Варианты фермента аспарагиназы и их применение
Номер патента: 18424
Опубликовано: 30.07.2013
Авторы: Морман Лисетте, Стор Марк Кристиаан, Ланге Де Илсе, Лаан Ван Дер Ян Метске


























Формула / Реферат
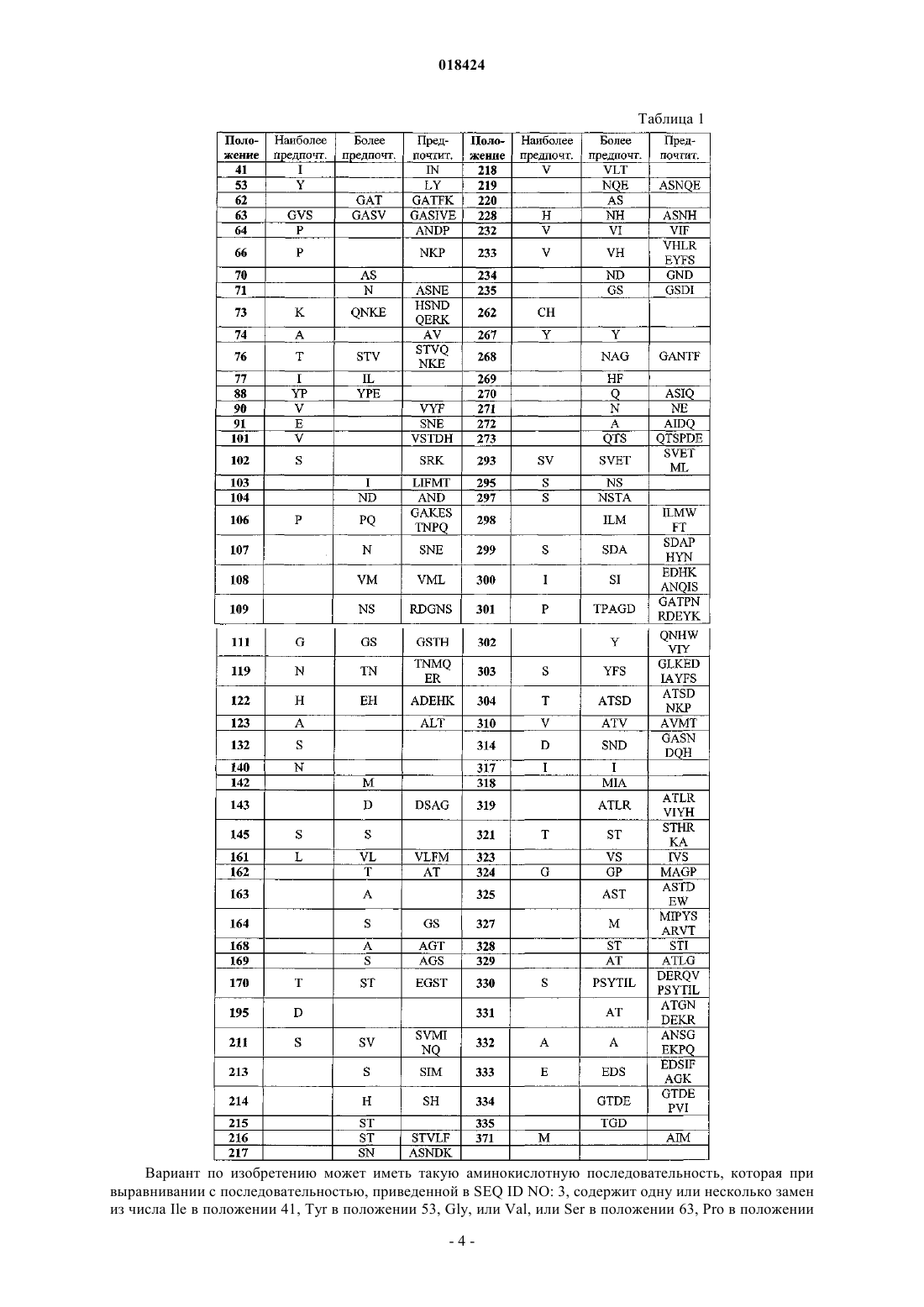
1. Вариант исходного полипептида, обладающего аспарагиназной активностью, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 63, 64, 66, 73, 74, 76, 77, 88, 90, 91, 101, 106, 111, 119, 122, 123, 132, 140, 161, 170, 195, 228, 262, 293, 297, 299, 300, 310, 314, 319, 321, 324 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид по меньшей мере на 80% гомологичен SEQ ID NO: 3, где вариант обладает удельной активностью, которая выше, чем у исходного полипептида при измерении при одном и том же значении pH, по крайней мере при одном значении pH.
2. Вариант по п.1, у которого исходный полипептид имеет последовательность, приведенную в SEQ ID NO: 3.
3. Вариант по любому из пп.1, 2, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит одну или более замен из числа Ile в положении 41, Tyr в положении 53, Gly, или Val, или Ser в положении 63, Pro в положении 64, Pro в положении 66, Lys в положении 73, Ala в положении 74, Thr в положении 76, Ile в положении 77, Tyr или Pro в положении 88, Val в положении 90, Glu в положении 91, Val в положении 101, Pro в положении 106, Gly в положении 111, Asn в положении 119, His в положении 122, Ala в положении 123, Ser в положении 132, Asn в положении 140, Leu в положении 161, Thr в положении 170, Asp в положении 195, His в положении 228, Cys или His в положении 262, Ser или Val в положении 293, Ser в положении 297, Ser в положении 299, Ile в положении 300, Val в положении 310, Asp в положении 314, Ala в положении 319, Thr в положении 321, Gly в положении 324 или Met в положении 371, причем данные положения определяются по отношению к SEQ ID NO: 3.
4. Вариант по любому из пп.1-3, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего любой из аминокислот: 53, 63, 64, 66, 73, 74, 76, 77, 88, 101, 140, 170, 293, причем данные положения определяются по отношению к SEQ ID NO: 3.
5. Вариант по любому из пп.1-4, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит одну или более замен из числа Tyr в положении 53, Gly или Val в положении 63, Pro в положении 64, Pro в положении 66, Lys в положении 73, Ala в положении 74, Thr в положении 76, Ile в положении 77, Tyr или Pro в положении 88, Val в положении 101, Asn в положении 140, Thr в положении 170, Ser или Val в положении 293, причем данные положения определяются по отношению к SEQ ID NO: 3.
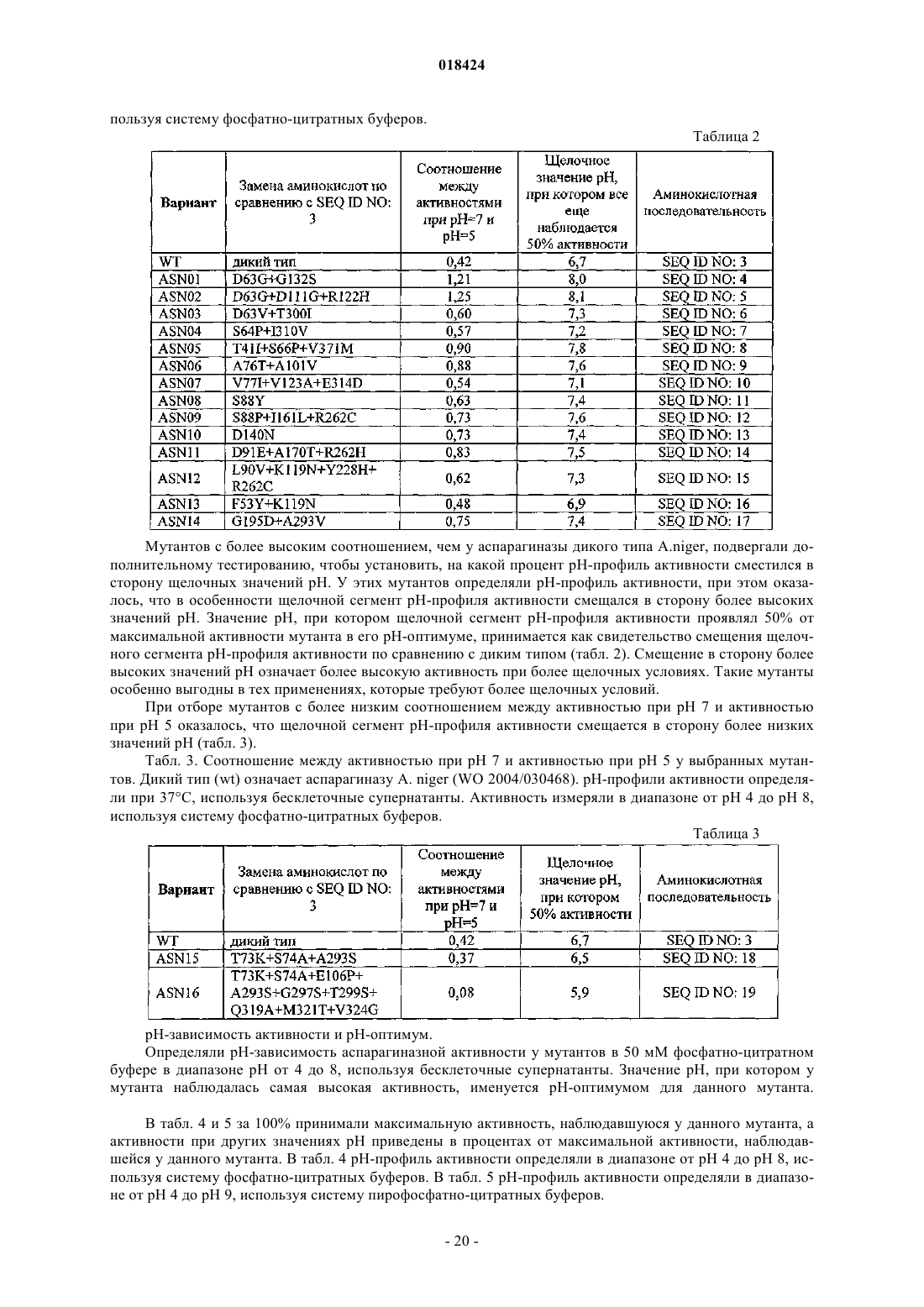
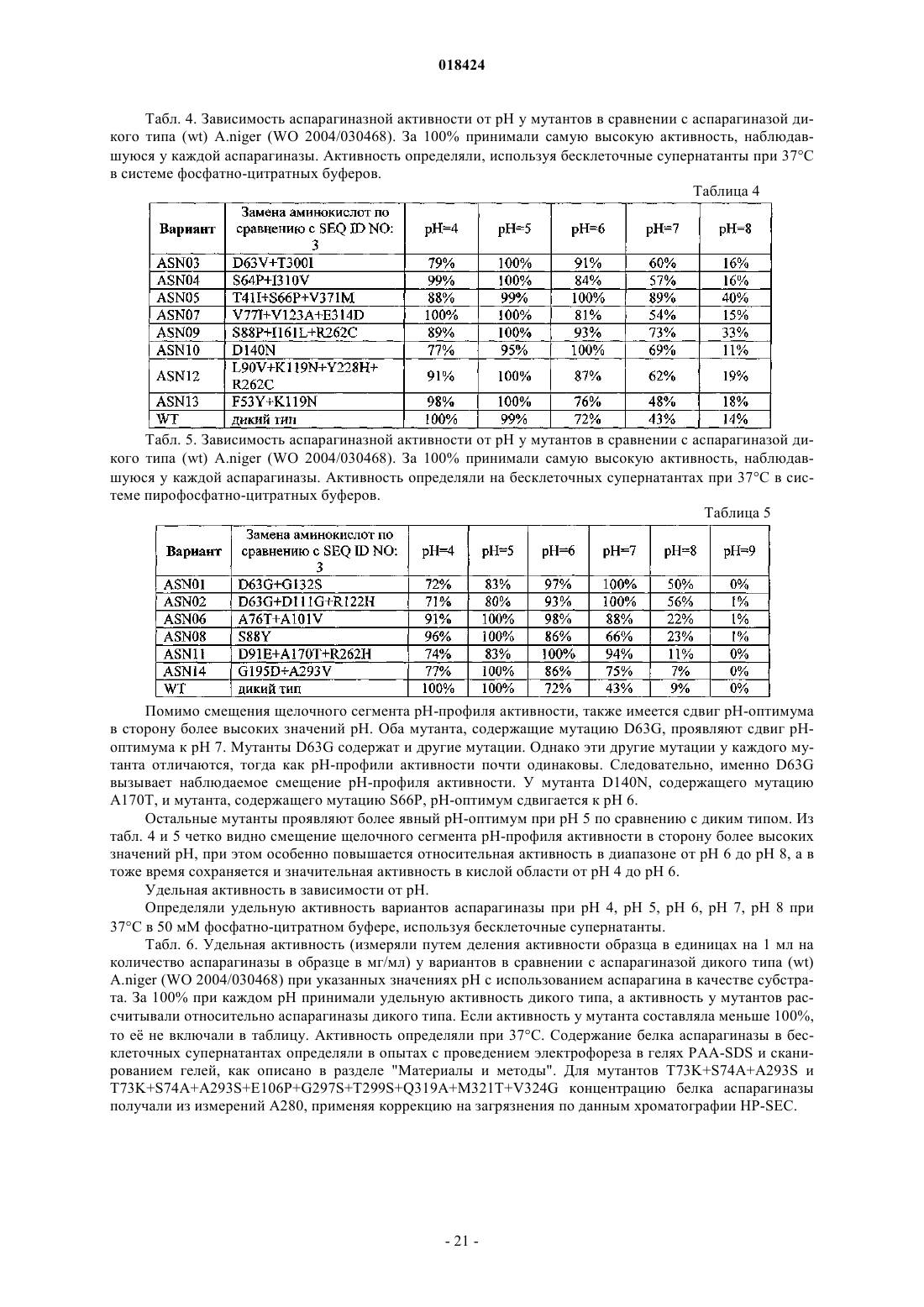
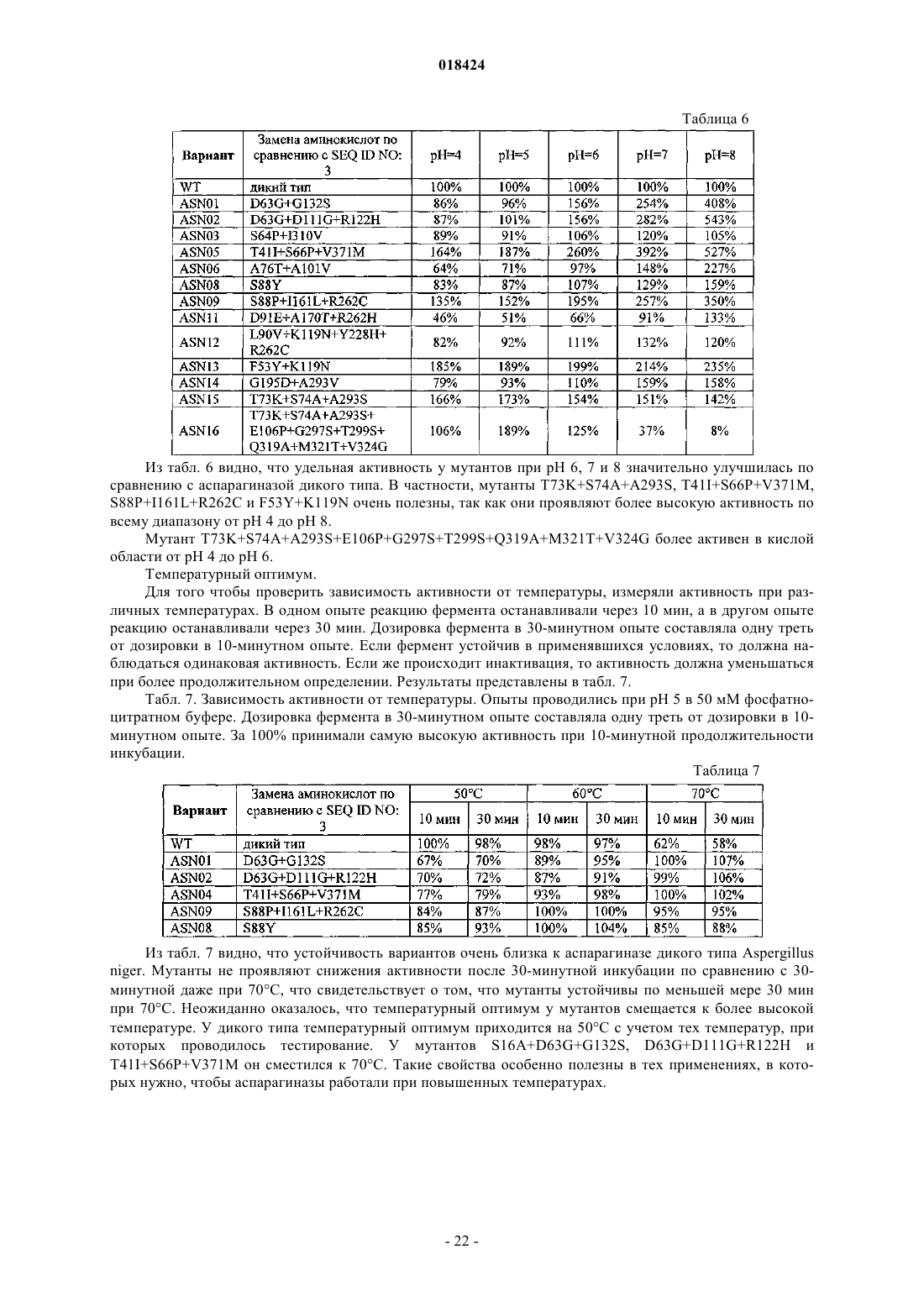
6. Вариант по любому из пп.1-5, причем вариант имеет аминокислотную последовательность согласно любой из SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19.
