Модифицированные экзосомы и их применение
Номер патента: 4428
Опубликовано: 29.04.2004
Авторы: Рапосо Граса, Штумптнер Памела, Бенарош Филипп, Боннеро Кристиан, Амигорена Себастьян, Венсан-Шнейдер Элен

























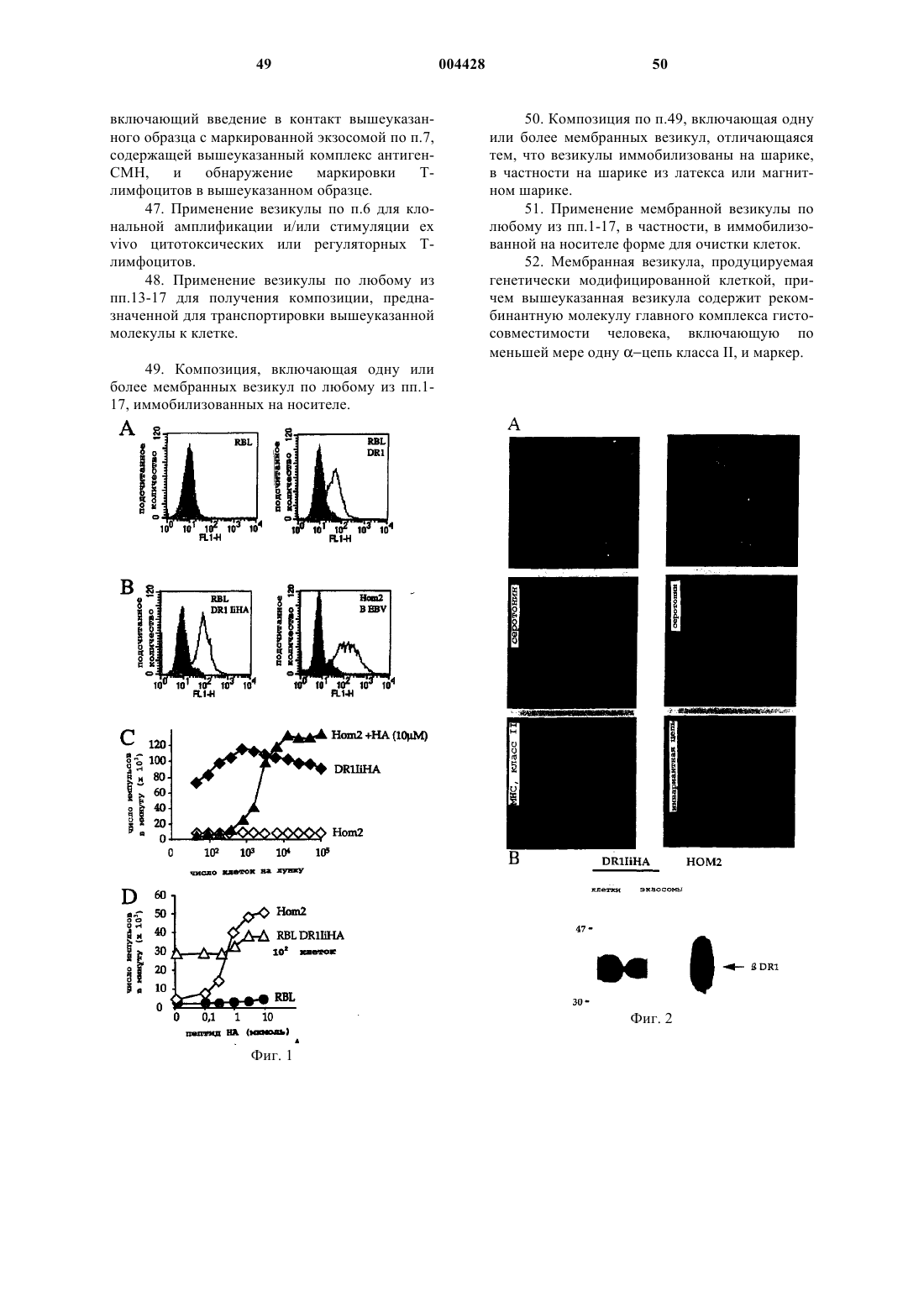
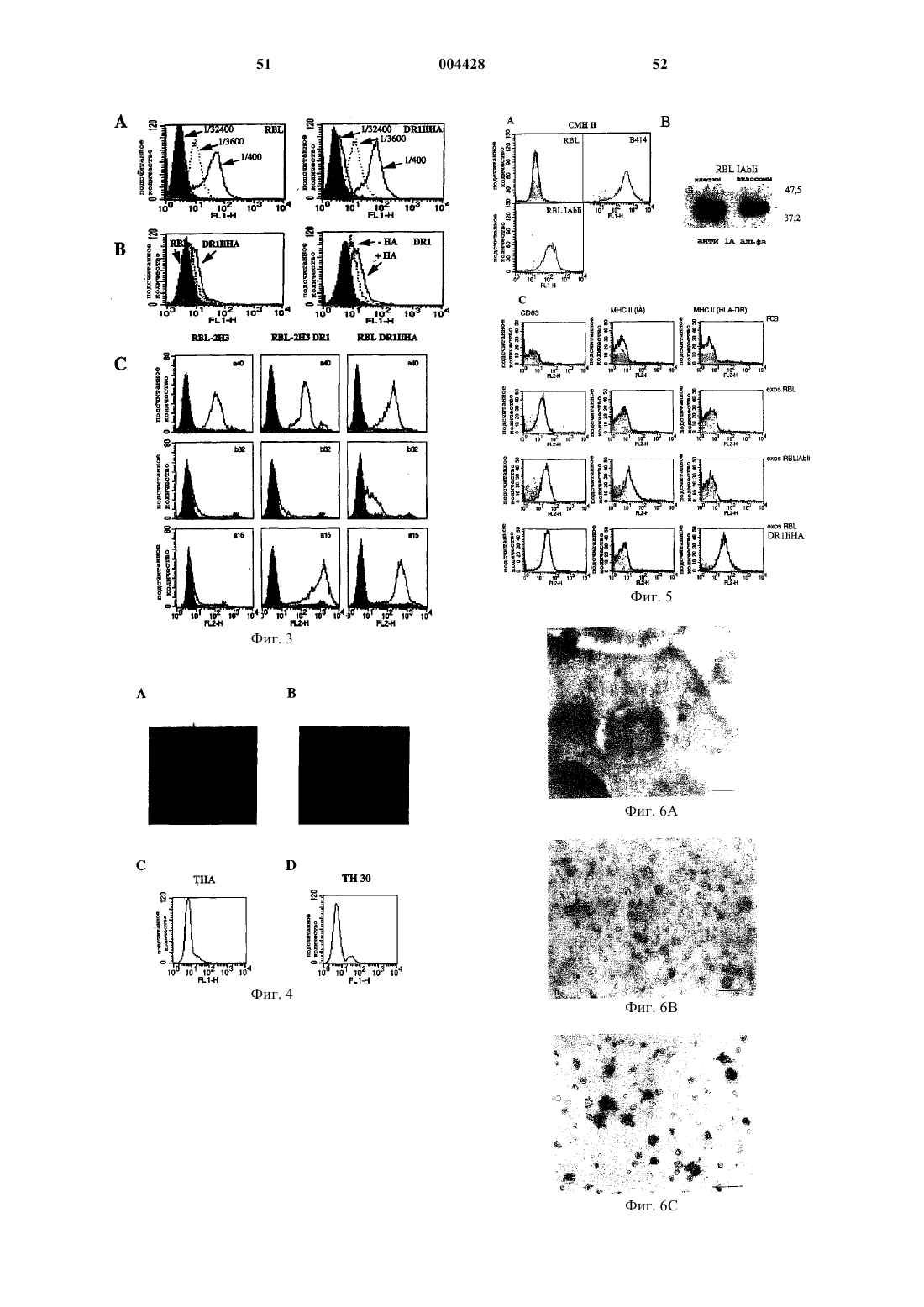
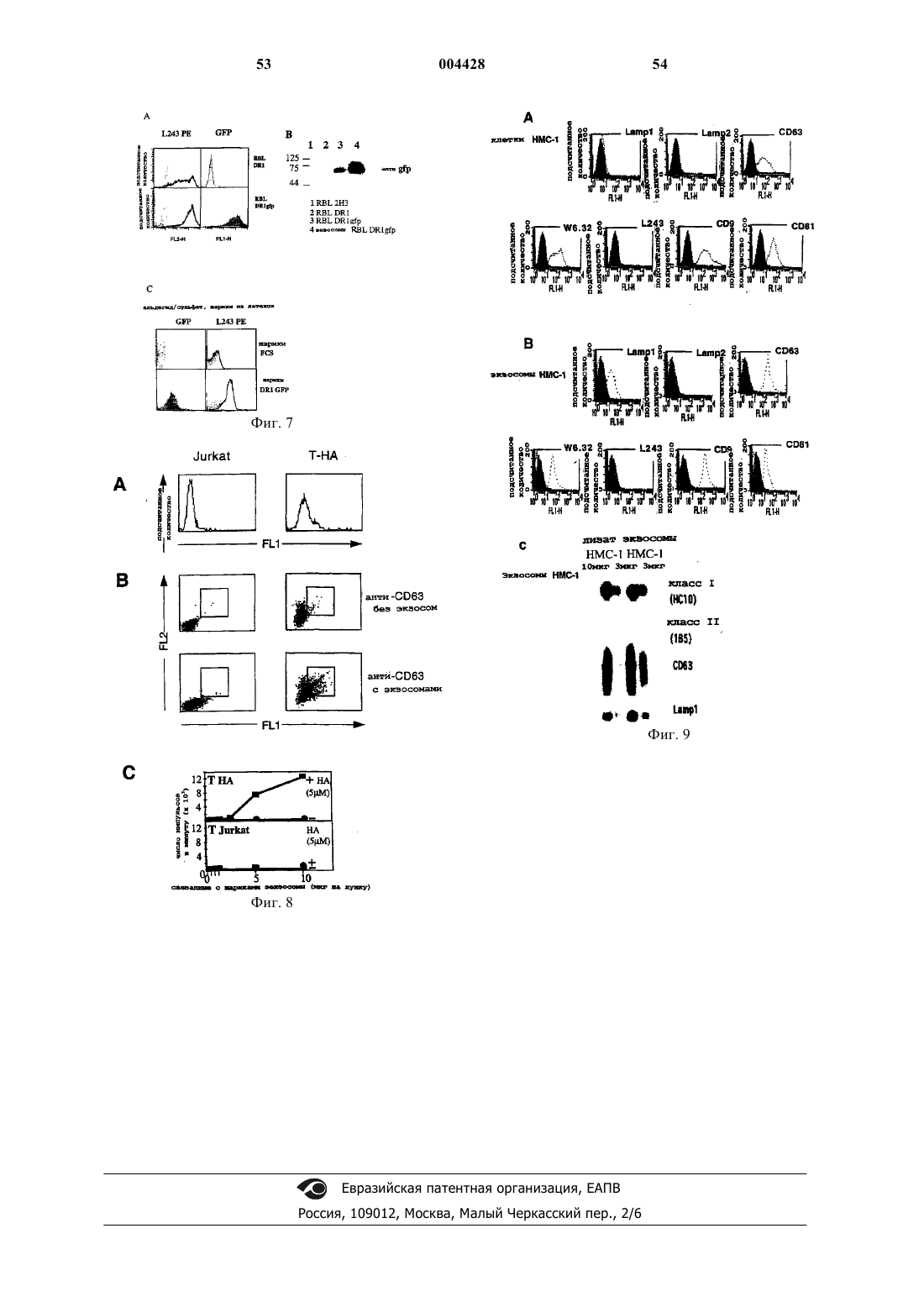
Формула / Реферат
1. Мембранная везикула, продуцируемая генетически модифицированной клеткой, причем вышеуказанная везикула содержит рекомбинантную молекулу главного комплекса гистосовместимости (СМН) человека, включающую по меньшей мере одну a-цепь класса II.
2. Мембранная везикула, продуцируемая генетически модифицированной клеткой, причем вышеуказанная везикула содержит рекомбинантную молекулу главного комплекса гистосовместимости человека, включающую по меньшей мере одну b -цепь класса II.
3. Везикула по п.1 или 2, отличающаяся тем, что рекомбинантная молекула главного комплекса гистосовместимости класса II включает a-цепь и b -цепь.
4. Везикула по любому из пп.1-3, отличающаяся тем, что рекомбинантную молекулу главного комплекса гистосовместимости класса II выбирают среди серотипов DR1-DR13, предпочтительно DR1-DR7.
5. Везикула по любому из пп.1-4, отличающаяся тем, что она дополнительно содержит молекулу главного комплекса гистосовместимости класса I.
6. Везикула по любому из пп.1-5, отличающаяся тем, что она содержит комплекс между определенным пептидом и рекомбинантной молекулой главного комплекса гистосовместимости.
7. Везикула по любому из пп.1-6, отличающаяся тем, что она дополнительно содержит одну или более представляющих интерес гетерологичных молекул.
8. Везикула по любому из пп.1-7, отличающаяся тем, что она дополнительно содержит рекомбинантный пептид или протеин, позволяющий осуществлять ее очистку.
9. Везикула по пп.1-8, отличающаяся тем, что она включает маркер.
10. Везикула по п.9, отличающаяся тем, что маркер представляет собой зеленый флуоресцентный белок (GFP).
11. Везикула по любому из пп.1-10, отличающаяся тем, что она, по существу, лишена эндогенных молекул СМН.
12. Мембранная везикула по любому из пп.1-11, отличающаяся тем, что она содержит рекомбинантную молекулу, образующуюся путем слияния между представляющим интерес пептидом и сигналом адресации.
13. Мембранная везикула, отличающаяся тем, что ее получают из генетически модифицированной тучной клетки или производной генетически модифицированной тучной клетки и она содержит одну или более представляющих интерес гетерологичных молекул.
14. Везикула по п.13, отличающаяся тем, что представляющей интерес молекулой является протеин, полипептид, пептид, нуклеиновая кислота, липид или вещество химической, биологической или синтетической природы.
15. Мембранная везикула по п.14, отличающаяся тем, что гетерологичной молекулой является молекула главного комплекса гистосовместимости, антиген, лиганд рецептора, рецептор лиганда, нуклеиновая кислота, фармакологический продукт, маркер и/или пептид для очистки.
16. Везикула по п.15, отличающаяся тем, что она экспрессирует рецептор лиганда и содержит другую представляющую интерес гетерологичную молекулу.
17. Мембранная везикула по любому из пп.13-16, отличающаяся тем, что она содержит рекомбинантную молекулу, образующуюся путем слияния между представляющим интерес пептидом и сигналом адресации.
18. Продуцирующая экзосомы клетка, отличающаяся тем, что она включает одну или более рекомбинантных нуклеиновых кислот, кодирующих молекулу главного комплекса гистосовместимости, включающую по меньшей мере одну a-цепь класса II СМН человека.
19. Клетка по п.18, отличающаяся тем, что она является тучной клеткой.
20. Клетка по п.19, отличающаяся тем, что она происходит от линии тучных клеток лейкоза базофилов, в частности от линии RBL, предпочтительно RBL-2H3.
21. Клетка по любому из пп.18-20, отличающаяся тем, что она включает рекомбинантную нуклеиновую кислоту, кодирующую a-цепь, или рекомбинантную нуклеиновую кислоту, кодирующую a-цепь и b-цепь молекулы главного комплекса гистосовместимости класса II и, необязательно, молекулу главного комплекса гистосовместимости класса I.
22. Способ получения мембранной везикулы по любому из пп. 1-17, содержащей определенную рекомбинантную молекулу, включающую по меньшей мере одну a-цепь класса II СМН человека, включающий следующие стадии:
a) культивирование тучной клетки или производной тучной клетки, включающей рекомбинантную нуклеиновую кислоту, кодирующую вышеуказанную определенную рекомбинантную молекулу;
c) рекуперацию продуцированных вышеуказанными клетками везикул, причем эти везикулы включают вышеуказанную определенную рекомбинантную молекулу.
23. Способ по п.22, отличающийся тем, что он включает промежуточную стадию b), в ходе которой клетки стимулируют для индукции и/или повышения секреции экзосом.
24. Способ по п.22 или23, отличающийся тем, что определенная рекомбинантная молекула экспонирована снаружи экзосомы или включена, частично или полностью, в цитозольную часть экзосомы.
25. Способ по любому из пп.22-24, отличающийся тем, что рекомбинантной молекулой является молекула главного комплекса гистосовместимости, антигенная молекула, лиганд рецептора, рецептор лиганда, пептид для очистки или другой представляющий интерес полипептид.
26. Способ по любому из пп.22-25, отличающийся тем, что нуклеиновая кислота включает, кроме того, область, кодирующую сигнал адресации к мембранным компартментам тучной клетки.
27. Способ получения экзосомы, содержащей комплекс пептид-СМН определенного состава, включающий
культивирование продуцирующей экзосомы клетки, включающей одну или более рекомбинантных нуклеиновых кислот, кодирующих определенную рекомбинантную молекулу СМН, включающую по меньшей мере одну a-цепь класса II СМН человека;
стимуляцию клеток для индукции высвобождения экзосом;
рекуперацию продуцированных вышеуказанными клетками экзосом, причем эти экзосомы экспрессируют на своей поверхности вышеуказанную определенную рекомбинантную молекулу СМН; и
введение в контакт экзосом с пептидом или пептидами.
28. Способ по п.27, отличающийся тем, что продуцирующей клеткой является тучная клетка или производная тучной клетки.
29. Способ по любому из пп.27-28, отличающийся тем, что продуцирующая клетка, по существу, лишена эндогенной молекулы СМН.
30. Способ получения экзосомы, содержащей комплекс пептид-СМН определенного состава, включающий
культивирование продуцирующей экзосомы клетки, включающей одну или более рекомбинантных нуклеиновых кислот, кодирующих определенную рекомбинантную молекулу СМН, и нуклеиновую кислоту, включающую область, кодирующую определенный рекомбинантный пептид;
стимуляцию клеток для индукции высвобождения экзосом;
рекуперацию продуцированных вышеуказанными клетками экзосом, причем эти экзосомы экспрессируют на своей поверхности вышеуказанную определенную рекомбинантную молекулу СМН, связанную с вышеуказанным рекомбинантным пептидом.
