Конструкции casb7439
Номер патента: 20617
Опубликовано: 30.12.2014
Авторы: Блэйс Норманд, Риу Клеман, Харви Мартин, Пилорге Энтони





























Формула / Реферат
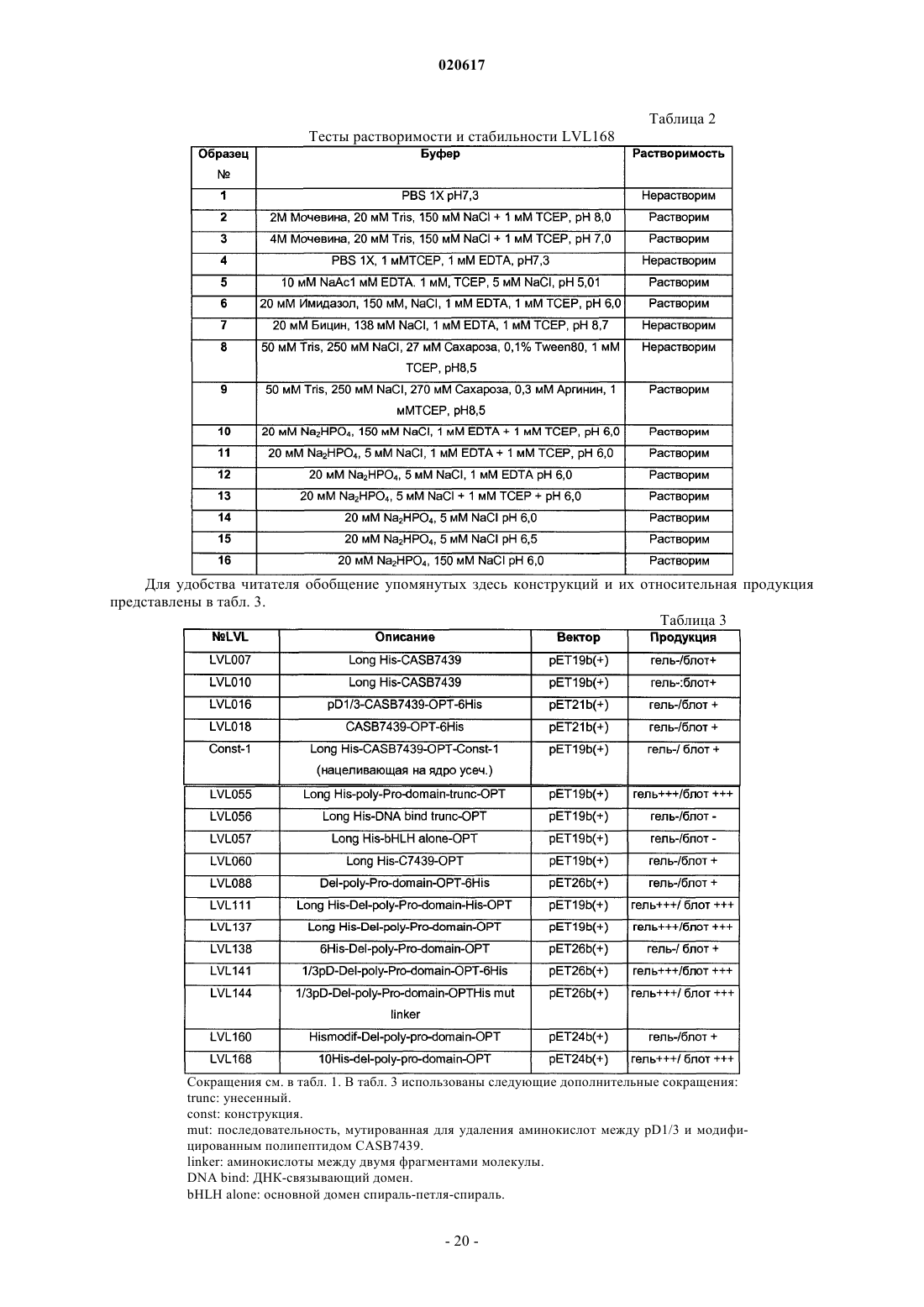
1. Белковая конструкция для индукции иммунного ответа против CASB7439, включающая модифицированный полипептид CASB7439 (SEQ ID NO: 13) и выбранная из:
(а) LVL055 (SEQ ID NO: 1);
(б) LVL111 (SEQ ID NO: 3);
(в) LVL137 (SEQ ID NO: 5);
(г) LVL141 (SEQ ID NO: 7);
(д) LVL144 (SEQ ID NO: 9);
(е) LVL168 (SEQ ID NO: 11).
2. Иммуногенная композиция, содержащая белковую конструкцию по п.1 и фармацевтически приемлемый носитель или эксципиент, где носитель или эксципиент могут, возможно, содержать буфер.
3. Иммуногенная композиция по п.2, дополнительно содержащая адъювант.
4. Иммуногенная композиция по п.3, где адъювант содержит по меньшей мере одно из следующего: агонист TLR-4 (Toll-подобный рецептор), иммунологически активная фракция сапонина, агонист TLR-9.
5. Иммуногенная композиция по п.4, где указанный агонист TLR-4 представляет собой 3D-MPL (монофосфориллипид), и/или указанный агонист TLR-9 представляет собой CpG олигонуклеотид, и/или указанная иммунологически активная фракция сапонина представляет собой QS21.
6. Иммуногенная композиция по п.5, содержащая 3D-MPL, и/или CpG, и/или QS21.
7. Иммуногенная композиция по п.6, дополнительно содержащая холестерин.
8. Молекула нуклеиновой кислоты, содержащая полинуклеотидную последовательность, кодирующую белковую конструкцию по п.1.
9. Способ индукции иммунного ответа на CASB7439 у человека или животного, отличного от человека, при котором человеку или животному, отличному от человека, вводят эффективное количество композиции, содержащей адъювант и белок, включающий полипептидную последовательность, выбранную из группы, состоящей из:
(а) полипептидной последовательности, представленной в SEQ ID NO: 9; и
(б) полипептидной последовательности, представленной в SEQ ID NO: 11.
10. Способ лечения человека или животного, отличного от человека, при котором осуществляют следующие стадии:
(а) выбор человека или животного, отличного от человека, имеющего раковые клетки, экспрессирующие CASB7439; и
(б) введение этому человеку или животному, отличному от человека, эффективного количества композиции, содержащей адъювант и белок, включающий полипептид, выбранный из группы, состоящей из:
(1) конструкции, представленной в SEQ ID NO: 9; и
(2) конструкции, представленной в SEQ ID NO: 11.
11. Применение иммуногенной композиции по любому из пп.2-7 в способе индукции иммунного ответа на CASB7439 у человека или животного, отличного от человека, где указанный способ включает введение эффективного количества указанной иммуногенной композиции.
12. Применение по п.11, где иммуногенная композиция содержит белок, включающий полипептид, выбранный из группы, состоящей из:
(а) конструкции, представленной в SEQ ID NO: 9; и
(б) конструкции, представленной в SEQ ID NO: 11.
13. Применение иммуногенной композиции по пп.2-7 в изготовлении лекарственного средства для использования в способе индукции иммунного ответа на CASB7439 у человека или животного, отличного от человека, где указанный способ включает введение эффективного количества указанной иммуногенной композиции.
Текст
Изобретение относится к соединениям и способам увеличения рекомбинантной продукции полипептидов CASB7439 и к способам их применения. Предшествующий уровень техники Гомологи Achaete-Scute млекопитающих представляют собой консервативных родственников млекопитающих комплекса Achaete-Scute дрозофилы. Они являются членами основного семейства генов спираль-петля-спираль (bHLH или HLH), которые действуют как специфичные для линий клеток транскрипционные факторы, необходимые для развития. Один из членов этого семейства MASH2, обнаруженный у мышей, необходим для развития плаценты, но не является важным для правильного развития эмбриона. HASH2, представляющий собой человеческий ортолог гена MASH2, был клонирован в 1997 году Alders et al. (обозначенный авторами как "ASCL2"). Alders et al. (1997), Hum. Molec. Genet. 6:859867. Как описано в WO 01/62778, исследования экспрессии выявили, что транскрипт HASH2 (называемый здесь как CASB7439) сверхэкспрессируется при колоректальных опухолях по сравнению с соседними нормальными участками толстого кишечника и другими исследованными нормальными тканями. Этот ген сверхэкспрессируется у пациентов, имеющих стадии I-IV аденокарциономы. Таким образом,белок можно рассматривать как раковый антиген, полезный при иммунотерапевтическом подходе для уменьшения интенсивности рака у субъекта, например путем использования рекомбинантногоCASB7439 в качестве антигенспецифического противоракового иммунотерапевтического агента (ASCI). Краткое изложение сущности изобретения В настоящем изобретении предложены соединения и способы увеличения рекомбинантной продукции полипептидов CASB7439. В некоторых воплощениях предложены модифицированные полипептидыCASB7439, а также белковые конструкции, содержащие такие модифицированные полипептидыCASB7439. Указанные модифицированные полипептиды CASB7439 включают по меньшей мере одну модификацию для усиленной продукции CASB7439. Авторы настоящего изобретения также раскрыли молекулы нуклеиновой кислоты, содержащие полинуклеотидную последовательность, кодирующую модифицированные полипептиды CASB7439 и белковые конструкции, содержащие такие описанные здесь модифицированные полипептиды CASB7439. В некоторых аспектах авторы настоящего изобретения раскрыли белковые конструкции, имеющие аминокислотные (аа) последовательности, и кодирующую их нуклеотидную последовательность (ДНК), представляющие собой следующие:LVL168 [SEQ ID NO: 11 (aa); SEQ ID NO: 12 (ДНК)]. Также раскрыты способы и процессы, в которых используются описанные здесь молекулы нуклеиновой кислоты, для продукции белковых конструкций. Авторы настоящего изобретения также раскрыли иммуногенные композиции, содержащие один или более чем один описанный здесь модифицированный полипептид CASB7439 или конструкции, и фармацевтически приемлемый носитель или эксципиент, где носитель или эксципиент могут возможно содержать буфер. В некоторых воплощениях раскрыто применение описанных здесь модифицированных полипептидов CASB7439 или белковых конструкций или кодирующей их молекулы нуклеиновой кислоты в изготовлении лекарственного средства для лечения колоректального рака. В некоторых воплощениях раскрыты белковые конструкции для применения в терапии, в частности терапии колоректального рака. В некоторых воплощениях раскрыты способы индукции иммунного ответа против CASB7439 у субъекта,страдающего от колоректального рака, включающие (а) выбор субъекта, страдающего от колоректального рака; и (б) введение этому субъекту эффективного количества иммуногенной композиции, содержащей описанные здесь модифицированные полипептиды CASB7439 или конструкцию и фармацевтически приемлемый носитель или эксципиент, где носитель или эксципиент могут возможно содержать буфер. Краткое описание графических материалов Фиг. 1. Схематическая диаграмма, изображающая различные области полипептида CASB7439, а именно ДНК-специфический (ДНК-связывающий) домен, домен bHLH и богатую пролином область. Подробную информацию см. в примере 2. Также изображены схематические представления следующих областей: нацеливающая на ядро (серая стрелка); альфа-спиральная (серые цилиндры); биспиральная(черные цилиндры); характерное расстройство (черные стрелки) и низкая интеграция (серая полоска). Фиг. 2. Схематическая диаграмма, изображающая различные усеченные варианты полипептидаCASB7439 (черные стрелки), совмещенные с полноразмерным полипептидом CASB7439. Подробную информацию см. в примере 4. Фиг. 3. Схематическая диаграмма CASB7439, демонстрирующая приблизительное расположение богатой пролином области, и идентифицированных и предсказанных эпитопов. Различные воплощения содержат полипептид CASB7439, модифицированный путем удаления некоторых или всех аминокислот в богатой пролином области. Такие модифицированные полипептиды CASB7439 сохраняют большую долю эпитопов в полипептиде CASB7439. Фиг. 4. Представлено совмещение между последовательностью немодифицированного полипептидаCASB7439 (SEQ ID NO: 13) и полипептидной последовательностью, возникающей в результате двух возможных модификаций полипептида CASB7439: первая, при которой удаляют 21 последовательный аминокислотный остаток (133-153 включительно); вторая, при которой удаляют 21 последовательный аминокислотный остаток (131-151 включительно). Кроме того, может быть описана третья модификация полипептида CASB7439, при которой удаляют 21 последовательный аминокислотный остаток (132-152 включительно). Как изображено на фигуре, получающаяся в результате модифицированная полипептидная последовательность CASB7439 (вставка, в круге) в области делеции является идентичной для всех трех модификаций. Последнее возникает в результате повторяющихся RG аминокислотных остатков в позициях 131-132 и 152-153 в SEQ ID NO: 13. Фиг. 5. Представлено совмещение аминокислотной последовательности между белковыми конструкциями в примере 6:LVL168 (SEQ ID NO: 11). Фиг. 6. Представлено совмещение между аминокислотной последовательностью HASH2LVL144 (SEQ ID NO: 9). Фиг. 7. Анализ при помощи qPCR (количественной ПЦР) в реальном времени экспрессии мРНКCASB7439 в нормальной ткани (12) и ткани колоректального рака (83), см. пример 9. Фиг. 8. Соответствие между мРНК CASB7439 и экспрессией белка в различных человеческих колоректальных образцах (CRC), черные круги демонстрируют экспрессию мРНК CASB7439 в образцах рака толстой кишки, тогда как пустые окружности (о) демонстрируют экспрессию мРНК CASB7439 в нормальных соседних тканях. Соответствующий уровень белка CASB7439, обнаруженный при помощи иммунофлуоресценции, указан над каждым образцом (- отсутствие экспрессии, +/- очень низкая экспрессия, + низкая экспрессия,сильная экспрессия иочень сильная экспрессия). CRC: первичные опухоли, METS: метастаз, NAT: нормальные соседние ткани. Все образцы представляют образцы аденокарциономы толстой кишки, за исключением образцов 22440 и 24762, которые представляют собой слизеобразующие карциномы, см. пример 9. Фиг. 9. В данной таблице представлен банк пептидов, охватывающий последовательность полноразмерного белка CASB7439, используемый для повторной стимуляции спленоцитов после иммунизации различными конструкциями CASB7439, см. пример 10. Фиг. 10. Изображена пептидная матрица CASB7439, используемая для исследований Т-клеточной иммуногенности, см. пример 10. Фиг. 11A-11D. CD4 ответ, выраженный в виде процентной доли дважды положительных(IFN/TNF) CD4 Т-клеток в четырех линиях инбредных мышей (C57BL6, BALBC, CB6F1 и С 3 Н), иммунизированных LVL111 + AS01B с последующей рестимуляцией перекрывающимися пептидамиCASB7439 (совокупности, матричный подход), см. пример 10, Эксперимент по мультилинейному сравнению инбредных мышей. Фиг. 12A, 12B. CD4 ответ, выраженный в виде процентной доли дважды положительных(IFN/TNF) CD4 Т-клеток в четырех линиях инбредных мышей, иммунизированных LVL111 + AS15 с последующей повторной стимуляцией CASB7439 перекрывающимися пептидами (совокупности, матричный подход), см. пример 10, Эксперимент по мультилинейному сравнению инбредных мышей. Фиг. 13. Идентификация CD4 иммуногенных пептидов CASB7439 в четырех инбредных линиях мышей, обсуждавшихся в предшествующем абзаце. Светло-серый = 0,2-0,4% дважды положительных(IFN/TNF) CD4 Т-клеток; темно-серый 0,5% дважды положительных (IFN/TNF) CD4 Т-клеток, см. пример 10, Эксперимент по мультилинейному сравнению инбредных мышей. Фиг. 14. Идентификация CD4 иммуногенных CASB7439 пептидов у аутбредных CD1 мышей(AS01B, фиг.14A; AS15, фиг. 14B). Светло-серый = 0,2-0,4% дважды положительных (IFN/TNF) CD4 Т-клеток; темно-серый 0,5% дважды положительных (IFN/TNF) CD4 T-клеток, см. пример 10, Иммуногенность CASB7439 у мышей: CASB7439 Т-клеточная иммуногенность у аутбредных мышей. Фиг. 15A, 15B. CD4 и CD8 Т-клеточные ответы, выраженные в виде процентной доли дважды положительных (IFN/TNF) у отдельных CD1 аутбредных мышей, иммунизированных LVL111, LVL168 или LVL144, приготовленных с AS15, с последующей повторной стимуляцией CASB7439 перекрывающимися пептидами (совокупности иммунодоминантных пептидов), см. пример 10, Исследования Фиг. 16. CD4 Т-клеточные ответы (выраженные в виде процентной доли дважды положительных(IFN/TNF в совокупности лейкоцитов периферической крови (PBL), выделенных у трансгенных мышей HLA A2.1/DR-1, иммунизированных LVL168 (SEQ ID NO: 11) или LVL144 (SEQ ID NO: 9), приготовленных с AS15, с последующей повторной стимуляцией CASB7439 перекрывающимися пептидами (7 совокупностей из 7 пептидов/совокупность + пептид 24 (SEQ ID NO: 76. Фиг. 17. CD8 Т-клеточные ответы (выраженные в виде процентной доли дважды положительных(IFN/TNF в объединенных PBL, выделенных у HLA A2.1/DR-1 трансгенных мышей, иммунизированных LVL168 (SEQ ID NO: 11) или LVL144 (SEQ ID NO: 9), приготовленных с AS15, с последующей повторной стимуляцией CASB7439 перекрывающимися пептидами (7 совокупностей из 7 пептидов + пептид 24 (SEQ ID NO: 76. Фиг. 18. CD4 Т-клеточные ответы (выраженные в виде процентной доли дважды положительныхLVL168 (SEQ ID NO: 11) или LVL144 (SEQ ID NO: 9), приготовленных с AS15, с последующей повторной стимуляцией CASB7439 перекрывающимися пептидами (7 совокупностей из 7 пептидов + пептид 24(SEQ ID NO: 76. Фиг. 19. CD8 Т-клеточные ответы (выраженные в виде процентной доли дважды положительныхLVL168 (SEQ ID NO: 11) или LVL144 (SEQ ID NO: 9), приготовленных с AS15, с последующей повторной стимуляцией CASB7439 перекрывающимися пептидами (7 совокупностей из 7 пептидов + пептид 24(SEQ ID NO: 76. Фиг. 20A-20D. Анализ в зависимости от времени ответа в виде CASB7439-специфических антител(IgG1 и IgG2a), инициируемого после иммунизации LVL111 + AS01 В или LVL111 +AS15 у CB6f1 инбредных мышей, см. пример 10, Иммуногенность CASB7439 у мышей: CASB7439-опосредованный гуморальный ответ. Фиг. 21A, 21B. CASB7439-специфический гуморальный ответ у инбредных CB6f1 мышей (IgG1 иIgG2a) после иммунизации LVL111, LVL168 или LVL144, приготовленных с адъювантом AS15 или только одним адъювантом AS15, см. пример 10, Иммуногенность CASB7439 у мышей: CASB7439 опосредованный гуморальный ответ. Фиг. 22. CASB7439-специфический гуморальный иммунный ответ (титр IgG) у CD1 аутбредных мышей, иммунизированных LVL111, LVL168 или LVL144, приготовленных с адъювантом AS15 или только одним адъювантом AS15; нативных мышей использовали в качестве контроля, см. пример 10,Исследования на мышах CD1. Фиг. 23. TC1/CASB7439 14 опухолевый трансплантат у мышей CB6f1, иммунизированныхLVL111, приготовленным с AS01B (фиг. 23A) или AS15 (фиг. 23B), см. пример 11, Исследование 1.1. На фиг. 23A порядок кривых на графике сверху вниз представляет собой "буфер", LVL111 (10 мкг),LVL111 (10 мкг) + AS01B и LVL111 (1 мкг) + AS01B. На фиг. 23B кривая внизу представляет собойLVL111 (10 мкг) + AS15. Фиг. 24. Кривые выживания для примера 11, исследование 1.2. TF: без опухоли. Фиг. 24A. Кривая выживания CB6f1 мышей, иммунизированных физиологическим растворомLVL111 без адъюванта или только одним адъювантом (AS01 или AS15) и сенсибилизированных клетками TC1/CASB7439 14. Последняя выжившая мышь из группы, иммунизированной LVL111, умерла первой, а затем последняя выжившая мышь из группы AS01B, а затем последняя выжившая мышь из группыAS15 и последняя выжившая мышь из группы, которой вводили буфер. Фиг. 24B. Кривая выживания CB6f1 мышей, иммунизированных только AS15 или указанными дозами LVL111, приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14. Последняя выжившая мышь из группы AS15 погибла первой. Из других групп: в группе 30 мкг было меньше всего выживших, а затем группа 1 мкг, затем группа 10 мкг, в которой было наибольшее количество выживших. Фиг. 24C. Кривая выживания CB6f1 мышей, иммунизированных только AS01B или указанными дозами LVL111, приготовленными с AS01 В, и сенсибилизированных клетками TC1/CASB7439 14. В этой панели последние выжившие мыши из группы 10 мкг и AS01B погибли в один и тот же день. Группы 1 и 30 мкг имели одинаковую процентную долю выживших к заключительным суткам. Фиг. 25. Кривые выживания для примера 11, представлено исследование 1.3. TF: без опухоли. Фиг. 25A. Кривая выживания для CB6f1 мышей, иммунизированных LVL111, LVL144 или LVL168,приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB743914. Фиг. 25B. Кривая выживания для CB6f1 мышей, иммунизированных LVL111, LVL144 или LVL168,приготовленных с AS15 или только AS15, и сенсибилизированных клетками MC38/CASB7439 35. Фиг. 26. Кривые выживания для примера 11, представлено исследование 1.4. TF: без опухоли. Фиг. 26A. Кривая выживания CB6f1 мышей, иммунизированных LVL144 или LVL168, приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14. В группе, иммунизированной LVL144, наблюдали несколько больший процент выживания. Фиг. 26B. Кривая выживания для CB6f1 