7. Вариант по любому из пп.1-6, который по меньшей мере на 95% гомологичен исходному полипептиду аспарагиназы.
8. Вариант по любому из пп.1-7, который имеет более высокую удельную активность, чем у исходного полипептида, измеренную при одном и том же значении pH, по меньшей мере при одном значении pH от 4 до 8.
9. Вариант по любому из пп.1-8, который имеет более высокий pH-оптимум, чем у исходного полипептида.
10. Вариант по любому из пп.1-9, который дополнительно содержит одну или более замен аминокислотных остатков, помимо тех, что определены в п.1.
11. Вариант по п.10, который содержит от 1 до 5 дополнительных замен, помимо тех, что определены в п.1.
12. Последовательность нуклеиновой кислоты, кодирующая вариант по любому из пп.1-11.
13. Конструкция нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты по п.12, функционально соединенную с одной или несколькими контрольными последовательностями, способными управлять экспрессией аспарагиназы в подходящем экспрессионном хозяине.
14. Рекомбинантный экспрессионный вектор, содержащий конструкцию нуклеиновой кислоты по п.13.
15. Рекомбинантная клетка-хозяин, содержащая экспрессионный вектор по п.14.
16. Способ получения аспарагиназы, включающий культивирование клетки-хозяина по п.15 в условиях, способствующих продукции аспарагиназы, и выделение аспарагиназы.
17. Способ получения варианта полипептида аспарагиназы, который включает:
a) выбор исходного полипептида аспарагиназы;
b) замену по меньшей мере одного аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 63, 64, 66, 73, 74, 76, 77, 88, 90, 91, 101, 106, 111, 119, 122, 123, 132, 140, 161, 170, 195, 228, 262, 293, 297, 299, 300, 310, 314, 321, 324 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид по меньшей мере на 80% гомологичен SEQ ID NO: 3;
c) необязательно, замену одной или нескольких дополнительных аминокислот, приведенных в b);
d) получение варианта, являющего результатом стадий а)-с);
e) определение удельной активности по меньшей мере при одном значении pH и/или pH-оптимума этого варианта и
f) отбор варианта, обладающего повышенной удельной активностью по меньшей мере при одном значении pH по сравнению с исходным полипептидом аспарагиназы и/или более высоким pH-оптимумом по сравнению с исходным полипептидом аспарагиназы, получая посредством этого вариант полипептида аспарагиназы.
18. Способ по п.17, в котором исходный полипептид аспарагиназы имеет последовательность, приведенную в SEQ ID NO: 3.
19. Способ по п.18, в котором на стадии b) заменяемые аминокислотные остатки соответствуют одной или нескольким аминокислотам из числа Ile в положении 41, Tyr в положении 53, Gly, или Val, или Ser в положении 63, Pro в положении 64, Pro в положении 66, Lys в положении 73, Ala в положении 74, Thr в положении 76, Ile в положении 77, Tyr или Pro в положении 88, Val в положении 90, Glu в положении 91, Val в положении 101, Pro в положении 106, Gly в положении 111, Asn в положении 119, His в положении 122, Ala в положении 123, Ser в положении 132, Asn в положении 140, Leu в положении 161, Thr в положении 170, Asp в положении 195, His в положении 228, Cys или His в положении 262, Ser или Val в положении 293, Ser в положении 297, Ser в положении 299, Ile в положении 300, Val в положении 310, Asp в положении 314, Ala в положении 319, Thr в положении 321, Gly в положении 324 или Met в положении 371, причем данные положения определяются по отношению к SEQ ID NO: 3.
20. Композиция, обладающая аспарагиназной активностью, содержащая эффективное количество аспарагиназы, являющейся вариантом по любому из пп.1-11 или вариантом, полученным способом по любому из пп.17-19, и другой ингредиент.
21. Применение аспарагиназы по любому из пп.1-11 или композиции по п.20 для получения пищевого продукта.
22. Применение аспарагиназы по любому из пп.1-11 или композиции по п.20 для снижения количества акриламида, образующегося в термически обработанном пищевом продукте на основе содержащего аспарагин исходного материала.
23. Способ обработки пищевого продукта для уменьшения в нем количества аспарагина, включающий по меньшей мере одну стадию нагревания, который включает добавление одного или нескольких ферментов аспарагиназы по любому из пп.1-11 или композиции по п.20 в промежуточную форму указанного пищевого продукта, при этом фермент добавляют перед указанной стадией нагревания в количестве, эффективном для снижения уровня аспарагина, содержащегося в указанной промежуточной форме пищевого продукта.
24. Применение аспарагиназы по любому из пп.1-11 или композиции по п.20 для получения медикамента для лечения опухолей.
Текст
ВАРИАНТЫ ФЕРМЕНТА АСПАРАГИНАЗЫ И ИХ ПРИМЕНЕНИЕ Изобретение касается недавно идентифицированных вариантов полипептида аспарагиназы согласно SEQ ID NO: 3 и последовательностей полинуклеотидов, кодирующих эти новые варианты аспарагиназы. Кроме того, изобретение касается применения этих новых вариантов аспарагиназы в промышленных процессах. Лаан ван дер Ян Метске, Стор Марк Кристиаан, Ланге Де Илсе, Морман Лисетте (NL) Соболев А.Ю. (RU)(71)(73) Заявитель и патентовладелец: ДСМ АйПи АССЕТС Б.В. (NL) Область техники, к которой относится изобретение Изобретение касается вариантов полипептида аспарагиназы и последовательностей полинуклеотидов, содержащих гены, кодирующие эти варианты аспарагиназы. В изобретении представлен способ идентификации подходящих вариантов аспарагиназы. Изобретение также касается способов применения этих вариантов белка в промышленных процессах. Также охвачены изобретением клетки, трансформированные полинуклеотидами по изобретению, пригодными для получения этих белков и клеток, причем белки согласно изобретению генетически модифицированы для повышения или снижения активности и/или уровня экспрессии. Изобретение также касается способов применения этих вариантов аспарагиназы в промышленных процессах. Сведения о предшествующем уровне техники Недавно сообщали о появлении акриламида во многих пищевых продуктах при нагревании (Tarekeet al., Chem. Res. Toxicol. 13, 517-522 (2000. Поскольку акриламид считается возможным канцерогеном для животных и человека, эти данные вызвали озабоченность во всем мире. Дальнейшие исследования показали, что значительные количества акриламида обнаруживаются в целом ряде распространенных продуктов при выпекании, жарке и приготовлении в печи, и было показано, что наличие акриламида в продуктах является результатом процесса нагревания. В работе Mottram et al., Nature. 419: 448 (2002) было предположено, что акриламид образуется из аминокислот: и восстанавливающих сахаров в результате реакции Майяра (Maillard). Согласно этой гипотезе акриламид может образоваться при реакции Майяра. За цвет, запах и вкус при выпекании и поджаривании главным образом ответственна реакция Майяра. На пути, ведущему к акриламиду, с реакцией Майяра связано и расщепление аминокислот по Штреккеру (Strecker). Образование акриламида становилось заметным при повышении температуры выше 120C, а максимальная скорость его образования наблюдалась примерно при 170C. Максимальный уровень акриламида наблюдался в присутствии аспарагина и глюкозы, тогда как глутамин и аспарагиновая кислота давали только следовые количества. Действующий в ЕС предел для переноса акриламида в продукты из контактирующих с ними пластиков составляет 10 ppb (10 мкг/кг). Хотя пока еще не установлен предел для акриламида, образующегося при приготовлении пищи, однако тот факт, что многие продукты превышают эту величину, особенно блюда из круп, хлебопродукты и продукты из картофеля или кукурузы, вызывает озабоченность. Известно, что некоторые растительные материалы содержат значительное количество аспарагина. В картофеле аспарагин является преобладающей свободной аминокислотой (940 мг/кг, что соответствует 40% от общего содержания аминокислот), а в пшеничной муке содержание аспарагина составляет около 167 мг/кг, что соответствует 14% от общего содержания свободных аминокислот (Belitz and Grosch inFood Chemistry - Springer New York, 1999). To, что акриламид в основном образуется из аспарагина (вместе с восстанавливающими сахарами), может объяснить высокий уровень акриламида в жареных, испеченных или поджаренных растительных продуктах. Следовательно, в интересах здоровья населения существует настоятельная потребность в том, чтобы пищевые продукты содержали существенно меньше акриламида, а предпочтительно его совсем не содержали. Предлагались различные способы снижения содержания акриламида как путем изменения параметров обработки, например температуры или продолжительности нагревания, так и путем химического или энзиматического предотвращения образования акриламида либо путем удаления образовавшегося акриламида. В некоторых патентных заявках изложено применение аспарагиназы для снижения содержания аспарагина и тем самым количества образующегося акриламида. Подходящие для этой цели аспарагиназы были получены из некоторых грибков, например из Aspergillus niger в WO 2004/030468 и Aspergillusoryzae в WO 04/032648. Хотя все L-аспарагиназы катализируют одно и то же химическое превращение, но это не значит,что все они подходят для одинакового применения. При различных применениях будут разные требования к условиям, в которых ферментам придется работать. Физические и химические параметры, которые могут повлиять на скорость энзиматического превращения, - это температура (которая оказывает положительный эффект на скорость химической реакции, но может оказывать отрицательный эффект на стабильность фермента), влагосодержание, pH, концентрация солей, структурная целостность пищевого матрикса, присутствие активаторов или ингибиторов фермента, концентрация субстрата и продуктов реакции и др. Таким образом, все еще существует потребность в улучшении аспарагиназ для некоторых применений с улучшенными свойствами. Цель изобретения Целью изобретения является получение новых вариантов полипептидов аспарагиназы и полинуклеотидов, кодирующих эти варианты. Другой целью является получение рекомбинантных штаммов, продуцирующих такие варианты. Составной частью изобретения является и способ идентификации вариантов, а также способы получения и применения полинуклеотидов и полипептидов по изобретению. Сущность изобретения Изобретением предусмотрены полипептиды, являющиеся вариантами исходного полипептида, обладающего аспарагиназной активностью, причем эти варианты имеют аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит замены аминокислотных остатков, соответствующих любым из аминокислот: 41, 53, 62, 63, 64, 66, 70, 71, 73,74, 76, 77, 88, 90, 91, 101, 102, 103, 104, 106, 107, 108, 109, 111, 119, 122, 123, 132, 140, 142, 143, 145, 161,162, 163, 164, 168, 169, 170, 195, 211, 213, 214, 215, 216, 217, 218, 219, 220, 228, 232, 233, 234, 235, 262,267, 268, 269, 270, 271, 272, 273, 293, 295, 297, 298, 299, 300, 301, 302, 303, 304, 310, 314, 317, 318, 319,321, 323, 324, 325, 327, 328, 329, 330, 331, 332, 333, 334, 335 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3. Изобретением также предусмотрены последовательности нуклеиновых кислот, кодирующих такие варианты; конструкции из нуклеиновой кислоты, содержащие последовательности нуклеиновых кислот, кодирующих варианты по изобретению, функционально соединенные с одной или несколькими контрольными последовательностями, способными управлять экспрессией аспарагиназы в подходящем для экспрессии организме хозяина; рекомбинантные экспрессионные векторы, содержащие такие конструкции из нуклеиновой кислоты; рекомбинантные клетки-хозяева, содержащие такие экспрессионные векторы; способ получения аспарагиназы, включающий культивирование таких клеток-хозяев в условиях,способствующих продуцированию аспарагиназы, и выделение аспарагиназы; способ получения вариантов полипептида аспарагиназы, который включает:a) выбор исходного полипептида аспарагиназы,b) замену по меньшей мере одного аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 62, 63, 64, 66, 70, 71, 73, 74, 76, 77, 88, 90, 91, 101, 102, 103, 104, 106, 107, 108, 109, 111,119, 122, 123, 132, 140, 142, 143, 145, 161, 162, 163, 164, 168, 169, 170, 195, 211, 213, 214, 215, 216, 217,218, 219, 220, 228, 232, 233, 234, 235, 262, 267, 268, 269, 270, 271, 272, 273, 293, 295, 297, 298, 299, 300,301, 302, 303, 304, 310, 314, 317, 318, 319, 321, 323, 324, 325, 327, 328, 329, 330, 331, 332, 333, 334, 335 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3,c) необязательно, замену одной или нескольких дополнительных аминокислот, приведенных в п. b),d) получение препарата варианта, являющегося результатом стадий а)-с),e) определение удельной активности и/или pH-оптимума этого варианта иf) отбор варианта, обладающего повышенной удельной активностью и/или более высоким pHоптимумом по сравнению с исходным полипептидом аспарагиназы, получая при этом вариант полипептида аспарагиназы; композиции, содержащие вариант по изобретению или вариант, полученный способом изобретения; применение композиций по изобретению при получении пищевых продуктов и применение композиций по изобретению для уменьшения количества акриламида, образующегося при термической обработке пищевых продуктов из содержащих аспарагин исходных материалов. Сведения, подтверждающие возможность осуществления изобретения По всему настоящему описанию и прилагаемой формуле изобретения слова "содержать" и "включать" и их варианты, такие как "содержит", "содержащий", "включает" и "включающий", следует понимать включительно. А именно, эти слова предполагают возможность включать другие элементы или нечто целое, и то и другое может быть не указано специально, если контекст это позволяет. Настоящее изобретение касается полипептидов, являющихся вариантами исходного полипептида,обладающего аспарагиназной активностью. Эти варианты имеют аминокислотную последовательность,которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит замены аминокислотных остатков, соответствующих любым из аминокислот: 41, 53, 62, 63, 64, 66, 70, 71, 73, 74, 76,77, 88, 90, 91, 101, 102, 103, 104, 106, 107, 108, 109, 111, 119, 122, 123, 132, 140, 142, 143, 145, 161, 162,163, 164, 168, 169, 170, 195, 211, 213, 214, 215, 216, 217, 218, 219, 220, 228, 232, 233, 234, 235, 262, 267,268, 269, 270, 271, 272, 273, 293, 295, 297, 298, 299, 300, 301, 302, 303, 304, 310, 314, 317, 318, 319, 321,323, 324, 325, 327, 328, 329, 330, 331, 332, 333, 334, 335 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3. Это значит, что при выравнивании последовательности варианта аспарагиназы с последовательностью аспарагиназы из SEQ ID NO: 3 этот вариант должен содержать по меньшей мере одну замену в положении (у варианта), которое соответствует одному из положений, приведенных выше в SEQ ID NO: 3. В этом контексте "замена" означает, что то положение у варианта, которое соответствует одному из положений, приведенных выше в SEQ ID NO: 3, содержит остаток аминокислоты, которая не значится в этом положении у исходного полипептида (причем исходным полипептидом может быть иSEQ ID NO: 3). Те положения у варианта полипептида аспарагиназы по изобретению, которые соответствуют положениям, приведенным выше в SEQ ID NO: 3, могут быть идентифицированы путем выравнивания по-2 018424 следовательности варианта полипептида с последовательностью SEQ ID NO: 3, например методом(подробно об этой программе см. ниже). При этом можно идентифицировать те положения у варианта,которые соответствуют вышеприведенным положениям в SEQ ID NO: 3 и именуются положениями, определенными по отношению к SEQ ID NO: 3. Исходным полипептидом аспарагиназы, который может быть использован в настоящем изобретении, может быть любая аспарагиназа (ЕС 3.5.1.1). Подходящий полипептид аспарагиназы может быть получен из различных источников, таких, к примеру, как растения, животные или микроорганизмы. Например, аспарагиназа может быть получена из каких-либо видов Escherichia, Erwinia, Streptomyces, Pseudomonas, Aspergillus и Bacillus. Примером подходящего вида Escherichia является Escherichia coli. Примером подходящего вида Erwinia является Erwinia chrysanthemi. Примерами подходящих видов Streptomyces являются Streptomyces lividans и Streptomyces murinus. Примерами подходящих видов Aspergillus являются Aspergillus oryzae, Aspergillus nidulans или Aspergillus niger. Примерами подходящих видов