Активаторы плазминогена, имеющие сниженную способность связывать лизин







Формула / Реферат
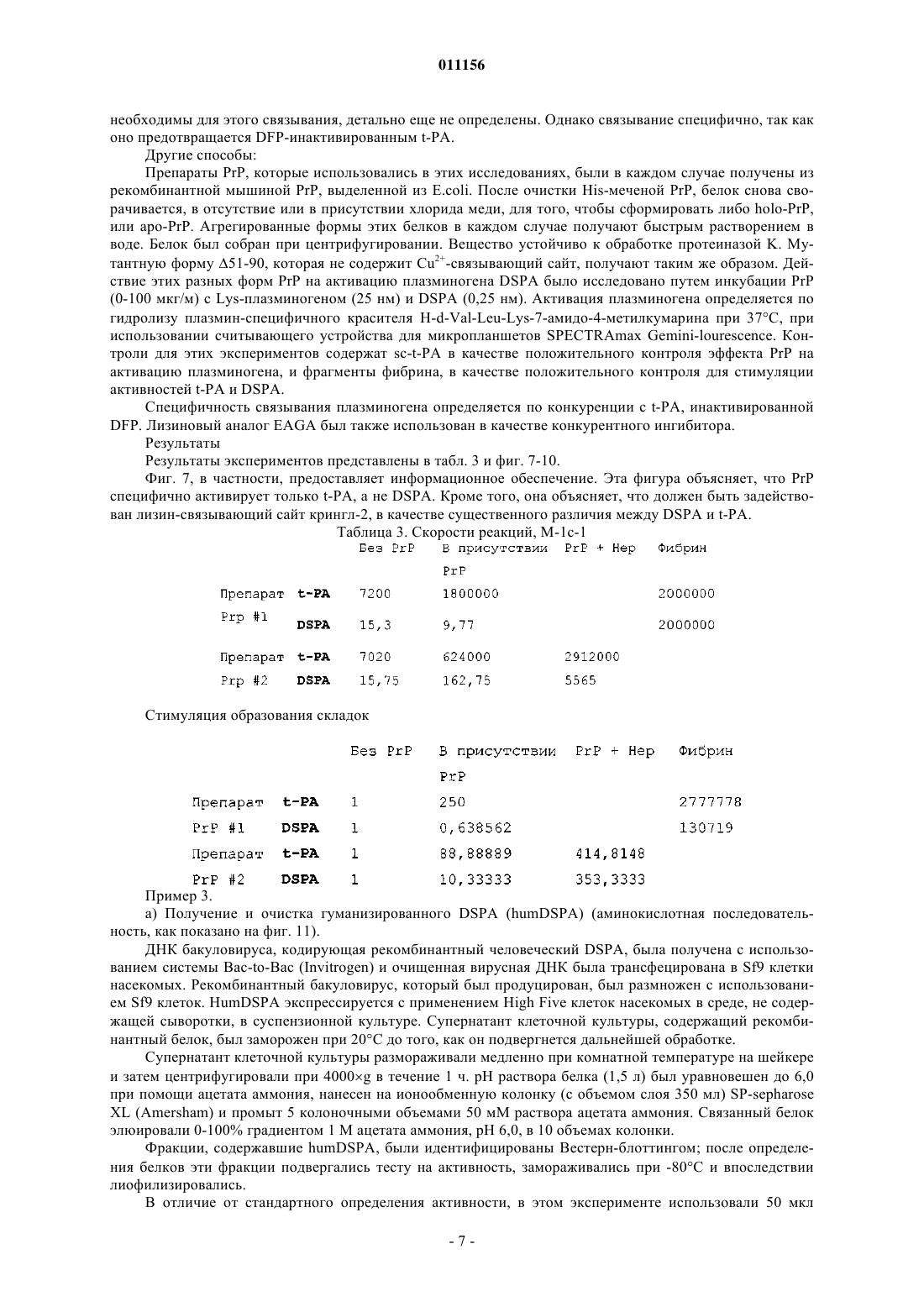
1. Активатор плазминогена, характеризующийся идентичностью по крайней мере на 70% с последовательностями, приведенными на одной из фиг. 3, 11 и 13.