мышей, иммунизированных LVL144 или LVL168, приготовленных с AS15 или только AS15, и сенсибилизированных клональными клетками TC1/CASB7439 14-2. Мыши, инокулированные LVL144, демонстрировали несколько больший процент выживания. Фиг. 26C. Кривая выживания для мышей CB6f1, иммунизированных LVL144 или LVL168, приготовленных с AS15 или только AS15, и сенсибилизированных клетками MC38/CASB7439 35. Среди мышей, инокулированных LVL144, не было выживших. Фиг. 27. Кривые выживания для примера 11, представлено исследование 2.1. TF: без опухоли. Фиг. 27A. Кривая выживания мышей CB6f1, иммунизированных 6,25 мкг LVL168 и 10 мкг LVL144,приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, инокулированная LVL168, имела несколько более высокую процентную долю выживших. Фиг. 27B. Кривая выживания мышей CB6f1, иммунизированных 3,1 мкг LVL168 и 5 мкг LVL144,приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, инокулированных LVL144, имела несколько более высокую процентную долю выживших. Фиг. 27C. Кривая выживания мышей CB6f1, иммунизированных 1,55 мкг LVL168 и 2,5 мкгLVL144, приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, инокулированных LVL144, имела самую высокую процентную долю выживших. Фиг. 27D. Кривая выживания мышей CB6f1, иммунизированных 0,77 мкг LVL168 и 1,25 мкгLVL144, приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, инокулированных LVL168, имела несколько более высокую процентную долю выживших. Фиг. 28. Кривые выживания для примера 11, представлено исследование 2.2. TF: без опухоли. Фиг. 28A. Кривая выживания мышей C57Bl/6, иммунизированных 6,25 мкг LVL168 или 10 мкгLVL144, приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Фиг. 28B. Кривая выживания мышей C57Bl/6, иммунизированных 3,1 мкг LVL168 или 5 мкгLVL144, приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Фиг. 28C. Кривая выживания мышей C57Bl/6, иммунизированных 1,55 мкг LVL168 или 2,5 мкгLVL144, приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Фиг. 28D. Кривая выживания мышей C57Bl/6, иммунизированных 0,77 мкг LVL168 или 1,25 мкгLVL144, приготовленных с AS15 или только AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Фиг. 29. Кривые выживания для примера 11, представлено исследование 2.3. TF: без опухоли. Фиг. 29A. Кривая выживания мышей CB6f1, иммунизированных 6,25 мкг LVL168 или 10 мкгLVL144, приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, получавшая LVL168, имела самую высокую процентную долю выживших, а затем группа, получавшая LVL144. Выжившие отсутствовали в группе, получавшей AS15. Фиг. 29B. Кривая выживания мышей CB6f1, иммунизированных 3,1 мкг LVL168 и 5 мкг LVL144,приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, получавшая LVL168, имела самую высокую процентную долю выживших, а затем группа, получавшаяLVL144. Выжившие отсутствовали в группе, получавшей AS15. Фиг. 29C. Кривая выживания мышей CB6f1, иммунизированных 1,55 мкг LVL168 и 2,5 мкгLVL144, приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, получавшая LVL168, имела самую высокую процентную долю выживших, а затем группа, получавшая LVL144. Выжившие отсутствовали в группе, получавшей AS15. В качестве контролей использовали физиологический раствор или AS15. Фиг. 30. Кривые выживания для примера 11, исследование 2.3. TF: без опухоли. Фиг. 30A. Кривая выживания мышей CB6f1, иммунизированных 0,77 мкг LVL168 или 1,25 мкгLVL144, приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, получавшая LVL168, имела самую высокую процентную долю выживших, а затем группа, получавшая LVL144. Выжившие отсутствовали в группе, получавшей AS15. Фиг. 30B. Кривая выживания мышей CB6f1, иммунизированных 0,19 мкг LVL168 или 0,31 мкгLVL144, приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, получавшая LVL168, имела самую высокую процентную долю выживших. Последняя выжившая мышь из группы, получавшей LVL144, пережила последнюю выжившую из группы AS15 и группы, получавшей буфер. Фиг. 30C. Кривая выживания мышей CB6f1, иммунизированных 0,048 мкг LVL168 или 0,078 мкгLVL144, приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. Группа мышей, получавшая LVL168, имела самую высокую процентную долю выживших. Последняя выжившая мышь из группы, получавшей LVL144, пережила последнюю выжившую из группы AS15 и группы, получавшей буфер. Физиологический раствор или AS15 использовали в качестве контролей. Фиг. 31. TC1/CASB7439 14-2 опухолевый трансплантат у самцов и самок мышей CB6f1, иммунизированных 1 мкг LVL168, приготовленных с AS15; только AS15 использовали в качестве контроля (фиг. 31A). TC1/CASB7439 14-2 опухолевый трансплантат у мышей C57Bl/6, иммунизированных 1 мкгLVL168, приготовленных с AS15; AS15 использовали в качестве контроля (фиг. 31B). Фиг. 32A. Кривая выживания самцов или самок мышей CB6f1, иммунизированных 1 мкг LVL168,приготовленных с AS15, и сенсибилизированных клетками TC1/CASB7439 14-2. AS15 использовали в качестве контроля у самцов и самок мышей CB6f1. Среди мышей CB6f1, иммунизированных LVL168 вместе с AS15, только 7 самцов и 3 самки не имели опухоли (TF). TF отсутствовали среди мышей, получавших только AS15. Фиг. 32B. Кривая выживания мышей C57Bl/6, иммунизированных 1 мкг LVL168, приготовленных сAS15, и сенсибилизированных клетками TC1/CASB7439 14-2. AS15 использовали в качестве контроля у самцов и самок мышей C57Bl/6. Среди мышей C57Bl/6, иммунизированных LVL168 вместе с AS15, присутствовали 2 TF самца и 1 TF самка. Среди мышей, получавших только AS15, присутствовали 5 TF самцов и 1 TF самка. Фиг. 33. CASB7439-специфический гуморальный иммунный ответ (титр IgG) у самцов (черные кружки) и самок (серые кружки) мышей CB6F1, иммунизированных 1 мкг LVL168, приготовленных сAS15 или только AS15 (слева). CASB7439-специфический гуморальный иммунный ответ (титр IgG) у самцов (черные кружки) и самок (серые кружки) мышей C57Bl/6, иммунизированных 1 мкг LVL168,приготовленных с AS15, или только AS15 (справа). Фиг. 34. CD4 Т-клеточные ответы (выраженные в виде процентной доли дважды положительных(SEQ ID NO: 11), приготовленных с AS15 или только AS15, с последующей повторной стимуляцией банком из 46 пептидов (см. фиг. 9), покрывающим последовательность полноразмерного белка CASB7439(фиг. 34A). CD4 Т-клеточные ответы (выраженные в виде процентной доли дважды положительных(SEQ ID NO: 11), приготовленных с AS15 или только AS15, с последующей повторной стимуляцией совокупными пептидами CASB7439 из банка 46 пептидов (см. фиг. 9), покрывающего последовательность полноразмерного белка CASB7439 (фиг. 34B, слева); CASB7439 пептид 39 (SEQ ID NO: 91) (фиг. 34B,середина); или RPMI (фиг. 34B, справа). Фиг. 35. CD8 Т-клеточные ответы (выраженные в виде процентной доли дважды положительных(SEQ ID NO: 11), приготовленных с AS15 или только AS15, с последующей повторной стимуляцией банком из 46 пептидов (см. фиг. 9), покрывающим последовательность полноразмерного белка CASB7439(фиг. 35A). CD8 Т-клеточные ответы (выраженные в виде процентной доли дважды положительных(SEQ ID NO: 11), приготовленных с AS15 или только AS15, с последующей повторной стимуляцией банком 46 пептидов (см. фиг. 9), покрывающим последовательность полноразмерного белка CASB7439(фиг. 35B, слева); CASB7439 пептид 39 (SEQ ID NO: 91) (фиг. 35B, середина) или RPMI (фиг. 35B, справа). Подробное описание изобретения Единственным препятствием для коммерчески приемлемых иммунотерапевтических агентов является получение коммерчески приемлемых количеств рекомбинантного ракового антигена. В общем и без ограничения, коммерчески приемлемый уровень продукции рекомбинантного полипептида составляет примерно 5% от общего белка, продуцируемого клеткой-хозяином, что оценивают при помощи окрашивания геля SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) красителем Кумасси. Существует множество причин того, почему нативная полипептидная последовательность может обеспечивать выход только небольших количеств при получении рекомбинантными способами. Подходы для увеличения продукции полипептида включают изменение системы экспрессии хозяина или оптимизацию кодона. Тем не менее, эти стандартные подходы не всегда приводят в результате к приемлемой продукции полипептида, и продукция некоторых полипептидов остается невозможной в коммерческих целях. Как более подробно описано ниже, заявители изучали, будет ли модифицированный полипептидCASB7439 продуцировать приемлемые белковые уровни при использовании рекомбинантных средств. В некоторых воплощениях предложены модифицированные полипептиды CASB7439, где модификация состоит из или включает C-концевое усечение полипептида CASB7439, так что удаляют весь или фрагмент богатой пролином области. В некоторых воплощениях модификация полипептида CASB7439 состоит из или включает делецию всей или фрагмента богатой пролином области. В некоторых воплощениях предложены модифицированные полипептиды CASB7439, которые сохраняют, по меньшей мере, аминокислотную последовательность 193 из SEQ ID NO: 13, но где вся богатая пролином область или ее фрагмент удален. В других воплощениях предложены белковые конструкции, содержащие модифицированный полипептид CASB7439, описанный ниже. В некоторых воплощениях такие белковые конструкции дополнительно содержат один или более гетерологичный полипептид. В некоторых воплощениях гетерологичный полипептид представляет собой партнер слияния, как описано здесь более подробно. В особенно подходящих воплощениях такие белковые конструкции содержат гетерологичный полипептид, располагающийся с N-конца относительно модифицированного полипептида CASB7439. В других особенно подходящих воплощениях предложены белковые конструкции, где гетерологичный полипептид представляет собой фрагмент белка D Н.influenzae (как более подробно описано ниже), располагающийся сN-конца модифицированного полипептида CASB7439. В некоторых воплощениях предложены белковые конструкции, где гетерологичный полипептид содержит полигистидиновую область. В дополнительных воплощениях полигистидиновая последовательность располагается в C-концевой области белковой конструкции. В других воплощениях полигистидиновая последовательность располагается на C-конце белковой конструкции. В дополнительных воплощениях полигистидиновая последовательность располагается в N-концевом фрагменте белковой конструкции. В других воплощениях полигистидиновая последовательность располагается на N-конце белковой конструкции. В подходящих воплощениях полигистидиновая область содержит десять или более последовательных гистидиновых остатков. В особенно подходящих воплощениях полигистидиновая область содержит десять или более последовательных гистидиновых остатков, располагающихся наN-конце модифицированного полипептида CASB7439. В некоторых воплощениях более чем одна полигистидиновая область может быть включена по одной или более из описанных здесь областей. В некоторых воплощениях предложены белковые конструкции, где гетерологичный полипептид представляет собой белок-носитель, химически конъюгированный с модифицированным полипептидомCASB7439. В некоторых воплощениях предложены белковые конструкции, содержащие гетерологичный полипептид, представляющий собой полигистидиновую область, и гетерологичный полипептид, представляющий собой белок-носитель. В некоторых воплощениях предложены белковые конструкции, содержащие гетерологичный полипептид, представляющий собой полигистидиновую область, и гетерологичный полипептид, представляющий собой партнер слияния. В некоторых воплощениях предложены белковые конструкции, содержащие гетерологичный полипептид, представляющий собой полигистидиновую область, гетерологичный полипептид, представляющий собой партнер слияния, и гетерологичный полипептид, представляющий собой белок-носитель. Заявители также раскрыли воплощения каждой из конструкций, описанных здесь, в которых некоторые или все из гетерологичных полипептидов, таких как полигистидиновая область, удалены. Композиции и способы удаления гетерологичных полипептидов известны в области техники, включая, без ограничения, расщепление эндо- или экзопептидазами, химическую модификацию или расщепление связи между гетерологичным полипептидом и оставшейся частью молекулы и т.п. Заявители также раскрыли здесь иммуногенные композиции, содержащие любой модифицированный полипептид CASB7439 или белковую конструкцию, включающую описанный здесь модифицированный полипептид CASB7439, и фармацевтически приемлемый носитель или эксципиент, где носитель или эксципиент могут возможно содержать буфер. В некоторых аспектах эти композиции дополнительно содержат адъювант. В некоторых аспектах эти композиции содержат адъювант, который вызывает, по меньшей мере, Th1 иммунный ответ. В некоторых аспектах описанный здесь адъювант содержит по меньшей мере одно из следующего: 3D-MPL (монофосфориллипид), QS21 и CpG. В некоторых воплощениях композиция содержит 3D-MPL. В других воплощениях композиция содержит CpG. В некоторых воплощениях композиция содержит QS21. В некоторых воплощениях композиция содержит QS21 и холестерин. В некоторых воплощениях композиция содержит холестерин, 3D-MPL и QS21 в липосомальной композиции. Заявители также раскрыли здесь молекулы нуклеиновой кислоты, содержащие полинуклеотидные последовательности, кодирующие модифицированные CASB7439 полипептиды и описанные здесь конструкции. В некоторых воплощениях раскрыт вектор, содержащий такие описанные здесь молекулы нуклеиновой кислоты. В некоторых воплощениях вектор содержит прокариотический экспрессирующийся вектор. В некоторых воплощениях вектор содержит эукариотический экспрессирующийся вектор. В некоторых воплощениях полинуклеотидная последовательность, кодирующая конструкцию, оптимизирована по кодонам для экспрессии в клетке-хозяине. Заявители также раскрыли клетки-хозяева, содержащие (1) описанную здесь молекулу нуклеиновой кислоты или (2) вектор, содержащий описанную здесь молекулу нуклеиновой кислоты. В некоторых воплощениях клетка-хозяин выбрана из группы: бактериальных клеток, клеток насекомых и клеток млекопитающих. В некоторых воплощениях предложена иммуногенная композиция, содержащая (1) описанную здесь молекулу нуклеиновой кислоты или (2) вектор, содержащий описанную здесь молекулу нуклеиновой кислоты, и фармацевтически приемлемый носитель или эксципиент. В некоторых воплощениях предложено применение модифицированных полипептидов CASB7439 или описанных здесь белковых конструкций (или кодирующих их молекул нуклеиновой кислоты) в изготовлении лекарственного средства для лечения колоректального рака. В некоторых воплощениях раскрыты описанные здесь модифицированные полипептиды CASB7439 или белковые конструкции (или кодирующие их молекулы нуклеиновой кислоты) для применения в терапии, в частности для терапии колоректального рака. В некоторых воплощениях предложены способы индукции иммунного ответа против CASB7439 у субъекта, страдающего от колоректального рака, где в способе осуществляют (1) выбор субъекта, страдающего от колоректального рака; и (2) введение субъекту эффективного количества иммуногенной композиции, содержащей описанный здесь модифицированный полипептид CASB7439 или белковую конструкцию, и фармацевтически приемлемый носитель или эксципиент, где носитель или эксципиент могут возможно содержать буфер. В некоторых воплощениях раскрытых здесь способов иммуногенная композиция дополнительно содержит адъювант. В некоторых воплощениях описанного здесь способа адъювант вызывает, по меньшей мере, Th1 иммунный ответ. В некоторых воплощениях раскрытого здесь способа адъювант содержит по меньшей мере один из: 3D-MPL, QS21 и CpG. В некоторых воплощениях описанного здесь способа субъект представляет собой человека. Белковые конструкции Полипептиды CASB7439.CASB7439, как он здесь назван, также известен как HASH2 или ASCL2. HASH2 представляет собой полипептид человеческого происхождения из 193 аминокислотных остатков (SEQ ID NO: 13). См., например,ААВ 86993. Здесь ссылка на CASB7439 сделана с точки зрения полипептидной последовательности, предложенной в SEQ ID NO: 13. Богатая пролином область. Область высокой пролиновой периодичности обнаружена в SEQ ID NO: 13 в области аминокислот 127-158. Заявители назвали область SEQ ID NO: 13 с аминокислоты 133 по аминокислоту 153 включительно как "богатая пролином", "полипролиновая", "полипролин-подобная" или "полипро" область или домен. Соответственно, используемые здесь термины "богатая пролином", "полипролиновая", "полипролин-подобная" или "полипро" относятся к этой области (или домену) белка CASB7439. Модификации. В этом описании предложены белковые конструкции, содержащие полипептид CASB7439, содержащий по меньшей мере одну усиливающую экспрессию модификацию. В некоторых воплощениях фрагмент полипептида CASB7439 удаляют, где указанный фрагмент содержит полноразмерную или фрагмент богатой пролином области. В некоторых воплощениях полноразмерный C-концевой фрагмент полипептида CASB7439 удаляют, что приводит в результате к усеченному полипептиду CASB7439, содержащему аминокислотные остатки 1-117 из SEQ ID NO: 13. В некоторых воплощениях гетерологичный полипептид расположен на N-конце полипептида CASB7439. Используемый здесь термин "усиление экспрессии" или "усиленная экспрессия" в контексте модифицированного полипептида CASB7439 относится к модификации, предназначенной для увеличения уровней экспрессии белковой конструкции, содержащей указанный модифицированный полипептидCASB7439 по сравнению с экспрессией иным образом эквивалентной белковой конструкции, содержащей немодифицированный полипептид CASB7439 SEQ ID NO: 13, т.