Bacillus являются Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacilluscoagulans, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus stearothermophilus, Bacillus subtilis или Bacillus thuringiensis. Примеры методов, пригодных для получения аспарагиназы из штаммов Bacillus, Streptomyces,Escherichia или Pseudomonas, описаны в WO 03/083043. Примеры методов, пригодных для получения аспарагиназы из Aspergillus, описаны в WO 2004/030468 и WO 04/032468. Предпочтительным исходным полипептидом аспарагиназы, подходящим для использования в настоящем изобретении, является полипептид, имеющий последовательность, приведенную вSEQ ID NO: 3, либо по меньшей мере на 80% гомологичную SEQ ID NO: 3, например по меньшей мере на 85% гомологичную SEQ ID NO: 3, по меньшей мере на 90% гомологичную SEQ ID NO: 3, к примеру по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% гомологичнуюSEQ ID NO: 3. Аминокислотные остатки у варианта по изобретению, которые подвергаются замене по сравнению с последовательностью, приведенной в SEQ ID NO: 3, соответствуют положениям: 41, 53, 62, 63, 64, 66,70, 71, 73, 74, 76, 77, 88, 90, 91, 101, 102, 103, 104, 106, 107, 108, 109, 111, 119, 122, 123, 132, 140, 142, 143,145, 161, 162, 163, 164, 168, 169, 170, 195, 211, 213, 214, 215, 216, 217, 218, 219, 220, 228, 232, 233, 234,235, 262, 267, 268, 269, 270, 271, 272, 273, 293, 295, 297, 298, 299, 300, 301, 302, 303, 304, 310, 314, 317,318, 319, 321, 323, 324, 325, 327, 328, 329, 330, 331, 332, 333, 334, 335 и 371, которые определяются по отношению к последовательности SEQ ID NO: 3. Вариант может содержать замены по одному или нескольким из указанных положений, например по 2, 3, 4, по меньшей мере 5, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 55, по меньшей мере 60, по меньшей мере 65, по меньшей мере 70 или по всем из указанных положений. Предпочтительное подмножество положений для замены задается следующими положениями: 41,53, 62, 63, 64, 66, 70, 71, 73, 74, 76, 77, 88, 90, 91, 101, 102, 103, 104, 106, 107, 108, 109, 111, 119, 122, 123,132, 140, 142, 143, 145, 161, 162, 163, 164, 168, 169, 170, 195, 211, 214, 215, 216, 218, 220, 228, 232, 233,262, 267, 293, 295, 297, 298, 299, 300, 301, 303, 304, 310, 314, 317, 319, 321, 324, 330, 332, 333, 334 и 371,которые определяются по отношению к последовательности SEQ ID NO: 3. Вариант может содержать замены по одному или нескольким из указанных положений, например по 2, 3, 4, по меньшей мере 5, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45 или по всем из указанных положений. Более предпочтительное подмножество положений для замены задается следующими положениями: 41, 53, 63, 64, 66, 73, 74, 76, 77, 88, 90, 91, 101, 106, 111, 119, 122, 123, 132, 140, 145, 161, 170, 195,211, 218, 228, 232, 233, 262, 267, 293, 295, 297, 299, 300, 301, 303, 304, 310, 314, 317, 321, 324, 330, 332,333 и 371, которые определяются по отношению к последовательности SEQ ID NO: 3. Еще более предпочтительное подмножество положений для замены задается следующими положениями: 53, 63, 64, 66, 73, 74, 76, 77, 88, 101, 140, 170, 293, которые определяются по отношению к последовательности SEQ ID NO: 3. Вариант может содержать замены по одному или нескольким из указанных положений, например по 2, 3, 4, по меньшей мере 5, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20 или по всем из указанных положений. Вариант по изобретению содержит одну или несколько замен, как определено выше. В этом контексте "замена" означает, что то положение у варианта, которое соответствует одному из положений,приведенных выше в SEQ ID NO: 3, содержит остаток аминокислоты, которая не значится в этом положении у исходного полипептида (причем исходным может быть и SEQ ID NO: 3). Предпочтительные замены приведены в табл. 1 (при этом положения определяются по отношению к последовательности, приведенной в SEQ ID NO: 3). Вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит одну или несколько замен из числа Ile в положении 41, Tyr в положении 53, Gly, или Val, или Ser в положении 63, Pro в положении 64, Pro в положении 66, Lys в положении 73, Ala в положении 74, Thr в положении 76, Ile в положении 77, Tyr или Pro в положении 88, Val в положении 90, Glu в положении 91, Val в положении 101, Pro в положении 106, Gly в положении 111, Asn в положении 119, His в положении 122, Ala в положении 123,Ser в положении 132, Asn в положении 140, Ser в положении 145, Leu в положении 161, Thr в положении 170, Asp в положении 195, Ser в положении 211, Val в положении 218, His в положении 228, Val в положении 232, Val в положении 233, Cys или His в положении 262, Tyr в положении 267, Ser или Val в положении 293, Ser в положении 295, Ser в положении 297, Ser в положении 299, Ile в положении 300, Pro в положении 301, Ser в положении 303, Thr в положении 304, Val в положении 310, Asp в положении 314,Ile в положении 317, Thr в положении 321, Gly в положении 324, Ser в положении 330, Ala в положении 332, Glu в положении 333 или Met в положении 371, причем данные положения определяются по отношению к SEQ ID NO: 3. В предпочтительном воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3,содержит одну или несколько замен из числа Tyr в положении 53, Gly или Val в положении 63,Pro в положении 64, Pro в положении 66, Lys в положении 73, Ala в положении 74, Thr в положении 76,Ile в положении 77, Tyr или Pro в положении 88, Val в положении 101, Asn в положении 140,Thr в положении 170, Ser или Val в положении 293, причем данные положения определяются по отношению к SEQ ID NO: 3. В предпочтительном воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3,содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 63,предпочтительно содержит Gly или Val в положении 63, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствуетSEQ ID NO: 3. Примерами такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 63, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, являются полипептид, содержащий замены D63G и G132S (условно названный ASN01), полипептид, содержащий замены D63G, D111G и R122H (условно названныйASN02), и полипептид, содержащий замены D63V и T300I (условно названный ASN03). В другом воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 64, предпочтительно содержит Pro в положении 64, причем данное положение определяется по отношению кSEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 64, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замены S64P и I310V (условно названный ASN04). В следующем воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 66, предпочтительно содержит Pro в положении 66, причем данное положение определяется по отношению кSEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 66, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замены T41I, S66P и V371M(условно названный ASN05). В следующем воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотных остатков, соответствующих аминокислотам 73, 74, 293,предпочтительно содержит одну или несколько замен из числа Lys в положении 73, Ala в положении 74,Ser или Val в положении 293, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примерами такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотных остатков, соответствующих аминокислотам 73, 74, 293, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, являются полипептид, содержащий замены G195D иA293V (условно названный ASN14), полипептид, содержащий замены T73K; S74A и A293S (условно названный ASN15), и полипептид, содержащий замены T73K, S74A, E106P, A293S, G297S, T299S,Q319A, M321T и V324G (условно названный ASN16). В следующем воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотных остатков, соответствующих аминокислотам 76 или 101,предпочтительно содержит одну или несколько замен из числа Thr в положении 76 или Val в положении 101, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотных остатков, соответствующих аминокислотам 76 или 101, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замены А 76 Т иA101V (условно названный ASN06). В следующем воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 77, предпочтительно содержит Ile в положении 77, причем данное положение определяется по отношению кSEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 77, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замены V77I, V123A иE314D (условно названный ASN07). В одном воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 88, предпочтительно содержит Tyr или Pro в положении 88, причем данное положение определяется по отношению кSEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примерами такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 88, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замену S88Y (условно названный ASN08), или полипептид, содержащий замены S88P, I161L и R262C (условно названныйASN09). В другом воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 140, предпочтительно содержит Asn в положении 140, причем данное положение определяется по отношению кSEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 140, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замену D140N (условно названный ASN10). В одном воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 170, предпочтительно содержит Thr в положении 170, причем данное положение определяется по отношению кSEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 170, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замены D91E, А 170 Т и В еще одном воплощении вариант по изобретению может иметь такую аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 53, предпочтительно содержит Tyr в положении 53, причем данное положение определяется по отношению кSEQ ID NO: 3, а исходный полипептид данного варианта предпочтительно соответствует SEQ ID NO: 3. Примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью, приведенной вSEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего аминокислоте 53, причем данное положение определяется по отношению к SEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замены F53Y и K119N (условно названный ASN13). Дальнейшим примером такого варианта исходного полипептида, обладающего аспарагиназной активностью, у которого аминокислотная последовательность при выравнивании с последовательностью,приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 63, 64, 66, 73, 74, 76, 77, 88, 90, 91, 101, 106, 111, 119, 122,123, 132, 140, 145, 161, 170, 195, 211, 218, 228, 232, 233, 262, 267, 293, 295, 297, 299, 300, 301, 303, 304,310, 314, 317, 321, 324, 330, 332, 333 и 371, причем данные положения определяются по отношению кSEQ ID NO: 3, а исходный полипептид соответствует SEQ ID NO: 3, является полипептид, содержащий замены L90V, K119N, Y228H и R262C (условно названный ASN12). Вариант по изобретению также может содержать дополнительные модификации по сравнению с исходным в других положениях, чем указанные выше, например одну или несколько дополнительных замен, вставок или делеций. Вариант по изобретению может содержать комбинацию из различных типов модификаций такого рода. Вариант может содержать 1, 2, 3, 4, по меньшей мере 5, по меньшей мере 10,по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30 или больше таких модификаций (которые все могут быть одного и того же типа или же различных типов модификаций). Как правило, дополнительные модификации могут представлять собой замены. Вариант по изобретению может быть по меньшей мере на 80% гомологичен исходному полипептиду аспарагиназы, например по меньшей мере на 85% гомологичен исходному полипептиду, на 90% гомологичен исходному полипептиду, по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен исходному полипептиду. Вариант по изобретению, как правило, сохраняет активность аспарагиназы (ЕС 3.5.1.1). Это значит,что вариант по изобретению будет способен катализировать гидролиз аспарагина до аспарагиновой кислоты. Следовательно, вариант по изобретению будет способен модифицировать боковую цепь аспарагина, участвующую в образовании акриламида при получении пищевого продукта с участием по меньшей мере одной стадии нагревания. Предпочтительно вариант по изобретению, как правило, проявляет улучшенные свойства по сравнению с исходным полипептидом аспарагиназы, из которого он получен. Такое улучшенное свойство должно быть существенным, если вариант будет применяться так, как изложено ниже, например, в способе получения пищевого продукта. К таким свойствам относятся повышение удельной активности (с тем, чтобы можно было использовать меньшее количество варианта в способе получения пищевого продукта по сравнению с требуемым количеством исходной аспарагиназы), повышение или понижение pH-оптимума, в частности такого pHоптимума, который более подходит для применения в способе получения пищевого продукта (по сравнению с исходной аспарагиназой), и повышение термостойкости. В одном воплощении вариант белка по изобретению может иметь более высокий pH-оптимум, чем у исходного полипептида, или более низкий, чем у исходного полипептида. Предпочтительно pHоптимум у варианта белка будет выше, чем у исходного полипептида. Предпочтительно исходным полипептидом является полипептид согласно SEQ ID NO: 3. Например, аспарагиназа дикого типа из A.niger(приведенная в SEQ ID NO: 3) имеет pH-оптимум от pH 4 до pH 5. Вариант белка по изобретению может быть более алкалофильным, чем тот же фермент дикого типа, т.е. он может, к примеру, иметь pHоптимум от pH 5 до pH 8, предпочтительно от pH 6 до pH 7. Необязательно, вариант белка по изобретению может быть более ацидофильным, чем аспарагиназа дикого типа. Предпочтительно у варианта белка аспарагиназы по изобретению может быть такое значение pH выше pH-оптимума, при котором сохраняется 50% активности аспарагиназы (в дальнейшем обозначается как щелочное значение pH), т.е. больше, чем у исходной аспарагиназы. Если исходной аспарагиназой является аспарагиназа согласно SEQ ID NO: 3, то у варианта белка щелочное значение pH, при котором наблюдается 50% активности, может составлять по меньшей мере 6,9, предпочтительно по меньшей мере 7,0, по меньшей мере 7,5, предпочтительно по меньшей мере 8. Вариант, проявляющий улучшенное свойство по отношению к исходной аспарагиназе, должен проявлять измеримое уменьшение или увеличение соответствующего свойства, как правило, такое, чтобы вариант лучше подходил для применения так, как это изложено ниже, например, в способе получения пищевого продукта. Предпочтительно вариант белка по изобретению может обладать более высокой удельной активностью, чем у исходного полипептида при одном и том же значении pH. При этом под удельной активностью варианта белка подразумевается аспарагиназная активность варианта белка, измеренная в единицах на 1 мг чистого белка. Предпочтительно удельная активность варианта белка по изобретению будет выше по крайней мере при одном значении pH, предпочтительно при pH между 4 и 8, чем у исходного полипептида при измерении при одном и том же значении pH. В другом воплощении изобретения вариант аспарагиназы является более термофильным, чем исходный полипептид аспарагиназы. При этом свойство может уменьшаться по меньшей мере на 10, 20, 30, 40, 50, 60, 70, 80, 90, 95 или на 99%. С другой стороны, свойство может увеличиваться по меньшей мере на 10, 25, 50, 100, 200, 500 или на 1000%. Процент уменьшения или увеличения в этом контексте означает процент уменьшения или увеличения в сравнении с исходным полипептидом аспарагиназы. Специалистам хорошо известно, как измеряется процент таких изменений - сравнением активности исходной аспарагиназы и варианта аспарагиназы. Описанные варианты собирательно охватываются терминами "полипептид по изобретению" или"вариант по изобретению". Термины "пептид" и "олигопептид" считаются синонимами (что общепринято), причем каждый термин может использоваться взаимозаменяемым образом в зависимости от контекста и означает цепочку по меньшей мере из двух аминокислот, соединенных пептидными связями. Слово "полипептид" в настоящем изобретении применяется в отношении цепочек, содержащих более 7 аминокислотных остатков. Все формулы или последовательности олигопептидов и полипептидов пишутся слева направо и в направлении от N- к С-концу. Однобуквенная кодировка аминокислот, используемая в настоящем изобретении, широко