31. Способ по п.30, отличающийся тем, что кодирующая рекомбинантный пептид нуклеиновая кислота кодирует производное инвариантной цепи li, в котором участок CLIP подвергнут делеции и заменен вышеуказанным пептидом.
32. Способ по любому из пп.30-31, отличающийся тем, что продуцирующей клеткой является тучная клетка или производная тучной клетки.
33. Способ по любому из пп.30-32, отличающийся тем, что продуцирующая клетка, по существу, лишена эндогенной молекулы СМН.
34. Способ модификации состава экзосомы, включающий введение в продуцирующую экзосомы клетку нуклеиновой кислоты, кодирующей определенную молекулу, связанную с сигналом адресации в мембранных компартментах, и получение экзосом из вышеуказанной клетки.
35. Композиция, включающая одну или более мембранных везикул по любому из пп.1-17.
36. Применение везикулы по любому из пп.1-17 для получения поликлональных и/или моноклональных антител.
37. Способ получения антител, включающий иммунизацию животного с помощью везикулы по п.7 и рекуперацию антител и/или клетюъ, продуцирующих антитела или принимающих участие в иммунном ответе.
38. Способ по п.37, отличающийся тем, что он используется для получения моноклональных антител, в частности, специфичных к комбинации СМН-пептид.
39. Применение антитела или фрагмента антитела, полученного способом по п.37 или 38, для детекции в биологическом образце наличия соответствующих специфических антигенов.
40. Применение антитела или фрагмента антитела, полученного способом по п.37 или 38, для получения терапевтической композиции, предназначенной для ингибирования взаимодействия между рецептором Т-лимфоцита и комплексом СМН-пептид, к которому он специфичен.
41. Применение везикулы по п.1 или 2 для получения терапевтической композиции, предназначенной для ингибирования взаимодействия между рецептором Т-лимфоцита и комплексом СМН-пептид, к которому он специфичен.
42. Применение мембранной везикулы по любому из пп.1-17 для детекции in vitro или ex vivo в биологическом образце партнеров, специфичных к белковой молекуле.
43. Применение по п.42, отличающееся тем, что экзосома несет комплекс СМН-пептид для детекции специфичных к этому комплексу Т-лимфоцитов в биологическом образце.
44. Применение по п.42, отличающееся тем, что экзосома несет рецептор TcR для детекции специфичных к этому рецептору комплексов пептид-СМН в биологическом образце.
45. Применение по п.42, отличающееся тем, что экзосома несет рецептор лиганда для детекции наличия вышеуказанного лиганда в биологическом образце.
46. Способ детекции in vitro или ex vivo наличия специфичных к комплексам антиген-СМН Т-лимфоцитов в биологическом образце, включающий введение в контакт вышеуказанного образца с маркированной экзосомой по п.7, содержащей вышеуказанный комплекс антиген-СМН, и обнаружение маркировки Т-лимфоцитов в вышеуказанном образце.
47. Применение везикулы по п.6 для клональной амплификации и/или стимуляции ex vivo цитотоксических или регуляторных Т-лимфоцитов.
48. Применение везикулы по любому из пп.13-17 для получения композиции, предназначенной для транспортировки вышеуказанной молекулы к клетке.
49. Композиция, включающая одну или более мембранных везикул по любому из пп.1-17, иммобилизованных на носителе.
50. Композиция по п.49, включающая одну или более мембранных везикул, отличающаяся тем, что везикулы иммобилизованы на шарике, в частности на шарике из латекса или магнитном шарике.
51. Применение мембранной везикулы по любому из пп.1-17, в частности, в иммобилизованной на носителе форме для очистки клеток.
52. Мембранная везикула, продуцируемая генетически модифицированной клеткой, причем вышеуказанная везикула содержит рекомбинантную молекулу главного комплекса гистосовместимости человека, включающую по меньшей мере одну a-цепь класса II, и маркер.
Текст
1 Настоящее изобретение относится к областям биологии и иммунологии. Оно относится к мембранным везикулам, содержащим молекулы,в частности антигенные молекулы, с предопределенной структурой, и к их применению. В особенности оно относится к везикулам, содержащим рекомбинантные молекулы главного комплекса гистосовместимости, и их применению в качестве иммуногена или в качестве диагностического или терапевтического средства. Изобретение также относится к способам получения этих везикул, генно-инженерным конструкциям, клеткам и композициям, используемым для осуществления способов согласно изобретению. Специфичность антигенного распознавания представляет собой одну их главных характеристик клеток иммунной системы. ВЛимфоциты распознают антигены в нативной форме. Т-Лимфоциты распознают комплексы,образующиеся за счет ассоциации пептидов,получающихся в результате деградации антигенов, с молекулами главного комплекса гистосовместимости (СМН). Пептиды, происходящие от синтезируемых клетками организма антигенов (опухолевых или вирусных антигенов), ассоциируются с молекулами класса I СМН, которые распознаются цитотоксическими Тлимфоцитами. Пептиды, происходящие от экзогенных антигенов, ассоциируются с молекулами класса II СМН, которые распознаются комплементарными Т-лимфоцитами. Идентификация пептидов, представляемых молекулами СМН и распознаваемых цитотоксическими (CD8) или комплементарными (CD4) Т-лимфоцитами, положена в основу новых терапевтических и вакцинальных стратегий. Эта эволюция иммунотерапии требует развития методов оценки специфичного к антигенам иммунного ответа. Антигенные пептиды ассоциируются с молекулами СМН во внутриклеточных компартментах. В случае молекул класса II эти компартменты образованы везикулами, содержащимися в большего размера грануле, относящейся к эндоцитарному пути (Peters и др Nature, 349,669 (1991. Их слияние с цитоплазматической мембраной приводит, с одной стороны, к экспрессии комплексов пептид-СМН на клеточной поверхности и, с другой стороны, к секреции этих везикул, называемых экзосомами. В работах Raposo и др. (J. Exp. Med., 183,1161 (1996 показано, что В-лимфоциты способны выделять везикулы-экзосомы, несущие молекулы класса II СМН. Кроме того, Zitvogel и др. (Nature Medicine, 4, 594 (1998 выявили продукцию особых мембранных везикул дендритными клетками (называемых декзосомами),которые обладают полезными свойствами. Так,эти везикулы экспрессируют молекулы классовI и II СМН и способны, после сенсибилизации соответствующими антигенами, стимулировать 2 лимфоцитов и вызывать полную или частичную регрессию опухолей. Настоящее изобретение относится к новым способам и композициям, используемым в областях биологии и иммунологии. Более конкретно, в настоящем изобретении описываются новые мембранные везикулы, состав которых модифицирован определенным образом. В частности, в настоящем изобретении описывается новый способ, позволяющий получать, например, везикулы, экспрессирующие молекулы главного комплекса гистосовместимости известного состава, возможно скомплексованные с антигенным пептидом определенной структуры. Настоящее изобретение, следовательно, позволяет модифицировать состав мембранных везикул контролируемым образом и, следовательно, создавать продукты, особенно предпочтительные в терапевтическом, диагностическом или даже в экспериментальном плане. Таким образом, описанные до настоящего времени везикулы включают, в наилучшем из случаев, эндогенные молекулы СМН, то есть молекулы СМН, экспрессированные клеткой, из которой они происходят. В связи с этим, эти молекулы имеют различную структуру, не всегда идентифицируются и обычно многочисленны, в зависимости от типа HLA организма, от которого они происходят. Напротив, согласно настоящему изобретению можно получать мембранные везикулы, несущие молекулы СМН определенного состава. Кроме того, везикулы согласно изобретению обладают тем преимуществом, что они обогащены таким образом определенными молекулами СМН и придают сильную иммуногенную способность. Настоящее изобретение относится, в частности, к мембранным везикулам, содержащим молекулы предопределенной структуры, в частности, молекулы СМН предопределенной структуры. Настоящее изобретение относится, в частности, к мембранным везикулам, содержащим комплексы СМН-пептид предопределенной структуры. Настоящее изобретение относится также к способу модификации состава мембранной везикулы, включающему введение в продуцирующую такую везикулу клетку нуклеиновой кислоты, содержащей гибридный участок, который образован слитой с сайтом адресации кодирующей областью, или нуклеиновой кислоты, кодирующей протеин или полипептид,который, индивидуально или в ассоциации с одним или несколькими протеинами, по своей природе, направлен в эти мембранные везикулы. Настоящее изобретение относится также к мембранным везикулам, содержащим определенные антигенные молекулы, заякоренные в мембранной части. Такие молекулы могут находиться на поверхности везикул или, напротив,могут быть включены в цитозольную часть. Настоящее изобретение относится также к мем 3 бранным везикулам, содержащим молекулы предопределенной структуры, экспонированные на их поверхности, что позволяет осуществлять их очистку, в частности, аффинными способами. Настоящее изобретение относится также к мембранным везикулам, таким, как определенные выше, включающим, кроме того, маркр. Такой маркр позволяет, в частности, детектировать везикулы в образце, например, осуществлять их контроль in vivo. Изобретение также относится к способу получения вышеуказанных везикул, а также к применению этих везикул. Так, эти везикулы могут быть использованы в качестве иммуногена для получения антитела. Особенно предпочтительно эти везикулы используют для продукции ограниченных СМН антител, то есть специфичных к комплексу пептид-молекула СМН. Первым предметом изобретения является,в частности, мембранная везикула, отличающаяся тем, что она содержит рекомбинантную молекулу главного комплекса гистосовместимости. Термин "мембранная везикула" в смысле изобретения означает, в частности, любую везикулу, образованную включающим цитозольную часть липидным бислоем. Эти везикулы обычно образуются путем высвобождения из клеток и в силу этого также называются в настоящей заявке термином "экзосома". Мембранные везикулы(или экзосомы) согласно изобретению обычно имеют диаметр примерно 60-80 нм. Кроме того,эти везикулы несут преимущественно мембранные протеины, которые находятся в той же самой ориентации, что и в цитоплазматической мембране клеток, из которых они происходят. Согласно настоящему изобретению в настоящее время показано, что контролируемо и специфически можно модифицировать состав экзосом. Более конкретно, согласно настоящему изобретению показано, что можно получать мембранные везикулы, экспрессирующие молекулярные рекомбинантные комплексы (пред) определенного состава. Как показано дальше в настоящем описании, такие везикулы обладают особенно благоприятными свойствами как в терапевтическом, так и в диагностическом и экспериментальном плане. Согласно настоящему изобретению прежде всего возможна селекция особых клеточных популяций для продукции мембранных везикул. Согласно настоящему изобретению также выявлено, что в эти клетки генно-инженерным путем можно вводить рекомбинантные молекулы и что эти молекулы затем функционально и интенсивно экспрессируются в экзосомах. Один из первых признаков изобретения,следовательно, заключается в определении и идентификации клеточной популяции, используемой для продукции мембранных везикул. Используемой клеткой преимущественно является клетка, включающая внутренние секретор 004428 4 ные везикулы, культивируемая, генетически модифицируемая и, предпочтительно, внутренние везикулы которой могут быть выделены под действием внешней стимуляции. Речь идет по существу о клетках млекопитающего, в частности, о клетках животных, но также о клетках человеческого происхождения. Кроме того, речь может идти о первичных культурах или иммортализованных линиях. Особенно предпочтительно исходные клетки по существу лишены молекул СМН, то есть не экспрессируют или в незначительной степени экспрессируют эндогенные молекулы СМН. Эта характеристика может оказаться очень важной при некоторых применениях, как будет показано дальше. В литературе описаны различные типы продуцирующих экзосомы клеток, таких, как,например, дендритные клетки или Влимфоциты. Тем не менее, эти клетки обычно трудно трансфицируемы и обогащены эндогенными молекулами СМН. В связи с этим, хотя они и могут быть использованы для осуществления изобретения, везикулы согласно настоящему изобретению более предпочтительно могут быть получены из мастоцитов (тучных клеток) или происходящих от мастоцитов клеток. Согласно особому варианту осуществления, мембранные везикулы согласно изобретению предпочтительно получают из мастоцитов или происходящих от мастоцитов клеток. Мастоциты объединяют совокупность клеточных типов, происходящих от медуллярных предшественников, остающихся после дифференцировки в эпителиях, таких как кожи, легкого, кишечника или селезенки (Smith и Weis, Immunology Today, 17, 60 (1996. Эти клетки характеризуются по существу тем, что их цитоплазма в большинстве своем образована гранулами, которые содержат гистамин, а также гепарин или протеазы, и тем, что они экспрессируют на своей поверхности рецепторы с высоким сродством к иммуноглобулинам Е (IgE). Кроме того, другое преимущество использования мастоцитов согласно изобретению заключается в способности вызывать (в особенности, сильно стимулировать) экзоцитоз (то есть высвобождение) экзосом путем различных обработок. Так,можно регулировать продукцию везикул путем обработки в присутствии кальций-ионофора или, более физиологическим путем, за счет стимуляции рецепторов с высоким сродством к IgE. Эти клетки обладают свойствами, особенно благоприятными для осуществления настоящего изобретения, а именно наличие внутренних секреторных везикул, возможность их культивирования и способность вызывать массивный экзоцитоз. Кроме того, в настоящее время показано, как описывается в примерах, что эти клетки также могут быть стабильно генетически модифицированы, что представляет собой осо 5 бенно благоприятное свойство для осуществления настоящего изобретения. Более специфически, везикулы согласно изобретению имеют диаметр около 60-80 нм и их получают из мастоцитов или происходящих от мастоцитов клеток. Согласно предпочтительному варианту,предлагаемые в изобретении мембранные везикулы по существу лишены эндогенных молекул СМН. Отсутствие эндогенных молекул СМН (то есть молекул СМН клетки, продуцирующей везикулы) может быть выявлено с помощью специфических антител классическими способами. Это отсутствие также может быть выявлено по селективности антител, получаемых путем иммунизации с помощью везикул. Как указано в примерах, предлагаемые в изобретении везикулы, согласно особенно предпочтительному варианту, способны вызывать у животного продукцию антител, специфичных к определенным рекомбинантным молекулам,экспрессируемым ими, без детекции антител,направленных против генетически немодифицированных клеток. Термин "по существу лишенный" означает то, что некоторые молекулы СМН могут быть представлены в очень незначительных количествах, трудно детектируемых классическими методами, и без заметной интерференции с антигенной специфичностью везикул согласно изобретению. Особые мембранные везикулы согласно изобретению более специфически характеризуются следующими свойствами: они по существу лишены эндогенных молекул СМН; они включают одну или несколько рекомбинантных молекул определенной структуры, например, рекомбинантные комплексы пептид-СМН определенного состава. Такие везикулы согласно изобретению преимущественно продуцируются происходящими от мастоцитов клетками, которые по существу лишены эндогенных молекул СМН. В этом отношении, известно, что мастоциты аккумулируют в своих гранулах секреции молекулы класса II СМН. В частности, мастоциты способны предпочтительно аккумулировать комплексы СМН-II-пептид в особых внутриклеточных мультивезикулярных компартментах, гранулах секреции (Raposo и др., Mol. Biol. Cell., 8,2619 (1997. Эти клетки, взятые у млекопитающего, включают, следовательно, эндогенные молекулы СМН. Особенно предпочтительно в рамках настоящего изобретения используют линии клеток, происходящих от мастоцитов,которые по существу лишены эндогенных молекул СМН. Различные линии мастоцитарных клеток описаны в литературе. Согласно настоящей заявке теперь показано, что некоторые из этих линий обладают незначительными содержаниями молекул СМН и, следовательно, являются особенно предпочтительными для осуществления изобретения. В качестве примера 6 можно назвать, в частности, линии, происходящие от клеток RBL (базофильный лейкоз крысы), депонированные в АТСС под номером(1974, линию KU-812 (Butterfield и др., Leukemia Res., 12, 345 (1988, или клетки линии человеческих незрелых мастоцитов, такой, как линия НМС (Nilsson и др., Scand. J. Immunol.,39, 489 (1994. Особым примером линии является линия RBL-2H3 (Basumian и др., Eur. J.Immunol., 11, 317 (1981. Разумеется, может быть использована любая другая клетка, обладающая вышеуказанными свойствами. В рамках настоящего изобретения выражение "определенный состав" более конкретно означает то, что везикулы согласно изобретению обладают, например, большой антигенной специфичностью и большой специфичностью в отношении гаплотипа. Так, везикулы, описанные в уровне техники, обычно экспрессируют молекулы СМН различных и неизвестных гаплотипов. Напротив, предпочтительные везикулы согласно изобретению экспрессируют рекомбинантные молекулы, гаплотип которых точно предопределен. Термин "рекомбинантный" указывает, что молекула получается в результате экспрессии, в продуцирующей везикулы клетке,рекомбинантной нуклеиновой кислоты, кодирующей эту молекулу. Мембранные везикулы согласно настоящему изобретению, следовательно, более предпочтительно получают из составляющих линии клеток, которые генетически модифицированы для экспрессии компонентов предопределенной структуры. Как указано выше, везикулы согласно изобретению экспрессируют преимущественно определенные молекулы СМН. Молекулы человеческого СМН группируют в виде двух различных классов: молекулы СМН класса I и молекулы СМН класса II. Согласно особому варианту осуществления, везикулы согласно изобретению экспрессируют одну или несколько рекомбинантных молекул главного комплекса гистосовместимости класса II. В этом отношении, молекулы человеческого СМН класса II образованы двумя цепями, -цепью и -цепью, причем -цель придает аллельную специфичность комплексу. Согласно одному особому варианту, везикулы согласно изобретению экспрессируют преимущественно рекомбинантную -цепь молекулы главного комплекса гистосовместимости класса II. Согласно другому особому варианту,везикулы согласно изобретению экспрессируют преимущественно рекомбинантные -цепь и цепь молекулы главного комплекса гистосовместимости класса II. Различные типы человеческих молекул СМН II идентифицированы, охарактеризованы и секвенированы (см., например, Immunogenetics, 36, 135 (1992. Предпочтительно можно 7 назвать молекулы типа DR1 - DR13, в частности, DR1, DR2, DR3, DR4, DR5, DR6 и DR7. Кодирующая человеческие DR, в особенностиDR1 -DR13, ДНК может быть легко выделена из клеток, банков или плазмид классическими методами молекулярной биологии. Эти последовательности описаны, в частности, у Bodmer и др.