2. Активатор плазминогена по п.1, отличающийся тем, что в области перехода между крингл-доменом и цистеиновым мостиком активатор плазминогена имеет сегмент аминокислотной последовательности, содержащий SKAT.
3. Активатор плазминогена по п.1 или 2, отличающийся тем, что сайт активации активатора плазминогена имеет последовательность LHST.
4. Активатор плазминогена по любому из пп.1-3, отличающийся замещением D236N или аминокислот, которые гомологичны ему.
5. Применение активатора плазминогена по любому из пп.1-4 для лечения тромботических заболеваний, в частности инсульта.
6. Активатор плазминогена по любому из пп.1-5, характеризующийся тромболитической активностью, которая является повышенной в сравнении с активностью нативного t-PA.
Текст
011156 Изобретение относится к преимущественному применению активаторов плазминогена, и для него испрашивается приоритет в соответствии с патентной заявкой Германии 103 42 518 7, содержание которой включено в настоящий документ в качестве ссылки. Назначение тромболизиса как варианта терапии при различных тромбоэмболических заболеваниях считается уже в значительной степени завершенным. Основной интерес в исследовании тромболизиса направлен поэтому на дальнейшее развитие и модификацию известных тромболитиков и/или улучшение сопутствующих методов терапии. Важным тромболитиком, служащим в качестве отправного пункта для разработки новых тромболитиков, является тканевой активатор плазминогена (t-PA), который обладает более высокой селективностью в отношении фибрина и более высокой активностью, чем стрептокиназа и урокиназа. Делеционные мутанты ретеплаза и ланотеплаза, а также тенектеплаза, являются дальнейшими разработками этого активатора плазминогена. Тканевой активатор плазминогена (t-PA) является одноцепочечным гликопротеином, состоящим из 527 аминокислот. Молекула, исходно представляющая собой одиночную цепь (sct-PA), протеолитически расщепляется на двухцепочечную форму (tct-PA). t-PA обладает определенными структурными и функциональными доменами. Так, N-концевая составляющая цепь содержит фингер-домен (F, аминокислотыSer1-Lys49), домен эпидермального фактора роста (Е, Ser50-Asp87) и два крингл-домена (K1, Thr88Gly176, и K2, Asn177-Cys261). С-концевая цепь, которая включает домен сериновой протеазы (Р), содержит аминокислоты с Ser262 по Pro527. Посредством аминокислотных остатков His322, Asp371 иSer478 С-концевая цепь формирует активный центр.t-PA и его рекомбинантные варианты и их получение, являются объектом, например, патента США 4766075 и большого числа публикаций (например, Bode and Renatus: Tissue-type plasminogen activator:variants and crystal/solution structures demarcate structural determinants of function, Current Opinion in Structural Biology 1997, 7:865-872). Диаграмма структуры t-PA прилагается на фиг. 1. Подобно большинству других сериновых протеаз sct-PA превращается в tct-PA форму при расщеплении. Расщепление происходит по связи между Arg275 и Ile276. После этого две цепи удерживаются вместе только посредством одного дисульфидного мостика между серином 264 и серином 395. Такая нумерация сайта расщепления t-PA соответствует нумерации, выбранной Bode и Renatus (loc.cit). Однако другие авторы берут за основу отличную от этой нумерацию и определяют сайт расщепления или активации как R15-I16 или R310-I311. Однако сайты, определенные таким образом, не отличаются функционально или структурно. Тканевой активатор плазминогена (t-PA, алтеплаза) способен активировать плазминоген, который переходит в плазмин. Однако из кинетических констант очевидно, что t-PA активирует циркулирующий плазминоген только в слабой степени. Обе формы t-PA, то есть и одно-, и двухцепочечная молекулы,демонстрируют в принципе одни и те же фармакологические свойства. Однако