известна в данной области и приведена в Sambrook et al. (Molecular Cloning: A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor,NY, 1989). Под "выделенным" полипептидом или белком подразумевается полипептид или белок, изъятый из его естественного окружения. Например, полученные рекомбинантным способом полипептиды и белки,экспрессируемые в клетках-хозяевах, для целей изобретения считаются выделенными, равно как и природные или рекомбинантные полипептиды, подвергавшиеся существенной очистке любым подходящим методом, таким, к примеру, как одноступенчатый метод очистки, приведенный в Smith and Johnson, Gene 67:31-40 (1988). Вариант полипептида по изобретению может быть выделен и очищен из культуры рекомбинантных клеток известными в этой области методами. Наиболее предпочтительно для очистки применяется метод высокоэффективной жидкостной хроматографии ("ВЭЖХ"). Полипептиды настоящего изобретения включают продукты методов химического синтеза и продукты, полученные рекомбинантными методами из прокариотических или эукариотических клетокхозяев, в том числе бактериальных, дрожжевых клеток, клеток высших растений, насекомых и млекопитающих. В зависимости от клеток-хозяев, используемых в рекомбинантном методе получения, полипептиды настоящего изобретения могут быть гликолизированными или негликолизированными. Кроме того,полипептиды по изобретению могут содержать и модифицированный остаток начального метионина, в некоторых случаях как результат опосредованных хозяином процессов. Изобретением также предусмотрены биологически активные фрагменты вариантов полипептидов по изобретению. Такие фрагменты считаются охваченными термином "вариант по изобретению". Биологически активные фрагменты вариантов полипептидов по изобретению включают полипептиды, имеющие аминокислотные последовательности, достаточно идентичные или происходящие от аминокислотной последовательности варианта белка по изобретению, которые содержат меньше аминокислот, чем полноразмерный белок, но проявляют по меньшей мере одну биологическую активность соответствующего полноразмерного белка. Как правило, биологически активные фрагменты содержат домен или мотив, обладающий по меньшей мере одной активностью варианта белка по изобретению. Биологически активный фрагмент белка по изобретению может представлять собой полипептид длиной, к примеру, в 10, 25, 50, 100 или больше аминокислот. Более того, рекомбинантными методами могут быть получены и другие биологически активные части, из которых удалены остальные участки белка, и исследованы на наличие одной или нескольких биологических активностей нативной формы полипептида по изобретению. Как правило, фрагмент белка по изобретению содержит одну или несколько замен, определенных в настоящем изобретении. Изобретением также предусмотрены фрагменты нуклеиновых кислот, кодирующие вышеприведенные биологически активные фрагменты (которые сами представляют собой варианты по изобретению). Как изложено выше, настоящим изобретением предусмотрены полинуклеотиды, кодирующие варианты полипептидов по изобретению. Изобретение также касается выделенных полинуклеотидов, кодирующих по меньшей мере один функциональный домен варианта полипептидов по изобретению. Как правило, такой домен содержит одну или несколько замен, описанных в настоящем изобретении. В одном воплощении изобретения последовательность нуклеиновой кислоты по изобретению кодирует полипептид, причем этот полипептид представляет собой вариант, который имеет аминокислотную последовательность, содержащую одно или несколько усечений и/или по меньшей мере одну замену,делецию и/или вставку аминокислоты по сравнению с последовательностью исходной аспарагиназы. Однако такой полипептид должен содержать одну или несколько замен, описанных в настоящем изобретении. В настоящем изобретении термины "ген" и "рекомбинантный ген" относятся к молекулам нуклеиновых кислот, включающим открытую рамку считывания, кодирующую вариант, как описано в настоящем изобретении. Ген может включать кодирующие последовательности, некодирующие последовательности, интроны и регуляторные последовательности. Это значит, что "ген" в настоящем изобретении может означать выделенную молекулу нуклеиновой кислоты, как определено в настоящем изобретении. Соответственно, термин "ген" в контексте настоящего изобретения относится не только к встречающимся в природе последовательностям. Молекула нуклеиновой кислоты настоящего изобретения может быть создана стандартными методами молекулярной биологии, хорошо известными специалистам, в сочетании с информацией о последовательности, приведенной в настоящем изобретении. Например, нужная молекула нуклеиновой кислоты может быть синтезирована de novo стандартными методами синтеза. Такой способ синтеза обычно представляет собой автоматизированный процесс. С другой стороны, молекула нуклеиновой кислоты по изобретению может быть создана при помощи направленного мутагенеза существующей молекулы нуклеиновой кислоты, к примеру молекулы нуклеиновой кислоты дикого типа. Направленный мутагенез может осуществляться с применением ряда методов, хорошо известных специалистам в этой области. В одном из таких методов, который здесь приводится просто для примера, на плазмидной матрице проводится реакция ПЦР с использованием олигонуклеотидных "праймеров", кодирующих нужную замену. Поскольку праймеры представляют собой концевые участки синтезируемых заново нитей, если во время первого цикла будет несоответствие при связывании с матричной нитью ДНК, то после этого первого цикла образующаяся на основе праймера нить (содержащая мутацию) сравняется по концентрации с исходной матрицей. После следующих циклов она будет возрастать экспоненциально и через 25 циклов превзойдет исходную, не несущую мутации нить в соотношении порядка 8 миллионов к 1, в результате чего образуется почти однородный раствор несущих мутацию амплифицированных фрагментов. Затем матричную ДНК можно удалить посредством энзиматического расщепления, например с помощью такого рестрикционного фермента, как Dpn1, который расщепляет только метилированную ДНК. Матрица,которая происходит из полученного методом щелочного лизиза препарата плазмид и поэтому метилирована, на этой стадии разрушается, а несущая мутацию плазмида сохраняется, так как она была создана invitro и поэтому не метилирована. В таком методе при одной реакции ПЦР можно вводить более чем одну мутацию (кодирующую замену, как описано в настоящем изобретении) в молекулу нуклеиновой кислоты, например используя один или несколько олигонуклеотидов, каждый из которых содержит одно или несколько несовпадений. С другой стороны, можно ввести более чем одну мутацию в молекулу нуклеиновой кислоты путем выполнения более чем одной реакции ПЦР, при каждой из которых вводится одна или несколько мутаций таким образом, что изменения в нуклеиновую кислоту вводятся последовательно, многократно. Нуклеиновая кислота по изобретению может быть создана с использованием в качестве матрицы кДНК, мРНК или же геномной ДНК и соответствующих олигонуклеотидных праймеров с несовпадениями согласно методу направленного мутагенеза, описанному выше. Полученная при этом молекула нуклеиновой кислоты может быть клонирована в соответствующий вектор и охарактеризована методами анализа последовательности ДНК. Последовательность нуклеиновой кислоты по изобретению может содержать одну или несколько делеций, т.е. брешей, в сравнении с исходной аспарагиназой. Такие делеции/бреши могут быть созданы и методом направленного мутагенеза с помощью соответствующих олигонуклеотидов. Методы создания таких делеций хорошо известны специалистам в этой области. Кроме того, олигонуклеотиды, соответствующие или гибридизирующиеся с последовательностями нуклеотидов по изобретению, могут быть получены стандартными методами синтеза, например с помощью автоматизированного синтезатора ДНК. Настоящим изобретением также охвачены комплементарные молекулы нуклеиновых кислот. Молекула нуклеиновой кислоты комплементарна другой последовательности нуклеотидов, если она настолько комплементарна этой другой последовательности нуклеотидов, что может гибридизироваться с ней с образованием устойчивой двойной спирали (дуплекса). Также комплементарные молекулы нуклеиновых кислот включены в настоящее изобретение. Молекула нуклеиновой кислоты, которая комплементарна другой нуклеотидной последовательности, является такой, которая достаточно комплементарна этой другой нуклеотидной последовательности, так что она может гибридизироваться с этой другой нуклеотидной последовательностью, тем самым образуя стабильный дуплекс. Один из аспектов изобретения касается выделенных молекул нуклеиновых кислот, кодирующих вариант по изобретению либо его биологически активный фрагмент или домен, а также молекул нуклеиновых кислот, пригодных для применения в качестве зондов для гибридизации при идентификации молекул нуклеиновых кислот, кодирующих полипептид по изобретению, и фрагментов таких молекул нуклеиновых кислот, пригодных для применения в качестве праймеров для ПЦР при амплификации или мутагенезе молекул нуклеиновых кислот, к примеру, для получения молекул нуклеиновых кислот по изобретению."Выделенный полинуклеотид" или "выделенная нуклеиновая кислота" означает такую ДНК или РНК, которая не соприкасается непосредственно с теми двумя кодирующими последовательностями (одной на 5'-конце и другой на 3'-конце), с которыми она непосредственно соприкасается в естественном геноме того организма, из которого она получена. Так, в одном воплощении выделенная нуклеиновая кислота включает часть или все 5'-некодирующие последовательности (например, промотор), которые непосредственно прилегают к кодирующей последовательности. Следовательно, этот термин охватывает,к примеру, рекомбинантную ДНК, встроенную в вектор, в автономно реплицирующуюся плазмиду или вирус либо в геномную ДНК прокариот или эукариот, либо существующую в виде отдельной молекулы(например, кДНК или фрагмента геномной ДНК, полученных методом ПЦР или при обработке рестрикционными эндонуклеазами) независимо от других последовательностей. Он также охватывает рекомбинантную ДНК, составляющую часть гибридного гена, кодирующего другой полипептид, которая практически свободна от клеточного материала, вирусного материала или культуральной среды (при получении методами рекомбинантной ДНК) либо химических предшественников или других химикатов (при химическом синтезе). Кроме того, "выделенный фрагмент нуклеиновой кислоты" означает такой фрагмент нуклеиновой кислоты, который не встречается в природе в виде фрагмента и не встречается в естественном состоянии. В настоящем изобретении термины "полинуклеотид" или "молекула нуклеиновой кислоты" служат для обозначения молекул ДНК (например, кДНК или геномной ДНК) и молекул РНК (например, мРНК),а также аналогов ДНК или РНК, полученных с помощью аналогов нуклеотидов. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но предпочтительно она представляет собой двухцепочечную ДНК. Нуклеиновая кислота может быть синтезирована с использованием аналогов нуклеотидов или их производных (например, инозина или фосфоротиоатных нуклеотидов). Такие олигонуклеотиды могут использоваться, к примеру, для получения нуклеиновых кислот, обладающих измененной способностью к спариванию оснований или повышенной устойчивостью к нуклеазам. В следующем воплощении изобретения предусмотрены выделенные молекулы нуклеиновых кислот, которые являются антисмысловыми по отношению к молекулам нуклеиновых кислот по изобретению. Термины "гомологичность" или "процент идентичности" в настоящем изобретении применяются взаимозаменяемым образом. В целях настоящего изобретения установлено, что для определения процента идентичности двух аминокислотных последовательностей или двух нуклеотидных последовательностей проводится выравнивание этих последовательностей с целью их оптимального сравнения (например, могут вводиться бреши в первую аминокислотную или нуклеотидную последовательность для оптимального выравнивания со второй аминокислотной или нуклеотидной последовательностью). Затем проводится сравнение аминокислотных или нуклеотидных остатков по соответствующим положениям аминокислот или нуклеотидов. Если какое-то положение в первой последовательности занято тем же аминокислотным или нуклеотидным остатком, что и соответствующее положение во второй последовательности, то эти молекулы идентичны в этом положении. Процент идентичности между двумя последовательностями является функцией от числа идентичных положений между этими последовательностями(т.е. процент идентичности = число идентичных положений/общее число положений (т.е. совпадающих положений)100. Предпочтительно две последовательности имеют одинаковую длину. Сравнение последовательностей может проводиться по всей длине двух сравниваемых последовательностей или по фрагменту этих двух последовательностей. Как правило, сравнение проводится по всей длине двух сравниваемых последовательностей. Однако идентичность последовательностей может устанавливаться по какому-то участку, например, из 20, 50, 100 или больше смежных аминокислотных остатков. Специалистам должно быть известно, что имеется несколько различных компьютерных программ для определения гомологичности между двумя последовательностями. Например, сравнение последовательностей и определение процента идентичности между двумя последовательностями могут осуществляться с помощью математического алгоритма. В предпочтительном воплощении процент идентичности между двумя аминокислотными или нуклеотидными последовательностями определяется с помощью алгоритма Needleman and Wunsch (J. Mol. Biol. (48): 444-453 (1970, встроенного в программу GAP в комплекте программного обеспечения Accelrys GCG (доступно на http://www.accelrys.com/products/gcg/),используя матрицу Blosum 62 либо матрицу РАМ 250 и весовой коэффициент для бреши 16, 14, 12, 10, 8,6 или 4 и весовой коэффициент для длины 1, 2, 3, 4, 5 или 6. Специалистам должно быть известно, что все эти различные параметры дают слегка отличные результаты, но в целом процент идентичности двух последовательностей существенно не меняется при использовании разных алгоритмов. Последовательности белков или нуклеиновых кислот настоящего изобретения также можно использовать в качестве "запрашиваемой последовательности" для проведения поиска в публичных базах данных, например для идентификации других представителей семейства или родственных последовательностей. Такой поиск может проводиться с помощью программ BLASTN и BLASTP (версия 2.0) поAltschul et al. (1990), J. Mol. Biol. 215:403-10. Поиск BLAST по белкам может проводиться с помощью программы BLASTP при значениях score = 50, wordlength = 3, получая аминокислотные последовательности, гомологичные молекулам белков по изобретению. Для получения выравниваний с брешами в целях сравнения можно использовать Gapped BLAST, как описано в Altschul et al. (1997), Nucleic Acids Res. 25(17): 3389-3402. При использовании программ BLAST и Gapped BLAST можно использовать параметры по умолчанию соответствующих программ (например, BLASTP и BLASTN). См. домашнюю страницу Национального центра информации по биотехнологии на http://www.ncbi.nlm.nih.gov/. Следующий аспект изобретения касается векторов, предпочтительно экспрессионных векторов, содержащих нуклеиновую кислоту, кодирующую вариант аспарагиназы по изобретению. В настоящем изобретении термин "вектор" обозначает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой он соединен. Одним из типов вектора является "плазмида", что означает кольцевую петлю двухцепочечной ДНК, в которую можно легировать (вставить) дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, при этом в вирусный геном можно легировать дополнительные сегменты ДНК. Некоторые векторы обладают способностью к автономной репликации в клетках-хозяевах, в которые они введены (например, бактериальные векторы, содержащие бактериальное начало репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) встраиваются в геном клеток-хозяев после введения в клетки и при этом реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны управлять экспрессией тех генов, с которыми они функционально связаны. Такие векторы именуются "экспрессионными векторами". Обычно экспрессионные векторы, имеющие применение в методах рекомбинантной ДНК, имеют вид плазмид. Термины "плазмида" и "вектор" в настоящем изобретении могут использоваться взаимозаменяющим образом, так как плазмида является наиболее распространенным типом вектора. Однако изобретение охватывает и другие разновидности экспрессионных векторов, такие как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции. Рекомбинантные экспрессионные векторы по изобретению содержат нуклеиновую кислоту по изобретению в виде, подходящем для экспрессии е в клетках-хозяевах, а это значит, что рекомбинантный экспрессионный вектор включает одну или несколько регуляторных последовательностей, выбранных исходя из клеток-хозяев, используемых для экспрессирования, которые функционально связаны с экспрессируемой последовательностью нуклеиновой кислоты. В рекомбинантном экспрессионном векторе"функционально связаны" означает то, что данная последовательность нуклеотидов связана с регуляторными последовательностями таким образом, который позволяет экспрессию этой последовательности нуклеотидов (например, в системе транскрипции/трансляции in vitro или в клетках-хозяевах при введении вектора в эти клетки). Термин "регуляторные последовательности" охватывает промоторы, энхансеры и другие контролирующие экспрессию элементы (например, сигнал полиденилирования). Такие регуляторные последовательности описаны, к примеру, в Goeddel, Gene Expression Technology: Methods inEnzymology. 