Способы инактивации клеток-мишеней, агенты и композиции, вызывающие цитолиз, и соединения, используемые для получения этих агентов
Номер патента: 2688
Опубликовано: 29.08.2002
Авторы: Ландо Петер, Форсберг Геран, Абрахмсен Ларс, Калланд Терье, Согорд Мортен, Дохлстен Микаэль

























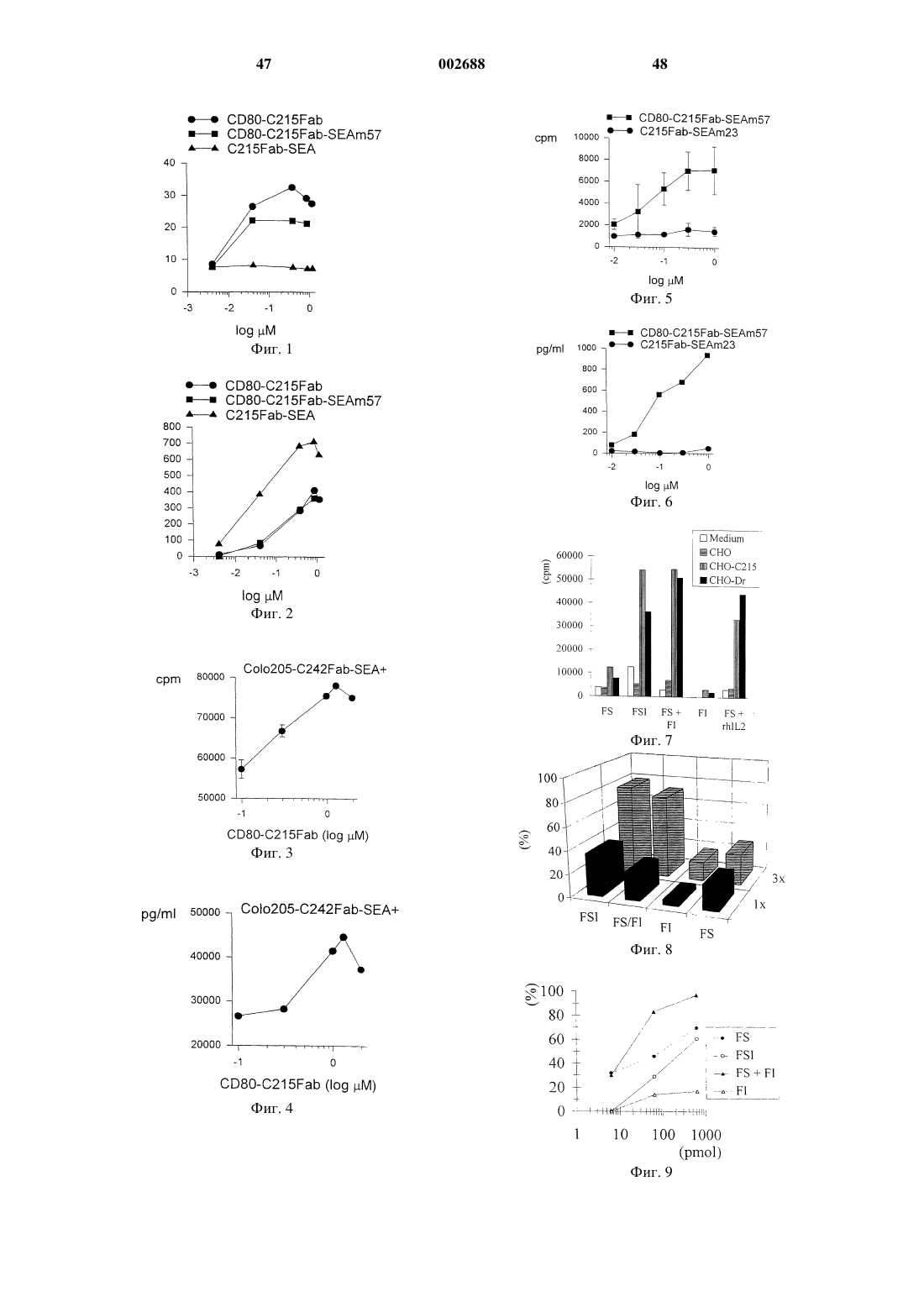
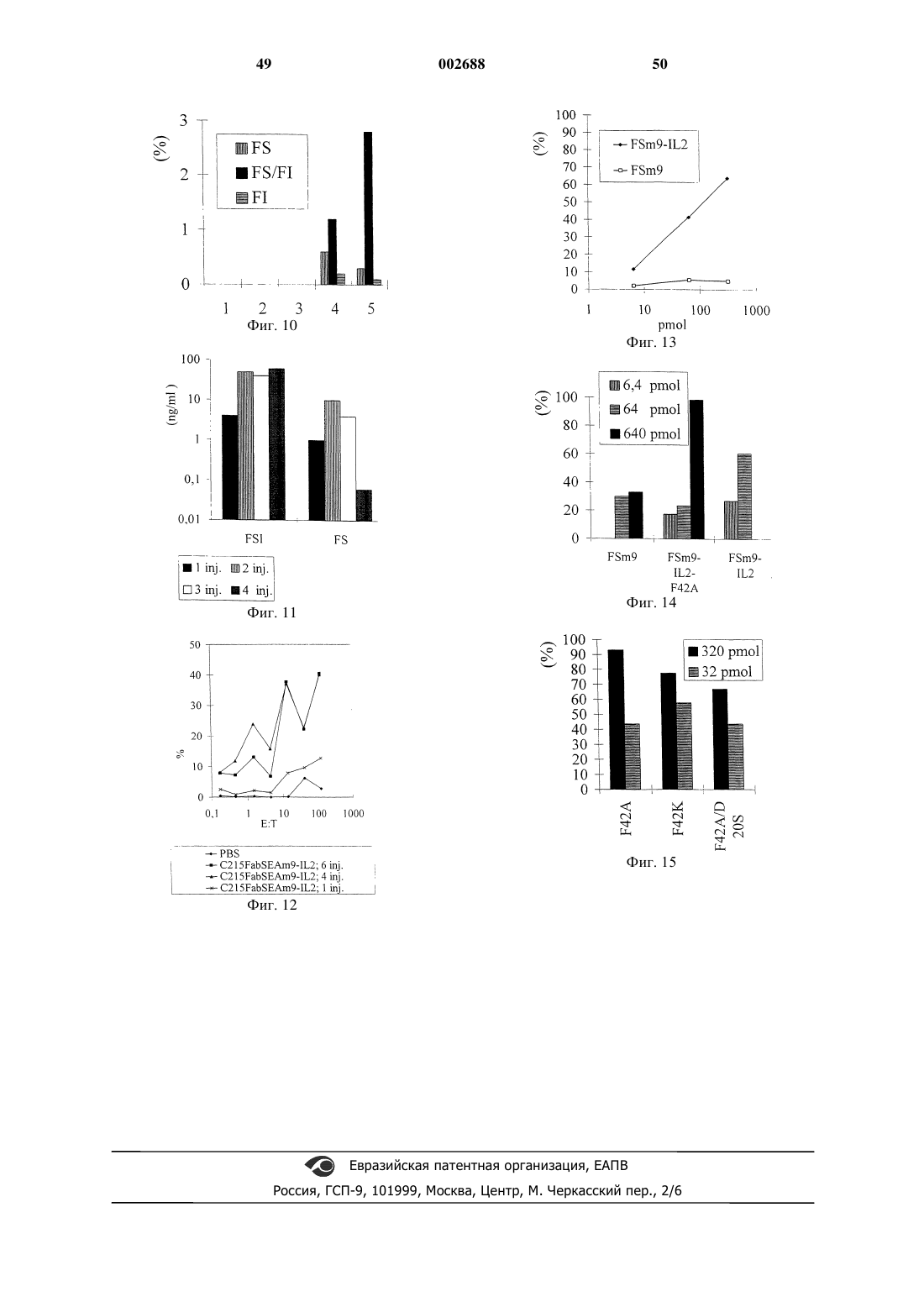
Формула / Реферат
1. Способ инактивации клеток-мишеней в присутствии Т-клеток, включающий контактирование клеток мишеней и Т-клеток с суперантигеном в присутствии иммуномодулятора, когда, по меньшей мере, один из элементов - суперантиген или иммуномодулятор - конъюгирован с нацеливающим компонентом, причем нацеливающий компонент выбран из группы, состоящей из антитела, антигенсвязывающего фрагмента антитела, Fab-фрагмента антитела, Fаb2-фрагмента антитела или одноцепочечного антитела, а указанный суперантиген модифицирован, таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида, и указанный иммуномодулятор выбирают из группы, содержащей цитокины, хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов.
2. Способ по п.1, отличающийся тем, что суперантиген и иммуномодулятор оба конъюгированы с одним и тем же компонентом мишени в форме тройного конъюгата.
3. Способ по п.1, отличающийся тем, что конъюгированы только суперантиген и нацеливающий компонент, в то время как иммуномодулятор не конъюгирован.
4. Способ по п.1, отличающийся тем, что клетки-мишени инактивируют in vivo в индивидууме, страдающем заболеванием, ассоциированным с клетками-мишенями.
5. Способ по п.4, отличающийся тем, что заболеванием является рак.
6. Способ по п.1, отличающийся тем, что суперантиген представляет собой химеру, содержащую последовательность, производную из двух или более суперантигенов дикого вида.
7. Способ по п.1, отличающийся тем, что иммуномодулятор представляет собой IL-2.
8. Способ по п.1, отличающийся тем, что иммуномодулятор представляет собой внеклеточную часть молекулы В7.
9. Способ по п.8, отличающийся тем, что внеклеточную часть молекулы В7 выбирают из группы, содержащей CD80 и CD86.
10. Способ по п.1, отличающийся тем, что иммуномодулятор модифицирован с целью уменьшения сродства рецепторов лимфоцитов по сравнению со сродством соответствующей нативной формы.
11. Конъюгат суперантигена, имеющий формулу
(T)x(Sag)y(IM)z,
где у>0 и z>0;
Т представляет собой нацеливающий компонент, выбран из группы, состоящей из антитела, антигенсвязывающего фрагмента антитела, Fab-фрагмента антитела, Fab2-фрагмента антитела или одноцепочечного антитела,
Sag представляет собой суперантиген, который модифицирован таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида,
IM представляет собой иммуномодулятор, выбранный из группы, содержащей цитокины, хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов.
12. Конъюгат суперантигена по п.11, где 10>х>0, 10>у>1 и 10>z>1.
13. Конъюгат суперантигена по п.12, где х, у и z представляют собой целые числа 1-3.
14. Конъюгат суперантигена по п.11, где Т содержит, по крайней мере, Т' и Т", суперантиген слит С-концом с Т', а иммуномодулятор слит С-концом с Т".
15. Конъюгат суперантигена по п.11, где суперантиген представляет собой химеру, содержащую последовательность, производную из двух или более суперантигенов дикого вида.
16. Конъюгат суперантигена по п.11, где иммуномодулятор представляет собой IL-6.
17. Конъюгат суперантигена по п.11, где иммуномодулятор представляет собой внеклеточную часть молекулы В7.
18. Конъюгат суперантигена по п.17, где внеклеточная часть молекулы В7 выбирают из группы, содержащей CD80 и CD86.
19. Конъюгат суперантигена по п.11, где иммуномодулятор модифицирован с целью уменьшения сродства рецепторов лимфоцитов по сравнению со сродством соответствующей нативной формы.
20. Конъюгат суперантигена по п.14, где суперантиген представляет собой стафилококковый энтеротоксин А, Т' является СH1 доменом С215 Fab, Т" является легкой цепью антитела С215, а иммуномодулятор представляет собой IL-2.
21. Конъюгат суперантигена по п.14, где суперантиген слит с Т' с помощью гибкого гидрофильного аминокислотного линкера В из 3-11 аминокислотных остатков, а иммуномодулятор слит с Т" через гидрофильный нейтральный или положительно заряженный аминокислотный линкер Q из 10-20 аминокислотных остатков.
22. Конъюгат суперантигена по п.21, где В выбирают из группы, состоящей из Gly-Gly-Pro и Pro-Ala-Ser-Gly-Gly-Gly-Gly-Ala-Gly-Gly-Pro (SEQ ID N 19), а Q выбирают из группы, состоящей из Gly-Pro-Arg-Gln-Ala-Asn-Glu-Leu-Pro-Gly-Ala-Pro-Ser-Gln-Glu-Glu-Arg (SEQ ID N 23), Gly-Pro-Arg-Gln-Ser-Asn-Glu-Thr-Pro-Gly-Ser-Pro-Ser-Gln-Glu-Glu-Arg (SEQ ID N 20), Gly-Pro-Arg-Gln-Ala-Lys-Thr-Leu-Pro-Gly-Ala-Pro-Ser-Gln-Thr-Thr-Arg (SEQ ID N 21) и Gly-Pro-Thr-Glu-Ala-Asp-Glu-Leu-Pro-Gly-Ala-Pro-Ser-Glu-Glu-Glu-Thr (SEQ ID N 22).
23. Конъюгат суперантигена по п.11, где х=0, у=1-3, a z=1-3.
24. Молекула ДНК, кодирующая суперантиген и иммуномодулятор не являющийся суперантигеном, которая находится в форме бицистронной конструкции, в которой
первый цистрон содержит последовательность, кодирующую суперантиген, модифицированный таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида,
и второй цистрон содержит последовательность, кодирующую иммуномодулятор, выбранный из группы, содержащей цитокины, хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов.
25. Молекула ДНК по п.24, где иммуномодулятором является IL-2.
26. Молекула ДНК по п.24, где указанный кодируемый иммуномодулятор представляет собой внеклеточную часть молекулы В7, выбранную из группы состоящей из CD80 и CD86.
27. Молекула ДНК, кодирующая суперантиген модифицированный таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида; иммуномодулятор, выбранный из группы, содержащей цитокины, хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов, нацеливающий компонент, выбранный из группы, состоящей из антитела, антигенсвязывающего фрагмента антитела, Fab-фрагмента антитела, Fab2-фрагмента антитела или одноцепочечного антитела.
28. Молекула ДНК по п.27, где иммуномодулятор является IL-2.
29. Молекула ДНК по п.27, где иммуномодулятор представляет собой внеклеточную часть молекулы В7 и выбран из группы, содержащей CD80 и CD86.
30. Фармацевтическая композиция для инактивации клеток-мишеней, содержащая суперантиген, иммуномодулятор и нацеливающий компонент, где, по меньшей мере, один из элементов - суперантиген или иммуномодулятор - конъюгирован с нацеливающим компонентом, причем указанный нацеливающий компонент выбран из группы, состоящей из антитела, антигенсвязывающего фрагмента антитела, Fab-фрагмента антитела, Fab2-фрагмента антитела или одноцепочечного антитела, а указанный суперантиген модифицирован таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида, и указанный иммуномодулятор выбирают из группы, содержащей цитокины, хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов, причем указанная композиция взята в эффективном количестве.
31. Фармацевтическая композиция по п.30, в которой суперантиген представляет собой химеру, содержащую последовательность, производную из двух или более суперантигенов дикого вида.
32. Фармацевтическая композиция по п.30, в которой иммуномодулятором является IL-2.
33. Фармацевтичесъря композиция по п.30, в которой иммуномодулятор представляет собой внеклеточную часть молекулы В7.
34. Фармацевтическая композиция по п.33, в которой внеклеточная часть молекулы В7 выбрана из группы, содержащей CD80 и CD86.
35. Фармацевтическая композиция по п.30, в которой иммуномодулятор модифицирован таким образом, чтобы уменьшить сродство рецепторов лимфоцитов по сравнению со сродством соответствующей нативной формы.
