Липополисахарид бактерий e. coli штамма dsm 6601 и способ его получения
Номер патента: 5217
Опубликовано: 30.12.2004
Авторы: Зонненборн Ульрих, Малинка Юрген, Ульмер Артур, Зарингер Ульрих, Шульце Юрген, Ритшель Эрнст Теодор, Пропперт Ханс






Формула / Реферат
1. Липополисахарид бактерий E.coli штамма DSM 6601, содержащий 8 фосфатных остатков и 0,5 моль P-Etn в расчете на молекулу, имеющий структуру, представленную на фиг. 7.
2. Способ получения липополисахарида по п.1, заключающийся в том, что промытую и высушенную бактериальную массу E.coli штамма DSM 6601 подвергают экстракции смесью фенол/вода и полученный экстракт обрабатывают РНКазой, ДНКазой и протеиназой K.
3. Применение липополисахарида бактерий E.coli штамма DSM 6601 по п.1, в частности полученного в соответствии с п.2, для модулирования иммунной системы человека и животных, изменения биологической активности других микроорганизмов, получения иммуномодулирующих лекарственных средств для человека и животных, а также в качестве маркера или эталонного вещества для микробиологической или медицинской диагностики in vitro, в качестве иммуностимулятора для вакцинации человека и животных, в качестве основного материала для биотехнологического синтеза и изготовления иммуномодулирующих веществ.
Текст
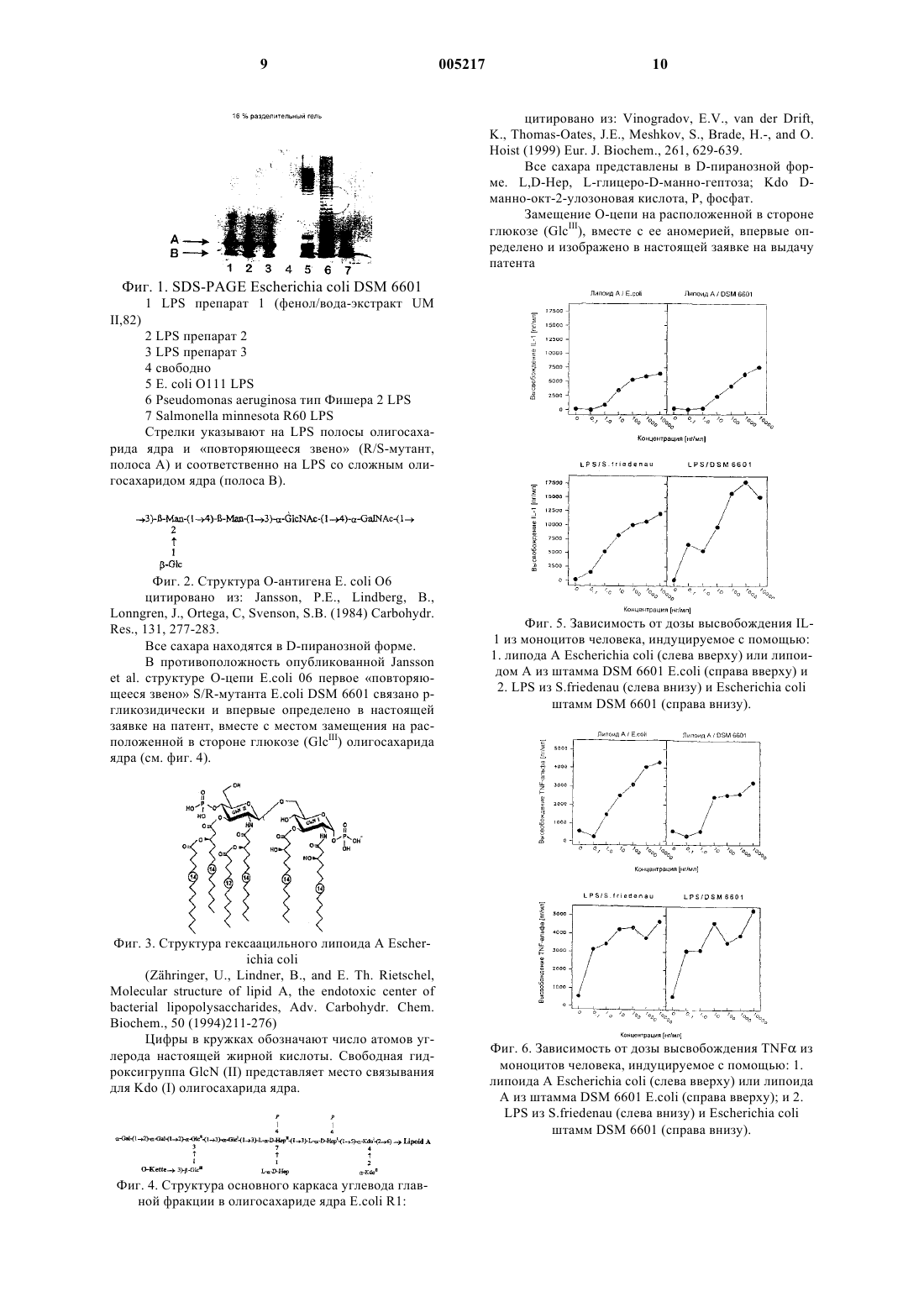
1 Изобретение касается новых липополисахаридов из E.coli. Под эндотоксинами понимают бактериальные структурные компоненты, которые, в отличие от экзотоксинов, не выделяются живыми бактериями, но главным образом освобождаются после аутолиза. В случае так называемых классических эндотоксинов речь идет о термостабильных липополисахаридах (далее LPS) из наружных клеточных мембран грамотрицательных бактерий. LPS состоят из так называемого липоида А, олигосахарида ядра, а также специфической О-цепи, причем липоид А отвечает за токсичное действие LPS. Эндотоксины стимулируют в макроорганизме продукцию медиаторов иммунной ситемы, например интерлейкина 1, (IL-1) и фактора некроза опухоли (TNF). В отношении состава эндотоксинов бактерий кишечной группы и, в частности, E.coli, уже проведен ряд исследований, причем было установлено, что S/R-мутанты, как правило, содержат только повторения, то есть повторяющееся звено" О-цепи (ср. фиг. 1). Исходят из того, что в этих случаях ген, который кодирует фермент полимеризации О-цепи, является дефектным и поэтому будет передавать только повторяющееся звено" на олигосахарид ядра. LPSструктуры подобного типа, но отличной структуры часто находятся также в патогенных для человека микроорганизмах, например Neisseria,Vibrio, Capmylobacter, Helicobacter и т.д. Эти микроорганизмы имеют LPS, которые дают им возможность лишать хозяина иммунной защиты путем особой молекулярной мимикрии и т.