185, Academic Press, San Diego, CA (1990). Регуляторные последовательности включают последовательности, управляющие конститутивной экспрессией последовательности нуклеотидов во многих типах клеток-хозяев, и последовательности, управляющие экспрессией последовательности нуклеотидов только в определенных клетках-хозяевах (например, тканеспецифичные регуляторные последовательности). Специалистам должно быть известно, что конструкция экспрессионного вектора может зависеть от таких факторов, как выбор подлежащих трансформации клеток-хозяев, уровень экспрессии нужного белка и т.д. Экспрессионные векторы по изобретению могут вводиться в клетки-хозяева и при этом продуцировать белки или пептиды, кодируемые нуклеиновыми кислотами, как описано в настоящем изобретении(например, вариант аспарагиназы согласно SEQ ID NO: 3 или его вариант, например функциональный эквивалент или фрагмент, либо слитый белок, включающий один или несколько таких вариантов). Рекомбинантные экспрессионные векторы по изобретению могут предназначаться для экспрессии вариантов белков по изобретению в прокариотических или эукариотических клетках. Например, можно экспрессировать вариант белка по изобретению в таких бактериальных клетках, как Е.coli, в клетках насекомых (с помощью бакуловирусных экспрессионных векторов), дрожжевых клетках или клетках млекопитающих. Подходящие клетки-хозяева обсуждаются более подробно в Goeddel, Gene ExpressionTechnology: Methods in Enzymology. 185, Academic Press, San Diego, CA (1990). С другой стороны, рекомбинантный экспрессионный вектор можно подвергнуть транскрипции и трансляции in vitro, например используя регуляторные последовательности промотора T7 и полимеразу T7. Экспрессионные векторы, применимые в настоящем изобретении, включают векторы хромосомного, эписомного и вирусного происхождения, например векторы, происходящие из бактериальных плаз- 11018424 мид, бактериофагов, дрожжевых эписом, дрожжевых хромосомных элементов, таких вирусов, как бакуловирусы, паповавирусы, вирус осповакцины, аденовирусы, птичьи поксвирусы, вирусы псевдобешенства и ретровирусы, а также вирусы, происходящие из их комбинаций, такие как вирусы, происходящие из генетических элементов плазмид и бактериофагов, такие как космиды и фагомиды. Вставленная ДНК должна быть функционально связана с соответствующим промотором, таким как промотор PL фага лямбда, промоторы lac, trp и tac E.coli, ранние и поздние промоторы SV40 и промоторы вирусных LTR - назовем лишь несколько. Другие подходящие промоторы должны быть известны специалистам. В одном конкретном воплощении предпочтительны такие промоторы, которые способны обеспечить высокий уровень экспрессии аспарагиназы в нитчатых грибах. Такие промоторы известны в данной области. Экспрессионные конструкции могут содержать сайты для инициации транскрипции,терминации и, в транскрибируемой области, сайт связывания с рибосомами для трансляции. Кодирующая часть готовых транскриптов, экспрессируемых этими конструкциями, должна включать инициирующий трансляцию кодон AUG в начале и терминирующий кодон в соответствующем месте на конце транслируемого полипептида. ДНК вектора может быть введена в прокариотические или эукариотические клетки стандартными методами трансформации или трансфекции. В настоящем изобретении термины "трансформация" и"трансфекция" служат для обозначения ряда признанных методов введения чужеродной нуклеиновой кислоты (например, ДНК) в клетки-хозяева, включая методы осаждения фосфатом кальция или хлоридом кальция, трансфекции с помощью ДЭАЭ-декстрана, трансдукции, инфицирования, липофекции,трансфекции с помощью катионных липидов или электропорации. Подходящие методы трансформирования или трансфецирования клеток-хозяев приведены в Sambrook et al. Molecular Cloning: A LaboratoryManual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY,1989, Davis et al., Basic Methods in Molecular Biology (1986) и других лабораторных пособиях. Известно, что при устойчивой трансфекции клеток млекопитающих, в зависимости от экспрессионного вектора и метода трансфекции, лишь небольшая часть клеток может встраивать чужеродную ДНК в свой геном. Обычно для идентификации и отбора этих интегрантов в клетки-хозяева вместе с заданным геном вводится ген, кодирующий селекционный маркер (например, устойчивость к антибиотикам). Предпочтительными селекционными маркерами являются маркеры, придающие устойчивость к лекарственным препаратам, таким как G418, гигромицин и метотрексат. Нуклеиновая кислота, кодирующая селекционный маркер, может вводиться в клетки-хозяева в том же самом векторе, который кодирует вариант белка по изобретению либо в отдельном векторе. Клетки, устойчиво трансфецированные введенной нуклеиновой кислотой, можно идентифицировать путем отбора в присутствии препарата (например, выживут клетки со встроенным геном селекционного маркера, тогда как остальные клетки погибнут). Экспрессирование белков в прокариотах зачастую проводится в Е.coli с помощью векторов, содержащих конститутивные или индуцибельные промоторы, управляющие экспрессией слитых либо неслитых белков. Вектор для слияния добавляет ряд аминокислот к кодируемому им белку, например наN-конец рекомбинантного белка. Как правило, такие векторы для слияния выполняют три задачи: 1) повышение экспрессии рекомбинантного белка; 2) повышение растворимости рекомбинантного белка и 3) облегчение очистки рекомбинантного белка, действуя в качестве лиганда при аффинной очистке. Часто в экспрессионные векторы для слияния вводится сайт протеолитического расщепления на стыке между сливаемой молекулой и рекомбинантным белком, который позволяет отделить рекомбинантный белок от сливаемой молекулы после очистки слитого белка. Как уже сказано, экспрессионные векторы предпочтительно должны содержать селекционные маркеры. Такие маркеры включают дигидрофолатредуктазу или устойчивость к неомицину при культивировании в эукариотических клетках и устойчивость к тетрациклину или ампициллину при культивировании в Е.coli и других бактериях. Репрезентативные примеры подходящих клеток-хозяев включают клетки бактерий, такие как Е.coli, Streptomyces, Salmonella typhimurium и некоторые виды Bacillus; клетки грибков типа Aspergillus, например A.niger, A.oryzae и A.nidulans; дрожжевые клетки типа Kluyveromyces, например K.lactis и/или Puchia, например P.pastoris; клетки насекомых, такие как Drosophila S2 иSpodoptera Sf9; клетки животных, такие как CHO, COS и меланомы Bowes; и клетки растений. Известны подходящие среды и условия культивирования вышеописанных клеток-хозяев. Предпочтительные векторы для бактерий приведены, к примеру, в WO-A1-2004/074468, включенной в настоящее изобретение путем отсылки. Другие подходящие векторы должны быть хорошо известны специалистам. Известные бактериальные промоторы, подходящие для настоящего изобретения, включают промоторы, приведенные в WO-A1-2004/074468, включенной в настоящее изобретение путем отсылки. Транскрипция ДНК, кодирующей вариант по настоящему изобретению у высших эукариот, может быть повышена при введении в вектор последовательности энхансера. Энхансеры представляют собойcis-элементы ДНК, обычно от 10 до 300 п.о., которые действуют, повышая транскрипционную активность промотора у данного типа клеток-хозяев. Примеры энхансеров включают энхансер SV40, который располагается на поздней стороне начала репликации в районе п.о. 100-270, энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне начала репликации и энхансеры аденовирусов. Для секреции транслируемого белка в просвет эндоплазматического ретикулума в периплазматическое пространство или во внешнюю среду можно вставить подходящий сигнал секреции в экспрессируемый полипептид. Сигналы могут быть эндогенными для полипептида либо гетерологичными. Вариант по изобретению может экспрессироваться в таком виде, что он будет включать дополнительные гетерологичные функциональные участки, например сигналы секреции. Вариант по изобретению также может содержать, к примеру, участок из добавочных аминокислот, в частности заряженных аминокислот, вставленный на N-конец полипептида, например, для улучшения стабильности и стойкости в клетках-хозяевах, во время очистки или при последующей обработке и хранении. К тому же к варианту по изобретению можно присоединить молекулы пептидов, способствующих очистке, например добавить остатки гистидина или Т 7-метку. Варианты по изобретению, такие как белки настоящего изобретения или их функциональные эквиваленты, например биологически активные части и фрагменты, могут быть функционально соединены с полипептидом не варианта (например, гетерологичной последовательностью аминокислот) с образованием слитых белков. В данном контексте "полипептид не варианта" означает полипептид, имеющий аминокислотную последовательность, соответствующую белку, который не является существенно гомологичным варианту аспарагиназы по изобретению. В слитом белке вариант по изобретению может соответствовать полноразмерной последовательности либо биологически активному фрагменту полипептида по изобретению. В предпочтительном воплощении слитый белок по изобретению включает по меньшей мере две биологически активные части. В отношении слитого белка термин "функционально соединен" служит для обозначения того, что полипептид варианта и полипептид не варианта слиты друг с другом в одной рамке считывания. Полипептид не варианта может быть пришит к N- или С-концу полипептида варианта. Например, в одном воплощении слитый белок представляет собой белок, у которого последовательность варианта пришита к С-концу последовательностей GST. Такие слитые белки могут способствовать очистке рекомбинантного варианта по изобретению. В другом воплощении слитый белок является вариантом по изобретению, содержащим гетерологичную сигнальную последовательность на N-конце. В некоторых клетках-хозяевах (например, клетках млекопитающих и дрожжевых клетках) при помощи гетерологичной сигнальной последовательности может быть повышена экспрессия и/или секреция варианта по изобретению. В другом примере в качестве гетерологичной сигнальной последовательности может применяться секреторная последовательность белка gp67 оболочки бакуловируса (Current Protocols in Molecular Biology, Ausubel et al., eds., John WileySons, 1992). Другие примеры эукариотических гетерологичных сигнальных последовательностей включают секреторные последовательности мелиттина и плацентарной щелочной фосфатазы человека (Stratagene; La Jolla, California). В еще одном примере полезные прокариотические гетерологичные сигнальные последовательности включают секреторный сигнал phoA(Sambrook et al., supra) и секреторный сигнал белка А (Pharmacia Biotech; Piscataway, New Jersey). Сигнальная последовательность может использоваться для облегчения секреции и выделения варианта по изобретению. Как правило, сигнальные последовательности характеризуются основой из гидрофобных аминокислот, которые обычно отщепляются от зрелого белка при секреции за один или несколько этапов отщепления. Такие сигнальные пептиды содержат сайты для процессинга, которые позволяют отщепление сигнальной последовательности от зрелого белка при прохождении через секреторный путь. Сигнальная последовательность может направлять секрецию варианта, скажем, из клеток эукариотического хозяина, трансформированных экспрессионным вектором, а затем сигнальная последовательность может быть отщеплена, потом или одновременно. После этого вариант по изобретению легко может быть выделен из внеклеточной среды известными методами. С другой стороны, сигнальная последовательность может быть присоединена к данному варианту с помощью последовательности, способствующей очистке, например, к домену GST. Так, например, последовательность, кодирующая вариант по изобретению, может быть слита с последовательностью маркера, такой как последовательность, кодирующая пептид, способствующий очистке слитого варианта по изобретению. В некоторых предпочтительных воплощениях этого аспекта изобретения маркерная последовательность представляет собой гексагистидиновый пептид типа метки, встроенной в вектор pQE (Qiagen, Inc.), среди прочих, многие из которых коммерчески доступны. Как описано в Gentz et al., Proc. Natl. Acad. Sci. USA. 86: 821-824 (1989), к примеру, гексагистидин обеспечивает удобную очистку слитого белка. Другим пептидом, полезным для очистки, является метка HA, которая соответствует эпитопу из белка гемагглютинина вируса гриппа,который описан в Wilson et al., Cell. 37: 767 (1984), к примеру. Слитый белок по изобретению может быть получен стандартными методами рекомбинантной ДНК. Например, фрагменты ДНК, кодирующие последовательности различных полипептидов, легируют друг с другом в одной рамке считывания в соответствии со стандартными методами, к примеру используя тупые концы или выступающие концы для легирования, расщепление рестрикционными ферментами для получения соответствующих концов, заполнение липких концов надлежащим образом, обработку щелочной фосфатазой для предотвращения нежелательного соединения и энзиматическое легирование. В другом воплощении можно синтезировать слитый ген стандартными методами, включая автоматизиро- 13018424 ванные синтезаторы ДНК. С другой стороны, можно провести ПЦР-амплификацию фрагментов генов с помощью якорных праймеров, создающих комплементарные выступы между двумя соседними фрагментами генов, которые затем можно подвергнуть отжигу и повторно амплифицировать, получая последовательность химерного гена (например, см. Current Protocols in Molecular Biology, Ausubel et al., eds., JohnWileySons, 1992). Более того, коммерчески доступны многие экспрессионные векторы, кодирующие готовые молекулы для слияния (например, полипептид GST). В такой экспрессионный вектор можно клонировать кодирующую вариант нуклеиновую кислоту таким образом, чтобы молекула для слияния соединялась с данным вариантом в одной рамке считывания. Термины "функциональные эквиваленты" и "функциональные варианты" в настоящем изобретении используются взаимозаменяемым образом. Функциональные эквиваленты по изобретению представляют собой выделенные фрагменты ДНК, которые кодируют полипептид, проявляющий определенную функцию варианта, как определено в настоящем изобретении. Следовательно, функциональные эквиваленты также охватывают биологически активные фрагменты и сами охватываются термином "вариант" по изобретению. Предпочтительно функциональный эквивалент по изобретению содержит одну или несколько замен, описанных в настоящем изобретении. Однако функциональный эквивалент может содержать одну или несколько модификаций вдобавок к описанным выше заменам. Функциональные эквиваленты нуклеиновой кислоты, как правило, могут содержать молчащие мутации или мутации, не изменяющие биологическую функцию кодируемого полипептида. Соответственно, изобретением предусмотрены молекулы нуклеиновых кислот, кодирующие