(Tissue antigens, 44, 1 (1994. Экзосомы согласно изобретению, следовательно, предпочтительно экспрессируют молекулу СМН класса II,включающую -цепь и -цепь, селекционированные из гаплотипов DR1 - DR13, и еще более предпочтительно DR1-DR7. Согласно особому примеру, изобретение относится к любой мембранной везикуле, содержащей рекомбинантную - и/или -цепь молекулы СМН гаплотипа DR1. Согласно другому варианту осуществления, везикулы согласно изобретению экспрессируют одну или несколько рекомбинантных молекул главного комплекса гистосовместимости класса I. Молекулы СМН класса I также образованы двумя цепями, трансмембранной и полиморфной -цепью и 2-микроглобулином,который является постоянным и растворимым. В случае человека -цепь кодируют три генных локуса, обозначаемых А, В и С. В классических молекулах CMH-I каждый локус А, В и С цепи подвергнут аллельной вариации. Таким образом, выявляют аллели А 1, А 2, A3, и т.д. А 10, В 1, В 7, В 37, В 54, и т.д., CW3, CW6, и т.д.(см., например, Bodmer и др., цитировано выше; и Immunogenetics, 36 (1992), цитировано выше). Экзосомы согласно изобретению предпочтительно экспрессируют -цепь классической,то есть трансмембранной и полиморфной, молекулы CMH-I. Еще более предпочтительно речь идет об -цепи молекулы CMH-I аллеля А 1, A2 или A3. Согласно особому варианту осуществления, экзосомы согласно изобретению экспрессируют -цепь неклассической, то есть неполиморфной, молекулы CMH-I. В самом деле, в противоположность так называемым "классическим" молекулам CMH-I, подвергнутым в значительной степени полиморфизму, у человека существуют "неклассические" молекулы CMH-I,которые по существу являются неполиморфными. Такие молекулы описаны, например, у Bendelac и др. (Ann. Rev. Immunol., 535 (1997. Предпочтительным примером неклассической молекулы CMH-I согласно изобретению является молекула Cdl. Разумеется, в рамках настоящего изобретения может быть экспрессирована любая другая молекула CMH-I человека. Согласно особому примеру, изобретение относится, следовательно, к любой мембранной везикуле, содержащей рекомбинантный протеин молекулы CMH-I. 8 Согласно одному особому варианту, везикулы согласно изобретению содержат несколько молекул СМН класса I и/или класса II. Так,предпочтительно везикула содержит, например,2 молекулы CMH-II разных гаплотипов или больше. Разумеется, возможна любая другая комбинация молекул СМН, такая как, например,CMH-I и СМН-II. Везикулы согласно изобретению, экспрессирующие один или несколько определенных комплексов СМН, особенно предпочтительны,поскольку они позволяют иметь данный антигенный пептид в определенной структуре МНС. В этом отношении, согласно более предпочтительному варианту осуществления изобретения,мембранные везикулы включают комплекс между определенным пептидом и рекомбинантной молекулой главного комплекса гистосовместимости. Везикулы согласно изобретению, кроме того, могут включать одну или несколько других, представляющих интерес, гетерологичных молекул, кроме или вместо вышеуказанных молекул СМН. В этом отношении, согласно особому варианту, изобретение относится к мембранным везикулам, продуцируемым мастоцитами или происходящими от мастоцитов клетками, отличающимся тем, что они содержат одну или несколько представляющих интерес гетеро-логичных молекул. Термин "происходящая от мастоцитов клетка" означает линии, трансформированные и/или иммортализованные и/или полученные из мастоцитов или базофилов и обладающие свойствами мастоцитов (аккумуляция внутренних секреторных везикул). Термин "гетерологичный" указывает, что представляющая интерес молекула не присутствует в этой форме в экзосомах согласно изобретению в естественном состоянии. Представляющие интерес молекулы, которые несут экзосомы согласно изобретению или которые содержатся в них, могут быть любым протеином, полипептидом, пептидом, нуклеиновой кислотой, липидом, а также любым,представляющим интерес веществом (химической, биологической или синтетической природы). Эти молекулы могут быть рекомбинантной природы и могут быть введены в продуцирующую клетку или прямо в/на экзосомы. В особенности предпочтительными типами представляющих интерес молекул являются, в частности,молекулы СМН, антигены (целые или в виде пептидов), лиганды рецепторов, специфичные к лигандам рецепторы, нуклеиновые кислоты,фармакологические продукты, маркры или пептиды или протеины, позволяющие осуществлять очистку везикул. В качестве антигена можно, в частности,назвать любой, например, цитоплазматический,протеин вирусного или опухолевого происхождения. В качестве предпочтительных примеров протеинов вирусного происхождения можно 9 назвать, в частности, любой цитоплазматический или мембранный протеин, процессированный вирусами EBV (вирус Эпштайна-Барра),CMV (вирус огуречной мозаики), VIH (вирус иммунного дефицита человека), кори, гепатита,и т.д. Более предпочтительно речь идет о цитоплазматических протеинах, то есть по существу не обнаруживаемых иммунной системой в процессе обычной инфекции, и, следовательно,слабо иммуногенных в естественных условиях,или также о протеинах или фрагментах мембранных протеинов. В качестве предпочтительных примеров протеинов опухолевого происхождения можно назвать, в частности, протеины р 53 (дикого типа или любой мутированной формы, присутствующей в опухоли), MAGE (в частности, MAGE 1, MAGE 2, MAGE 3, MAGE 4, MAGE 5 и MAGE 6), MART (в частности,MART 1), Gp100, ras-протеины (p21 дикого типа или мутированные), и т.д. Разумеется, что любой другой, представляющий интерес протеин может быть экспрес-сирован в экзосомах согласно изобретению или на их поверхности,следуя технической инструкции настоящей заявки. В этом отношении, рекомбинантные антигенные молекулы могут находиться либо на поверхности везикул (экспонированные), либо внутри везикул. В самом деле, что особенно неожиданно, изобретатели показали, что везикулы согласно изобретению, содержащие в своем цитозоле рекомбинантный антиген (в частности, р 53), способны индуцировать у животного в очень высокой степени продукцию антител,направленных против этого антигена. Из рецепторов лигандов можно назвать,как правило, любой природный или получаемый методами генной инженерии рецептор лиганда. В частности, речь может идти о любом рецепторе гормона, фактора роста, лимфокина, трофического фактора, антигена, и т.д. В особенности можно назвать рецепторы интерлейкинов IL1 IL15, рецептор гормона роста или рецептор факторов стимуляции колоний гранулоцитов и/или макрофагов (G-CSF, GM-CSF, CSF, и т.д.). Особый пример рецептора лиганда представляет собой соединение простое антителоцепь (ScFv), которая позволяет протекать взаимодействию со специфичным лигандом. Другим, особенно предпочтительным в смысле изобретения примером является антигенный рецептор Т-лимфоцитов (TcR). Экзосомы согласно изобретению, экспрессирующие на своей поверхности один или несколько определенныхTcR, представляют собой особенно предпочтительные средства для анализа и диагностики,как это будет подробно показано дальше. В качестве фармацевтического продукта можно назвать любое активное вещество химической природы, такое, как, например, фармацевтические продукты, получаемые обычными химическими способами. Также можно назвать 10 любой протеин, полипептид или пептид, обладающий биологической активностью, такой,как, например, токсин, гормон, цитокин, фактор роста, фермент, супрессор опухоли, и т.д. Нуклеиновой кислотой может быть любая ДНК или РНК, кодирующая протеин, полипептид или фармакологический пептид, такой, как указанный выше, а также любая другая нуклеиновая кислота, обладающая особым свойством(антисмысловая; антиген, промотор, репрессор,сайт связывания фактора транскрипции, и т.д.). Речь может также идти об олигонуклеотиде,кодирующей фазе, искусственной хромосоме и т.д. Везикулы согласно изобретению, несущие рецептор лиганда, могут быть использованы для детекции любого взаимодействия типа рецептор-лиганд, в особенности с низкой степенью сродства, в любом биологическом образце, как это будет пояснено более подробно в продолжении текста. С другой стороны, такие везикулы также могут быть использованы для транспортировки представляющих интерес веществ (протеин, пептид, нуклеиновая кислота, химическое вещество, и т.д.) к клетке. Таким образом, экзосомы согласно изобретению могут быть использованы, как правило, для транспортировки и переноса любой молекулы в клетки, in vitro, exvivo или in vivo. Изобретение, следовательно,относится к любой везикуле, такой, как описанная выше, содержащей представляющую интерес гетерологичную молекулу, используемому в качестве вектора для переноса вышеуказанной молекулы в клетку. Согласно более предпочтительному варианту, экзосомы согласно изобретению используют для направленного переноса представляющих интерес веществ к селекционированным клеточным популяциям. Так, можно получать везикулы согласно изобретению, содержащие представляющее интерес вещество (токсин,гормон, цитокин, рекомбинантная нуклеиновая кислота, и т.д.) и экспрессирующие на своей поверхности рецептор лиганда или лиганд рецептора, и вводить вышеуказанные везикулы в контакт с клетками, экспрессирующими лиганд или соответствующий рецептор. Этот подход,следовательно, позволяет осуществлять целенаправленный и эффективный перенос. В этом отношении, особым предметом изобретения является везикула, такая как указанная выше, отличающаяся тем, что она экспрессирует рецептор лиганда и содержит представляющую интерес гетерологичную молекулу. Везикулы согласно изобретению, кроме того, могут содержать рекомбинантный пептид или протеин, позволяющий осуществлять очистку везикул. Таким образом, на деле в изобретении описывается возможность генетической модификации состава экзосом и, следовательно,придание им способности экспрессировать особую "маркированную"' молекулу, позволяющую 11 ее очищать. В частности, можно получать экзосому, экспонирующую пептид особой структуры, который может быть легко детектирован и зафиксирован рецепторной молекулой. Согласно особому примеру, экзосома представляет собой продукт, включающий в свою структуру пептидную молекулу, содержащую звено His 6(то есть 6 последовательных остатков гистидина). Присутствие такого остатка на поверхности экзосом позволяет легко очищать их на функционализованном никелем носителе. Могут быть использованы другие рекомбинантные пептиды этого типа, как, например, tag c-myc,VSV или НА. Наконец, согласно особому варианту, везикулы согласно изобретению включают, кроме того, маркр. Маркр может быть различной природы (ферментативной, флуоресцентной,радиоактивной, и т.д.) и находится в везикуле или на ее поверхности. Предпочтительной маркировкой является нерадиоактивная, как, например, флуоресцентная маркировка. Используемым маркром более предпочтительно является флуорохром или фермент с хромогенным субстратом. Маркировка может быть реализована непосредственно в случае продуцирующей клетки или же продуцированных экзосом. Изобретение также относится к любой композиции, включающей одну или несколько мембранных везикул, таких, как указанные выше. Композиции согласно изобретению, кроме того, могут включать множество мембранных везикул, таких, как указанные выше, несущих различные рекомбинантные молекулы. Композиция согласно изобретению в особенности может включать мембранные везикулы, такие, как указанные выше, несущие рекомбинантные молекулы СМН разных гаплотипов в ассоциации с одним и тем же антигенным пептидом. Речь может также идти о композициях, включающих мембранные везикулы, такие, как указанные выше, несущие рекомбинантные молекулы СМН одного и того же гаплотипа, ассоциированные, например, с разными антигенными пептидами. Разумеется, возможны другие комбинации везикул согласно изобретению. Композиции согласно изобретению обычно включают носитель, такой, как буферный раствор, солевой раствор, физиологический раствор, и т.д., позволяющий сохранять структуру везикул. Кроме того, они могут включать любой стабилизатор, любое поверхностно-активное вещество, и т.д., предпочтительно приемлемое для биологического использования (in vitro илиin vivo). Эти композиции могут быть упакованы в любом соответствующем устройстве, таком,как тюбик, флакон, ампула, бутыль, мешочек и т.д. и могут храниться, например, при температуре 4 С или -20 С. Типичные композиции согласно изобретению включают 5-5000 мкг, например, 5-200 мкг, экзосом. 12 Везикулы согласно изобретению получают из генетически модифицированных клеток. Как указано выше, в самом деле, согласно настоящему изобретению выявлено, что в некоторые клетки можно вводить генно-инженерным путем рекомбинантные молекулы и что эти молекулы затем функционально и интенсивно экспрессируются в экзосомах. Для получения везикул согласно изобретению, несущих рекомбинантные молекулы определенного состава, первая стадия состоит, следовательно, во введении в продуцирующую везикулы клетку, такую, как указанная выше, генно-инженерных конструкций, позволяющих протекать экспрессии выбранной рекомбинантной молекулы (или выбранных рекомбинантных молекул). Используемые для получения клеток генно-инженерные конструкции могут включать,как правило, кодирующую область, находящуюся под контролем функционального промотора в используемой клетке (полигенный экспрессирующий кластер). Обычно, используемым промотором является, следовательно, функциональный промотор в клетках млекопитающих. Речь может идти,например, о вирусном, клеточном или бактериальном промоторе. Речь может идти о конститутивном или регуляционном промоторе, который позволяет протекать экспрессии до высоких содержаний протеина в клетке. Из используемых промоторов в качестве примера можно назвать непосредственный ранний промотор цитомегаловируса (CMV), промотор вируса SV40,промотор гена тимидинкиназы, в частности,HSV-1 ТК, промотор LTR ретровируса, в частности, LTR-RSV, или сильный эндогенный промотор мастоцитов. Особенно предпочтительный способ осуществления включает использование промотора SR, такого как описанный более подробно в примерах. Используемая кодирующая область обычно образована комплементарной, геномной или синтетической ДНК (например, модифицированной путем встраивания некоторых интронов или в отношении предпочтительного использования кодонов). В целом, речь идет о кДНК. Эта нуклеиновая кислота может быть получена любыми известными способами молекулярной биологии и, в частности, путем скрининга банка, амплификации, синтеза, ферментативных расщеплений и лигирований, и т.д. В зависимости от типа используемой кодирующей области, кроме того, в конструкцию могут быть внесены некоторые изменения. Так,в некоторых случаях особенно предпочтительным может быть встраивание в кодирующую область сигнальной последовательности, позволяющей направлять продукт экспрессии в особый компартмент клетки, в частности, в мембранный компартмент (внутренний, цитоплазматический, и т.д.). Этот сигнал адресации мо 13 жет быть расположен выше (5'), ниже (3') или внутри кодирующей области. Сигнал адресации предпочтительно находится у 3'-конца кодирующей области, более предпочтительно на ее цитоплазматическом участке, и в рамке считывания с кодирующей областью. Использование сигнала адресации может быть особенно полезно для того, чтобы благоприятствовать аккумуляции продукта экспрессии в данном внутриклеточном аппартменте или на его поверхности,в частности, в секреторных везикулах или на их поверхности. Этот способ осуществления особенно адаптирован для экспрессии молекулы,такой, как "маркированный" пептид, антиген,молекула CMH-I или еще лиганд рецептора. Однако, в настоящей заявке показано, что человеческие молекулы СМН-II непосредственно,без дополнительного введения особого сигнала,могут быть экспрессированы в секреторных везикулах мастоцитов, даже ксеногенных клеток. Для осуществления настоящего изобретения, в частности, в качестве сигнала адресации можно использовать фрагмент нуклеиновой кислоты, обладающий последовательностью части следующих генов: Lampl, CD63, LIMPII,Cdlc, FcR. Эти гены действительно включают области, кодирующее сигналы адресации протеина к эндосомным компартментам клеток(1994. Используемый согласно настоящему изобретению сигнал адресации отвечает, например, формуле G-Y-X-X-I, в которой X означает любой остаток аминокислоты. Особенно адаптированный для осуществления настоящего изобретения сигнал адресации образован сигнальным пептидом протеина LAMP1 последовательности SHAGYQTI. Другой тип сигнала, позволяющего осуществлять адресацию к мембранным компартментам, включает всю или часть трансмембранной области протеина. Адресация продукта экспрессии к клеточным компартментам, позволяющая присутствовать этому продукту в экзосомах, также может быть реализована путем слияния кодирующей области со всей или частью кодирующей области мембранного или трансмембранного протеина, в частности, мембранного или трансмембранного протеина, экспрессированного в экзосомах. В настоящем контексте, особый вариант осуществления изобретения включает введение рекомбинантного продукта в экзосому, путем экспрессии этого продукта в продуцирующей клетке, в слитой с мембранным или трансмембранным протеином форме. Особым примером такого протеина является, например, рекомбинантный протеин СМН, введенный в продуцирующую клетку, в частности, бета-цепь, предпочтительно бета-цепь СМН класса II. Так,представленные в примерах результаты показывают, что такое слияние позволяет эффективно аккумулировать в экзосоме любой представ 004428 14 ляющий интерес полипептид, без ухудшения его свойств и свойств молекулы СМН. Этот аспект настоящего изобретения представляет собой новую концепцию векторизации рекомбинантных продуктов в экзосоме и может быть применен к любому рекомбинантному продукту, вводимому в любой тип экзосомы. В этом отношении, изобретение, следовательно, относится к любой экзосоме, включающей рекомбинантную молекулу, получаемую путем слияния между представляющим интерес пептидом и сигналом адресации. Речь может идти об зкзосоме, получаемой, например, из мастоцита, дендритной клетки, опухолевой клетки или таже из В-лимфоцита. Представляющим интерес полипептидом может быть антиген (или фрагмент антигена) или любой другой, представляющий интерес биологический продукт. Сигналом адресации может быть любой пептид, полипептид или протеин, обладающий свойством направлять слитый продукт к мембранному, особенно внутриклеточному, компартменту, такому, как указанный выше. Речь идет преимущественно о цепи молекулы СМН. Согласно особому варианту осуществления изобретения, эти везикулы получают путем введения в продуцирующую клетку химерной нуклеиновой кислоты, кодирующей слитый протеин, который включает рекомбинантный продукт, связанный с С-концом бета-цепи молекулы СМН, предпочтительно СМН класса II. В используемых конструкциях кодирующая область функционально связана с промотором, чтобы могла происходить его экспресия в клетках. Кроме того, конструкции согласно изобретению преимущественно могут влючать участок, расположенный у 3'-конца кодирующей области, который точно определяет сигнал конца транскрипции (например, участок polyA). Полигенные экспрессирующие кластеры согласно изобретению преимущественно составляют часть вектора плазмидного, вирусного,эписомального, искусственного хромосомного,и т.