п.,благодаря наличию сиалиновых кислот и содержащих сиалиновые кислоты олигосахаридов,подобных гликопротеинам и гликолипидам млекопитающих. Для E.coli DSM 6601 был установлен О 6-Серотип. Эта структура исследована и опубликована P.E.Jansson et al.,Carbohydr. Res. 131 (1984) 277-283. Структура соответствует формуле, представленной на фиг. 2. Различные исследовательские группы изучали липоид А колибактерий. При этом было установлено, что структура липоида А, как правило, существует в гексаацильной форме и совпадает для всех серотипов из E.coli (фиг. 3). Структура гексаацильной связи была опубликована в 1984 г. Th. Rietschel et al., Structure andRietschel, ed.), Elsevier, Amsterdam (1984), pp. 187-220 и соответствует изображению на фиг. 3. Специфическая О-цепь и липоид А связаны друг с другом через олигосахарид ядра. До настоящего времени известны пять различных олигосахаридов ядра у E.coli. Здесь также производится ссылка на О.Holst et al., Chemicalstructure of the core region of lipopolysaccharide,в: Bacterial Endotoxic Lipopolysaccharides, Vol.1,Morrison D.S. and Ryan, J.L (eds.), Boca Raton,FL, USA (1992) pp. 135-170 (ср. фиг. 4). При исследованиях LPS из штамма DSM 6601 E.coli было обнаружено, что липоид А в структуре гексаацильной формы также соответствует липоиду А, описанному для Е. coli. Исследования в отношении высвобождения IL-1 и TNF в моноцитах человека подтверждают, что липоид А обладает такой же активностью и поэтому соответствует с высокой вероятностью известной структуре липоида AE.coli (ср. фиг. 5, 6). Это предположение подтверждалось химическими анализами. В структуре специфического О-антигенаDSM 6601 Е. coli поражает тот факт, что очевидно в цепи регулярно существует только единственное повторяющееся звено" (ср. фиг. 1). Отсюда можно заключить, что речь идет оS/R-мутанте у штамма DSM 6601, что является чрезвычайно необычным для человеческого изолята. Однако, как показывают серологические анализы, структура этого повторяющегося звена" соответствует основному образцу О-цепиE.coli О 6. Область ядра у штамма DSM 6601 соответствует известной R1-структуре. Структурные особенности состоят в том, что аналитически было установлено 8 фосфатных остатков на молекулу LPS, причем, как правило, на липоид А выпадают только 2 фосфатных остатка. В дальнейшем было обнаружено нестехиометрическое содержание пирофосфоэтаноламина. Таким образом, обобщение позволяет установить, что LPS штамма DSM 6601 значительно отличается от до сих пор известного LPS из E.coli, в частности, в фосфорилированной сахаридной части ядра и по степени полимеризации О-цепи. Липоид А структурно и биологически соответствует обычному для E.coli типу,что подчеркивает роль этого вещества в биологической активности. Описанный LPS предназначен не только для идентификации несущего его колиштамма, но и придает ему также пониженную патогенность при сохранении иммуномодулирующего действия. Тот факт, что О-цепь имеет -гликозидную связь вместо гликозидной, впервые стало возможным однозначно увидеть на примере S/R-мутанта DSM 6601 для E.coli. В случае липополисахарида (LPS) из Е.coli DSM 6601 речь идет о новой гладкошероховатой (S/R)-структуре, которая, с одной стороны, состоит из до сих пор известных частей структур (О-специфическая цепь, олигосахарид ядра и липоид А), с другой стороны,впервые и полностью структурно охарактеризована в представленной здесь комплексной форме (ср. фиг. 7). О-специфическая цепь, состоящая только из единственного повторения ("по 3 вторяющееся звено") серотипа О 6, имеет гликозидную связь с олигосахаридом ядра и,таким образом, иначе связана в пределах О-цепиR1-структуру, химическое состояние которой подтверждается серологическими