится к специфическому связывающему агенту, содержащему область 3,определяющую комплементарность (CDR 3), выбранную из группы, включающей Изобретение также относится к молекуле нуклеиновой кислоты, кодирующей специфический связывающий агент, заявленный в соответствии с настоящим изобретением. Кроме того, изобретение относится к способу определения уровня ангиопоэтина-2 в биологических образцах, включающему: (а) взаимодействие специфического связывающего агента, заявленного в соответствии с настоящим изобретением, с биологическим образцом; и (б) определение продолжительности связывания специфического агента, заявленного в соответствии с настоящим изобретением, и биологического образца. Изобретение также относится к способу определения уровня ангиопоэтина-2 в биологических образцах, включающему: (а) взаимодействие антитела, заявленного в соответствии с настоящим изобретением, и биологического образца; и (б) определение продолжительности связывания антитела с биологическим образцом. Изобретение относится к способу ингибирования нежелательного ангиогенеза у млекопитающих,заключающемуся во введении терапевтически эффективного количества полипептида или композиций,описанных выше. Изобретение также относится к способу модулирования ангиогенеза у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или композиций, описанных выше. Изобретение также относится к способу ингибирования опухолевого роста, характеризующегося нежелательным ангиогенезом у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или композиций, описанных выше. Кроме того, изобретение относится к способу лечения рака у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или композиций, описанных выше, и химиотерапевтического агента. Наиболее подходящим химиотерапевтическим агентом может быть или 5-FU, или СРТ-11,или Taxotere. Однако возможно использование других подходящих химиотерапевтических агентов и других вариантов лечения рака. Необходимо отметить, что специфические связывающие агенты, заявленные в соответствии с настоящим изобретением, могут использоваться для лечения целого ряда заболеваний, ассоциированных с дерегулируемым и нежелательным ангиогенезом. К таким заболеваниям относятся, но не ограничиваются указанными, глазные неоваскулярные патологии, такие как ретинопатии (включая диабетическую ретинопатию и старческую атрофию макулы), псориаз, гемангиобластома, гемангиома, артериосклероз,воспалительные заболевания, такие как ревматоидные или ревматические воспалительные заболевания, в особенности артриты (включая ревматоидный артрит), или другие хронические воспалительные расстройства, такие как хроническая астма, артериальный или посттрансплантационный атеросклероз, эндометриоз и неопластические заболевания, например, так называемые твердые опухоли и гемопоэтические злокачественные заболевания ("жидкие" опухоли, такие как лейкемия). Другие заболевания, для терапии которых возможно применение специфических связывающих агентов, очевидны для специалистов в данной области. К таким заболеваниям относятся, но не ограничиваются указанными, ожирение,проницаемость сосудов, утечка плазмы и заболевания костной системы, включая остеопороз. Таким образом, изобретение также относится к способам лечения вышеуказанных заболеваний, ассоциированных с дерегулируемым и нежелательным ангиогенезом. Другие аспекты данного изобретения очевидны из описания.-6 010726 Краткое описание чертежей Фиг. 1 демонстрирует графическую зависимость размеров опухоли (ось у) от времени (ось х) при изучении опухолей мышей, обработанных антителами к Ang-2 (клон 533, 537 или 544), полученными в соответствии с настоящим изобретением, и контрольным антителами или фосфатным буферным раствором (PBS). Детали описаны в примерах. Фиг. 2 А, 2 В и 2 С иллюстрируют данные по эпитопному картированию (O.D. 370) полноразмерногоAng-2 человека (hAng-2), N-концевого участка Ang-2 и С-концевого участка Ang-2 соответственно, с помощью пептидных антител TN8-Con4-C, L1-7-N и 12-9-3-С, заявленных в соответствии с настоящим изобретением, а также с помощью контрольных пептидных антител Tie-2-Fc, С 2 В 8 или 5 В 12. Детали описаны в примерах. Детальное описание изобретения Указанный раздел включен в описание исключительно в технических целях и ни в какой степени не ограничивает сущности раскрытых в нем изобретений. Для получения рекомбинантной молекулы ДНК, белка и антител могут использоваться стандартные способы, так же, как и для культуры ткани и трансформации клеток. Ферментные реакции и способы очистки осуществляются стандартно, согласно приведенным инструкциям, или обычно используются известные в этой области методики, например описанные у Sambrook et al. (Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY [1989]), или раскрытые в настоящем изобретении. Если не указано иное, например специфические термины, в описании раскрываются номенклатурные обозначения и лабораторные методики аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, которые очевидны для специалистов в данной области. Стандартные способы могут использоваться для химического синтеза, химического анализа, получения, приготовления и доставки фармацевтических препаратов и лечения пациентов. Термины В соответствии с настоящим изобретением описанные ниже термины, если специальным образом не оговорено иное, имеют приведенное ниже значение. Термин "Ang-2" относится к полипептиду, как показано на фиг. 6 патента США 6166185 ("Tie-2 лиганд-2), или его фрагментам, а также к полипептидам, которые представляют собой аллельные варианты, сплайсинговые варианты, варианты, основанные на заменах, делециях и/или вставках аминокислот,производные, пептиды и полипептиды слияния и межвидовые гомологи указанного полипептида. Ang-2 полипептид может включать или не включать дополнительные концевые аминокислотные остатки, например, лидерные последовательности, направляющие последовательности, аминоконцевой метионин,аминоконцевой метионин и остаток лизина и/или аминокислотные "метки" или последовательности белков слияния, что зависит от способа получения полипептида Ang-2. Термин "биологически активный", когда используется совместно со специфическими связывающими Ang-1 и/или Ang-2 агентами, означает пептид или полипептид, имеющий по крайней мере одну активность, характерную для Ang-1 и/или Ang-2 специфического связывающего агента. Специфический связывающий агент Ang-2 может представлять собой агонист, антагонист, нейтрализующий или блокирующий агент, соответствующий по крайней мере одной биологической активности Ang-2. Понятие "специфический связывающий агент" означает молекулу, предпочтительно молекулу белка, которая связывает Ang-2 (а также его варианты и производные, как определено выше) с наибольшей аффиностью, чем другие ангиопоэтины. Специфическим связывающим агентом может быть белок, пептид, нуклеиновая кислота, углевод, липид или вещество с низкой молекулярной массой, преимущественно связывающееся с Ang-2. Согласно настоящему изобретению наилучшими специфическими связывающими агентами являются антитела, такие как поликлональные антитела, моноклональные антитела(mAb), химерные антитела, CDR-привитые антитела, мультиспецифические антитела, биспецифические антитела, каталитические антитела, гуманизированные антитела, человеческие антитела, антиидиотипические (anti-Id) антитела и антитела, которые могут быть помечены в растворимой и связанной форме, а также их фрагменты, варианты и производные, используемые как самостоятельно, так и в комбинации с другими аминокислотными последовательностями, обеспечиваемыми существующими технологиями. Такие технологии включают, но не ограничиваются указанными, ферментное расщепление, химическое расщепление, пептидный синтез и рекомбинантные методики. Специфические связывающие агенты Ang-2, заявленные в соответствии с настоящим изобретением, способны связывать участки Ang2, которые, например, модулируют, ингибируют или промотируют биологическую активность Ang-2 и/или другие активности, ассоциированные с Ang-2. Термин "поликлональные антитела" означает гетерогенную смесь антител, которые распознают и связывают различные эпитопы одного антигена. Поликлональные антитела могут быть получены из препаратов неочищенной сыворотки или могут быть очищены с помощью антигенной аффинной хроматографии или протеин А/протеин G аффинной хроматографии. Термин "моноклональные антитела" относится к антителам, кодируемым одной и той же молекулой нуклеиновой кислоты, которые могут вырабатываться определенной гибридомной или другой клеточной линией или синтезироваться в организме трансгенного млекопитающего. При этом каждое антитело спе-7 010726 цифически распознает один и тот же эпитоп антигена. Понятие "моноклональный" не ограничивается каким-либо определенным способом получения антитела, так же, как и не ограничивается антителами,продуцируемыми каким-либо определенным видом животного, например мышью, крысой и т.д. Термин "химерные антитела" относится к антителам, у которых участки тяжелых и/или легких цепей идентичны или гомологичны соответствующим последовательностям в антителах, выделенных из организма животных определенного вида и принадлежащих к определенному классу или подклассу, в то время как остальные участки цепей идентичны или гомологичны соответствующим последовательностям в антителах, выделенных из организма животных другого вида и принадлежащих к другому классу или подклассу. В объем изобретения также включены фрагменты указанных антител, которые обеспечивают желательную биологическую активность (т.е. способность специфически связывать Ang-2). См. патент США 4816567 и Morrison et al., Proc Natl. Acad Sci (USA), 81:6851-6855 [1985]. Термин "CDR-привитые антитела" означает антитела, в которых CDR одного антитела определенного вида млекопитающего или изотипа рекомбинантно встроен в каркасную область другого антитела того же или другого вида млекопитающего или изотипа. Термин "мультиспецифические антитела" означает антитела, имеющие вариабельные участки, которые распознают более чем один эпитоп одного или более антигена. Подклассом этого типа антител являются "биспецифические антитела", которые распознают два отдельных эпитопа одного и того же или разных антигенов."Каталитические" антитела относятся к антителам, в состав которых входят структуры, обладающие одной или более цитотоксической или, более точно, одной или более биологической активностью. Термин "гуманизированные антитела" относится к специфическому типу CDR-привитых антител, в которых каркасный участок получен из человеческих антител, но при этом каждая CDR заменена перенесенным участком, полученным из антител другого вида млекопитающего, например, мыши. ТерминCDR раскрывается ниже. Термин "полные человеческие" антитела относится к антителам, у которых как CDR, так и каркасная часть кодируются одной или более одной молекулой ДНК человека. Термин "антиидиотипические" антитела относится к любым антителам, которые специфически связываются с другими антителами, распознающими антигены. Для получения любого антиидиотипического антитела может быть использован любой способ, описанный здесь, для получения Ang-2 специфических антител, за исключением того, что указанные антиидиотипические антитела получают, например,иммунизацией животных Ang-2 специфическими антителами или их фрагментами, связывающими Ang2, а не самим Ang-2 полипептидом или его фрагментом. Термин "варианты", используемый здесь, относится к полипептидам, полученным из природной(или по крайней мере известной) аминокислотной последовательности связывающего агента путем вставок, делеций и/или замещений аминокислотных остатков. Варианты, используемые в соответствии с настоящим изобретением, включают белки слияния, как описано ниже. Понятие "производные" относится к связывающим агентам, которые были химически модифицированы определенным образом и отличаются от вариантов, полученных путем инсерции, делеций и/или замещения аминокислотных остатков природного связывающего агента. Понятие "специфическое связывание Ang-2" относится к способности специфических связывающих агентов (таких, как антитела или их фрагменты), заявленных в соответствии с настоящим изобретением,распознавать и связывать зрелый, полноразмерный или усеченный полипептид Ang-2 человека или его ортологи таким образом, что их аффинность (определяемая, например, методом аффинного иммуноферментного анализа или методом BIAcore, как описано здесь) или нейтрализующая способность (определяемая, например, методом нейтрализационного иммуноферментного анализа, как описано здесь) является по крайней мере в 10 раз больше, но предпочтительнее в 50 раз больше, 100, 250 или 500 раз больше или даже в 1000 раз больше, чем их аффинность или нейтрализующая способность в отношении другого ангиопоэтина, или другого пептида или полипептида. Термин "антигенсвязывающий домен" или "антигенсвязывающий участок" относится к области специфического связывающего агента (такого, как молекула антитела), которая содержит специфические аминокислотные остатки (или другие структуры) и взаимодействует с антигеном и определяет специфичность и аффинность специфического связывающего агента по отношению к антигену. В молекуле антитела антигенсвязывающий домен обычно обозначается как CDR ("область, определяющая комплементарность", от англ. complementarity-determining region). Термин "эпитоп" относится к тому участку любой молекулы, который может распознаваться и с которым может связываться один или более антигенсвязывающий участок специфического связывающего агента, например, антитела. Эпитоп обычно состоит из химически активных поверхностно сгруппированных молекул, таких как, например, молекулы аминокислот или боковые цепи молекулы углевода, и имеет специфическую трехмерную пространственную структуру и определенный заряд. Эпитопы, как описано здесь, могут быть смежными и несмежными. Кроме того, эпитопы могут быть миметиками в том смысле, что они включают трехмерную пространственную структуру, которая идентична таковой эпитопа, используемого для получения антител, или содержат только некоторые из аминокислотных ос-8 010726 татков, обнаруживаемых в Ang-2, который используется для стимуляции антительного иммунного ответа. Термин "ингибирующий и/или нейтрализующий эпитоп" относится к эпитопу, связывание которого со специфическим связывающим агентом, таким как антитело, приводит к утрате (или по крайней мере снижению) биологической активности молекулы, клетки или организма, содержащего данный эпитоп, invivo, in vitro, in situ. Согласно настоящему изобретению нейтрализующий эпитоп находится в биологически активном участке Ang-2 или связан с ним. В противоположность этому, термин "активирующий эпитоп" относится к эпитопу, связывание которого со специфическим связывающим агентом, заявленным в соответствии с настоящим изобретением, приводит к стимуляции или, по крайней мере, поддержанию биологической активности Ang-2. Термин "фрагмент антитела" относится к пептиду или полипептиду, который представляет собой меньше по размеру соединение, чем целое интактное антитело. Целые антитела содержат два функционально независимых фрагмента или участка: антигенсвязывающий участок, известный как "Fab", и карбоксильный терминальный кристаллизующийся участок, известный как "Fc"-участок. Fab-участок включает первый константный домен как тяжелой, так и легкой цепи (СН 1 и CL1) вместе с вариабельными участками как тяжелой, так и легкой цепи, которые связывают специфический антиген. Каждый вариабельный участок тяжелой и легкой цепей включает три области, определяющие комплементарность(CDRs), и каркас из аминокислотных остатков, который отделяет CDRs друг от друга. Fc-участок содержит второй и третий константные участки (СН 2 и СН 3) тяжелой цепи и задействован в активации комплемента и атаки фагоцитирующих клеток. У некоторых антител Fc и Fab участки разделены "шарнирной областью"; в зависимости от того, каким образом протеолитически расщепляется полноразмерное антитело, она может быть связана как с Fab, так и с Fc участком. Например, в результате расщепления антитела протеазой протамином образуется фрагмент, состоящий из шарнирной области и Fc-участка, а расщепление антитела пепсином приводит к образованию фрагмента, в котором шарнирная область связана с обоими Fab-участками одновременно. Поскольку два Fab-участка остаются ковалентно связанными после воздействия пепсина, образовавшийся фрагмент обозначается как F(ab')2 фрагмент.Fc-домен может иметь сравнительно большое время полужизни в сыворотке, в то время как Fab является короткоживущим. [Capon et al., Nature, 337:525-31 (1989)]. Будучи экспрессируемым как часть белка слияния, Fc-домен может приобретать более длительное время полужизни в сыворотке, а также проявлять такие функциональные особенности, как связывание Fc-рецептора, связывание белка А, связывание комплемента и, возможно, даже перенос через плаценту того белка, с которым он был слит. Fcучасток может быть природным участком или может быть изменен для улучшения определенных свойств антител, таких, как их терапевтические свойства и время циркуляции. Термин "вариабельный участок" или "вариабельный домен" относится к части легкой и/или тяжелой цепи антитела, обычно включающей приблизительно 120-130 аминоконцевых аминокислот в тяжелой цепи и около 100-110 аминоконцевых аминокислот в легкой цепи. Вариабельные участки сильно различаются по аминокислотным последовательностям даже в антителах животных одного вида. Вариабельный участок антитела обычно определяет специфичность связывания каждым определенным антителом своего определенного антигена. Вариабельность последовательностей наблюдается в областях,определяющих комплементарность (CDRs), в то время как более консервативные участки вариабельного домена называются каркасными (FR от англ. Framework Region). CDRs легких и тяжелых цепей содержат аминокислоты, которые в значительной степени определяют взаимодействие антитела с антигеном, тем не менее, аминокислоты в FRs участках могут значительно влиять на связывание/распознавание антигена, как обсуждается здесь ниже. Термин "легкая цепь", когда используется по отношению к антителу, относится сразу к двум различным типам цепей, обозначаемым каппаи лямбда , причем С-концевые участки практически одинаковы (константны) у легких цепей всех типов. Термин "тяжелая цепь", когда используется по отношению к антителу, относится сразу к пяти различным типам цепей, обозначаемым альфа, дельта, ипсилон, гамма и мю, которые также имеют константные С-концевые участки. Комбинации тяжелых и легких цепей дают начало пяти известным классам антител: IgA, IgD, IgE, IgG и IgM, соответственно, включая четыре известных подкласса IgG, обозначаемых как IgG1, IgG2, IgG3 и IgG4. Термин "природный", когда используется по отношению к биологическому материалу, такому, как молекулы нуклеиновых кислот, полипептиды, клетки-хозяева и т.