Текст
1 ОБЛАСТЬ ТЕХНИКИ Настоящее изобретение касается инактивации/цитолиза клеток-мишеней, вызываемых Т-клетками (Т-лимфоцитами), активированными функциональными суперантигенами. Цитолиз можно использовать в терапии и в проводимых in vitro исследованиях. ОПРЕДЕЛЕНИЯ Суперантигены. Согласно самому первому определению (примерно в 1988-1993 гг.),суперантигены представляют собой бактериальные или вирусные белки, способные к связыванию с антигенами ГКГ (главного комполекса гистосовместимости) класса II без предварительного осуществления внутриклеточных процессов, и активирующие Т-клетки посредством связывания с вариабельной областью (V) Vцепи рецептора Т-клетки (TCR). Это связывание ведет к ограниченной активации V-семейства у относительно большой части/субпопуляции Тклеток и к лизису клеток, экспрессирующих ГКГ класса II (т.е. к цитолизу, опосредованному зависимыми от суперантигена клетками SDCC). Обычно активируемая суперантигеном субпопуляция Т-клеток составляет около 1-30% от общего числа Т-клеток индивидуума. Хорошо известные суперантигены дикого типа, согласно вышеприведенному определению, представляют собой стафилококковые энтеротоксины (SEA, SEB, SEC1, SEC2, SED, SEE и SEH). Другими их примерами являются токсин синдрома токсического шока 1 (TSST-1,также стафилококкового происхождения), отслаивающие токсины (Exft), стрептококковый пирогенный экзотоксин А, В и С (SPE А, В, С),вирус опухоли молочной железы мышей(MMTV), стрептококковые М-белки, энтеротоксин клостридиального озноба (СРЕТ), суперантигены микоплазменного артрита и т.п. Обзор суперантигенов и их свойств см. в публикацииKotzin et al. 1993. Суперантигены дикого типа и химерные суперантигены также могут быть подвергнуты мутагенезу, чтобы придать им свойства уменьшенного или отсутствующего связывания с ГКГ класса II и/или связывания с TCRV ( Kappler etal., WO 9736932; Antonsson et al., 1997). Этот тип суперантигенов становится менее токсичным. В случае, если они полностью теряют способность связываться с ГКГ класса II или сTCRV, то они больше не являются функциональными суперантигенами, поскольку в этом случае они теряют свою способность к активации Т-клеток. Посредством мутирования структурно подобных суперантигенов дикого типа оказалось возможным сконструировать химерные функционально активные суперантигены (гибридные 2 суперантигены) (Lamphaer et al., 1996 и Antonsson et al., WO 9736932). Было установлено, что активация и последующий лизис клеток могут происходить независимо от ГКГ класса II, в случае, если суперантиген дикого типа был конъюгирован с частью,обеспечивающей связывание с мишенью, способной связываться со структурами поверхности клетки (Dohlsten et al., WO 9201470). Этот новый эффекторный механизм получил название цитолиз, опосредованный клетками, зависимыми от суперантигена и антитела(=SADCC). Он включает аналогичные механизмы для обеспечивающих связывание с мишенями частей, иных чем антитела (Abrahmsn et al.,WO 9601650; Antonsson et al., WO 9736932). В соответствии с принятой в настоящее время концепцией суперантигенов, она охватывает любые соединения (предпочтительно полипептидной структуры), которые без осуществления внутриклеточных процессов способны связываться со структурой клеточной поверхности (структурой-мишенью) и с одной или более полиморфными TCR-цепями, в частности с Vцепью, активируя тем самым субпопуляцию Тклеток, экспрессирующих конкретную TCRцепь, участвующую в связывании. После этого Т-клетки становятся цитотоксичными и направляют свою цитотоксичность против клеток, несущих поверхностные структуры (структурымишени, клетки-мишени). Определение суперантигена (SAG), как оно используется в контексте настоящего изобретения, кроме особо оговоренных случаев, таким образом, охватывает конъюгаты между частью, обеспечивающей связывание с мишенью, и свободным суперантигеном, как описано выше для SADCC. Под термином суперантиген подразумеваются, кроме особо оговоренных случаев,только функциональные суперантигены. Свободный суперантиген (Sag) представляет собой суперантиген дикого типа, возможно мутированный или другим образом модифицированный, который не конъюгирован с частью,обеспечивающей связывание с мишенью или с иммуномодулятором. Способность свободных суперантигенов к связыванию с ГКГ класса II является наследственным свойством связывания с мишенью. Поскольку у свободных суперантигенов отсутствуют конъюгированные части,обеспечивающие связывание с мишенью, то они могут использовать только механизм SDCC (цитолиза, опосредованного зависимыми от суперантигена клетками). Коньюгированный суперантиген представляет собой конъюгат между свободным суперантигеном и частью, обеспечивающей связывание с мишенью, или с иммуномодулятором. Конъюгированный суперантиген использует механизмы как SDCC (цитолиза, опосредованного зависимыми от суперантигена клетками),так и SADCC (цитолиза, опосредованного клет 3 ками, зависимыми от суперантигена и антитела). Иммуномодулятор (IM) представляет собой соединение, обладающее способностью регулировать иммунную систему. В контексте настоящего изобретения суперантигены рассматриваются отдельно и не охватываются термином Иммуномодулятор, когда он используется здесь. Иммуномодулятор часто имеет наследственно обусловленную способность к связыванию с мишенью, такую, как для соответствующего рецептора лимфоцита. Кроме особо оговоренных случаев, иммуномодулятор находится в неконъюгированной форме. Обеспечивающая связывание с мишенью часть (Т) представляет собой часть, способную связываться со структурой клеточной поверхности и/или со структурой ткани. Коньюгат состоит из двух или более частей Sag, IM, Т и т.п., которые ковалентно соединены друг с другом. Под растворимыми формами активных ингредиентов имеются в виду такие формы, которые растворимы в текучих средах, существующих в организме, таких как сыворотка и плазма крови. Известный уровень техники терапевтическое использование суперантигенов Неконъюгированные суперантигены дикого типа и мутантные суперантигены предложены для использования в терапии, при этом их лечебный эффект преимущественно осуществляется посредством активации иммунной системы либо локальной активации по отношению к клеткам, экспрессирующим класс II, связанным с подлежащей лечению болезнью либо посредством системной активации (Kalland et al., WO 9104053; Terman et al., WO 9110680 и WO 9324136; Antonsson et al., WO 9736932; и Newell at al., 1991). Из-за очень сильной токсичности суперантигенов дикого типа, этот подход в отношении лечения рака может применяться только для очень малой части от всех форм рака. Было также предложено использовать суперантигены, конъюгированные с частями,обеспечивающими связывание с мишеньюOchi et al., 1993), причем все эти три публикации включены в настоящее описание в качестве ссылок. В связи с исследованиями по предотвращению индуцированной суперантигенами регуляции опосредованной Т-клетками цитотоксической активности по типу обратной связи, проявляемой IL-2 in vivo, было высказано предположение, что было бы полезно вводить IL-2 совместно с неконъюгированными суперантигенами дикого типа и с суперантигенами дикого типа, конъюгированными с антителами (Belfrage Thesis Augusti/September 1996; Belfrage et 4 1997a; Belfrage et al., 1977b (суперантигены дикого типа. Предположили также, что заякоренный на клеточной мембране CD80 играет роль в активации суперантигеном Т-клеток в отсутствие антигенов ГКГ класса II (Lando et al., 1993 и 1996). Фиг. 4 в публикации Lando et al., 1996, иллюстрирует эксперимент, в котором анализировали способность суперантигена, конъюгированного только с антителом или с антителом в комбинации с IL-2, индуцировать пролиферацию покоящихся Т-клеток человека. В этом продолжавшемся 4 суток эксперименте конъюгированный суперантиген присутствовал на исходной СНО-клетке и СНО-клетках, трансфецированных для экспрессии класса II или С 215 или класса II плюс С 215. Эффект IL-2 был незначительным.Kappler et al. (WO 9314264) предположили,что неконъюгированные SEB дикого типа мутировали, потеряв в результате этого свою способность связываться с V или ГКГ класса II (в контексте вакцин и в качестве агента, нейтрализующего токсические эффекты суперантигенов).Abrahmsen et al. (WO 9601650) предложили способ терапии рака с помощью конъюгированных суперантигенов, обладающих модифицированной, предпочтительно уменьшенной способностью связывать антигены класса II. Antonsson etal. (WO 9736932) предложили способ терапевтического лечения рака с помощью химерных суперантигенов и суперантигенов с уменьшенной серологической реактивностью (см. такжеAbrahmsen et al). Мутации, такие как описанные в публикациях Abrahmsn et al. WO 9601650) иAntonsson et al. (WO 9736932), включают суперантигены с пониженной системной токсичностью, с пониженной иммуногенностью и/или с пониженной серологической реактивностью у подвергавшихся лечению млекопитающих. Предложен также способ терапии с помощью введения нуклеиновых кислот, кодирующих суперантигены дикого типа (Terman et al., WO 9110680;WO 9324136; и Dow et al., WO 9636366). Dow et al. пошли дальше и предложили совместное введение нуклеиновой кислоты, кодирующей цитокин или хемокин, с нуклеиновой кислотой, кодирующей суперантиген. Без экспериментального подтверждения в публикации WO 9636366 предлагаются также конструкции в форме бисцистронных генных конструкций, в которых один цистрон содержит ген, кодирующий суперантиген, а другой цистрон содержит ген, кодирующий цитокин или хемокин. Без экспериментального подтверждения,Pouletty P. (Sangstat, ЕР 510949) высказал предположение, что конъюгаты между частями, обеспечивающими связывание с мишенью, такими какIL-2, и суперантигенами дикого типа могут быть полезны для инактивации клеток, экспрессирующих рецептор IL-2. 5 Известный уровень техники - терапевтическое использование иммуномодуляторов в комбинации с антителами, специфичными для клеток, подлежащих инактивации Ранее было предложено конъюгировать антитела с модификаторами биологических реакций, например с хемокином или цитокином,такими как интерлейкин-2 (Fell at al., ЕР 439095;Rosenblum et al., EP 396387; Pancook et al., 1996; и Becker et al., 1996). Проблемы, решение которых является целью настоящего изобретения Настоящее изобретение имеет целью добиться улучшений в суперантигенной терапии,включающих активацию иммунной системы,для того, чтобы инактивировать нежелательные клетки-мишени у млекопитающего, подвергаемого лечению. В частности, эти улучшения касаются следующего: 1) продления периода активации локально, например в опухоли, во время первого цикла лечения; 2) противодействия появлению сверхчувствительности из-за тенденции активированных Т-клеток приобретать толерантность; 3) усиления активации Т-клеток,независимых от ГКГ класса II, в области опухоли; и 4) расширения терапевтического окна для цитолиза посредством активации суперантигенов. В настоящее время установлено, что эти улучшения можно осуществить полностью или частично, в случае если введение суперантигена(SAG) объединяют с введением иммуномодулятора в растворимой форме, причем по, меньшей мере, один из них (суперантиген или иммуномодулятор) находится в форме конъюгата с частью, обладающей свойством связывания с мишенью для подлежащей инактивации клетки. Первый важный аспект изобретения: способ инактивации клеток-мишеней Первый аспект изобретения касается как терапии, так и исследований in vitro и представляет собой способ инактивации нежелательных клеток-мишеней в присутствии Т-клеток посредством приведения этих двух типов клеток в контакт с суперантигеном (SAG), в частности с суперантигеном, активирующим Т-клетки путем связывания с TCR V, в присутствии иммуномодулятора (IM), не являющегося суперантигеном (Sag). В своем наиболее широком аспекте этот способ отличается тем, что, по меньшей мере, один элемент: либо суперантиген, либо иммуномодулятор, находится в форме конъюгата с частью (Т), обладающей свойством связывания с мишенью для клетки, подлежащей инактивации. В своем подаспекте данный способ отличается тем, что а) суперантиген (SAG) и иммуномодулятор используют в форме тройного конъюгата, включающего суперантиген (Sag), обеспечивающую связывание с клетками-мишенями часть (Т) и иммуномодулятор (IM) (Т, IM, Sag-конъюгат);b) суперантиген (SAG) используют в форме двойного конъюгата между суперантигеном(Sag) и обеспечивающей связывание с клеткамимишенями частью (Т), в комбинации с двойным конъюгатом между иммуномодулятором (IM) и обеспечивающей связывание с клеткамимишенями частью (Т') (Т,Sag-конъюгат + Т',IMконъюгат); с) суперантиген (SAG) используют в форме двойного конъюгата между суперантигеномd) суперантиген (SAG) используют в свободной форме (Sag), a иммуномодулятор используют в форме конъюгата, т.е. двойного конъюгата между иммуномодулятором (IM) и суперантигеном (Sag) (Sag + Т,IМ-конъюгат); и е) суперантиген (SAG) и иммуномодулятор используют в форме двойного конъюгата между суперантигеном (Sag) и иммуномодулятором(IM) (Sag/IM-конъюгат). Мишенями для суперантигена и иммуномодулятора могут быть клетки одного и того же типа, например идентичные или реагирующие друг с другом структуры/эпитопы, или разные типы клеток в пределах одной и той же ткани. Мишенями могут быть нормальные клетки или больные клетки, связанные с одной и той же тканью. Любой элемент - суперантиген или иммуномодулятор - или они оба могут связываться с мишенью с помощью одного или нескольких антител. Болезни, которые можно лечить с помощью способа но настоящему изобретению Болезни, которые можно лечить с помощью данного способа, в принципе те же самые,которые ранее было предложено лечить с помощью суперантигенов. См., например, выше, в разделе под заголовком Известный уровень техники. В качестве иллюстрирующих примеров можно назвать различные формы рака,аутоиммуные болезни, паразитарные инфекции,вирусные инфекции и прочие заболевания, связанные с клетками, экспрессирующими на своей поверхности антигены ГКГ класса II и/или или другие структуры, специфичные для соответствующей болезни и связывающиеся с обеспечивающими связывание с мишенями частями,включенными в суперантиген в соответствии с концепцией настоящего изобретения (формулаI). С помощью настоящего изобретения можно также бороться с бактериальными инфекциями. Важными формами рака в контексте настоящего изобретения являются следующие: меланомы, карциномы, новообразования кроветворной системы и фибросаркомы, а также такие специфические формы рака, как плоскоклеточная карцинома, различные формы рака молочной железы, карциномы головы и шеи, карциномы щитовидной железы, карциномы мяг 7 ких тканей, костные саркомы, тестикулярный рак, рак простаты, раковые заболевания яичников, раковые заболевания мочевого пузыря, раковые заболевания кожи, раковые заболевания мозга, ангиосаркомы, гемангиосаркомы, опухоли клеток молочной железы, первичные раковые заболевания печени, раковые заболевания легких, раковые заболевания шеи, карциномы клеток почек, лейкозы и лимфомы. Сюда входят также и любые типы злокачественных или доброкачественных опухолей, а также формы рака,устойчивые ко многим лекарствам, метастатические формы рака, различные формы рака, индуцированные химическим или вирусным путем(герпес, SV40, ВИЧ и