е. тот же самый вектор, клеткахозяин и т.д Любые из способов, известных в области техники для определения количества белка, приемлемы для определения того, обладает ли заданная конструкция, содержащая модифицированный полипептид CASB7439, увеличенным уровнем экспрессии по сравнению с иным образом эквивалентными белковыми конструкциями, содержащими немодифицированный полипептид CASB7439. В некоторых воплощениях продукция может быть оценена с использованием SDS-PAGE и окрашивания при помощи Кумасси голубого. Удаление богатой пролином области. В некоторых воплощениях модификация полипептида CASB7439 включает делецию части богатой пролином области. В некоторых воплощениях модификация полипептида CASB7439 включает делецию всей богатой пролином области. В некоторых воплощениях модификация полипептида CASB7439 включает делецию аминокислот за пределами богатой пролином области дополнительно к делеции богатой пролином области. В некоторых воплощениях модификацию осуществляют путем усечения C-концевого фрагментаSEQ ID NO: 13. Таким образом, в некоторых воплощениях получают белковую конструкцию, в которойC-концевой аминокислотный остаток модифицированного полипептида CASB7439 соответствует остатку, расположенному по N-концу богатой пролином области SEQ ID NO: 13. В некоторых воплощениях получают белковую конструкцию, в которой C-концевой аминокислотный остаток модифицированного полипептида CASB7439 соответствует остатку, расположенному в богатой пролином областиSEQ ID NO: 13. В некоторых воплощениях C-концевой аминокислотный остаток модифицированного полипептида CASB7439 соответствует аминокислотному остатку 117, 118, 119, 120, 121, 122, 123, 124,125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146,147, 148, 149, 150, 151 или 152 в SEQ ID NO: 13. В некоторых воплощениях модификация включает делецию части или всей богатой пролином области. В некоторых воплощениях модифицированная полипептидная последовательность CASB7439 соответствует SEQ ID NO: 13, из которого удален 21 или более непрерывный аминокислотный остаток. Этот 21 или более последовательный остаток соответствуют непрерывной аминокислотной последовательности SEQ ID NO: 13, начиная по любому из остатков 117, 118, 119, 120, 121, 122, 123, 124, 125, 126,127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148,149, 150, 151 или 152 включительно. В дополнительных воплощениях модифицированная полипептидная последовательность CASB7439 соответствует SEQ ID NO: 13, из которой удален 21 последовательный аминокислотный остаток, где указанный 21 последовательный остаток начинается по любому из остатков 131,132 и 133. В некоторых воплощениях модифицированная полипептидная последовательность CASB7439 соответствует SEQ ID NO: 13, из которой удален один или более остаток 133, 134, 135, 136, 137, 138, 139,140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153. В некоторых воплощениях модифицированная полипептидная последовательность CASB7439 соответствует SEQ ID NO: 13, из которой удалены два или более последовательных аминокислотных остатка в области аминокислотных остатков 133153 включительно. В некоторых воплощениях модифицированная полипептидная последовательность CASB7439 соответствует SEQ ID NO: 13, из которой удалены остатки с 133 по 153 последовательности SEQ ID NO: 13 включительно. Примеры не ограничивающих объем изобретения воплощений включают следующие:LVL168, SEQ ID NO: 11 (aa). Гетерологичные последовательности. В некоторых воплощениях модифицированный полипептид CASB7439 комбинирован с гетерологичным полипептидом. Термин "гетерологичный" в отношении молекулы нуклеиновой кислоты, полипептида или другого клеточного компонента указывает на то, что имеется компонент, в норме не обнаруживаемый в природе, и/или что он происходит из отличающегося от референсной молекулы источника или видов. Использованные здесь "гетерологичные" молекулы включают, без ограничения, слитые белки, белки-носители и последовательности для очистки. В некоторых воплощениях гетерологичный полипептид может быть химически конъюгирован с модифицированным полипептидом CASB7439, т.е. белком-носителем. В других воплощениях гетерологичный полипептид и модифицированный полипептид CASB7439 могут экспрессироваться в виде единичного рекомбинантного слитого белка. В других воплощениях модифицированный полипептидCASB7439 и гетерологичный полипептид экспрессируются в виде единичного рекомбинантного слитого белка и химически конъюгированного с другим гетерологичным полипептидом. Гетерологичный полипептид может способствовать получению Т-хелперных эпитопов, включая Т-хелперные эпитопы, распознаваемые у людей. В случае белковой конструкции, содержащей полипептид CASB7439 и гетерологичный полипептид, который представляет собой слитый белок, гетерологичный полипептид (партнер слияния) может способствовать получению таких эпитопов или способствовать экспрессии белка (усилитель экспрессии) с более высокими выходами по сравнению с нативным рекомбинантным белком. Партнер слияния может представлять собой иммунологический партнер слияния и партнер, усиливающий экспрессию. Белки-носители. Белок-носитель химически конъюгирован с другим целевым полипептидом. Химическое связывание полипептида с белком-носителем может быть осуществлено при помощи любого из способов, известных в области техники. Таким образом, для прямого ковалентного связывания можно использовать карбодиимид, глутаровый альдегид или N-[-малеимидобутирилокси]сукцинимидный эфир с использованием обычных имеющихся в продаже гетеробифункциональных линкеров, таких как CDAP и SPDP (с использованием указаний производителей). После реакции связывания конъюгат полипептид-белокноситель легко может быть выделен и очищен при помощи способов диализа, способа гель-фильтрации,способа фракционирования и т.д. Типы белков-носителей, используемых в некоторых воплощениях, известны специалистам в данной области техники. Функция некоторых белков-носителей заключается в том, чтобы обеспечить помощь цитокинам для того, чтобы способствовать индукции иммунного ответа против связанного полипептида. Например, неполный перечень белков-носителей, которые могут быть использованы в настоящем изобретении, включает: гемоцианин лимфы улитки (KLH), сывороточные альбумины, такие как бычий сывороточный альбумин (BSA), инактивированные бактериальные токсины, такие как столбнячный или дифтерийный токсины (ТТ и DT), или их рекомбинантные фрагменты (например, домен 1 фрагмента С из ТТ или домен транслокации DT), или очищенный белок, происходящий из туберкулина (PPD). Партнеры слияния. Несмотря на то что имеется множество генетических слитых систем, экспрессия рекомбинантного белка в Escherichia coli остается затруднительной. Разработка партнера слияния, который в наилучшей степени усиливает экспрессию сложных для экспрессии белков, остается эмпирической задачей. Обычные партнеры слияния включают C-концы белка, связывающего мальтозу (MBP), глутатионS-трансферазу (GST), тиоредоксин (TRX), NUS A, убиквитин (Ub) и небольшие последовательности убиквитин-подобного модификатора (SUMO), каждый из которых описан в литературе. См., например,Marblestone (2006), Protein Sci. 15:182-7; Hunt (2005), Protein Exp.Purif, 40:1-22; Hammarstom et al.(2002), Protein Sci. 11:313-321. Иммуногенные партнеры слияния включают белок D Haemophilus influenza B и неструктурный белок вируса гриппа, NS1 (гемагглютинин). Другой иммунологический партнер слияния представляет собой белок, известный как LYTA. Может быть использован C-концевой фрагмент молекулы. LYTA происходит из Streptococcus pneumonia, синтезирующего N-ацетил-L-аланинамидазу, амидазу LYTA (кодируемую геном lytA Garcia et al. (1986), Gene, 43:265-272). Можно использовать повторяющийся фрагмент молекулы Lyta, обнаруженный на C-конце, начиная с остатка 178, например остатки 188-305. Другой подходящий партнер слияния представляет собой белок аденилатциклазу (CyaA) или его фрагмент, где указанный фрагмент СуаА сохраняет свойство указанного белка аденилатциклазы нацеливания на антигенпрезентирующие клетки, см. WO 2005/089792. В одном из воплощений изобретения гетерологичный полипептид представляет собой белок DIgD-связывающий белок Haemophilus influenzae (WO 91/18926, выданный как ЕР 0594610 B1). В некоторых случаях, например, в системах экспрессии рекомбинантного иммуногена может быть желательно использовать N-концевые фрагменты белка D, например, содержащие от приблизительно 100 до приблизительно 110 N-концевых аминокислот белка D (GB 9717953.5) (pD1/3, SEQ ID NO: 39). Белок D H.influenzae синтезируется в виде предшественника с 18 аминокислотной сигнальной последовательностью (SEQ ID NO: 41, см. также номер ААА 24998). Когда сигнальная последовательность процессируется во время секреции, тогда цистеиновый остаток в положении 19 в молекулепредшественнике становится N-концевым остатком. В одном из воплощений гетерологичная полипептидная последовательность содержит приблизительно первую треть процессированной молекулы белка D H.influenzae или N-концевые 100-110 аминокислот процессированного белка D. В одном из воплощений гетерологичный белок содержит аминокислотные остатки Met-Asp-Pro, связанные с N-концевыми 109 аминокислотами процессированного белкаD. В еще одном воплощении гетерологичный белок содержит аминокислотные остатки Met-Asp-Pro, связанные с аминокислотными остатками 2-109 процессированного белка D (SEQ ID NO: 39). В одном из воплощений модифицированный полипептид CASB7439 или белковая конструкция химически связана с белком СуаА или его фрагментом, см. WO 2005/054851. В еще одном воплощении модифицированный полипептид CASB7439 или белковая конструкция химически связана с B субъединицей токсина Шига или его иммунологически функциональным эквивалентом, см. патент США 6613882; WO 02/060937; WO 2005/112991. Полигистидиновые последовательности, неродственные аминокислоты. Часто благоприятным является включение гетерологичного полипептида, содержащего секреторную или лидерную последовательности, пропоследовательности, последовательности, способствующие очистке, такие как гистидиновая метка, например, полигистидиновая последовательность (состоящая из множественных гистидиновых остатков), или дополнительную последовательность для стабильности во время рекомбинантной продукции. Векторы и наборы с гистидиновыми метками имеются в продаже. Например, векторы для добавления к полипептиду метки из шести гистидиновых молекул доступны от Roche. Наборы для приготовления и применения полипептидов с гистидиновыми метками доступны от Qiagen, Sigma, Thermo Scientific,GE Healthcare и т.п. За гистидиновой последовательностью может также следовать подходящая аминокислотная последовательность, облегчающая удаление полигистидиновой последовательности с использованием эндопептидаз. Эндопептидаза, например Qiagen TAGZyme, может быть использована для удаления концевой полигистидиновой последовательности без дополнительной последовательности. В некоторых воплощениях полигистидиновая последовательность располагается в N-концевой области белковой конструкции. В дополнительных воплощениях полигистидиновая последовательность располагается на N-конце белковой конструкции. В некоторых воплощениях полигистидиновая последовательность содержит 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или больше последовательных гистидинов. В дополнительных воплощениях полигистидиновая последовательность содержит десять последовательных гистидинов. В некоторых воплощениях гетерологичный полипептид включает полигистидиновую последовательность и одну или более неродственную аминокислоту. Под "неродственными аминокислотами" подразумевают одну или более аминокислоту, которая не является частью полигистидиновой последовательности или слитого белка и в природе не является частью полипептидной последовательностиCASB7439. Например, неродственные аминокислоты могут представлять собой аминокислоты, включенные в предложенный пептидазный сайт, они могут просто представлять собой внешнюю последовательность, такую как артефакт клонирования и т.п. Молекулы нуклеиновой кислоты, кодирующие конструкции CASB7439 Другие воплощения этого описания относятся к рекомбинантным нуклеиновым кислотам, кодирующим описанные здесь модифицированные полипептиды CASB7439 и белковые конструкции. В некоторых воплощениях рекомбинантные нуклеиновые кислоты оптимизированы по кодонам для экспрессии в выбранных прокариотических или эукариотических клетках-хозяевах. Для облегчения репликации и экспрессии нуклеиновые кислоты могут быть включены в вектор, такой как прокариотический или эукариотический экспрессирующийся вектор. Клетки-хозяева, включающие рекомбинантные модифицированные CASB7439 полипептиды или нуклеиновые кислоты, кодирующие белковую конструкцию, также представляют собой характеристику данного описания. Благоприятные клетки-хозяева включают прокариотические (т.е. бактериальные) клетки-хозяева, такие как Е.coli, а также многочисленные эукариотические клетки-хозяева, включая клетки грибов (например, дрожжи), клетки насекомых и клетки млекопитающих (такие как клетки HEK-293 (человеческие эмбриональные почечные клетки), CHO (клетки яичника китайского хомячка) и VERO (клетки почки африканской зеленой мартышки. Экспрессирующиеся векторы. Для облегчения репликации и экспрессии нуклеиновые кислоты могут быть включены в вектор, такой как прокариотический или эукариотический экспрессирующийся вектор. Хотя раскрытые здесь нуклеиновые кислоты могут быть включены в любой из множества векторов (включая, например, бактериальные плазмиды; фаговую ДНК; бакуловирус; дрожжевые плазмиды; векторы, происходящие из комбинаций плазмид и фаговой ДНК, ДНК вирусов, таких как вирус оспы, аденовирус, вирус оспы птиц, вирус псевдобешенства, аденовирус, аденоассоциированный вирус, ретровирусы и множество других), наиболее обычный вектор представляет собой экспрессирующийся вектор, подходящий для получения продуктов полипептидной экспрессии. В экспрессирующемся векторе нуклеиновая кислота, кодирующая белковую конструкцию, типично расположена поблизости и ориентирована к соответствующей последовательности, контролирующей транскрипцию (промотор, и возможно одному или более энхансеру) для управления синтезом мРНК. То есть, интересующая полинуклеотидная последовательность функционально связана с соответствующей последовательностью, контролирующей транскрипцию. Примеры таких промоторов включают предранний промотор CMV, LTR или промотор SV40, полигедроновый промотор бакуловируса, промотор lac или trp E.coli, промотор фага Т 7 и лямбда PL и другие промоторы,известные для контроля экспрессии генов в прокариотических или эукариотических клетках или соответствующих вирусах. Экспрессирующийся вектор типично также содержит сайт связывания с рибосомой для инициации трансляции и терминатор транскрипции. Вектор возможно включает соответствующие последовательности для увеличения экспрессии. Кроме того, экспрессирующиеся векторы возможно содержат один или более селективный маркерный ген для обеспечения фенотипического признака для отбора трансформированных клеток-хозяев, такой как дигидрофолатредуктаза или резистентность к неомицину для культуры эукариотических клеток или такой как резистентность к тетрациклину или ампициллину у Е coli. Примеры способов, достаточных для ознакомления специалиста в данной области техники с продукцией нуклеиновых кислот рекомбинантного CASB7439, можно найти в Sambrook et al., Moleculared., WileySons, 1999. Примеры молекул нуклеиновых кислот, кодирующих белковые конструкции, представленыSEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10 и SEQ ID NO: 12. Дополнительные варианты последовательности, которые обладают идентичной последовательностью с взятой для примера молекулой нуклеиновой кислоты, могут быть получены специалистами в данной области техники. Типично, варианты нуклеиновой кислоты кодируют полипептиды, отличающиеся не более чем на 1, или 2, или 5, или 10, или 15, или 20% по аминокислотным остаткам. То есть, кодируемые полипептиды обладают по меньшей мере 80 или 85%, более типично по меньшей мере приблизительно 90% или большей, такой как 95, 98, 99 или 99,5%, идентичностью последовательности. Специалистам в данной области техники понятно, что полинуклеотидные последовательности, кодирующие белковые конструкции, сами могут обладать меньшей идентичностью последовательности вследствие вырожденности генетического кода. В некоторых случаях модифицированный полипептид CASB7439 