варианты белка аспарагиназы, которые содержат замены аминокислотных остатков, не существенных для определенной биологической активности. Такие варианты белка отличаются по аминокислотной последовательности от исходной последовательности аспарагиназы, из которой они происходят, но сохраняют по меньшей мере одну его биологическую активность, предпочтительно, по крайней мере, сохраняют аспарагиназную активность. В одном воплощении выделенная молекула нуклеиновой кислоты содержит последовательность нуклеотидов, кодирующую белок, причем этот белок имеет существенно гомологичную аминокислотную последовательность, которая по меньшей мере на 60, 65, 70, 75, 80, 85, 90, 95, 96, 97, 98, 99% или больше гомологична исходной аминокислотной последовательности (например, приведенной вSEQ ID NO: 3). В настоящем изобретении термин "существенно гомологичная" относится к первой аминокислотной или нуклеотидной последовательности, содержащей достаточное или минимальное количество идентичных или эквивалентных (например, с аналогичной боковой цепью) аминокислот или нуклеотидов ко второй аминокислотной или нуклеотидной последовательности с тем, что и первая, и вторая аминокислотная или нуклеотидная последовательность содержит общий домен. Например, аминокислотные или нуклеотидные последовательности, содержащие общий домен и идентичные на 60%, предпочтительно на 65%, более предпочтительно на 70%, еще более предпочтительно на 75, 80, 85, 90, 95, 96, 97,98, 99% или больше, в настоящем изобретении определяются как достаточно идентичные. Специалисты должны понимать, что в нуклеотидные последовательности по изобретению могут вводиться изменения при помощи мутагенеза, которые приводят к изменению аминокислотной последовательности получаемого белка без существенного изменения функции такого белка. Соответственно, вариант аспарагиназы по изобретению предпочтительно представляет собой такой белок, аминокислотная последовательность которого по меньшей мере на 60, 65, 70, 75, 80, 85, 90, 95, 96,97, 98, 99% или больше гомологична исходной аминокислотной последовательности, например, приведенной в SEQ ID NO: 3, и обычно также сохраняет по меньшей мере одну функциональную активность исходного полипептида. Варианты по изобретению, например функциональные эквиваленты белка по изобретению, также можно идентифицировать, например, путем скрининга комбинаторной библиотеки мутантов, например укороченных мутантов белка по изобретению на наличие аспарагиназной активности. В одном воплощении создается разнородная библиотека вариантов путем комбинаторного мутагенеза на уровне нуклеиновой кислоты. Разнородная библиотека вариантов может быть получена, к примеру, путем энзиматического легирования смеси синтетических олигонуклеотидов с последовательностями гена таким образом,чтобы вырожденный набор потенциальных последовательностей белков можно было экспрессировать в виде индивидуальных полипептидов или же в виде набора более крупных слитых белков (например, при фаговом дисплее). Существует целый ряд методов, которые можно использовать для получения библиотек потенциальных вариантов полипептидов по изобретению из последовательности вырожденных олигонуклеотидов. Методы синтеза вырожденных олигонуклеотидов известны в этой области (например,см. Narang (1983), Tetrahedron. 39: 3; Itakura et al. (1984), Annu. Rev. Biochem. 53: 323; Itakura et al. (1984),Science. 198: 1056; Ike et al. (1983), Nucleic Acids Res. 11: 477). Кроме того, библиотеки фрагментов последовательности, кодирующей полипептид по изобретению, можно использовать для получения разнородной популяции полипептидов для скрининга при последующем отборе вариантов. Например, можно создать библиотеку фрагментов кодирующей последовательности путем обработки нуклеазой двухцепочечного ПЦР-фрагмента заданной кодирующей после- 14018424 довательности в таких условиях, когда точечный разрыв происходит только примерно один раз на молекулу, денатурации двухцепочечной ДНК, ренатурации этой ДНК с образованием двухцепочечной ДНК,которая может включать пары смысловых/антисмысловых нитей из продуктов с разными точечными разрывами, удаления одноцепочечных участков из новообразованных дуплексов при обработке нуклеазой S1 и легирования полученной библиотеки фрагментов в экспрессионный вектор. Таким способом можно получить экспрессионную библиотеку, кодирующую N-концевые и внутренние фрагменты заданного белка различного размера. Известно несколько методов скрининга генных продуктов из комбинаторных библиотек, полученных путем точечных мутаций или усечения, и скрининга библиотек кДНК на наличие генных продуктов,обладающих заданным свойством. Наиболее распространенные методы, которые поддаются высокопроизводительному анализу для скрининга больших генных библиотек, обычно включают клонирование генной библиотеки в реплицирующиеся экспрессионные векторы, трансформацию подходящих клеток полученной библиотекой векторов и экспрессию комбинаторных генов в условиях, при которых детектирование требуемой активности способствует выделению вектора, кодирующего тот ген, продукт которого был детектирован. Для идентификации вариантов белка по изобретению в сочетании с методами скрининга может применяться метод рекурсивного группового мутагенеза (REM), который повышает частоту функциональных мутантов в библиотеках (Arkin and Yourvan (1992), Proc. Natl. Acad. Sci. USA,89: 7811-7815; Delgrave et al. (1993), Protein Engineering, 6(3): 327-331). Фрагменты полинуклеотидов по изобретению также могут включать полинуклеотиды, не кодирующие функциональные полипептиды. Такие полинуклеотиды могут функционировать как зонды или праймеры для реакции ПЦР. Нуклеиновые кислоты по изобретению, независимо от того, кодируют они функциональные или нефункциональные полипептиды, могут использоваться в качестве зондов для гибридизации или праймеров для полимеразной цепной реакции (ПЦР). Применение молекул нуклеиновых кислот настоящего изобретения, не кодирующих полипептид, обладающий аспарагиназной активностью, включает, среди прочего, (1) гибридизацию in situ (например, FISH) расправленных метафазных хромосом для уточнения хромосомной локализации кодирующего аспарагиназу гена, как описано в Verma et al., Human Chromosomes: a Manual of Basic Techniques, Pergamon Press, New York (1988); (2) Northern-гибридизацию для детектирования экспрессии мРНК аспарагиназы в определенных тканях и/или клетках и (3) зонды и праймеры, которые могут применяться как средства диагностики при анализе на присутствие нуклеиновой кислоты, гибридизирующейся с таким зондом или праймером в данном биологическом образце (например, образце ткани). Варианты заданного исходного фермента аспарагиназы могут быть получены по следующей стандартной методике: мутагенез (вызывающие ошибки и сбои олигонуклеотиды); первичный скрининг; идентификация улучшенных вариантов (например, в отношении удельной активности); хранение (например, в культуре с глицерином, на чашках с LB-Amp, в виде мини-препарата); рассев штрихом на другую чашку для анализа - вторичный скрининг; секвенирование ДНК; трансформация, к примеру, клеток Aspergillus; культивирование, к примеру, в объеме 100 мл, очистка, DSC. В одном воплощении изобретение касается способа получения варианта полипептида аспарагиназы по изобретению, который включает:a) выбор исходного полипептида аспарагиназы;b) замену по меньшей мере одного аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 62, 63, 64, 66, 70, 71, 73, 74, 76, 77, 88, 90, 91, 101, 102, 103, 104, 106, 107, 108, 109, 111,119, 122, 123, 132, 140, 142, 143, 145, 161, 162, 163, 164, 168, 169, 170, 195, 211, 213, 214, 215, 216, 217,218, 219, 220, 228, 232, 233, 234, 235, 262, 267, 268, 269, 270, 271, 272, 273, 293, 295, 297, 298, 299, 300,301, 302, 303, 304, 310, 314, 317, 318, 319, 321, 323, 324, 325, 327, 328, 329, 330, 331, 332, 333, 334, 335 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3;c) необязательно, замену одной или нескольких дополнительных аминокислот, приведенных в пункте b);d) получение препарата варианта, являющегося результатом стадий а)-с);e) определение удельной активности по меньшей мере при одном значении pH и/или pH-оптимума этого варианта иf) отбор варианта, обладающего повышенной удельной активностью по меньшей мере при одном значении pH по сравнению с исходным полипептидом аспарагиназы и/или более высоким pH-оптимумом по сравнению с исходным полипептидом аспарагиназы, получая при этом вариант полипептида аспарагиназы. В предпочтительном воплощении в способе получения варианта полипептида аспарагиназы по изобретению исходный полипептид аспарагиназы имеет последовательность, приведенную в SEQ ID NO: 3. Более предпочтительно на стадии b) способа по изобретению производится замена по меньшей мере одного аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 63, 64, 66, 73, 74,76, 77, 88, 90, 91, 101, 106, 111, 119, 122, 123, 132, 140, 145, 161, 170, 195, 211, 218, 228, 232, 233, 262, 267,293, 295, 297, 299, 300, 301, 303, 304, 310, 314, 317, 321, 324, 330, 332, 333 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид по меньшей мере на 80% гомологичен SEQ ID NO: 3. Еще более предпочтительно на стадии b) способа по изобретению заменяемые аминокислотные остатки соответствуют одной или нескольким аминокислотам из числа Ile в положении 41,Tyr в положении 53, Gly, или Val, или Ser в положении 63, Pro в положении 64, Pro в положении 66,Lys в положении 73, Ala в положении 74, Thr в положении 76, Ile в положении 77, Tyr илиPro в положении 88, Val в положении 90, Glu в положении 91, Val в положении 101,Pro в положении 106, Gly в положении 111, Asn в положении 119, His в положении 122,Ala в положении 123, Ser в положении 132, Asn в положении 140, Ser в положении 145,Leu в положении 161, Thr в положении 170, Asp в положении 195, Ser в положении 211,Val в положении 218, His в положении 228, Val в положении 232, Val в положении 233,Cys или His в положении 262, Tyr в положении 267, Ser или Val в положении 293,Ser в положении 295, Ser в положении 297, Ser в положении 299, Ile в положении 300,Pro в положении 301, Ser в положении 303, Thr в положении 304, Val в положении 310,Asp в положении 314, Ile в положении 317, Thr в положении 321, Gly в положении 324,Ser в положении 330, Ala в положении 332 ,Glu в положении 333 или Met в положении 371, причем данные положения определяются по отношению к SEQ ID NO: 3. В одном воплощении способа по изобретению на стадии е) определяется удельная активность приpH между 4 и 8. В другом воплощении на стадии е) перед определением удельной активности по меньшей мере при одном значении pH и/или pH-оптимума у варианта можно измерить соотношение, при определенной температуре, между аспарагиназной активностью при pH 7 и аспарагиназной активностью при pH 5 и выбрать такой вариант, у которого данное соотношение будет выше, чем у исходного полипептида аспарагиназы. В другом воплощении способа по изобретению на стадии f) производится отбор варианта, обладающего повышенной удельной активностью по меньшей мере при одном значении pH, предпочтительно при pH между 4 и 8, по сравнению с исходным полипептидом, и/или обладающего более высоким pHоптимумом по сравнению с исходным полипептидом. Предпочтительно вариант обладает повышенной удельной активностью по меньшей мере при одном значении pH, предпочтительно при pH между 4 и 8 по сравнению с исходным полипептидом, и более высоким pH-оптимумом по сравнению с исходным полипептидом. В следующем воплощении способа по изобретению на стадии f) производится отбор варианта, обладающего повышенной удельной активностью по меньшей мере при одном значении pH,предпочтительно при pH между 4 и 8, по сравнению с исходным полипептидом. В следующем воплощении изобретения представлены клетки, например трансформированные клетки-хозяева или рекомбинантные клетки-хозяева, содержащие нуклеиновую кислоту согласно изобретению. "Трансформированные клетки" или "рекомбинантные клетки" представляют собой клетки, в которые (или в предшественниках которых) методами рекомбинантной ДНК была введена нуклеиновая кислота по изобретению. Они охватывают как прокариотические, так и эукариотические клетки, например бактерии, грибки, дрожжи и др., но особенно предпочтительны клетки нитчатых грибов, в особенности Aspergillus niger. Можно выбрать такие клетки-хозяева, которые модулируют экспрессию введенных последовательностей либо модифицируют и подвергают процессингу продукт, кодируемый вставленной последовательностью нуклеиновой кислоты определенным, заданным образом. Такие модификации (например,гликолизирование) и процессинг (например, расщепление) белковых продуктов могут способствовать оптимальному функционированию кодируемого белка. Различные клетки-хозяева обладают характерными и специфическими механизмами посттрансляционного процессинга и модификации белков и генных продуктов. Можно выбрать подходящие линии клеток или системы хозяев, известные специалистам в области молекулярной биологии и микробиологии, чтобы обеспечить требуемую и правильную модификацию и процессинг экспрессируемого чужеродного белка. Для этой цели можно использовать эукариотические клетки-хозяева, обладающие клеточными механизмами для правильного процессинга первичного транскрипта, гликозилирования и фосфорилирования генного продукта. Такие клетки-хозяева хорошо известны в этой области. Клетки-хозяева охватывают и клеточные линии млекопитающих, такие как линии клеток CHO,VERO, BHK, HeLa, COS, MDCK, 293, 3T3, WI38 и хориоидного сплетения. При желании, устойчиво трансфецированная линия клеток может продуцировать вариант по изобретению. Имеется целый ряд общедоступных векторов, подходящих для устойчивой трансфекции клеток млекопитающих, а способы конструирования таких клеточных линий также общеизвестны, например см. Ausubel et al. (supra). В настоящем изобретении также представлены композиции, включающие варианты аспарагиназы по изобретению. Композиции могут, необязательно, включать и другие ингредиенты, например другие ферменты. Варианты аспарагиназы по изобретению или композиции, содержащие данные аспарагиназы,могут применяться при получении пищевых продуктов. В одном воплощении изобретения варианты аспарагиназы по изобретению или композиции, содержащие данные аспарагиназы, могут применяться для снижения количества акриламида, образующегося при термической обработке пищевых продуктов на основе содержащих аспарагин исходных материалов. Например, они могут применяться в способе получения пищевых продуктов, включающем по меньшей мере одну стадию нагревания, который включает добавление одного или нескольких ферментов аспарагиназы в промежуточную форму данного пищевого продукта в данном способе получения, при этом фермент добавляется перед стадией нагревания в количестве, эффективно снижающем уровень аспарагина в данной промежуточной форме пищевого продукта. Такой способ раскрыт в WO 04/030468, причем сам способ и все его преимущества включены в настоящее изобретение путем ссылки. Также и в WO 04/026043 описаны подходящие способы, в которых может применяться аспарагиназа по изобретению. Раскрытые в WO 04/026043 способы и все их преимущества включены в настоящее изобретение путем ссылки. Промежуточная форма пищевого продукта определяется в настоящем изобретении как любая форма, возникающая в процессе получения до получения конечной формы пищевого продукта. Промежуточная форма может включать индивидуальные исходные материалы и/или их смеси и/или смеси с добавками и/или технологическими добавками либо их подвергнутые последующей обработке формы. Например, промежуточные формы пищевого продукта - хлеба включают, к примеру, пшеницу, пшеничную муку, исходную смесь е с другими ингредиентами хлеба, такими, к примеру, как вода, соль, дрожжи и композиции для улучшения хлеба, замешанное тесто, вымешенное тесто, поднявшееся тесто и частично испеченное тесто. Например, для некоторых продуктов из картофеля промежуточными продуктами являются обезвоженные картофельные хлопья или гранулы, а промежуточным продуктом для кукурузных чипсов является кукурузная масса. Пищевой продукт может быть изготовлен по меньшей мере из одного исходного материала, имеющего растительное происхождение, например картофеля, табака, кофе, какао, риса, зерновых, к примеру