др.) Второй важный аспект изобретения: конъюгаты суперантигена по настоящему изобретению Этот аспект изобретения включает конъюгаты, соответствующие формуле(T)X(Sag)Y(IM)Z I Т представляет собой часть, обеспечивающую связывание с мишенью, Sag соответствует свободному суперантигену, a IM представляет собой иммуномодулятор, не являющийся суперантигеном. Т, Sag и IM связаны вместе с помощью органических линкеров В, которые могут быть разными или одинаковыми в пределах одной и той же молекулы конъюгата или вещества. Конъюгаты в соответствии с формулой I охватывают химические конъюгаты, а также конъюгаты, полученные путем рекомбинации (слитые белки), x, y и z представляют собой целые числа, которые обычно выбирают среди чисел от 0 до 10, такие как 0-5, и представляют собой количество частей Т, Sag и IM соответственно в молекуле данного конъюгата, при условии, что у 0 и каждое из чисел х и z или оба эти числа 0. Химические конъюгаты обычно представляют собой конъюгированные вещества, содержащие смесь различных молекул конъюгата. Соответственно в химических конъюгированных веществах х, у и z могут также представлять собой нецелые числа в диапазоне от 0 до 10, такие как в диапазоне от 0 до 5. В первом подаспекте формулы I, Sag, IM и Т присутствуют в конъюгате (х, у и z 0; Т Sag,IM-конъюгаты), х, у и z обычно представляют собой целые числа от 1 до 3, предпочтительно 1-2. Типичные взаимоотношения между х, у и z следующие: х=у=z; х=у=0,5z; х=0,5 у=0,5z; и х=0,5 у=z. Во втором подаспекте конъюгатов в соответствии с формулой I, обеспечивающая связывание с мишенью часть отсутствует (IM, Sagконъюгаты, х=0), у и z обычно представляют собой целые числа от 1 до 3. Предпочтительные взаимоотношения между х и у следующие: z=у;z=0,5 у; 0,5z=у; z=1/3 у; и 1/3z=у. В обоих подаспектах целые числа или соотношения прежде всего относятся к слитым белкам, в которых часть, обеспечивающая свя 002688 8 зывание с мишенью, может представлять собой белок, содержащий 1, 2, 3 или 4 полипептидных цепи и в котором имеется одна часть Т в каждой молекуле конъюгата. Формула I для конъюгатов в соответствии со вторым подаспектом сокращается до нижеследующего:II Этот тип конъюгатов главным образом подходит для лечения болезней, связанных с клетками, экспрессирующими антигены ГКГ класса II, в частности класс II-экспрессирующих форм рака, такие как раковые заболевания кроветворной системы, а также некоторые аутоиммунные болезни, вирусные инфекции и паразитарные инфекции, но также и подходит для лечения болезней, связанных с заякоренными на клеточной мембране рецепторами иммуномодуляторов, например Т-клеточной лимфомы, экспрессирующей, например, рецептор IL-2. А. ИММУНОМОДУЛЯТОР IM В ФОРМУЛЕ IIM обозначает иммуномодулятор, который не является свободным или конъюгированным суперантигеном. Этот иммуномодулятор может представлять собой цитокин или хемокин. Примерами цитокинов являются: стимулирующий факторTNF), стимулирующий фактор (M-CSF) колонии макрофага, стимулирующий фактор (GCSF) зернистого лейкоцита, IL-1, IL-2, IL-4, IL6, IL-7, IL-12, IL-15, IL-18 и IGF. Примерами хемокинов являются С 5 а, IL-8, моноцитный хемотаксический белок 1 альфа (MIPlalpha) или моноцитный хемотаксический белок 1 (MIPl),моноцитный хемоаттрактантный белок 1 (МСР 1), моноцитный хемоаттрактантный белок 2(МСР-2), моноцитный хемоаттрактантный белок 3 (МСР-3), фактор активации тромбоцитов(PAFR),N-формил-метионил-лейцилфенилаланин (FMLPR), лейкотриен В 4 (LTB4R),высвобождающий гастрин пептид (GRP), RANTES, эотаксин, лимфотактин, IP10, I-309,ENA78, GCP-2, NAP-2, MGSA/gro, DC-CK1,Flt3L (эктопический домен), фракталькин, PF-4 и т.п. Другой тип иммуномодуляторов представляют собой такие иммуномодуляторы, которые происходят из заякоренных на клеточной мембране пар рецептор/лиганд, участвующих в модуляции запускаемого иммунного ответа, такого как костимуляция (например, рецепторы, связанные с поверхностью лимфоцитов и связанные с соответствующими клетками лиганды). В качестве примеров можно привести элементы,выбранные из пар CD40L/CD40, 4-BB1/4-BB1L,CD28/B7, CTLA-4/B7 и т.п. В 7 включает варианты, такие как CD80 и CD86, причем предпочтение отдается первому из них. Предпочтительными формами являются растворимые, содержащие внеклеточную часть (эктопический до 9 мен) и не имеющие внутриклеточных и заякоренных на мембране частей. Особо предпочтительные иммуномодуляторы способны обеспечивать действие супермутагенов in vivo, например, путем противодействия приобретению толерантности активированными супермутагеном Т-клетками. Типичными подходящими связанными с клетками рецепторами/лигандами являются CD28/B7, включая аналоги и фрагменты, как указано выше. Типичными цитокинами этой группы являются IL2, поскольку он представляет собой главный расположенный в прямом направлении считывания эффектор передачи сигнала CD28/B7, и подобные IL-2 цитокины IL-7 и IL-15. В парах,связанных с поверхностью Т-клеток рецепторов/лигандов, тот элемент, который не связан с Т-клеткой, подлежащей активированию, является предпочтительным для включения в конъюгат в соответствии с настоящим изобретением. Для CD40L/CD40, 4-BB1/4-BB1L и CD28/B7 это означает растворимые формы CD40, 4-BB1L и В 7 с указанными выше предпочтениями. В экспериментальной части настоящего описания приводятся примеры вариантов иммуномодуляторов, для которых было установлено, что они являются оптимальными для данного изобретения на момент даты приоритета. Иммуномодуляторы предпочтительно должны происходить из того же самого биологического вида, что и индивидуум, подлежащий лечению. Нативные иммуномодуляторы, такие как цитокины и хемокины, часто проявляют высокую системную токсичность и относительно короткий период полураспада у млекопитающих. Существуют многочисленные публикации, посвященные тому, как модифицировать иммуномодуляторы, чтобы они приобрели повышенную устойчивость к окислению, более длительный период полураспада in vivo, более низкую токсичность и улучшенную способность к разворачиванию при получении с помощью рекомбинантных технологий и т.п. Например, в патенте США 5229109 (Grim et al.) описываются малотоксичные аналоги IL-2, имеющие уменьшенное сродство к рецептору IL-2 с высоким сродством (IL-2R) за счет того, что у них отсутствует связывание с субъединицей р 55 этого рецептора. Эти аналоги получают в основном посредством мутирования кодона, кодирующего аминокислоту в позиции 33-46 в IL-2(например, Аrg38 Аlа и Phe42Lys или Phe42Ala). Мутант Asp20Lys имеет уменьшенное в 100-500 раз сродство к р 75/-цепи рецептора IL-2, без изменения его связывания с р 55 (Collins et al.,1988). Другие мутации, например, Asp20Ser,менее жесткие (Berndt et al., 1994). Изучение мышиного IL-2 указывает на то, что Asp84 и Аsn88 человеческого IL-2 также участвуют в связывании р 75 (Zuravski et al., 1993). Это предположение подтверждается моделированием 10 связывания между человеческим IL-2 и его рецептором (Bamborough et al., 1994). Ожидаемое уменьшение сродства этих мутаций выражается как Asp20Asn88Asp84. Объединение мутаций в Аrg38 и Phe42 с мутациями в положениях 88 и 20 приводит к еще более низкому сродству к IL-2R. Другая возможная и на дату приоритета предпочтительная мутация IL-2 - мутация Thr51Pro, в результате которой образуется аналог IL-2 с пониженной скоростью интернализации клеток и с увеличенной продолжительностью его иммуномодулирующего эффекта (Chang et al., 1996). Нумерация аминокислотных позиций приведена в соответствии с Taniguchi et al., 1983. Публикации авторов Grim et al., Collins etBamborough et al., 1994; Chang et al., 1996 и Taniguchi et al., 1983, включены в данное описание в качестве ссылки. Использование аналогов цитокинов и хемокинов с уменьшенным сродством к их нормальным рецепторам в описанных выше конъюгатах усиливает их связывание с предварительно выбранной клеткой-мишенью. Понятно, что цитокины и хемокины,в результате мутаций проявляющие уменьшенную скорость интернализации клеток в отношении связывания с их соответствующим клеточным рецептором и включенные в конъюгат в соответствии с формулой I, приводят к продлению активности суперантигенов по сравнению с соответствующими конъюгатами с нативной формой иммуномодулятора. Термин иммуномодулятор (IM), таким образом, охватывает любые модифицированные формы, например любые мутированные формы,которые способны агонизировать или антиагонизировать эффекты соответствующей нативной формы иммуномодулятора. В. Sag-ЧАСТЬ, ПРЕДСТАВЛЯЮЩАЯ СУПЕРАНТИГЕН В ФОРМУЛЕ ISag в формуле I представляет суперантиген,как это определено выше для свободных суперантигенов и в разделе под заголовком Известный уровень техники, т.е. суперантигены дикого типа, возможно модифицированные, например,путем мутации, для того, чтобы они а) имели уменьшенную способность связываться с антигенами ГКГ класса II по сравнению с соответствующим суперантигеном дикого типа (см., например, в публикации Abrahmsen etb) имели уменьшенную серологическую реактивность в сыворотке крови человека, по сравнению с соответствующим суперантигеном дикого типа (см., например, Abrahmsen et al.Antonsson et al., 1997); с) имели уменьшенную иммуногенность для человека по сравнению с соответствующим суперантигеном дикого типа (см., например, 11d) являлись химерами между двумя или более аналогичными суперантигенами дикого типа, в которых одна область в одном первом суперантигене дикого типа заменена соответствующей областью второго аналогичного суперантигена. Область, о которой идет речь, может представлять собой область, определяющую связывание с TCRV, например, как это определено для химер SEE/SEA и SEA/SEE (см. например, и Antonsson et al., WO 9736932; Antonsson et al., 1997; и Lamphaer et al., 1996). Сюда входят также и другие модификации/мутации, которые могут быть признаны пригодными, например, для того, чтобы не допустить нежелательного гликозилирования при продуцировании в эукариотных клетках. Типичными мутациями для суперантигенов, подобных SEA/SEE, в порядке их очередности, были следующими (нумерация согласно использованной Antonsson et al., 1997 и Antonsson et al., WO 9736932): а) уменьшенное связывание с ГКГ классаb и d) химеры между SEA и SEE, направленные на уменьшение серологической реактивности у человека и в то же время сохраняющие возможности SADCC соответствующего конъюгированного суперантигена: SEE с нижеследующими замещениями:Gly20Arg,Thr21Asn, Gly24Ser, Lys27Arg. В экспериментальной части использовали следующие мутанты: SEA (Asp227Ala) =Asp227Ala) = SEAm57. Предпочтительные суперантигены в названном порядке первоочередности выбирали из нижеследующих: 1) суперантигены (Sag), проявляющие сайты связывания с ГКГ класса II (например, стафилококковые энтеротоксины А и Е),2) суперантигены (Sag), которые в своей немутированной форме требуют ионов Zn для оптимального связывания с ГКГ класса II (например, SEA, SEE и SEH),3) cтафилококковые энтеротоксины. Нет необходимости в том, чтобы молекулаSag, которая должна быть включена в конъюгат по настоящему изобретению, была функциональным суперантигеном, главное требование чтобы конечный конъюгат был способен использовать механизмы SADCC и/или SDCC, как описано выше. С. Т-часть, обеспечивающая связывание с мишенью, в формуле I Т в принципе может представлять из себя любую структуру, способную связываться с поверхностными структурами клетки, предпоч 002688 12 тительно со структурами, специфичными для болезни. Структура, против которой направлена Т,обычно отличается (а) от эпитопа полиморфной цепи TCR, с которым связывается Sag, и (б) от эпитопов ГКГ класса II, с которыми связываетсяSag. Часть, обеспечивающую связывание с мишенью, можно выбрать среди интерлейкинов(например, интерлейкин-2), гормонов, антител,включая связывающиеся с гормонами фрагменты антител, ростовых факторов и т.п. См., например, публикацию Woodworth 1993 (включенную в данное описание в качестве ссылки). Часть, обеспечивающая связывание с мишенью, может, таким образом, представлять собой белок, содержащий 1, 2, 3 или 4 полипептидных цепи. На момент даты приоритета предпочтительным являлось, чтобы Т представляла собой антитело (полноразмерное антитело, Fab, F(ab)2,Fv, ScFv (одноцепочечное антитело), множественные одноцепочечные антитела (ScFv)n и любой другой связывающий антиген фрагмент антитела), включая любые функционально активные усеченные формы антител, упомянутые выше. Другие варианты являются моноспецифичными и биспецифичными. Антитело в принципе может быть направлено против любой болезни, связанной с/специфичной для поверхностных структур клетки, например для структур, связанных с любыми вышеприведенными формами рака, при этом особое значение имеют активные фрагменты антител (такие как Fab). Обычно антитело может быть направлено против эпитопа, специфичного для толстой кишки и/или поджелудочной железы, например против так называемого эпитопа С 242 (Lindholm et al,.WO 93/01303), против специфичного для рака легких эпитопа, например эпитопа для антитела 5 Т 4 (Stern et al., WO 89/07947), против специфичного для лимфомы эпитопа, например CD19,против специфичного для меланомы эпитопа,например HMW-MAA, и т.п. Термин антитело включает как моноклональные, так и поликлональные варианты,причем предпочтение отдается моноклональным препаратам. В случае, если часть, обеспечивающая связывание с мишенью, представляет собой фрагмент Fab, предпочтительно, чтобы остатки цистерны, в норме связывающие между собой тяжелые и легкие цепи Fab, были замещены аминокислотой, предотвращающей образование дисульфида, например серином. См. также Antonsson et al., WO 97/36932. Вышесказанное также включает в себя то,что Т может быть направлена и против необычных структур на более или менее здоровых клетках, которые регулируют или контролируют развитие болезни. Линкер В может быть выбран так, как описано ранее (Dohlsten et al., WO 92/01470;Abrahmsen et al. WO 96/01650; и Antonsson et al.,WO 97/36932), т.е. В предпочтительно должен быть гидрофильным и иметь одну или более структур, выбранных из амида, тиоэфира, дисульфида и т.п. Наиболее эффективными линкерами являются те, которые получены с помощью рекомбинантных технологий, т.е. конъюгация у них происходит на геномном уровне,приводя к получению олигопептидных линкеров. Как правило, олигопептидные линкеры содержат 1-30, например 1-20, аминокислотных остатков, которые предпочтительно выбирают так, чтобы линкер в целом был гидрофильным. Линкерные остатки поэтому предпочтительно выбирают из остатков гидрофильных аминокислот, таких как Gln, Ser, Gly, Glu, Pro, His и Аrg. Типичные олигопептидные линкеры включают трипептид GlyGlyPro или так называемый Qлинкер (публикация Wootton et al., 1989, включенная в настоящее описание в качестве ссылки), который может быть модифицирован с помощью gly-pro на аминном конце. Е. ТОЧКИ ПРИСОЕДИНЕНИЯ ДЛЯ Т, Sag и IM. Химические конъюгаты обычно содержат смесь молекул конъюгата, различающихся позициями связывания. Вещество конъюгата содержит как гетеро-, так и гомоконъюгаты. Для рекомбинатных конъюгатов (слитых белков) полученное вещество конъюгата является однородным в отношении позиции связывания. Для каждой отдельной субъединицы (Т,Sag, IM) аминный конец слит с карбоксильным концом другой субъединицы или наоборот,предпочтительно через встроенный олигопептидный мостик. Комбинации для случаев, когда конъюгат содержит по одной субъединице Т,IM, Sag, могут быть следующими: T-IM-Sag,IM-T-Sag, Sag-IM-T, Sag-T-IM, T-Sag-IM, IMSag-T (наличие линкерной структуры В не показано). В случае если одна или более субъединиц содержат две или более полипептидных цепей,количество возможностей увеличивается. Для случая, когда Т представляет собой фрагмент антитела Fab, эти возможности таковы (олигопептидные линкеры В не показаны): 1. Sag-Fab (легкая цепь)IМ В дополнительных вариантах иммуномодулятор и суперантиген могут быть слиты последовательно на любом конце любой из цепей антитела. 