или белковая конструкция имеют модификацию одной или более аминокислоты относительно аминокислотной последовательности, изложенной здесь, используемой в качестве примера конструкции (например, дополнительно к вышеупомянутым модификациям). Такие отличия могут представлять собой добавление, делецию или замену одного или более нуклеотида или аминокислоты соответственно. Вариант, как правило, отличается не более чем приблизительно 1, или 2, или 5, или 10, или 15, или 20, или 25, или 30% нуклеотидных остатков. Например, вариант нуклеиновой кислоты, кодирующей полипептид CASB7439 или белковую конструкцию, может включать 1, или 2, или до 5, или до приблизительно 10, или до приблизительно 15,или до приблизительно 50, или до приблизительно 100 нуклеотидных различий по сравнению с используемыми в качестве примера нуклеиновыми кислотами, кодирующими полипептид CASB7439 или белковую конструкцию в соответствии с последовательностями SEQ ID NO: 2, SEQ ID NO: 4,SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10 и SEQ ID NO: 12. Таким образом, вариант в контексте нуклеиновой кислоты, кодирующей полипептид CASB7439 или белковую конструкцию, типично обладает по меньшей мере 70, или 75, или 80, или 85, или 90, или 95, или 98, или 99, или 99,5% идентичностью последовательности с референсной последовательностью, например референсной последовательностью,проиллюстрированной как SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10 иSEQ ID NO: 12, или любые другие из раскрытых здесь примеров нуклеиновых кислот, кодирующих полипептид CASB7439 или белковую конструкцию. Дополнительные варианты могут возникать в результате генетического дрейфа или могут быть получены искусственно с использованием сайтнаправленного или произвольного мутагенеза или путем рекомбинации двух или более предварительно существующих вариантов. Такие дополнительные варианты также подходят в контексте раскрытого здесь полипептида CASB7439 или белковых конструкций. Дополнительно к ранее описанному варианту нуклеиновой кислоты, нуклеиновые кислоты, которые гибридизуются с одним или более из примеров нуклеиновых кислот, представленных SEQ ID NO: 2,SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10 и SEQ ID NO: 12, также могут быть использованы для того, чтобы кодировать раскрытые здесь полипептиды CASB7439 или белковые конструкции. Специалисту в данной области техники понятно, что дополнительно к обсуждающейся здесь процентной доле (%) меры идентичности последовательности другой признак сходства последовательности между двумя нуклеиновыми кислотами заключается в способности гибридизоваться. Чем ближе последовательности двух нуклеиновых кислот, тем более строгими являются условия, в которых они гибридизуются. Строгость условий гибридизации зависит от последовательности и отличается при различных параметрах окружающей среды. Таким образом, условия гибридизации, приводящие в результате к конкретной степени строгости, варьируют в зависимости от природы выбранного способа гибридизации и композиции и длины гибридизуемой последовательности нуклеиновой кислоты. Как правило, температура гибридизации и ионная сила (в особенности концентрация Na+ и/или Mg) буфера для гибридизации определяют строгость условий гибридизации, хотя значения времени промывания также влияют на строгость. Как правило, строгие условия выбирают такими, что они приблизительно на 5-20C ниже температуры плавления (Tm) специфической последовательности при определенной ионной силе и pH. Tm представляет собой температуру (при определенной ионной силе и pH), при которой 50% последовательности-мишени гибридизуется с наиболее подходящим зондом. Условия для гибридизации нуклеиновой кислоты и расчет строгости условий можно найти, например, в Sambrook et al., Molecular Cloning: Ain Molecular Biology, 4th ed., John WileySons, Inc., 1999. Для задач настоящего описания "строгие условия" охватывают условия, при которых гибридизация происходит только в том случае, если имеется меньше 25% несовпадений между гибридизуемой молекулой и последовательностью-мишенью. "Строгие условия" могут быть классифицированы до конкретной степени строгости для более точного определения. Таким образом, используемый здесь термин "умеренно строгие" условия представляют собой такие условия, при которых молекулы с более чем 25% несовпадениями последовательности не будут гибридизоваться; условия "средней строгости" представляют собой условия, в которых молекулы с более чем 15% несовпадений не будут гибридизоваться, и условия"высокой строгости" представляют собой условия, при которых последовательности с более чем 10% несовпадениями не будут гибридизоваться. Условия "очень высокой строгости" представляют собой условия, в которых последовательности с более чем 6% несовпадениями не будут гибридизоваться. Наоборот, нуклеиновые кислоты, которые гибридизуются в "условиях низкой строгости", включают кислоты с гораздо меньшей идентичностью последовательности или с идентичностью последовательности только на очень коротких подпоследовательностях нуклеиновой кислоты. Таким образом, понятно, что различные варианты нуклеиновых кислот, которые охватываются этим изобретением, способны гибридизоваться в строгих условиях по меньшей мере с одной из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQID NO: 8, SEQ ID NO: 10 и SEQ ID NO: 12, по существу, по всей их длине. Продукция белковых конструкций Раскрытые здесь белковые конструкции могут быть получены с использованием признанных способов экспрессии и очистки рекомбинантных белков. Способы, достаточные для ознакомления специалиста в данной области техники, можно найти в следующих ссылках: Sambrook et al., Molecular Cloning:Short Protocols in Molecular Biology, 4th ed., John WileySons, Inc., 999. Дополнительная и конкретная подробная информация приведена ниже. Рекомбинантные нуклеиновые кислоты, кодирующие белковые конструкции, могут быть введены в клетки-хозяева при помощи множества хорошо известных способов, таких как электропорация, опосредованная липосомами трансфекция (например, с использованием имеющегося в продаже реагента для липосомальной трансфекции, такого как LIPOFECTAMINE 2000 или TRANSFECTIN), осаждение фосфатом кальция, инфицирование, трансфекция и т.п., в зависимости от выбранных векторов и клетокхозяев. Клетки-хозяева можно выращивать в обычных питательных средах, модифицированных таким образом, чтобы они подходили для активации промоторов, выбранных трансформантов или амплификации встроенных полинуклеотидных последовательностей. Условия выращивания, такие как температура, pH и т.п., типично являются такими, какие использовались ранее для клеток-хозяев, выбранных для экспрессии, и будут понятны специалистам в данной области техники и в цитированных здесь ссылках, включая,например, Freshney (1994), Culture of Animal Cells, a Manual of Basic Technique, third edition, Wiley-Liss,New York и цитированные там ссылки. В некоторых воплощениях подходящая температура культивирования составляет 16C; в других - 37C. Продукты экспрессии, соответствующие нуклеиновым кислотам по изобретению, также могут продуцироваться в клетках, отличающихся от клеток животных, таких как растения, дрожжи, грибы, бактерии и т.п. Дополнительно к Sambrook, Berger and Ausubel, подробную информацию, относящуюся к клеточной культуре, можно найти в Payne et al. (1992), Plant Cell and TissueHeidelberg New York) и Atlas and Parks (eds), The Handbook of Microbiological Media (1993), CRC Press,Boca Raton, FL. Клетки-хозяева. Таким образом, клетки-хозяева, которые включают нуклеиновые кислоты, кодирующие рекомбинантную белковую конструкцию, также представляют собой признак этого изобретения. Благоприятные клетки-хозяева включают прокариотические (т.е. бактериальные) клетки-хозяева, такие как E.coli, а также многочисленные эукариотические клетки-хозяева, включая клетки грибов (например, дрожжей, таких как Saccharomyces cerevisiae и Picchia pastoris), клетки насекомых, клетки растений и клетки млекопитающих (такие как клетки СНО). Нуклеиновые кислоты, кодирующие рекомбинантный белок, вводят(например, трансдуцируют, трансформируют или трансфицируют) в клетки-хозяева, например, при помощи вектора, такого как экспрессирующий вектор. Как здесь описано, вектор наиболее типично представляет собой плазмиду, но такие векторы также могут представлять собой, например, вирусную частицу, фаг и т.д. Примеры подходящих экспрессирующих хозяев включают бактериальные клетки, такие как E.coli, Streptomyces и Salmonella typhimurium; клетки грибов, такие как Saccharomyces cerevisiae, Pichia pastoris и Neurospora crassa; клетки насекомых,такие как Drosophila и Spodoptera frugiperda; клетки млекопитающих, такие как 3 Т 3, COS, CHO, BHK,HEK293 или Bowes melanoma; клетки растений, включая клетки водорослей, и т.д. Клетка-хозяин возможно выбрана по ее способности модулировать экспрессию встроенной последовательности или процессировать экспрессированный белок желаемым образом. Такие модификации белка включают гликозилирование (а также, например, ацетилирование, карбоксилирование, фосфорилирование, липидизацию и ацилирование), но не ограничиваются ими. Посттрансляционный процессинг,например, расщепляющий форму-предшественника на зрелую форму белка (например, посредством фуриновой протеазы), возможно осуществляют в контексте клетки-хозяина. Различные клетки-хозяева, такие как 3 Т 3, COS, CHO, HeLa, BHK, MDCK, HEK293, WI38 и т.д., обладают специфическим клеточным аппаратом и характерными механизмами для таких посттрансляционных активностей и могут быть выбраны для обеспечения правильной модификации и процессинга введенного чужеродного белка. Для длительной продукции с высоким выходом раскрытых здесь рекомбинантных белковых конструкций типично используют стабильные системы экспрессии. Например, молекулы нуклеиновой кислоты, которые стабильно экспрессируют белковую конструкцию, вводят в клетку-хозяина с использованием экспрессирующих векторов, которые содержат вирусные ориджины репликации или эндогенные экспрессирующие элементы и селективный маркерный ген. После введения вектора клетки оставляют расти в течение 1-2 суток в обогащенных средах, а затем переводят в селективные среды. Задача селективного маркера заключается в том, чтобы придать резистентность для отбора, и его присутствие дает возможность для роста и выделения клеток, которые успешно экспрессируют введенные последовательности. Например, резистентные группы или колонии стабильно трансформированных клеток могут быть распространены с использованием способов обращения с культурой ткани, подходящих для типа клеток. Клетки-хозяева, трансформированные нуклеиновой кислотой, кодирующей белковую конструкцию, возможно выращивают в условиях, подходящих для экспрессии и выделения кодируемого белка из клеточной культуры. Экспрессированные белковые конструкции могут быть выделены и очищены из рекомбинантных клеточных культур при помощи любого из множества способов, хорошо известных в области техники,включая осаждение сульфатом аммония или этанолом, кислотную экстракцию, фильтрацию, ультрафильтрацию, центрифугирование, анион- или катионообменную хроматографию, фосфоцеллюлозную хроматографию, хроматографию на основе гидрофобного взаимодействия, аффинную хроматографию(например, с использованием любой из указанных здесь систем маркирующих последовательностей),гидроксилапатитную хроматографию и лектиновую хроматографию. Могут быть использованы желаемые стадии рефолдинга белка для заключительной конфигурации зрелого белка. Наконец, высокоэффективная жидкостная хроматография (HPLC) может быть использована для конечных стадий очистки. Дополнительно к указанным здесь ссылкам, в области техники хорошо известны различные способы очистки, включая, например, изложенные в Sandana (1997), Bioseparation of Proteins, Academic Press, Inc.;Applications, Second Edition Wiley-VCH, NY и Walker (1998), Protein Protocols on CD-ROM Humana Press,NJ. В некоторых примерах нуклеиновые кислоты вводят в клетки посредством векторов, подходящих для введения и экспрессии в прокариотических клетках, например клетках E.coli. Например, нуклеиновая кислота, включающая полинуклеотидную последовательность, кодирующую белковую конструкцию,может быть введена в любой из множества имеющихся в продаже или индивидуально изготовленных векторов, таких как серии рЕТ экспрессирующих векторов (например, рЕТ 9b и pET2d). Экспрессию кодирующей последовательности вызывает изопропил -D-1-тиогалактопиранозид (IPTG), что приводит в результате к высоким уровням экспрессии белка. Полинуклеотидная последовательность, кодирующая белковую конструкцию CASB7439, транскрибируется под управлением промотора фага T7. Также подходят альтернативные векторы, такие как pURV22, которые включают индуцируемый теплом промотор лямбда pL. Экспрессирующий вектор вводят (например, путем электропорации) подходящему хозяинубактерии. Существуют многочисленные подходящие штаммы Е.coli, и они могут быть выбраны специалистом в данной области техники (например, штаммы BLR DE3, BL21 DE3 и Rosetta DE3 признаны благоприятными для экспрессии рекомбинантных векторов, содержащих полинуклеотидные последовательности, которые кодируют белковые конструкции). Возможно, что полинуклеотиды, кодирующие белковые конструкции, включены в экспрессирующие векторы, подходящие для введения и экспрессии у эукариот (например, насекомых или клетках млекопитающих). Благоприятным образом, такие нуклеиновые кислоты оптимизированы по кодонам для экспрессии в выбранном векторе/клетке хозяина. Выбранные клетки могут быть клонированы и охарактеризованы в отношении экспрессии желаемой белковой конструкции. Способы оптимизации кодона известны в области техники. Кроме того, коммерческие организации, обеспечивающие сервис в области молекулярной биологии, предлагают оптимизацию кодона, кроме других стандартных технических сервисов. Прокариотические. В бактериальных системах множество экспрессирующих векторов может быть выбрано в зависимости от предполагаемого применения экспрессируемого продукта. Например, когда большие количества полипептида или его фрагментов необходимы для продукции антител, тогда благоприятно использование векторов, которые обеспечивают высокий уровень экспрессии слитых белков, которые легко очищаются. Такие векторы включают мультифункциональные клонирующие и экспрессирующие векторыE.coli, такие как BLUESCRIPT (Stratagene), в которых интересующая кодирующая последовательность,например описанный здесь полинуклеотид по изобретению, может быть лигирована в векторе внутри рамки с последовательностями для N-концевой трансляции, инициируя метионин и последующие 7 остатков бета-галактозидазы продуцировать каталитически активный слитый с бета-галактозидазой белок;Wl), в которых N-концевой метионин лигирован внутри рамки с гистидиновой последовательностью; и т.п., но не ограничиваются ими. В некоторых воплощениях подходящий вектор представляет собой рЕТ 19; в других рЕТ 24; в других рЕТ 26. Эукариотические. Аналогично, в дрожжах, таких как Saccharomyces cerevisiae, множество векторов, содержащих конститутивные или индуцируемые промоторы, такие как альфа-фактор, алкогольоксидаза и PGH, может быть использовано для продукции желаемых продуктов экспрессии. Обзор см. в Berger, Ausubel, и, например, Grant et а/. (1987; Methods in Enzymology 153:516-544). В клетках-хозяевах млекопитающих может быть использовано множество экспрессирующих систем, включая плазмиды и вирусные системы. Иммуногенные композиции Фармацевтически приемлемые носители или эксципиенты. Фармацевтически приемлемые носители и эксципиенты хорошо известны и могут быть выбраны специалистом в данной области техники. Например, носитель или эксципиент благоприятным образом может включать буфер. Возможно, носитель или эксципиент также содержит по меньшей мере один компонент, который стабилизирует растворимость и/или стабильность. Примеры солюбилизато- 13020617 ров/стабилизаторов включают детергенты, например лаурилсаркозин и/или tween. Альтернативные солюбилизаторы/стабилизаторы включают аргинин и стеклообразующие полиолы (такие как сахароза, трегалоза и т.