д., характеризует то, что может быть обнаружено в природе и еще не модифицировано человеком. Термин "изолированный", используемый по отношению к Ang-2 или к специфическому связывающему агенту Ang-2, относится к соединению, которое не загрязнено по крайней мере одним контаминирующим полипептидом, или к соединению, которое обнаруживается в своем природном окружении и,предпочтительно, практически свободно от каких-либо контаминирующих полипептидов млекопитающих, способных препятствовать его терапевтическому и диагностическому применению. Термин "зрелый", когда он используется по отношению к Ang-2, анти-Ang-2-антителам или любому другому специфическому связывающему агенту Ang-2 белковой природы, относится к пептиду или по-9 010726 липептиду, у которого отсутствуют лидерные или сигнальные последовательности. Когда связывающий агент, заявленный в соответствии с настоящим изобретением, экспрессируется, например, в прокариотической клетке-хозяине, "зрелый" пептид или полипептид может также включать дополнительные аминокислотные остатки (но по-прежнему не иметь лидерной последовательности), такие как аминоконцевой метионин или один или более остаток метионина и лизина. Пептид или полипептид, полученный таким образом, может быть использован как с дополнительными аминокислотными последовательностями, так и без них; в последнем случае они удаляются. Термины "эффективное количество" и "терапевтически эффективное количество", когда они используются по отношению к специфическим связывающим агентам Ang-2, означают такое количество специфического связывающего агента, которое является полезным или необходимым для поддержания заметных изменений уровня одной или более биологических активностей Ang-2. Изменения могут выражаться как в повышении, так и в снижении уровня Ang-2-активности. Наиболее предпочтительным изменением является снижение Ang-2-активности. Специфические связывающие агенты и антитела. Используемый термин "специфический связывающий агент" относится к молекуле, которая обладает способностью специфично распознавать и связывать Ang-2, как описано здесь. Подходящими специфическими агентами могут быть, но не ограничиваются указанными, антитела и их производные, полипептиды и малые молекулы. Подходящие специфические связывающие агенты могут быть получены с помощью способов, известных специалистам в данной области. Показательный специфический связывающий агент полипептида Ang-2, заявленный в соответствии с настоящим изобретением, способен связывать определенное количество полипептида Ang-2 и, предпочтительно, модулировать активность и функцию такого полипептида. Специфические связывающие агенты, такие как антитела и их фрагменты, которые специфически связывают Ang-2-полипептид, входят в объем настоящего изобретения. Антитела могут быть поликлональными, включая моноспецифические поликлональные антитела, моноклональными (mAbs), рекомбинантными, химерными, гуманизированными, такими как CDRs-привитые антитела, человеческими, одноцепочечными, каталитическими, мультиспецифическими и/или биспецифическими, а также их фрагментами, вариантами и производными. Поликлональные антитела, направленные к Ang-2-полипептиду, обычно получают путем множественных подкожных или внутрибрюшинных инъекций животным (например, кроликам, хомякам, козам,овцам, лошадям, свиньям, крысам, песчанкам, морским свинкам, мышам или любым другим подходящим видам млекопитающих, впрочем как и видам, не являющимся млекопитающими) Ang-2 полипептида или его фрагмента совместно с адъювантом или без него. Такие адъюванты включают, но не ограничиваются указанными, полные или неполные адъюванты Фрейнда; минеральные гели, такие как гидроксид алюминия, и поверхностно-активные вещества, такие как лизолецитин, поверхностноактивные полиолы, полианионы, пептиды, масляные эмульсии, гемоцианин моллюска фиссуреллы и динитрофенол. БЦЖ (бациллы Кальметта-Герена, от англ. bacilli Calmette-Guerein) и Corynebacterium parvum могут потенциально использоваться в качестве адъювантов у человека. Часто оказывается полезным соединение антигенного полипептида с белком-носителем, который является иммунногенным для иммунизируемого вида животного, таким, как гемоцианин моллюска фиссуреллы, сывороточный альбумин,бычий тиреоглобулин или ингибитор трипсина, выделенный из бобов сои. Кроме того, для усиления иммунного ответа используются агрегирующие агенты, например, квасцы. После иммунизации у животных берут кровь и исследуют сыворотку на предмет наличия в ней антител к Ang-2-полипептиду; титр антител может быть определен с помощью методик, описанных здесь в примерах. Поликлональные антитела могут использоваться вместе с сывороткой, из которой они были выделены, или могут быть очищены от нее, с помощью, например, антигенной аффинной хроматографии или протеин А/протеин G аффинной хроматографии. Моноклональные антитела, направленные к Ang-2-полипептиду, могут быть получены, например(без ограничений указанным), с помощью гибридомной технологии или современной "фаговодисплейной" методики. Например, моноклональные антитела, заявленные в соответствии с настоящим изобретением, могут быть получены с помощью гибиридомной технологии, как описано у Kohler et al.,Nature 256:495 [1975]; технологии человеческой В-клеточной гибридомы [Kosbor et al., Immunol Today 4:72 (1983); Cote et al., Proc Natl. Acad Sci (USA) 80:2026-2030 (1983); Brodeur et al., Monoclonal AntibodyN.Y., pp. 77-96 (1985)]. Также настоящее изобретение предлагает гибридомные клеточные линии, продуцирующие моноклональные антитела, реагирующие с Ang-2-полипептидом. Когда используется гибридомная технология, могут применяться миеломные клеточные линии. Такие клеточные линии, используемые в методике гибридомного слияния, предпочтительно, не продуцируют антитела, обладают эффективной способностью к слиянию и ферментной недостаточностью, позволяющей им расти на определенных селективных средах, поддерживающих рост только необходимых клеток, полученных путем слияния (гибридом). Например, мышиные клеточные линии, используемые- 10010726 для слияния, представляют собой Sp-20, P3-X63/Ag8, P3-X63-Ag8.653, NS1/1.Ag41, Sp210-Ag14, FO,NSO/U, MPC-11, MPO11-X45-GTG1.7 и S194/5XXO Bul; крысиные клеточные линии, используемые для слияния, представляют собой R210.RCY3, Y3-Ag 1.2.3, IR983F и 4 В 210. К другим клеточным линиям,полезным для слияния, относятся U-266, GM1500-GRG2, LICR-LON-HMy2 и UC729-6. Гибридомы и другие клеточные линии, продуцирующие моноклональные антитела, также могут быть новыми и входить в объем настоящего изобретения. Фагово-дисплейная методика может также использоваться для получения моноклональных антител любого вида животного. Преимущественно, данная технология применяется для получения полноразмерных моноклональных антител человека, причем полинуклеотид, кодирующий одиночный Fab или Fv фрагмент антитела, экспрессируется на поверхности фаговой частицы. [Hoogenboom et al., J Mol Biol 227:381 (1991); Marks et al., J Mol Biol 222:581 (1991); см. также патент США 5885793)]. Каждый фаг может быть "скринирован" с помощью методов связывания, описанных здесь, для идентификации тех фрагментов антител, которые имеют аффинность к Ang-2. Таким образом, эти подходы имитируют иммунную селекцию путем представления набора антительных фрагментов на поверхности нитевидного бактериофага и последующую селекцию фага путем обеспечения связывания этих фрагментов с Ang-2. Подобная процедура описана в заявкеPCT/US98/17364, поданной Adams et al., раскрывающей выделение высокоаффинных и функционально агонистических фрагментов антител к MPL- и msk-рецепторам при использовании указанных подходов, согласно которым полный набор генов антител человека может быть создан путем клонирования природных перегруппированных человеческих V генов из лимфоцитов периферической крови, как уже раньше описано. [Mullinax et al., Proc Natl. AcadSci (USA) 87:8095-8099(1990)]. После того, как полинуклеотидные последовательности, кодирующие каждую цепь полноразмерного моноклонального антитела или Fab или Fv фрагмента (фрагментов), в соответствии с настоящим изобретением, идентифицированы, клетки-хозяева, как эукариотические, так и прокариотические, могут быть использованы для экспрессии полинуклеотидов моноклональных антител с помощью рекомбинантных технологий, хорошо известных и обычных для данной области. В противоположность этому,трансгенных животных получают путем встраивания в геном животного-реципиента, которым может быть, например, мышь, кролик, коза или корова, полинуклеотида, кодирующего нужный специфический связывающий агент, таким образом, что обеспечивается экспрессия молекулы полинуклеотида, кодирующей моноклональное антитело или другой специфический связывающий агент. В одном случае полинуклеотиды, кодирующие моноклональное антитело или другой специфический связывающий агент,могут быть лигированы с регуляторными последовательностями, специфичными для молочной железы млекопитающих, и такие химерные полинуклеотиды могут быть встроены в клетки зародышевой линии животного-мишени. Полученное таким образом трансгенное животное продуцирует необходимое антитело с молоком [Pollok et al., J Immunol Meth 231:147-157 (1999); Little et al., Immunol Today 8:364-370(2000)]. К тому же для экспрессии и получения Ang-2 специфических связывающих агентов, таких как моноклональные антитела, могут использоваться растения; в подходящие растения путем трансфекции переносят полинуклеотид, кодирующий моноклональные антитела или другие специфические связывающие агенты. Согласно другому аспекту настоящего изобретения моноклональные или поликлональные антитела или их фрагменты, полученные из других млекопитающих, кроме человека, могут быть "гуманизированы" или "химеризованы". Методы для химеризации нечеловеческих антител хорошо известны в данной области. (см. патенты США Nos. 5859205, 5585089 и 5693762). Гуманизация производится, например, с помощью технологий, также хорошо известных в данной области знаний [Jones et al., Nature 321:522-525(1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science 239:1534-1536 (1988)], например, путем замещения, по крайней мере, области, определяющей комплементарность (CDRs), в антителе грызуна соответствующими участками антител человека. Изобретение также относится к вариантам и производным таких антител человека, которые описаны здесь и хорошо известны специалистам в данной области. Изобретение также относится к полноразмерным антителам человека, которые связывают Ang-2 полипептид, а также к их фрагментам, вариантам и/или производным. Такие антитела могут быть получены с помощью "фагово-дисплейной" технологии, описанной выше. Кроме того, в случае отсутствия эндогенного синтеза иммуноглобулинов для их получения могут быть использованы трансгенные животные (например, мыши), способные продуцировать набор человеческих антител. Это может быть достигнуто путем иммунизации животных Ang-2-антигеном или его фрагментами, причем эти фрагменты имеют аминокислотные последовательности, специфичные для Ang-2. Такие иммуногены могут быть соединены с носителем, но это необязательно. См., например, Jakobovits et al., Proc Natl. AcadSci (USA),90:2551-2555 (1993); Jakobovits et al., Nature 362:255-258 (1993); Bruggermann et al., Year in Immuno, 7:33(1993). Согласно одной такой методике указанных трансгенных животных получают путем повреждения эндогенного локуса, кодирующего тяжелые и легкие цепи иммуноглобулинов, и втраивания вместо него в геном этих животных локуса, кодирующего человеческие протеины тяжелых и легких цепей. Частично модифицированные животные, не имеющие полного набора соответствующих модификаций, затем пе- 11010726 рекрестно скрещиваются для получения животных, имеющих все необходимые модификации иммунной системы. После введения иммуногена эти животные становятся способными продуцировать антитела с человеческими вариабельными участками, включая человеческие (а не, например, мышиные) аминокислотные последовательности, которые являются иммуноспецифичными для заданных антигенов. См. заявки РСТ Nos., PCT7US96/05928 и PCTVUS93/06926. Дополнительные методики описаны в патенте США 5545807, заявках РСТ Nos. PCT/US91/245, PCT/GB89/01207 и в ЕР 546073 В 1 и ЕР 546073 А 1. Человеческие антитела также могут быть получены путем экспрессии рекомбинантной ДНК в клеткаххозяевах или путем экспрессии в гибридомных клетках, как описано здесь. Трансгеноз может быть достигнут разными способами. См., например, Bruggeman et al., Immunol Today 17:391-7 (1996). Согласно одному подходу минилокус конструируют таким образом, что сегменты гена, перенесенные в клетки зародышевой линии, тесно прилегают друг к другу. За счет ограничений по размеру (т.