14 На момент даты приоритета были предпочтительными рекомбинантные конъюгаты, причем наибольшее предпочтение отдавалось Fabфрагментам в качестве частей, обеспечивающих связывание с мишенью, и связыванию аминного конца свободного суперантигена с первым константным доменом тяжелой (СHl) или легкой цепи антитела, а иммуномодулятора - с оставшимся карбоксильным концом (это справедливо для формулI-IV). Для оптимального продуцирования и функционирования, слитый белок экспрессируется рекомбинантно как двухцепочечный продукт, в котором суперантиген слит на С-конце с СHl-доменом Fab-фрагмента антитела через гибкий гидрофильный аминокислотный линкер,состоящий из 3-11 остатков. Этот линкер может иметь последовательность Gly-Gly-Pro или ProAla-Ser-Gly-Gly-Gly-Gly-Ala-Gly-Gly-Pro (Последов.19), либо 4-9 остатков, основанных на Последов.19, при этом Последов.19 предпочтительна. Часть, представляющая собой иммуномодулятор, сливается на С-конце с легкой цепью с помощью гидрофильного и нейтрального или положительно заряженного линкера, состоящего из 10-20 остатков (линкер Q). Предпочтительно, чтобы линкер Q имел следующие последовательности: Gly-Pro-Arg-GlnAla-Asn-Glu-Leu-Pro-Gly-Ala-Pro-Ser-Gln-GluGlu-Arg (Последов.23), Gly-Pro-Arg-Gln-SerAsn-Glu-Thr-Pro-Gly-Ser-Pro-Ser-Gln-Glu-GluArg (Последов.20), Gly-Pro-Arg-Gln-Ala-LysThr-Leu-Pro-Gly-Ala-Pro-Ser-Gln-Thr-Thr-Arg(Последов.22), причем Последов.20 и 21 являются наиболее предпочтительными (подробности см. в примере 2). Аналогичные комбинации в аминном конце или комбинацию мест присоединения в аминном и карбоксильном концах домена VH иVL на данной стадии исследований авторы считают приводящими к получению активных, но менее эффективных конъюгатов.F. АКТИВНЫЕ ЧАСТИ, НЕ СООТВЕТСТВУЮЩИЕ ФОРМУЛЕ I, НО ИСПОЛЬЗУЕМЫЕ СОГЛАСНО ВЫШЕУКАЗАННЫМ КОМБИНАЦИЯМ А-Е В СПОСОБЕ ПО НАСТОЯЩЕМУ ИЗОБРЕТЕНИЮ Свободные суперантигены (Sag): cм. в разделе под заголовком Определения. Типичные Sag приведены в разделах под заголовками Существующий уровень техники и В. Sagчасть, представляющая суперантиген в формулеI. Неконъюгированные иммуномодуляторы: cм. в разделе под заголовком Определения. В принципе можно использовать те же самые иммуномодуляторы, которые приведены в разделе под заголовком А. Иммуномодулятор 15 хемокины, особенно IL-2 и подобные IL-2 цитокины. Иммуномодуляторы по настоящему изобретению (Т, IМ-конъюгаты): эти конъюгаты соответствуют общей формулеIII в которой Т' и IM' связаны между собой с помощью органического линкера В'. Т', IM' и В' выбирают из тех же самых групп соединений/структур, что и Т, IM и В, х и z определяются таким же образом, как и в формуле I (у=0),при этом предпочтительно, чтобы приходилось по одному или два IM' на каждую Т' и каждую молекулу конъюгата. Места соединения между Т' и IM' таковы, как они определены для формулы I. См. также раздел под заголовком Е. Точки присоединения, в котором формулы 1-8 и комментарии к ним применимы также и к Т, IMконъюгатам, за исключением того, что опускается Sag. Конъюгаты формулы III можно получить в соответствии с известными способами,т.е. путем обычного химического связывания или с помощью рекомбинатных технологий(слитые белки), причем предпочтительно последнее (Fell et al., EP 439095; Rosenblum et al.,EP 396387). Особо важные Т, IM-конъюгаты включают IM'-часть, которая является модифицированной, например мутированной, получив в результате этого свойство уменьшенного сродства и/или уменьшенной скорости интернализации, как определено выше. Суперантигены по настоящему изобретению (Т, Sag-конъюгаты). Эти конъюгаты соответствуют общей формулеIV в которой Т и IM связаны между собой с помощью органических линкеров В. Т, Sag и В выбирают из тех же самых групп соединений/структур, что и Т, IM и В, х и у определяются таким же образом, как и в формуле I (z=0),при этом предпочтительно, чтобы приходилось по одному или два Sag на каждую Т и каждую молекулу конъюгата. Места соединения между Т' и Sag такие же, как они определены для формулы I. См. также раздел под заголовком Е. Точки присоединения, в котором формулы 18 и комментарии к ним применимы также и к Т,Sag-конъюгатам, за исключением того, что в формуле опускается IM. Конъюгаты формулы II можно получить в соответствии с известными способами, т.е. путем обычного химического связывания или с помощью рекомбинатных технологий (слитые белки), причем предпочтительно первое. См., например, Dohlsten et al.,WO 92/01470; Abrahmsen et al., WO 96/01650;Antonsson et al., WO 97/36932. Третий важный аспект изобретения: конъюгаты, содержащие модифицированный иммуномодулятор Эти новые конъюгаты соответствуют формулеV в которой T и Sag выбирают из тех же самых соединений, что и Т и Sag для формулы I. IM представляет собой иммуномодулятор, который был модифицирован, например, посредством мутации, таким образом, что он получил свойство проявлять пониженное сродство к своему заякоренному на клеточной мембране рецептору и/или пониженную скорость интернализации путем связывания с его рецептором (по сравнению с соответствующими нативными формами).IM предпочтительно представляет собой цитокин или хемокин. См. также раздел под заголовком А. Иммуномодулятор IM в формуле I. Одним из важных иммуномодуляторов для этого аспекта изобретения является модифицированный IL-2, х, у и z определяются таким же образом, как и в формуле I. В первом подаспекте конъюгатов в соответствии с формулой V, Т, Sag и IM всегда присутствуют (все х, у и z0), т.е. Т, Sag, IMконъюгаты, х, у и z обычно представляют собой целые числа от 1 до 3, предпочтительно 1-2. Обычные взаимосвязи между х, у и z таковы: х=у=z; х=у=0,5z; х=0,5 у=0,5z; и х=0,5 у=z. Во втором подаспекте конъюгатов в соответствии с формулой V часть, представленная суперантигеном, отсутствует (у=0), т.е. Т, IMконъюгаты, х и z обычно представляют собой целые числа от 1 до 3. Предпочтительные взаимосвязи между х и z следующие: х=z; х=0,5z; 0,5 х=z; х=1/3z и 1/3 х=z. В обоих подаспектах конъюгатов в соответствии с формулой V диапазоны, приведенные для целых чисел и их взаимосвязей, в первую очередь, относятся к слитым белкам, в которых часть, обеспечивающая связывание с мишенью, может представлять собой белок, содержащий 1, 2, 3 или 4 полипептидных цепи, и в которых имеется одна Т в каждой молекуле конъюгата. Точки присоединения в слитых белках формулы V те же самые, что и для соответствующих слитых белков формулы I. См. также раздел под заголовком Е. Точки присоединения, в котором формулы 1-8 и комментарии к ним применимы также и к Т, IM-конъюгатам, за исключением того, что для второго подаспектаSag опускается. Получение описанных выше конъюгатов Конъюгаты можно в принципе получать двумя основными способами. 1. Химическое связывание между собой отдельных субъединиц Т, Sag и IM. Каждую отдельную субъединицу или комбинацию их можно получить с помощью рекомбинантных технологий. 2. С помощью рекомбинантных технологий непосредственно получают конъюгат, описанный в любой из вышеприведенных формул. Фактически применяемые способы хорошо известны специалистам средней квалификации и включают большое количество вариантов. 17 При химическом связывании обычно используют функциональные группы (например первичные аминогруппы, карбоксильные группы, меркаптогруппы, углеводородные группы),которые, как правило, нативно присутствуют в нескольких позициях в суперантигенах, белковых иммуномодуляторах и белковых частях,обеспечивающих связывание с мишенью. Эти способы хорошо известны в данной области техники. Основной клеткой-хозяином для крупномасштабного рекомбинантного получения конъюгатов между суперантигеном и иммуномодулятором по настоящему изобретению (как слитых форм, так и неконъюгированных форм) является Е.coli. Этот хозяин в принципе обеспечивает получение рекомбинантов двумя путями: внутриклеточное продуцирование и секреция. Последний вариант предпочтителен, поскольку он предлагает выделение правильно уложенных складками белков из периплазмы и из культуральной среды. Вышесказанное не исключает того, что активные конъюгаты можно получать также и при использовании других клетокхозяев, например клеток эукариотов, таких как дрожжи или клетки млекопитающих. Предварительные результаты указывают на то, что эукариотные клетки, такие как клетки млекопитающих, могут быть предпочтительными для встраивания иммуномодуляторов, взаимодействующих с CD28, например В 7 и его аналогов. Четвертый аспект изобретения: генные конструкции, кодирующие новые конъюгаты по настоящему изобретению Четвертый аспект изобретения представляет собой рекомбинантную молекулу нуклеиновой кислоты, предпочтительно ДНК, такой, как кДНК или РНК, кодирующую описанные выше суперантиген и иммуномодулятор, предпочтительно IL-2, IL-7, IL-12, IL-15, IL-18, RANTES,эктопический домен CD80, эктопический доменCD86, 4-BB1L и Flt3L и их аналоги, описанные выше. В этом аспекте изобретения нуклеиновая кислота обычно содержит регуляторные регулярные последовательности, такие как трансляционные регуляторные последовательности,область начала репликации, последовательности секреторного сигнала, либо для одной из субъединиц - иммунорегулятора или суперантигена,либо для обеих этих субъединиц и т.п., совместимых с клеткой-хозяином, в котором должны экспрессироваться суперантиген и иммуномодулятор. Область между частями последовательностей, кодирующих суперантиген и иммуномодулятор, может включать последовательность, кодирующую аминоацильные сайты рибосом, такие как бисцистронная генная конструкция. Последнее позволяет осуществлять отдельную экспрессию каждой полипептидной цепи, включенной в конъюгат. В случае многоцепочечных комбинаций конъюгатов, содержащих IM,Sag,Т-конъюгат и соответствующие 18 сигнальные последовательности, бисцистронные конструкции облегчают складчатое сворачивание и сборку до полностью активных конъюгатов с IM, присоединенным к одной из цепей. Это важно для случаев, когда часть,обеспечивающая связывание с мишенью, представляет собой двухцепочечную молекулу антитела, и, по меньшей мере, один из суперантигенов (Sag) и иммуномодулятор сливают с терминальным концом цепи антитела (см. выше). ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, ДОЗИРОВКИ И СПОСОБЫ ВВЕДЕНИЯ Четвертый аспект настоящего изобретения представляет собой фармацевтическую композицию, содержащую комбинацию по данному изобретению суперантигена (Sag), обеспечивающую связывание с мишенью части (Т) и иммуномодулятора (IM). Характерным признаком этого аспекта изобретения является то, что, по меньшей мере, одна субъединица - суперантиген или иммуномодулятор - находится в форме конъюгата, позволяющего осуществлять связывание с клетками-мишенями, которые подлежат инактивации, как описано выше. Рассматриваемые композиции известны в данной области техники, за исключением того,что в данном случае они содержат комбинацию конъюгата по настоящему изобретению. В конкретной композиции эта комбинация может включать: а) тройной конъюгат, включающий суперантиген (Sag), обеспечивающую связывание с клетками-мишенями часть (Т) и иммуномодулятор (IM) (Т, IМ, Sag-конъюгат);b) два двойных конъюгата - один между обеспечивающим связывание с мишенью антителом (Т) и суперантигеном, а второй - между обеспечивающим связывание с мишенью антителом (Т') и иммуномодулятором (т.е. Т, Sagконъюгат + Т', IM-конъюгат). Т и Т' могут быть разными или одинаковыми, в зависимости от конкретного эпитопа; с) один двойной конъюгат между суперантигеном (Sag), обеспечивающей связывание с мишенью частью (Т) и иммуномодуляторомd) один двойной конъюгат между иммуномодулятором (IM), обеспечивающей связывание с мишенью частью (Т) и суперантигеном в свободной форме (Sag) (Т, IM-конъюгат + Sag); или е) суперантиген (Sag) и иммуномодулятор(IM) в форме двойного конъюгата (Sag, IMконъюгат). См. также выше, в контексте способов по настоящему изобретению. В случае если композиции содержат два активных компонента (b-d выше), то такая композиция позволяет поддерживать компоненты отдельными друг от друга вплоть до ее введения. Композиции могут быть в форме состоящего из частиц лиофилизированного материала, стерильного или асептически изготовленного раство 19 ра, таблеток, ампул и т.п. Могут также присутствовать носители, такие как вода, или другие, предпочтительно забуференные до физиологически приемлемого значения рН, например с помощьюPBS (физиологического раствора с фосфатным буфером), или другие инертные твердые или жидкие материалы. Как правило, композиции готовят из конъюгата, можно в комбинации с неконъюгированным активным компонентом, которые смешивают, растворяют, связывают или иным способом соединяют с одним или более водорастворимых или нерастворимых в воде водным или неводным носителем, если требуется, то вместе с подходящими добавками и адъювантами. Эти носители и условия ни в коем случае не должны отрицательно влиять на активность активных компонентов, определенных выше в пунктах а-е. Как правило, суперантигены (SAG) для использования по настоящему изобретению продают и вводят в виде предварительно диспергированных дозировок, каждая из которых содержит эффективное количество SAG, которое основано на представленных в настоящем описании результатах и которое находится в диапазоне от 1 пг до 50 мг. Точные дозировки разные для разных случаев и зависят от веса и возраста пациента, а также от пре-титра антител, специфичных для используемого SAG, способа введения, типа заболевания, от части,обеспечивающей связывание с мишенью, суперантигена, типа соединения (-В-), иммуномодулятора и т.п. Важным фактором, который следует принимать во внимание при определении дозы для комбинации, используемой в соответствии со способом по настоящему изобретению, является то, что суперантигены и иммуномодуляторы имеют оптимальные диапазоны концентраций. Слишком низкая доза приведет к отсутствию эффекта или к эффекту ниже оптимального, а слишком высокая доза приведет к нежелательным побочным действиям, таким как токсичность, которая может быть летальной. Таким образом, необходимо заострить внимание на том, что вышеприведенный широкий диапазон представляет собой лишь попытку охватить все возможные диапазоны для всех вариантов способа по настоящему изобретению. Таким образом, каждая конкретная комбинация в соответствии со способом по настоящему изобретению имеет свой субдиапазон доз в пределах диапазона от 0,1 пг до 50 мг. Это не исключает того,что будущие исследования и результаты могут привести к дозировкам, выходящим за пределы этого диапазона. Способы введения такие же, какие обычно используются в данной области, т.