п.). Многочисленные фармацевтически приемлемые носители и/или фармацевтически приемлемые эксципиенты известны в области техники и описаны, например, в Remington's Pharmaceutical Sciences, by E.W. Martin, Mack Publishing Co., Easton, PA, 5th Edition (975). Соответственно, подходящие эксципиенты и носители могут быть выбраны специалистами в данной области техники для получения композиции, подходящей для доставки субъекту при помощи выбранного пути введения. Подходящие эксципиенты включают, без ограничения, глицерин, полиэтиленгликоль (PEG), сорбит, трегалозу, N-лауроилсаркозин натриевую соль, L-пролин, недетергентный сульфобетаин, гуанидин гидрохлорид, мочевину, оксид триметиламина, KCl, Ca2+, Mg2+, Mn2+, Zn2+ и другие относящиеся к двухвалентному катиону соли, дитиотреитол, дитиоэритрол и -меркаптоэтанол. Другие эксципиенты могут представлять собой детергенты (включая Tween 80, Tween 20, Triton X-00, NP-40, Empigen BB,октилглюкозид, лаурилмальтозид, Zwittergent 3-08, Zwittergent 3-0, Zwittergent 3-2, Zwittergent 3-4,Zwittergent 3-6, CHAPS, дезоксихолат натрия, додецилсульфат натрия, бромид цетилтриметиламмония). Адъюванты. Возможно, иммуногенные композиции также включают адъювант. В контексте иммуногенной композиции, подходящей для введения субъекту, такому как субъект-человек, для задач индукции иммунного ответа против CASB7439, адъювант выбран таким образом, чтобы индуцировать Th1 иммунный ответ. Адъювант типично выбран таким образом, чтобы усилить Th1 иммунный ответ у субъекта или популяции субъектов, которым вводят композицию."Th1" тип иммунного ответа характеризуется индукцией CD4+ Т-хелперных клеток, которые продуцируют IL-2 и IFN. Наоборот,"Th2" тип иммунного ответа характеризуется индукцией CD4+ хелперных клеток, продуцирующих IL-4, IL-5 и IL-13. Аффекторы TLR, включая, без ограничения, 3D-MPL. Один из подходящих адъювантов для применения в комбинации с модифицированными полипептидами CASB7439 представляет собой модулятор TLR-4. Один из примеров представляет собой нетоксичное производное липида А, такое как монофосфориллипид А или, конкретней, 3-деацилированный монофосфориллипид A (3D-MPL). 3D-MPL продается под названием MPL от GlaxoSmithKline BiologicalsS.A. и в документе обозначается как MPL или 3D-MPL, см., например, патенты США 4436727; 4877611; 4866034 и 4912094. 3D-MPL прежде всего способствует CD4+ Т-клеточным ответам с фенотипом IFN (Th1). 3D-MPL может быть продуцирован в соответствии со способами, раскрытыми вGB 2220211 А. Химически он представляет собой смесь 3-деацилированного монофосфориллипида А с 3, 4, 5 или 6 ацилированными цепями. В композициях по настоящему изобретению может быть использован 3D-MPL с небольшим размером частиц. 3D-MPL с небольшим размером частиц имеет такой размер частиц, что он может быть подвергнут стерильной фильтрации через фильтр 0,22 мкм. Такие препараты описаны в WO 94/21292. В других воплощениях липополисахарид может представлять собой (1-6) глюкозаминовый дисахарид, описанный в патенте США 6005099 и европейском патенте 0729473 В 1. Специалист в данной области техники способен легко получить различные липополисахариды, такие как 3D-MPL, на основе информации, изложенной в указанных ссылках. Дополнительно к вышеупомянутым иммуностимуляторам (близким по структуре с LPS, или MPL, или 3D-MPL), ацилированные моносахаридные и дисахаридные производные, представляющие собой субфрагмент приведенной здесь структуры MPL, также представляют собой подходящие адъюванты. В других воплощениях адъювант представляет собой синтетические производные липида А, некоторые из которых описаны как агонисты TLR-4 и включают ОМ 174 (2-дезокси-6-о-[2-дезокси-2-[(R)-3-додеканоилокситетрадеканоиламино]-4-офосфоD-глюкопиранозил]-2-[(R)-3-гидрокситетрадеканоиламино]Dглюкопиранозилдигидрофосфат), WO 95/14026; ОМ 294OM197 MP-Ac DP (3S-,9R)-3-[(R)-додеканоилокситетрадеканоиламино]-4-оксо-5-аза-9-[(R)-3 гидрокситетрадеканоиламино]декан-1,10-диол, 1-дигидрофосфат 10-(6-аминогексаноат), WO 01/46127,но не ограничиваются ими. Другие лиганды TLR-4, которые могут быть использованы, представляют собой алкил глюкозаминидфосфаты (AGP), такие как раскрытые в WO 98/50399 или патенте США 6303347 (также раскрыты способы получения), подходящим образом RC527 или RC529, или фармацевтически приемлемые солиAGP, как раскрыто в патенте США 6764840. Некоторые AGP представляют собой агонисты TLR-4, а некоторые представляют собой антагонисты TLR-4. Полагают, что и те и другие полезны в качестве адъювантов. Другие подходящие лиганды TLR-4, способные вызывать сигнальный ответ посредством TLR-4(Sabroe et al., J1, 2003, р. 1630-5), представляют собой, например, липополисахарид грамотрицательных бактерий и его производные или его фрагменты, в частности нетоксичное производное LPS (такое как 3D-MPL). Другие подходящие агонисты TLR представляют собой белок теплового шока (HSP) 10, 60, 65,70, 75 или 90; поверхностно-активный белок А, гиалуронановые олигосахариды, гепарансульфатные фрагменты, фибронектиновые фрагменты, фибриногеновые пептиды, -дефенсин-2 и мурамилдипептид(MDP). В одном из воплощений агонист TLR представляет собой HSP 60, 70 или 90. Другие подходящие лиганды TLR-4 являются такими, как описано в WO 2003/011223 и в WO 2003/099195, такие как соединение I, соединение II и соединение III, раскрытые на стр. 4, 5 в WO 2003/011223 или на стр. 3, 4 вWO 2003/099195 и, в частности, соединения, раскрытые в WO 2003/011223 как ER803022, ER803058,ER803732, ER804053, ER804057, ER804058, ER804059, ER804442, ER804680 и ER804764. Например,один из подходящих лигандов TLR-4 представляет собой ER804057. Сапониновые адъюванты. Другие адъюванты, которые могут быть использованы в иммуногенных композициях с CASB7439,например, сами по себе или в комбинации с 3D-MPL или другим описанным здесь адъювантом, представляют собой сапонины, такие как QS21. Сапонины обсуждают в Lacaille-Dubois, M. and Wagner H. (1996), A review of the biological andpharmacological activities of saponins. Phytomedicine, vol 2, p. 363-386). Сапонины представляют собой стероидные или тритерпеновые гликозиды, широко распространенные среди растений и морских животных. Сапонины известны образованием коллоидных растворов в воде, которые вспениваются при встряхивании, и осаждением холестерина. Когда сапонины находятся рядом с клеточными мембранами, они формируют структуры типа пор в мембране, что приводит к нарушению мембраны. Гемолиз эритроцитов представляет собой пример этого явления, представляющего собой свойство некоторых, но не всех сапонинов. Сапонины, в частности иммунологически активные фракции сапонинов, известны в качестве адъювантов в вакцинах для системного введения. Адъювант и гемолитическая активность индивидуальных сапонинов широко исследуются в области техники (Lacaille-Dubois and Wagner, выше). Например, Quil А(полученный из коры южно-американского дерева Quillaja Saponaria Molina) и его фракции описаны вQS17 (фракции Quil А, очищенные путем ВЭЖХ) описаны в качестве сильных системных адъювантов, и способ их получения раскрыт в патенте США 5057540 и ЕР 0362279 В 1. Другие сапонины, используемые в исследованиях по системному введению вакцин, включают вакцины, происходящие из других видов растений, таких как Gypsophila и Saponaria (Bomford et al., Vaccine, 10(9):572-577,1992). Показано, что такие композиции, содержащие QS21 и холестерин, являются хорошими Th1 стимулирующими адъювантами при приготовлении вместе с антигеном. Таким образом, например, CASB7439 благоприятно может быть использован в иммуногенных композициях с адъювантом, содержащим комбинацию QS21 и холестерина. Иммуностимулирующие олигонуклеиновые кислоты. Один из подходящих адъювантов для применения в комбинации с CASB7439 представляет собой агонист TLR бактериальной ДНК, способный вызывать сигнальный ответ посредством TLR-9, т.е. агонист TLR-9, более конкретно ДНК, содержащей неметилированные CpG нуклеотиды, в конкретных контекстах последовательностей, известных как мотивы CpG. CpG-содержащие олигонуклеотиды вызывают преимущественно Th1 ответ. Такие олигонуклеотиды хорошо известны и описаны, например, вCpG представляют собой олигонуклеотиды CpG. Подходящие олигонуклеотиды для применения в иммуногенных композициях по настоящему изобретению представляют собой олигонуклеотиды, содержащие CpG, возможно содержащие два или более динуклеотидных CpG мотива, разделенных по меньшей мере тремя, подходящим образом по меньшей мере шестью или более нуклеотидами. Мотив CpG представляет собой цитозиновый нуклеотид с последующим гуаниновым нуклеотидом. ОлигонуклеотидыCpG по настоящему изобретению типично представляют собой дезоксинуклеотиды. В конкретном воплощении межнуклеотидный компонент в олигонуклеотиде представляет собой фосфородитиоат, или подходящим образом фосфоротиоатную связь, хотя фосфодиэфир и другие межнуклеотидные связи входят в объем изобретения. Некоторые воплощения представляют собой олигонуклеотиды со смешанными межнуклеотидными связями. Способы получения фосфоротиоатных олигонуклеотидов или фосфородитиоата описаны в патентах США 5666153, 5278302 и WO 95/26204. В альтернативном воплощении используют агонист TLR, способный вызывать сигнальный ответ посредством TLR-9. В одном из воплощений агонист TLR, способный вызывать сигнальный ответ посредством TLR-9, представляет собой HSP90. Комбинации различных адъювантов, таких как упомянутые здесь, также могут быть использованы в композициях с CASB7439. Например, как уже указывалось,QS21 может быть приготовлен вместе с 3D-MPL. Отношение QS21:3D-MPL, как правило, имеет порядок от 1:10 до 10:1; такой как от 1:5 до 5:1 и часто, по существу, 1:1. Как правило, отношение находится в диапазоне от 2,5:1 до 1:1 3D-MPL:QS21. Еще один адъювант комбинации представляет собой 3D-MPL вместе с QS21 в липосомальной композиции, также в комбинации с CpG. Субъекты, имеющие колоректальный рак. Выбор субъекта, имеющего колоректальный рак, для участия в описанных здесь способах может быть осуществлен при помощи клинической диагностики, молекулярной диагностики или их комбинации. В некоторых воплощениях субъекта тестируют для определения того, экспрессируют ли его раковые клетки CASB7439 (HASH2). Клинические способы диагностики колоректального рака известны в области техники. Молекулярные способы и способы определения того, экспрессирует ли раковая клетка или ткань CASB7439 (HASH2), раскрыты в WO 01/62778. Экспериментальные примеры Пример 1. Две молекулы нуклеиновой кислоты, кодирующие белковые конструкции, содержащие полипептидCASB7439, конструировали в векторе pET19b. His-метку располагали рядом с N-концом последовательности CASB7439. Эти клоны назвали LVL007 [SEQ ID NO: 15(aa); SEQ ID NO: 16 (ДНК)] и LVL010[SEQ ID NO: 17 (aa); SEQ ID NO: 18 (ДНК)] соответственно). Наблюдали, что имела место непреднамеренная мутация в клоне LVL007, приводя в результате к одной аминокислотной замене пролина на лейцин (по аминокислотному остатку 17 относительно SEQ ID NO: 13). Молекулы нуклеиновой кислоты использовали с тремя различными типами клеток-хозяев, а именно штаммами E.coli BLR DE3, BL21 DE3 и Rosetta DE3. Индукции осуществляли при двух температурах на индукцию (16 или 37C) и с 1 мМIPTG (изопропилтиогалактозид). См. нижеприведенный раздел "Материалы и способы" в отношении подробной информации о протоколе. Белки собирали для каждой индукции и анализировали при помощи вестерн-блоттинга и при помощи SDS-PAGE с окрашиванием Кумасси голубым. В этих экспериментах белок обнаруживается только на вестерн-блоте. Пример 2. Осуществляли биоинформатические анализы гена CASB7439, выявившие следующие структурные особенности: структурный элемент РНК типа гребенки, расположенный между нуклеотидами 2 и 27 вSEQ ID NO: 14; содержание GC в нуклеотидной последовательность, составляющее 62%; нуклеотидную последовательность, богатую аргининовыми кодонами (26/193 кодированных аминокислот); аминокислотную последовательность с высоким уровнем основных аминокислотных остатков; аминокислотную последовательность с высокой изоэлектрической точкой (11, 18); область с низкой вариабельностью (low complexity) в аминокислотной последовательности (остатки 7-27 в SEQ ID NO: 13; домен спираль-петля-спираль (HLH) (IPR000014) в аминокислотной последовательности (остатки 56-108 в SEQ ID NO: 13); первую область присущего расстройства в аминокислотной последовательности (остатки 118-164SEQ ID NO: 13); вторую область присущего расстройства в аминокислотной последовательности (остатки 118-164 вSEQ ID NO: 13). Авторы изобретения также идентифицировали область с высокой периодичностью пролина от аминокислоты 127 по 158. Заявители обозначили область аминокислот 133-153 включительно вSEQ ID NO: 13 (аа) как богатую пролином область. Богатая пролином область вместе с некоторыми другими особенностями, упомянутыми в этом примере, схематически изображены на фиг. 1 вместе с упомянутыми здесь областями. Пример 3. Немодифицированная нуклеотидная последовательность ДНК CASB7439 (SEQ ID NO: 14) оптимизирована по кодонам фирмой, осуществляющей услугу синтеза ДНК на коммерческой основе, GeneartRegensburg, Germany. Нуклеотидную последовательность CASB7439 получали из клона при помощи амплификации путем ПЦР и использовали для построения двух конструкций. В первой конструкцииLVL016 [SEQ ID NO: 29 (аа), SEQ ID NO: 30 (ДНК)] последовательность CASB7439 встраивали в вектор рЕТ 21b (+) в слиянии с белком pD1/3 белок (из Haemophilus influenzae) по N-концевой последовательности CASB7439. Во второй конструкции LVL018 [SEQ ID NO: 31 (аа); SEQ ID NO: 32 (ДНК)] CASB7439 встраивали в вектор рЕТ 21b (+) (без какого-либо компонента белка D). Эти конструкции оценивали в отношении продукции в BLR DE3 E.coli при 16 и 37C с 1 мМ IPTG. Продукцию обнаруживали при помощи вестерн-блоттинга в виде нерастворимого белка, в основном продуцируемого при 16C. Четыре домена CASB7439 выбирали для дальнейшего исследования для улучшения продукции белка: нацеливающий на ядро домен, богатую пролином область, ДНК-связывающий домен и доменHLH. Пример 4. Нацеливающий на ядро домен, ДНК-связывающий домен, домен bHLH и богатую пролином область CASB7439 выбирали для исследования продукции белка конструкциями, содержащими нуклеотидные последовательности, кодирующие полипептиды CASB7439. Различные участки нуклеотидной последовательности CASB7439 амплифицировали при помощи ПЦР из оптимизированного по кодонам клона Дженеарта, описанного здесь и встроенного в вектор рЕТ 19b (+), приводя в результате к пяти конструкциям, имеющим отличающуюся последовательность CASB7439 (усеченную некоторым образом или полноразмерную); каждая конструкция с полигистидиновой последовательностью по N-концевой последовательности CASB7439, см. фиг. 2. Эти конструкции, представленные на фиг. 2, оценивали в отношении экспрессии белка в штаммеBLR DE3 E.coli при 37 и 16C с 1 мМ IPTG. Как изображено на фиг. 2, эти конструкции содержали полноразмерную CASB7439 [LVL060, SEQ ID NO: 19 (аа), SEQ ID NO: 20 (ДНК)]; CASB7439, усеченную для удаления нацеливающего на ядро домена [Const-1, SEQ ID NO: 21 (аа), SEQ ID NO: 22 (ДНК)];CASB7439, усеченную для удаления богатой пролином области [LVL055, SEQ ID NO: 1 (аа),SEQ ID NO: 2 (ДНК)]; CASB7439, усеченную для удаления ДНК-связывающего домена [LVL056,SEQ ID NO: 23 (аа), SEQ ID NO: 24 (ДНК)]; CASB7439, усеченную для высвобождения основного домена спираль-петля-спираль (bHLH) [LVL057, SEQ ID NO: 25 (аа), SEQ ID NO: 26 (ДНК)]. Вследствие экспериментальной ошибки Const-1 был лишен стоп-кодона, что приводило в результате к трансляции фрагмента нуклеотидной последовательности вектора (приводя в результате к дополнительной 21 аминокислоте в C-концевой области, начиная с LED). Продукцию LVL055 после инкубации при 16C (усеченная по C-концу) ясно обнаруживали на окрашенном Кумасси голубым геле для SDS-PAGE. Пример 5. На основе результатов, описанных в примере 4, разработаны новые конструкции для (1) удаления богатой пролином области при (2) сохранении других областей C-концевого фрагмента полипептида. См. описание на фиг. 3 и 4, в которых обсуждаются (1) различные идентифицированные или предсказанные эпитопы в полипептиде и (2) различные альтернативные подходы к удалению богатой пролином области. Оптимизированная по кодонам синтетическая нуклеиновая генная последовательность, кодирующая модифицированный полипептид CASB7439, получена затем из Geneart (уникальный идентификатор 0706840), расщеплена и встроена в параллели в два отличающихся вектора для построения следующих конструкций:(1) LVL088, содержащей ген CASB7439 ген с делецией богатой пролином области из 21 аминокислоты, слитый с полигистидиновой последовательностью по C-концу в векторе рЕТ 26b (+)(2) LVL111, содержащей ген CASB7439 с делецией богатой пролином области из 21 аминокислоты и полигистидиновой последовательностью как по N-, так и C-концу в векторе рЕТ 19b (+)(SEQ ID NO: 3 (аа), SEQ ID NO: 4 (ДНК. Конструкцию LVL111 (см. SEQ ID NO: 3) клонировали в соответствии с той же самой стратегией,используемой для LVL055, т.