е., в целом, менее 30 kb) полученные в результате этого минилокусы содержат ограниченное число различных генных сегментов, но все еще способны продуцировать большой спектр антител. Минилокусы, содержащие только последовательности ДНК человека, включая промоторы и энхансеры, полностью функциональны в организме трансгенных мышей. Для получения большого числа генных сегментов в организме трансгенных животных используют искусственные хромосомы дрожжей (YACs, от англ. yeast artificial chromosomes). YACs могут иметь размер от нескольких сот килобаз до 1 Mb и встраиваются в геном мыши (или любого другого подходящего животного) путем микроинъекции непосредственно в яйцеклетку или путем их переноса в эмбриональные линии стволовых клеток (ES). Обычно YACs переносятся в ES клетки путем липофекции очищенной ДНК или путем слияния дрожжевого сферопласта, при том что очищенная ДНК переносится в мицелий,и процедуру слияния осуществляют так же, как и при получении гибридом. Отбор необходимых ES клеток, несущих трансфицированную ДНК, производится путем включения в YAC любого селективного маркера, известного специалистам в области данных технологий. Альтернативой данному способу является использование векторов бактериофага Р 1, которые амплифицируют в бактериях-хозяевах Е. coli. В то время как эти векторы несут меньше встроенной ДНК, чем YACs, соответствующие клоны вырастают быстро и имеют высокую продуктивность, что позволяет осуществить прямую микроинъекцию векторов в яйцеклетку мыши. Показано, что использование наборов различных Р 1 векторов приводит к эффективной гомологичной рекомбинации. После того, как подходящая трансгенная мышь (или другое подходящее животное) была идентифицирована на предмет определения сывороточных уровней циркулирующих антител с помощью методики, известной из уровня техники (например, ELISA), трансгенную мышь скрещивают с мышью, у которой был разрушен эндогенный локус Ig. В результате получается потомство, у которого большинство Вклеток экспрессируют человеческие антитела. Согласно другой методике полноразмерный Ig локус животного замещают человеческим Ig локусом, после чего животное начинает экспрессировать только человеческие антитела. Еще один подход заключается в замещении участков локуса животного специфическими и соответствующими участками локуса человека. Обычно животные, полученные после этой процедуры, могут экспрессировать химерные антитела (в противоположность полным человеческим антителам), чья структура зависит от природы перенесенного в геном мыши Ig локуса. Человеческие антитела также могут быть получены путем экспозиции спленоцитов человека (Вили Т-клеток) антигену in vitro с последующим восстановлением экспонированных клеток в иммунокомпромиссных мышах, например SCID или nod/SCID. См. Brams et al., J Immunol, 160:2051-2058 [1998];Carballido et al., Nat Med, 6: 103-106 [2000]. Согласно одному подходу трансплантация человеческой эмбриональной ткани SCID мышам (SCID-hu) приводит к длительному гемопоэзу и созреванию человеческих Т-клеток. [McCune et al., Science 241:1532-1639 (1988); Ifversen et al., Sem Immunol 8:243-248(1996)]. Любой гуморальный иммунный ответ химерных мышей полностью зависит от развития Т-клеток в организме этих животных. [Martensson et al., Immunol 83:1271-179 (1994)]. Согласно другому подходу лимфоциты периферической крови человека интраперитонеально (или другим образом) трансплантируются SCID мышам. [Mosier et al., Nature 335:256-259 (1988)]. Когда трансплантируемые клетки обрабатывают праймирующим агентом, таким как Стафилококковый Энтеротоксин A (SEA) [Martensson et al.,Immunol 84:224-230 (1995)], или моноклональными антителами к CD40 человека [Murphy et al., Blood 86:1946-1953 (1995)], регистрируется высокий уровень продукции В-клеток. Альтернативно, конструируют полный набор синтетических тяжелых цепей антитела человека из неперестроенных сегментов V-гена путем объединения каждого VH-сегмента с D-сегментом выборочных нуклеотидов вместе с J-сегментом [Hoogenboom et al., J Mol Biol 227:381-388 (1992)]. Таким же образом, набор легких цепей создается путем объединения каждого V-сегмента человека с J-сегментом[Griffiths et al., EMBO J 13:3245-3260 (1994)]. Нуклеотиды, кодирующие полное антитело (т.е. оба вида цепей, легкие и тяжелые), объединяются в один одноцепочечный Fv-фрагмент и получившийся полинуклеотид связывают с полинуклеотидом, кодирующим малый белок оболочки нитевидного бактериофага. Когда этот белок слияния экспрессируется на поверхности фага, полинуклеотид, кодирующий специфическое антитело, идентифицируют методом селекции, используя иммобилизованный антиген.- 12010726 Согласно другому подходу антительные фрагменты конструируют как два Fab-фрагмента путем слияния одной цепи с фаговым протеином и помещения другой цепи в периплазму бактерий [Hoogenboom et al., Nucl Acids Res 19:4133-4137 [1999]; Barbas et al., Proc Natl. AcadSci (USA) 88:7978-7982(1991)]. Промышленное получение химерных, гуманизированных, CDR-привитых и полных антител человека или их фрагментов обычно осуществляется с помощью рекомбинантных технологий. Полинуклеотидные молекулы, кодирующие тяжелые и легкие цепи каждого антитела и их фрагменты, могут быть встроены в клетки-хозяева и экспрессированы с помощью технологий и процедур, описанных здесь. Предпочтительным способом является получение антител в клетках-хозяевах млекопитающих железы,таких как СНО-клетки. Детали данного метода описаны ниже. Партнеры-слияния специфических связывающих агентов. Еще одним объектом изобретения являются полипептиды, содержащие аминокислотные последовательности вариабельных доменов Ang-2-антител. Такие последовательности, как описанные здесь аминокислотные последовательности вариабельных участков тяжелых цепей, или как описанные здесь аминокислотные последовательности вариабельных участков легких цепей, могут быть слиты как по Nконцевому участку так и по С-концевому участку с одним или более доменом Fc-фрагмента IgG человека. Будучи соединенным с терапевтическим белком, таким как Fab-фрагмент Ang-2 специфического антитела, Fc-домен может приобретать более длительное время полужизни в сыворотке, а также проявлять такие функциональные особенности, как связывание Fc-рецептора, связывание белка А, связывание комплемента и даже возможно перенос через плаценту того белка, с которым он был слит [Capon et al., Nature, 337:525-531 (1989)]. В одном случае, шарнирная область антитела, СН 2- и СН 3-участки могут быть слиты как с Nконцевыми, так и с С-концевыми участками полипептидов специфических связывающих агентов, таких,как анти-Ang-2 Fab- или Fv-фрагментов (полученных, например, с помощью фагово-дисплейной методики), с помощью способов, хорошо известных специалистам в данной области. Полученный пептид слияния может быть очищен методом аффинной колоночной хроматографии с использованием протеина А или протеина G сефарозы. Пептиды и протеины, слитые с Fc-фрагментом, обнаруживают гораздо большее время полужизни in vivo, чем их "неслитые" аналоги. Также, слияние с Fc-фрагментом делает возможным димеризацию/мультимеризацию полипептида слияния. Fc-участок может быть природным участком или может быть изменен для улучшения определенных свойств антител, таких, как их терапевтические свойства, время циркуляции и способность к агрегации. Другие примеры, известные в данной области, включают слияние Fc-фрагмента, который может быть человеческим или принадлежать любому другому виду животного, а также синтетическим, с N-концевым участком CD30L с целью лечения болезни Ходжкина, анапластической лимфомы и Т-клеточной лейкемии (патент США 5480981), а также слияние Fc-фрагмента с TNF-рецептором с целью лечения септического шока [Fisher et al., N Engl J Med,334: 1697-1702 (1996)] и слияние Fc-фрагмента с Cd4-рецептором с целью лечения СПИДа [Capon et al.,Nature, 337:525-31 (1989)]. Каталитические антитела представляют собой другой тип слитых молекул и включают антитела, к которым присоединено одно или более соединений, обладающих цитотоксичностью, или, наиболее часто, одно или более биологически активных соединений. См., например, Rader et al., Chem Eur J 12:20912095 (2000). Цитотоксические соединения такого рода улучшают цитотоксичность, опосредованную антителами, и включают такие вещества, как цитокины, прямо или косвенно стимулирующие клеточную гибель, радиоизотопы, химиотерапевтические препараты (включая пролекарства), бактериальные токсины (например, рицин, гелонин и т.д.), химические соединения (например, маутанзиноидные токсины,калехемицин и т.д.), радиоконъюгаты, ферментные конъюгаты [конъюгаты РНК-азы, направляемая антителами ферментная/пролекарственная терапия (ADEPT, от англ. antibody-directed enzyme/prodrug therapy)] и т.д. Согласно одному подходу цитотоксический агент может быть "прикреплен" к одному компоненту биспецифического или мультиспецифического антитела путем связывания данного агента с одним из альтернативных сайтов распознавания антигена данного антитела. Как альтернатива, белковые цитотоксины могут экспрессироваться как белки слияния со специфическим связывающим агентом после слияния полинуклеотида, кодирующего токсин, с полинуклеотидом, кодирующим связывающий агент. Согласно другому подходу специфический связывающий агент может быть ковалентно модифицирован,чтобы он включал желаемый цитотоксин. Примерами таких белков слияния являются иммуногенные полипептиды, белки с длительным временем полужизни, такие как константные участки иммуноглобулинов, белковые маркеры, протеины и полипептиды, обеспечивающие очистку необходимого полипептида специфического связывающего агента, и полипептидные последовательности, обеспечивающие формирование мультимерных белков(таких, как лейциновые застежки-молнии, необходимые для формирования димеров и их стабильности). Такие инсерционные варианты обычно включают целую нативную молекулу первого полипептида или ее существенную часть, связанную с N- или С-концевыми участками целой молекулы или ее части второго полипептида. Например, белки слияния обычно содержат лидерные последовательности других видов, чтобы предотвратить рекомбинантную экспрессию белка в гетерологичном хозяине. Другой по- 13010726 лезный белок слияния включает дополнительный иммунологически активный домен, такой как эпитоп антитела, для того, чтобы обеспечить очистку белка слияния. Включение сайта расщепления по месту слияния или рядом с ним будет способствовать удалению постороннего полипептида после очистки. Другие полезные белки слияния включают функциональные домены, такие как активные сайты ферментов, домены гликозилирования, клеточные сигнальные молекулы или трансмембранные участки. Существует множество коммерчески доступных систем для экспрессии белков слияния, которые могут быть использованы в настоящем изобретении. Наиболее полезные системы включают, но не ограничиваются указанными, систему глутатион-S-трансферазы (GST) (Pharmacia), мальтозную систему связывания белка (NEB, Beverly, MA), FLAG систему (IBI, New Haven, СТ) и 6His систему (Qiaqen, Chatsworth, CA). Эти системы способны продуцировать рекомбинантные полипептиды, используя лишь небольшое количество дополнительных аминокислот, которые практически не влияют на антигенные свойства рекомбинантных полипептидов. Например, обе FLAG система и 6His система добавляют только короткие последовательности, причем известно, что каждая из них почти не обладает антигенными свойствами и не влияет значительным образом на восстановление нативной конформации полипептида. Другое рассматриваемое N-концевое слияние, которое может оказаться полезным, - это слияние дипептида"метионин-лизин" с N-концевым участком белка или полипептида. Такой вариант слияния может способствовать выгодному увеличению экспрессии и активности белка. Особенно полезным вариантом слияния может быть такая конструкция слияния, в которой пептид специфического связывающего агента соединяется с гаптеном с целью увеличения иммуногенности этого специфического связывающего агента; такая конструкция может оказаться полезной, например, для получения антиидиотипических антител согласно настоящему изобретению. Такие конструкции слияния хорошо знакомы специалистам в данной области, например, слияние специфического связывающего агента с хелперным антигеном, таким как hsp70, или белковой последовательностью, например, цепью дифтерийного токсина, или с цитокином, таким как ИЛ 2, и могут быть полезны для получения иммунного ответа. Согласно другому подходу может быть сделана такая конструкция слияния, которая будет способствовать увеличению нацеливания антигенных композиций специфического связывающего агента на специфическую область или сайт. Также рассматриваются другие конструкции слияния, включающие гетерологичные полипептиды с заданными свойствами, например, константный участок Ig для продления времени полужизни в сыворотке, или антитело или его фрагмент для нацеливания. Другие конструкции слияния используются для получения полипептидных гибридов, когда желательно отделить партнер слияния от нужного пептида. Согласно одному подходу партнер слияния связывается с рекомбинантным полипептидом специфического связывающего агента пептидной последовательностью, содержащей специфический сайт распознавания для протеазы. Примером подходящих последовательностей являются последовательности, распознаваемые протеазой вируса табачной мозаики (Life Technologies, Gaithersburg,MD) или Фактором Ха(New England Biolabs, Beverly, MA). Изобретение также относится к полипептидам слияния, содержащим целый вариабельный доменAng-2-антитела или его часть, таким, как вариабельный участок тяжелой цепи с аминокислотными последовательностями, как описано выше, или вариабельный участок легкой цепи с аминокислотными последовательностями, как также описано выше, в комбинации с усеченным тканевым фактором (tTF), нацеливающим агентом, состоящим из усеченной формы человеческого белка, индуцирующего свертывание, который действует как фактор свертывания в опухолевых сосудах. Слияние tTF с антителом к Ang-2 или его фрагментами может способствовать доставке анти-Ang-2-антител к клеткам-мишеням. Варианты специфических связывающих агентов. Варианты специфических связывающих агентов, заявленных в соответствии с настоящим изобретением, включают инсерционные, делеционные варианты и/или варианты, основанные на замещении. В одном аспекте изобретения инсерционные варианты подразумевают замещение одним или более аминокислотным остатком аминокислотных последовательностей специфических связывающих агентов. Инсерции могут быть локализованы как в одном, так и в обоих концевых участках белка, или могут быть расположены во внутренних участках аминокислотных последовательностей специфических связывающих агентов. Инсерционные варианты с дополнительными аминокислотными последовательностями, как в одном, так и в обоих концевых участках белка, могут включать, например, белки слияния и белки,включающие аминокислотные "ярлыки" и "метки". Инсерционные варианты включают полипептиды специфических связывающих агентов, в которых один или более аминокислотный остаток добавляется к аминокислотным последовательностям специфических связывающих агентов или их фрагментам. Варианты специфических связывающих агентов, заявленные в соответствии с настоящим изобретением, также включают зрелые специфические связывающие агенты. У таких специфических связывающих агентов удалена лидерная или сигнальная последовательность, тем не менее полученный в результате этого полипептид имеет дополнительные аминоконцевые остатки по сравнению с полипептидом Ang2 дикого типа. Вариантами специфических связывающих агентов являются полипептид с дополнительным остатком метионина в положении-1 (Мет-1-специфический связывающий агент) и полипептид с до- 14010726 полнительными метионином и лизином в положениях -2 и -1 (Мет-2-Лиз-1-специфический связывающий агент). Варианты специфических связывающих агентов с дополнительными остатками Мет, Мет-Лиз,Лиз (или с одним или более основными остатками) особенно полезны для увеличения рекомбинантной продукции в бактериальных клетках-хозяевах. Изобретение также охватывает специфические связывающие агенты, имеющие дополнительные аминокислотные остатки, которые являются результатом применения специфических экспрессирующих систем. Например, использование коммерчески доступных векторов, которые экспрессируют нужный полипептид как часть продукта слияния, полученного с помощью системы глутатион-S-трансферазы(GST), обеспечивает получение такого полипептида, имеющего дополнительный остаток глицина в аминокислотном положении-1, после того, как этот полипептид был отделен от компонента GST. Также рассматриваются варианты, полученные после экспрессии в других векторных системах, включая такие, у которых полигистидиновые "метки" встраиваются в аминокислотные последовательности, обычно в карбоксильный и/или аминоконцевой участок последовательности. Инсерционные варианты также включают белки слияния, описанные выше, у которых амино- и/или карбоксиконцевые участки полипептида специфического связывающего агента слиты с другим полипептидом или его фрагментом, или аминокислотными последовательностями, которые обычно не распознаются как часть какой-либо специфической белковой последовательности. Изобретение также относится к делеционным вариантам полипептидов специфических связывающих агентов, из которых удален один или более аминокислотный остаток. Делеции могут быть осуществлены как в одном, так и в обоих концевых участках полипептида специфического связывающего агента,а также могут заключаться в удалении одного или более аминокислотного остатка из аминокислотной последовательности специфического связывающего агента. Делеционные варианты непременно включают все фрагменты полипептида специфического связывающего агента, ответственные за его функцию. Антительные фрагменты включают те участки антитела, которые связываются с эпитопом антигенного полипептида. Примерами таких фрагментов являются Fab- и F(ab')2-фрагменты, полученные, например, путем ферментативного или химического расщепления полноразмерных антител. К другим связывающим фрагментам относятся, например, фрагменты, полученные с помощью рекомбинантных ДНКтехнологий, таких как экспрессия рекомбинантных плазмид, содержащих нуклеотидные последовательности, кодирующие вариабельные участки антител. Изобретение также охватывает полипептидные фрагменты Ang-2 специфического связывающего агента, причем эти фрагменты сохраняют способность специфически связывать Ang-2-полипептид. Фрагменты, содержащие по меньшей мере 5, 10, 15, 20, 25,30, 35, 40, 45 или 50 или более следующих друг за другом аминокислот белка или полипептида, заявленного в соответствии с настоящим изобретением, представлены здесь. Выделенные полипептидные фрагменты демонстрируют уникальные и специфические иммунологические свойства, характерные для специфического связывающего агента, заявленного в