исследованиями с R1-специфическими антителами. Компонент липоида А обладает характерной определенной для липоида А Е. coli химической конституцией.LPS из штамма DSM 6601 E.coli обладает удивительной гомогенностью. Гетерогенность может быть установлена только в отношении заместителей фосфатов (РР и P-Etn vs. P и Р),что впервые описывается в этой форме. Посредством комплексных NMR-анализов можно четко определить P-Etn-заместитель в олигосахариде ядра R1-олигосахарида ядра в позиции 2 второй гептозы (Hep"). Окончательная структура LPS штаммаDSM 6601 представлена на фиг. 7. В дальнейшем изобретение более подробно поясняется с помощью примеров: Пример 1. Получение LPSLPS получали из промытой и высушенной бактериальной массы после модифицированной экстракции фенол/вода; как указано O.Westphalpp.83-91. 47 г лиофилизированных бактерий, которые перед этим дважды промывали дистиллированной водой, экстрагировали по Westphal иJann в соответствии с модифицированной инструкцией. Модификация состояла в дополнительной ферментативной обработке (ДНКазой,РНКазой, протеиназой К) водного экстракта,которая служит для удаления случайных чужеродных белков и ДНК/РНК-составных частей. Далее к водной фазе (около 1,2 л) при комнатной температуре примешивали 20 мг ДНКазы(рибонуклеаза А, бычья поджелудочная железа,Sigma) и 20 мг ДНКазы (ДНКаза I, бычья поджелудочная железа, степень II, Sigma), смесь перемешивали при комнатной температуре в течение 30 ч. Далее добавляли 20 мг протеиназы К (Tritirachium album, Boehringer, Mannheim),затем перемешивали 12 ч. Полученную суспензию трижды подвергали диализизу через 15 л дистиллированной воды при 4 С в течение 24 ч и затем лиофилизировали. Ферментсодержащий экстракт промывали в дистиллированной воде до тех пор, пока не была достигнута окончательная концентрация около 50 мг/мл. Суспензию три раза центрифугировали с ультразвуком в холоде (155.000 х г, 4 С, 4 ч). Осадок (LPS) помещали в 150 мл дистиллированной воды и еще один раз подвергали диализу через воду в течение трх дней и после этого лиофилизировали (выход LPS: 1,45 г, 3.1% м/м). Пример 2. Анализ LPS из штамма DSM 6601 E.coli. Гексозамин (HexN) (обозначаемый здесь как глюкозамин + галактозамин, GlcN+GaLN) определяли по модифицированному тесту Моргана-Эльсона(в 1959, и кроме того также посредством HPLS(PICO-TAG, Waters). В противоположность тесту Моргана-Эльсона при этом методе анализа можно не только определить количественно и по-отдельности GlcN и GalN, но также можно параллельно установить еще и наличие GlcNфосфата, 2-этаноламина (Etn) и 2-этаноламинфосфата (Etn-P), которые часто встречаются в LPS. Газожидкостную хроматографию (GC) проводили на Varian 3700 GC или Hewlett Packard (HP 5890 серия II) газовом хроматографе на капиллярной колонке (заполненной диоксидом кремния SPB-5, 30 м, Supelco). Комбинированную газожидкостную хроматографию/массспектрометрию (GC-MS) проводили в массспектрометре (HP модель 5989), который оснащен НР-1 капиллярной колонкой (30 м, HewlettPackard). GC- и соответственно GC-MS анализы применяли для определения нейтрального сахара (Glc, Gal, Hep, Man) в качестве их алдитолацетатов (Sawardeker, J.S., Slonerker, J.H.Jeanes,A. Anal. Chem. 37, 1602-1604 (в 1967, а также для