пшеницы, ржи, кукурузы, ячменя, крупы, гречихи и овса. Здесь и далее пшеница означает все известные виды из рода Triticum, например Т.aestivum, durum и/или spelta. Также в рамки изобретения входят пищевые продукты, изготовленные более чем из одного исходного или промежуточного материала, например пищевые продукты, содержащие и пшеницу (муку и/или крахмал), и картофель. Примерами пищевых продуктов, для которых может подойти способ по изобретению, являются мучные продукты, такие как хлеб, выпечка, пирожные, крендели, бублики, медовые торты, печенье, пряники, имбирное печенье и хлебцы, и картофельные продукты, такие как картофель фри, жареный картофель, картофельные чипсы, крокеты. Вышеприведенные исходные материалы, как известно, содержат значительное количество аспарагина, который в процессе производства участвует в образовании акриламида на стадии нагревания. С другой стороны, аспарагин может происходить из других источников, чем исходные материалы, например из гидролизатов белков, таких как дрожжевые экстракты, гидролизат сои, гидролизат казеина и др.,которые применяются как добавки в процессе производства пищевых продуктов. Предпочтительным производственным процессом является выпекание хлеба и других хлебобулочных изделий из пшеничной муки и/или муки из других злаков. Другим предпочтительным производственным процессом является обжаривание во фритюре картофельных чипсов из ломтиков картофеля. Предпочтительными стадиями нагревания являются те, при которых по меньшей мере часть промежуточного пищевого продукта, например поверхность пищевого продукта, подвергается воздействию температур, при которых происходит образование акриламида, например, 110C или выше, 120C или выше. Стадия нагревания в способе по изобретению может проводиться в печи, например, при температуре 180-220C, как при выпекании хлеба и других хлебобулочных изделий, либо в масле, как при обжаривании картофельных чипсов, например, при 160-190 С. В следующем аспекте изобретения предусмотрены пищевые продукты, получаемые способом изобретения, как описано выше, или с применением новых аспарагиназ, как описано выше, для получения пищевых продуктов. Эти пищевые продукты отличаются значительным снижением уровня акриламида по сравнению с продуктами, получаемыми такими способами, которые не включают добавления одного или нескольких ферментов в количестве, эффективно снижающем уровень аминокислот, участвующих в образовании акриламида на данной стадии нагревания. Способ по изобретению может применяться для снижения содержания акриламида в приготовленном пищевом продукте предпочтительно до 50%, более предпочтительно до 20%, еще более предпочтительно до 10% и наиболее предпочтительно до 5% по сравнению с пищевым продуктом, полученным традиционным способом. Другим применением для вариантов аспарагиназы по изобретению является их применение при лечении опухолей. Метаболизм раковых клеток требует L-аспарагина, который может быть быстро разрушен аспарагиназами. Аспарагиназа по изобретению также может применяться как вспомогательное средство при лечении некоторых разновидностей лейкемии у человека. Применение аспарагиназы на экспериментальных животных и людях приводит к регрессии некоторых лимфом и лейкемии. Поэтому в одном воплощении изобретение касается аспарагиназ или композиций по изобретению для применения в качестве медикамента, например, при лечении опухолей, например при лечении лимфомы или лейкемии у животных или человека. Варианты аспарагиназы по изобретению можно удобно продуцировать в микроорганизмах. В вышеописанных способах предпочтительно применяются аспарагиназы, полученные методами рекомбинантной ДНК. Такие рекомбинантные ферменты обладают рядом преимуществ: продукция при низкой себестоимости, с высоким выходом, отсутствие таких загрязнений, как бактерии или вирусы, а также отсутствие бактериальных токсинов или примесей активности других ферментов. Далее изобретение раскрывается на следующих неограничивающих примерах. Примеры Материалы и методы. Определение аспарагиназы для измерения pH-зависимости в диапазоне pH 4-9. Аспарагиназную активность определяли, используя в качестве субстрата L-аспарагин. Количество выделяющегося под действием фермента аммиака измеряли согласно реакции Berthelot. Готовые к употреблению реагенты фенол нитропруссид и щелочной гипохлорит получали от фирмы Sigma. Образец фермента в 100 мкл смешивали с 2000 мкл 100 мМ L-аспарагина в буфере из смеси 50 мМ лимонной кислоты и 50 мМ пирофосфата натрия с требуемым значением pH. После инкубации при 37C в течение 30 мин реакцию останавливали добавлением 400 мкл 25% трихлоруксусной кислоты, после чего добавляли 2500 мкл воды. Во время инкубации температуру поддерживали при 37C, если не указано иначе. Специалистам должно быть понятно, что дозировку фермента выбирали таким образом, чтобы после инкубации при вышеуказанных условиях полученный сигнал был значительно выше уровня фона, но попадал в такой диапазоне, в котором полученные сигналы пропорциональны количеству добавленного фермента. Предпочтительно реакция была нулевого порядка. После остановки реакции 4 мкл инкубационной смеси вносили в 156 мкл воды. После этого добавляли 34 мкл раствора фенола/нитропруссида (Sigma P6994) и 34 мкл щелочного раствора гипохлорита(Sigma A1727). После инкубации в течение 676 с при 37C измеряли экстинкцию при 600 нм. Показания подвергали коррекции на фоновый сигнал по соответствующим холостым пробам. В качестве холостой пробы использовали образец с инактивированным ферментом (TXY). Определения проводили на автоматическом анализаторе Konelab Arena 30 (Thermo Scientific). Активность определяли по калибровочной прямой, которую составляли, нанося на график значения поглощения при 600 нм в зависимости от известных концентраций сульфата аммония из стандартного ряда. Активность выражали в единицах, причем 1 единица определялась как количество фермента, необходимое для выделения 1 мкмоль аммиака изL-аспарагина за 1 мин при заданных условиях определения. Определение аспарагиназы для измерения pH-зависимости в диапазоне pH 4-8. Выполнялась такая же методика, как описанная выше методика для измерения pH-зависимости активности в диапазоне pH 4-9 с тем отличием, что образец фермента в 100 мкл смешивали с 2000 мкл 100 мМ L-аспарагина в 50 мМ фосфатно-цитратном буфере с требуемым значением pH. Определение аспарагиназы вручную для измерения активности при pH 5 и pH 7. Определение проводили в микропланшетах или пробирках. Для идентификации аспарагиназы со смещенным pH-профилем активности измеряли активность при pH 5 и 7. Образец фермента в 10 мкл смешивали со 190 мкл 100 мМ L-аспарагина в 100 мМ цитратном буфере pH 5 или 100 мМ фосфатном буфере pH 7. После инкубации при комнатной температуре в течение 1 ч реакцию останавливали добавлением 100 мкл 12,5% трихлоруксусной кислоты. Дозировку фермента выбирали таким образом, чтобы после инкубации 1 ч при комнатной температуре полученный сигнал был значительно выше уровня фона. После остановки реакции к 8 мкл инкубационной смеси добавляли 95 мкл воды. Затем добавляли 70 мкл раствора фенола/нитропруссида (Sigma P6994) и 70 мкл щелочного раствора гипохлорита (SigmaA1727). После инкубации в течение 60 мин при комнатной температуре измеряли экстинкцию при 620 нм. Показания подвергали коррекции на фоновый сигнал по соответствующим холостым пробам,например инактивированным образцам и/или супернатантам из образцов ферментации пустых штаммов хозяина. Пустой штамм означает такой штамм хозяина, который не был трансформирован и поэтому не содержит гена аспарагиназы. Активность определяли по калибровочной прямой, которую составляли,нанося на график значения поглощения при 620 нм в зависимости от известных концентраций сульфата аммония из стандартного ряда. Активность выражали в единицах, причем 1 единица определялась как количество фермента, необходимое для выделения 1 мкмоль аммиака из L-аспарагина за 1 мин при заданных условиях определения. При всех определениях активность аспарагиназы в образцах выражали в единицах на 1 мл. Пример 1. Ферментация, выделение и очистка аспарагиназ по изобретению. Аспарагиназы по изобретению получали путем конструирования экспрессионных плазмид, содержащих последовательность ДНК, кодирующую аспарагиназу по изобретению, трансформации этой плазмидой штамма Aspergillus niger и культивирования штаммов Aspergillus niger, как описано вWO 2004/030468. После культивирования клеток Aspergillus niger, содержащих соответствующие экспрессионные плазмиды, получали бесклеточные супернатанты путем центрифугирования ферментационной жидкости при 5000g в течение 30 мин при 4C. При необходимости супернатанты дополнительно фильтровали через фильтр Miracloth (Calbiochem, кат.475855) и микроволоконный фильтр GF/A Glass фирмыWhatmann (150 мм ) соответственно для удаления любых частиц. Чтобы избавиться от грибков, супернатанты доводили до pH 5 с помощью 4 н. KOH и стерилизировали фильтрованием через фильтр с диаметром пор 2 мкм (бутылочный) с отсасыванием. Супернатанты хранили до использования при 4C или замораживали при -20C, если нужно. В том случае, когда примеси составляли более 60 мас.%, аспарагиназы очищали методом анионообменной хроматографии, исходя из бесклеточных супернатантов или ccUF, обессоленных пропусканием через колонку PD-10 (Amersham Biosciences). Обессоленный материал наносили на колонку с сефарозой Mono-Q или Q-сефарозой, уравновешенной в 20 мМ гистидиновом буфере pH 5,96. После исчерпывающей промывки аспарагиназы элюировали из колонки с помощью градиента от 0 до 1 М NaCl. Чистоту фракций супернатанта, содержащих аспарагиназную активность, или очищенных фракций аспарагиназы (при определении в мг белка/мл) проверяли методом аналитической эксклюзионной хроматографии (HP-SEC: высокоэффективная эксклюзионная хроматография, TSKgel 3000SW-XL, колонка 3007,8 мм; диапазон мол. весов 10-300 кДа, 100 мМ фосфатный буфер pH 7 и 5,96). Скорость потока составляла 1 мл/мин (за исключением инжекции проб на колонке с Q-сефарозой, которое проводилось при 5 мл/мин). Детектирование элюированного белка проводили при 280 нм. Концентрацию элюированной аспарагиназы дикого типа Aspergillus niger рассчитывали по экстинкции при 280 нм (А 280), используя молярный коэффициент экстинкции 10240 М-1 см-1 (2801 см, 1 мг/мл = 0,28, где 2801 см, 1 мг/мл означает экстинкцию при 280 нм, измеренную при длине пути в 1 см и при концентрации очищенного белка в 1 мг/мл). Измерение А 280 проводили на спектрофотометре Uvikon XL Secomam (Beun de Ronde,Abcoude, Нидерланды). Для аспарагиназ, соответствующих ASN15 и ASN16, использовали тот же самый коэффициент экстинкции, что и для аспарагиназы дикого типа Aspergillus niger. При наличии примесей,поглощающих при 280 нм, концентрацию аспарагиназ подвергали коррекции по хроматограмме HP-SEC,умножая значение А 280 в образце аспарагиназы на отношение площади под пиком аспарагиназы к общей площади пиков, поглощающих при 280 нм. Если пики аспарагиназ не были четко отделены от других пиков, то вместо площади пика использовали его высоту. Для аспарагиназ, соответствующих ASN01-ASN14, содержание аспарагиназ (в мг белка/мл) можно определить методом гель-электрофореза в DDCNa-ПААГ, используя 12-луночные гели NuPAGE Novex 4-12% Bis-Tris (Invitrogen, NP0322BOX). Образец в 1 мкл супернатанта культуры инкубировали с 1 мкл восстанавливающего реагента 10NuPAGE (Invitrogen, NP0004), 2,5 мкл буфера для образцов 4NuPAGE LDS (Invitrogen, NP0007) и 5,5 мкл воды milliQ в течение 10 мин при 70C. Полученный восстановленный образец наносили на гель. В качестве маркера размера использовали готовый окрашенный стандарт SeeBlue Plus2 (Invitrogen, LC5925). Кроме того, наносили 0,5 мкг БСА (Sigma А 9418) для калибровки количества белка. Гели разгоняли в рабочем буфере NuPAGE MES SDS (Invitrogen, NP0002),содержащем антиоксидант NuPAGE (Invitrogen, NP0005), в течение 35 мин при 200 В. После электрофореза гели фиксировали 230 мин в растворе Fix (7% НАс (об./об.) и 10% этанола (об./об., окрашивали в течение ночи красителем на белок в геле SYPRO Ruby (Invitrogen S12000) и отмывали от красителя 230 мин в растворе Fix. После этого гели промывали деминерализованной водой и сканировали на сканере Typhoon 9200 (GE Healthcare). Вычисляли площади пиков с помощью программы Image QuantTLv0203.02 и рассчитывали концентрацию белка, исходя из полосы белка БСА. Пример 2. Функционирование вариантов аспарагиназы по изобретению. Библиотеку случайных мутантов аспарагиназы A.niger (у которой исходный полипептид соответствовал SEQ ID NO: 3) подвергали скринингу на мутантов с измененным pH-профилем активности. Для того чтобы обнаружить мутантов с лучшей активностью при более щелочных значениях pH, определяли активность аспарагиназы при pH 5 и 7. После этого определяли соотношение между активностью при pH 7 и активностью при pH 5. Эти соотношения представлены в табл. 2. Более высокое соотношение указывает на смещение pH-профиля активности в сторону pH 7. Табл. 2. Третий столбец: соотношение между активностью при pH 7 и активностью при pH 5 у выбранных мутантов. Дикий тип (wt) означает аспарагиназу А.niger (WO 2004/030468). Четвертый столбец: смещение щелочного сегмента pH-профиля активности представлено значением pH, при котором мутант проявляет 50% от своей максимальной каталитической активности. pH-профили активности определяли при 37C, используя бесклеточные супернатанты. Активность измеряли в диапазоне от pH 4 до pH 8, ис- 19018424 Мутантов с более высоким соотношением, чем у аспарагиназы дикого типа А.niger, подвергали дополнительному тестированию, чтобы установить, на какой процент pH-профиль активности сместился в сторону щелочных значений pH. У этих мутантов определяли pH-профиль активности, при этом оказалось, что в особенности щелочной сегмент pH-профиля активности смещался в сторону более высоких значений pH. Значение pH, при котором щелочной сегмент pH-профиля активности проявлял 50% от максимальной активности мутанта в его pH-оптимуме, принимается как свидетельство смещения щелочного сегмента pH-профиля активности по сравнению с диким типом (табл. 2). Смещение в сторону более высоких значений pH означает более высокую активность при более щелочных условиях. Такие мутанты особенно выгодны в тех применениях, которые требуют более щелочных условий. При отборе мутантов с более низким соотношением между активностью при pH 7 и активностью при pH 5 оказалось, что щелочной сегмент pH-профиля активности смещается в сторону более низких значений pH (табл. 3). Табл. 3. Соотношение между активностью при pH 7 и активностью при pH 5 у выбранных мутантов. Дикий тип (wt) означает аспарагиназу A. niger (WO 2004/030468). pH-профили активности определяли при 37C, используя бесклеточные супернатанты. Активность измеряли в диапазоне от pH 4 до pH 8,используя систему фосфатно-цитратных буферов. Таблица 3pH-зависимость активности и pH-оптимум. Определяли pH-зависимость аспарагиназной активности у мутантов в 50 мМ фосфатно-цитратном буфере в диапазоне pH от 4 до 8, используя бесклеточные супернатанты. Значение pH, при котором у мутанта наблюдалась самая высокая активность, именуется pH-оптимумом для данного мутанта. В табл. 4 и 5 за 100% принимали максимальную активность, наблюдавшуюся у данного мутанта, а активности при других значениях pH приведены в процентах от максимальной активности, наблюдавшейся у данного мутанта. В табл. 4 pH-профиль активности определяли в диапазоне от pH 4 до pH 8, используя систему фосфатно-цитратных буферов. В табл. 5 pH-профиль активности определяли в диапазоне от pH 4 до pH 9, используя систему пирофосфатно-цитратных буферов. Табл. 4. Зависимость аспарагиназной активности от pH у мутантов в сравнении с аспарагиназой дикого типа (wt) A.niger (WO 2004/030468). За 100% принимали самую высокую активность, наблюдавшуюся у каждой аспарагиназы. Активность определяли, используя бесклеточные супернатанты при 37 С в системе фосфатно-цитратных буферов. Таблица 4 Табл. 5. Зависимость аспарагиназной активности от pH у мутантов в сравнении с аспарагиназой дикого типа (wt) A.niger (WO 2004/030468). За 100% принимали самую высокую активность, наблюдавшуюся у каждой аспарагиназы. Активность определяли на бесклеточных супернатантах при 37 С в системе пирофосфатно-цитратных