соответствии с настоящим изобретением. Полипептидные фрагменты, заявленные в соответствии с настоящим изобретением и имеющие желательные иммунологические свойства, могут быть получены любым из способов, хорошо известных и рутинных для специалистов в данной области. Изобретение также относится к вариантам специфических связывающих агентов, заявленных в соответствии с настоящим изобретением, основанным на замещении аминокислот. Варианты, основанные на замещении, обычно считаются "сходными" с первоначальным полипептидом или имеют определенную "процентную идентичность" с первоначальным полипептидом и включают такие полипептиды, у которых один или более аминокислотный остаток удален и замещен альтернативным остатком. С одной стороны, замены консервативны по своей природе, но тем не менее изобретение охватывает и неконсервативные замены. Идентичность и схожесть родственных полипептидов может быть легко подсчитана с помощью хорошо известных методов. Такие методы включают, но не ограничиваются указанными, описанные, например, в Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York (1988); Biocomputing: Informatics and Genome Projects Smith D.W., ed., Academic Press, New York (1993); ComputerCarillo et al., SIAM J. Applied Math., 48:1073 (1988). Предпочтительные методы для определения степени родства или процентной идентичности двух полипептидов разработаны таким образом, чтобы обеспечить наибольшее перекрывание между двумя тестируемыми последовательностями. Способы определения идентичности раскрыты в общедоступных компьютерных программах. Предпочтительные методы для определения идентичности двух последовательностей, основанные на компьютерных программах, включают, но не ограничиваются указанными,GCG программный пакет, включающий GAP (Devereux et al., Nucl Acid Res., 12:387 (1984); GeneticsBiol., 215:403-410 (1990. Программа BLASTX общедоступна и может быть получена в Национальном Центре Биотехнологической Информации (NCBI) и из других источников (BLAST Manual., Altschul et al.NCB/NLM/NIH Bethesda, MD 20894; Altschul et al., supra (1990. Хорошо известный алгоритм Смита Ва- 15010726 термана также может быть использован для определения идентичности. Некоторые схемы выравнивания для сравнения двух аминокислотных последовательностей могут привести к перекрыванию только короткого участка и эти небольшие выровненные участки могут иметь очень высокую степень идентичности, даже несмотря на то, что не существует значительного родства между двумя полноразмерными последовательностями. Соответственно, в определенных случаях, выборочный метод выравнивания (программа GAP) может привести к выравниванию, которое охватывает по меньшей мере 10% полной длины сравниваемого полипептида; т.е. по крайней мере 40 последовательно расположенных аминокислот из по крайней мере 400 аминокислот полноразмерного полипептида подлежат сравнению, по крайней мере 30 последовательно расположенных аминокислот из по крайней мере от 300 до около 400 аминокислот полноразмерного полипептида подлежат сравнению, по крайней мере 20 последовательно расположенных аминокислот из 200 до около 300 аминокислотных последовательностей подлежат сравнению и по крайней мере 10 последовательно расположенных аминокислот из около 100 до 200 аминокислотных последовательностей подлежат сравнению. Например, при использовании компьютерного алгоритма GAP (Genetics Computer Group, Universityof Wisconsin, Madison, WI) два полипептида, для которых необходимо определить процентную идентичность последовательностей, выравниваются для оптимального перекрывания их соответствующих аминокислотных последовательностей ("шаг перекрывания", как определено алгоритмом). В определенных вариантах штрафная функция за открытие щели (которая обычно рассчитывается как средняя 3 Х диагональ; "средняя диагональ" есть средняя величина диагонали матрицы сравнения, подлежащей использованию; "диагональ" - это очко или номер, определяемый для каждого предполагаемого перекрывания аминокислот с помощью определенной матрицы сравнения) и штрафная функция за расширение щели(которая обычно составляет 1/10 штрафной функции за открывания щели), так же, как и матрица сравнения, такая как РАМ 250 или BLOSUM 62, используется совместно с алгоритмом. В определенных случаях стандартная матрица сравнения (см. Dayhoff et al., Atlas of Protein Sequence and Structure, 5(3)(1978 для матрицы сравнения РАМ 250 [Henikoff et al., Proc. Natl. Acad. Sci. USA, 89:10915-10919 (1992) для матрицы сравнения BLOSUM 62] также используется алгоритмом. В определенных случаях параметры для сравнения полипептидных последовательностей включают следующее: Алгоритм: Needlemann et al., J. Mol. Biol., 48:443-453 (1970) Матрица сравнения: BLOSUM 62 из Henikoff et al., supra (1992) Штрафная функция: 12 Штрафная функция за длину щели: 4 Пороговая величина подобия: 0 Программа GAP с вышеуказанными параметрами может оказаться приемлемой. В определенных ситуациях вышеуказанные параметры могут являться недостаточными для сравнения полипептидов(вместе с отсутствием штрафной функции за конец щелей) при использовании алгоритма GAP. В определенных случаях параметры для сравнения последовательностей полипептидных молекул включают следующее: Алгоритм: Needlemann et al., supra (1970) Матрица сравнения: совпадения = +10, несовпадения = 0 Штрафная функция: 50 Штрафная функция за длину щели: 3 Программа GAP также может оказаться приемлемой для вышеуказанных параметров. Вышеуказанные параметры являются недостаточными для сравнения полинуклеотидных молекул. Другие показательные алгоритмы, gap opening penalties, gap extension penalties, матрицы сравнения,пороговые величины подобия и т.д. могут использоваться, в том числе и включенные в программное руководство, Wisconsin Package, Версия 9, сентябрь 1997. Какую программу предпочесть, будет очевидным для специалистов в этой области и будет зависеть от того специфического сравнения, которое необходимо осуществить, например, сравнения ДНК и ДНК, белка и белка, белка и ДНК; и, кроме того, даже сравнения заданных пар последовательностей (в этом случае предпочтительными являются GAP или BestFit программы) или одной последовательности и большой базы данных о последовательностях (в этой ситуации предпочтительно использовать FASTA или BLASTA). В соответствии с настоящим изобретением используется двадцать основных аминокислот и их производных. См. Immunology-A Synthesis (2nd Edition, E.S. Golub and D.R. Gren, Eds., Sinauer Associates,Sunderland, Mass. (1991, данные источники приведены здесь в качестве ссылки. Аминокислоты могут иметь как L-, так и D-стереоконфигурацию (кроме глицина, который не является ни D-, ни L-изомером), и полипептиды, и соединения, заявленные в соответствии с настоящим изобретением, могут содержать комбинации стереоизомеров. Однако L-стереоконфигурация является предпочтительной. Изобретение также относится к реверсивным молекулам, у которых последовательности аминоконцевого участка аминокислоты и последовательности карбоксиконцевого участка аминокислоты поменяны местами. Например, реверсивная молекула для молекулы, имеющей нормальную последовательность X1-Х 2-Х 3, будет выглядеть как X3-X2-X1. Изобретение также относится к ретро-реверсивным- 16010726 молекулам, у которых, как указано выше, аминоконцевой участок аминокислоты и карбоксиконцевой участок поменяны местами, и остатки, которые обычно являются "L"-энантиомерами, становятся "D"изомерами. Стереоизомеры (например, D-аминокислоты) двадцати основных аминокислот, неприродные аминокислоты, такие как -,-замещенные аминокислоты, N-алкилированные аминокислоты, молочная кислота и другие необычные формы аминокислот, также могут быть подходящими компонентами для полипептидов, заявленных в соответствии с настоящим изобретением. Примерами необычных аминокислот являются аминоадипиновая кислота, -аланин, -аминопропионовая кислота, аминомасляная кислота,пиперидиновая кислота, аминокапроновая кислота, аминогептановая кислота, аминоизомасляная кислота, аминопимелиновая кислота, диаминомасляная кислота, десмозин, диаминопимелиновая кислота,диаминопропионовая кислота, N-этилглицин, N-этиласпарагин, ксироксилизин, алло-ксироксилизин,гидроксипролин, изодесмозин, алло-изолейцин, N-метилглицин, саркозин, N-метилизолейцин, Nметилвалин, норвалин, норлейцин, оритин, 4-гидроксипролин, -карбоксиглутамат, -N,N,N,триметиллизин,-N-ацетиллизин, О-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин,5-гидроксилизин, -N-метиларгинин и другие подобные аминокислоты и аминокислоты (например, 4 гидроксипролин). Так же, если специально не указано иное, левый конец одноцепочечной полинуклеотидной последовательности является 5' концом; левостороннее направление двуцепочечной полинуклеотидной последовательности также рассматривается как 5' направление. Направление от 5' конца к 3' концу, в котором идет добавление транскриптов к образующейся цепи РНК, называется транскрипционным направлением; участки последовательности одной цепи ДНК, имеющие ту же самую структуру, что и РНК и соответствующие 5'-5' концевому участку РНК-транскрипта, называются "вышележащими последовательностями"("upstream seguences"); участки последовательности одной цепи ДНК, имеющие ту же самую структуру,что и РНК и соответствующие 3'-3' концевому участку РНК-транскрипта, называются "нижележащими последовательностями" ("downstream seguences"). Консервативные аминокислотные замены могут относится к неприродным остаткам, которые обычно включаются в структуру белка путем химического пептидного синтеза, по сравнению с синтезом в биологических системах. Сказанное относится к пептидомиметикам и другим белкам, содержащим реверсивные или инвертированные формы аминокислотных остатков. Существующие в природе остатки могут быть разделены на классы, основываясь на свойствах их боковых цепей: 1) гидрофобные: Met, Ala, Val., Leu, Ile; 2) нейтрально гидрофильные: Cys, Ser, Thr, Asn, Gln; 3) кислые: Asp, Glu; 4) основные: His, Lys, Arg; 5) остатки, влияющие на пространственную ориентацию цепей: Gly, Pro; и 6) ароматические: Trp, Tyr, Phe. Например, неконсервативные замены могут включать замену представителя одного такого класса представителем другого класса. Такие замещенные остатки могут быть встроены в участки антител человека, гомологичные нечеловеческим антителам, или в негомологичные участки молекулы антитела. При производстве таких замен, согласно определенным подходам, должен учитываться гидропатический индекс аминокислот. Для каждой аминокислоты установлен гидропатический индекс на основании ее заряда и гидрофобности. Вот эти индексы: изолейцин (+4.5); валин (+4.2); лейцин (+3.8); фенилаланин (+2.8); цистеин/цистин (+2.5); метионин (+1.9); аланин (+1.8); глицин (-0.4); треонин (-0.7); серин(-0.8); триптофан (-0.9); тирозин (-1.3); пролин (-1.6); гистидин (-3.2); глутамат (-3.5); глутамин (-3.5); аспартат (-3.5); аспарагин (-3.5); лизин (-3.9) и аргинин (-4.5). Роль гидропатического индекса в обеспечении биологической функции взаимодействия белков понятно из уровня техники. Kyte et al., J. Mol. Biol., 157:105-131 (1982). Известно, что определенные аминокислоты могут быть заменены другими аминокислотами, имеющими такой же гидропатический индекс; при этом белок будет сохранять свою биологическую активность. Согласно одному подходу при производстве замен, основанных на гидропатическом индексе, в такие замены включают аминокислоты,чей гидропатический индекс лежит в диапазоне от -2 до +2. Другие подходы включают замены аминокислот, чей индекс лежит в диапазоне от -1 до +1; третьи - замены аминокислот с индексом от -0.5 до+0.5. Также из области техники известно, что замена схожих аминокислот будет эффективной, если она основывается на гидрофильности этих аминокислот, особенно в тех случаях, когда биологически функциональный белок или пептид, полученный таким образом, предназначен для использования в иммунологических целях, как в настоящем изобретении. В определенных случаях наибольшая локальная гидрофильность белка, складывающаяся из гидрофильности составляющих его аминокислот, коррелирует с иммуногенностью и антигенностью, т.е. биологическими свойствами этого белка. Приводим результаты оценки гидрофильности для следующих аминокислотных остатков: аргинин(-1.3); валин (-1.5); лейцин (-1.8); иэолейцин (-1.8); тирозин (-2.3); фенилаланин (-2.5) и триптофан (-3.4). При производстве замен, основанных на одинаковой гидрофильности, при определенных подходах, производятся замены аминокислот, чья гидрофильность варьирует в пределах 2; при определенных подходах, в замены включают аминокислоты гидрофильностью 1, при других подходах - аминокислоты с гидрофильностью 0.5. Возможно также идентифицировать эпитопы в основной аминокислотной последовательности, основываясь на гидрофильности. Такие участки также называются "основными участками эпитопа". Примеры аминокислотных замен приведены в табл. 1 Таблица 1 Аминокислотные замены Специалист без труда сможет выделить подходящие варианты полипептида, как будет показано ниже, используя хорошо известные технологии. В определенных случаях специалист может также идентифицировать подходящие участки молекулы, которые могут быть заменены участками-мишенями, не влияющими на активность, соответственно, без нарушения активности данной молекулы. В определенных случаях специалист может идентифицировать остатки и части молекулы, которые являются постоянными у сходных полипептидов. При определенном подходе даже участки, являющиеся необходимыми для биологической активности или сохранения структуры молекулы, могут быть задействованы в консервативных заменах без нарушения биологической активности и без серьезного влияния на структуру полипептида. Кроме того, специалист может провести структурно-функциональные исследования для идентификации тех остатков сходных полипептидов, которые являются необходимыми для активности и структуры полипептида. На основе такого сравнения можно предсказать важность аминокислотных остатков в белке и соотнести их с аминокислотными остатками, имеющими существенное значение для активности и структуры сходного белка. Специалист в данной области может выбирать, на основе такой предсказан- 18010726 ной значимости аминокислотных остатков, химически сходные остатки, которые подлежат замене. Специалист в этой области также может анализировать трехмерную пространственную структуру и аминокислотную последовательность сходных полипептидов. На основе таких данных специалист в данной области может предсказывать расстановку аминокислотных остатков в молекуле антитела, зависящую от трехмерной пространственной структуры антитела. При таком подходе специалист в данной области может не делать радикальных изменений в аминокислотных остатках, для которых предсказана их локализация на поверхности белка, поскольку эти остатки могут быть вовлечены в важное взаимодействие с другими молекулами. Кроме того, специалист в данной области может создавать тестовые варианты, содержащие определенную аминокислотную замену по каждому аминокислотному остатку. Эти варианты могут быть потом скринированы с использованием методик, хорошо известных специалистам в данной области. Такие тестовые варианты могут использоваться для получения информации о подходящих вариантах. Например, можно выяснить, что замена определенного аминокислотного остатка приводит к нарушенной, нежелательно сниженной или неподходящей активности; варианты с такой заменой не будут использоваться. С другой стороны, основываясь на информации, полученной с помощью таких рутинных экспериментов, специалист в данной области может точно определить аминокислоты, замещения которых в дальнейшем следует избегать как самих по себе, так и в сочетании с другими мутациями. Многочисленные научные публикации были посвящены предсказанию вторичной структуры. См.(1978); Chou et al., Ann. Rev. Biochem., 47:251-276 и Chou et al., Biophys. J., 26:367-384 (1979). Кроме того,постоянно доступны специальные компьютерные программы, позволяющие предсказать вторичную структуру. Один метод предсказания вторичной структуры базируется на гомологичном моделировании. Например, два полипептида или белка, имеющие последовательности, совпадающие более чем на 30% или совпадающие более чем на 40%, очень часто имеют общую структурную топографию. Недавнее расширение базы данных о структуре белков позволило увеличить предсказуемость вторичной структуры, включая потенциальное число складок в структуре белков или полипептидов. См. Holm et al., Nucl.