качественного и количественного определения жирных кислот в качестве производных сложных эфиров жирных кислот и метилового эфира после сильного метанолиза (2 Мand Rietschel, E.Th., Analysis of lipopolysaccharide (lipid A) fatty acids, J. Microbiol. Meth. 11,(1990) 195-211) и экстракции с хлороформом. Оба GC-метода анализа начинали при 150 С(изотерма для 3 мин) и посредством линейного температурного градиента повышали от 5 С/мин до 320 С. Фосфат определяли по Lowry(1954 и 2-кето-3-дезокси-D-маннооктулозоновую кислоту (Kdo) посредством теста тиобарбитуровой кислоты (Waravdekar, V.C.Saslaw,L.D., J. Biol. Chem. 234, 1945-1950 (1959. Получение и очистка свободного липоида А и олигосахарида ядраLPS (258,8 мг) суспендировали в 25 мл 0,1 М NaOAc/HOAc (pH 4.4) и подвергали мягкому кислотному гидролизу при 100 С в течение 1 ч. Затем экстрагировали три раза липофильную часть (липоид А) из гидролизата с 25 мл хлороформа (выход 23,2 мг). Далее липоид А из органической фазы очищали с помощью препаративной слоевой хроматографии (PSC) (2 мм PSC силикагеля 60 пластин, E.Merck, Daamstadt),хроматографировали с хлороформом-метанолом-водой 100:75:15 (объем/объем/объем) и вы 5 деляли погружением в дистилированную воду. Таким образом получали 6 фракций, из которых основная фракция (Rf=0,4) представляла собой очищенный дифосфорилированный гексаациллипоид A (DPHLA-Ec6601). Очищенный DPHLAEc6601 (выход 2,06 мг) растворяли в хлороформметаноле 8:2 (объем/объем) и обрабатывали перед MALDI-TOF-MS с ионообменником (Amberlite IRA 120, Н+-форма). Аликвоту (250 г) очищенного DPHLA-Ec6601 использовали для биологических экспериментов. Водную фазу экстракции хлороформа лиофилизировали (выход: 272 мг) и далее очищали олигосахарид с помощью TSK-колонки[3,5 х 90 см, TSK HW-40(S), E.Merck] в пиридине - уксусной кислоте - воде 8:20:2000 (объем/объем/объем). Отдельные фракции олигосахаридов (пул А, В, С и D) анализировали с помощью GC-MS и NMR-спектроскопии. Далее очищали основные фракции (пул А, 28-41; 49,05 мг), которые содержали как составные части сахара О-цепи (Man, GalNac), так и такие же части олигосахарида ядра (Hep, Kdo). Другие фракции содержали моносахариды, не исследованные ближе артефакты Kdo (ангидро- и лактон) и, наконец, соль. В основной фракции TSKразделения все составные части олигосахарида ядра (Kdo, Gal, Hep) и О-цепи (Man, GalNAc) были обнаружены не только при GC-MSанализе, но и при NMR-анализе. Поэтому обработку продолжали далее. Кроме того, в первую очередь проверяли,подходит ли для очистки олигосахарида до гомогенности аналитическая анионно-обменная хроматография при высоком давлении НРАЕС(high pressure anion exchange chromatography). Для этого применяли специфическую HPLCметодику для анализа комплексных структур сахара (DIONEX-система) с аналитической CarboPac PA1 колонкой; (4,6 мм х 250 мм) и линейными градиентами соли (5 мин при 0, затем при 50 мин на 0,52 NaOAc) при скорости тока 1 мл/мин. Элюат определяли с помощью пульсамперометрического детектора (PAD) на восстановительном эквиваленте (молекула сахара). Таким образом получали четыре олигосахаридные фракции, которые далее очищали с помощью семипрепаративной НРАЕС аналогичным способом. Семипрепаративную НРАЕС осуществляли с