е. уничтожающее мишень эффективное количество или терапевтически активное количество комбинации суперантигена и иммуномодулятора в соответствии с настоящим изобретением приводят в контакт с клетками-мишенями. В соответствии с 20 вышеуказанным, это чаще всего означает парентеральное введение, такое как инъекции или вливания (подкожно, внутривенно, внутриартериально, внутримышечно, внутрибрюшинно) млекопитающему, такому как человек. Комбинацию конъюгата по настоящему изобретению можно вводить локально или системно. Под термином уничтожающее мишень эффективное количество имеется в виду такое количество, которое эффективно для активации Т-клеток и стимулирования их к разрушению клеток-мишеней. Предпочтительные способы введения на момент даты приоритета такие же самые, как и для конъюгатов суперантигенов согласно Dohlsten et al., WO 92/01470; Abrahmsen et al., WO 96/01650; и Antonsson et al., PCT 97/00537. Это означает 1-5-ти часовое внутривенное вливание(предпочтительно 4-х часовое) в день с жаропонижающим средством (парацетамолом). Введение повторяют в течение нескольких дней, например 5-8 дней, при этом тщательно учитывают возможный риск бустеринга антител, направленных против конъюгата. Оптимальным является проведение нескольких циклов терапии, причем каждый цикл включает введение лекарства в течение одного или более дней, за которым следует период отдыха, составляющий один или более дней, например циклы с введением лекарства и отдыхом, длящиеся 2 и 5 дней соответственно. Композиции по настоящему изобретению можно вводить либо в качестве основной терапии, либо в предпочтительных режимах в виде дополнительной терапии в комплексе с хирургическим лечением или с другими лекарствами. ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ Пояснения к фигурам Фиг. 1. FACS-анализ клеток CHO-CD28,окрашенных с CD80-C215Fab, CD80-C215FabSEAm57 или C215Fab-SEA с последующей инкубацией с антимышиным каппа-цепочечным моноклональным антителом, меченным РЕ. Окрашивание проводили при 4 С без промывок. Ордината: главный канал. Фиг. 2. FACS-анализ клеток Соlо 205, окрашенных с CD80-C215Fab, CD80-C215FabSEAm57 или C215Fab-SEA с последующей инкубацией с антимышиным каппа-цепочечным моноклональным антителом, меченным РЕ. Окрашивание проводили при 4 С с тремя промывками между стадиями окрашивания. Ордината: главный канал. Абсцисса: отношение эффекторных клеток (Е) к клеткам-мишеням (Т). Фиг. 3. Пролиферация. Т-клетки инкубировали с клетками Соlо 205 и 4 мкМ C242FabSEA, а также с различными количествамиCD80-C215Fab в течение 4 суток, после чего подсчитывали количество включенного 3Hтимидина. Активность, полученная без CDC215-Fab, составила 20039 импульсов/мин. +/1750.C242Fab-SEA, а также с различными количествами CD80-C215Fab в течение 4 суток, после чего собирали надосадочную жидкость и определяли количество IL-2. Количество IL-2, полученное без CD80-C215-Fab, составило 2849 пг/мл. Фиг. 5. Пролиферация. Т-клетки инкубировали с клетками Соlо 205 и с различными количествамиC215Fab-SEAm23 в течение 4 суток, после чего подсчитывали количество включенного 3Hтимидина. Фиг. 6. Продуцирование IL-2. Т-клетки инкубировали с клетками Соlо 205 и с различными количествами CD80-C215Fab-SEAm-57 или C215Fab-SEAm23 в течение 4 суток, после чего собирали надосадочную жидкость и определяли содержание IL-2. Фиг. 7. Пролиферативная способность Тклеток крови человека, инкубированных в течение 7 суток с указанными белками и с облученными трансфецированными клетками СНО (для представления SEA). Ордината: включение 3Hтимидина (импульсов/мин.). Фиг. 8. Одновременное введение SEA и IL2 ведет к повышению активации Т-клеток in vivo. Цитотоксичность (выраженная в процентах) против покрытых SEA Raji-клеток ГКГ класса II спленоцитов, взятых у мышей, получивших от 1 до 3 инъекций C215FabSEA (FS), C215-Fab-Q-hIL2(FSI). Отношение эффекторных клеток к клеткаммишеням составляло 30:1, а цитотоксичность измеряли с помощью стандартного анализа высвобождения 51 Сr за 4 ч. Фиг. 9. Терапия 5-суточных опухолей В 16 С 21- у мышей линии С 57 В 1/6, осуществлявшаяся с помощью трех инъекций (сутки 5, 6 и 7 после инокуляции опухоли) C215FabSEA (FS),C215Fab-Q-hIL2 (FI), C215FabSEA + C215FabQ-hIL2 (FS+FI) или C215FabSEA-Q-hIL2 (FSI). Использовали эквимолярные количества FabSEA и Fab-Q-hIL2. Абсцисса: количество инъецированного белка. Ордината: уменьшение опухоли, выраженное в процентах. Фиг. 10. Увеличенная инфильтрация опухоли Т-клетками CD-25+ после обработкиC215FabSEA-Q-hIL2. СD25-положительные клетки, инфильтрирующие легкие мышей,имеющих индуцированные опухоли легких B16GA733, определяли биоиммуногистохимически. Ордината: процент окрашенной области. Абсцисса: 1 = PBS; 2 = первая инъекция; 3 = вторая инъекция; 4 = третья инъекция; 5 = четвертая инъекция. Фиг. 11. Уровни окрашивания интерферона после проведения до 4 инъекций,один раз в день (37 мг на инъекцию),C215FabSEA-Q-hIL2 (FSI) (37 мг/инъекцию). 22 Через четыре часа после последней инъекции брали пробы крови и определяли содержание интерферона (показано на ординате) с помощью твердофазного иммуноферментного анализа(ИФА), с использованием в качестве стандарта рекомбинантного мышиного -интерферона(Pharmingen). Подобный эксперимент выполнили и с эквимолярными количествами (30 мг на инъекцию) C215FabSEA (FS). Фиг. 12. Цитотоксичность (выраженная в процентах и показанная на ординате) против покрытых SEA Raji-клеток ГКГ класса II+ спленоцитов, взятых у мышей, получивших PBS или 1, 4 или 6 инъекций C215FabSEAD227A-Q-hIL2(=FSm9-Il2). Цитотоксичность измеряли с помощью стандартного анализа высвобождения 51 Сr за 4 ч. PBS использовали в качестве отрицательного контроля. Фиг. 13. Терапия 3-х суточных опухолей В 16-С 215 у мышей линии С 57 В 1/6, после введения 8 инъекций C215FabSEAD227A (=FSm9) или C215FabSEAD227A-Q-hIL2 (=FSm9-Il2). Лечение проводили ежедневно в течение 8 последовательных дней. На 21 сутки животных умертвили и подсчитали легочные метастазы. Ордината: уменьшение опухоли, выраженное в процентах. Фиг. 14. Терапия 3-х суточных опухолей В 16-С 215 у трансгенных мышей линии Vb3(F42A, C215FabSEAD227A-Q-hIL2 (=FSm9-Il2) или C215FabSEAD227A (=FSm9). Лечение проводили ежедневно в течение 8 последовательных дней. На 21 сутки животных умертвили и подсчитали легочные метастазы. Ордината: уменьшение опухоли, выраженное в процентах. Фиг. 15. Терапия 3-суточных опухолей В 16-С 215 у трансгенных мышей линии Vb3C215FabSEAD227A-Q-hIL2F42A/D20S (F42A/D20S). Лечение проводили ежедневно в течение 8 последовательных дней. На 21 сутки животных умертвили и подсчитали легочные метастазы. Ордината: уменьшение опухоли, выраженное в процентах. ПРИМЕР 1. БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ СЛИТЫХ БЕЛКОВ CD80-C215Fab ДЛЯ ИСПОЛЬЗОВАНИЯ В КОСТИМУЛЯЦИИ ТЕРАПИИ ОПУХОЛИ С ПОМОЩЬЮ Fab, СВЯЗЫВАНИЕ КОТОРОГО С МИШЕНЬЮ БЫЛО ОБЕСПЕЧЕНО С ПОМОЩЬЮ Sag.CD80-C215Fab и CD80-C215Fab-SEAm57, содержащие внеклеточный домен человеческогоCD80. Эти слитые белки продуцировали в клетках млекопитающего и испытывали на связывание с CD28 с помощью клеточных трансфектантов СНО и антигена С 215, а также с помощью клеток Соlо 205. Оба слитых белка связывались 23 с CD28 и антигеном С 215, причем связывание с последним было уменьшенным в 100 раз по сравнению со связыванием с C215Fab-SEA. Поскольку CD80 присоединяется к N-концевой части L-цепи, то возможно, что CD80 препятствует связыванию с C215Fab. Оба слитых белка костимулировалиSAG-активированные Тклетки человека, что указывает на то, что растворимый CD80 сохраняет свои биологические функции. МАТЕРИАЛЫ И МЕТОДЫ Технологии рекомбинантной ДНК и ферменты: получение плазмидной ДНК и другие операции выполняли, по существу, согласно Сэмбруку и др. (Sambrook et al., 1989). В качестве штамма-хозяина использовали Е. coli HB101(Boyer et al., 1969). Рестрикционные эндонуклеазы и фрагмент Кленова ДНК-полимеразы I получили от Boehringer Mannhelm из New England Biolabs и использовали согласно рекомендациям поставщика. Taq-полимеразу получили от Perkin Elmer. кДНК получали из тотальной РНК с помощью набора GeneAmp RNA PCR kit(Perkin Elmer). Олигонуклеотиды синтезировали на установке Gene Assembler (Pharmacia BiotechAB) или на синтезаторе ABI 392 DNA/RNA(Applied Biosystems) и очищали с помощью хроматографии с обращенной фазой на системеFPLC (Pharmacia Biotech). Секвенирование осуществляли в соответствии с принципом дидезокситерминирования цепей (Sanger et al., 1977) с помощью набора Applied Biosystems DyeDeoxy Termination Cycle Sequencing Kit, а полученные продукты разделяли и определяли на секвенаторе ДНК модели ABI 373 А (AppliedBiosystems). Бактерии, несущие различные плазмиды, отбирали в чашках, содержащих среду 2 х YT и 15 г агаровой основы на литр, с добавкой 70 мг/л канамицина или 100 мг/л амплимицина. Жидкая питательная среда представляла собой 2 х YT (с содержанием на литр: 10 г дрожжевого экстракта (Difco), 16 г триптона Плазмидные конструкции Гены, кодирующие фрагмент Fab, содержащий вариабельные домены мышиного антитела С 215, собрали, как описано выше (Dohlstenet al., 1994). Клонирование и мутагенез гена стафилококкового энтеротоксина A (SEA), которое привело к получению замен F47A иD227A, описано ранее (Abrahmsen et al., 1995). Три другие мутации ввели в ген SEA с помощью праймеров LAKQ88-93 (все использованные олигонуклеотиды представлены в табл. 1) и получили мутант SEA, которому дали номер 57.LAKQ5 и LAKQ7 использовали в реакции ПЦР с обратной транскриптазой для введения в обратном направлении рамки Козака и для клонирования гена, кодирующего сигнальный пептид и внеклеточную часть человеческого CD80 из тотальной РНК селезенки человека. Было установлено, что эта нуклеотидная последовательность соответствует последовательности CD80 из банка генов (номер образца М 27533). Получили два варианта гена CD80, имеющие различные сайты клонирования на 3' конце: в отдельных реакциях ПЦР использовали праймерыBssHII или MroI соответственно. Группу слитых генов, кодирующих CD80, слитых перед каппацепью с помощью Q-линкера (Wootton et al.,1989), сконструировали путем встраивания ДНК-линкера LAKQ37/38 BssHII-Esp3I между соответствующим геном CD80 и сайтом DsaI,расположенным непосредственно перед геном каппа. Плазмиду pKGE987 получили путем встраивания слитых генов, кодирующих CD80(Q-линкер)-каппа, перед которым находится сигнальный пептид CD80, в вектор, который в дополнение к промотору CMV и поли-А-хвосту,содержит ген неомицина, используемый для отбора трансформантов. Последнюю версию гена CD80 использовали для конструирования группы слитых генов, в которой он предшествует группе слитых генов мутанта Fd-SEA за но 25 мером 57: фрагмент ДНК (LAKQ117/118), кодирующий Q-линкер, встроили между сайтом MroI в гене CD80 и сайтом PstI в кодоне 4 и 5 в гене С 215 VH. Эту группу слитых генов встроили во второй вектор CMV-промотора и получили плазмиду рМВ 189, таким образом закодировав сигнальный пептид CD80 и внеклеточную часть,вслед за которыми идет мутант Fd и SEA за номером 57, соединенный с помощью состоящего из трех остатков спейсера GGP. В плазмидеpKGE961 ген Fd вставили после сигнальной последовательности (происходящей из другого мышиного гена VH). Плазмида рМВ 165 кодирует нативную каппа-цепь, которой предшествует ее нативный сигнальный пептид, и содержит ген неомицина. Продуцирование Клетки эмбриональной почки 293 хомяка трансфецировали либо плазмидами pKGE961 иpKGE987, либо рМВ 156 и рМВ 189, в результате чего получили клеточные линии, продуцирующие CD80-C215Fab и CD80-C215Fab-SEAm57 соответственно. Для того, чтобы получить стабильные клеточные линии, использовали среду для отбора DMEM без красителя фенолового красного (Pharmacia no MS 0127) с добавкой Lглютамина (GIBCO BRL no 25030-24), 10% телячьей сыворотки и 1 мг/л генетицина. Средой для продуцирования была DMEM без фенолового красного с добавкой L-глютамина и 0,1%(PharmaciaUpjohn АВ, Швеция). Слитые белки выделяли из отфильтрованной (Sartobran 0,65-0,4 um) культуральной среды с помощью аффинной хроматографии на протеин Gсефарозе FF (Pharmacia Biotech AB), вслед за чем осуществляли aнти-CD80 аффинную очистку (иммобилизованное антитело против человеческого CD80, Camfolio L307.4) или ионнообменную хроматографию на SP-сефарозе FFKG, Berlin, Germany), 0,1 мг/л сульфата гентамицина (Biological Industries, Kibbutz Beit Haemek, Israel), 1 мМ пирувата натрия (JRH Biosciencies Industries, USA) и 10% инактивированной высокой температурой фетальной телячьей сыворотки (Gibco Middlesex, UK) использовали в качестве полной среды для всех клеточных культур. Антитела. Моноклональные антителаCD57 (HNK1) и CD56 (НВ 55), получили из продуцирующих mАВ клеток гибридомы (Американская коллекция тканевых культур (АТСС),Rockville,MD). Антимышиные каппацепочечные моноклональные антитела, меченные РЕ, получили от Becton Dickinson (San Jose,CA). 26 Клетки. Клетки K1 яичников китайского хомячка (СНО) трансфецировали человеческой кДНК, кодирующей ген CD28, в Pharmacia andUpjoin, Stockholm, Швеция. Трансфектанты обычным способом проанализировали на экспрессию CD28 и поддерживали путем FACSсортинга на близких уровнях экспрессии антигена. Клеточную линию карциномы толстой кишки человека Соlо 205 получили из АТСС. Все клеточные линии были свободны от микоплазмы. Анализ на пролиферацию Т-лимфоцитов. Т-клетки получали из мононуклеаров периферической крови (РВМ), как описано ранееCD57, HLA-DR4, CD14 и CD56. Все испытания на Т-клетках проводили при 0,1 х 106 клеток/лунку в объемах по 200 мкл в плоскодонных 96-луночных планшетах (Nunc, Roskilde,Дания). Синтез ДНК изучали после экспозиции культур с [3H]-тимидином [3H]TdR (0,5mCi/лунку), как описано ранее (10). Анализ с помощью проточной цитометрии. Проточный цитометрический анализ и сортинг осуществляли в соответствии со стандартной настройкой на проточном цитометре модели FACStarPlus (Becton Dickinson, Mountain View,CA). Из-за низкого сродства CD80 к CD28 окрашивание клеток CHO-CD28 со слитыми белками CD215Fab осуществляли, опуская промывку клеток. Анализ на цитокины. Продуцирование IL-2 анализировали методом ИФА, с помощью набора IL-2 ELIZA kit (huIL-2 Duoset, Genzyme). Результаты. Описание слитых белков CD80-Fab. Fabчасть представляет собой изотип мышиногоIgGl/k, хотя она содержит вариабельные доменыIgG2A/k-моноклонального антитела С 215 (Dohlsten et al., 1995). Трипептидная последовательность GGP следует за внутрице-почечным дисульфидом, образуя цистеин в домене СН 1. В тройном слитом белке CD80-C215Fab-SEAm57 это функционирует как спейсер между Fd и мутантом стафилококкового энтеротоксина A(SEA; Betley et al., 1988), имеющий шесть замещений. Замены F47A и D227A встроили для того, чтобы уменьшить сродство к ГКГ класса II(Abrahmsen et al., 1995), а замены N102Q,N149D и T218V ввели для того, чтобы не допустить гликозилирования при продуцировании в эукариотных клетках. Эти последние замены выбраны с помощью рентгеновской структуры(Sundstrom et al., 1996). Окончательный пентамутант обозначили как SEA-мутант 57. Оба слитых белка содержат внеклеточный доменCD80 человека (обозначенный как FPDN). Использовали нативный сигнальный пептид CD80 и установили, что зрелый белок запускаетVIHV, что определили с помощью секвенирования аминокислот очищенного CD80-C215Fab. 27 Состоящий из 