е. с использованием вектора рЕТ 19 (+), который включает полигистидиновую последовательность, состоящую из 10 гистидинов с последующим сайтом энтерокиназы. Это приводило в результате к образованию партнера слияния из 23 аминокислот (SEQ ID NO: 42) на N-конце. Другие конструкции конструировали с оптимизированным по кодонам синтетическим геном, обсуждающимся в примере 3 (уникальный идентификатор 0606597), включающие следующие:LVL114 (белковая конструкция, содержащая (с N-конца) белок SUMO, CASB7439, усеченный по аминокислоте 117, в векторе pSUMO) (SEQ ID NO: 49 (aa); SEQ ID NO: 50 (ДНК;CASB7439 с удаленной богатой пролином областью, в векторе pSUMO) (SEQ ID NO: 51 (aa);He обнаружено никакого улучшения продукции, хотя осуществляли только один повтор (данные не представлены). Продукцию LVL088 анализировали после индукции в течение ночи с использованием 1 мМ IPTG при 37 и при 16C. При анализе путем электрофореза продукцию LVL088 визуализировали только при помощи вестерн-блоттинга. Одна партия, очищенная из четырех культуральных колб по 800 мл, приводила в результате к 0,4 мг белка. С другой стороны, LVL111 легко обнаруживали после окрашивания Кумасси голубым. Рекомбинантный белок LVL111 продуцировали и очищали в 8 партиях. Несколько партий LVL111 (каждая из четырех культуральных колб по 800 мл) оценивали в отношении продукции в диапазоне от 3,3 мг/партию до 10 мг/партию очищенного белка. Таким образом, продукция LVL088 была ниже по сравнению с обнаруженной для LVL111. Очищенный белок также тестировали в отношении его реактивности с несколькими моноклональными антителами против CASB7439 и во всех случаях обнаружили положительную реактивность. Пример 6. Исследовали удаление C-концевой полигистидиновой последовательности из конструкции LVL111. Молекулу нуклеиновой кислоты, кодирующую полипро(-)полипептид CASB7439 без фрагмента, кодирующего C-концевой полигистидин, амплифицировали при помощи ПЦР из конструкции LVL111 и клонировали в вектор рЕТ 19b. Эту конструкцию обозначали как LVL137. Конструкция LVL137, по существу, представляет собой LVL111 без нуклеотидной последовательности, кодирующей C-концевую полигистидиновую последовательность. SEQ ID NO: 5 (aa); SEQ ID NO: 6 (ДНК). Удаление C-концевой полигистидиновой последовательности, по-видимому, не оказывает действия на уровень продукции. В соответствии с анализом при помощи окрашенного Кумасси геля после SDS-PAGE LVL137 продуцировал тот же самый уровень белка, как и LVL111. Молекулу нуклеиновой кислоты, кодирующую полипро(-)полипептид CASB7439, затем амплифицировали при помощи PCR из синтетической последовательности Geneart (0706840) CASB7439 и клонировали в вектор рЕТ 26 с концевой последовательностью из шести полигистидинов на N-конце (LVL138).SEQ ID NO: 33 (аа); SEQ ID NO: 34 (ДНК). Эта конструкция демонстрировала значительно уменьшенный уровень продукции белка и ее сложно обнаружить при помощи вестерн-блоттинга. Молекулу нуклеиновой кислоты, кодирующую последовательность шести полигистидинов поN-концу полипро(-)полипептида CASB7439, конструировали в устойчивом к канамицину векторе рЕТ 24. Новую конструкцию линкера использовали для удаления структуры типа гребенки, сформировавшейся между полигистидиновой концевой последовательностью и 5' концом гена CASB7439. Конкретно, кодоны для первых восьми аминокислотных остатков белковой конструкции заменяли следующей нуклеотидной последовательностью: 5' ATG CAT CAT CAT CAT CAT CAT GAC3' (SEQ ID NO: 35). (Исходные кодоны представляли собой 5' ATG CAC CAT CAC CAT CAC CAT GAT3' (SEQ ID NO: 36).) Эту конструкцию обозначали как LVL160. SEQ ID NO: 37 (aa); SEQ ID NO: 38 (ДНК). Как определено анализом на геле и вестерн-блоттингом, продукция белка, по-видимому, является столь же слабой, как и дляLVL138. Молекулу нуклеиновой кислоты, кодирующую белковую конструкцию с концевой последовательностью из десяти полигистидинов по N-концу полипро(-)полипептида CASB7439, конструировали в устойчивом к канамицину векторе рЕТ 24. Эту конструкцию идентифицировали как LVL168.SEQ ID NO: 11 (аа); SEQ ID NO: 12 (ДНК). Уровень продукции сравним с уровнем, обнаруженным дляLVL111. Сопоставление нескольких конструкций, обсуждавшихся в этом примере, представлено на фиг. 5. Результаты для конструкций, обсуждавшихся в этом примере и примере 1, обобщены в табл. 1. Таблица 1. Уровни экспрессии для конструкции, обсуждавшейся в примерах 1, 3, и 6. Конструкцию,которая приводит к продукции "гель-/блот+" в общем, обнаруживают только при помощи вестернблоттинга, когда лизат целых клеток подвергают SDS-PAGE. Конструкцию, которая приводит к продукции "гель/блот ", ясно обнаруживают при помощи окрашивания Кумасси голубым, когда лизат целых клеток подвергают SDS-PAGE. Таблица 1 6His: концевая последовательность из гистидинов = 6 гистидиновых остатков.Del-poly-Pro-domain: CASB7439, модифицированный путем делеции богатой пролином области. ОРТ: оптимизированный по кодонам.Hismodif: гистидиновая концевая последовательность, модифицированная для выбора кодона (см. пример 6). 1/3pD: конструкция белка D, описанная здесь. 10His: гистидиновая последовательность с 10 гистидиновыми остатками. Пример 7. Еще одну конструкцию CASB7439 конструировали в слиянии с фрагментом белка D изMet-Asp-Pro сливали с 108 N-концевыми аминокислотами процессированного Белка D (аминокислотные остатки 2-109 процессированного Белка D). SEQ ID NO: 39 названа здесь как 1/3pD или 1/3 Белок D. Синтетическую нуклеотидную последовательность CASB7439, полученную из Geneart (0706840) (удалена пролинбогатая область), субклонировали в pET26-1/3pD, представляющей собой устойчивую к канамицину плазмиду, которая содержит 1/3pD, с получением LVL141. Последовательность LVL141 представлена в SEQ ID NO: 7 (аа) и SEQ ID NO: 8 (ДНК). Продукция сравнима с продукцией LVL111. Затем производили мутацию этого клона при помощи ПЦР для удаления линкера из четырех аминокислот (Ala-Ala-Ala-His) между компонентами 1/3pD и CASB7439. Последовательность получающейся в результате конструкции, обозначенной LVL144, представлена на SEQ ID NO: 9 (аа) и SEQ ID NO: 10(SEQ ID NO: 13); LVL168 (SEQ ID NO: 12); PD1/3 (SEQ ID NO: 39) и LVL144(SEQ ID NO: 9). Пример 8. Тесты солюбилизации LVL168. В общем, очищенные белковые конструкции, содержащие CASB7439, солюбилизировали в 6 М гуанидиновом буфере, переносили в буфер на основе 8 М мочевины и хранили в 4 М мочевине, 20 мМHEPES, 1 мМ TCEP (трихлорэтилфосфат), 150 мМ NaCl, pH 7,0. Предварительные результаты, полученные на небольших объемах, свидетельствуют о том, что LVL168 также может быть солюбилизирован в фосфатном буфере без мочевины, а также каких-либо восстановителей, таких как EDTA (этилендиаминтетрауксусная кислота) или TCEP, и с NaCl или без него. Никакого осаждения не обнаруживали после диализа в этих буферах. Анализы солюбилизации осуществляли для того, чтобы охарактеризовать белокLVL168 в 16 буферах, представленных в табл.2. 100 мкл очищенных образцов белка LVL168 в буфере на основе 8 М мочевины, 20 мМ HEPES, 150 мМ NaCl, 1 мМ TCEP, pH 7,0 помещали в блок для минидиализа 10000 MWCO (Biolynx), предварительно уравновешенный буфером путем двух промываний. Диализ осуществляли в течение ночи в 100 мл каждого из 16 буферов, представленных в табл. 2, при встряхивании при 4C. Образцы затем отбирали из блоков, помещали в пробирки eppendorfs и центрифугировали в течение 5 мин при 20000g. Растворимые белки оставались в суспензии, а нерастворимые белки осаждались и формировали осадок после центрифугирования. Буферы, в которых растворим белокLVL168, представлены в табл. 2. Супернатанты тестировали при помощи SDS-PAGE и концентрации оставшихся растворимых белков измеряли при помощи способа RC DC в соответствии с указаниями производителя. Набор для осуществления способа RC DC, представляющего собой модифицированный метод Лоури, получен изBioRad Laboratories, 1000 Alfred Nobel Drive, Hercules, CA 94547). Образцы оставляли при 4C для тестирования стабильности. Тесты стабильности LVL168. Белки LVL168 после анализа растворимости содержали в течение 7 суток при 4C. 20 мкл каждого из образцов 2, 3, 5, 6, 9-16 переносили в другие пробирки и тестировали в отношении стабильности при 4C, 37C или комнатной температуре в течение еще 2 суток. Образцы центрифугировали в течение 5 мин при 20000g. Супернатанты разделяли при помощи SDS-PAGE и профили анализировали в отношении разрушения, агрегации или утраты белка путем сравнения с исходным очищенным белком. Белок оставался стабильным в большинстве исследованных буферов. Для удобства читателя обобщение упомянутых здесь конструкций и их относительная продукция представлены в табл. 3. Таблица 3 Сокращения см. в табл. 1. В табл. 3 использованы следующие дополнительные сокращения:mut: последовательность, мутированная для удаления аминокислот между pD1/3 и модифицированным полипептидом CASB7439.linker: аминокислоты между двумя фрагментами молекулы. Материалы и способы. Экспрессия - мелкомасштабные индукции. Штаммы E.coli BLR (DE3) трансформировали упомянутыми здесь конструкциями, помещали в планшеты с агаром Луриа-Бертани (LB), содержащие соответствующий антибиотик (40 мкг/мл канамицина или 100 мкг/мл карбеницилина), и инкубировали в течение ночи при 37C. Образующийся в результате бактериальный газон использовали для инокуляции 20 мл LB альтернативной белковой среды (APS) подходящим антибиотиком (40 мкг/мл канамицина или 100 мкг на 1 мл карбеницилина) и помещали на 37C для достижения OD600 от 0,5 до 1,0. Индукции осуществляли путем добавления 1 мМ IPTG при 16 или 37C. Для индукции при 16C растущую культуру охлаждали на льду за 1 ч перед индукцией экспрессии рекомбинантного белка. Культуру затем возвращали назад для роста при 16C и поддерживали при этих условиях в течение 16 ч. Для индукции при 37C экспрессию рекомбинантных белков немедленно индуцировали путем добавления IPTG к растущей культуре. Индукцию осуществляли при 37C в течение 3 ч. Небольшие аликвоты по 1 мл отбирали до и после индукции. Аликвоты затем центрифугировали (10 мин, 4C, 20000g), и бактериальный осадок после центрифугирования хранили при -20C до анализа. Анализ осуществляли после экстракции белка BugBuster в соответствии с рекомендациями производителя. BugBuster, производимый Novagen, имеется в продаже от VWR. Уровень экспрессии определяли путем окрашивания гелей после SDS-PAGE Кумасси голубым и при помощи вестерн-блоттинга. Растворимость оценивали после обнаружения белка в осадках или в супернатантах после экстракции BugBuster. Экспрессия - крупномасштабные индукции. Штаммы E.coli BLR (DE3) трансформировали описанными здесь конструкциями, помещали на планшеты с агаром LB, содержащим соответствующий антибиотик, и инкубировали в течение ночи при 37C. Образующийся в результате бактериальный газон использовали для инокуляции желаемого количества среды LB APS в колбах объемом 800 мл с подходящим антибиотиком (40 мкг/мл канамицина или 100 мкг на 1 мл карбеницилина) и помещали на 37C для достижения OD600 от 0,5 до 1,0. Колбы объемом 2,5 л, содержащие 800 мл ростовой среды, затем охлаждали на льду за 1 ч перед индукцией экспрессии рекомбинантного белка путем добавления 1 мМ IPTG. Для индукции при 16C растущую культуру охлаждали на льду за один час перед индукцией. Культуры затем помещали назад для роста при 16C и поддерживали при этих условиях в течение 16 ч. Культуры центрифугировали (15 мин, 4C, 6000g), и бактериальный осадок после центрифугирвоания хранили при -80C до очистки. Небольшие алквоты по 1 мл также отбирали до и после индукцииBugBuster и анализировали при помощи SDS-PAGE и вестерн-блоттинга перед очисткой. Очистка. Бактериальный осадок ресуспендировали в буфере для лизиса (20 мМ буфер Tris (pH 8,0), содержащий 500 мМ NaCl, 10 мМ TCEP) и смесь ингибиторов протеаз (полные без EDTA (этилендиаминтетрауксусная кислота. Бактерии лизировали путем ультразвуковой обработки с использованием 13 мм зонда на ультразвуковом процессоре Vibra cell, от Emulsiflex C3 (Avestin) или при помощи системы разрушения Constant Cell (Constant System). Растворимые (супернатант) и нерастворимые (осадок) компоненты разделяли путем центрифугирования при 20000g в течение 20 мин при 4C. Поскольку рекомбинантные белки CASB7439 ожидали обнаружить в осадках, супернатанты удаляли. Обычно нерастворимые компоненты (осадок) повторно солюбилизировали в 50 мМ бициновом буфере pH 8,0, содержащем 6 М гуанидин HCl, 500 мМ NaCl, 10 мМ TCEP, и затем центрифугировали(20000g в течение 20 мин). Супернатант наносили на колонку His Trap объемом 5 мл (GE Healthcare),уже предварительно уравновешенную в предшествующем буфере, но концентрацию TCEP уменьшали до 1 мМ. После загрузки колонку однократно промывали тем же самым буфером. 6 М GnHCl хаотропный агент замещали 8 М мочевиной (8 М мочевина, 20 мМ HEPES, 500 мМ NaCl, 1 мМ TCEP, pH 8,0). Элюцию осуществляли с использованием 20 мМ буфера HEPES (pH 8,0), содержащего 8 М мочевину,500 мМ NaCl, 1 мМ TCEP и 250 мМ имидазол. Образцы из элюируемых фракций анализировали при помощи SDS-PAGE и фракции, содержащие интересующий белок, объединяли, при необходимости концентрировали на центрифуге и использовали на второй стадии очистки. Осуществляли одну или две стадии SEC (гель-фильтрация) (Superdex 200, GE Healthcare) в зависимости от требуемой чистоты в 8 М мочевине, 20 мМ HEPES, 500 мМ NaCl, 1 мМ TCEP, pH 8,0 перед последней стадией обессоливания. Обессоливание осуществляли на G25 обессоливающей колонке, предварительно уравновешенной конечным буфером 20 мМ HEPES (pH 7,0), содержащим 4 М мочевину,150 мМ NaCl и 1 мМ TCEP. Белковые образцы аликвотировали по 1 мл пробирки и хранили при -80C. Анализ белка - определение концентрации. Концентрацию белка определяли с использованием RC DC белкового анализа (модифицированный метод по Лоури от Bio-Rad). Этот анализ использовали из-за несовместимости некоторых компонентов конечного буфера с классическими анализами Лоури или BSA. Количество препарата конечного белка также анализировали при помощи SDS-PAGE, вестерн-блоттинга и в отношении содержания LAL для обеспечения или деградации белка E.coli и уровня контаминации LPS. Пример 9. Подтверждение экспрессии CASB7439 в нормальных тканях и тканях, пораженных колоректальным раком (CRC). Заявители проводили анализ экспрессии мРНК CASB7439 с использованием qPCR (количественной ПЦР) в режиме реального времени в 83 образцах CRC и 12 нормальных образцах толстой кишки, полученных из Corixa Corporation. Значения экспрессии мРНК CASB7439 были по меньшей мере в 10 раз выше в CRC по сравнению с нормальной толстой кишкой в 63% тестируемых образцов CRC по сравнению с нормальными тканями (данные приведены в виде усредненного отношения экспрессииCASB7439/-актин) (фиг. 7). Кроме того, заявители обнаружили белок CASB7439 путем иммунофлуоресценции (IF) в человеческих CRC тканях с использованием кроличьего моноклонального антитела против CASB7439 (40-12) (обсуждаемое здесь антитело); никакого количества белка CASB7439 не обнаружено на срезах нормальных соседних тканей. Дополнительные данные по экспрессии для CASB7439 получены при помощи анализа GeneLogic(Gene Microarray) с использованием стандартных способов. Обнаружено, что мРНК CASB7439 сильно экспрессируется в образцах CRC со всех стадий заболевания (данные не представлены). Заявители провели анализ экспрессии мРНК CASB7439 qPCR в режиме реального времени в 83 образцах CRC и 12 образцах нормальной толстой кишки, полученных из Corixa Corporation. Наблюдали сверхэкспрессию мРНК CASB7439 по меньшей мере в 10 раз выше в CRC по сравнению с нормальной толстой кишкой в 63% тестированных образцов CRC по сравнению с нормальными тканями (данные приведены в виде усредненного отношения экспрессии CASB7439/-актин) (фиг. 7). В независимых литературных источниках сообщается об обнаружении мРНК CASB7439, т.