помощью CarboPac PA1 колонки [(9 мм х 250 мм) Dionex система] с такими же градиентами соли, как при аналитической НРАЕС (5 мин при 0, затем 50 мин на 0,5 М NaOAc) и скорости тока 4 мл/мин. Нанесение олигосахарида(42 мг; пул А из TSK-колонки) при семипрепаративной НРАЕС осуществляли в двух аналогичных НРАЕС-направлениях. Элюат собирали в фракции каждую минуту, и их по отдельности исследовали с помощью аналитической НРАЕС. Таким образом, с помощью семипрепаративной НРАЕС получали две основных фракции (фрак 005217 6 ция I, время удержания tR12 мин и фракция II,tR15 мин). Обе НРАЕС-фракции должны были обессоливать перед MALDI-TOF-MS и NMRанализом с помощью G-10-колонки (2,5 х 120 см) (выход: фракция I 4,68 мг; фракция II 4,39 мг). Усиленная матрицей лазерная десорбция/ионизация времяпролетной (MALDI-TOF) масс-спектрометрии Усиленную матрицей лазерную десорбцию/ионизацию времяпролетной (MALDI-TOF) масс-спектрометрии проводили в Bruker-ReflexII времяпролетном спектрометре (Bruker-FranzenAnalytik, Bremen), a также в линейной конфигурации и в отрицательном режиме при ускоряющем напряжении 20 кВ и запаздывающей экстракции ионов". Пробы вначале растворяли в хлороформе (липоид А) или дистиллированной воде (олигосахаридные фракции) в концентрации 10 мкг/мкл, и 2 мкл их аликвоты растворяли в 2 мкл раствора матрицы, состоящего из 0,5 м 2,4,6-тригидроксиацетофенона (Aldrich, Steinheim) в метаноле. Аликвоты (0,5 мкл) этой смеси наносили на металлический держатель и высушивали феном.Avance DRX-600 спектрометре (Bruker, Rheinstetten) и 13C NMR-спектры на Bruker АМХ-360 спектрометре при 300 К в 2 Н 2 О. Перед каждым измерением пробы дважды лиофилизировали с дейтеризированной водой (2 Н 2 О). В качестве внешнего контрольного сигнала использовали ацетон (Н 2,225 частей на миллион, С 31,45 частей на миллион) или 85% Н 3 РО 4 ( 0 частей на миллион). Для получения NMR-данных использовали стандартное программное обеспечение Bruker (XWINNMR 1.3). Продолжительность перемешивания для TOCSY (total correlated spectroscopy - общей коррелированной спектроскопии) и соответственно NOESY (Nuclear Overhauser enhancement spectroscopy ядерной супер-увеличенной спектроскопии) составляла 100 и соответственно 500 мс. Серологические анализы Серологические анализы проводили как вестерн-блоттинг анализы, которые разрабатывали с тремя различными антителами. 1. Поликлональную анти-О 6-антисыворотку (кролик) получали от штамма DSM 6601 Е. coli (серотип О 6:К 5:Н 1) в Институте гигиены в Гамбурге (профессор Бокемюль). 2. Поликлональную анти-E.coli R1 антисыворотку (кролик, внутреннее обозначение: K299/d58) получали иммунизацией с мутантами шероховатой формы, один из которых имел R1-ядро (анти-R1). 3. Использовали моноклональное антитело(WN1-222-5, внутреннее обозначение F 167),которое широко перекрестно реагирует против 7 всех олигосахаридов ядра Е. coli с минимальной структурой (Rd). Таблица. Анализ компонентов, экстрагированных из Е. coli LPS из штамма DSM 6601 Молярное соотношение отдельных компонентов (в скобках) было стандартным благодаря наличию GalNAc и ClcNAc в О-цепи, при значении миристиновой кислоты (14:0) (1.0 моль 14:0/моль LPS).GlcN определено с помощью анализатора аминокислоты. Значение получено из суммыn.b. не определено. Полученные согласно описанному способуLPS-препараты подвергали электрофорезу на полиакриаламидном геле (PAGE) вместе со сравниваемыми LPS (ср. фиг. 1). Для полученияLPS-полосы окрашивали с помощью чувствительного щелочного метода серебрения ( С.