д., типа/ В этом отношении, такой вектор преимущественно включает систему, позволяющую осуществлять селекцию содержащих его клеток. В частности, векторы преимущественно включают ген, кодирующий продукт,придающий резистентность к агенту, например,к антибиотику (ампициллину, гигромицину,генетицину, неомицину, зеоцину, и т.д.). Согласно особому варианту осуществления, каждый вектор включает один полигенный экспрессирующий кластер, такой, как описанный выше. Согласно этому варианту осуществления, клетки, следовательно, модифицируют путем введения нескольких векторов, когда несколько молекул экспрессируются в везикулах (например,-цепь и -цепь CMH-II). Согласно этому варианту осуществления, каждый тип используемого вектора включает преимущественно разную 15 систему селекции, позволяющую легко селекционировать многочисленные трансфектанты. Согласно другому варианту осуществления, вектор может включать несколько полигенных экспрессирующих кластеров, таких, как указанные выше, например, один, кодирующий-цепь, и другой, кодирующий -цепь CMH-II. Используемые векторы предпочтительно являются векторами плазмидного типа и включают, например, бактериальный источник репликации, позволяющий манипуляцию с ними и их легкое получение in vitro. Такие векторы, в частности, могут быть сконструированы из плазмид типа pBR322, pUC, pBS, pSR, и т.д. Для получения экзосом согласно изобретению, следовательно, используют генетически модифицированные клетки, экспрессирующие селекционированные молекулы. Эти генетически модифицированные клетки получают путем введения в клетки, выбранные как указано выше, вышеуказанных генно-инженерных конструкций. Введение генно-инженерных конструкций может быть осуществлено различными способами, главным образом в зависимости от используемого типа клетки. Так, перенос нуклеиновых кислот может быть осуществлен любым известным способом, таким как электропорация,преципитация при использовании фосфата кальция, химического агента (катионный пептид, полимеры, липиды, и т.д.), баллистический способ, и т.д. В случае вирусных векторов перенос обычно осуществляют путем простой инфекции клеток. Используемые количества вектора, кроме того, могут быть адаптированы специалистом в зависимости от типа переноса и используемых клеток. В этом отношении, особенно эффективным методом введения нуклеиновых кислот в мастоциты является электропорация векторов. С другой стороны, когда в клетки должны быть введены несколько конструкций (векторов), они могут быть перенесены одновременно или последовательно. После переноса, клетки, в которые действительно были введены нуклеиновые кислоты,селекционируют и клонируют на основании их резистентности к соединению (например, к антибиотику) благодаря гену устойчивости, присутствующему в перенесенной ДНК. Эти клетки могут быть использованы незамедлительно для получения экзосом согласно изобретению или же могут быть сохранены для позднейшего использования. В этом случае, клетки могут храниться при температуре 4 С в обычной кондиционированной среде в течение периода времени, достаточного для получения различных партий продуцированных экзосом. Клетки также можно хранить в замороженной форме (например, в жидком азоте) для позднейшего использования. В этом отношении, согласно настоящему изобретению, также можно создавать бан 004428 16 ки клеток, продуцирующих экзосомы, которые обладают особыми свойствами. В частности,согласно изобретению можно создавать банки клеток, экспрессирующих основные типы HLA молекул класса II СМН. В связи с этим, в зависимости от предусматриваемых применений и в зависимости от типа HLA пациента тогда можно выбирать в банке клетки, продуцирующие молекулы соответствующего СМН без необходимости реконструировать эти клетки от случая к случаю. В этом отношении, особым предметом изобретения является продуцирующая экзосомы клетка, такая, как указанная выше, в частности,мастоцит, отличающаяся тем, что она включает рекомбинантную нуклеиновую кислоту, кодирующую молекулу главного комплекса гистосовместимости. Изобретение относится также к любой продуцирующей экзосомы клетке, такой,как указанная выше, в частности, к мастоциту,отличающейся тем, что она включает рекомбинантную нуклеиновую кислоту, кодирующую инвариантную, в частности, модифицированную, цепь li, для включения антигенного пептида вместо области CLIP или пептида, позволяющего осуществлять очистку экзосомы. Речь идет преимущественно о клетке млекопитающего, в особенности, приходящей от животного, в частности о клетке грызуна. Речь также может идти о клетке человеческого происхождения. Согласно более особому варианту,речь идет о клеточной линии, происходящей от мастоцита, такой, как, в частности, мастоцитарная линия базофильного лейкоза. В качестве особого примера можно назвать клетки линииRBL, в частности, RBL-2H3, клетки линии KU812 или НМС-1. Рекомбинантная нуклеиновая кислота предпочтительно кодирует - и/или -цель молекулы главного комплекса гистосовместимости класса II и/или молекулу главного комплекса гистосовместимости класса I. Согласно другому варианту осуществления, клетка включает несколько нуклеиновых кислот, кодирующих, соответственно, -цепь и -цепь молекулы главного комплекса гистосовместимости класса II. Согласно настоящему изобретению, просто и воспроизводимо можно получать значительные количества экзосом известного состава. Для получения экзосом, вышеописанные, генетически модифицированные клетки культивируют в соответствующей среде и экзосомы рекуперируют. Особым предметом изобретения является,таким образом, способ получения экзосомы,содержащей определенную рекомбинантную молекулу, включающий следующие стадии: а) культивирование мастоцита или происходящей от мастоцита клетки, включающей рекомбинантную нуклеиновую кислоту, коди 17 рующую вышеуказанную определенную рекомбинантную молекулу; с) рекуперация продуцированных вышеуказанными клетками экзосом, причем эти экзосомы включают вышеуказанную определенную рекомбинантную молекулу. Способ согласно изобретению предпочтительно включает, кроме того, промежуточную стадию b), в процессе выполнения которой клетки стимулируют для индуцирования и/или повышения секреции экзосом. С другой стороны, способ согласно изобретению позволяет получать везикулы, в которых определенная рекомбинантная молекула находится снаружи экзосомы или включена,частично или полностью, в цитозольную часть экзосомы. Как указано выше, согласно способу изобретения, рекомбинантной молекулой может быть, например, молекула главного комплекса гистосовместимости, антигенная молекула, лиганд рецептора, рецептор лиганда или пептид для очистки или любой другой представляющий интерес полипептид. Кроме того, как указано выше, согласно некоторым вариантам осуществления, используемая в способе нуклеиновая кислота включает, кроме того, область, кодирующую сигнал адресации к мембранным компартментам, в частности, внутренним секреторным везикулам, мастоцита. Другим особым предметом изобретения является способ получения мембранной везикулы, включающий культивирование продуцирующей экзосомы клетки, включающей рекомбинантную нуклеиновую кислоту, кодирующую рекомбинантную, в частности, человеческую, молекулу СМН, в особенности класса I или класса II; и рекуперацию продуцированных экзосом, в случае необходимости, после стимуляции экзоцитоза. В этом отношении, изобретение также относится к способу получения экзосомы, содержащей комплекс пептид-СМН определенного состава, отличающемуся тем, что он включает культивирование продуцирующей экзосомы клетки, включающей одну или несколько рекомбинантных нуклеиновых кислот, кодирующих определенную рекомбинантную молекулу СМН; стимуляцию клеток для индукции высвобождения экзосом; рекуперацию продуцированных вышеуказанными клетками экзосом, причем эти экзосомы экспрессируют на своей поверхности вышеуказанную определенную рекомбинантную молекулу СМН; и введение в контакт экзосом с пептидом или пептидами. Для осуществления изобретения, используемый пептид или используемые пептиды могут быть синтетическими пептидами, смесями 18 пептидов, клеточными экстрактами, например,смесью пептидов, экстрагированной из опухолевых клеток. Пептид или пептиды могут быть в изолированной форме или очищенной форме или, как указано выше, в виде смеси. Кроме того, после введения в контакт экзосом с пептидами, экзосомы могут быть выделены или очищены обычными способами. Согласно другому варианту, изобретение относится к способу получения экзосомы, содержащей комплекс пептид-СМН определенного состава, отличающемуся тем, что он включает культивирование продуцирующей экзосомы клетки, включающей одну или несколько рекомбинантных нуклеиновых кислот, кодирующих определенную рекомбинантную молекулу СМН, и нуклеиновую кислоту, включающую область, кодирующую определенный рекомбинантный пептид; стимуляцию клеток для индукции высвобождения экзосом; рекуперацию продуцированных вышеуказанными клетками экзосом, причем эти экзосомы экспрессируют на своей поверхности вышеуказанную определенную рекомбинантную молекулу СМН, ассоциированную с вышеуказанным рекомбинантным пептидом. Более предпочтительно согласно этому способу нуклеиновая кислота, включающая кодирующую рекомбинантный пептид область,кодирует производное инвариантной цели 1i, в котором участок CLIP подвергнут делении и заменен вышеуказанным пептидом. Этот вариант реализации обеспечивает большую специфичность в отношении образования комплекса пептид-СМН. Согласно другому варианту, нуклеиновая кислота включает кодирующую пептид область и область адресации к внутриклеточным компартментам. Кроме того, нуклеиновая кислота может включать несколько областей, кодирующих один и тот же или разные антигенные пептиды. Используемые согласно способу продуцирующие клетки представляют собой предпочтительно мастодиты или происходящие от мастоцитов клетки. Согласно этому варианту осуществления, стимуляцию клеток для индукции высвобождения экзосом предпочтительно реализуют с помощью одного или нескольких кальций-ионофоров или с помощью IgE. Согласно особенно предпочтительному варианту осуществления, используемые в способе продуцирующие клетки по существу лишены эндогенных молекул СМН. Следующий предмет изобретения относится к способу модификации состава экзосомы,включающему введение в продуцирующую экзосомы клетку нуклеиновой кислоты, кодирующей оп 19 ределенную молекулу, связанную с сигналом адресации в мембранных компартментах; и получение экзосом из вышеуказанной клетки. Этот способ позволяет преимущественно получать экзосомы, экспрессирующие определенные и различные рекомбинантные молекулы. Экзосомы согласно изобретению могут быть использованы для многочисленных применений, таких, как, например, в качестве средств для анализа, диагностики, терапии или эксперимента. Так, они могут быть использованы для анализа Т-специфичного антигенного ответа; для изучения взаимодействий рецептор/ лиганд с низкой степенью сродства рецептора к лиганду, где необходима мультимеризация разных партнеров для повышения авидности этих молекулярных комплексов, превышающей таким образом область иммунологических применений; в диагностическом или терапевтическом плане, а также для продуцирования особых антител, в частности, ограниченных МНС антител. Эти различные и другие применения проиллюстрированы ниже. а) Использование для продуцирования антител. Одно из первых применений экзосом согласно изобретению состоит в продуцировании антител. В самом деле, в соответствии с определенным составом экзосом согласно изобретению, можно получать антитела определенной специфичности. Кроме того, как это показывают примеры, зкзосомы согласно изобретению обладают очень сильными иммуногенными свойствами, в частности, в соответствии с высокой плотностью комплексов СМН-пептид на их поверхности, их функциональностью и их эффективной презентацией в иммунной системе. Таким образом продуцированные антитела могут быть поликлональными или моноклональными. Они могут быть получены классическими методами иммунологии, включающими иммунизацию животных и отбор сывороток(полигенные антитела) и/или слияние селезеночных лимфоцитов с клетками миелом, непродуцирующими иммуноглобулины (для генерирования продуцирующих монокло нальные антитела гибридом). Следующий предмет изобретения относится, следовательно, к антителам или фрагментам антител, продуцируемым путем иммунизации экзосом, таких, как описанные выше. Фрагментами антител могут быть, например, Fab,(Fab')2, ScFv, и т.д., и, вообще, любой фрагмент,сохраняющий специфичность антитела. В особенности изобретение относится к способу получения антител, включающему иммунизацию животного с помощью экзосомы, такой, как указанная выше, несущей определенный комполекс пептид-СМН, и рекуперацию антител и/или клеток, продуцирующих антитела или вовлеченных в иммунный ответ. Способ согласно изобрете 004428 20 нию преимущественно позволяет получать моноклональные, в частности, ограниченные СМН, антитела, то есть специфичные к ассоциации СМН-пептид. Согласно предлагаемому в изобретении способу, предпочтительно используют экзосомы, по существу лишенные эндогенных молекул СМН, которые экспрессируют рекомбинантные комплексы СМН-пептид и которые получены из клетки, аутологичной по отношению к животному, которое было иммунизировано. Таким образом, как показано в примерах, этот способ позволяет получать, без необходимости стимулятора, многочисленные антитела , направленные против пептида, в частности, ограниченные СМН антитела, то есть специфичные к пептиду в его конформации,ассоциированной с определенной молекулой СМН. Такие антитела особенно предпочтительны в экспериментальном, диагностическом и терапевтическом плане. Кроме того, антитела согласно изобретению могут быть маркированы любым известным (ферментативным, флуоресцентным, радиоактивным и т.д.) способом, согласно известным специалисту методам.b) Диагностические применения. Экзосомы и антитела согласно изобретению обладают полезными свойствами для диагностического использования. Так, полученные согласно изобретению антитела или фрагменты антител могут быть использованы для любого диагностического применения, для детекции в биологическом образце присутствия соответствующих специфичных антигенов, благодаря применению различных классических методов, таких, как, например, проточная, цитометрия, иммуногистохимия или иммунофлуоресценция. В особом случае ограниченных МНС антител, они позволяют преимущественно детектировать соответствующие комплексы СМН-пептиды и, следовательно, осуществлять диагностику соответствующих патологий. Эти антитела, в частности,могут быть использованы в диагностике патологий, причастных к отсутствию ответа или несоответствующему ответу иммунной системы, для определения экспрессии предварительно определенного антигена в форме, распознаваемой Тлимфоцитами. Например, и не исчерпывающим образом, можно осуществлять диагностику: опухолевых патологий, где детекция в опухолевых образцах различных пептидов, происходящих от протеинов, как р 53, Неr2, MAGE,BAGE, Mart, GP100, ассоциированных с молекулами класса I СМН, может позволять осуществлять фенотипирование опухоли и облегчать выбор терапии; вирусных заболеваний на прединфекционной или латентной стадии, где вирион не может быть определен (гепатит, инфекция ВИЧ, вируса огуречной мозаики (CMV) и других вирусов); аутоиммунных заболеваний, как рассеянный склероз, аутоиммунный диабет, аутоим 21 мунный тиреоидит, ревматоидный артрит, системная красная волчанка, где детекция молекул СМН, связанных с происходящими от аутоантигенов пептидами, может служить сигналом перед ускорением эволюционной вспышки заболевания. Экзосомы согласно изобретению также можно использовать для детекции специфичных к протеиновой молекуле партнеров в биологическом образце. Так, экзосомы согласно изобретению, несущие комплексы СМН-пептиды,можно использовать для детекции специфичных к этим комплексам Т-лимфоцитов в биологических образцах, например, в различных патологических ситуациях, в частности, в случае вышеуказанных патологий. В этом отношении,экзосомы могут быть маркированы с помощью известной специалисту любой системы для маркировки (ферментативной, флуоресцентной,радиактивной, и т.