плазминоген, связанный с фибрином, активируется с каталитической активностью, на три порядка превосходящей активность, с которой активируется свободный плазминоген. Соответственно, тромболитические свойства тканевого активатора плазминогена значительно усиливаются в присутствии фибрина. Сообщается, что относительная селективность t-PA по отношению к фибрину находится в интервале от 500 до 1000. Каталитический эффект может также быть увеличен при взаимодействии активатора плазминогена с бета-амилоидом или фибриногеном. Способность t-PA активироваться фибриногеном сравнима с его способностью активироваться бета-амилоидом. Эффект активатора плазминогена контролируется физиологически ингибиторами, причем ингибитор I активатора плазминогена (PAI-1) является важным антагонистом. Связывание PAI-1 с легкой цепью, входящей в состав молекулы t-PA, изменяет структуру каталитического центра таким образом, что реакция активации плазминогена не может далее осуществляться (Bennet WF, Paoni NF, Keyt BA, Botstein D, Jones AJS, Presta L, Wurm FM, Zoller M: High resolution analysis of functional determinants in humantissue-type plasminogen activator. Journal of Biological Chemistry 1991; 266: 5191-5201). Активатор тканевого плазминогена быстро метаболизируется в печени. Поэтому наличие печеночной недостаточности у пациента увеличивает период полужизни этого вещества в плазме крови (EmeisDNA technology Circulation 1985; 71: 110-116). Активаторы плазминогена разработаны для лечения тромботических заболеваний, таких как инфаркт миокарда и инсульт. t-PA является в настоящее время единственным тромболитиком, одобреннымFood and Drug Administration (FDA) в США для лечения инсульта. Тем не менее, в прошлом возросли подозрения, что, хотя t-PA, с одной стороны, проявляет ожидаемые положительные тромболитические эффекты при лечении инсульта, с другой стороны он является ответственным за нежелательное повреждение тканей. Таким образом, инфузия t-PA t-PA-дефицитным мышам приводила к возрастанию числа инфарктов (Wang Y.F., Tsirka S.E., Strickland S, Stieg PE, Sorianowild-type and tPA-deficient mice. Nat Med 1998; 4 (2): 228-231). Предполагается, что это возросшее число повреждений обусловлено стимуляцией NMDA-зависимых глутаматных рецепторов (Liberatore GT, Samson A, Bladin С, Schleuning WD, Medcalf RL: Vampire bat salivary plasminogen activator (desmoteplase): aunique fibrinolitic enzyme that does not promote neurodegeneration. Stroke 2003; 34 (2) : 537-543). Этот эффект может быть вызван протеолитическим расщеплением NMDA-рецепторов под действием t-PAplasminogen activator enhances NMDA receptor-mediated signaling. Nat Med 2001; 7 (1): 59-64). Следовательно, цель настоящего изобретения состоит в разработке новых методов терапии тромботических болезней, в частности инсульта. Эта цель достигается посредством применения плазминоген-активирующего фактора, который проявляет способность связывать лизин, сниженную по сравнению со связывающей способностью нативного активатора плазминогена. В особенно преимущественном варианте осуществления активатор плазминогена демонстрирует модифицированный крингл-домен, предпочтительно крингл-2 домен, который изменен по сравнению с доменом нативного t-PA. Последний домен может полностью или частично отсутствовать, так, что способность связывать лизин снижена.K2 домен или области, которые функционально или структурно, по существу, гомологичны ему,могут быть предпочтительно модифицированы таким образом, что остатки лизина более не связываются или связываются только с низкой аффинностью. В особенно преимущественном варианте осуществления, активатор плазминогена, который может использоваться в соответствии с изобретением, представляет собой модифицированный t-PA. Значение K2-домена, включая получение K2-делеционных