М.gels, Anal. Biochem., 119, 1982, 115-119). Результат исследований представлен на фиг. 1. Пример 3. Биологическая активностьIL-1-активность определяли с помощью анализа MNC-пролиферации в надосадочной жидкости культуры. Моноциты человека (MNC) изолировали из периферической крови добровольных доноров (8 х 105 MNC/200 мл) и их переводили на стекло и одновременно перемещали с тестовым веществом. Для тестирования биологической активности in vitro клетки вначале стимулировали с LPS (10 нг/мл). После периода инкубации продолжительностью 8 ч исследовали 150 мл надосадочной жидкости культуры на высвобождение цитокинов. IL-1 активность определяли с помощью анализа пролиферации фибробластов в надосадочной жидкости культуры. Необходимые для этого фибробласты получали из крайней плоти человека. Пролиферация этих фибробластов повышалась 8 благодаря IL-1. Сравнением кривой действия дозы надосадочной жидкости культуры с кривой стандарта в пробит-анализе определяли биологическую активность в надосадочной жидкости культуры. LPS из известных эндотоксически активных штаммов бактерий (Salmonella friedenau) служит как ссылка (положительный контроль) и поэтому представлен на фиг. 5.TNF-активность в надосадочной жидкости культуры определяли анализом цитотоксичности с помощью TNF-чувствительной клеточной линии L929. Сравнение кривой действия дозы надосадочной жидкости культуры с кривой стандарта в пробит-анализе позволяет определить TNF-активность. Также здесь в качестве положительного контроля служит известный эндотоксически активный LPS из Salmonellafriedenau. Результаты представлены графически на фиг. 6. Результаты показывают, что в отношенииIL-1- и TNF-выделения нет никаких значительных различий между LPS из Salmonella friedenau, который служит в качестве стандарта и положительного контроля, и LPS, обнаруженном в штамме DSM 6601 (фиг. 5 и 6 изображения внизу). Это подтверждается также тем, что липоид А штамма DSM 6601 имеет почти одинаковую активность по покрытию как и высоко очищенный липоидом А из Е. coli (фиг. 5 и 6,изображения сверху). ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Липополисахарид бактерий E.coli штамма DSM 6601, содержащий 8 фосфатных остатков и 0,5 моль P-Etn в расчете на молекулу,имеющий структуру, представленную на фиг. 7. 2. Способ получения липополисахарида по п.1, заключающийся в том, что промытую и высушенную бактериальную массу E.coli штаммаDSM 6601 подвергают экстракции смесью фенол/вода и полученный экстракт обрабатывают РНКазой, ДНКазой и протеиназой K. 3. Применение липополисахарида бактерий E.coli штамма DSM 6601 по п.1, в частности полученного в соответствии с п.2, для модулирования иммунной системы человека и животных, изменения биологической активности других микроорганизмов, получения иммуномодулирующих лекарственных средств для человека и животных, а также в качестве маркера или эталонного вещества для микробиологической или медицинской диагностики in vitro, в