д.), чтобы можно было их детектировать в биологических образцах. Согласно особому варианту, изобретение,следовательно, относится к применению маркированных, в частности, флуоресцентных, экзосом, таких, как описанные выше, для детекции Т-лимфоцитов, специфичных к комплексам антигенный пептид-СМН в биологическом образце. Биологическим образцом может быть любой образец крови, сыворотки, ткани, опухоли, биопсии, кожи, мочи и т.д. Кроме того, биологический образец может быть предварительно обработан, например, для отделения клеток, амплификации клеток путем культивирования, получения мембранных фракций и т.д. Биологический образец преимущественно происходит из человеческого организма. В этом отношении,изобретение относится также к способу детекции наличия Т-лимфоцитов, специфичных к комплексам антиген-СМН, в биологическом образце, включающему введение в контакт вышеуказанного образца с маркированной экзосомой, такой, как указанная выше, содержащей вышеуказанный комплекс антиген-СМН, и обнаружение маркировки Т-лимфоцитов в вышеуказанном образце. Более того, детекция этих Т-лимфоцитов позволяет осуществлять не только обнаружение и, следовательно, диагностику физиопатологического состояния, но и также следить, например, за эффективностью протоколов иммунизации и состоянием иммунного ответа в различных стадиях заболевания и оценивать таким образом эффективность терапевтических воздействий. Согласно особому применению, экзосомы согласно изобретению, несущие рецептор TcR,используют для детекции специфичных к этому рецептору комплексов пептид-СМН в биологическом образце. Кроме того, флуоресцентные экзосомы согласно изобретению, несущие любой тип протеина определенного состава, также представ 004428 22 ляют собой флуоресцентные зонды, позволяющие обнаруживать потенциальные рецепторы. Таким образом, новая область использования экзосом может быть широко применена для обнаружения in vivo любого взаимодействия протеин/протеин со слабым сродством. Следовательно, предметом изобретения также является применение экзосом, предпочтительно маркированных, в частности, флуоресцентных, таких как описанные выше: для детекции специфичных к протеиновой молекуле рецепторов в биологическом образце. Согласно этому варианту осуществления, используемые экзосомы, следовательно, содержат на своей поверхности вышеуказанную биологическую молекулу определенной структуры; для детекции наличия лиганда в биологическом образце. Согласно этому варианту осуществления, используемые экзосомы, следовательно, содержат на своей поверхности специфичный к вышеуказанному лиганду рецептор. с) Терапевтические применения. Ограниченные антитела или фрагменты этих антител потенциально способны ингибировать взаимодействие между рецептором одного из Т-лимфоцитов и комплексом СМН-пептид, к которому он специфичен. Параллельно, экзосомы, несущие на своей поверхности один тип комплекса СМН-пептид, за счет взаимодействия со специфичными к этим комплексам Тлимфоцитами, могут вступать в конкуренцию с их природными лигандами, Т-лимфоцитами, и вызывать их инактивацию. Ограниченные антитела и экзосомы, следовательно, могут быть использованы в любых ситуациях, где хотят уменьшить или ликвидировать иммунный ответ, опосредованный Тлимфоцитами, который оказывается пагубным для организма, как это имеет место, например, в случае трансплантаций органов или пересадки костного мозга, при которых стремятся нейтрализовать ответ хозяина против трансплантата обычно с помощью сильных доз иммуносупрессоров; аутоиммунных заболеваний или вирусных патологий, во время которых иммунный ответ ТCD8 или CD4 постоянно приводит к разрушению ткани; аллергий и астмы. В случае этого типа патологий, экзосомы согласно изобретению, экспрессирующие на своей поверхности определенный комплекс пептид-МНС, вовлечение которого в развитие патологического состояния известно, следовательно, могут быть использованы для блокирования развития иммунного ответа и, следовательно, развития патологического ответа. Экзосомы согласно изобретению, несущие комплексы СМН-пептиды, также могут быть использованы для амплификации (экспансии) exvivo популяции цитотоксических Т-лимфо 23 цитов. Используемые непосредственно из образца крови, они также могут быть базой клеточных терапий против различных клетокмишеней. Таким образом, экзосомы могут служить для отбора Т-клеток, специфичных к различным составам комплексов, экспрессируемых клетками, которые представляют собой терапевтическую мишень, как опухолевые или инфицированные вирусом клетки. Другим предметом изобретения, следовательно, является применение вышеописанных экзосом для клональной амплификации и/или стимуляции цитотоксических и/или комплементарных лимфоцитов. Изобретение также относится к способу клональной амплификации (или экспансии) ex vivo Тлимфоцитов, особенно цитотоксических Тлимфоцитов, включающему введение в контакт содержащего Т-лимфоциты биологического образца с экзосомами, такими, как описанные выше, включающими определенный комплекс пептид-СМН; рекуперацию специфичных Тлимфоцитов и их амплификацию. Этот способ особенно предпочтителен для клональной амплификации цитотоксических Т-лимфоцитов,специфичных к комплексам между молекулами СМН и пептидами опухолевых или вирусных антигенов. Другое, особенно представляющее интерес применение везикул согласно изобретению состоит в переносе молекул к клеткам. В самом деле, в силу своего состава, везикулы согласно изобретению способны играть роль вектора переноса молекул к клеткам in vitro, ex vivo и invivo. В этом отношении, изобретение относится к применению экзосом, таких как описанные выше, содержащих представляющее интерес вещество, для получения композиции, предназначенной для переноса вышеуказанного вещества в клетку. Речь идет преимущественно об экзосоме, содержащей на своей поверхности рецептор лиганда или лиганд рецептора, позволяющей таким образом направлять перенос к выбранной клеточной популяции или нескольким выбранным клеточным популяциям. Изобретение также относится к способу переноса вещества в клетку in vitro, ex vitro или in vivo,включающему введение в контакт вышеуказанной клетки с содержащей вышеуказанное вещество везикулой согласно изобретению. Более предпочтительно,используемая везикула зкспрессирует, кроме того, рецептор лиганда, и способ согласно изобретению позволяет осуществлять направленный перенос вещества к экспрессирующим сооответствующий лиганд клеткам. Для осуществления способа in vivo, везикулы согласно изобретению вводят пациенту(предпочтительно млекопитающему, в частности, человеку) любым классическим путем введения(внутривенная,внутриартериальная,внутримышечная, подкожная и т.д. инъекция). Для использования способа in vitro или ex vivo,клетки вводят в контакт путем инкубации в со 004428 24 ответствующем приспособлении (чашка, бутыль, мешочек, ампула и т.д.), предпочтительно в стерильных условиях. Параметры стадии контакта (количество везикул, продолжительность контакта, температура, среда, и т.д.) могут быть легко установлены специалистом в зависимости от преследуемых целей и технической инструкции настоящей заявки.d) Применения в области исследования. Эти применения, разумеется, относятся ко всем вышеуказанным использованиям для анализа молекулярных механизмов антигенной презентации путем использования антител, позволяющих детектировать и анализировать различные стадии образования комплексов СМНпептиды в различных нормальных или патологических ситуациях. Кроме того, они относятся также к анализу и молекулярной характеризации популяций Тлимфоцитов, способных распознавать определенный комплекс СМН-пептид, путем использования флуоресцентных экзосом в отношении их способности детектировать Т-рецепторы к комплексам СМН-пептид. При этих различных (диагностических, терапевтических, экспериментальных, для продуцирования Т-лимфоцитов, и т.д) применениях экзосомы согласно изобретению могут быть использованы либо в чистом виде, либо в иммобилизованной на носителе форме. Так, представленные в примерах результаты в самом деле показывают, что экзосомы можно фиксировать на носителях без ухудшения их функциональных свойств, в частности, например, их антигенной специфичности. В этом отношении, особым предметом настоящего изобретения является композиция, включающая иммобилизованные на носителе экзосомы. Носителем предпочтительно является твердый или полутвердый носитель типа шарика, фильтра или аналогичных носителей. Речь идет предпочтительно о носителе из пластмассы полимерного типа, например, о шариках из латекса или магнитных шариках. Разумеется, может быть использован любой другой синтетический или биологический материал, если он не вызывает существенного ухудшения качеств экзосом или клеток. Преимущественно используют шарики диаметром 1-10 мкм, например 2-5 мкм. Иммобилизации экзосом на носителях преимущественно достигают за счет ковалентной связи, например,путем активации альдегидом или любым другим реагентом для химического связывания. Как правило, иммобилизацию экзосом осуществляют путем инкубации экзосом с носителем в растворе, в условиях, позволяющих осуществлять фиксацию, затем носители извлекают путем центрифугирования. Таким образом полученные функционализированные носители могут быть использованы для охарактеризовывания экзосом или для детекции или амплификации in vitro Т 25 лимфоцитов, как это будет подробно описано в экспериментальной части. Другие аспекты и преимущества настоящего изобретения показаны в нижеследующих примерах, которые нужно рассматривать как показательные и не ограничивающие объема охраны изобретения. Кроме того, все цитированные в заявке публикации включены в настоящую заявку в виде ссылки. Пояснения к фигурам Фиг. 1 - продукция функциональных комплексов СМН-пептид DRI-HA в линии RDL2H3.A. Анализ экспрессии на поверхности путем проточной цитометрии молекул х человеческого СМН класса II DR1 до (слева) и после-цепи DR1, в линии RBL 2H3. Молекулы DRI детектированы с помощью антитела L243 (заштриховано черным), которое в свою очередь обнаруживают с помощью связанной с флуоросцеинизотиоцианатом (FITC) козьей сыворотки против мышиного IgG.B. Экспрессия на поверхности DR1 в линии, экспрессирующей инвариантную цепь(liHA), где пептид СLIP заменен пептидом 308319, происходящим от гемагглютинина вируса гриппа. Молекулы DR1 детектированы в линииDR1-HA Т-лимфоцитов с помощью линии RBL,экспрессирующей этот комплекс или B-EBV того же гаплотипа. Линии RBL DRlliHA и BEBV Hom2 разводили в культуральных планшетах специфичным к комплексу DR1 НА Тлимфоцитом. Линия B-EBV Hom2 также была инкубирована в присутствии насыщающей концентрации (10 мМ) пептида НА. Продукция IL2 в супернатантах культуры позволяет оценивать стимуляцию ТНА-лимфоцитов (специфичных к пептиду НА Т-лимфоцитов). IL2 определяют посредством теста включения меченого тритием тимидина в линию CTLL2, пролиферация которой является IL2-зависимой.D. Анализ насыщения лептидом НА линииRBL DR1 liHA. Клетки Ноm2 и RBL DR1 1iНА были инкубированы (100 клеток на лунку) в присутствии возрастающих концентраций пептида НА и ТНА-лимфоцитов. Стимуляция лимфоцитов была оценена как указано выше. Фиг. 2. Аккумуляция молекул DR1 в секреторном компартменте RBL 2 Н 3.A. Анализ внутриклеточного сайта аккумуляции молекул DR1 в RBL 2H3. Клетки RBLDRLHA были фиксированы с помощью 0,5% глутарового альдегида, затем сделаны проницаемыми с помощью 0,05% сапонина. Молекулы DR1 и инвариантная цепь были детектированы, соответственно, с помощью антител L243 иPIN1, затем связанной с FITC ослиной сыворотки против мышиного IgG. Серотонин был детектирован благодаря специфичной кроличьей сыворотке, обнаруживаемой с помощью связанной с техасским красным (Texas red) ослиной сыворотки против кроличьего IgG. Изображения были получены с помощью кофокальной микроскопии(Leica). Толщина плоскостей среза составляла 0,5 мкм.B. Очистка экзосом RDL DR1liHA. После промывки в модифицированной по способу Дульбекко среде Игла (DMEM) клетки были инкубированы в течение 30 мин в присутствии 1 ммоль иономицина при температуре 37 С. Экзосомы были очищены из супернатанта клеток путем дифференциального центрифугирования. Полученный после центрифугирования осадок экзосом, суспендированный в забуференном фосфатом физиологическом растворе (PBS),был подвергнут разделению (5 мг) путем электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), затем перенесен на мембрану из нейлона. -Цепь DR1 была детектирована с помощью монокловального антитела IB5 в экзосомном препарате и в качестве контроля в лизатах клеток RDLDRLHA и Ноm2(эквивалент 105 клеток на лунку), мигрировавших в тех же самых условиях. Фиг. 3. Использование экзосом для продукции антител к DR1 НА.A. Возрастающие разведения сывороток мышей, иммунизированных с помощью экзосом, были инкубированы с клетками RBL, экспрессирующими (справа) или нет (слева) молекулы DR1 НА. Таким образом полученная маркировка была проанализирована путем проточной цитометрии.B. Сыворотки крыс (разведенные в соотношении 1/100), иммунизированные экзосомами, были инкубированы с клетками RBL, экспрессирующими или нет (слева) молекулы DR1 НА. Справа экспрессирующие DR1 клетки были предварительно инкубированы или нет в течение двух часов при температуре 37 С с 10 ммоль пептида НА, затем, при том же самом разведении, сыворотки иммунных крыс.C. Селезенку иммунной крысы подвергали слиянию с линией Х 63 А 8 в классических условиях получения моноклональных антител. Супернатант различных гибридом был тестирован путем иммунофлуоресценции при использовании клеток RBL2H3, экспрессируюяцих или нет молекулы DR1 или DR1 НА. Клоны а 40, b82 и а 15 являются типичными примерами полученных антител. Фиг. 4. Использование экзосом для детекции специфичных к комплексу DR1 НА Тлимфоцитов. 27 лизосомных компартментах клеток) в течение 30 минут при температуре 37 С, затем были промыты и снова инкубированы в тение часа при температуре 37 С в отсутствие флуоресцентного маркра. Клетки были фиксированыB. Параллельно, экзосомы DR1 НА были очищены из описанных в п.А клеток. Имеющаяся в образцах флуоресценция была определена количественно с помощью флуориметра и прямо визуализирована с помощью кофокальной микроскопии.C.D. Флуоресцентные экзосомы DR1 НА были инкубированы, в концентрациии 50 мг/мл,с ТНА-лимфоцитами, специфичными к комплексу DR1 НА, или ТН 30-лимфоцитами, специфичными к другому комплексу (D), в течение двух часов при температуре 37 С в присутствии азида для блокирования интернализации. Флуоресценция клеток была оценена путем проточной цитометрии. Фиг. 5. Продукция экзосом, несущих молекулы класса II СМН.A. Экспрессию молекул класса II IАb детектировали с помощью моноклонального антитела Y3P и анализировали путем проточной цитометрии. Полученные в клетке RBL2H3 трансфектанты экпрессируют подобные количества молекул класса II, распознаваемых антителомB. Анализ путем вестерн-блотирования экспрессии молекулы IАb в RBL. 10 мг Клеточного лизата и препарата экзосом, происходящих из клетки RBL IAbIi, были проанализированы путем вестерн-блотирования с кроличьей сывороткой, специфичной к цитоплазматической области -цепи молекулы IA.C. Анализ путем проточной цитометрии композиции экзосом. Шарики из латекса были покрыты либо фетальной телячьей сывороткой(FCS), либо экзосомами, происходящими из клеток RDL 2H3 (exos RBL), либо трансфектантом этой клетки с мышиными молекулами класса II IAbIi (exos IAbIi), либо человеческимDR1IiHA (exos DR1IiHA). Молекула CD63 крысы была детектирована с помощью антителаAD1, молекулы IАb - с помощью антитела Y3P,тогда как молекулы DR1 были детектированы с помощью антитела L243. Эти различные антитела были обнаружены с помощью вторичных антител, связанных с фикоэритрином. Фиг. 6. Морфологическая характеризация продуцируемых RBL-2H3 экзосом. А. Трансфицированные с помощью HLADR1 клетки RBL-2H3 были фиксированы с помощью параформальдегида. Ультратонкие срезы в замороженном состоянии были получены и иммуномаркированы с помощью поликлональных антител, направленных против молекулHLA-DR. Эти антитела были визуализированы с помощью протеина А, связанного с частицами 28 коллоидального золота размером 10 нм. Молекулы класса II были детектированы по существу в компартментах, заполненных мембранами везикулярного вида. Риска: 250 нм.B.C. Морфологическая характеризация экзосом, секретированных клетками RBL-2H3. Экзосомы фиксировали с помощью 2% параформальдегида в 0,2 М буфере, рН 7,4, (буфер РВ) и помещали на сетки электронного микроскопа, покрытые пленкой из углеродсодержащего формвара. Экзосомы либо делали контрастными и покрывали оболочкой в 4% растворе уранилацетата и метилцеллюлозы (b), либо иммуномаркировали с помощью антител, направленных против молекул класса II перед нанесением покрытия (с). Как показано на фигуре (а),антитела визуализировали с помощью протеина А, связанного с частицами коллоидального золота размером 10 нм. Риски: 250 нм. Фиг. 7. Манипуляция с внутренним составом экзосом, введение рекомбинантного протеина.A. Экспрессию молекул класса II DR1 детектировали с помощью моноклонального антитела L243 и анализировали путем проточной цитометрии. Трансфекция молекул в клеткеRBL2H3 индуцирует экспрессию подобных количеств молекул класса II, распознаваемых антителом Y3P, В-лимфома которого контролирует В 414.B. Анализ путем вестерн-блотирования экспрессии молекулы DR1 GFP в RBL. 10 мкг Клеточного лизата и 20 мкг препарата экзосом, происходящих из клетки RBLDR1 GFP, были проанализированы путем вестерн-блотирования с парой моноклональных антител, специфичных к GFP.C. Анализ путем проточной цитометрии состава экзосом. Шарики из латекса были покрыты либо фетальной телячьей сывороткой(FCS), либо экзосомами, происходящими из клеток RBL DR1 GFP. Молекулы DR1 детектировали с помощью антитела L243 и вторичных антител, связанных с фикоэритрином, тогда как наличие GFP детектировали прямо в каналеFL1. Фиг. 8. А. Связывания флуоресцентных экзосом специфичными Т-клетками. Получаемые из клеток RBL DR1 IiНА экзосомы, меченые зеленым клеточным маркером, были инкубированы в присутствии двух типов Т-клеток: ТНА,которые обладают TCR, специфичным к комплексам YLA-DR1/HA, и T-Jurkat дикого типа,лишенными такого рецептора. Флуоресцентные экзосомы были инкубированы в течение 3 ч при температуре 37 С с двумя линиями Тлимфоцитов, затем получающиеся в результате маркировки анализировали в клеточном сортере с возбуждением флуоресценции (FACS).B. Маркировка зкзосом с помощью специфически маркированных Т-клеток. Были осуществлены такие же маркировки, что и в п. А, но 29 связывание экзосом (без маркировки зеленым клеточным маркером) с Т-клетками детектировали с помощью моноклонального мышиного антитела AD1 против крысиного CD63, обнаруживаемого затем с помощью ослиных антител против мышиного IgG, маркированных фикоэритрином (Jackson Immuno Research, WestGrove, PA). Такая же маркировка была реализована параллельно в случае Т-клеток, не контиктировавших в экзосомах.C. Стимуляция лимфоцитов экзосомами. Очищенные экзосомы из клеток DR1 GFP были подвергнуты перекрестному связыванию с шариками из латекса, полученными таким же образом, как и для проточной цитофлуориметрии,но промытых полной средой. Каждый осадок шариков был обработан 100 мкл среды, 50 мкл было помещено в первую лунку 96-луночного планшета и 50 мкл было разведено дважды в два раза. Т-Клетки (T-Jurkat и THA1.7) были доведены до концентрации 106 клеток/мл и были внесены по 50 мкл на лунку в присутствии или в отсутствие пептида НА 307-319 в количестве 5 мкмоль. Культуральный планшет был помещен в инкубатор (37 С, 5% СО 2, Н 2 О) на 20 ч, затем супернатант был отобран и концентрация IL2 была оценена с помощью теста CTL.L2. Фиг. 9. Характеризация клеток и экзосом НМС-1.B. Анализ экзосом НМС-1, связанных с шариками из латекса, путем проточной цитофлуориметрии: полностью заштриховано темным: одни шарики из латекса+экзосомы НМС-1; полностью незаштриковано: одно антитело + антимышь-FITC; частые пунктирные линии: контрольные шарики из латекса-SVF + специфическое антитело + антимышь-FITC; редкие пунктирные линии: шарики из латекса-зкзосомы+ специфическое антитело + антимышь-FITC. С. Анализ путем вестерн-блотирования лизата клеток HМС-1 по сравнению с их экзосомами; 10 или 3 мкг протеинов на лунку; НС 10: супернатант в 1/10 е; 1 В 5: супернатант в 1/10 е;CD63: 5 мкг/мл; Lamp 1: 2 мкг/мл; Н 68.4: супернатант в 1/10 е. Экзосомы оказываются обогащенными CD63, Lampl/ TfR. Отсутствие молекул СМН класса II как в лизате, так и в экзосомах подтверждает анализ путем цитофлуориметрии. Доля СМН класса I оказывается идентичной для клеточного лизата и экзосом. 