мутантов t-PA, детально описано в Horrevoets AJ, Smilde A, de Vries С, Pannekoek H (The specific roles of finger and kringle 2 domains of tissuetype plasminogen activator during in vitro fibrinolysis: Journal of Biological Chemistry, 269, 17, 12639-12644,1994). Поскольку их способность связывать лизин снижена или отсутствует, плазминоген-активирующие факторы, которые могут использоваться в соответствии с изобретением, проявляют повышенную селективность по отношению к фибрину и сниженную способность активироваться фибриногеном или бетаамилоидом. Эта сниженная способность активироваться фибриногеном является, возможно, причиной селективности по отношению к фибрину. Наличие у активатора плазминогена высокой селективности по отношению к фибрину имеет важное значение в связи с лечением инсульта, в частности, поскольку, например, нативный t-PA может быть активирован фибриногеном, который преодолевает поврежденный гематоэнцефалический барьер и затем стимулирует опосредованную глутаматом эксцитотоксичность,посредством последующей активации NMDA рецепторов. Соответственно, снижение в соответствии с изобретением способности активатора плазминогена активироваться фибриногеном также ведет к снижению нейротоксичности. Важность лизин-связывающих участков на t-PA крингл-2 выяснилась, в частности, в результате сравнительных исследований структурных и функциональных доменов t-PA в сравнении с DSPA. DSPA представляет собой активатор плазминогена, который был первоначально выделен из слюнной железы летучей мыши-вампира (в этой связи см. патент США 6008019; ЕР 0 383 417). DSPA может быть выделен в виде четырех изоформ, из которых DSPAalpha1 можно получить в рекомбинантном виде, используя клетки СНО. В отличие от t-PA, DSPA содержит только один крингл-домен. Функционально и структурно этот домен соответствует скорее K1 домену t-PA, чем K2 домену, и не имеет участков связывания лизина(Bringmann P, Gruber D, Liese A, Toschi L, Kratzschmar J, Schleuning WD, Dormer P: Structural features mediating fibrin selectivity of vampire bat plasminogen activators; Journal of Biological Chemistry 1995, 270(43): 25596-25603). Поэтому в литературе встречаются утверждения, что DSPA не содержит крингл-2 домена. Кроме того, DSPA всегда представлен одноцепочечными молекулами, потому что в нем отсутсвует сайт активации плазмина, такой как в t-PA. В сравнении с t-PA активность DSPA стимулируется в присутствии фибрина в 45000 раз, в то время как, согласно Gardell SJ, Duong LT, Diehl, York JD, Hare TR,Register RB, Jacobs JW, Dixon RA, Friedman PA (Isolation, characterization and cDNA cloning of a vampirebat salivary plasminogen activator: Journal of Biological Chemistry 198 9; 264(30): 17947-17952) это значение составляет 205. Диаграмма структуры DSPA прилагается в виде фиг. 2a. Фиг. 2b показывает сравнение аминокислотных последовательностей t-PA и DSPA (SEQ ID Nos. 1+2). Прежде считалось, что способность t-PA связывать фибрин функционально обусловлена фингер- и крингл-2 доменами (van Zonnenfeld AJ, Veerman H, Pannekoek H: (1986) Proc. Natl. Acad. Sci. USA., 83: 4670-4674). Однако в недавних публикациях предполагается определенное участие и протеазного домена Р (Bennett loc. cit.). Функции индивидуальных доменов исследовались также Bakker AHF, Jacoline Е.,Weening-Verhoeffet D., Verheijen JH (The role of Lysyl-Binding site of tissue type plasminogen activator in theBakker с соавторами получили различные модификации t-PA, например два делеционных мутанта,которые лишены либо крингл-2, фингер-домена, или домена эпидермального фактора роста. Эти мута-2 011156 ции были получены также в сочетании, и некоторые из них были дополнены точечной мутацией в крингл-2 домене, конкретно включающей замену D236N. Эта избирательная аминокислотная замена