качестве иммуностимулятора для вакцинации человека и животных, в качестве основного материала для биотехнологического синтеза и изготовления иммуномодулирующих веществ.Hoist (1999) Eur. J. Biochem., 261, 629-639. Все сахара представлены в D-пиранозной форме. L,D-Hep, L-глицеро-D-манно-гептоза; Kdo Dманно-окт-2-улозоновая кислота, Р, фосфат. Замещение О-цепи на расположенной в стороне глюкозе (GlcIII), вместе с ее аномерией, впервые определено и изображено в настоящей заявке на выдачу патентаII,82) 2 LPS препарат 2 3 LPS препарат 3 4 свободно 5 E. coli O111 LPS 6 Pseudomonas aeruginosa тип Фишера 2 LPS 7 Salmonella minnesota R60 LPS Стрелки указывают на LPS полосы олигосахарида ядра и повторяющееся звено (R/S-мутант,полоса А) и соответственно на LPS со сложным олигосахаридом ядра (полоса В).Res., 131, 277-283. Все сахара находятся в D-пиранозной форме. В противоположность опубликованной Janssonet al. структуре О-цепи E.coli 06 первое повторяющееся звено S/R-мутанта E.coli DSM 6601 связано ргликозидически и впервые определено в настоящей заявке на патент, вместе с местом замещения на расположенной в стороне глюкозе (GlcIII) олигосахарида ядра (см. фиг. 4). Фиг. 3. Структура гексаацильного липоида A Escherichia coliBiochem., 50 (1994)211-276) Цифры в кружках обозначают число атомов углерода настоящей жирной кислоты. Свободная гидроксигруппа GlcN (II) представляет место связывания для Kdo (I) олигосахарида ядра. Фиг. 4. Структура основного каркаса углевода главной фракции в олигосахариде ядра E.coli R1: Фиг. 5. Зависимость от дозы высвобождения IL1 из моноцитов человека, индуцируемое с помощью: 1. липода A Escherichia coli (слева вверху) или липоидом А из штамма DSM 6601 E.coli (справа вверху) и 2. LPS из S.friedenau (слева внизу) и Escherichia coli штамм DSM 6601 (справа внизу). Фиг. 6. Зависимость от дозы высвобождения TNF из моноцитов человека, индуцируемое с помощью: 1. липоида A Escherichia coli (слева вверху) или липоида А из штамма DSM 6601 E.coli (справа вверху); и 2. Фиг. 7. Структура сложного липополисахарида (LPS) из Е. coli DSM 6601. В олигосахариде ядра фосфатные заместители Ри Etn в обеих гептозах HepI и HepII не являются стехиметрическими и поэтому обозначены закрашенными линиями. Позиция и аномерное связывание KdoI с липоидом А происходит по аналогии с другими LPS-структурами Е.
МПК / Метки
МПК: A61P 37/02, C12P 19/04, C07H 15/00, A61K 39/02
Метки: липополисахарид, штамма, способ, бактерий, получения
Код ссылки
<a href="https://eas.patents.su/7-5217-lipopolisaharid-bakterijj-e-coli-shtamma-dsm-6601-i-sposob-ego-polucheniya.html" rel="bookmark" title="База патентов Евразийского Союза">Липополисахарид бактерий e. coli штамма dsm 6601 и способ его получения</a>
Слитый белок, вектор экспрессии слитого белка, рекомбинантный штамм бактерий escherichia coli и вакцинный препарат

Номер патента: 307
Опубликовано: 29.04.1999
Авторы: Леал Ангуло Мария Де Хесус, Насабаль Гальвес Консуэло, Альварес Акоста Анабель, Кинтана Васкес Диогенес, Гомес Родригес Кармен Элена, Гильен Ньето Херардо Энрике, Карпио Муньос Эмилио Луис, Сильва Родригес Рикардо Де Ла Каридад, Дуарте Кано Карлос Антонио, Мартин Дунн Алехандро Мигель
МПК: C07K 14/16, C12N 15/62, A61K 39/095...