30 Используемые для продукции экзосом в экспериментальной части клетки представляют собой мышиные или человеческие мастоциты. В частности, была использована опухолевая линия базофилов с фенотипом слизистых мастоцитов,обозначаемая RBL-2 Н 3 (Barsumian и др., Eur. J.Immunol, 11, 317 (1981, а также линия незрелых человеческих мастоцитов (НМС-1). Могут быть использованы другие мастоциты, в особенности составляющие линию, такие, как линии, происходящие от клеток RBL (базофильный лейкоз крысы), депонированные в АТСС под номером CRL1378 (Kulczycki и др., J. Exp.Med., 139, 600 (1974. Также были использованы Т-лимфоцитарные линии, способные распознавать особый антиген в структуре человеческого СМН IIJurkatt, транс-фицированная с помощью кДНК,кодирующей рецептор Т-клеток ("Т-НА"), специфичный к пептиду 306-318 гемагглютинина вируса гриппа, в ассоциации с HLA-DRI (Sidhu и др., J. Exp. Med., 176, 875 (1992. Линия человеческих В-клеток, трансформированных вирусом Эпштайна-Барра (линия Ноm-2) была использована в качестве контроля для ограниченного HLA-DR1 ответа. Клетки были культивированы в средеRPMI, дополненная 10% фетальной телячьей сыворотки (Sigma), 1 мг/мл пенициллинастрептомицина, 1 мг/мл глутамина, 5 ммоль пирувата натрия и 50 мкмоль меркаптоэтанола. Разумеется, может быть использована любая другая среда, приспособленная для культивирования эукариотических клеток, особенно клеток млекопитающих. Клетки главным образом культивировали в культуральном флаконе объемом 25 см 3 или 15 см 3. Клетки RBL-2H3, являющиеся сросшимися клетками, отделяли от носителя благодаря воздействию трипсинаэтилендиаминтетрауксусной кислоты (ЭДТК) (Seromed). Для получения этих последних в больших количествах также можно их культивировать в "центрифуге" при плотности 106 клеток/мл. Плазмиды Для генетической модификации мастоцитов были реализованы следующие генноинженерные конструкции. Были выделены кДНК, кодирующие человеческую -цепь HLA-DR1 (Larhammer и др.,Cell, 30, 153 (1982, человеческую -цепь HLADR1 (Bell и др., PNAS, 82, 3405 (1985 и человеческую инвариантную цепь р 33li (Ciaesson и др., PNAS, 80, 7395 (1983. кДНК, кодирующая инвариантную цепь p331i, затем была модифицирована с помощью полимеразной цепной реакции (PCR) для замены кодирующей пептидCLIP (остатки 87-102) области сайтом рестрикции. Эта кДНК позволяет таким образом 31 встраивать, вместо пептида CLIP, любой, представляющий интерес фрагмент кДНК, кодирующей антигенный пептид (Stumtner и др.,EMBO J., 16, 5807 (1997. В конкретном примере, фрагмент ДНК, кодирующий пептид НА 308319 гемагглютинина вируса гриппа, был встроен в эту кДНК, кодирующую химерный полипептид li HA308-319). Вышеуказанные нуклеиновые кислоты затем были клонированы раздельно в плазмидepSRct под контролем промотора SR (Takebe и др., Mol. Cell Bio, 8, 466 (1988. Каждая из плазмид затем была модифицирована для встраивания другого гена устойчивости, позволяющего осуществлять селекцию для каждой из плазмид, и, следовательно, для каждой из цепей: цепь с геном устойчивости к неомицину; цепь с геном устойчивости к гигромицину; и инвариантная цепь с геном устойчивости к зеоцину. Трансфекции Для введения различных нуклеиновых кислот в мастоциты, соответствующие плазмидные векторы были линеаризованы с помощью фермента рестрикции Scal. 50 мкг каждой плазмиды были линеаризованы, затем подвергнуты преципитации в этаноле и осадки после центрифугирования были ресуспендированы в присутствии клеток RBL-2H3 в концентрации 1107 клеток/мл. Стабильные трансформанты были получены путем электропорации 5106 клеток с помощью "gene pulser" (Bio-Rad, Ричмонд, Канада) в следующих условиях: 260 V, 960 F. Спустя 72 ч после электропорации трансфектанты селекционировали путем культивирования в среде для селекции, содержащей 250 мкг/млG418 (Geneticine, Gibco), 1 мг/мл гигромицина и 500 мкг/мл зеоцина. После культивирования в течение 8 дней в среде для селекции были трансфицированы 60-90% присутствующих клеток. Затем был произведен посев трансфектантов в чашке Петри в среду для селекции в концентрации, позволяющей появляться индивидуализированным сросшимся колониям. Таким образом полученные клоны были отобраны и раздельно культивированы. Эти клоны могут быть сохранены в замороженном виде для последующего использования. АнтителаY3P (МНС II (IA) ) представляет собой мышиное моноклональное антитело, распознающее комплекс IAb a (Janeway и др., 1984). Анти-IA представляет собой кроличью сыворотку, направленную против цитоплазматической части -цепи IA. Анти-GFP представляет собой смесь двух моноклональных антител(клоны 7.1 и 13.1), направленную против "флуоресцирующего зеленым протеина", выпускаемого фирмой Boehringer Mannheim. В экспериментах с использованием проточной цитофлуориметрии используемыми вторыми антителамиImmunoresearch Laboratories) . Шарики Шарики из латекса: не содержащий поверохностно-активного вещества белый альдегид/ сульфатный латекс; диаметр (D): 3,9 мкм; Interfacial Dynamics Corp., Portland, Or. USA. Электронная микроскопия Трансфицированные с помощью HLA-DR1 клетки RBL-2H3 были фиксированы с помощью 2%-ного параформальдегида в 0,2 М фосфатном буфере, рН 7,4 (буфер РВ). После фиксации клетки были промыты с помощью 50 мМ раствора глицина в буфере РВ, затем покрыты 10% желатином. После затвердевания блоки были препарированы, введены в 2,3 М раствор сахарозы и заморожены в жидком азоте. Были получены ультратонкие срезы в замороженном состоянии и иммуномаркированы с помощью поликлональных антител, направленных к молекулам HLA-DR. Эти антитела визуализированы с помощью протеина А, связанного с частицами коллоидального золота размером 10 нм. Экзосомы фиксировали с помощью 2%ного параформальдегида в 0,2 М фосфатном буфере, рН 7,4 (буфер РВ) и помещали на сетки электронного микроскопа, покрытые пленкой из углеродсодержащего формвара. Экзосомы либо делали контрастными и покрывали защитной оболочкой в растворе 4% уранилацетата и метилцеллюлозы, либо перед покрытием оболочкой иммуномаркировали с помощью антител, направленных против молекул класса II. Антитела визуализировали с помощью А-протеина, связанного с частицами коллоидального золота размером 10 нм. Результаты 1. Продукция экзосом DR1 НА. 1.1. Конструкция и характеризация генетически модифицированных продуцирующих клеток. С целью продуцирования контролируемым образом экзосом, несущих комплексы СМНпептид определенного состава, - и -цепи молекул класса II СМН, DR1, были экспрессированы в мастоцитарной 1 о линии RBL2H3, происходящей от базофильного лейкоза крысы. Для этого, два вектора, несущие, соответственно,кодирующую каждую цепь нуклеиновую кислоту, под контролем промотора SRa были одновременно трансфицированы в клетки (см. Материалы и методы). Полученные путем проточной цитометрии результаты показывают, что трансфицированные клетки хорошо экспрессируют молекулы DR1 (фиг. 1 А). Эти клетки RBL-2H3 затем были сенсибилизированы данным пептидом точного состава,чтобы генерировать комплексы СМН-пептид определенного состава. Для этого могут быть предусмотрены различные способы. Согласно 33 одному способу реализации, пептид может быть инкубирован непосредственно с экзосомами. Согласно другому варианту, в клетки может быть введена кодирующая пептид нуклеиновая кислота, чтобы также экпрессировать этот петид. В этом отдельном примере осуществления,для получения антигенпредставляющей клетки,включающей одну антигенную специфичность,выбранный антигенный пептид был введен в клетки в виде слитого путем генной инженерии с инвариантной человеческой цепью 1i. Более предпочтительно, пептид СLIP инвариантной цепи был заменен последовательностью выбранного пептида, происходящего от гемагглютинина (НА 308-319) вируса гриппа, способного связываться с молекулой DR1. Эта конструкцияDR1 гибридная цепь позволила сконструировать клетки, которые экспрессируют молекулы DR1,распознаваемые антителом L243, в подобном количестве, как и контрольная линия B-EBV(Ноm2) гаплотипа DR1 (фиг. 1 В). Эти результаты, следовательно, показывают, что мастоциты согласно изобретению хорошо экспрессируют человеческий функциональный комплекс пептид-СМН предопределенного и контролируемого состава. Функциональный характер комплексов пептид-СМН, экспрессированных клетками согласно изобретению, был подтвержден в тесте стимуляции Т-лимфоцитов, специфичных к комбинации DR1-HA, которую несут эти клетки. Для этого клетки согласно изобретению были инкубированы в присутствии ТНАлимфоцитов и стимуляция была детектирована путем определения в супернатанте высвободившегося интерлейкина-2 с помощью теста роста IL-2-зависимой клеточной линии. В качестве контроля была использована линия Влимфоцитов, трансформированная с помощьюEBV (Hom-2), гаплотипа DR1, вытесняемая насыщающей концентрацией (10 ммоль) пептида НА. Полученные результаты представлены на фиг. 1C и 1D. Они показывают, что мастоциты согласно изобретению экспрессируют комплексDR1-HA, способный стимулировать специфичный к этой комбинации Т-лимфоцит. Они также показывают, что достигаемая в присутствии клеток согласно изобретению стимуляция является более эффективной, чем таковая, достигаемая с помощью контрольных клеток (B-EBV гаплотипа DR1), вытесняемых насыщающей концентрацией (10 ммоль) пептида НА. Наконец, полученные результаты показывают, что молекулы DR1 по-видимому презентируют пептид НА, так как добавление насыщающей концентрации пептида не увеличивает заметным образом способности клеток RBL DR1 liHA стимулировать ТНА-лимфоциты (фигура 1D). 34 Совокупность этих результатов, следовательно, свидетельствует о функциональном характере продуцированных комплексов пептидСМН. Они также иллюстрируют специфичекий характер полученных клеток и, следовательно,специфический характер способа согласно изобретению, который позволяет получать клетки(и экзосомы), несущие молекулы определенного и контролируемого состава. 1.2. Продукция функционализированных экзосом. Изучение путем иммунофлуоресценции позволило показать, что рекомбинантные комплексы СМН-пептид (DR1 НА) аккумулируются в гранулах секреции клеточной линии RBL-2H3. На фиг. 2 А в самом деле показана солокализация молекул DR1 с серотонином во внутриклеточных везикулярных структурах. Следовательно, была изучена возможность того, что функциональные экзосомы могут быть высвобождены из этих клеток. Для этого клетки культивировали в присутствии кальцийионофора и наблюдали за продукцией мембранных везикул. Более предпочтительно, клетки были центрифугированы при ускорении 300g в течение 5 мин при комнатной температуре. К каждому осадку клеток после центрифугирования было добавлено примерно 300 мкл раствора кальций-ионофора (1 ммоль иономицина) и инкубация была продолжена в течение 30 мин при температуре 37 С. Экзоцитоз был прекращен путем быстрого охлаждения на льду и добавления 300 мкл 1 мМ холодного раствора ЭДТК в забуференном фосфатом физиологическом растворе. Клетки затем были подвергнуты центрифугированию при ускорениии 300g при температуре 4 С в течение 5 мин. Супернатанты были извлечены и снова подвергнуты центрифугированию сначала в течение 5 мин при ускорении 1200g, затем в течение 5 мин при ускорении 10000g, и, наконец, потом в течение 1 ч при ускорении 70000g. После этого дифференциального центрифугирования осадки после центрифугирования (содержащие экзосомы) извлекали и солюбилизировали в буферном растворе (30 мкл буфера Laemmli-DDT (1X или 2 Х. Часть осадков после центрифугирования также солюбилизировали в лизирующем буфере для определения концентрации протеина. Растворы экзосом могут быть разделены путем миграции на геле ( 12% полиакриламидный минигель) при 20 мА ,затем перенесены на устройство Иммобилен. После этого проводили анализ экзосом путем вестерн-блотирования с антителами, специфичными к разным цепям молекул класса II СМН. Полученные результаты показывают, что экзосомы могут быть в значительной степени высвобождены из линии RBL-2H3 после стимуляции соответствующим агентом. Эти экзосомы могут быть выделены и очищены, например,путем дифференциального ультрацентрифугирования для получения композиции из экзосом. 35 Наконец, представленные на фигуре 2 В результаты показывают, что эти экзосомы являются функциональными. В самом деле, анализы путем вестерн-блотирования показывают, что полученные экзосомы экспрессируют разные рекомбинантные цепи молекул класса II СМН. Кроме того, эти результаты показывают также высокую плотность комплексов пептид-СМН на поверхности экзосом согласно изобретению. Нижеследующие примеры поясняют, в частности, использование экзосом DR1 НА для продуцирования специфичных к этой комбинации антител и способность экзосом DR1 НА связывать Т-лимфоциты, специфичные к этой же самой комбинации. 2. Генерирование антител, специфичных к комплексу DR1 НА. Этот пример иллюстрирует использование экзосом согласно изобретению для продуцирования антител, в частности, так называемых"ограниченных" антител, то есть специфичных к антигенному пептиду в ассоциации с молекулой СМН. Этот пример иллюстрирует, в частности,очень значительную иммуногенную способность экзосом согласно изобретению, так как они позволяют осуществлять продукцию антител в отсутствие всякого стимулятора. Очищенные из супернатанта клеток RBLDR1 НА (пример 1) экзосомы были суспендированы в забуференном фосфатом физиологическом растворе (PBS). Эти экзосомы затем были использованы для иммунизации мышей линииBalb/c или крыс линии LOU в отсутствие всякого стимулятора согласно следующим протоколам. Мышам вводили путем инъекции подкожно 10 мкг экзосом, два раза с интервалом в три недели, затем 30 мг интраперитонеально и, наконец, 30 мкг внутривенно за 3 дня до отбора сывороток. Крысам вводили путем инъекции интраперитонеально 10 мкг экзосом, два раза с интервалом в три недели, затем 50 мкг внутривенно за три дня до отбора сывороток. Как показано на фиг. 3 А, взятые у иммунизированных крыс сыворотки обладали очень сильной реактивностью против линии RBL, экспрессирующей или нет комплекс DR1 НА, но которая была детектируема вплоть до разведения свороток в тридцать тысячных только в случае клетки DR1 НА. Как показано на фиг. 3 В, сыворотки таким образом иммунизированных крыс неожиданно проявили реактивность против линии BRL, экспрессирующей комплекс DR1 НА, тогда как те же самые сыворотки реагировали с более слабой интенсивностью с первоначальной линией RBL2H3. Кроме того, добавление пептида НА к экспрессирующим DR1 клеткам (RBL-2H3 DR1) в значительной степени увеличивает реактивность таким образом продуцированных антисывороток (фиг. 3 В). 36 Эти результаты, следовательно, показывают, что экзосомы линии RBL способны индуцировать ответ антитела, которое неожиданно у крысы направлено главным образом к комплексам DR1 НА. Селезенки иммунных крыс были подвергнуты слиянию с клетками линии Х 63 А 8. Таким образом полученные гибридомы были сортированы путем клонального разведения согласно классическим методам иммунологии, затем селекционированы путем иммунофлуоресценции по специфичности продуцированных монклональных антител. Таким образом были получены различные моноклональные антитела, из которых некоторые направлены к протеинам линии RBL, другие - к мономорфическим детерминантам молекул класса II человека гаплотипа DR1, и, наконец, к комплексу, образованному молекулами DR1, ассоциированными с пептидом, происходящим от протеина НА вируса гриппа (фиг. 3 С). Эти последние моноклональные антитела представляют собой ограниченные антитела и обладают, следовательно,особенно благоприятными свойствами для диагностических или терапевтических применений. 