(1997, что существует ограниченное количество складок в заданном полипептиде или белке, и установление критического числа таких складок позволит предсказывать структуру белка или полипептида намного более точно. Дополнительные методы предсказания вторичной структуры включают "нанизывание" ("threading")Gribskov et al., Proc. Natl. Acad. Sci., 84 (13):4355-4358 (1987 и анализ "эволюционной связи" (См. Holm,supra (1999), and Brenner, supra (1997. В определенных случаях варианты антител включают гликозилированные антитела, в которых число и/или тип сайта гликозилирования изменено по сравнению с аминокислотной последовательностью исходного полипептида. В частности, белковые варианты включают большее или меньшее число Nсвязанных сайтов гликозилирования по сравнению с нативным белком. N-связанный сайт гликозилирования характеризуется последовательностью: Asn-X-Ser или Asn-X-Thr, при этом аминокислотный остаток, обозначенный как X, может представлять собой любую аминокислоту, за исключением пролина. Замена аминокислотных остатков для создания таких последовательностей обеспечивает потенциально новый сайт для добавления N-связанной углеводной цепи. Альтернативно, замена, которая элиминирует такую последовательность, будет удалять существующую N-связанную углеводную цепь. Изобретение также предусматривает реорганизацию N-связанных углеводных цепей, при этом один или более Nсвязанный сайт гликозилирования (обычно тот, который существует в природе) элиминируется, а вместо него один или более N-связанный сайт создается. Дополнительные предпочтительные варианты антител включают цистеиновые варианты, в которых один или более остаток цистеина удален или заменен другим аминокислотным остатком (например, серином) по сравнению с аминокислотной последовательностью исходного белка. Цистеиновые варианты могут оказаться полезными в тех случаях, когда должна быть восстановлена конформация антител, обеспечивающая их биологическую активность, например,после выделения нерастворимых телец включения. Цистеиновые варианты в общем имеют меньше цистеиновых остатков по сравнению с нативным белком, и обычно их четное число для того, чтобы минимизировать взаимодействия, обусловленные неспаренными цистеиновыми остатками. Согласно определенному подходу аминокислотные замены: (1) снижают восприимчивость к протеолизу, (2) сокращают восприимчивость к окислению, (3) меняют связывающую аффинность для формирования белковых комплексов, (4) изменяют связывающую аффинность и/или (5) придают новые свойства или изменяют функциональные свойства таких полипептидов. Согласно такому подходу одиночные или множественные аминокислотные замены (в определенных вариантах консервативные аминокислотные замены) могут быть произведены в природно-существующих последовательностях (в определенных случаях, в участке полипептида вне домена(ов), формирующих межмолекулярные контакты). В определенных случаях консервативные аминокислотные замены могут существенно не изменять структурные- 19010726 характеристики исходной последовательности (например, перемещение аминокислот не должно приводить к нарушению спирали, характерной для исходной последовательности, а также не должно изменять другие типы вторичной структуры, характерные для исходной структуры). Примеры хорошо известных из области техники вторичных и третичных структур описаны в следующих работах: "Proteins, Structuresal. Nature 354:105 (1991). Молекулы специфических связывающих агентов, заявленные в соответствии с настоящим изобретением, представляющие собой пептидные или полипептидные варианты замен, могут иметь от 10 до 20% замещенных оригинальных аминокислотных последовательностей. Для вариантов антител тяжелая цепь может иметь 50, 49, 48, 47, 46, 45, 44, 43, 42, 41, 40, 39, 38, 37, 36, 35, 34, 33, 32, 31, 30, 29, 28, 27, 26,25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 замещенную аминокислоту, в то время как легкая цепь может иметь 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9,8, 7, 6, 5, 4, 3, 2 и 1 замещенную аминокислоту. Производные специфических связывающих агентов Изобретение также относится к производным полипептидов специфически связывающих агентов. Производные включают полипептиды специфических связывающих агентов, имеющие модификации в виде инсерций, делеций или замен аминокислотных остатков. Предпочтительно модификации являются ковалентными по своей природе, и включают, например, химическое связывание с полимерами, липидами, другими органическими и неорганическими молекулами. Производные полипептидов специфических связывающих агентов, полученные в соответствии с настоящим изобретением, могут обладать увеличенным временем полужизни в сыворотке или обладать повышенной тропностью к нужным клеткам,тканям или органам. Изобретение также охватывает производные связывающих агентов, ковалентно модифицированные для включения одного или более водорастворимого полимера, такого, как полиэтиленгликоль, полиоксиэтиленгликоль или полипропиленгликоль, как описано в патентах США Nos: 4640835, 4496689, 4301144,4670417, 4791192 и 4179337. Другие хорошо известные специалистам полимеры включают монометоксиполиэтиленгликоль, декстран, целлюлозу или другие полимеры, поли-(N-винил пирролидон)полиэтиленгликоль, пропиленгликоль, гомополимеры, сополимер полипропилен-оксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерол) и поливиниловый спирт, так же как смеси указанных полимеров. Особенно предпочтительными продуктами являются специфические связывающие агенты, ковалентно модифицированные субъединицами полиэтиленгликоля (PEG). Водорастворимые полимеры могут быть присоединены к специфическим участкам, например, к аминоконцу полипептида специфического связывающего агента, или могут быть произвольно распределены на боковых цепях полипептида. Использование PEG для улучшения терапевтической активности специфических связывающих агентов и, в особенности, гуманизированных антител, описано в патенте США 6133426,Gonzales et al., от 17 октября 2000 г. Сайты-мишени для мутагенеза антител Для манипулирования присущими Ang-2 специфическому антителу свойствами, такими как, например, аффинность антитела к клетке-мишени, могут быть использованы определенные подходы. Эти подходы включают применение сайт-специфического или произвольного мутагенеза полинуклеотидной молекулы, кодирующей антитело, с целью получения вариантов антител, с последующей стадией скрининга для отбора вариантов антител, которые обнаруживают нужные изменения, т.е. увеличенную или сниженную аффинность. К аминокислотным последовательностям, наиболее часто являющимся мишенями для мутагенеза, в первую очередь относятся последовательности CDRs. Как описано выше, именно эти участки содержат остатки, непосредственно взаимодействующие с Ang-2, и аминокислоты, влияющие на пространственную организацию этих остатков. Кроме того, показано, что аминокислоты в каркасных участках вариабельных доменов, расположенные вне CDR участков, также могут вносить существенный вклад в антигенсвязывающие свойства антитела и поэтому могут быть мишенями мутагенеза для изменения этих свойств. См. Hudson, Curr Opin Biotech, 9:395-402 (1999) и ссылки к ним. Меньшие и более эффективно скринируемые библиотеки вариантов антител могут быть получены путем ограничивающего случайного или сайт-специфического мутагенеза в отношении сайтов в CDRs,которые соответствуют областям, склонным к "гипермутировнию" в процессе созревания соматической аффинности. См. Chowdhury и Pastan, Nature Biotech, 17:568-572 [1999] и ссылки к ним. К типам элементов ДНК, для которых известна их способность определять сайты гипермутагенеза, относятся прямые и инвертированные повторы, определенные согласованные последовательности, втроичные структуры и палиндромы. Согласованные последовательности ДНК включают четырехосновную последовательность Пурин-G-Пиримидин - А/Т (т.е. А или G-G-C или Т-А или Т) и сериновый кодон AGY (в котором вместоY может быть С или Т). Таким образом, подходы, заявленные в соответствии с настоящим изобретением, включают стратегии мутагенеза, имеющие своей целью увеличить аффинность антитела к его мишени. Эти стратегии- 20010726 включают мутагенез целых вариабельных тяжелых и легких цепей, мутагенез только CDR участков, мутагенез согласованных сайтов гипермутаций с CDRs, мутагенез каркасных участков, а также различные комбинации этих вариантов ("мутагенез" в этом случае может быть произвольным или сайтспецифичным). Точное описание CDR участков и идентификация остатков, входящих в антигенсвязывающий участок антитела, может быть достигнута путем определения структуры заданного антитела и комплекса антитело-лиганд с помощью методов, хорошо известных специалистам в данной области, таких, как, например, рентгеновская кристаллография. Различные методы, основанные на анализе и характеристике кристаллических структур антител, хорошо известны специалистам в данной области и могут быть применены для приблизительного определения структуры CDR участков. Примерами таких широко применяемых методов являются методы Kabat, Chothia, AbM и контактное определение. Метод Kabat основан на вариабельности последовательностей и является наиболее часто применяемым методом определения CDRs [Johnson and Wu, Nucleic Acids Res, 28:214-8 (2000)]. Метод Chothia основан на определении местонахождения структурных петельных участков. [Chothia et al., J Mol Biol.,196: 901-17(1986); Chothia et al., Nature, 342: 877-83 (1989)]. Метод AbM является компромиссом между методами Kabat и Chothia. AbM - это интегральный набор программ для моделирования структуры антител, разработанный Оксфордской Молекулярной Группой (Oxford Molecular Group) [Martin et al., ProcNatl Acad Sci (USA) 86:9268-9272 (1989); Rees et al., ABM - компьютерная программа для моделирования вариабельных участков антител, Oxford, UK; Oxford Molecular, Ltd.]. AbM создает модели третичной структуры антитела из первичных последовательностей, используя комбинации известных баз данных и методов. Дополнительный метод определения, известный как метод контактного определения, был представлен выше. [MacCallum et al., J Mol Biol, 5:732-45 (1996)]. Этот метод основан на анализе доступных комплексов кристаллических структур. Принято, что CDR участки в тяжелых цепях обычно обозначаются H1, H2 и Н 3 и нумеруются последовательно от аминоконцевого участка к карбоксиконцевому участку. CDR участки в легких цепях обычно обозначаются L1, L2 и L3 и последовательно нумеруются от аминоконцевого участка и карбоксиконцевому.CDR-H1 обычно имеет примерно от 10 до 12 остатков в длину и обычно начинается через 4 аминокислотных остатка после Cys, в соответствии с данными Chothia и AbM, или через 5 аминокислотных остатка после Cys, согласно Kabat. За H1 обычно следует Trp, чаще Trp-Val, то также может быть последовательность Trp-Ile или Trp-Ala. Длина H1 приблизительно от 10 до 12 аминокислотных остатков согласно AbM, в то время как Chothia исключает 4 последних остатка.CDR-H2 обычно начинается через 15 аминокислотных остатков после окончания H1, в соответствии с данными Kabat и AbM. Остатками, предшествующими Н 2, обычно являются Leu-Glu-Trp-Ile-Gly,хотя существует множество вариантов. После Н 2 обычно следует аминокислотная последовательностьLys/Arg-Leu/Ile/Val/Phe/Thr/Ala-Thr/Ser/Ile/Ala. Согласно Kabat длина H1 составляет приблизительно от 16 до 19 остатков, в то время как по данным AbM она обычно включает от 9 до 12 остатков.CDR-H3 участок обычно начинается через 33 аминокислотных остатка после окончания Н 2 и обычно ему предшествует аминокислотная последовательность Cys-Ala-Arg. После Н 3 обычно следует аминокислотный остаток Gly. Длина Н 3 может быть любой, в промежутке от 3 до 25 аминокислотных остатков.CDR-L1 обычно начинается приблизительно после 24-го аминокислотного остатка сразу за Cys. Остатком, следующим за CDR-L1, всегда является Trp и с него начинаются последовательности Trp-TyrGln, Trp-Leu-Gln, Trp-Phe-Gln или Trp-Tyr-Leu. Длина CDR-L1 обычно составляет от 10 до 17 аминокислотных остатков. Структура CDR-L1 для антител, заявленных в соответствии с настоящим изобретением,полностью соответствует указанной схеме, поскольку этот участок содержит 15 аминокислот и следует за Cys, а за ним идет последовательность Trp-Tyr-Gln.CDR-L2 начинается приблизительно через 16 аминокислотных остатков после окончания L1. Он следует за аминокислотными последовательностями Ile-Tyr, Val-Tyr, Ile-Lys или Ile-Phe. Длина CDR-L2 составляет приблизительно 7 аминокислотных остатков.CDR-L3 обычно начинается через 33 аминокислотных остатка после окончания L2 и обычно следует за Cys. После L3 обычно следуют аминокислотные последовательности Phe-Gly-XXX-Gly. ДлинаCDR-L3 составляет приблизительно от 7 до 13 аминокислотных остатков. Различные способы для модификации антител были описаны в уровне техники. Например, патент США 5530101 (Queen et al., June 25, 1996) описывает методы получения гуманизированных антител, в которых последовательность каркасного участка вариабельного домена тяжелой цепи гуманизированного иммуноглобулина на 65-95% идентична аналогичной последовательности донорного иммуноглобулина. Каждая цепь гуманизированного иммуноглобулина обычно содержит, в дополнение к CDRs, аминокислоты, взятые из каркасного участка донорного иммуноглобулина, которые,например, способны взаимодействовать с CDRs для изменения связывающей аффинности; это такие аминокислоты, которые располагаются точно рядом в CDR участке донорного иммуноглобулина, или аминокислоты, чей размер предсказывается вплоть до 3 ангстремов с помощью молекулярного моделирования. Как тяжелые, так и легкие цепи могут быть созданы с помощью использования одного или не- 21010726 скольких различных позиционных критериев. В комбинации с интактными антителами гуманизированные иммуноглобулины, заявленные в соответствии с настоящим изобретением, могут быть по большей части неимуногенными в человеческом организме, но сохранять такую же аффинность к антигенам, таким, как белки и другие соединения, содержащие эпитоп, как и донорные иммуноглобулины. См. также родственные методики, изложенные в патенте США 5693761, Queen et al., от 2 декабря 1997 ("Полинуклеотиды, кодирующие гуманизированные иммуноглобулины"); патенте США 5693762, Queen et al., от 2 декабря 1997 ("Гуманизированные иммуноглобулины"); патенте США 5585089, Queen et al., от 17 декабря 1996 ("Гуманизированные иммуноглобулины"). В одном примере, см. патент США 5565332, Hoogenboom et al., от 15 октября 1996 ("Получение химерных антител - комбинаторный подход") описываются методы получения антител, фрагментов антител, которые имеют такую же специфичность связывания, как и исходное антитело, но в большей степени сходны с антителами человека. Гуманизированные антитела получают с помощью комбинирования цепей, используя, например, фагово-дисплейную технологию, причем полипептид, содержащий тяжелые или легкие цепи с вариабельными доменами антитела нечеловеческого происхождения, специфического к заданному антигену, комбинируется с набором комлементарных вариабельных доменов (легкой или тяжелой) цепи антител человека. Гибридные пары, которые являются специфичными для заданного антигена, идентифицируются, а цепи антител человека из выборочных пар комбинируются с набором комплементарных вариабельных доменов (легкой или тяжелой) цепи антител человека. Согласно другому подходу часть CDR участка антитела нечеловеческого происхождения комбинируется с набором частейCDR участка антител человека. Из полученной библиотеки полипептидных димеров антител отбирают гибриды и используют их на второй стадии гуманизирования путем комбинирования цепей. Альтернативно, указанная вторая стадия может опускаться, если гибрид уже обладает в достаточной степени свойствами антител человека, чтобы было возможно его терапевтическое использование. Методы модификации антител с целью придания им свойств человеческих антител также описаны. См. также Winter,FEBS Letts 430:92-92 (1998). В другом примере, см. патент США 6054297, Carter et al., от 25 апреля 2000 г., описывается метод получения гуманизированных антител путем замещения аминокислотной последовательности CDR соответствующей аминокислотной последовательностью CDR антитела человека и/или замещения аминокислотной последовательности FR соответствующей аминокислотной последовательностью FR участка антитела человека. Еще один пример, описанный в патенте США 5766886 Studnicka et al., от 16 июня 1998 г. ("Модифицированные вариабельные домены антител"), включает методы идентификации аминокислотных остатков вариабельных доменов, которые могут быть модифицированы без уменьшения нативной аффинности антиген-связывающего домена, но со снижением его иммуногенности по отношению к гетерологичным видам, а также методы получения таких модифицированных вариабельных доменов антител,которые могут быть полезными для применения у гетерологичных видов. См. также патент США 5869619 Studnicka от 9 февраля 1999 г. Как уже обсуждалось, модификация антител любым из способов, известных специалистам, обычно производится для достижения увеличенной связывающей аффинности к антигену и/или снижения иммуногенности антитела в организме реципиента. Согласно одному подходу гуманизированные антитела модифицируются таким образом, что у них удаляются сайты гликозилирования с целью увеличения их аффиности к специфическому антигену [Co et al., Mol Immunol 30:1361-1367 (1993)]. Технологии такие,как "решейпинг" ("reshaping"), "гиперхимеризация" ("hyperchimerization"), "вениринг/ресурфэйсинг"("veneering/resurfacing"), позволяют получать антитела со значительным терапевтическим потенциалом.