18 аминокислот спейсер соединяет CD80 с каппа-цепью в CD80-C215Fab или сFd-частью Fab-фрагмента в тройном слитом белке CD80-CD215Fab-SEAm57. Эти спейсеры напоминают Q-линкер (Wooton et al., 1996) и имеют последовательности SARQANELPGAPSQEERA (Последов.15) и SARQANELPGAPSQEERP (Последов.16), соответственно.CHO-CD28 окрашивали с CD80-C215Fab, CD80C215Fab-SEAm57 или с C215Fab-SEA, а затем инкубировали с антимышиным каппацепочечным моноклональным антителом, меченным РЕ. Окрашивание проводили при 4 С без промывок. Как CD80-C215Fab, так и тройной слитый белок связывались с CD28, экспрессируемым на клетках СНО, при этом связывание зависело от дозы. Как и ожидалось, не наблюдалось окрашивания с контрольным слитым белкомC215Fab-SEA. С 215-положительные клетки. Связывание слитых белков против антигена С 215 оценивали против клеток Соlо 205. Клетки Соlо 205 окрашивали с CD-80-C215Fab, CD80-C215FabSEAm57 или с C215Fab-SEA, а затем инкубировали с антимышиным каппа-цепочечным моноклональным антителом, меченным РЕ. Окрашивание проводили при 4 С с тремя промывками между стадиями окрашивания. Результаты (фигуры 1-2): Как CD80 C215Fab, так и CD80-C215Fab-SEAm57 связывались с антиген С 215-положительными клетками Соlо 205, при этом связывание зависело от дозы. Связывание было в 50-100 раз ниже, чем для C215Fab-SEA. Это указывает на то, что встраивание CD80 в N-концевую часть слитого белка может препятствовать связываниюC215Fab с антигеном С 215. Двойные слияния. Костимуляция активированных суперантигеном Т-клеток. Для того, чтобы определить биологическую активность слитых белков, их испытывали на костимуляцию активированных суперантигеном Т-клеток. Пролиферация: Т-клетки инкубировали с клетками Соlо 201 и с 4 мкМ C242Fab-SEA, а также с различными количествами CD80C215Fab, в течение 4 суток, после чего измеряли пролиферацию, по количеству включенного 3Hтимидина. Активность, полученная без CD80C215Fab составляла 20039 отсчетов/мин. +/1750. Продуцирование IL-2: Т-клетки инкубировали с клетками Соlо 205 и с 4 мкМ C242FabSEA, а также с различными количествами 28 собирали надосадочную жидкость и определяли количество IL-2. Количество IL-2, полученное без CD80-C215Fab, составляло 2849 пг/мл. Результаты (фигуры 3-4): CD80-C215Fab оказывал костимулирующее действие на индуцированную C242Fab-SEA активацию Т-клеток,причем степень стимуляции зависела от дозы,как для пролиферации, так и для продуцирования IL-2. Тройные слияния. Костимуляция активированных суперантигеном Т-клеток. Для того, чтобы определить биологическую активность тройного слитого белка, очищенные Т-клетки инкубировали с C215FabSEAm23 (причем m23 представлял собой такой же мутант SEA, влияющий на связывание с ГКГ класса II, как и тот, что использовался для тройного слитого белка, т.е. Phe47Ala/Asp227Ala),или с CD80-C215Fab-SEAm57, представленным на клетках Соlо 205. Пролиферация: количество включенного 3H-тимидина подсчитывали через 4 дня. Продуцирование IL-2: Собирали надосадочную жидкость и определяли содержание IL-2 через 4 дня. Результаты (фигуры 5-6): Тройной слитый белок индуцировал активацию Т-клеток и продуцирование IL-2, когда он присутствовал на клетках Соlо 205. Подобной активности сC215Fab-SEAm23 не наблюдалось, что указывает на важность костимуляции с помощью CD80 для активации. Из-за более низкого сродства тройного слитого белка (в 50-100 раз ниже, чем для C215Fab-SEA, см. данные анализа FACS),фактическое количество C215Fab-SEAm23, связанного с клетками, существенно выше, чем количество тройного слитого белка. ПРИМЕР 2. IL-2 no настоящему изобретению усиливает и продлевает действие FabSEA на активацию Т-клеток и на лечение опухолей Методы и материалы: Конструирование плазмид, экспрессирующих IL2. кДНК IL2 клонировали с помощью ПЦР с обратной транскрипцией, используя мРНК, изолированную из мононуклеаров периферической крови человека (РВМ), которые были простимулированы суперантигеном SEA в течение 24 ч. мРНК выделяли из 5 х 106 клеток, с помощью набора mRNA Direct kit Dynal, Oslo, в соответствии с инструкциями изготовителя. мРНК, отожженную до покрытых олиго(dТ)26 магнитных шариков, элюировали путем нагревания до 95 С. Вслед за этим с помощью ПЦР с обратной транскрипцией получали продукт ПЦР, обладающий преимуществами обратнотранскрип-тазной и ДНК-полимеразной активности Taq-полимеразы. 1/10 элюированной мРНК смешивали с ПЦР-праймерами IL2-1 иIL2-2 и проводили стандартную реакцию ПЦР(30 раундов). Продукт ПЦР подвергали электрофорезу в агарозном геле, полосу извлекали из геля и очищали с помощью набора Prep-a 29EcoRI/BamHI. Эту вставку секвенировали и подтверждали ее идентичность кДНК IL2, ранее описанной Taniguchi et al.(1983). Однако в результате ПЦР-реакции сегмент ДНК, кодирующий Gly-Pro-Arg-Gln-Ala-Asn-Glu-Leu-Pro-GlyAla-Pro-Ser-Gln-Glu-Glu-Arg (Последов.23) Gly-Pro-Q-линкер - был добавлен в позиции 5' к сегменту, кодирующему зрелый человеческийIL-2. QhIL2 клонировали в плазмиду из собственной коллекции (разрезанную с помощьюC215FabSEA-Q-hIL2 в периплазму E.coli. Линкеры между этими частями далее оптимизировали следующим образом. Кодирующую линкер область, которая кодирует последовательность Рrо-Ala-Ser-Gly-Gly-Gly-Gly-Ala-Gly-Gly-Pro(Последов.19) (заменяющую исходную GlyGly-Pro), встроили между суперантигеном и областями, кодирующими Fab-части, с помощью сайт-направленного мутагенеза. Подобным же образом, линкеры Gly-Pro-Arg-Gln-Ser-AsnGlu-Thr-Pro-Gly-Ser-Pro-Ser-Gln-Glu-Glu-Arg(Последов.20), Gly-Pro-Arg-Gln-Ala-Lys-ThrLeu-Pro-Gly-Ala-Pro-Ser-Gln-Thr-Thr-Arg (Последов.21) или Gly-Pro-Thr-Glu-Ala-Asp-GluLeu-Prp-Gly-Ala-Pro-Ser-Glu-Glu-Glu-Thr (Последов.22) заменили оригинальный Q-линкер между IL-2 и Fab-частью. Для комбинаций двойных слитых белков сравнивали также конструкции с IL-2, слитым с тяжелой и легкой цепью, соответственно. Праймеры: Экспрессия слитых белков в ферментере. Слитые белки экспрессировали в E.coli K-12,штамм U1 635 (xyl-7, ara-14, T4R, deltaompT), с помощью плазмиды с геном устойчивости к канамицину и lacUV5-промотором. Бактерии из замороженного образца инкубировали при 25 С примерно в течение 21 ч на встряхивателе, в колбах, содержащих, г/л: (NH4)2SO4, 2,5;MgSO4.7H2O, 1; моногидрат глюкозы, 11, 0,11 мМ канамицина и 1 мг/л раствора микроэлементов (Forsberg et al., 1989), но без Na2MoO42 Н 2 О. Клетки выращивали до оптической плотности ОП 600, равной 1-2, и 450 мл культуральной среды использовали для инокуляции ферментера(Chemap, Швейцария), доведя до окончательного объема в 5 л. Среда в ферментере содержала,г/л: (NH4)2SO4, 2,5; KH2PO4, 9; K2HPO4, 6; цит 002688MgSO47H2O, 1; 0,11 мМ канамицина; 1 мл адеканола (Asahi Denka Kogyo K.K., Япония) и 1 мг/л раствора микроэлементов. рН поддерживали на уровне 7,0, с помощью титрования 25%ным аммонием, температура составляла 25 С, а аэрацию осуществляли с помощью атмосферного воздуха, со скоростью 5 л/мин. Парциальное давление растворенного O2 доводили до 30% путем увеличения скорости перемешивания с 300 до 1000 оборот./мин во время периодической фазы культивирования и регулирования подачи 60% (по объему) глюкозы во время фазы дозирования. Образование продукта индуцировали при ОП 600, равном 50, путем добавления 0,1 мМ изопропилD-тиогалактопиранозида(IPTG). После ферментации клетки удаляли путем центрифугирования при 8000 хg в течение 40 мин при 4 С. Осветленную среду либо анализировали и осуществляли выделение сразу,либо хранили при -20 С. Выделение слитых белков. ДНК, присутствующую в осветленной среде, удаляли из нее с помощью осаждения 0,19%-ным полиэтиленимином и 0,2 М NaCl в течение 30 мин (Atkinson and Jack, 1973). После центрифугирования,как описано выше, собирали над-осадочную жидкость, а концентрацию NaCl доводили до 0,5 М. Эту среду использовали для колонки с протеин G-сефарозой (Pharmacia Biotech AB,Uppsala, Швеция), уравновесив ее с помощью 10 мМ фосфата натрия, 150 мМ NaCl с рН 7,4 и сPBST с 0,05%-ным содержанием Твина 80. Затем колонку дважды промывали PBST в количестве 5 объемов колонки, и связанный белок элюировали 0,1 М уксусной кислотой с 0,02% Твина при рН 3,2. рН образца доводили до 5,0 с помощью 1 М Tris-HCl с рН 8,0, а затем помещали в HP-колонку с SP-сефарозой (PharmaciaBiotech), уравновешенную с помощью 50 мМ ацетата аммония, 0,02% Твина 80. Затем колонку промывали уравновешивающим буфером в количестве 2 объемов колонки, и слитый белок элюировали, используя линейный градиент от 50 до 500 мМ ацетата аммония, количество которого составляло свыше 10 объемов колонки. Для слитых белков C215Fab-IL2 использовали рН 6,0, а для тройных слитых белков C215FabSEA-IL2 значение рН во время разделения составляло 5,7. Слитые белки фильтровали через фильтр 0,22 um и хранили при -70 С. Если элюат был недостаточно концентрированным, его концентрировали до окончательной концентрации в 0,5-1 мг/мл с помощью устройства Centricon 30 (Amicon) в соответствии с инструкциями изготовителя. Анализы на цитотоксичность. Анализы на цитотоксичность, зависимую и независимую от ГКГ класса II, проводили как описано ранее(Dohlsten et al., 1990). Вкратце, для анализов на зависимую от ГКГ класса II цитотоксичность 31 меченные 51 Сr клетки Raji (по 2500 клеток на лунку в окончательном объеме в 200 мкл) смешивали с SEA-зависимой линией эффекторных клеток, полученной путем инкубации РВМ человека в присутствии низких уровней рекомбинантного hIL-2. Отношение эффекторных клеток к клеткам-мишеням (Е:Т) составляло 30:1. Для анализов на независимую от ГКГ класса II цитотоксичность меченные 51 Сr клетки С 215 соlо 205SEAзависимыми эффекторными клетками при соотношении Е:Т, составлявшем 45:1. Количество выделившегося в среду 51 Сr определяли после 4 ч инкубации с помощью сцинтилляционного счетчика. Анализ костимуляции in vitro на очищенных Т-клетках человека. Не подвергавшиеся воздействиям Т-клетки человека очищали в основном так, как описано в публикацииLando et al. (1993). Вкратце, не подвергавшиеся воздействиям Т-клетки человека очищали от РВМ человеческой крови с помощью центрифугирования с градиентом фиколла, с последующим разделением на желатиновых колонках, и,наконец, путем пэннинга с негативной селекцией в чашках Петри, содержащих моноклональные антитела HNK1 и HLA-DR. Эксперименты по пролиферации с использованием не подвергавшихся воздействиям Т-клеток человека проводили в основном так, как описано в публикации Lando et al. (1993). Вкратце, не подвергавшиеся воздействиям Т-клетки (по 100000 клеток на лунку) объединяли с облученными СНОтрансфектантами (по 10000 клеток на лунку) в общем объеме в 200 мкл среды RPMI-1640, с добавками (Lando et al., 1993). Для экспериментов со слитыми белками, содержащими IL-2,клетки инкубировали в течение 7 суток в присутствии 1 нМ указанных веществ. В последний день эксперимента клетки метили 3Hтимидином, чтобы измерить включение его в ДНК делящихся клеток. Терапия опухолей В 16-С 215. Терапию проводили по существу так, как описано ранее(Dohlsten et al., 1994; Hansson et al., 1997). В нулевой день эксперимента мышам линии С 57 Вl/6 внутривенно (в хвостовую вену) инъецировали 75000-150000 сингенных клеток меланомы B16F10. Эти клетки В-16 экспрессировали человеческий антиген GA-733, распознаваемый моноклональными антителами С 215. В дни 1, 3 или 5 начинали лечение белками C215Fab. На 21-й день эксперимент закончили, и в этот день удалили легкие и подсчитали количество рассеянных опухолей легких. Иммуногистохимические исследования проводили на легких животных, несущих опухоли В 16-С 215 18-го дня, по существу так, как описано ранее (Dohlsten et al., 1995). Образцы брали через 4 ч после последней инъекции. Окрашенные области определяли вручную. 32 Иммунофармакологические исследования проводили по существу так, как описано в публикации Rosendahl et al. (1996). Селезенки мышей линии С 57 В 1/6, получивших от 1 до 3 инъекций, по одной в день, удаляли через 48 ч после последней инъекции и определяли SEAзависимую цитотоксичность, используя Rajiклетки в качестве мишеней, в стандартном анализе выделения 51 Сr, как описано выше. Отношение Е:Т составляло 100:1. РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ: Продуцирование и выделение IL-2 содержащих слитых белков. Сконструировали вектор экспрессии Е.coli,кодирующий тройной слитый белокC215FabSEA-Q-hIL2. Этот вектор кодирует две субъединицы тройного слитого белка на бицистронной мРНК, транскрибированной с промотора LacUV5. Каждой из этих субъединиц предшествует сигнальный пептид, направляющий экспорт к периплаз-матическому пространству Е.coli. Первая субъединица представляет собойVH белка C215Fab, за которым следует линкерGly-Gly-Pro (VH-СHl-Gly-Gly-Pro-SEA). Другая субъединица представляет собой VK белкаC242Fab, за которым следует СK белка(Последов.23) представляет собой слегка модифицированную версию естественного линкера, найденного в белке OmpR E.coli (Woottonet al., 1989). Более полную информацию см. в разделе Материалы и методы. Получили также слитый белок C215Fab-Q-hIL2. Он идентичен слитому белку C215FabSEA-Q-hIL2, за исключением того, что часть Gly-Gly-Pro-SEA из этого белка удалена. Соответствующую последовательность ДНК в векторе экспрессии также удалили. Мутантные производные слитого белка C215FabSEA-Q-hIL2 получили с помощью опосредованного реакцией ПЦР сайтнаправленного мутагенеза, с помощью стандартных способов, при этом получили ряд белков, таких как C215FabSEAD227A-Q-hIL2 иC215FabSEAD227A-Q-hIL2F42A. В более поздних вариантах тройного слитого белка внутрисубъединичный цистеин заменили двумя остатками серина. Это изменение не повлияло на биологическую активность. Плазмидами,кодирующимиIL2 содержащие белки, трансформировали продуцирующий штамм Е.coli UL635, а затем осуществили ферментацию. Слитые белки выделили из культуральной среды с помощью протеин Gаффинной хроматографии. Деградированные варианты слитых белков удалили с помощью ионообменной хроматографии. Полученные продукты представляли собой, по меньшей мере, на 90% полноразмерный слитый белок, что определили с помощью электрофореза в полиакриламидном геле (SDS-PAGE) (см. раздел 33 Материалы и методы). С помощью оптимальной схемы получения слитого белка в ростовой среде получали до 130 мг/л слитого белка и, как правило, из 1 л среды получали 70 мг тройного слитого белка. Функциональная характеризация IL-2 содержащих Fab-слитых белков. СпособностьIL2-содержащих слитых белков, таких как слитые белки C215FabSEA-Q-hIL2 и C215Fab-QhIL2, индуцировать пролиферацию IL-2 зависимой линии CTLL-2 клеток мышей была,по существу, такой же, как у рекомбинантного человеческого IL2 на молярной основе (данные не показаны). Более того, связывание с антигеном и SEA-активность C215FabSEA-Q-hIL2 иC215FabSEA оказались неразличимыми в ряде анализов, что подтверждает, что введение IL2 в молекулу не оказывает отрицательного действия. Эти анализы включали исследование возможности индуцировать независимое от ГКГ класса II уничтожение клеточной линии соlо 205 рака толстой кишки человека С 215 с помощьюSEA-реактивной эффекторной клеточной линии Т-клеток, в анализе четырехчасового выделения 51 Сr. Зависимое от ГКГ класса II уничтожениеSEA-реактивными эффекторными Тклетками также происходило с подобной же эффективностью (данные не показаны). Более прямые доказательства безусловного связывания антигена C215 и ГКГ класса II получили с помощью FACS-анализа. Установили, что степень зависимости от дозы связыванияC215FabSEA и C215FabSEA-Q-hIL2 с Rajiклетками подобны между собой на молярной основе (данные не показаны). Установили также, что зависимое от дозы связываниеC215FabSEA (данные не показаны), что указывает на то, что связывание с антигеном в IL2 содержащих слитых белках не вызывает сомнений.IL-2-зависимая пролиферация, индуцированная FabSEA. Для запуска оптимальной пролиферации и активации Т-клеток человека,покоящимся Т-клеткам требуются как сигнал 1,так и сигнал 2 (Schwartz, 1990). Суперантигены,если они присутствуют на клетке, могут доставить сигнал 1. При этом предполагают, что IL-2,будучи основным расположенным в прямом направлении считывания генетического кода эффектором сигнализации B7/CD28, подает сигнал 2. В данном разделе авторы показывают с помощью покоящихся Т-клеток, выделенных из крови человека, что тройной слитый белокC215FabSEA-Q-hIL2 или C215FabSEA в комбинации с C215Fab-Q-hIL2 или с рекомбинантным человеческим IL2 действительно индуцирует пролиферацию Т-клеток in vitro (фиг. 7).