3. Детекция Т-лимфоцитов, специфичных к комплексу DR1 НА. Этот пример иллюстрирует использование экзосом согласно изобретению для детекции специфических Т-лимфоцитов в биологическом образце. Этот пример также показывает, как экзосомы могут быть использованы для селекции и амплификации популяции особых Тлимфоцитов, в частности, с целью их повторной инъекции субъекту (клеточная терапия). Этот подход, разумеется, может быть распространен на использование описанных в примере 2 ограниченных антител, а также на детекцию любого рецептора, специфичного к лиганду. Для осуществления этого применения были получены маркированные экзосомы. Для этого, перед очисткой экзосом линии DR1 НА,ее инкубировали с флуоресцентным меченым веществом, очень сильно аккумулирующимся в содержащихся в гранулах секреции экзосомах. Используемый маркер, "Green Tracker" (зеленый клеточный маркер), представляет собой флуоресцентный липид, который аккумулируется в лизосомах клеток. Анализ с помощью кофокальной микроскопии клеток, после фиксации,показывает наличие флуоресцентной маркировки в гранулах секреции клеток (фиг. 4 А). Затем были получены и очищены из этих клеток, в описанных в примере 1 условиях, флуоресцентные экзосомы. Сделанные таким образом флуоресцентными, эти экзосомы (фиг. 4 В) были использованы для детекции в биологическом образце наличия Т-лимфоцитов, специфичных к комбинации DR1 НА (ТНА- лимфоциты). Понятно, что в рамках изобретения может быть использована любая другая маркировка, нане 37 сенная либо на продуцирующие клетки, либо продуцируемые экзосомы. Для этого экзосомы были инкубированы в присутствии образца ТНА-лимфоцитов и ТН 30 лимфоцитов другого комплекса. Полученные путем проточной цитофлуориметрии результаты показывают, что экспрессирующие DR1 НА экзосомы специфически связываются с THAлимфоцитами (фиг. 4 С), тогда как эти же самые экзосомы неспособны распознавать лимфоциты,обладающие другой специфичностью (фиг. 4D). Эти результаты свидетельствуют об исключительной и неожиданной способности экзосом,продуцируемых мастоцитарной линией согласно изобретению, детектировать специфичные к этому же самому комплексу Т-лимфоциты, то применение может быть осуществлено в случае любого типа биологического образца. Кроме того, эта технология может быть применена таким же образом к получению и использованию экзосом, экспрессирующих комплексы СМН I-пептид. Настоящее изобретение позволяет также детектировать на поверхности антигенпредставляющих клеток, даже опухолевых клеток, представление молекулами класса I и класса II СМН пептидов, происходящих от экспрессированных опухолями антигенов. Настоящее изобретение также позволяет детектировать, даже очищать, Т-лимфоциты,способные распознавать эти же самые комплексы. Таким образом они могут делать возможным амплификацию популяции Т-лимфоцитов, специфичных к индивидуальному комплексу пептид-СМН, например, популяцииCTL-лимфоцитов, для их терапевтического использования. Действительно, разработаны различные подходы иммунотерапии, например, в случае раковых заболеваний или вирусных инфекций, базирующиеся на отборе у пациента лимфоцитов и на экспансии ех vivo отдельных клонов Т-лимфоцитов, специфичных к вовлеченному в патологию антигену (например, опухолевый или вирусный антиген). Эти амплифицированные клоны затем снова вводят пациенту в качестве терапевтического агента. Настоящее изобретение позволяет в высокой степени облегчать селекцию и амплификацию специфичных клонов Т-лимфоцитов и, следовательно,потенциал и осуществление этих терапевтических подходов. 4. Продукция экзосом, несущих мышиные молекулы класса II СМН (фиг. 5 А, 5 В). Комплементарные ДНК, кодирующие - и-цепи мышиных молекул класса II гаплотипаIАb, а также мышиную инвариантную цепь, были встроены в эукариотные экспрессирующие векторы NT, в которых транскрипция кДНК находится под контролем промотора SR. Каждая из плазмид, кроме того, несет ген устойчивости к гигромицину (для -цепи IАb), к неомицину (для -цепи IАb) или к зеоцину (для инва 004428 38 риантной цепи). После трансфекции клеток RBL 2 Н 3 путем электропорации (см. раздел "Материалы и методы") клетки были селекционированы на основе их резистентности к трем антибиотикам, затем резистентные клетки были клонированы путем ограниченного разведения и охарактеризованы в отношении экспрессии молекул IAb путем проточной цитометрии при использовании специфичных антител Y3P. На фиг. 5 представлены некоторые из таким образом полученных результатов. Анализ путем проточной цитометрии показывает, что клетка RBL IАbIi распознается антителом Y3P ( специфичным к молекулам IАb) эквивалентно лимфоцитарной линии В В 414, тогда как никакая маркировка не определяется в случае исходной линии RBL (фиг. 5 А). Установленное согласно морфологическому анализу с помощью микроскопии, что мышиные молекулы класса IIIАb, как и человеческие молекулы DR1, аккумулированы в гранулах секреции клетки (не показано), явилось причиной поиска их локализации в экзосомах. Экзоцитоз клеток был прекращен путем добавления 1 ммоль иономицина, таким образом полученные экзосомы были очищены путем дифференциального ультрацентрифугирования (см. пример 1.2.). Анализ путем вестерн-блотирования этих препаратов зкзосом показывает, что они содержат мышиные молекулы класса II СМН, идентичные детектированным в контрольном клеточном лизате (фиг. 5 В). Эти результаты свидетельствуют о том,что человеческие (DR1), а и также мышиные(1 Аb) молекулы класса II могут быть экспрессированы и могут аккумулироваться в экзосомах соответствующих линий RBL 2H3. 5. Морфологическая характеризация экзосом, продуцированных RBL 2H3 (фиг. 6). Наблюдения с помощью электронной микроскопии иммуно-маркированного среза в замороженном состоянии клеток RBL показывают наличие многочисленных внутриклеточных компартментов, заполненных мембранами. Преобладающая часть этих мембран соответствует везикулам, которые заполняют полость компартментов (фиг. 6 А). Трансфицированные в этих клетках молекулы класса II аккумулируются в содержащих внутренние везикулы компартментах и визуализируются, в частности, в ассоциации с мембраной этих везикул (фиг. 6 А). Когда клетки RBL стимулированы таким образом, чтобы индуцировать их дегрануляцию(IgE-антиген или иономицин), внутриклеточные компартменты сливаются с цитоплазматической мембраной. Внутренние везикулы, с которыми ассоциированы молекулы класса II, высвобождаются во внеклеточную среду. Эти везикулы тогда называют экзосомами. Выбранный метод электронной микроскопии для изучения морфологии экзосом, а также содержащихся в них протеинов представляет собой метод "целого гистологического среза(препарата)". Эта методика позволяет визуализировать целые экзосомы, лишенные всякого другого клеточного содержимого. Этот метод также позволяет детектировать с большой эффективностью молекулы, ассоциированные с мембраной экзосом. Используя эту методику,наблюдали, что секретированные клетками RBL экзосомы имеют неоднородный размер от 30 до 120 нм и изменяемую электронную плотность(фиг. 6 В) . Молекулы класса II преобладают в популяции везикул с размером 80-100 нм,имеющей среднюю электронную плотность. Подавляющее большинство этих везикул, обогащенных молекулами класса II, имеет форму тарелки (фиг. 6 С). 6. Иммобилизация экзосом на носителях(фиг. 5 С). В этом примере описывается фиксация экзосом на твердых носителях и показано, что таким образом фиксированные экзосомы сохраняют свои функциональные свойства. Эти новые продукты (экзосомы на носителях) могут быть использованы для характеризации и анализа экзосом или в качестве диагностических или реактивных продуктов, например, для детекции и/или стимуляции in vitro Т-лимфоцитов. Различные препараты экзосом, продуцированных клетками RBL, экспрессирующими или нет человеческие или мышиные молекулы класса II, были инкубированы с шариками из латекса размером 4 микрона, активированными альдегидсульфатом. В частности, экзосомы, очищенные из супернатантов дегрануляции RBL 2H3, были промыты в PBS (центрифугирование со скоростью 50000 об./мин на TLA 100.4 в течение 30 мин). Экзосомы в количестве 30 мкг смешивали с 10 мкл шариков из латекса, предварительно стерилизовали, гомогенизировали,затем инкубировали в течение 10-15 мин при комнатной температуре. Объем шариков затем дополняли до 1 мл с помощью PBS 1 х, затем инкубировали во вращающемся инкубаторе при комнатной температуре в течение двух часов. После этого "перекрестно связанные" с экзосомами шарики: насыщали путем добавления окончательно 100 мМ глицина (30 мин при комнатной температуре); центрифугировали при ускорении 2200g в течение двух минут при температуре 4 С, затем осадок шариков обрабатывали с помощью 1 мл РВS lx 3%SVF 0,01%NaN3. Для использования шариков в цитофлуориметрии: полученный после центрифугирования осадок шариков промывают два-три раза с помощью PBS 1x 3%SVF0,01%NaN3; обрабатывают с помощью 1 млPBS1x0,01%NaN3; используют от 5 до 20 мкл на точку и инкубируют обычным образом с первым, затем со 40 вторым антителом. Считывание осуществляют на приборе Facscalibur (Becton Dickinson). Эта методика позволяет покрывать поверхность этих шариков из латекса экзосомами,полностью сохраняя их структуру. Когда экзосомы находятся на шариках, облегчается манипуляция с ними. В самом деле,они могут быть центрифугированы с низкой скоростью и детектированы методами классической проточной цитометрии. На фиг. 5 С приведено несколько примеров детекции путем проточной цитометрии различных протеинов, входящих в состав экзосом. Активированные альдегидсульфатом шарики из латекса таким образом были инкубированы либо с экзосомами,продуцированными нетрансфицированными клетками RBL 2 Н 3, либо с экзосомами клетокRBL 2H3, экспрессирующих человеческие молекулы класса II DR1 и конструкцию IiHA или мышиные молекулы класса II IАb, либо с фетальной телячьей сывороткой (FCS) в качестве контроля. Таким образом приготовленные шарики из латекса затем были инкубированы с различными моноклональними антителами:AD1, распознающее молекулу CD63 крысы,находится в гранулах секреции RBL 2H3; специфичное к молекулам IАb антитело Y3P; и специфичное к молекулам DR1 антитело L243. Эти различные антитела были обнаружены с помощью вторичных, связанных фикоэритрином антител, затем полученные маркировки были проанализированы путем проточной цитометрии при использовании прибора FacscaliburCD63 несомненно присутствует во всех экзосомах, происходящих из клетки RBL, экспрессирующей или нет молекулы класса II, тогда как покрытые экзосомами IАb шарики из латекса специфически распознаются антителом Y3P и не распознаются антителом L243, тогда как, в противоположность, экзосомы DR1IiHA распознаются антителом L243 и не распознаются антителом Y3P. Ни одно из этих антител не распознает шариков из латекса, покрытых фетальной телячьей сывороткой (фиг. 5 С). Эти результаты показывают, что этот способ позволяет точно и специфически детектировать экспрессию различных протеинов, входящих в состав экзосом. Пример 8, кроме того,показывает, что такие продукты (экзосомы на носителе) также позволяют детектировать или стимулировать пролиферацию специфичных Тлимфоцитов. 7. Манипуляция с составом экэосом (фиг. 7). Экзосомы представляют собой везикулы,ограниченные липидным бислоем, в который введено большое количество молекул, как вышеуказанные молекулы класса II СМН илиCD63. Внутри этих везикул находятся цитоплазматические области вышеуказанных трансмембранных молекул, однако, также раствори 41 мые протеины, происходящие из цитозоля клеток. Для доказательства того, что можно по желанию модифицировать содержимое этих везикул, заявителями в качестве метки был использован "флуоресцирующий зеленым протеин"(GFP). Кодирующая GFP ДНК была подвергнута слиянию с СООН-концом -цепи человеческой молекулы DR1. Эта генно-инженерная конструкция, встроенная в экспрессирующие векторыNT, несущие ген устойчивости к гигромицину,была котрансфицирована в клетку RBL 2H3 с вектором, несущим -цепь DR1 и ген устойчивости к неомицину. Резистентные к этим двум антибиотикам клетки были селекционированы,затем точно отобраны для экспрессии GFP. В частности была реализована (генноинженерная) конструкция, связывающая цитоплазматическую часть DR-цепи (по С-концу) сN-концом GFP. кДНК DR включает сайт PstI в положении 565. Фрагмент размером примерно 200 пар оснований со стороны 3' этой кДНК был амплифицирован путем PCR, исходя из вектораpcDNA3/RSV/DRa, с помощью двух олигонуклеотидов, включающих сайт PstI в 5'и сайт NcoI в 3', из которого, кроме того, удален стоп-кодон. Полученный фрагмент PCR был подвергнут рестрикции с помощью PstI и NcoI и клонированию в тех же сайтах вектора pEGFP N1(Clontech.). Полученная в результате плазмида,рестриктированная с помощью PstI и XbaI, позволила высвободить фрагмент, соответствующий последним 30 аминокислотам DR, за которой следует GFP. Параллельно, плазмидаpcDNA3/ RSV была подвергнута рестрикции с помощью EcoRV/PstI, высвобождающего таким образом фрагмент, соответствующий остаткуDR-цепи (от начала вплоть до сайта PstI). Оба фрагмента затем были сгруппированы, потом клонированы в pcDNA3/CMV между EcoRV иXbaI. Анализ этих клеток (DR1 GFP) путем проточной цитометрии показывает, что клетки,распознаваемые антителом L243, специфичным к молекулам DR1, также излучают зеленую флуоресценцию, детектируемую в канале FL1,тогда как клетки, трансфицированные альфа- и бета-цепями DR1, не включающей GFP, не излучают никакой флуоресценции в канале FL1(фиг. 7 А). Для того, чтобы показать, что GFP действительно содержится в экзосомах клетки RBL 2H3, происходящие из клеток RBL DR1 GFP экзосомы были получены путем дифференциального ультрацентрифугирования, затем проанализированы путем вестерн-блотирования(фиг. 7 В) и проточной цитометрии после "перекрестного связывания" с шариками из латексаDR1 GFP и в ее экзосомах протеин массой 65 42 кДа, соответствующий молекулярной массе бета-цепи DR1, слитой с GFP (фиг. 7 В), тогда как никакого сигнала не было обнаружено в клеточных лизатах клетки RDL 2 Н 3, экспрессирующей или нет только молекулу DR1. Путем сравнения между шариками из латекса, "перекрестно связанными" с фетальной телячьей сывороткой или происходящими из клетки DR1 GFP экзосомами, наблюдают, что только покрытые экзосомами шарики индуцируют флуоресценцию в канале FL1 (FITC) и распознаются антителом L243,специфичным к DR1, детектируемой вторичным, связанным с фикоэритрином антителом. Эти результаты, следовательно, свидетельствуют о том, что можно вводить экзогенный протеин в экзосомы, продуцируемые клеткойRBL 2H3. Эти результаты также показывают,что можно направлять протеин в экзосому путем экспрессии в продуцирующей клетке в форме, получаемой путем слияния с трансмембранной молекулой, такой, как молекула СМН. 8. Функциональная характеризация экзосом (фиг. 8). Этот пример показывает, что экзосомы,продуцируемые клеткой RBL 2H3, несущей молекулы класса II СМН, способны связывать антигенный пептид и стимулировать Т-лимфоцит,экспрессирующий рецептор, специфичный к этому комплексу пептид-СМН класса II. Экзосомы, продуцируемые клетками RBLDR1 IiHA, мечеными зеленым клеточным маркером, были инкубированы в присутствии двух типов Т-клеток: ТНА, которые обладают TCR,специфичным к комплексам HLA-DR1/HA, и TJurkat дикого типа, которые не содержат такого рецептора. Эксперименты в отношении связываний реализовали в 96-луночном планшете с круглодонными лунками в среде RPMI 1640,дополненной 10% фетальной телячьей сыворотки, забуференной с помощью 10 мМ ГЕПЕСа,по 50 мкл окончательно на лунку, 105 Т-клеток на лунку и с изменяемыми количествами флуоресцирующих экзосом, в течение 3 ч при температуре 37 С. Затем осуществляли две промывки в той же самой среде перед анализом обработанных 400 мкл PBS клеток в клеточном сортере с возбуждением флуоресценции. Была реализована гамма-эффективная доза, показывающая, что интенсивность маркировки ТНА пропорциональна используемому количеству экзосом. 108 Клеток RBL DR1 IiHA продуцировали 700 мкл экзосом. Были тестированы количества экзосом от 32 до 2 мкл на лунку. Была принята во внимание маркировка, получаемая с 16 мкл на точку, что соответствует продуцированию экзосом, осуществляемому 2,3 миллионами клеток (результаты не представлены). Полученные результаты показывают, что популяция ТНА маркирована флуоресцентными экзосомами, что дает детектируемую в FL1 путем цитофлуориметрии флуоресценцию, тогда 43 как Jurkats дикого типа не модифицированы(фиг. 8 А). Был реализован такой же тип маркировки,но после связывания экзосом; Т-клетки были инкубированы с моноклональным мышиным антителом AD1 против CD63 крысы, обнаруживаемым затем с помощью ослиных антител против мышиного IgG, маркированных фикоэритрином (Jackson Immuno Research, West Grove,PA) . Такая же маркировка была реализована параллельно в случае Т-клеток, которые не были доступны экзосомам. Наблюдают (фиг. 8 В) ,что только ТНА-клетки, связанные с экзосомами, как свидетельствует об этом их флуоресценция в FL1, также замечены в FL2, что указывает на новое присутствие CD63 крысы на их поверхности. Полученные заявителями результаты, следовательно, показывают, что флуоресцентные экзосомы можно использовать для визуализации путем проточной цитометрии популяций Тлимфоцитов, специфичных к комплексам HLADR/антигенный пептид, которые несут экзосомы. Для оценки способности экзосом стимулировать Т-лимфоцитзависимым от наличия пептида образом, полученные из клетки DR1 GFP экзосомы были подвергнуты перекрестному связыванию с шариками из латекса, затем инкубированы в присутствии клетки лимфоцитарной линии Т Jurkat, экспрессирующей или нет Трецептор, специфичный к комплексу DR1 пептид НА 307-319. Шарики из латекса были приготовлены таким же образом, как и для проточной цитофлуориметрии, но промыты в полной среде (RPMI,10% фетальной телячьей сыворотки, 1% пенициллин-стрептомицин-глутамина,0,1% меркаптоэтанола, 4% пирувата натрия). Для стимуляции Т-лимфоцитов, каждый, полученный после центрифугирования осадок шариков был обработан 100 мкл среды: 50 мкл среды было помещено в первую лунку 96-луночного планшета и 50 мкл среды было разведено дважды в 2 раза. Контроль был осуществлен в соответствии с той же методикой работы с шариками, инкубированными в фетальной телячьей сыворотке. Т-Клетки (Т Jurkat Pasteur и ТНА 1.7) были доведены до концентрации 106 клеток/мл и были внесены по 50 мкл на лунку. Разведенный до 15 мкмоль в полной среде пептид также был добавлен по 50 мкл на лунку. В случае серий без пептида на лунку были добавлены 50 мкл полной среды. Культуральный планшет был помещен в инкубатор (37 С, 5% СO2, Н 2O) на 20 ч, затем был отобран супернатант и концентрация IL2 была оценена с помощью тестаCTL.L2. Таким образом наблюдают, что добавление пептида НА (5 мМ) специфически индуцирует стимуляцию клетки Т Jurkat, экспрессирующей рецептор комплекса DR1-HA (ТНА), 004428 44 тогда как не оказано никакого воздействия на контрольный Т-лимфоцит (Т Jurkat). Кроме того, экзосомы не способны стимулировать ТНА в отсутствие пептида НА (фиг. 8 С). 