[Vaswami et al., Annals of Allergy, Asthma,Immunol 81:105 (1998); Roguska et al., Prot Engineer 9:895904 (1996)]. См. также патент США 6072035 Hardman et al., от 6 июня 2000 г., который описывает методы восстановления исходной структуры антител ("reshaping"). В то время как эти технологии снижают иммуногенность антител путем сокращения числа инородных остатков, они не препятствуют антиидиотипическому и антиаллотипическому иммунному ответу, появляющемуся после повторных введений антител. Альтернативные методики для снижения иммуногенности антител описаны в источнике:Gilliland et al., J Immunol 62(6):3663-71 (1999). Во многих случаях гуманизированные антитела имеют сниженную способность связывать антиген. В таком случае является предпочтительным "ремутировать" гуманизированное антитело с тем, чтобы включить в его структуру один или более аминокислотный остаток, обнаруживаемый в оригинальном антителе (чаще всего в антителе грызуна), для того, чтобы попытаться восстановить связывающую аффинность антитела. См., например, Saldanha et al., Mol Immunol 36:709-19 (1999). Непептидные аналоги специфических связывающих агентов/белковые миметики Изобретение также относится к небелковым аналогам специфических связывающих агентов, обеспечивающих стабильную структуру и минимальную биодеградацию. Пептидные аналоги/миметики специфических связывающих агентов могут быть получены на основе выбранного ингибиторного пептида путем замещения одного или более аминокислотного остатка непептидными молекулами. Желательно,чтобы непептидные молекулы способствовали сохранению естественной конформации пептида или ста- 22010726 билизировали предпочтительную, т.е. биоактивную, конформацию, обеспечивающую способность распознавать и связывать Ang-2. С одной стороны, полученные аналоги/миметики демонстрируют увеличенную связывающую аффинность к Ang-2. Один пример метода, используемого для получения непептидных миметиков/аналогов из белковых специфических связывающих агентов, описан в источнике:Nachman et al., Regul Pept 57:359-370 (1995). При необходимости, пептиды специфических связывающих агентов, заявленные в соответствии с настоящим изобретением, могут быть модифицированы, например,путем гликозилирования, аминирования, карбоксилирования или фосфорилирования, а также путем создания аддитивных солей кислоты, дополнительных кислых солей, амидов, эфиров, в особенности Сконцевых эфиров и N-ацилированных производных пептидов, которые тоже входят в объем настоящего изобретения. Пептиды специфических связывающих агентов также могут быть модифицированы для создания белковых производных путем формирования ковалентных и нековалентных комплексов с другими молекулами. Ковалентно-связанные комплексы могут быть получены путем соединения химических молекул с функциональными группами боковых цепей аминокислот, входящих в состав пептидов специфических связывающих агентов, или путем присоединения этих химических молекул к N- или Сконцевым участкам. В особенности, предполагается, что пептиды специфических связывающих агентов могут быть соединены с маркерной группой, включающей, но не ограничивающейся указанными, радиоактивную метку, флуоресцентную метку, фермент (например, катализирующий колориметрическую или флуоресцентную реакцию), подложку, твердый матрикс или носитель (например, биотин или авидин). Изобретение,соответственно, обеспечивает получение соединения, содержащего молекулы антител, причем это соединение предпочтительно содержит маркерную группу, выбранную из группы, включающей радиоактивную метку, флуоресцентную метку, фермент, подложку, твердый матрикс или носитель. Такие метки хорошо известны специалистам в данной области, например, чаще всего применяются биотиновые метки, описанные, например, в патенте США 3817837; в патенте США 3850752; патенте США 3996345 и патенте США 4277437. Другие метки, которые могут оказаться полезными, включают,но не ограничиваются указанными, радиоактивные метки, флуоресцентные метки и хемилюминесцентные метки. Патенты США, касающиеся использования таких меток, включают, например, патент США 3817837; патент США 3850752; патент США 3939350 и патент США 3996345. Каждый из пептидов, заявленных в соответствии с настоящим изобретением, может содержать одну, две или более указанных меток. Методы получения специфических связывающих агентов Специфические связывающие агенты настоящего изобретения, являющиеся по своей структуре белками, могут быть получены путем химического синтеза в растворе или на твердом субстрате в соответствии с существующими технологиями. Существующий лимит для твердофазного синтеза составляет 85-100 аминокислот в длину. Однако технологии химического синтеза могут быть применены для химического связывания серии небольших пептидов для получения полноразмерного полипептида. Различные автоматические синтезаторы являются коммерчески доступными и могут быть использованы в соответствии с известным протоколом. См., например, Stewart и Young et al., Solid Phase Peptide Synthesis, 2d.ed., Pierce Chemical Co., (1984); Tarn et al., J Am Chem Soc, 105:6442, (1983); Merrifield, Science, 232:341347, (1986) и Barany and Merrifield, The Peptides, Gross and Meienhofer, eds, Academic Press, New York, 1284; Barany et al., Int. J. Peptide Protein Res., 30, 705-739 (1987) и патент США 5424398); каждая из этих работ приведена здесь в качестве ссылки. Методы твердофазного синтеза подразумевают использование сополимера (стиролдивинилбензола), содержащего 0,1-1,0 мМ аминов на один грамм полимера. Такие методы пептидного синтеза используют бутилоксикарбонил (t-BOC) или 9-фторенилметилоксикарбонил (FMOC) для защиты альфа-аминогрупп. Оба метода подразумевают ступенчатый синтез, при котором на каждом шагу добавляется единичная аминокислота, начиная с С-концевого участка пептида (см. Coligan et al., Current Protocols in Immunology, Wiley Intersciense, 1991, Unit 9). После завершения химического синтеза, с синтезированного пептида снимается защита для удаления блокирующих аминокислоты групп t-BOC илиFMOC, после чего пептид выделяют из полимера путем обработки кислотой при пониженной температуре (например, жидкой HF-10% анизолом в течение 25 мин-1 ч при температуре 0 С). После упаривания реагентов, пептиды специфического связывающего агента экстрагируют из полимера 1%-ным раствором уксусной кислоты и затем лиофизилируют с получением неочищенного материала. Такой материал может быть очищен с помощью такой технологии, как гель-фильтрация на Сефадексе G-15 с использованием 15%-ной уксусной кислоты в качестве растворителя. Лиофилизация подходящих фракций, полученных с колонки, приведет к получению гомогенных пептидов специфического связывающего агента или пептидных производных, которые могут быть определены с помощью таких стандартных технологий как аминокислотный анализ, тонкослойная хроматография, жидкостная хроматография высокого разрешения, ультрафиолетовая абсорбционная спектроскопия, молярная ротация, и подсчитаны с помощью твердофазовой деградации по Эдману. Химический синтез антител к Ang-2, а также их производных, вариантов и фрагментов, также, как и других специфических связывающих агентов Ang-2 белковой природы, допускает включение в структу- 23010726 ру агента аминокислот, не являющихся природными. Технологии рекомбинантных ДНК являются подходящими для получения полноразмерных антител или других крупных белковых специфических связывающих агентов и их фрагментов, заявленных в соответствии с настоящим изобретением. Молекула кДНК, кодирующая антитело или его фрагмент, может быть встроена в экспрессирующий вектор, который, в свою очередь, будет помещен в клетку-хозяин для продукции антител или их фрагментов. Понятно, что молекулы кДНК, кодирующие такие антитела, могут быть модифицированы по сравнению с исходными молекулами кДНК (которые были транслированы с мРНК), чтобы обеспечить вырожденность кодонов или включить в их состав предпочтительные кодоны, использующиеся в определенных типах клеток-хозяев. В целом, молекулы ДНК, кодирующие антитела, могут быть получены с помощью методик, описанных здесь в примерах. Когда необходимо получить Fab молекулы или CDRs, родственные оригинальной молекуле антитела, можно скринировать подходящую библиотеку (фагово-дисплейную библиотеку; лимфоцитарную библиотеку и т.д.), используя стандартные технологии для идентифицирования и клонирования родственных Fabs/CDRs. ДНК-зонды, используемые для такого скрининга, могут быть полноразмерными или усеченными нуклеотидными последовательностями, кодирующими Fab-фрагменты исходного антитела, один или более CDRs Fab-фрагмента исходного антитела, или другие подходящие зонды. Когда ДНК фрагменты используются в качестве зондов, стандартными условиями гибридизации являются условия, описанные у Ausubel et al. (Current Protocols in Molecular Biology, Current ProtocolsPress [1994]). После гибридизации блоты отмывают в условиях подходящей строгости, в зависимости от таких факторов, как размер зонда, ожидаемая гомология зонда и клона, тип библиотеки, подлежащей скринингу, и количество клонов, подлежащих скринингу. Примерами условий с высокой строгостью использующихся для скрининга являются 0,1 X SSC и 0,1% SDS при температуре 50-65 С. Различные системы "вектор экспрессии/хозяин" могут быть использованы для экспрессии полинуклеотидных молекул, кодирующих специфические связывающие агенты, заявленные в соответствии с настоящим изобретением. Такие системы включают, но не ограничиваются указанными, микроорганизмы, такие как бактерии, трансформированные рекомбинантным бактериофагом; плазмидные или космидные экспрессионные ДНК векторы; дрожжи, трансформированные дрожжевыми экспрессионными векторами; системы клеток насекомых, трансформированные вирусными экспрессионными векторами(например, бакуловирусными векторами); системы растительных клеток, трансформированные вирусными экспрессионными векторами (например, векторами на основе вируса мозаики цветной капусты,CaMV; вируса табачной мозаики, TMV) или трансформированные бактериальными экспрессионными векторами (например, Ti или pBR322 плазмидами); или системы клеток животных. Клетки млекопитающих, которые являются полезными в рекомбинантной продукции специфических связывающих агентов, включают, но не ограничиваются указанными, VERO-клетки, HeLa-клетки,клеточные линии яичника китайского хомячка (СНО), COS клетки (такие как COS-7), W138, BHK,HepG2, 3 Т 3, RIN, MDCK, A549, РС 12, K562 и клетки 293, так же, как и гибридомные клеточные линии,как описано здесь. Клетки млекопитающих являются подходящими для получения таких специфических связывающих агентов и фрагментов антител, которые обычно являются гликозилированными и требуют правильного восстановления структуры для своей активности. Предпочтительными клетками млекопитающих являются CHO клетки, гибридомные клетки и миелоидные клетки. Некоторые примерные протоколы для рекомбинантной экспрессии специфических связывающих агентов описаны здесь ниже. Термин "экспрессионный вектор" относится к плазмиде, фагу, вирусу или вектору, предназначенному для экспрессии полипептида, кодируемого нуклеотидной последовательностью ДНК (РНК) в составе такого вектора. Экспрессионный вектор может содержать транскрипционную единицу, включающую: (1) генетический элемент или элементы, играющие регуляторную роль в генной экспрессии, например, промоторы и энхансеры, (2) структуры или последовательности, кодирующие связывающий агент, транскрибируемые с образованием мРНК и транслируемые в белки, и (3) подходящие последовательности, инициирующие и терминирующие транскрипцию. Структурные единицы, предназначенные для использования в дрожжевых или эукариотических экспрессионных системах, предпочтительно включают лидирующую последовательность, делающую возможной внеклеточную секрецию транслируемого белка клеткой-хозяином. Альтернативно, когда рекомбинантный белок специфического связывающего агента экспрессируется без лидирующей или транспортной последовательности, он может включать аминоконцевой остаток метионина. Этот остаток может быть впоследствии выщеплен или не выщеплен из экспрессированного рекомбинантного белка для получения конечного продукта специфического связывающего агента. Например, специфические связывающие агенты могут быть рекомбинантно экспрессированы в дрожжах с использованием коммерчески доступной экспрессионной системы, например, с помощью дрожжевой экспрессионной системы [Pichia Expression System (Invitrogen, San Diego, CA)], следуя инструкции производителя. Эта система также задействует для направления секреции пре-про-альфа последовательность, но транскрипция вставки контролируется промотором алкогольоксидазы (АОХ 1), который индуцируется метанолом.- 24010726 Полученный пептид специфического связывающего агента очищается от дрожжевой ростовой среды с помощью, например, методов, используемых для очистки белка от супернатантов бактериальных клеток и клеток млекопитающих. Альтернативно, кДНК, кодирующая белок специфического связывающего агента, может быть клонирована в бакуловирусном экспрессионном векторе pVL1393 (PharMingen, San Diego, CA). Такой вектор может быть использован, в соответствии с инструкцией производителя (PharMingen) для заражения клеток насекомого Spodoptera frugiperda в sF9 небелковой среде и для получения рекомбинантного белка. Протеин специфического связывающего агента может быть очищен и выделен из среды с помощью гепариново-сефарозной колонки (Pharmacia). Альтернативно, пептид может быть экспрессирован в системе клеток насекомых. Такие системы для экспрессии белков хорошо известны специалистам в данной области. В одной такой системе в качестве вектора для экспрессии чужеродных генов в клетках Spodoptera frugiperda или в клетках Trichoplusialarvae может использоваться вирус ядерного полиэдроза Autographa californica (AcNPV). Последовательность, кодирующая специфический связывающий агент, может быть клонирована в несущественном участке вируса, таком как ген полиэдрина, и помещена под контроль полиэдринового промотора. Успешное встраивание гена, кодирующего пептид специфического связывающего агента, приведет к тому,что ген полиэдрина будет инактивирован, и полученный рекомбинантный вирус утратит способность синтезировать белок оболочки. Рекомбинантные вирусы могут быть использованы для заражения клеток S. frugiperda или клеток Т.larvae, в которых экспрессируется белок [Smith et al., J Virol 46:584 (1983); Engelhard et al., Proc Nat AcadSci (USA) 91:3224-7 (1994)]. В другом примере последовательность ДНК, кодирующая пептид специфического связывающего агента, может быть амплифицирована с помощью ПЦР и клонирована в подходящем векторе, например,pGEX-3X (Pharmacia). PGEX вектор создан для получения белков слияния, содержащих глутатион-Sтрансферазу (GST), кодируемую вектором, и белков специфических связывающих агентов, кодируемых фрагментом ДНК, встроенным в клонирующий сайт вектора. Праймеры для ПЦР могут быть получены таким образом, чтобы обеспечить включение подходящих сайтов расщепления. Когда компонент, используемый для слияния со специфическим связывающим агентом, применяется исключительно для обеспечения экспрессии или нежелательно его прикрепление к нужному пептиду, рекомбинантный белок слияния специфического связывающего агента может быть вырезан из GST участка белка слияния.PGEX-3X конструктом пептида специфического связывающего агента трансформируют XL-1 Blue клетки (Stratagene, La Jolla CA) E. coli и отдельные трансформанты изолируют и выращивают. Плазмидные ДНК индивидуальных трансформантов могут быть очищены и частично секвенированы, с использованием автоматического секвенсора для того, чтобы обнаружить включение вставки, кодирующей специфический связывающий агент, в нужной ориентации. Экспрессия полинуклеотидов, кодирующих антитела к Ang-2 и их фрагменты, с использованием рекомбинантных систем, описанных выше, может привести к получению антител и их фрагментов, нуждающихся в восстановлении структуры (правильном восстановлении различных дисульфидных мостиков) для того, чтобы они были биологически активными. Типичные методы восстановления структуры антител описаны здесь в примерах и в последующем разделе. Специфические связывающие агенты, полученные в бактериальных клетках, продуцируются в составе бактериальных нерастворимых телец включения и могут быть очищены, как описано ниже. Клетки-хозяева центрифугируют, отмывают в 