(C215FabSEA или C215FabSEA-Q-hIL2) в присутствии IL2 (в форме C215Fab-Q-hIL2,C215FabSEA-Q-hIL2 или рекомбинантного человеческого IL-2). SEA был представлен на облученных клетках СНО, трансфецированных антигеном С 215 (через Fab-часть), с ГКГ классаII/Dr, который связывает SEA. Контролем служили нетрансфецированные СНО-клетки. После 7 суток инкубации Т-клеток с трансфектантами СНО и с упомянутыми выше веществами измеряли пролиферацию путем включения 3Hтимидина в ДНК.C215FabSEA-Q-hIL2 индуцировал пролиферацию Т-клеток человека, когда он присутствовал на СНО-клетках, трансфецированных антигеном С 215 человека (СНО-С 215), против которых направлено действие Fab (фиг. 7). Подобным же образом пролиферация индуцировалась, когда белок присутствовал на CHO-Dr(ГКГ человека, класс II), в то время как в присутствии нетрансфецированных СНО-клетокC215Fab-Q-hIL2 не индуцировали скольконибудь заметной пролиферации сами по себе,когда они присутствовали на СНО-клетках, что указывает на то, что как SEA, так и IL-2 действительно необходимы для индукции пролиферации. Это было подтверждено тем, что комбинация C215FabSEA и C215Fab-Q-hIL2 илиIL-2 является необходимым, но в отличие от SEA,он не обязательно должен быть связанным с клеткой.C215FabSEA-Q-hIL2 так же, как и комбинация C215FabSEA и C215Fab-Q-hIL2, вызывают усиленную и длительную активацию Т-клеток и улучшенную инфильтрацию опухолей in vivo. Как C215Fab-Q-hIL2 +C215FabSEA, так и C215FabSEA-Q-hIL2 гораздо более эффективно индуцировали активацию Тклеток, чем C215FabSEA, что измерялось способностью индуцировать SEA-зависимое уничтожение клеток-мишеней (фиг. 8). Вкратце,мыши получали от 1 до 3 инъекций указанных белков в сутки. Через 2 суток после последней инъекции у подвергавшихся лечению мышей извлекали селезенки и определяли цитотоксическую активность против Raji-клеток, покрытыхSEA, с помощью стандартного анализа, основанного на четырехчасовом выделении 51Cr. Как C215FabSEA-Q-hIL2, так и 215FabSEA/C215Fab-Q-hIL2 индуцировали не только повышенную, но и более длительную активацию Тклеток. Уровень содержания сывороточных цитокинов, таких как IFNgamma, содержание которого резко падало после четвертой инъекцииC215FabSEA, оставалось на очень высоком уровне даже после четвертой инъекции C215FabSEA-QhIL2 (данные не показаны). Как правило, уровни содержания IFNgamma и TNFalpha были гораздо выше (до 10 раз), чем в случае C215FabSEA. Улучшенная терапия индуцированных опухолей В 16-С 215 при лечении с помощью комбинации C215FabSEA и C215Fab-Q-hIL2. Влияние IL-2 на повышение эффективности FabSag-терапии опухолей исследовали на модели мышиной меланомы В 16 (фиг. 9). В показанном примере 8-12-недельных самок мышей линии С 57 В 1/6 в нулевые сутки эксперимента инокулировали клетками В 16 (150000 шт.),трансфецированными антигеном GA-733 опухоли человека, против которого направлено антитело С 215 (ниже называемыми В 16-С 215). Указанные вещества инъецировали на 5, 6 и 7 сутки. На каждую дату обработано по 7 животных. На 21 сутки животных умертвили и подсчитали количество пигментированных меланином опухолей В-16, колонизировавших легкие. В нескольких экспериментах показали, что комбинация C215FabSEA и C215Fab-Q-hIL2 оказывает более высокий терапевтический эффект, чемC215FabSEA, по сравнению с не получавшими лечения контрольными животными (фиг. 9). В указанном эксперименте комбинацияC215FabSEA/C215Fab-Q-hIL2 давала более высокий терапевтический эффект, чем тройной слитый белок C215FabSEA-Q-hIL2. Последующие иммунохимические исследования показали, что C215FabSEA в отдельности или в комбинации C215FabSEA и C215FabQ-hIL2 приводил к образованию близких количеств CD4- и CD8-T-клеток, инфильтрующих опухоль В 16-С 215. Однако интересно отметить,что количество CD25 (IL2Ra)-положительных клеток - хороший маркер активации Т-клеток резко увеличивалось между 3-ей и 4-ой инъекциями C215FabSEA/C215Fab-Q-hIL2 (фигура 10). В противоположность этому, это количество уменьшалось в случае, если появлялись признаки того, что C215FabSEA начинает вызывать толерантность. Таким образом, качество инфильтрующих Т-клеток, по-видимому, выше в случае комбинационного лечения, чем при применении одного только C215FabSEA, что может служить объяснением улучшенного терапевтического эффекта. Подобным же образом, количество сывороточных цитокинов, таких как интерферон, продуцируемых вторично по отношению к активации Т-клеток, заметно уменьшалось после четвертой инъекцииC215FabSEA (фиг. 11). Однако при применении тройного слитого белка FabSEA-Q-hIL2 содержание интерферона оставалось на высоком уровне даже после четвертой инъекции. Это наблюдение дает дополнительные доказательства того, что включение IL-2 в конструкцию может противодействовать появлению толерантности Т-клеток, индуцируемых Fab-SEA. 36 Улучшенная терапия опухолей В 16 С 215 с помощью С 215FabSEAD227A-Q-hIL2. Суперантигены гораздо более токсичны для человека, чем для мышей, в частности из-за того, что сродство к ГКГ класса II у человека значительно выше (Hansson et al., 1997). Поэтому вполне вероятно, что системная токсичность белков FabSEA является главным ограничением для лечения человека. Одним из способов усиления локальной активации в опухоли (Fab-зависимой) в противовес системной иммуноактивации (зависимой от SEA-ГКГ класса II) является уменьшение сродства SEA к ГКГ класса II. Основываясь на кристаллической структуре SEA, авторы настоящего изобретения получили мутантC215FabSEAD227A, с уменьшенным в 100 раз сродством к ГКГ класса II. В отличие от дикого типа C215FabSEA этот мутант не обладает свойством поперечно связываться с молекулами ГКГ класса II, что считают основной причиной опосредованной SEA системной токсичности(Hansson et al., 1997). Окно между эффективной терапией и токсичностью в первые сутки лечения опухолей В 16-С 215 у трансгенных мышей линии Vb3TCR было, по меньшей мере, в 50 раз шире для мутанта C215FabSEAD227A по сравнению сC215FabSEA (Hansson et al., 1997). В соответствии с этим наблюдением иммуногистохимически установили, что при дозах этих двух белков,приводящих к близким терапевтическим результатам, наблюдалась сравнимая иммуноактивация в опухоли, в то время как при применении C215FabSEAD227A наблюдалась намного меньшая системная иммуноактивация в селезенке (Hansson et al., 1997). Кроме того, фармакокинетические исследования на кроликах показали резкое уменьшение связыванияC215FabSEAD227A с мишенями в селезенке и других лимфоидных органах, где расположено большинство клеток ГКГ класса II, по сравнению с C215FabSEA (данные не показаны). Полагают, что для клинического использования тройной слитый белок Fab-SEA-IL2 должен содержать мутированный суперантиген. Поэтому авторы настоящего изобретения получили тройной слитый белок C215FabSEAD227AQ-hIL2. Полученный белок был способен индуцировать пролиферацию покоящихся Т-клеток человека (данные не показаны) и индуцировал длительную активность SEA-зависимых ЦТЛ у мышей в продолжение до 6 инъекций (фиг. 12). В противоположность этому фоновая активность ЦТЛ наблюдалась при применении С 215FabSЕАD227 А (10% - данные не показаны) и C215Fab-Q-hIL2 (15% - данные не показаны). Терапия индуцированных опухолей B16-GA733(3-и сутки) у нормальных мышей линии С 57 В 1/6, получивших по 8 инъекций этого белка,вводимых ежедневно, привела к таким же хо 37 рошим терапевтическим результатам, как и при оптимальной дозе C215FabSEA (фиг. 13). В этой системе С 215FabSЕАD227 А, который предназначен для получения максимальной активности у человека, а не у мышей, давал только очень невысокий эффект (фиг. 13). Однако при самых высоких дозах была отмечена некоторая токсичность C215FabSEAD227A-Q-hIL2. В противоположность этому несколько экспериментов по лечению указывают на то, что комбинацияC215FabSEAD227A и C215Fab-Q-hIL2 (8 инъекций) также ведет к существенно улучшенному терапевтическому эффекту на 3-и сутки для опухолей B16-GA733 у трансгенных мышей линии VB3 TCR по сравнению с применением только одного C215FabSEAD227A (данные не показаны). У трансгенных мышей линии VB3 TCR 90% Т-клеток реагировали с SEA по сравнению с 10-20% у нормальных мышей. Интересно отметить, что для кроликов повторные инъекции (до 8 инъекций) C215FabSEAD227A-Q-hIL2 (0 из 4 кроликов погибли после повторных инъекций в дозе 20 мкг/кг), по-видимому, не более токсичны, чем C215FabSEAD227A без IL-2 (0 из 4 кроликов погибли после повторных инъекций в дозе 20 мкг/кг; 4 из 4 погибли при дозе 20 мкг/кг), и гораздо менее токсичны, чемC215FabSEA (1 из 4 кроликов погиб после повторных инъекций в дозе 1 мкг/кг). В то же самое время длительная им-муноактивация (эффект повторного связывания лимфоцитов) наблюдалась только после лечения с помощьюC215FabSEAD227 или C215FabSEA, что указывает на то, что включение IL-2 действительно помогает противодействовать индуцируемой FabSEA толерантности Т-клеток (данные не показаны). Терапия опухолей В 16-С 215 с помощью мутированных по IL-2 белков C215FabSEAD227A-Q-hIL2. Более высокая токсичность, а в случае с C215FabSEAD227A-Q-hIL2 и более низкая терапевтическая эффективность тройных слитых белков, содержащих IL-2, может быть частично обусловлена перенацеливанием активности SEA на лимфоидные ткани. Этот эффект перенацеливания приписывают высокому сродству IL2 (Kd = 10 пМ) к его рецептору, который в основном находится на Т-клетках в селезенке и в других лимфоидных тканях. Чтобы изучить этот вопрос, авторы настоящего изобретения получили производные отC215FabSEAD227A-Q-hIL2, в которых IL-2-часть является мутированной, для того, чтобы уменьшить сродство к клеткам IL2R+ и таким образом уменьшить системную активацию. В принципе,это такой же самый подход, какой был использован для улучшения терапевтического окна для белка C215FabSEAD227A. Взаимодействие между IL-2 и его высокоаффиным рецептором изучено очень хорошо. Этот рецептор состоит из трех субъединиц: а, b 38 и g. Для биологической активности требуется поперечное связывание b и g, а субъединица а необходима для оптимального связывания. Стратегия авторов изобретения состояла в том,чтобы уменьшить сродство IL-2 к его высокоафинному рецептору (abg) путем устранения связывания с а-субъединицей, а затем уменьшения, но не устранения, связывания с субъединицами b и g по мере необходимости. Три таких мутанта белкаC215FabSEAD227A-Q-hIL2 создали с целью устранить связывание с ILRa (F42A, F42K) и, кроме того, чтобы ослабить связывание с IL2Rb(F42A/D20S). Эти белки имеют в 100 раз(F42A/D20S) уменьшенную активность соответственно в отношении индукции пролиферацииCTLL-2. В настоящее время продолжаются эксперименты, направленные на более подробное изучение свойств этих белков, в частности, в отношении их сродства и интенсивности связывания с активированными Т-клетками человека и с рецептором IL2. Первые эксперименты по терапии показывают, что такая стратегия является перспективной (фиг. 14, 15). В показанных примерах по 8 инъекций ввели трансгенным мышам линии Vb3-TCR (где 90% Т-клеток были активированы SEA), несущим 3-х суточные опухоли B16-GA733. Хотя при самой высокой дозе все же наблюдалась некоторая токсичность при использованииC215FabSEAD227A-Q-hIL2F42A или С 215FаbSЕАD227A-Q-hIL2F42K, она отмечалась позже (8-ая инъекция), чем при использованииC215FabSEAD227A-Q-hIL2 (6-ая инъекция), а лечение с использованием самой высокой дозы постоянно приводило к уменьшению опухолей более чем на 90% по сравнению с контрольными животными, получавшими PBS (физиологический раствор с фосфатным буфером) (фиг. 14). Авторы изобретения в настоящее время применяют этот многообещающий подход путем введения дополнительных мутаций в частиSЕА и IL2 для того, чтобы еще более улучшить окно между терапевтическими и токсичными дозировками. Кроме того, в настоящее время проводится работа по созданию тройных слитых белковThr51Pro в IL-2-части (Chang et al., 1996). Эта мутация может служить для блокировки опосредованной IL2R интернализации слитых белков без снижения биоактивности IL-2. Это представляется очень важным, поскольку при этом уменьшится удаление слитого белка этим путем и за счет этого увеличится локальная концентрация и эффективность лекарства. Белки с мутацией Thr51Pro могут также содержать и другие мутации в частях SEA и IL-2, которые уменьшают сродство к ГКГ класса II и к рецептору IL2 соответственно. 39 ПРИМЕР 3. Эксперименты для подтверждения действия (Sag,IM)-конъюгатов.IM-рецептор-положительные клеткимишени или ГКГ класс II-положительные клетки. Молекула Sag-IM может связываться с клетками, экспрессирующими ГКГ класса II,таким образом способствуя SDCC. И наоборот,клетки, экспрессирующие рецептор для IM, могут быть мишенями. Поэтому возможно, что молекула Sag-IM может быть полезной для индуцирования уничтожения нежелательных клеток, экспрессирующих рецептор для IM. Тройной слитый белок C215FabSEA-hIL2 можно рассматривать как молекулу Sag-IM, в случае, если ни эффектор, ни клетка-мишень не экспрессируют антиген, который С 215Fаb может распознать. Эффекторные клетки должны быть Т-клетками правого субтипа V. Клеткимишени могут быть, например, аберрантными кроветворными клетками, экспрессирующими рецептор IL-2, например, такими, которые имеются в некоторых лейкозах или лимфомах. Эксперимент А: Доказательство концепции in vitro: Эффекторные Т-клетки (Т-клетки человека, неоднократно стимулированные SEA) и клетки-мишени (например, Raji В-клетки лимфомы человека) инкубируют с тройным слитым белком C215FabSEA-IL2, подвергнутым мутагенезу для уменьшения связывания с ГКГ класса I. Это стандартный набор для анализов наSDCC (эффекторные Т-клетки, Raji-клетки, экспериментальное вещество). Авторы настоящего изобретения установили, что белок С 215FabSЕАD227 АhIL2 с резко уменьшенным сродством к ГКГ классаII имеет примерно в 10 раз более высокую эффективность уничтоженияRaji-клеток,чем С 215FаbSЕАD227 А. Очевидное объяснение этой повышенной эффективности состоит в том, что тройной слитый белок представлен на рецепторах IL2, экспрессируемых на Raji-клетках. Эксперимент В: Доказательство концепции in vivo. Модели лимфомы мышей (такие,как лимфома RBL-5 мышей линии С 57 В 1/6) хорошо разработаны (Hoglund et al., J. Exp. Med. 168, 1469-1474). Связывание с мишенями рецептора IL-2 или другого IM-R с белком Sag-IM в таких моделях может обеспечить доказательство концепции о связывании с мишенями IMрецепторположитель-ных клеток in vivo. Связывание с Sag-IM IM-R-положительных клеток осложняется тем фактом, что как эффекторные клетки, так и клетки-мишени часто экспрессируют рецептор для IM. Тем не менее, в определенных обстоятельствах такой режим терапии может быть эффективным. Повидимому, имеют значение такие факторы, как плотность экспрессии IM-R на клеткахмишенях, количество и расположение клетокмишеней и т.