9. Экзосомы человеческой мастоцитарной линии НМС 1 (фиг. 9). Этот пример показывает, что модифицированные согласно изобретению экзосомы могут быть получены из других клеток, в частности,человеческих клеток. Полученные заявителями результаты в частности свидетельствуют о том,что мастоцитарная линия человеческого происхождения НМС 1 способна продуцировать экзосомы под импульсным действием повышения количества внутриклеточного кальция и в условиях, идентичных тем, которые индуцируют секрецию экзосом крысиной линией RBL 2H3. Характеризация линии НМС 1 путем проточной цитометрии указывает на то, что поверхность этих клеток экспрессирует молекулы класса I CMH (W6.32), но не молекулы класса II(L243). Кроме того, они являются положительными для молекул CD9, CD63 и CD81, но отрицательными для молекул Lamp1 и Lamp2 (фиг. 9 А). Экзосомы были получены из линии НМС 1 путем добавления иономицина (1 ммоль), затем очищены из супернатантов путем дифференциального ультрацетрифугирования. Их состав был проанализирован проточной цитометрией после "перекрестного связывания" с шариками из латекса и путем вестерн-блотирования. Анализ экзосом методом с использованием шариков из латекса показывает, что они несут молекулы Lamp1, CD9, CD63, CD81 и молекулы класса I СМН, но не содержат ни молекул класса II, ни молекулы Lamp2 (фиг. 9 В) . Кроме того, наблюдаемая путем электронной микроскопии морфология этих экзосом идентична таковым, продуцируемым линией RВL 2 Н 3. Наконец, наблюдаемый путем вестерн-блотирования протеиновый состав подтверждает эти результаты, так как согласно этому способу найдены молекулы CD63, Lamp1, СМН класса I, тогда как Lamp2, CMH класса II не обнаружены (фиг. 9 С). В заключение, линия НМС 1 представляется человеческой гомологией крысиной линииRBL-2H3 в отношении ее способности, после повышения количества внутриклеточного кальция, продуцировать экзосомы, которые имеют такие же структурные и молекулярные характеристики. Эти клетки, следовательно, могут продуцировать человеческие рекомбинантные экзосомы, экспрессирующие молекулы класса II СМН или любую другую молекулу, которая будет специфически направлена. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Мембранная везикула, продуцируемая генетически модифицированной клеткой, при 45 чем вышеуказанная везикула содержит рекомбинантную молекулу главного комплекса гистосовместимости (СМН) человека, включающую по меньшей мере одну -цепь класса II. 2. Мембранная везикула, продуцируемая генетически модифицированной клеткой, причем вышеуказанная везикула содержит рекомбинантную молекулу главного комплекса гистосовместимости человека, включающую по меньшей мере одну -цепь класса II. 3. Везикула по п.1 или 2, отличающаяся тем, что рекомбинантная молекула главного комплекса гистосовместимости класса II включает -цепь и -цепь. 4. Везикула по любому из пп.1-3, отличающаяся тем, что рекомбинантную молекулу главного комплекса гистосовместимости классаII выбирают среди серотипов DR1-DR13, предпочтительно DR1-DR7. 5. Везикула по любому из пп.1-4, отличающаяся тем, что она дополнительно содержит молекулу главного комплекса гистосовместимости класса I. 6. Везикула по любому из пп.1-5, отличающаяся тем, что она содержит комплекс между определенным пептидом и рекомбинантной молекулой главного комплекса гистосовместимости. 7. Везикула по любому из пп.1-6, отличающаяся тем, что она дополнительно содержит одну или более представляющих интерес гетерологичных молекул. 8. Везикула по любому из пп.1-7, отличающаяся тем, что она дополнительно содержит рекомбинантный пептид или протеин, позволяющий осуществлять ее очистку. 9. Везикула по пп.1-8, отличающаяся тем,что она включает маркер. 10. Везикула по п.9, отличающаяся тем,что маркер представляет собой зеленый флуоресцентный белок (GFP). 11. Везикула по любому из пп.1-10, отличающаяся тем, что она, по существу, лишена эндогенных молекул СМН. 12. Мембранная везикула по любому из пп.1-11, отличающаяся тем, что она содержит рекомбинантную молекулу, образующуюся путем слияния между представляющим интерес пептидом и сигналом адресации. 13. Мембранная везикула, отличающаяся тем, что ее получают из генетически модифицированной тучной клетки или производной генетически модифицированной тучной клетки и она содержит одну или более представляющих интерес гетерологичных молекул. 14. Везикула по п.13, отличающаяся тем,что представляющей интерес молекулой является протеин, полипептид, пептид, нуклеиновая кислота, липид или вещество химической, биологической или синтетической природы. 46 15. Мембранная везикула по п.14, отличающаяся тем, что гетерологичной молекулой является молекула главного комплекса гистосовместимости, антиген, лиганд рецептора, рецептор лиганда, нуклеиновая кислота, фармакологический продукт, маркер и/или пептид для очистки. 16. Везикула по п.15, отличающаяся тем,что она экспрессирует рецептор лиганда и содержит другую представляющую интерес гетерологичную молекулу. 17. Мембранная везикула по любому из пп.13-16, отличающаяся тем, что она содержит рекомбинантную молекулу, образующуюся путем слияния между представляющим интерес пептидом и сигналом адресации. 18. Продуцирующая экзосомы клетка, отличающаяся тем, что она включает одну или более рекомбинантных нуклеиновых кислот,кодирующих молекулу главного комплекса гистосовместимости, включающую по меньшей мере одну -цепь класса II СМН человека. 19. Клетка по п.18, отличающаяся тем, что она является тучной клеткой. 20. Клетка по п.19, отличающаяся тем, что она происходит от линии тучных клеток лейкоза базофилов, в частности от линии RBL, предпочтительно RBL-2H3. 21. Клетка по любому из пп.18-20, отличающаяся тем, что она включает рекомбинантную нуклеиновую кислоту, кодирующую цепь, или рекомбинантную нуклеиновую кислоту, кодирующую -цепь и -цепь молекулы главного комплекса гистосовместимости классаII и, необязательно, молекулу главного комплекса гистосовместимости класса I. 22. Способ получения мембранной везикулы по любому из пп. 1-17, содержащей определенную рекомбинантную молекулу, включающую по меньшей мере одну -цепь класса II СМН человека, включающий следующие стадии: а) культивирование тучной клетки или производной тучной клетки, включающей рекомбинантную нуклеиновую кислоту, кодирующую вышеуказанную определенную рекомбинантную молекулу; с) рекуперацию продуцированных вышеуказанными клетками везикул, причем эти везикулы включают вышеуказанную определенную рекомбинантную молекулу. 23. Способ по п.22, отличающийся тем, что он включает промежуточную стадию b), в ходе которой клетки стимулируют для индукции и/или повышения секреции экзосом. 24. Способ по п.22 или 23, отличающийся тем, что определенная рекомбинантная молекула экспонирована снаружи экзосомы или включена, частично или полностью, в цитозольную часть экзосомы. 47 25. Способ по любому из пп.22-24, отличающийся тем, что рекомбинантной молекулой является молекула главного комплекса гистосовместимости, антигенная молекула, лиганд рецептора, рецептор лиганда, пептид для очистки или другой представляющий интерес полипептид. 26. Способ по любому из пп.22-25, отличающийся тем, что нуклеиновая кислота включает, кроме того, область, кодирующую сигнал адресации к мембранным компартментам тучной клетки. 27. Способ получения экзосомы, содержащей комплекс пептид-СМН определенного состава, включающий культивирование продуцирующей экзосомы клетки, включающей одну или более рекомбинантных нуклеиновых кислот, кодирующих определенную рекомбинантную молекулу СМН,включающую по меньшей мере одну -цепь класса II СМН человека; стимуляцию клеток для индукции высвобождения экзосом; рекуперацию продуцированных вышеуказанными клетками экзосом, причем эти экзосомы экспрессируют на своей поверхности вышеуказанную определенную рекомбинантную молекулу СМН; и введение в контакт экзосом с пептидом или пептидами. 28. Способ по п.27, отличающийся тем, что продуцирующей клеткой является тучная клетка или производная тучной клетки. 29. Способ по любому из пп.27-28, отличающийся тем, что продуцирующая клетка, по существу, лишена эндогенной молекулы СМН. 30. Способ получения экзосомы, содержащей комплекс пептид-СМН определенного состава, включающий культивирование продуцирующей экзосомы клетки, включающей одну или более рекомбинантных нуклеиновых кислот, кодирующих определенную рекомбинантную молекулу СМН,и нуклеиновую кислоту, включающую область,кодирующую определенный рекомбинантный пептид; стимуляцию клеток для индукции высвобождения экзосом; рекуперацию продуцированных вышеуказанными клетками экзосом, причем эти экзосомы экспрессируют на своей поверхности вышеуказанную определенную рекомбинантную молекулу СМН, связанную с вышеуказанным рекомбинантным пептидом. 31. Способ по п.30, отличающийся тем, что кодирующая рекомбинантный пептид нуклеиновая кислота кодирует производное инвариантной цепи li, в котором участок CLIP подвергнут делеции и заменен вышеуказанным пептидом. 32. Способ по любому из пп.30-31, отличающийся тем, что продуцирующей клеткой 48 является тучная клетка или производная тучной клетки. 33. Способ по любому из пп.30-32, отличающийся тем, что продуцирующая клетка, по существу, лишена эндогенной молекулы СМН. 34. Способ модификации состава экзосомы, включающий введение в продуцирующую экзосомы клетку нуклеиновой кислоты, кодирующей определенную молекулу, связанную с сигналом адресации в мембранных компартментах, и получение экзосом из вышеуказанной клетки. 35. Композиция, включающая одну или более мембранных везикул по любому из пп.117. 36. Применение везикулы по любому из пп.1-17 для получения поликлональных и/или моноклональных антител. 37. Способ получения антител, включающий иммунизацию животного с помощью везикулы по п.7 и рекуперацию антител и/или клеток, продуцирующих антитела или принимающих участие в иммунном ответе. 38. Способ по п.37, отличающийся тем, что он используется для получения моноклональных антител, в частности, специфичных к комбинации СМН-пептид. 39. Применение антитела или фрагмента антитела, полученного способом по п.37 или 38,для детекции в биологическом образце наличия соответствующих специфических антигенов. 40. Применение антитела или фрагмента антитела, полученного способом по п.37 или 38,для получения терапевтической композиции,предназначенной для ингибирования взаимодействия между рецептором Т-лимфоцита и комплексом СМН-пептид, к которому он специфичен. 41. Применение везикулы по п.1 или 2 для получения терапевтической композиции, предназначенной для ингибирования взаимодействия между рецептором Т-лимфоцита и комплексом СМН-пептид, к которому он специфичен. 42. Применение мембранной везикулы по любому из пп.1-17 для детекции in vitro или exvivo в биологическом образце партнеров, специфичных к белковой молекуле. 43. Применение по п.42, отличающееся тем, что экзосома несет комплекс СМН-пептид для детекции специфичных к этому комплексу Т-лимфоцитов в биологическом образце. 44. Применение по п.42, отличающееся тем, что экзосома несет рецептор TcR для детекции специфичных к этому рецептору комплексов пептид-СМН в биологическом образце. 45. Применение по п.42, отличающееся тем, что экзосома несет рецептор лиганда для детекции наличия вышеуказанного лиганда в биологическом образце. 46. Способ детекции in vitro или ex vivo наличия специфичных к комплексам антигенСМН Т-лимфоцитов в биологическом образце, 49 включающий введение в контакт вышеуказанного образца с маркированной экзосомой по п.7,содержащей вышеуказанный комплекс антигенСМН,и обнаружение маркировки Тлимфоцитов в вышеуказанном образце. 47. Применение везикулы по п.6 для клональной амплификации и/или стимуляции exvivo цитотоксических или регуляторных Тлимфоцитов. 48. Применение везикулы по любому из пп.13-17 для получения композиции, предназначенной для транспортировки вышеуказанной молекулы к клетке. 49. Композиция, включающая одну или более мембранных везикул по любому из пп.117, иммобилизованных на носителе. 50 50. Композиция по п.49, включающая одну или более мембранных везикул, отличающаяся тем, что везикулы иммобилизованы на шарике,в частности на шарике из латекса или магнитном шарике. 51. Применение мембранной везикулы по любому из пп.1-17, в частности, в иммобилизованной на носителе форме для очистки клеток. 52. Мембранная везикула, продуцируемая генетически модифицированной клеткой, причем вышеуказанная везикула содержит рекомбинантную молекулу главного комплекса гистосовместимости человека, включающую по меньшей мере одну цепь класса II, и маркер.
МПК / Метки
МПК: G01N 33/50, C07K 16/00, C12N 5/10, A61K 39/385
Метки: экзосомы, модифицированные, применение
Код ссылки
<a href="https://eas.patents.su/28-4428-modificirovannye-ekzosomy-i-ih-primenenie.html" rel="bookmark" title="База патентов Евразийского Союза">Модифицированные экзосомы и их применение</a>
Гидрофобно-модифицированные белки, способы их получения и их применение

Номер патента: 3739
Опубликовано: 28.08.2003
Авторы: Пепински Р.Блэйк, Уилльямс Кевин П., Тэйлор Фредерик Р., Вен Дингайи, Гэлдес Элфонс, Бейкер Даррен П., Гэрбер Эллен Э., Портер Джеффри
МПК: A61P 25/00, C07K 14/00, A61K 38/00...
Метки: применение, способы, гидрофобно-модифицированные, получения, белки
Формула / Реферат:
1. Изолированный белок, включающий в себя N-концевую аминокислоту и C-концевую аминокислоту, который выбран из (a) белка с N-концевым цистеином, соединенным, по меньшей мере, с одной гидрофобной структурной единицей; (b) белка с N-концевой аминокислотой, отличной от цистеина, соединенного, по меньшей мере, с одной гидрофобной структурной единицей; и (c) белка, по меньшей мере, с одной гидрофобной структурной единицей, заменяющей N-концевую...
Модифицированные аминокислоты и лекарственное средство на их основе

Номер патента: 4037
Опубликовано: 25.12.2003
Авторы: Халлермайер Герхард, Винен Вольфганг, Эберлайн Вольфганг, Энцерот Михаель, Энгель Вольфхард, Доодс Хенри, Рудольф Клаус, Пипер Хельмут
МПК: C07D 401/12, A61P 3/10, C07K 5/06...
Метки: основе, лекарственное, модифицированные, средство, аминокислоты
Формула / Реферат:
1. Модифицированные аминоксилоты общей формулы (I) в которой R означает неразветвленный алкил с 1-3 атомами углерода, который на конце замещен циклоалкилом с 5-7 атомами углерода, одной или двумя фенильными группами, 1-нафтилом, 2-нафтилом или 4-бифенилилом, причем указанные ароматические остатки дополнительно могут быть замещены атомом фтора, хлора или брома, метилом, метоксигруппой, аминогруппой или ацетиламиногруппой, 2-пирролилом,...
Водорастворимые или вододиспергируемые повторно используемые смолы на основе полиэтилентерефталата или терефталатов, модифицированные сульфоарильными группами

Номер патента: 2279
Опубликовано: 28.02.2002
Автор: Сэлсмэн Роберт Кит
МПК: D02G 3/00
Метки: полиэтилентерефталата, используемые, группами, сульфоарильными, терефталатов, повторно, вододиспергируемые, основе, водорастворимые, смолы, модифицированные
Формула / Реферат:
1. Водорастворимая или вододиспергируемая повторно используемая полиэфирная смола, представляющая собой продукт реакции 1-50 маc.% сульфоарилдикарбоновой кислоты, 10-50 маc.% терефталатного полимера, содержащего полиалкилентерефталатные звенья, 1-50 маc.%, по меньшей мере, одного гликоля и, необязательно, 1-50 маc.%, по меньшей мере, одного оксиалкилированного полиола. 2. Смола по п.1, в которой сульфоарилдикарбоновая кислота находится в форме...
Клеточная везикула, называемая “экзосома”, ее получение и применение для стимуляции иммунного ответа

Номер патента: 2827
Опубликовано: 31.10.2002
Авторы: Амигорена Себасьян, Рапосо Граса, Реньо Армелль, Зитвожель Лоранс
МПК: A61K 39/00, C12N 5/08
Метки: получение, ответа, называемая, применение, везикула, стимуляции, клеточная, экзосома, иммунного
Формула / Реферат:
1. Мембранная везикула, называемая декзосомой, выделенная из своего естественного окружения, отличающаяся тем, что она (i) происходит от дендритной клетки, (ii) включает билипидный слой, окружающий цитозольную часть, (iii) имеет на своей поверхности молекулы класса I и/или II главного комплекса гистосовместимости (СМН), (iv) содержит одну или несколько молекул CD63 и (v) имеет диаметр примерно от 60 до 90 нм. 2. Везикула по п.1, отличающаяся...
Клеточная везикула, называемая “текзосома”, ее получение и применение для стимуляции иммунного ответа

Номер патента: 4300
Опубликовано: 26.02.2004
Авторы: Рапосо Граса, Амигорена Себасьян, Реньо Армелль, Зитвожель Лоранс
МПК: C12N 5/08, A61K 39/00
Метки: стимуляции, текзосома, получение, применение, называемая, везикула, иммунного, ответа, клеточная
Формула / Реферат:
1. Везикула, называемая текзосомой, освобожденная от своего естественного окружения, отличающаяся тем, что она происходит от опухолевой клетки, тем, что она включает липидный бислой, окружающий цитозольную фракцию, содержащую опухолевые антигенные молекулы и/или иммуномодуляторы, и/или хемоаттрактанты, и/или гормоны, и/или нуклеиновые кислоты, тем, что она имеет на своей поверхности молекулы класса I и/или класса II главного комплекса...
Предыдущий патент: Замещенные 3-цианохинолины в качестве ингибиторов протеинтирозинкиназ
Следующий патент: Полиэфирные нити с высоким модулем и с малой усадкой и способ их формования
Случайный патент: Набор для переноса гена в клетки-мишени с помощью ретровируса (варианты)