0,15 М NaCl, 10 мМ Tris, pH 8; 1 мМ ЭДТА; и обрабатывают 0,1 мг/мл лизоцима (Sigma, St. Louis, МО) в течение 15 мин при комнатной температуре. Лизат осветляют обработкой ультразвуком и клеточный дебрис осаждают центрифугированием в течение 10 мин при 12000 X g. Осадок, содержащий специфический связывающий агент, может быть ресуспендирован в 50 мМ Tris, pH 8, и 10 мМ ЭДТА, нанесен на раствор 50%-го глицерина и центрифугирован в течение 30 мин при 6000 X g. Осадок может быть также ресуспендирован в стандартном фосфатном буферном растворе (PBS), свободном от ионов Mg+2 и Ca+2. Специфический связывающий агент может быть также выделен путем фракционирования ресуспендированного осадка в денатурирующем полиакриламидном геле с додецилсульфатом натрия (SDS) (Sambrook et al., supra). Гель вымачивают в 0,4 М KCl для визуализации белка, который вырезают из геля и элюируют под действием электрического тока в буфере для гель-электрофореза, не содержащем SDS. Если GST белки слияния продуцируются в бактериях как растворимые белки, их центрифугируют с помощью системы очистки GST (GST Purification Module, Pharmacia). Системы клеток млекопитающих для экспрессии рекомбинантных белков хорошо известны специалистам в данной области. Штаммы клеток-хозяев могут быть выбраны на основе их способности процессировать экспрессируемый белок или обеспечивать посттрансляционные модификации белка, которые необходимы для проявления белком нужной активности. Такие модификации полипептида включают, но не ограничиваются указанными, ацетилирование, карбоксилирование, гликозилирование, фосфорилирование, ацилирование и связывание с липидами. Различные клетки-хозяева, такие как CHO, HeLa, MDCK,293, W138, гибридомные клеточные линии и подобные им клетки, имеют специфические клеточные сис- 25010726 темы и характерные механизмы для обеспечения такой посттрансляционной модификации белка и могут быть использованы для обеспечения модификации и процессинга чужеродного белка, получаемого с помощью таких клеток. Существует целый ряд систем селекции для выделения клеток, трансформированных с целью рекомбинантного получения белка. Такие системы селекции включают, но не ограничиваются указанными,гены HSV тимидинкиназы, гипоксантин-гуанин фосфорибозилтрансферазы и аденин фосфорибозилтрансферазы в tk-, hgpt- или aprt-клетках, соответственно. Также, антиметаболическая устойчивость может быть использована как основа селекции по DHFR, обеспечивающей резистентность к метотрексату;gpt, обеспечивающей резистентность к мукофеноловой кислоте; neo, обеспечивающей резистентность к аминогликозиду G418 и хлорсульфону; и hydro, обеспечивающей резистентность к гигромицину. Дополнительные гены, кодирующие продукты, по которым может быть проведена селекция, включают генtrpB, продукт которого позволяет клеткам использовать индол вместо триптофана; и ген hisD, продукт которого позволяет клеткам использовать гистинол вместо гистидина. Маркеры, обеспечивающие визуальную индикацию для идентификации трансформантов, включают антоциан, -глюкуронидазу и ее субстрат, GUS, и люциферазу и ее субстрат, люциферин. Очистка и восстановление структуры (рефолдинг) специфических связывающих агентов В определенных случаях специфические связывающие агенты, полученные с помощью методов,описанных выше, нуждаются в "рефолдинге" и окислении для придания им правильной третичной структуры и в создании дисульфидных мостиков для того, чтобы быть биологически активными. Рефолдинг может быть достигнут с помощью набора методов, хорошо известных специалистам в данной области. Такие методы включают, например, экспозицию растворимого полипептидного агента при pH обычно выше 7 в присутствии хаотропного агента. Выбор хоатропного агента аналогичен выбору растворителя телец включения, однако хаотропный агент применяется в меньшей концентрации. Примером хаотропного агента является гуанидин. В большинстве случаев раствор для рефолдинга/окисления также содержит восстанавливающий агент и его окисленную форму в специфической пропорции для создания определенного восстановительно-окислительного потенциала, чтобы перемешанные дисульфиды образовали цистеиновые мостики. Некоторые обычно используемые восстановительно-окислительные пары включают цистеин/цистамин, глутатион/дитио-бис-GSH, хлорид меди, дитиотреитол DTT/дитиан DTT, и 2-меркаптоэтанол (bME)/ дитио- bME. Во многих случаях для повышения эффективности рефолдинга может быть использован сорастворитель. Как правило, в качестве сорастворителей используют глицерол,полиэтиленгликоль различного молекулярного веса и аргинин. Было бы желательно произвести очистку белковых специфических связывающих агентов и их вариантов, заявленных в соответствии с настоящим изобретением. Методы очистки белков хорошо известны специалистам в данной области. Такие методы, например, включают грубое фракционирование полипептидных и неполипептидных фракций. После отделения полипептида специфического связывающего агента от других белков, нужный полипептид может быть дополнительно очищен с помощью хроматографии и электрофореза для достижения частичной или полной очистки (или очистки до гомогенизации). Аналитическими методами, применяемыми для получения очищенного пептида специфического связывающего агента, являются ионообменная хроматография, эксклюзионная хроматография, электрофорез в полиакриламидном геле, изоэлектрическое фокусирование. Наиболее эффективным методом очистки пептидов является жидкостная хроматография быстрого разделения белков или жидкостная хроматография высокого давления (HPLC). Определенные аспекты настоящего изобретения касаются очистки, и в определенных случаях, существенной очистки белка или пептида специфического связывающего агента. Термин "очищенный белок или пептид специфического связывающего агента", используемый здесь, означает композицию, изолированную от других компонентов, в то время как белок или пептид специфического связывающего агента очищен до уровня, который соотносится с той степенью чистоты, в которой он существует в природе. Термин "очищенный белок или пептид специфического связывающего агента" также относится к белку и пептиду специфического связывающего агента, свободному от окружения, в котором он существует в природе. В целом, термин "очищенный" относится к специфическому связывающему агенту, который подвергся фракционированию с удалением примесных компонентов, но который в значительной степени сохраняет свою биологическую активность. Термин "по существу очищенный" относится к белку или пептиду специфического связывающего агента, который составляет большую часть препарата специфического связывающего агента, а именно около 50, около 60, около 70, около 80, около 90, около 95%. Методы для количественной оценки уровня очистки специфического связывающего агента будут понятны специалистам в данной области из настоящего изобретения. Эти методы включают, например,определение специфической связывающей активности активной фракции или определение количества специфического связывающего агента во фракции, полученной в результате электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS/PAGE). Предпочтительным методом для оценки чистоты фракции специфического связывающего агента является вычисление связывающей активности фракции по сравнению со связывающей активностью первоначального экстракта, и на основе- 26010726 этого определяют степень очистки, которую выражают понятием "кратность очистки". Выбор современных приборов, использующихся для определения связывающей активности, безусловно, зависит от методики исследования, применяемой для определения активности после очистки, и от того, обнаруживает ли экспрессируемый белок или пептид специфического связывающего агента выявляемую активность. Различные технологии, являющиеся подходящими для очистки белка специфического связывающего агента, хорошо известны специалистам в данной области знаний. Они включают, например, преципитацию с сульфатом аммония, полиэтиленгликолем, антителами (иммунопреципитацию) и другими подобными соединениями или тепловую денатурацию с последующим центрифугированием; хроматографию, такую как аффинная хроматография (например, на Протеин-А-Сефарозе), ионообменная хроматография, гель-фильтрационная хроматография, обратнофазовая хроматография, гидроксиапатитная и аффинная хроматография; изоэлектрическое фокусирование, электрофорез в геле и комбинации любых из этих методов. Из области техники известно, что порядок проведения различных стадий очистки может быть изменен, или можно пропускать определенные стадии, и при этом метод все равно останется подходящим для получения очищенного специфического связывающего агента. Не существует такого общего требования, что специфический связывающий агент всегда должен быть получен в максимально возможной очищенной форме. В действительности, показано, что не по существу очищенные специфические связывающие агенты могут использоваться в определенных случаях. Частичная очистка может быть достигнута путем использования нескольких стадий очистки в комбинации или путем применения различных вариантов той же общей схемы очистки. Например, известно,что катионообменная колоночная хроматография, осуществленная с помощью HPLC системы, приводит к значительному повышению "кратности" очистки по сравнению с хроматографией, проведенной в системе низкого давления. Методы, приводящие к низкой степени очистки, могут оказаться полезными для восстановления белкового продукта специфического связывающего агента или поддержания связывающей активности экспрессируемого белка специфического связывающего агента. Известно, что перемещение полипептида в геле может варьировать иногда значительным образом при различных условиях осуществления SDS/PAGE [Capaldi et al., Biochem BiophysRes Comm, 76:425(1977)]. Таким образом, можно предположить, что при различных условиях электрофореза явная молекулярная масса выявленных продуктов очищенных или частично очищенных специфических связывающих агентов будет различна. Исследования связывания Иммунологические исследования связывания обычно предполагают использование захватывающего агента для специфического связывания и часто иммобилизации анализируемого антигена-мишени. Захватывающий агент - это соединение, которое специфически связывается с анализируемым антигеном. В одном варианте настоящего изобретения захватывающим агентом является антитело или его фрагмент,специфически связывающий Ang-2. Такие иммунологические исследования связывания хорошо известны специалистам [см. Asai, ed., Methods in Cell Biology, Vol. 37, Antibodies in Cell Biology, AcademicPress, Inc., New York (1993)]. Иммунологические исследования связывания часто подразумевают использование маркерного агента, который просигнализирует о наличии комплекса связывания, сформированного захватывающим агентом и антигеном. Маркерным агентом может быть одна из молекул, содержащаяся в комплексе связывания; т.е. это может быть маркерный специфический связывающий агент или маркерный антиген к специфическому связывающему агенту. Альтернативно, маркерным агентом может быть третья молекула, обычно другое антитело, связывающееся с образовавшимся комплексом. Маркерным агентом может быть, например, антитело к специфическому связывающему агенту, несущее метку. Второе антитело,специфичное к комплексу связывания, может не иметь метки, но может быть связано четвертой молекулой, специфичной к тому типу антител, к которому принадлежит второе антитело. Например, второе антитело может быть модифицировано с помощью выявляемой метки, такой как биотин, которая затем может быть связана с четвертой молекулой, например меченным ферментом стрептавидином. Другие белки, способные специфично связываться с константными участками иммуноглобулина, такие, как белок А или белок G, могут также использоваться в качестве маркерных агентов. Эти связывающие белки являются нормальными структурными белками клеточной стенки бактерии стрептококка и демонстрируют сильную неиммуногенную реактогенность по отношению к константным участкам иммуноглобулинов различных видов животных [см., в основном, Akerstrom, J Immunol, 135:2589-2542 (1985); andChaubert, Mod Pathol, 10:585-591 (1997)]. Во время исследования каждая комбинация реагентов может потребовать стадии инкубации и/или отмывания. Стадия инкубации может длиться по времени от нескольких секунд до нескольких часов,предпочтительно от около 5 мин до около 24 ч. Однако время инкубации зависит от модификации способа, анализируемого антигена, объема раствора, концентрации и т.п. Обычно исследования проводят при температуре окружающей среды, хотя они могут проводиться и при различных температурах.A. Неконкурентные исследования связывания. Иммунологические исследования связывания могут быть неконкурентного типа. Эти исследования подразумевают прямое измерение количества захваченного антигена. Например, согласно одной пред- 27010726 почтительной модификации "сэндвич-метода", захватывающий агент (антитело) может быть непосредственно связан с твердым субстратом, на котором он иммобилизуется. Такие иммобилизованные антитела затем связываются с антигеном (захватывают антиген) в тестируемом образце. Далее захваченный антиген связывают с меченным агентом, например, вторым антителом, соединенным с меткой. Согласно другой предпочтительной модификации "сэндвич-метода" второе антитело не имеет метки, но может быть связано со специфическим к нему меченным антителом того же видового происхождения. Второе антитело также может быть модифицировано с помощью выявляемого соединения, например, биотина, с которым способен связываться третий меченный агент, такой как стрептавидин. [См. Harlow and Lane, Antibodies, A Laboratory Manual., Ch 14, Cold Spring Harbor Laboratory, NY (1988); работа приведена здесь в качестве ссылки].B. Конкурентные исследования связывания. Иммунологические исследования связывания могут быть конкурентного типа. Количество захваченного агента, присутствующее в образце, определяют непрямым образом путем измерения количества добавленного агента, конкурирующего с захватываемым соединением, присутствующим в образце, за связывание с захватывающим агентом. В одном предпочтительном варианте конкурентного исследования связывания, известное количество захватываемого агента, обычно меченного, добавляется к образцу и затем этот образец связывают с антителом (захватывающим агентом). Количество меченного соединения, связавшегося с антителом, обратно пропорционально концентрации тестируемого вещества в образце (См. Harlow and Lane, Antibodies, A Laboratory Manual., Ch 14, pp. 579-583 supra). В другом предпочтительном конкурентном исследовании связывания антитело иммобилизировано на твердой подложке. Количество белка, связывающегося с антителом, может быть определено как путем измерения количества белка, присутствующего в комплексе белок/антитело, так и альтернативно этому способу, путем измерения количества не связавшегося в комплекс белка. Количество белка может быть определено с помощью метки. (См. Harlow and Lane, Antibodies, A Laboratory Manual., Ch 14, supra). В другом предпочтительном конкурентном исследовании связывания используется ингибирование гаптеном. Так, известное количество захватываемого агента иммобилизируется на твердой подложке. Известное количество антитела добавляется к образцу, и этот образец соединяют с иммобилизируемым захватываемым агентом. Количество антитела, связавшееся с иммобилизуемым захватываемым агентом,обратно пропорционально количеству этого захватываемого агента, присутствующему в образце. Количество иммобилизованного антитела может быть определено как путем оценки непосредственно иммобилизованной фракции антитела, так и путем оценки фракции, оставшейся в растворе. Определение может быть прямым, когда само антитело связано с меткой, или непрямым, когда добавляют меченное соединение, которое специфически связывается с антителом, как описано выше. С. Применение конкурентных исследований связывания. Конкурентные исследования связывания могут применяться для определения перекрестной реактивности, чтобы специалист смог установить, входит ли в состав белкового или ферментного комплекса,который был выявлен с помощью заявленного специфического связывающего агента, нужный белок, а не вещество, обладающее с ним перекрестной реактивностью, и что используемое антитело действительно специфично по отношению к этому белку и не связывает других антигенов. В исследованиях такого рода антиген может быть иммобилизован на твердой подложке и к образцу добавляют смесь неизвестных белков, которые будут конкурировать с иммобилизованным белком за связывание со специфическими связывающими агентами. Способность белков конкурировать с иммобилизованным антигеном за связывание с антителами специфических связывающих агентов сравнивают со связыванием этих же белков, которые были иммобилизованы