е.HASH-2, путем гибридизации in situ (ISH) на очень низком уровне на дне кишечных крипт, но ни в какой из других нормальных тканей, за исключением плаценты. Jubb et al. (2006), Oncogene 25:3445-3457. В тканях, пораженных колоректальным раком (CRC), показано, что мРНК CASB7439 в 15 раз сверхэкспрессируется в 70% тестируемых образцов человеческих тканей. Экспрессия белка CASB7439 в CRC и нормальных соседних тканях толстой кишки. Для анализа экспрессии белка CASB7439 с использованием иммунофлуоресценции в нормальных иCRC тканях сконструировали специфическое кроличье моноклональное антитело против CASB7439 (4012). Кроличьи гибридомы, наработанные против CASB7439, получены из коммерческого источникаEpitomics Inc. и отобраны заявителями. Моноклональные антитела (mAb), продуцируемые гибридомами,сначала отбирали при помощи ELISA (иммуноферментного анализа) с использованием рекомбинантного белка LVL111 (SEQ ID NO: 3). Специфичность моноклональных антител подтверждали при помощи вестерн-блоттинга с использованием LVL111 и клеточного лизата, полученного из рекомбинантной клеточной линии, сконструированной для экспрессии CASB7439 (см. ниже TC1/CASB7439). Экспрессию белкаCASB7439 в опухолях TC1/CASB7439 (выращенных у мышей), клетках SW-620 и SW-480 (положительная в отношении CASB7439 человеческая линия клеток колоректального рака) и в клеткахHCT-116 (отрицательная в отношении CASB7439 человеческая линия клеток колоректального рака) оценивали при помощи иммунофлуоресценции с использованием mAb. mAb 40-12 выбраны для подтверждения экспрессии белка CASB7439. Заявители получили из NDRI (National Disease Research Interchange (Национальная организация по обмену информации по исследованию заболеваний), 8 Penn Center, 8th Floor, 1628 JFK Boulevard, Philadelphia, PA 19103) залитые в OCT замороженные образцы ткани CRC и нормальной соседней ткани 17 различных индивидов; для 5 из этих индивидов также получены метастатические ткани. Заявители отобрали образцы 17 первичных опухолей, 17 нормальных соседних тканей и 5 метастатических тканей. Из 17 проанализированных первичных опухолей CRC 13 оказались положительными в отношении экспрессии белка CASB7439 (76,5%). Все эти положительные в отношении CASB7439 образцы представляли собой аденокарциномы. Ни в одной из нормальных соседних тканей не экспрессировались обнаруживаемые уровни белка CASB7439. Среди метастатических тканей 3 из 5 были положительными (60%) в отношении экспрессии белка CASB7439. Корреляцию между белком CASB7439 и экспрессией мРНК исследовали в 16 тканях CRC при помощи анализа qPCR в режиме реального времени, проводимого с использованием стандартных способов в отношении раковых и нормальных соседних тканей толстой кишки. Окрашивание Ki67 использовали в качестве маркера пролиферации. Корреляция между белком CASB7439 и экспрессией мРНК представлена на фиг. 8. Образцы ткани, которые демонстрировали наличие белка CASB7439 в соответствии с иммунофлуоресценцией, также продемонстрировали самые высокие уровни мРНК CASB7439. Пример 10. Иммуногенность CASB7439 у мышей: Т-клеточная иммуногенность CASB7439 у инбредных мышей. Для исследования Т-клеточной иммуногенности CASB7439 у мышей банк пептидов получали при помощи стандартных способов синтеза для того, чтобы покрыть всю последовательность белкаCASB7439 (фиг. 9). Каждый пептид представлял собой 15-мер, перекрывающийся со следующим пептидом по 11 аминокислотам. Для исследования иммунодоминантных областей 14 совокупностей пептидов расположили в соответствии с матрицей таким образом, что любые две совокупности пептидов имели только один общий пептид (фиг. 10). Для исследования Т-клеточной иммуногенности у мышей до 15 инбредных мышей и до 10 аутбредных мышей на группу получали четыре внутримышечные иммунизации каждые две недели различными дозами (от 0 до 30 мкг) различных рекомбинантных белков CASB7439, приготовленных с AS01B илиAS15, оба из которых представляют собой патентованные адъюванты GSK. После приготовления с антигеном AS01B состоял из MPL (100 мкг/мл) и QS21 (100 мкг/мл) в липосомах). После приготовления с антигеном AS15 состоял из MPL (100 мкг/мл), QS21 (100 мкг/мл) и CpG7909 (840 мкг/мл) в липосомах. Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439, смешанного с 25 мкл 2 концентрированного адъюванта. Таким образом, каждая мышь в группе AS01 В получала 5 мкг MPL и 5 мкг QS21. Каждая мышь в группе AS15 получала 5 мкг MPL, 5 мкг QS21 и 5 мкг CpG7909. После последней иммунизации из мышей отбирали образцы селезенок или PBL (лейкоциты периферической крови). Для анализа иммуногенности CASB7439 у инбредных мышей образцы селезенок или PBL анализировали индивидуально или объединяли. Для анализа иммуногенности CASB7439 у CD1 аутбредных мышей каждый образец селезенки обрабатывали индивидуально.PBL или выделенные спленоциты ресуспендировали в конечной концентрации 10106 клеток на 1 мл в культуральной среде RPMI 1640 со следующими добавками: пенициллин-стрептомицин (1), ненезаменимые аминокислоты (1), 2-меркаптоэтанол (55 мМ), пируват натрия (100 мМ), L-глутамин(200 мМ). Клетки стимулировали пептидами из последовательности CASB7439 (используемого в конечной концентрации 1 мкг/мл/пептид в RPMI 1640 + добавки + 5% инактивированная теплом FBS (фетальная телячья сыворотка. Внутриклеточное окрашивание (ICS) в отношении CD4, CD8, IL-2, IL-5, IFN иTNF осуществляли в соответствии с классическими способами. После стимуляции клетки обрабатывали брефельдином А и фиксировали. Окрашивание поверхностных CD4 и CD8 осуществляли при помощи специфических в отношении CD4 и CD8 моноклональных антител. Затем клетки пермеабилизовали и окрашивали специфическими моноклональными антителами против белков IL-2, IL-5, IFN и TNF. Клетки анализировали при помощи проточной цитометрии с использованием BD FACSCanto II. Данные выражены в процентной доле CD4 или CD8 Т-клеток, положительных в отношении как IFN+, так иTNF+. Эксперимент по сравнению мультилинейной Т-клеточной иммуногенности у инбредных мышей. Заявители сравнили иммуногенность белка CASB7439, приготовленного с AS01B или AS15, у инбредных мышей C57Bl/6, Balb/C, CB6f1 (потомство первого поколения мышей C57Bl/6, скрещенных сBalb/C) и С 3 Н, приобретенных в Charles River, Senneville, QC, Canada. Пять мышей на группу иммунизировали 10 мкг рекомбинантного белка CASB7439 (LVL111), приготовленного с AS01B или AS15. Образцы селезенок собирали и объединяли (один пул из пяти образцов селезенок на группу). CD4 Т-клеточный ответ (положительный в отношении как IFN+, так и TNF+) на LVL111 для каждого штамма представлен на фиг. 11 для композиции AS01B и на фиг. 12 для композиции AS15. Никакой CD8 Т-клеточный(положительный как в отношении IFN+, так и TNF+) ответ не обнаружен в какой-либо из четырех инбредных линий мышей. Иммунодоминантные пептиды CASB7439, идентифицированные у инбредных мышей, иммунизированные CASB7439+AS15, представлены на фиг. 13. Иммунодоминантные пептиды и соответствующие области SEQ ID NO: 13 представляют собой:(а.к. 161-183 в SEQ ID NO: 13). Исследования Т-клеточной иммуногенности LVL111, LVL168 и LVL144 у инбредных мышей. Иммуногенность LVL111, LVL168 и LVL144 исследовали у инбредных мышей CB6f1. В этом эксперименте по пять мышей на группу (Charles River, Senneville, QC, Canada) иммунизировали четыре раза каждые две недели рекомбинантными белками CASB7439 (LVL111, LVL168 и LVL144), приготовленными с AS15. Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439, смешанного с 25 мкл 2 концентрированного описанного здесь адъюванта. CD4 и CD8 Т-клеточные ответы (положительные в отношении как IFN, так и TNF) измеряли путем внутриклеточного окрашивания и при помощи анализа путем проточной цитометрии у индивидуальных инбредных мышей CB6f1 после повторной стимуляции спленоцитов матрицей пептидов CASB7439 (данные не представлены). Иммунизации всеми тремя конструкциями запускали CD4 Т-клеточный иммунный ответ против тех же самых иммуногенных областей (а.к. с 25 по 43 в SEQ ID NO: 13 и а.к. с 89 по 107 в SEQ ID NO: 13) у инбредных мышейCD1 аутбредных мышей (Charles River, Senneville, QC, Canada) иммунизировали четыре раза каждые две недели рекомбинантным белком CASB7439 (LVL111), приготовленным с AS01B или AS15. Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439, смешанного с 25 мкл 2 концентрированного описанного здесь адъюванта. CD4 и CD8 Т-клеточные ответы (положительные в отношении как IFN так и TNF) измеряли путем внутриклеточного окрашивания и при помощи анализа путем проточной цитометрии у индивидуальных CD1 мышей после повторной стимуляции спленоцитов при помощи матрицы пептидов CASB7439. Таблицы CASB7439 CD4-иммуногенных пептидов, идентифицированных у аутбредных мышей,иммунизированных CASB7439+AS01 В или CASB7439+AS15, представлены на фиг. 14 вверху и внизу,соответственно. CD4 Т-клеточные иммуногенные области, идентифицированные у CD1 аутбредных мышей, покрывают следующие области CASB7439 (выраженные с точки зрения аминокислотных позиций вSEQ ID NO: 13) и пептиды 24-26 (а.к. с 93 по 115 в SEQ ID NO: 13). CD8 Т-клеточные иммунодоминантные области, идентифицированные у CD1 аутбредных мышей, покрывают следующие части CASB7439LVL144, тестировали у индивидуальных аутбредных CD1 мышей, приобретенных в Charles River,Senneville, QC, Canada. Пять мышей на группу получали по четыре внутримышечных иммунизации каждые две недели, состоящие из 10 мкг рекомбинантного белка CASB7439 (LVL111, LVL168 или LVL144),приготовленного с AS15. Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439,смешанного с 25 мкл 2 концентрированного описанного здесь адъюванта. Пять мышей в контрольной группе иммунизировали физиологическим раствором. Через две недели после четвертой иммунизации мышей умерщвляли. CD4 и CD8 Т-клеточные ответы (положительные в отношении как IFN так и TNF) измеряли путем внутриклеточного окрашивания и при помощи анализа путем проточной цитометрии после повторной стимуляции спленоцитов четырьмя совокупностями пептидов (пул 1: пептиды 8-9-10, пул 2: пептиды 16-17-18, пул 3: пептиды 24-25-26 и пул 4: пептиды 30-31-32). Данные представлены на фиг. 15. Те же самые CD4 Т-клеточные иммуногенные области SEQ ID NO: 13, а именно пептиды 16-18 (а.к. с 61 по 83 в SEQ ID NO: 13) и пептиды 24-26(а.к. с 93 по 115 в SEQ ID NO: 13) активировали после иммунизации каждой из трех конструкцийCASB7439 + AS15-опосредованные CD4 и CD8 T-клеточные ответы, вызванные двумя конструкциями (LVL168 или LVL144), тестировали у группы HLA A2.1/DR1-трансгенных мышей (3 совокупности из 3 мышей), приобретенных в Pasteur Institute, Paris, France. 9 мышей на группу получали четыре внутримышечные иммунизации каждые две недели, состоящие из 6,25 мкг LVL168 или 10 мкг LVL144,приготовленных с AS15. Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439,смешанного с 25 мкл 2 концентрированного описанного здесь адъюванта. Двух мышей в контрольной группе иммунизировали только адъювантом. Через две недели после четвертой иммунизации мышей умерщвляли. CD4 и CD8 Т-клеточные ответы (положительные в отношении как IFN так и TNF) измеряли путем внутриклеточного окрашивания и при помощи анализа путем проточной цитометрии после повторной стимуляции PBL и спленоцитов 7 совокупностями пептидов, покрывающих всю последовательность CASB7439 [пул 1: пептиды 1-7(SEQ ID NO: 53-59), пул 2: пептиды 8-14 (SEQ ID NO: 60-66), пул 3: пептиды 15-21 (SEQ ID NO: 67-73),пул 4: пептиды 22-28 (SEQ ID NO: 74-80), пул 5: пептиды 29-35 (SEQ ID NO: 81-87), пул 6: пептиды 3642 (SEQ ID NO: 88-94) и пул 7: пептиды 43-46 (SEQ ID NO: 95-98)]. Кроме того, человеческий HLA DR иммунодоминантный пептид 24 (SEQ ID NO: 76) тестировали индивидуально. Результаты, представленные на фиг. 16-19, демонстрируют, что одна и та же CD4 и CD8 Т-клеточная иммунодоминантная область (пептиды 22-28 + пептид 24) активируется после иммунизации каждой из двух конструкцийCASB7439, приготовленных с AS15. Таким образом, пептид 24 также представляет собой человеческийHLA A2.1 иммунодоминантный пептид. Эти результаты подтверждены во втором эксперименте, в котором те же самые пептиды CASB7439 использовали для повторной стимуляции спленоцитов, полученных у 5 индивидуальных мышей на группу, иммунизированных LVL168 или LVL144, приготовленных сCASB7439-опосредованный гуморальный ответ на 10 мкг CASB7439 (LVL111), приготовленного с(Charles River, Senneville, QC, Canada). 19 мышей на группу получали четыре внутримышечные иммунизации каждые две недели. Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439,смешанного с 25 мкл адъюванта (2 концентрация). У мышей отбирали кровь непосредственно перед каждой иммунизацией и через две недели после четвертой иммунизации. В еще одном экспериментеCASB7439-опосредованный гуморальный ответ на LVL111, LVL168 или LVL144 оценивали у инбредных мышей CB6f1 (Charles River, Senneville, QC, Canada). 50 мышей на группу получали четыре внутримышечные иммунизации каждые две недели 10 мкг рекомбинантного белка CASB7439 (LVL111,LVL168 или LVL144), приготовленного с AS15 или только AS15 в качестве контрольной группы. Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439, смешанного с 25 мкл 2 концентрированного описанного здесь адъюванта. У мышей отбирали кровь через две недели после четвертой иммунизации. Титры антител IgG1 и IgG2a в сыворотке крови против CASB7439 определяли с использованиемELISA в соответствии со стандартными способами. Кратко, рекомбинантный белок CASB7439 наносили в 96-луночный планшет в физиологическом растворе (PBS). После блокирования бычьим сывороточным альбумином 1% в PBS-Tween (0,1%) сыворотки крови, иммунизированных мышей инкубировали в планшетах в течение 1 ч при 37C. После промывания планшета на 1 ч при 37C добавляли специфическое козье антитело против мышиного антитела IgG1 или IgG2a, меченого HRP (перокисдаза хрена)(Southern Biotech Associates, Birmingham, AL, USA). После промывания планшета добавляли субстрат ТМВ (тетраметилбензидин) (BD Pharmingen, Mississauga, ON, Canada) на 15 мин. Колориметрическую реакцию останавливали путем добавления 1 М серной кислоты. OD измеряли при 450 нм с использованием спектрофотометра Spectramax (Molecular Device, Sunnyvale, CA, USA). Титр антитела определяли при помощи стандартной кривой с использованием специфических общих антител IgG, IgG1 или IgG2a и мышиной сыворотки крови с CASB7439. На фиг. 20 представлен анализ зависимости от времени титров антител IgG1 и IgG2a в сыворотках крови мышей, иммунизированных CASB7439 (LVL111), приготовленным с AS01B или AS15. Данные демонстрируют, что AS15 запускает более высокий Th1 (приблизительно в 10 раз выше отношениеIgG2a/IgG1 после 4 иммунизации по сравнению с AS01B (IgG2a представляет собой суррогатный маркер иммунного ответа 1 типа). Как представлено на фиг. 21, белок CASB7439, приготовленный с AS15, запускает очень сильный ответ в виде антител IgG1 и IgG2a у инбредных мышей CB6f1 независимо от используемой конструкции (LVL111, LVL168 или LVL144). Иммуногенность CASB7439 у мышей: исследования CASB7439-опосредованного гуморального ответа у мышей CD1.CASB7439-опосредованный гуморальный ответ после иммунизации CASB7439 (LVL111, LVL168 или LVL144) оценивали у аутбредных мышей CD1 (Charles River, Senneville, QC, Canada). Десять мышей на группу получали четыре внутримышечные иммунизации каждые две недели 10 мкг рекомбинантного белка CASB7439 (LVL111, LVL168 или LVL144), приготовленного с AS15 (или только AS15 в качестве контрольной группы). Каждая иммунизация содержала 25 мкл рекомбинантного белка CASB7439, смешанного с 25 мкл 2 концентрированного описанного здесь адъюванта. Через две недели после четвертой иммунизации у мышей отбирали кровь и титры в сыворотке крови общих антител IgG против CASB7439 определяли при помощи ELISA, как описано ранее. У мышейCD1, иммунизированных различными конструкциями CASB7439 (LVL111, LVL168 или LVL144), развивался очень сильный CASB7439-специфический гуморальный иммунный ответ (фиг. 22). Данные указывают на то, что в 8-10-раз более высокие титры общего IgG против CASB7439 индуцировались при помощи LVL144 по сравнению с LVL168 и LVL111 соответственно у аутбредных мышей CD1. Пример 11. Противоопухолевая эффективность CASB7439 ASCI у мышей: разработка модели опухоли у мышей. Модель опухоли у мыши, экспрессирующей белок CASB7439, с использованием мышиных опухолевых клеток TC-1 или мышиных клеток колоректального рака MC38 разработали для исследованияCASB7439-опосредованной защиты от опухоли у мышей in vivo. Клетки опухоли TC-1 клеток любезно предоставлены Т.С. Wu (Johns Hopkins University, Baltimore, MD). Они получены из первичных легочных клеток мышей C57Bl/6 путем успешного переноса генов HPV16 E6 и E7 и активированного онкогена ras,как описано ранее (Lin et al. (1996), Cancer Res. Jan 1; 56(1):21-6. Клетки MC38, представляющие собой мышиную клеточную линию колоректальной аденокарциономы, приобретали в ATCC (Manassas, VA). Эти клетки стабильно трансфицировали конструкцией, кодирующей CASB7439, с использованием нелипосомального реагента для трансфекции Fugene-6 (Roche, Mississauga, ON, Canada) в соответствии с рекомендациями производителя. Кратко, плазмиду (рсДНК 3.1/Zeocine), кодирующую полноразмерную последовательность белка CASB7439 и содержащую ген резистентности к зеоцину, смешивали с ночи. После трансфекции клетки оставляли для восстановления в течение 24 ч в культуральной среде + 10% инактивированной теплом FBS (фетальная телячья сыворотка). Клетки затем собирали и распределяли в 96-луночных планшетах (50 мкл клеточной суспензии на лунку) для ограниченных разведений. После высевания клеток среду заменяли на культуральную среду + 10% инактивированной теплом FBS,содержащую 100 мкг/мл зеоцина (Invitrogen, Burlington, ON, Canada). После отбора каждый клон собирали и оставляли расти в 24-луночных планшетах и замораживали. Экспрессию мРНК CASB7439 в клеткахTC1/CASB7439 и MC38/CASB7439 контролировали посредством qPCR в реальном времени с использованием CASB7439-специфических праймеров и зонда TaqMan. Реакции qPCR в реальном времени осуществляли с использованием реакционного набора TaqMan с использованием аппарата TaqMan 7900(Applied Biosystem, Foster City, CA, USA). Экспрессию белка CASB7439 оценивали при помощи вестернблоттинга и иммунофлуоресценции с использованием специфического моноклонального антитела против CASB7439 (40-12). Клеточная популяция TC-1/CASB7439 14 не является полностью клональной, но скорее всего состоит по меньшей мере из двух отличающихся субклонов. Таким образом, еще одну стадию ограниченного разведения осуществляли для получения полностью клональной клеточной популяцииTC-1/CASB7439: клетки TC-1/CASB7439 14-2. Для определения оптимальной дозы клеток TC-1 или MC38 для опухолевой сенсибилизации у мышей различные дозы клеток (от 1105 до 2106 клеток на мышь) инъецировали инбредным мышам CB6f1. Оптимальная определенная доза составляет 2,5105 клеток на мышь для сенсибилизацииTC-1/CASB7439 14 и TC1/CASB7439 14-2; обнаружено, что 1105 клеток на мышь является оптимальной для сенсибилизации клетками MC38/CASB7439 35. Противоопухолевая эффективность у мышей: эксперименты по иммунизации и сенсибилизации. Серии из четырех исследований (1.1-1.4) разработаны для оценки CASB7439-опосредованной опухолевой защиты in vivo. В общем, в каждом из этих четырех исследований группам мышей CB6f1 осуществляли четыре внутримышечные иммунизации с двухнедельными интервалами, с различными дозами (0, 1, 10 или 30 мкг) специфического рекомбинантного белка CASB7439, выбранными для исследования, приготовленными с AS01B или AS15 в описанной здесь конечной концентрации, с физиологическим раствором, только белком CASB7439 или только адъювантами. Через две недели после четвертой иммунизации мышей CB6f1 сенсибилизировали путем подкожной инъекции 250000 клетокTC-1/CASB7439 14-2. После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. В соответствии с протоколом мышей с опухолями больше чем 200 мм 2 умерщвляли. Затем строили кривые выживания. Дополнительно осуществляли три исследования (2.1-2.4) для изучения доз, отличающихся от 0, 1,10 или 30 мкг рекомбинантного белка CASB7439. В исследовании 2.4 оценивали корреляцию между иммуногенностью и опухолевой защитой при помощи CASB7439 у самцов и самок мышей C57Bl/6; параллельно оценивали мышей CB6f1. Исследования 1.1-2.4 более подробно описаны ниже. Исследование 1.1 Заявители исследовали противоопухолевую защиту, обеспечиваемую иммунизацией LVL111 (1 или 10 мкг), приготовленным с AS01B или AS15, по сравнению с сенсибилизацией TC1/CASB7439 14 у мышей CB6f1. 