Метки: препарат, слитый, рекомбинантный, вакцинный, штамм, escherichia, белок, вектор, бактерий, белка, слитого, экспрессии
Формула / Реферат:
1. Слитый белок, содержащий пептид-стабилизатор, представляющий собой модифицированный N-концевой участок из 47 аминокислот антигена Р64К Neisseria meningitidis B:4:P1.15 структуры и гетерологичный белок или полипептид. 2. Слитый белок по п.1, отличающийся тем, что гетерологичный белок представляет собой белок внешней мембраны Neisseria meningitidis. 3. Слитый белок по п.2, отличающийся тем, что белок внешней мембраны N. meningitidis...
Способ биосинтеза кобаламинов, штаммы бактерий, продуцирующие кобаламины, и способ их получения

Номер патента: 2588
Опубликовано: 27.06.2002
Авторы: Тибо Дени, Дебюсш Лоран, Камерон Беатрис, Крузе Жоэль, Бланш Франсис, Реми Элизабет
МПК: C12N 15/52, C12P 19/42
Метки: получения, биосинтеза, кобаламины, штаммы, бактерий, продуцирующие, способ, кобаламинов
Формула / Реферат:
1. Способ биосинтеза кобаламинов путем ферментации прокариотического микроорганизма, продуцирующего кобаламины, отличающийся тем, что в качестве указанного микроорганизма используют рекомбинантный штамм микроорганизма, трансформированный фрагментом ДНК, содержащим гены bluЕ и bluF Rhodobacter capsulatus, кодирующие фермент, вовлеченный в биосинтез O-фосфо-L-треонина, или гомологичным фрагментом и/или фрагментом, который гибридизуется с...
Применение бактериального штамма lactobacillus casei для получения композиции, предназначенной для воздействия на пролиферацию т-клеток

Номер патента: 5081
Опубликовано: 28.10.2004
Авторы: Бонавида Бенджамин, Постер Эрик
МПК: A61P 31/16, A23C 9/123, A61K 35/74...
Метки: применение, получения, воздействия, композиции, пролиферацию, lactobacillus, предназначенной, штамма, casei, бактериального, т-клеток
Формула / Реферат:
1. Применение бактериального штамма вида L. casei для получения композиции, которая может вводиться перорально, для воздействия на пролиферацию Т-клеток, специфичных в отношении антигенов патогенных микроорганизмов, для усиления специфического системного иммунного ответа на патогенный микроорганизм. 2. Применение по п.1, характеризующееся тем, что указанный патогенный микроорганизм представляет собой патоген дыхательных путей. 3. Применение по...
Штамм бактерий bifidobacterium bifidum 791/баг, используемый для получения жидкого концентрата бифидобактерий, кисломолочных лечебно-диетических продуктов и биологически активных добавок

Номер патента: 2614
Опубликовано: 27.06.2002
Авторы: Молокеев Алексей Владимирович, Никулин Леонид Георгиевич, Карих Татьяна Леонидовна, Байбаков Владимир Иванович
МПК: A61K 35/74, C12N 1/20, A23C 9/12...
Метки: концентрата, жидкого, лечебно-диетических, продуктов, активных, bifidobacterium, кисломолочных, bifidum, бактерий, штамм, бифидобактерий, получения, биологически, используемый, добавок
Формула / Реферат:
Штамм Bifidobacterium bifidum ь 791/БАГ (коллекционный ь В-686 НИИ коллекций культур микроорганизмов ГНЦ ВБ "Вектор"), используемый для получения жидкого концентрата бифидобактерий, кисломолочных лечебно-профилактических продуктов и биологически активных добавок.
Способ выщелачивания руды в насыпной массе с помощью бактерий

Номер патента: 3499
Опубликовано: 26.06.2003
Автор: Хантер Колин Джон
МПК: C22B 3/18
Метки: насыпной, массе, выщелачивания, бактерий, способ, помощью, руды
Формула / Реферат:
1. Способ выщелачивания в насыпной массе с помощью бактерий, отличающийся тем, что сооружают насыпную массу руды для окисления содержащихся в ней сульфидных минералов, сооружают биологический контактор с посевом бактерий, окисляющих двухвалентное железо, сооружают, по меньшей мере, один бассейн для подачи выщелачивающего раствора к насыпной массе и биологическому контактору и приема выщелачивающего раствора из насыпной массы и биологического...
Предыдущий патент: Синергетические инсектицидные композиции
Следующий патент: Прицеп-роспуск
Случайный патент: Сухопорошковый ингалятор