на твердой подложке, с данными антителами, для определения перекрестной реактивности смеси белков.D. Другие исследования связывания. Настоящее изобретение также относится к методам Вестерн-блоттинга для определения количества присутствующего в образце Ang-2. Технология обычно заключается в разделении белков образца с помощью электрофореза в геле на основе их молекулярного веса и в последующем переносе их на подходящую твердую подложку, такую как нитроцеллюлозный фильтр, нейлоновый фильтр или производные нейлонового фильтра. Образец инкубируют с антителами или их фрагментами, специфически связывающими Ang-2, и выявляют полученный комплекс. Указанные антитела могут быть непосредственным образом соединены с меткой или, альтернативно, могут быть впоследствии выявлены с помощью меченных вторых антител, связывающихся с первыми. Исследования связывания для определения Ang-2 специфических связывающих агентов, препятствующих взаимодействию Ang-2 с его рецептором, описаны ниже в примерах. Диагностические исследования Антитела и их фрагменты, заявленные в соответствии с настоящим изобретением, могут быть полезными для диагностики заболеваний и состояний, характеризующихся экспрессией Ang-2 или его субъединиц, или для исследований, применяющихся для контролирования состояния пациентов, принимающих индукторы Ang-2, его фрагменты, агонисты или ингибиторы активности Ang-2. Диагностические исследования Ang-2 включают методы, использующие специфический связывающий агент и метку- 28010726 для определения Ang-2 в жидких средах тела человека или в клеточных или тканевых материалах. Специфические связывающие агенты, заявленные в соответствии с настоящим изобретением, могут использоваться как с модификацией, так и без нее. В предпочтительном диагностическом исследовании специфические связывающие агенты метят, например, путем присоединения метки или репортерной молекулы. Большое разнообразие меток и репортерных молекул хорошо известно, некоторые из них были уже описаны здесь. В особенности, настоящее изобретение наиболее полезно для диагностики заболеваний человека. Разнообразные методы для выявления белков Ang-2, использующие как моноклональные, так и поликлональные специфичные к ним антитела, хорошо известны специалистам. Примеры включают твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA) и флуоресцентный метод разделения клеток (FACS). Двухсайтовое, иммунологическое исследование с помощью моноклональных антител, реагирующих с двумя неперекрывающимися эпитопами Ang-2, является предпочтительным, но может быть применено и конкурентное исследование связывания. Такие исследования описаны, например, Maddox et al., J Exp Med. 158:1211 [1983]. Чтобы создать основу для диагностики, обычно устанавливают нормальные или стандартные уровни экспрессии человеческого Ang-2. Это может быть осуществлено путем объединения жидкостей организма или клеточных экстрактов здорового субъекта, предпочтительно, человека со специфическим связывающим агентом, например, антителом к Ang-2, в условиях, подходящих для формирования комплекса, что хорошо известно специалистам в данной области. Количество образовавшихся стандартных комплексов может быть подсчитано путем сравнения связывания специфических связывающих агентов с известным количеством белка Ang-2 как в контрольном, так и в исследуемом образце. Затем стандартные уровни, полученные при исследовании нормального образца, сравнивают с уровнями, полученными при исследовании образца, взятого из организма потенциально больного субъекта. Отклонение между стандартным и выявленным в образце уровнями Ang-2 и определяет его роль в возникновении данного заболевания. Для диагностического использования, в определенных случаях, специфические связывающие агенты соединяют с выявляемой меткой, которой может быть любая молекула, способная генерировать, как непосредственно, так и опосредованно, регистрируемый сигнал. Например, меткой может быть радиоизотоп, такой как 3 Н, 14 С, 32 Р, 35S или 125I, флуоресцентное или хемилюминесцентное соединение, такое как флуоресцеина изотиоцианат, родамин, люциферин, или фермент, такой как щелочная фосфатаза, галактозидаза, или пероксидаза хрена [Bayer et al., Meth Enz, 184: 138-163, (1990)]. Заболевания Изобретение также относится к специфическому связывающему агенту, который связывается сAng-2 и является полезным для лечения заболеваний и патологических состояний человека. Агенты, модулирующие связывающую активность Ang-2, или другую клеточную активность, могут использоваться в комбинации с другими терапевтическими агентами для увеличения их терапевтических эффектов или снижения возможных побочных эффектов. В одном аспекте, настоящее изобретение относится к реагентам и методам, полезным для лечения заболеваний и состояний, характеризующихся нежелательной и отклоняющейся от нормального уровня активностью Ang-2 в клетках. Такие заболевания включают рак и другие гиперпролиферативные состояния, такие как гиперплазия, псориаз, контактный дерматит, иммунологические нарушения, а также бесплодие. Изобретение также относится к способам лечения рака у животных и человека, заключающимся во введении животным эффективного количества специфического связывающего агента, который ингибирует или снижает активность Ang-2. Изобретение, кроме того, относится к способам ингибирования роста раковых клеток, включая процессы клеточной пролиферации, инвазии и метастазирования в биологических системах. Способы подразумевают использование соединений, заявленных в соответствии с настоящим изобретением, в качестве ингибиторов клеточного роста раковых клеток. Предпочтительно эти способы применяют для ингибирования или снижения роста раковых клеток, инвазии, метастазирования или для снижения частоты опухолевых заболеваний у животных, преимущественно млекопитающих. Способы, заявленные в соответствии с настоящим изобретением, также могут применяться в исследовательских целях, например, для исследования роста раковых клеток и изучения их свойств, а также для исследования соединений, способных влиять на рост раковых клеток. Рак, для лечения которого возможно применение методов, заявленных в соответствии с настоящим изобретением, встречается преимущественно у млекопитающих. К таким млекопитающим относятся человек и другие приматы, домашние животные, такие как кошки и собаки, лабораторные животные,такие как крысы, мыши и кролики, и животные, выращиваемые на фермах, такие как лошади, свиньи,овцы и крупный рогатый скот. Опухоли или неоплазии представляют собой такую форму тканевого роста, при которой клеточное деление является неконтролируемым и прогрессирующим. В некоторых случаях такой рост может быть благоприятным, но в других он является заведомо злокачественным и может привести к гибели организма. Злокачественные опухоли или раковые опухоли отличаются от доброкачественных тем, что для них- 29010726 характерна агрессивная клеточная пролиферация, они могут прорастать в окружающие ткани и обладают способностью к метастазированию. Кроме того, для злокачественных опухолей характерна сходная структура и очень низкая степень клеточной дифференцировки (недифференцированные опухоли); кроме того, по внешнему виду они похожи на окружающие их ткани. Это свойство также называют "аплазией". Неоплазии, для лечения которых могут применяться способы, заявленные в соответствии с настоящим изобретением, также включают солидные опухоли, т.е. карциномы и саркомы. К карциномам относят злокачественные опухоли, происходящие из эпителиальных клеток, инфильтрирующие (инвазирующие) окружающие ткани и дающие метастазы. Аденокарциномы - это карциномы, происходящие из железистого эпителия или из определенной железистой структуры. Другая широкая категория включает саркомы, опухоли, происходящие из соединительной ткани или гомогенной субстанции, такой как эмбриональная соединительная ткань. Изобретение также относится к лечению опухолей, происходящих из миелоидной или лимфоидной ткани, таких как лейкозы, лимфомы и других опухолей, которые обычно не представляют собой опухолевую массу, но поражают сосудистую и лимфоцитарную системы. К типам рака, которые подлежат лечению с помощью способов, заявленных в соответствии с настоящим изобретением, относятся, например, АКТГ-продуцирующая опухоль, острый лимфоцитарный лейкоз, острый нелимфоцитарный лейкоз, рак коры надпочечников, рак мочевого пузыря, опухоли головного мозга, рак молочной железы, рак шейки матки, хронический лимфоцитарный лейкоз, хронический миелоцитарный лейкоз, рак прямой кишки, кожная Т-клеточная лимфома, рак эндометрия, рак пищевода, саркома Юинга, рак желчного пузыря, волосатоклеточный лейкоз, рак головы и шеи, лимфома Ходжкина, саркома Капоши, рак почки, рак печени, рак легкого (мелко- и крупноклеточный), злокачественный канцероматоз брюшины, злокачественный канцероматоз плевры, меланома, мезотелиома, множественная миелома, нейробластома, глиома, неходжкинская лимфома, остеосаркома, рак яичников, рак яичников (из зародышевых клеток), рак поджелудочной железы, рак полового члена, рак простаты, ретинобластома, рак кожи, мягкотканная саркома, плоскоклеточный рак, рак ротовой полости, рак яичек, рак щитовидной железы, трофобластические неоплазмы, рак матки, рак влагалища, рак вульвы и опухоль Вильмса. Изобретение в особенности относится к лечению различных типов экспериментально определенных раковых заболеваний. В этих иллюстративных экспериментах используют стандартные модели in viro иin vivo, известные из уровня техники. Такие эксперименты проводятся для того, чтобы определить агенты, которые могут оказаться полезными для использования в лечебном режиме in vivo. Однако очевидно,что применение способа, заявленного в соответствии с настоящим изобретением, не ограничивается только лечением этих типов опухолей, но и распространяется на любые солидные опухоли, происходящие из любого органа или системы. Способ, заявленный в соответствии с настоящим изобретением, наилучшим образом подходит для ингибирования роста или, по крайней мере, снижения его темпов, злокачественных опухолей, инвазия и метастазирование которых ассоциированы с экспрессией или активностью Ang-2. Изобретение также включает применение специфического связывающего агента, такого как антитело, заявленного в соответствии с настоящим изобретением, в комбинации с противоопухолевым химиотерапевтическим агентом, которым может быть любой подходящий химиотерапевтический агент. Комбинирование специфического связывающего агента с другими подобными соединениями может служить основой для разработки химиотерапевтической схемы. Различные химиотерапевтические схемы могут быть разработаны практикующим врачом в данной области медицины и могут входить в способ, заявленный в соответствии с настоящим изобретением. Могут использоваться любые химиотерапевтические агенты, включая алкилирующие агенты, антиметаболиты, гормоны и антагонисты, радиоизотопы, а также натуральные препараты. Например, соединение, заявленное в соответствии с настоящим изобретением, может вводиться совместно с антибиотиком, таким как доксирубицин и другими аналогами антрациклина, с азотсодержащими алкалоидами растения горчицы, таким как циклофосфамид, аналогами пиримидина, такими как 5-фторурацил, цисплатин, гидроксимочевина, таксол и их природными или синтетическими аналогами, и им подобными. В качестве другого примера, в частности, в случаях смешанных опухолей, таких как аденокарцинома молочной железы, в состав которой входят гонадотропинзависимые и гонадотропин-независимые клетки, соединение, заявленное в соответствии с настоящим изобретением, может применяться совместно с лейпрорелином или гозерелином (синтетические белковые аналоги гонадотропин-рилизинг гормона). Другие антинеопластические схемы подразумевают использование соединений тетрациклинового ряда вместе с иными подходами, например, хирургией, радиационным облучением и т.д., которые называются здесь "сопутствующими антинеопластическими подходами". Таким образом, заявленный в соответствии с изобретением способ может применяться совместно с подходящими методиками для снижения побочных эффектов и повышения эффективности лечения. Соответственно, настоящее изобретение относится к композициям и способам, полезным для лечения различных видов злокачественных опухолей, в том числе солидных опухолей и лейкозов. К указанным заболеваниям относятся (но не ограничиваются указанными): аденокарцинома молочной железы,- 30
МПК / Метки
МПК: C12P 21/08, C07K 16/46, C07K 16/00, C07H 21/04, A61K 39/385, A61K 39/395
Метки: агенты, связывающие, специфически, ангиопоэтин-2
Код ссылки
<a href="https://eas.patents.su/30-10726-agenty-specificheski-svyazyvayushhie-angiopoetin-2.html" rel="bookmark" title="База патентов Евразийского Союза">Агенты, специфически связывающие ангиопоэтин-2</a>
Агенты, специфически связывающиеся с человеческим ангиопоэтином-2

Номер патента: 8248
Опубликовано: 27.04.2007
Авторы: Мин Хосунг, Олинер Джонатан Дэниэл
МПК: C07K 14/00, C07K 1/00, A61K 38/00...
Метки: специфически, агенты, ангиопоэтином-2, человеческим, связывающиеся
Формула / Реферат:
1. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность WDPWT (SEQ ID NO: 65) длиной в 5-50 аминокислот, и его физиологически приемлемые соли. 2. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность WDPWTC (SEQ ID NO: 66) и его физиологически приемлемые соли. 3. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность Cz2WDPWT (SEQ ID NO: 67) где...
Связывающие агенты, ингибирующие миостатин

Номер патента: 9056
Опубликовано: 26.10.2007
Авторы: Мин Хосунг, Хан Хкью, Бун Томас Чарлз
МПК: C07K 14/475, C07K 14/495, C07K 14/00...
Метки: связывающие, миостатин, агенты, ингибирующие
Формула / Реферат:
1. Связывающий агент, включающий по меньшей мере один пептид, способный связывать миостатин, причем пептид включает аминокислотную последовательность WMCPP (SEQ ID NO: 633). 2. Связывающий агент по п.1, отличающийся тем, что пептид содержит от 5 до 50 аминокислот. 3. Связывающий агент, включающий по меньшей мере один пептид, способный связывать миостатин, причем пептид включает аминокислотную последовательность Ca1a2Wa3WMCPP (SEQ ID NO: 352),...
Ингибиторы ароматазы и моноклональные антитела против her2 как противоопухолевые агенты

Номер патента: 5931
Опубликовано: 25.08.2005
Авторы: Массимини Джорджо, Пишителли Габриэлла, Пурандаре Дайнеш
МПК: A61K 39/395, A61P 35/00
Метки: моноклональные, антитела, противоопухолевые, ингибиторы, ароматазы, агенты, против
Формула / Реферат:
1. Применение ингибитора ароматазы для производства фармацевтической композиции для лечения гормонзависимого нарушения, характеризуемого сверхэкспрессией HER2. 2. Применение по п.1, где нарушение представляет собой рак молочной железы, шейки матки, яичников и эндометрия или эндометриоз. 3. Применение по п.2, где нарушение представляет собой рак молочной железы. 4. Применение по п.1, где ингибитор ароматазы выбран из эксеместана,...
Гуманизированный иммуноглобулин, который специфически связывается с бета-амилоидным пептидом, способы его применения

Номер патента: 10469
Опубликовано: 29.08.2008
Авторы: Шенк Дэйл Б., Салданха Джоз, Бэси Гарик
МПК: A61K 39/395, C07K 16/18, A61P 25/28...
Метки: способы, beta-амилоидным, связывается, применения, специфически, иммуноглобулин, гуманизированный, пептидом
Формула / Реферат:
1. Гуманизированный иммуноглобулин, который специфически связывается с бета-амилоидным пептидом (Ab), или его антигенсвязывающий фрагмент, содержащий: i) легкую цепь, содержащую гипервариабельные участки (CDR) последовательности вариабельной области лёгкой цепи иммуноглобулина 3D6, представленной как SEQ ID NO:2, и каркасные участки вариабельной области лёгкой цепи человеческого акцепторного иммуноглобулина, и ii) тяжелую цепь, содержащую...
Антитело или антигенсвязывающий домен, специфически связывающиеся с полипептидом потенциалзависимого воротного ионного канала

Номер патента: 8252
Опубликовано: 27.04.2007
Авторы: Коэн Даниель, Абдеррахим Хади, Симон Анн-Мари, Чумаков Илья
МПК: A61P 25/28, C07K 16/18, A61K 39/395...
Метки: антигенсвязывающий, полипептидом, потенциалзависимого, связывающиеся, специфически, ионного, воротного, домен, канала, антитело
Формула / Реферат:
1. Антитело или антигенсвязывающий домен, специфически связывающийся с полипептидом с последовательностью SEQ ID NO: 5. 2. Антитело или антигенсвязывающий домен по п.1, который является химерным, гуманизированным антителом, или антигенсвязывающим доменом, или моноклональным или поликлональным антителом, или антигенсвязывающим доменом человека. 3. Антитело или антигенсвязывающий домен по п.1 или 2, которое является антителом или...
Предыдущий патент: Гербицидное средство
Следующий патент: Аминогетероарильные соединения в качестве ингибиторов протеинкиназ
Случайный патент: Предохранительная крышка