10 мышей на группу внутримышечно иммунизировали четыре раза с двухнедельными интервалами рекомбинантным белком CASB7439 (LVL111), приготовленным с AS01B или AS15 в описанной здесь конечной концентрации. Также тестировали две контрольные группы, иммунизированные физиологическим раствором или только белком CASB7439. Через две недели после четвертой иммунизации каждую мышь подкожно сенсибилизировали 250000 клеток TC-1/CASB7439 14. За опухолевым ростом следили в течение 42 суток, но кривые выживания не строили, поскольку слишком мало мышей умерщвляли на 42 сутки. Тем не менее, кривые опухолевого роста строили для каждой группы. На фиг. 23 на верхней и нижней панелях представлены кривые опухолевого роста, построенные соответственно для композиций с AS01B и AS15 (замечание: AS15 неправильно идентифицирован на этом фиг. как "AS015".) Кривые демонстрируют, что у мышей CB6f1, получающих белокCASB7439, приготовленный с AS01B или AS15, рост опухоли TC1/CASB743914 замедляется. Исследование 1.2. Группы из 14 мышей получали отличающуюся дозу LVL111 (1, 10 и 30 мкг) или использовались в качестве контроля. Мышей внутримышечно иммунизировали четыре раза с двухнедельными интервалами различными дозами LVL111, приготовленными с AS01B или AS15 в описанной здесь конечной концентрации. Четыре контрольные группы также иммунизировали только AS01B, только AS15, только физиологическим раствором или только 30 мкг LVL111. Через две недели после четвертой иммунизации каждую мышь подкожно сенсибилизировали 250000 клеток TC-1/CASB7439 14. После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. В соответствии с протоколом мышей,имеющих опухоль больше чем 200 мм 2, умерщвляли. Строили кривую выживания. Результаты указывают на то, что мыши, иммунизированные LVL111, приготовленным с AS15,лучше защищены против опухолевой сенсибилизации TC1/CASB7439 14 по сравнению с мышами, иммунизированными LVL111, приготовленным с AS01B. Оптимальные результаты у мышей CB6f1 получены для 10 мкг LVL111, приготовленного с AS15, см. фиг. 24. Исследование 1.3. 38 мышей CB6f1 на группу внутримышечно иммунизировали четыре раза с двухнедельными интервалами 10 мкг LVL111, LVL168 или LVL144, приготовленных с AS15 в описанной здесь конечной концентрации. Также тестировали контрольную группу, иммунизированную только AS15. Через две недели после четвертой иммунизации 28 мышей подкожно сенсибилизировали 250000 клеток TC-1/CASB7439 14 и 10 оставшихся мышей подкожно сенсибилизировали 100000 клетокMC38/CASB7439 35. После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. В соответствии с протоколом мышей, имеющих опухоль больше чем 200 мм 2, умерщвляли. Результаты указывают на то, что все из LVL111, LVL168 и LVL144, приготовленных с AS15, защищают против сенсибилизации TC1/CASB743914 или MC38/CASB743935, см. фиг. 25. Исследование 1.4. 66 мышей CB6f1 на группу внутримышечно иммунизировали четыре раза с двухнедельными интервалами 10 мкг LVL168 или LVL144, приготовленных с AS15 в описанной здесь конечной концентрации. Также тестировали контрольную группу, иммунизированную только AS15. Через две недели после четвертой иммунизации 22 мыши на группу подкожно сенсибилизировали 250000 клеток TC-1/CASB7439 14 (неклональная популяция), 250000 клеток TC-1/CASB7439 14-2(клональная популяция) или 100000 клеток MC38/CASB7439 35. После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. В соответствии с протоколом мышей, имеющих опухоль больше чем 200 мм 2, умерщвляли. По-видимому, сенсибилизация TC1/CASB7439 14-2 более агрессивна по сравнению с сенсибилизацией TC1/CASB7439 14, о чем свидетельствует более ранняя гибель контрольных мышей и отсутствие видимого защитного действия в этом конкретном исследовании. См. фиг. 26. Последующие эксперименты осуществляли более агрессивным TC-1/CASB7439 14-2, как здесь описано. Исследование 2.1. 15 мышей CB6f1 на группу внутримышечно иммунизировали четыре раза различными эквимолярными дозами LVL168 или LVL144, приготовленных с AS15 в описанной здесь конечной концентрации. Для LVL168 использовали следующие дозы антигена: 0,77, 1,5, 3,1 и 6,25 мкг/мышь. Для LVL144 использовали следующие дозы антигена: 1,25, 2,5, 5 и 10 мкг/мышь. Также тестировали контрольную группу, иммунизированную только AS15. Через две недели после четвертой иммунизации всех мышей подкожно сенсибилизировали 250000 клеток TC-1/CASB7439 14-2 (клональная популяция). После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. Мышей, имеющих опухоль больше чем 289 мм 2 (1717 мм), умерщвляли. Строили кривые выживания для каждой группы. См. фиг. 27, на которой изображены кривые выживания, полученные с эквимолярными дозами LVL168 иLVL144, приготовленных с AS15, после сенсибилизации опухолью TC1/CASB7439 14-2. TF обозначает мышей без опухоли. По сравнению с контрольной группой AS15 четыре набора эквимолярных дозLVL144 и LVL168, по-видимому, ингибируют рост опухоли у мышей CB6f1 в равной степени. Исследование 2.2. 15 мышей C57Bl/6 на группу внутримышечно иммунизировали четыре раза различными эквимолярными дозами LVL168 или LVL144, приготовленных с AS15 в описанной здесь конечной концентрации. Для LVL168 использовали следующие дозы антигена: 0,77, 1,5, 3,1 и 6,25 мкг/мышь. Для LVL144 использовали следующие дозы антигена: 1,25, 2,5, 5 и 10 мкг/мышь. В качестве контрольных групп мышей также иммунизировали только AS15 или буфером. Через две недели после четвертой иммунизации всех мышей подкожно сенсибилизировали 250000 клеток TC-1/CASB7439 14-2 (клональная популяция). После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. Мышей, имеющих опухоль больше чем 289 мм 2 (1717 мм), умерщвляли. Строили кривые выживания для каждой группы, см. фиг. 28, на котором изображены кривые выживания, построенные для различных эквимолярных доз LVL168 и LVL144, приготовленных с AS15, после сенсибилизации опухолью TC1/CASB7439 14-2. TF обозначает мышей без опухоли. В противоположность исследованиям, проведенным для мышей CB6f1, в настоящем исследовании отсутствует статистически значимое различие между выживанием мышей C57Bl/6 в иммунизированных группах, группой AS15 или группой, которой вводили буфер. Исследование 2.3. 15 мышей CB6f1 на группу внутримышечно иммунизировали четыре раза различными эквимолярными дозами LVL168 или LVL144, приготовленных с AS15 в описанной здесь конечной концентрации. Для LVL168 использовали следующие дозы: 0,04, 0,19, 0,77, 1,5, 3,1 и 6,25 мкг/мышь. Для LVL144 использовали следующие дозы: 0,078, 0,31, 1,25, 2,5, 5 и 10 мкг/мышь. Также тестировали контрольную группу, иммунизированную только AS15. В качестве второй контрольной группы также использовали наивных мышей. Через две недели после четвертой иммунизации всех мышей подкожно сенсибилизиро- 27020617 вали 250000 клеток TC-1/CASB743914-2 (клональная популяция). После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. Мышей, имеющих опухоль больше чем 289 мм 2(1717 мм), умерщвляли. Строили кривые выживания для каждой группы, см. фиг. 29 и 30, на которых изображены кривые выживания, построенные для различных эквимолярных доз LVL168 и LVL144, приготовленных с AS15, после сенсибилизации опухоли TC1/CASB7439 14-2. TF обозначает мышей без опухоли. Две наименьшие тестированные эквимолярные дозы, по-видимому, не оказывают значительного ингибирования опухолевого роста. При четырех наибольших дозах обнаружена большая защита от опухоли для мышей, иммунизированных LVL168 + AS15 по сравнению с LVL144 + AS15. Исследование 2.4. Дополнительно исследовали утрату защиты, обнаруженной для мышей C57Bl/6, иммунизированных CASB7439, приготовленным с AS15, против сенсибилизации опухолью TC1/CASB7439 14-2, как обнаружено в исследовании 2.2. 15 мышей CB6f1 или C57Bl/6 на группу внутримышечно иммунизировали четыре раза 1 мкг LVL168, приготовленного с AS15, в описанной здесь конечной концентрации. Группы самцов и самок мышей, иммунизированных 1 мкг LVL168 + AS15, исследовали по отдельности. Также тестировали отдельные контрольные группы самцов и самок, иммунизированных только AS15. Частичные заборы крови осуществляли через 7 суток после четвертой иммунизации и сывороточные титры общего антитела IgG против CASB7439 определяли при помощи ELISA, как описано в примере 10. Аналогичные CASB7439-специфические CD4 Т-клеточные ответы (приблизительно 1106 мкг/мл общего IgG) получены для самцов и самок мышей CB6f1, иммунизированных LVL168 +AS15; никакого ответа в виде антител не обнаружено у мышей C57Bl/6, см. фиг. 33. Частичные заборы крови осуществляли через 14 суток после четвертой иммунизации. CD4 и CD8 Т-клеточные ответы (положительные в отношении как IFN, так и TNF) измеряли путем внутриклеточного окрашивания и при помощи анализа путем проточной цитометрии, как описано в примере 10, после того, как PBL (лимфоциты периферической крови) мышей CB6f1 или C57Bl/6 (5 совокупностей из 3 мышей на группу) повторно стимулировали пулом пептидов, покрывающих полноразмерную последовательность CASB7439. Кроме того, пептид 39 (SEQ ID NO: 91) тестировали с PBL из C57Bl/6. Аналогичные CASB7439-специфические CD4 Т-клеточные ответы (частота приблизительно 0,1%) получали для самцов и самок мышей CB6f1, иммунизированных LVL168 + AS15; CASB7439-специфический CD8 Тклеточный ответ (частота 0,05%) получен только для самцов мышей, см. фиг. 34A и 35A. НикакойCASB7439-специфический CD4 или CD8 Т-клеточный ответ не обнаружен у мышей C57Bl/6, иммунизированных 1 мкг LVL168 + AS15 после повторной стимуляции PBL пулом пептидов, покрывающих полноразмерную последовательность CASB7439, или CASB7439 пептид 39 (SEQ ID NO: 91) (а.к. с 153 по 167 в SEQ ID NO: 13), см. фиг. 34A и 35A. Необходимо учитывать, что CASB7439 пептид 39(SEQ ID NO: 91) использовали независимо, поскольку ранее его идентифицировали как пептид, обладающий низкой CD4 Т-клеточной иммуногенностью, в спленоцитах мышей C57Bl/6, четыре раза иммунизированных 10 мкг LVL168 + AS15, см. фиг. 13 в примере 10. Через две недели после четвертой иммунизации всех мышей подкожно сенсибилизировали 250000 клеток TC-1/CASB7439 14-2. После сенсибилизации размер опухолей измеряли три раза в неделю для каждой мыши. Мышей, имеющих опухоль больше чем 289 мм 2 (1717 мм), умерщвляли. Строили кривые роста опухолей и кривые выживания для каждой группы. На фиг. 31 и 32 изображен соответственно рост опухоли и кривые выживания, полученные для самцов и самок мышей CB6f1 или C57Bl/6, иммунизированных 1 мкг LVL168 или LVL144, приготовленных с AS15, после сенсибилизации опухольюTC1/CASB7439 14-2. Результаты указывают на то, что CASB7439 + AS15 обладают тенденцией вызывать меньший опухолевый рост и более хорошее выживание у самцов мышей CB6f1 по сравнению с самками CB6f1, но не оказывают действие в отношении противоопухолевой защиты у мышей C57Bl/6. Утрата защиты против опухоли CASB7439 у мышей C57Bl/6 соответствует отсутствию обнаруживаемыхCASB7439-специфических CD4 и CD8 Т-клеток, а также отсутствию обнаруживаемого CASB7439 специфического ответа в виде антител у мышей C57Bl/6. Последнее указывает на то, что в используемых экспериментальных условиях мыши C57Bl/6 не представляют собой идеальную модель для исследования иммуногенности и противоопухолевой защиты в отношении CASB7439. С другой стороны, у мышейCB6f1 обнаруженная противоопухолевая защита в отношении CASB7439 соответствует присутствию
МПК / Метки
МПК: A61K 39/00, A61P 35/00, C07K 14/47
Метки: конструкции, casb7439
Код ссылки
<a href="https://eas.patents.su/30-20617-konstrukcii-casb7439.html" rel="bookmark" title="База патентов Евразийского Союза">Конструкции casb7439</a>
Способ обработки железобетонной конструкции со стальной арматурой посредством пропитки конструкции катионами ингибитора

Номер патента: 4161
Опубликовано: 26.02.2004
Автор: Люблинский Ефим Я.
МПК: C23F 13/00
Метки: посредством, катионами, пропитки, ингибитора, способ, конструкции, железобетонной, обработки, стальной, арматурой
Формула / Реферат:
1. Способ обработки железобетонной конструкции со стальной арматурой посредством пропитки конструкции катионами ингибитора, отличающийся тем, что сначала осуществляют измерение коррозионного потенциала стальной арматуры, затем размещают первичный анод смежно с внешней поверхностью железобетонной конструкции, затем подают на первичный анод водный раствор ингибитора для железобетона и осуществляют насыщение конструкции раствором ингибитора, после...
Устройство для формирования арматурной конструкции, способ сборки устройства для формирования арматурной, конструкции и способ изготовления арматурной конструкции

Номер патента: 10843
Опубликовано: 30.12.2008
Автор: Келли Стивен Эдвард
МПК: E04C 5/06, B21F 27/08, B23K 37/04...
Метки: устройство, конструкции, изготовления, способ, сборки, устройства, формирования, арматурной
Формула / Реферат:
1. Устройство для формирования арматурной конструкции, которое содержит несколько держателей (1, 7, 8) для крепления на каждом из них по меньшей мере одного фиксатора (3) для удержания по меньшей мере одного арматурного стержня (6), причем каждый держатель (1, 7, 8) имеет по меньшей мере одно отверстие или углубление (2) для установки фиксатора (3) и по меньшей мере одно отверстие или углубление (2, 4) для присоединения к другому держателю (1,...
Слитые конструкции и конъюгаты лекарственного средства

Номер патента: 18471
Опубликовано: 30.08.2013
Авторы: Холт Люси Дж., Хамилтон Брюс, Жеспер Лоран, Херринг Кристофер
МПК: C07K 16/46, A61K 47/48, C07K 19/00...
Метки: средства, слитые, лекарственного, конструкции, конъюгаты
Формула / Реферат:
1. Композиция слитой конструкции или конъюгата, содержащая или состоящая из следующего: (а) лекарственное средство, выбранное из эксендинов, PYY (пептида YY), GLP (глюкагоноподобного пептида), ОХМ (окситомодулина), VIP (вазоактивного интестинального пептида) или РР (панкреатического полипептида), присутствующее в виде слитой конструкции или конъюгата с (б) однодоменным антителом (dAb) DOM 7h-14, которое связывает сывороточный альбумин и которое...
Конструкции нуклеиновых кислот

Номер патента: 10056
Опубликовано: 30.06.2008
Автор: Фуллер Джеймс
МПК: A61J 1/22, A61K 31/7088, A61K 9/14...
Метки: кислот, конструкции, нуклеиновых
Формула / Реферат:
1. Конструкция нуклеиновой кислоты, включающая в себя: (i) последовательность химерного промотора, которая содержит: (a) последовательность предраннего промотора hCMV, (b) экзон 1 и по меньшей мере часть экзона 2 основного предраннего гена hCMV и (c) гетерологичный интрон, расположенный на месте участка интрона A в основном предраннем гене hCMV; (ii) кодирующую последовательность в функциональной связи с химерным промотором; (iii)...
Конструкции нуклеиновых кислот

Номер патента: 11557
Опубликовано: 28.04.2009
Автор: Фуллер Джеймс
МПК: A61K 39/245, A61K 39/145, A61K 39/39...
Метки: нуклеиновых, кислот, конструкции
Формула / Реферат:
1. Конструкция нуклеиновой кислоты, содержащая последовательность промотора и кодирующую последовательность, функционально связанную с промотором, где кодирующая последовательность кодирует антиген вируса гриппа, его иммуногенный фрагмент или иммуногенный вариант любого из них, обладающий по меньшей мере 80% аминокислотной гомологией с указанным антигеном или фрагментом; и/или ADP-рибозилирующую субъединицу бактериального токсина, ее фрагмент с...
Предыдущий патент: Кондиционирующая композиция для волос
Следующий патент: Способ производства фосфорной кислоты
Случайный патент: Устройство для порционной выдачи жидкого моющего средства