буферов. Таблица 5 Помимо смещения щелочного сегмента pH-профиля активности, также имеется сдвиг pH-оптимума в сторону более высоких значений pH. Оба мутанта, содержащие мутацию D63G, проявляют сдвиг pHоптимума к pH 7. Мутанты D63G содержат и другие мутации. Однако эти другие мутации у каждого мутанта отличаются, тогда как pH-профили активности почти одинаковы. Следовательно, именно D63G вызывает наблюдаемое смещение pH-профиля активности. У мутанта D140N, содержащего мутацию А 170 Т, и мутанта, содержащего мутацию S66P, pH-оптимум сдвигается к pH 6. Остальные мутанты проявляют более явный pH-оптимум при pH 5 по сравнению с диким типом. Из табл. 4 и 5 четко видно смещение щелочного сегмента pH-профиля активности в сторону более высоких значений pH, при этом особенно повышается относительная активность в диапазоне от pH 6 до pH 8, а в тоже время сохраняется и значительная активность в кислой области от pH 4 до pH 6. Удельная активность в зависимости от pH. Определяли удельную активность вариантов аспарагиназы при pH 4, pH 5, pH 6, pH 7, pH 8 при 37 С в 50 мМ фосфатно-цитратном буфере, используя бесклеточные супернатанты. Табл. 6. Удельная активность (измеряли путем деления активности образца в единицах на 1 мл на количество аспарагиназы в образце в мг/мл) у вариантов в сравнении с аспарагиназой дикого типа (wt)A.niger (WO 2004/030468) при указанных значениях pH с использованием аспарагина в качестве субстрата. За 100% при каждом pH принимали удельную активность дикого типа, а активность у мутантов рассчитывали относительно аспарагиназы дикого типа. Если активность у мутанта составляла меньше 100%,то е не включали в таблицу. Активность определяли при 37C. Содержание белка аспарагиназы в бесклеточных супернатантах определяли в опытах с проведением электрофореза в гелях PAA-SDS и сканированием гелей, как описано в разделе "Материалы и методы". Для мутантов T73K+S74A+A293S иT73K+S74A+A293S+E106P+G297S+T299S+Q319A+M321T+V324G концентрацию белка аспарагиназы получали из измерений А 280, применяя коррекцию на загрязнения по данным хроматографии HP-SEC. Из табл. 6 видно, что удельная активность у мутантов при pH 6, 7 и 8 значительно улучшилась по сравнению с аспарагиназой дикого типа. В частности, мутанты T73K+S74A+A293S, T41I+S66P+V371M,S88P+I161L+R262C и F53Y+K119N очень полезны, так как они проявляют более высокую активность по всему диапазону от pH 4 до pH 8. Мутант T73K+S74A+A293S+E106P+G297S+T299S+Q319A+M321T+V324G более активен в кислой области от pH 4 до pH 6. Температурный оптимум. Для того чтобы проверить зависимость активности от температуры, измеряли активность при различных температурах. В одном опыте реакцию фермента останавливали через 10 мин, а в другом опыте реакцию останавливали через 30 мин. Дозировка фермента в 30-минутном опыте составляла одну треть от дозировки в 10-минутном опыте. Если фермент устойчив в применявшихся условиях, то должна наблюдаться одинаковая активность. Если же происходит инактивация, то активность должна уменьшаться при более продолжительном определении. Результаты представлены в табл. 7. Табл. 7. Зависимость активности от температуры. Опыты проводились при pH 5 в 50 мМ фосфатноцитратном буфере. Дозировка фермента в 30-минутном опыте составляла одну треть от дозировки в 10 минутном опыте. За 100% принимали самую высокую активность при 10-минутной продолжительности инкубации. Таблица 7 Из табл. 7 видно, что устойчивость вариантов очень близка к аспарагиназе дикого типа Aspergillusniger. Мутанты не проявляют снижения активности после 30-минутной инкубации по сравнению с 30 минутной даже при 70C, что свидетельствует о том, что мутанты устойчивы по меньшей мере 30 мин при 70C. Неожиданно оказалось, что температурный оптимум у мутантов смещается к более высокой температуре. У дикого типа температурный оптимум приходится на 50C с учетом тех температур, при которых проводилось тестирование. У мутантов S16A+D63G+G132S, D63G+D111G+R122H иT41I+S66P+V371M он сместился к 70C. Такие свойства особенно полезны в тех применениях, в которых нужно, чтобы аспарагиназы работали при повышенных температурах. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Вариант исходного полипептида, обладающего аспарагиназной активностью, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 63, 64, 66, 73, 74, 76, 77, 88, 90, 91, 101, 106, 111, 119, 122, 123,132, 140, 161, 170, 195, 228, 262, 293, 297, 299, 300, 310, 314, 319, 321, 324 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид по меньшей мере на 80% гомологичен SEQ ID NO: 3, где вариант обладает удельной активностью, которая выше, чем у исходного полипептида при измерении при одном и том же значении pH, по крайней мере при одном значении pH. 2. Вариант по п.1, у которого исходный полипептид имеет последовательность, приведенную в SEQID NO: 3. 3. Вариант по любому из пп.1, 2, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит одну или более замен из числа Ile в положении 41, Tyr в положении 53, Gly, или Val, или Ser в положении 63, Pro в положении 64, Pro в положении 66, Lys в положении 73, Ala в положении 74, Thr в положении 76, Ile в положении 77, Tyr или Pro в положении 88, Val в положении 90, Glu в положении 91, Val в положении 101,Pro в положении 106, Gly в положении 111, Asn в положении 119, His в положении 122, Ala в положении 123, Ser в положении 132, Asn в положении 140, Leu в положении 161, Thr в положении 170, Asp в положении 195, His в положении 228, Cys или His в положении 262, Ser или Val в положении 293, Ser в положении 297, Ser в положении 299, Ile в положении 300, Val в положении 310, Asp в положении 314, Ala в положении 319, Thr в положении 321, Gly в положении 324 или Met в положении 371, причем данные положения определяются по отношению к SEQ ID NO: 3. 4. Вариант по любому из пп.1-3, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит по меньшей мере одну замену аминокислотного остатка, соответствующего любой из аминокислот: 53, 63, 64, 66, 73, 74,76, 77, 88, 101, 140, 170, 293, причем данные положения определяются по отношению к SEQ ID NO: 3. 5. Вариант по любому из пп.1-4, причем вариант имеет аминокислотную последовательность, которая при выравнивании с последовательностью, приведенной в SEQ ID NO: 3, содержит одну или более замен из числа Tyr в положении 53, Gly или Val в положении 63, Pro в положении 64, Pro в положении 66, Lys в положении 73, Ala в положении 74, Thr в положении 76, Ile в положении 77, Tyr или Pro в положении 88, Val в положении 101, Asn в положении 140, Thr в положении 170, Ser или Val в положении 293, причем данные положения определяются по отношению к SEQ ID NO: 3. 6. Вариант по любому из пп.1-5, причем вариант имеет аминокислотную последовательность согласно любой из SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19. 7. Вариант по любому из пп.1-6, который по меньшей мере на 95% гомологичен исходному полипептиду аспарагиназы. 8. Вариант по любому из пп.1-7, который имеет более высокую удельную активность, чем у исходного полипептида, измеренную при одном и том же значении pH, по меньшей мере при одном значенииpH от 4 до 8. 9. Вариант по любому из пп.1-8, который имеет более высокий pH-оптимум, чем у исходного полипептида. 10. Вариант по любому из пп.1-9, который дополнительно содержит одну или более замен аминокислотных остатков, помимо тех, что определены в п.1. 11. Вариант по п.10, который содержит от 1 до 5 дополнительных замен, помимо тех, что определены в п.1. 12. Последовательность нуклеиновой кислоты, кодирующая вариант по любому из пп.1-11. 13. Конструкция нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты по п.12, функционально соединенную с одной или несколькими контрольными последовательностями, способными управлять экспрессией аспарагиназы в подходящем экспрессионном хозяине. 14. Рекомбинантный экспрессионный вектор, содержащий конструкцию нуклеиновой кислоты по п.13. 15. Рекомбинантная клетка-хозяин, содержащая экспрессионный вектор по п.14. 16. Способ получения аспарагиназы, включающий культивирование клетки-хозяина по п.15 в условиях, способствующих продукции аспарагиназы, и выделение аспарагиназы. 17. Способ получения варианта полипептида аспарагиназы, который включает:a) выбор исходного полипептида аспарагиназы;b) замену по меньшей мере одного аминокислотного остатка, соответствующего любой из аминокислот: 41, 53, 63, 64, 66, 73, 74, 76, 77, 88, 90, 91, 101, 106, 111, 119, 122, 123, 132, 140, 161, 170, 195, 228,- 23018424 262, 293, 297, 299, 300, 310, 314, 321, 324 и 371, причем данные положения определяются по отношению к SEQ ID NO: 3, а исходный полипептид по меньшей мере на 80% гомологичен SEQ ID NO: 3;c) необязательно, замену одной или нескольких дополнительных аминокислот, приведенных в b);d) получение варианта, являющего результатом стадий а)-с);e) определение удельной активности по меньшей мере при одном значении pH и/или pH-оптимума этого варианта иf) отбор варианта, обладающего повышенной удельной активностью по меньшей мере при одном значении pH по сравнению с исходным полипептидом аспарагиназы и/или более высоким pH-оптимумом по сравнению с исходным полипептидом аспарагиназы, получая посредством этого вариант полипептида аспарагиназы. 18. Способ по п.17, в котором исходный полипептид аспарагиназы имеет последовательность, приведенную в SEQ ID NO: 3. 19. Способ по п.18, в котором на стадии b) заменяемые аминокислотные остатки соответствуют одной или нескольким аминокислотам из числа Ile в положении 41, Tyr в положении 53, Gly, или Val, илиVal в положении 293, Ser в положении 297, Ser в положении 299, Ile в положении 300, Val в положении 310, Asp в положении 314, Ala в положении 319, Thr в положении 321, Gly в положении 324 или Met в положении 371, причем данные положения определяются по отношению к SEQ ID NO: 3. 20. Композиция, обладающая аспарагиназной активностью, содержащая эффективное количество аспарагиназы, являющейся вариантом по любому из пп.1-11 или вариантом, полученным способом по любому из пп.17-19, и другой ингредиент. 21. Применение аспарагиназы по любому из пп.1-11 или композиции по п.20 для получения пищевого продукта. 22. Применение аспарагиназы по любому из пп.1-11 или композиции по п.20 для снижения количества акриламида, образующегося в термически обработанном пищевом продукте на основе содержащего аспарагин исходного материала. 23. Способ обработки пищевого продукта для уменьшения в нем количества аспарагина, включающий по меньшей мере одну стадию нагревания, который включает добавление одного или нескольких ферментов аспарагиназы по любому из пп.1-11 или композиции по п.20 в промежуточную форму указанного пищевого продукта, при этом фермент добавляют перед указанной стадией нагревания в количестве, эффективном для снижения уровня аспарагина, содержащегося в указанной промежуточной форме пищевого продукта. 24. Применение аспарагиназы по любому из пп.1-11 или композиции по п.20 для получения медикамента для лечения опухолей.
МПК / Метки
МПК: C12N 9/82, A23L 1/03, A23L 1/30, C07K 14/38
Метки: фермента, варианты, аспарагиназы, применение
Код ссылки
<a href="https://eas.patents.su/30-18424-varianty-fermenta-asparaginazy-i-ih-primenenie.html" rel="bookmark" title="База патентов Евразийского Союза">Варианты фермента аспарагиназы и их применение</a>
Новые аспарагиназы и их применение

Номер патента: 17305
Опубликовано: 30.11.2012
Авторы: Стор Марк Кристиаан, Морман Лисетте, Лаан Ван Дер Ян Метске, Ланге Де Илсе
МПК: A23L 1/03, A23L 1/30, C07K 14/38...
Метки: аспарагиназы, применение, новые
Формула / Реферат:
1. Аспарагиназа, которая представляет собой:a) полипептид, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 2 или SEQ ID NO: 4; илиb) полипептид, который по меньшей мере на 85% гомологичен полипептиду по п.а); илиc) полипептид, который кодируется последовательностью нуклеиновой кислоты по SEQ ID NO: 1 или SEQ ID NO: 3, кодирующей зрелый полипептид.2. Аспарагиназа по п.1, у которой в отношении SEQ ID NO: 2 полипептид по пп.b),...
Применение комбинации ингибитора конвертирующего фермента и диуретика для лечения нарушений микроциркуляции

Номер патента: 2968
Опубликовано: 26.12.2002
Авторы: Шиави Пьер, Леви Бернар, Гез Давид
МПК: A61K 38/55, A61P 9/00
Метки: фермента, комбинации, диуретика, микроциркуляции, нарушений, лечения, конвертирующего, применение, ингибитора
Формула / Реферат:
1. Применение комбинации ингибитора ангиотензин-конвертирующего фермента и диуретика для получения фармацевтических композиций, предназначенных для лечения нарушений артериоло-капиллярной микроциркуляции. 2. Применение по п.1, отличающееся тем, что ингибитор ангиотензин-конвертирующего фермента представлен периндоприлом, каптоприлом, эналаприлом, лизиноприлом, делаприлом, фозиноприлом, квинаприлом, рамиприлом, спираприлом, имидаприлом,...
Стабильная местная и/или мукозальная эмульсионная композиция (варианты), способ ее приготовления (варианты) и ее применение (варианты)

Номер патента: 17434
Опубликовано: 28.12.2012
Автор: Йорсаль Стен Бойе
МПК: A61K 47/30, A61K 9/107, A61K 8/06...
Метки: мукозальная, композиция, стабильная, эмульсионная, способ, варианты, приготовления, местная, применение
Формула / Реферат:
1. Стабильная местная и/или мукозальная эмульсионная композиция для фармацевтического и/или косметического применения, включающая водный компонент, неионный термообратимый блок-сополимер этиленоксида и пропиленоксида (полоксамер), масло чайного дерева и гидрофильный неионный эмульгатор на основе жирной кислоты, в которой сополимер, смешанный с водным компонентом, представляет собой текучую жидкость при температуре ниже 15°С, масло чайного дерева...
Замещенные амины (варианты), способ их получения и их применение в качестве ингибиторов транспорта глицина (варианты), композиция (варианты), промежуточный иодид и способ его получения, способ леченияпациента (варианты)

Номер патента: 6636
Опубликовано: 24.02.2006
Авторы: Дилейни Уильям, Шумахер Ричард, Мэддейфорд Шон, Техим Ашок, Ван Чжаоцин, Игл Иэн, Хоппер Аллан Т.
МПК: A61P 25/28, A61K 31/381, C07D 333/20...
Метки: промежуточный, глицина, замещенные, ингибиторов, применение, леченияпациента, композиция, иодид, варианты, способ, получения, транспорта, качестве, амины
Формула / Реферат:
1. Замещенные амины общей формулы I где Ar1 означает тиофеновую группу, выбранную из остатков 2- или 3-тиофена, необязательно содержащую один заместитель, выбранный из метила или этила, и Ar2 выбран из группы, содержащей тиофен, фуран и замещенный фенил, причем заместитель фенильной группы выбран из ряда C1-6алкил, галоген, C1-6галогеналкил, C1-6алкокси, C1-6галогеналкокси, циано; и его соли, сольваты и гидраты, или Ar1 означает 2-метилфенил, а...
Применение 4-хлор-5-{2-[4-(6-фтор-1,2-бенз[d]изоксазол-3-ил)пиперидин-1-ил]этиламино}-2-метил-3-(2h) пиридазинона или его фармацевтически приемлемых солей для нейропротекции (варианты), лекарственный препарат (варианты) и способ лечения (варианты)

Номер патента: 16142
Опубликовано: 28.02.2012
Авторы: Баркоци Йожеф, Харшинг Ласло Габор, Мориц Кристина, Кертес Саболч, Гачайи Иштван, Шимиг Дьюла, Сенаши Габор, Компагне Хайналка, Гиглер Габор, Леваи Дьёрдь, Агоштон Марта
МПК: A61P 25/28, A61K 31/501
Метки: применение, 4-хлор-5-{2-[4-(6-фтор-1,2-бенз[d]изоксазол-3-ил)пиперидин-1-ил]этиламино}-2-метил-3-(2h, лекарственный, солей, нейропротекции, препарат, лечения, пиридазинона, приемлемых, варианты, фармацевтически, способ
Формула / Реферат:
1. Применение 4-хлор-5-{2-[4-(6-фтор-1,2-бенз[d]изоксазол-3-ил)пиперидин-1-ил]этиламино}-2-метил-3-(2H)пиридазинона формулы (I)или его фармацевтически приемлемой соли для изготовления нейропротективных лекарственных препаратов.2. Применение 4-хлор-5-{2-[4-(6-фтор-1,2-бенз[d]изоксазол-3-ил)пиперидин-1-ил]этиламино}-2-метил-3-(2H) пиридазинона формулы (I) или его фармацевтически приемлемой соли для изготовления лекарственного препарата, пригодного...
Предыдущий патент: Способ определения пространственного распределения свойств геологической среды
Следующий патент: Фармацевтический, дезинфицирующий или консервирующий экстракт пажитника сенного (trigonella foenum-graecum), способ его получения и применение экстракта
Случайный патент: Селективные модуляторы рецептора, активируемого пролифератором пероксисом