п. Следует также подчеркнуть, что,например, IL2R alpha регулируется позитивно только в отношении активации Т-клеток, и, та 002688 40 ким образом, может существовать временное окно, во время которого IM-R экспрессируется в высокой степени на клетках-мишенях, но не на эффекторных клетках. Слитые белки Sag-IM, в которых Sag используется вместо суперантигена дикого типа,который был подвергнут мутагенезу для уменьшения сродства с ГКГ класса II, могут также использоваться подобным образом. ССЫЛКИ Статьи, отмеченные знаком , можно считать в большей степени относящимися к иммуномодуляторам, чем другие.Zurawski et al. (1993) EMBO J. 12:5113. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ инактивации клеток-мишеней в присутствии Т-клеток, включающий контактирование клеток мишеней и Т-клеток с суперантигеном в присутствии иммуномодулятора, когда, по меньшей мере, один из элементов - суперантиген или иммуномодулятор - конъюгирован с нацеливающим компонентом, причем нацеливающий компонент выбран из группы, состоящей из антитела, антигенсвязывающего фрагмента антитела, Fab-фрагмента антитела,Fаb2-фрагмента антитела или одноцепочечного антитела, а указанный суперантиген модифицирован, таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида, и указанный иммуномодулятор выбирают из группы, содержащей цитокины, хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов. 2. Способ по п.1, отличающийся тем, что суперантиген и иммуномодулятор оба конъюгированы с одним и тем же компонентом мишени в форме тройного конъюгата. 3. Способ по п.1, отличающийся тем, что конъюгированы только суперантиген и нацеливающий компонент, в то время как иммуномодулятор не конъюгирован. 4. Способ по п.1, отличающийся тем, что клетки-мишени инактивируют in vivo в индивидууме, страдающем заболеванием, ассоциированным с клетками-мишенями. 5. Способ по п.4, отличающийся тем, что заболеванием является рак. 6. Способ по п.1, отличающийся тем, что суперантиген представляет собой химеру, содержащую последовательность, производную из двух или более суперантигенов дикого вида. 7. Способ по п.1, отличающийся тем, что иммуномодулятор представляет собой IL-2. 8. Способ по п.1, отличающийся тем, что иммуномодулятор представляет собой внеклеточную часть молекулы В 7. 9. Способ по п.8, отличающийся тем, что внеклеточную часть молекулы В 7 выбирают из группы, содержащей CD80 и CD86. 10. Способ по п.1, отличающийся тем, что иммуномодулятор модифицирован с целью уменьшения сродства рецепторов лимфоцитов 44 по сравнению со сродством соответствующей нативной формы. 11. Конъюгат суперантигена, имеющий формулу(T)x(Sag)y(IM)z,где у 0 и z0; Т представляет собой нацеливающий компонент, выбран из группы, состоящей из антитела, антигенсвязывающего фрагмента антитела, Fab-фрагмента антитела, Fab2-фрагмента антитела или одноцепочечного антитела,Sag представляет собой суперантиген, который модифицирован таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида,IM представляет собой иммуномодулятор,выбранный из группы, содержащей цитокины,хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов. 12. Конъюгат суперантигена по п.11, где 10 х 0, 10 у 1 и 10z1. 13. Конъюгат суперантигена по п.12, где х,у и z представляют собой целые числа 1-3. 14. Конъюгат суперантигена по п.11, где Т содержит, по крайней мере, Т' и Т", суперантиген слит С-концом с Т', а иммуномодулятор слит С-концом с Т". 15. Конъюгат суперантигена по п.11, где суперантиген представляет собой химеру, содержащую последовательность, производную из двух или более суперантигенов дикого вида. 16. Конъюгат суперантигена по п.11, где иммуномодулятор представляет собой IL-6. 17. Конъюгат суперантигена по п.11, где иммуномодулятор представляет собой внеклеточную часть молекулы В 7. 18. Конъюгат суперантигена по п.17, где внеклеточная часть молекулы В 7 выбирают из группы, содержащей CD80 и CD86. 19. Конъюгат суперантигена по п.11, где иммуномодулятор модифицирован с целью уменьшения сродства рецепторов лимфоцитов по сравнению со сродством соответствующей нативной формы. 20. Конъюгат суперантигена по п.14, где суперантиген представляет собой стафилококковый энтеротоксин А, Т' является СH1 доменом С 215 Fab, Т" является легкой цепью антитела С 215, а иммуномодулятор представляет собойIL-2. 21. Конъюгат суперантигена по п.14, где суперантиген слит с Т' с помощью гибкого гидрофильного аминокислотного линкера В из 3-11 аминокислотных остатков, а иммуномодулятор слит с Т" через гидрофильный нейтральный или положительно заряженный аминокислотный линкер Q из 10-20 аминокислотных остатков. 45 22. Конъюгат суперантигена по п.21, где В выбирают из группы, состоящей из Gly-Gly-Pro и Pro-Ala-Ser-Gly-Gly-Gly-Gly-Ala-Gly-Gly-Pro(SEQ ID N 19), а Q выбирают из группы, состоящей из Gly-Pro-Arg-Gln-Ala-Asn-Glu-LeuPro-Gly-Ala-Pro-Ser-Gln-Glu-Glu-Arg (SEQ ID N 23), Gly-Pro-Arg-Gln-Ser-Asn-Glu-Thr-Pro-GlySer-Pro-Ser-Gln-Glu-Glu-Arg (SEQ ID N 20), GlyPro-Arg-Gln-Ala-Lys-Thr-Leu-Pro-Gly-Ala-ProSer-Gln-Thr-Thr-Arg (SEQ ID N 21) и Gly-ProThr-Glu-Ala-Asp-Glu-Leu-Pro-Gly-Ala-Pro-SerGlu-Glu-Glu-Thr (SEQ ID N 22). 23. Конъюгат суперантигена по п.11, где х=0, у=1-3, a z=1-3. 24. Молекула ДНК, кодирующая суперантиген и иммуномодулятор не являющийся суперантигеном, которая находится в форме бицистронной конструкции, в которой первый цистрон содержит последовательность, кодирующую суперантиген, модифицированный таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида,и второй цистрон содержит последовательность, кодирующую иммуномодулятор,выбранный из группы, содержащей цитокины,хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов. 25. Молекула ДНК по п.24, где иммуномодулятором является IL-2. 26. Молекула ДНК по п.24, где указанный кодируемый иммуномодулятор представляет собой внеклеточную часть молекулы В 7, выбранную из группы состоящей из CD80 и CD86. 27. Молекула ДНК, кодирующая суперантиген модифицированный таким образом, чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида; иммуномодулятор, выбранный из группы,содержащей цитокины, хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов, нацеливающий компонент, выбранный из группы, состоящей из антитела, антигенсвязывающего фрагмента ан 002688 46 титела, Fab-фрагмента антитела, Fab2-фрагмента антитела или одноцепочечного антитела. 28. Молекула ДНК по п.27, где иммуномодулятор является IL-2. 29. Молекула ДНК по п.27, где иммуномодулятор представляет собой внеклеточную часть молекулы В 7 и выбран из группы, содержащей CD80 и CD86. 30. Фармацевтическая композиция для инактивации клеток-мишеней, содержащая суперантиген, иммуномодулятор и нацеливающий компонент, где, по меньшей мере, один из элементов - суперантиген или иммуномодулятор конъюгирован с нацеливающим компонентом,причем указанный нацеливающий компонент выбран из группы, состоящей из антитела, антигенсвязывающего фрагмента антитела, Fabфрагмента антитела, Fab2-фрагмента антитела или одноцепочечного антитела, а указанный суперантиген модифицирован таким образом,чтобы он имел уменьшенную способность к связыванию с антигеном ГКГ класса II и уменьшенную серологическую реактивность и иммуногенность в сыворотке крови человека по сравнению с соответствующим суперантигеном дикого вида, и указанный иммуномодулятор выбирают из группы, содержащей цитокины,хемокины и внеклеточные части связанных с поверхностью лимфоцита рецепторов и лигандов, причем указанная композиция взята в эффективном количестве. 31. Фармацевтическая композиция по п.30,в которой суперантиген представляет собой химеру, содержащую последовательность, производную из двух или более суперантигенов дикого вида. 32. Фармацевтическая композиция по п.30,в которой иммуномодулятором является IL-2. 33. Фармацевтическая композиция по п.30,в которой иммуномодулятор представляет собой внеклеточную часть молекулы В 7. 34. Фармацевтическая композиция по п.33,в которой внеклеточная часть молекулы В 7 выбрана из группы, содержащей CD80 и CD86. 35. Фармацевтическая композиция по п.30,в которой иммуномодулятор модифицирован таким образом, чтобы уменьшить сродство рецепторов лимфоцитов по сравнению со сродством соответствующей нативной формы.
МПК / Метки
МПК: A61K 39/385, A61K 39/085, C07K 14/31, A61K 47/48
Метки: клеток-мишеней, используемые, способы, агентов, вызывающие, соединения, агенты, получения, инактивации, этих, цитолиз, композиции
Код ссылки
<a href="https://eas.patents.su/27-2688-sposoby-inaktivacii-kletok-mishenejj-agenty-i-kompozicii-vyzyvayushhie-citoliz-i-soedineniya-ispolzuemye-dlya-polucheniya-etih-agentov.html" rel="bookmark" title="База патентов Евразийского Союза">Способы инактивации клеток-мишеней, агенты и композиции, вызывающие цитолиз, и соединения, используемые для получения этих агентов</a>
Трициклические соединения, обладающие активностью в отношении интегринов, в частности, в отношении интегрина альфаvбета 3, способ их получения и промежуточные соединения, используемые в этом способе,их применение в качестве медикаментов и содержащие их фармацевтические композиции.

Номер патента: 2271
Опубликовано: 28.02.2002
Авторы: Бернар Серж, Питти Роберт М., Бодари Сара С., Карниато Дени, Гадек Томас Р., Штильц Ханс-Ульрих, Кнолле Йохен, Гурвест Жан-Франсуа, Венер Фолькмар, Тетш Жан-Жорж, Макдауэлл Роберт С.
МПК: A61K 31/19, A61P 9/10, C07C 281/12...
Метки: соединения, способе,их, качестве, способ, используемые, трициклические, активностью, интегринов, этом, частности, получения, содержащие, медикаментов, композиции, промежуточные, применение, альфаvбета, фармацевтические, обладающие, отношении, интегрина
Формула / Реферат:
1. Соединения общей формулы (I) в которой R1 обозначает группу -О-[А]-[В]-COR6, в которой R6 обозначает -ОН, C1-С6алкокси, -О-СН2-СН(ОН)-СН2OН, [A] обозначает группу C1-С6алкилен, возможно замещенный оксогруппой, [B] обозначает радикал -CH(Z)- или простую связь, Z обозначает группу -NHCO2Rc, или -NHSO2Rc, где Rc обозначает радикал фенил(C1-С4)алкил-, хинолинил или пиридинилимидазолил(C1-С4)алкил-, R2 и R3, одинаковые или разные, обозначают атом...
Четырехциклические конденсированные соединения с гетероатомами, замещенные арилом, промежуточные соединения, способы получения, композиции и методы лечения

Номер патента: 1649
Опубликовано: 25.06.2001
Автор: Гриз Тимоти А.
МПК: A61P 5/30, A61K 31/35, C07D 471/00...
Метки: гетероатомами, композиции, конденсированные, четырехциклические, соединения, получения, способы, арилом, промежуточные, замещенные, лечения, методы
Формула / Реферат:
1. Соединение формулы I или II где Y обозначает -О-, -S-, -СН2-, -СН2СН2-, -СН=СН- или -NR4-; В обозначает -СН2- или -СО-; R1 и R2 обозначают каждый независимо -Н, -ОН, -O(C1-C4алкил), -ОСОС6Н5, -ОСО(С1-С6алкил), -ОSО2(С4-С6алкил), OSO2СF3, Сl или F; n равно 1 или 2; W обозначает -СН2- или >С=O; R3 обозначает 1-пиперидинил, 2-оксо-1-пиперидинил, 1-пирролидинил, метил-1-пирролидинил, диметил-1-пирролидинил, 2-оксо-1-пирролидинил,...
Мутантный il-6 человека и его внутренний фрагмент, кодирующие их последовательности днк, способы их получения, содержащие их фармацевтические композиции, содержащие их векторы, линии клеток- хозяев испособ лечения il-6 опосредованных заболеваний

Номер патента: 852
Опубликовано: 26.06.2000
Авторы: Гротзингер Йоахим, Элерс Марк, Розе-Йон Штефан
МПК: A61P 19/10, A61P 35/00, A61K 38/20...
Метки: внутренний, лечения, хозяев, человека, испособ, кодирующие, мутантный, получения, линии, композиции, последовательности, содержащие, фрагмент, клеток, днк, заболеваний, векторы, способы, фармацевтические, опосредованных
Формула / Реферат:
1. Мутантный интерлейкин-6 (IL-6) человека, имеющий аминокислотную последовательность содержащую следующие точечные мутации по сравнению с природным IL-6 человека: Pro 54, Glu 159, Pro 162, Leu 170 и Аrg 176. 2. Внутренний фрагмент мутантного IL-6 человека по п.1 формулы, обладающий аналогичной биологической активностью. 3. Последовательность ДНК, кодирующая мутантный IL-6 человека по п.1 формулы. 4. Последовательность ДНК, кодирующая...
Производные хиноксалин-2,3-диона и их фармацевтически приемлемые соли, фармацевтические композиции, обладающие антагонистической активностью в отношении рецептора глютамата и противосудорожной активностью, способы лечения пациентов, страдающих от удара или заболеваний, при помощи этих соединений.

Номер патента: 762
Опубликовано: 24.04.2000
Авторы: Никам Шам, Рафферти Майкл Фрэнсис, Корнберг Брайэн Эдвард
МПК: C07D 241/44
Метки: композиции, пациентов, соединений, отношении, приемлемые, хиноксалин-2,3-диона, обладающие, противосудорожной, способы, помощи, фармацевтические, производные, соли, заболеваний, страдающих, активностью, удара, фармацевтически, глютамата, этих, антагонистической, рецептора, лечения
Формула / Реферат:
1. Производные хиноксалин-2,3-диона общей формулы I где R - группа - NR4R5, где R4 и R5 независимо друг от друга означают водород, низший алкил, незамещенный или замещенный низшим алкилом, низший циклоалкил, который может содержать 1 или 2 атома кислорода в качестве гетероатома, R1 и R2 независимо друг от друга означают водород или нитрогруппу, R3 - низший алкил, при этом радикалы R3 и R-CH2- могут каждый находиться в положениях 5 или...
Изолируемые смеси из семян eugenia jambolana lamarck, способы их получения, способ получения оксамата натрия, лекарственные средства для профилактики и лечения диабета и применение указанных смесей, их компонентов и оксамата натрия для получения этих лекарственных средств

Номер патента: 1463
Опубликовано: 23.04.2001
Авторы: Лебуль Жан, Ракото Ратсимаманга Сюзанн, Рейсдорф Даниель, Ракото Ратсимаманга Альберт, Расоанаиво Филипп, Прово Жан
МПК: A61K 35/78, A61P 5/50
Метки: смесей, указанных, оксамата, eugenia, лекарственных, способы, диабета, способ, компонентов, jambolana, lamarck, натрия, средства, лечения, профилактики, средств, лекарственные, этих, смеси, семян, применение, изолируемые, получения
Формула / Реферат:
1. Смесь, не содержащая полифенольных и стерольных производных и изолируемая посредством измельчения семян EUGENIA JAMBOLANA LAMARCK, мацерации полученного порошка низшим алифатическим спиртом в горячем состоянии, фильтрации, выделения нерастворимой фракции, не содержащей более полифенольных и стерольных соединений, мацерации нерастворимой фракции с помощью низшего алифатического спирта, фильтрации и выделения нерастворимой фракции, не...
Предыдущий патент: Производные бензотиофена, их применение и композиции
Следующий патент: Способ лечения оппозиционного расстройства
Случайный патент: Способ получения гранулированного сорбента