заключается в замещении аспарагиновой кислоты в положении 236 аспарагином и, таким образом, уничтожении участка связывания лизина (LBS в K2 домене). Для процедуры получения мутации читателю настоятельно рекомендуют обратиться к упомянутой выше публикации Bakker et al., включающей цитируемые в ней ссылки. В этих исследованиях Bakker с соавторами продемонстрировали, что занятие лизин-связывающего участка в K2 домене ЕАСА (-аминокапроновой кислотой) заметно ослабляет связывание нативного t-PA с фибрином. То же самое относится к модификации LBS заменой D236N, за тем исключением, что ослабление меньше. Даже делеционный мутант, который состоял из одного K2 домена и протеолитического С-концевого фрагмента, все еще связывался с фибрином, но в небольшой степени. И только K2 Р мутант, в котором был делетирован LBS, не проявлял более способности связывать фибрин. Хотя эти результаты демонстрируют, с одной стороны, важность F и K2 доменов, а также LBS в K2,они также демонстрируют, что взаимодействие t-PA и фибрина не опосредуется исключительно F и K2 доменами. Есть еще функция у LBS в домене K2, причем LBS отвечает предположительно за стабилизацию конформации t-PA, наиболее благоприятную для связывания фибрина. Важность K2 домена, включая его лизин-связывающий участок, выяснилась в исследованиях, проведенных Stewart RJ, Fredenburgh JG и Witz JI (Characterization of the interactions of plasminogen and tissuelinked to fragment E: Journal of Biological Chemistry 273, 29, 18292-18299, 1998). В экспериментах по связыванию Stewart et al. изучают, среди прочего, сродство t-PA и DSPA к фибрину и фибриногену. В исследовании сродство изучалось в присутствии или в отсутствие аналога лизина ЕАСЕ для того, чтобы проанализировать значение крингл-зависимого взаимодействия. На основании исследований они заключили, что DSPA не способен связывать фибриноген из-за отсутствия связывающих лизин сайтов в крингл-домене. Они поэтому приписывают функцию, существенную для способности t-PA связывать фибрин, LBS в крингл-2-домене t-PA в сочетании с фингер-доменом. Результаты экспериментов, представленные в примере 1, подтверждают разное сродство DSPAalpha1 и рекомбинантного t-PA человека к кофакторам бета-амилоиду (1-42), фибриногену и фибрину. Для получения этих данных, кинетические параметры Kкат и Km, и отношение Kкат/Km, были определены для каждой комбинации активатор плазминогена/кофактор. В отсутствие кофактора способностьDSPAalpha1 к активации плазминогена была приблизительно в 100 раз ниже, чем способность к активации плазминогена t-PA, в то время как оба соединения одинаково способны активировать плазминоген в присутствии фибрина. DSPAalpha1 был примерно в 30 раз менее эффективным, чем t-PA в присутствии кофактора фибриногена или бета-амилоида. При физиологической концентрации плазминогена 2 мкМ,из этих данных выводится отношение эффективности фибрина как кофактора к эффективности фибриногена или бета-амилоида как кофактора, 480 в случае DSPAalpha1 и 16 в случае t-PA человека. Как было уже разъяснено выше, плазминоген-активирующий фактор, который используется в соответствии с изобретением, характеризуется тем фактом, что его связывающий лизин сайт крингл-2-домена или отсутствует, или является измененным. В одном варианте осуществления крингл-2 удален. Однако также возможно, в другом варианте осуществления сохранить крингл-2 и заменить аспарагиновую кислоту в 236 позиции на аспарагин. Соответственно, для изобретения не критично, в соответствии с изобретением, отсутствует ли структура крингл-домена t-PA, но это имеет место только для лизинсвязывающего сайта, подлежащего модификации таким образом, что взаимодействие, повышающее активность t-PA, с кофактором более невозможно. В другом предпочтительном варианте осуществления активатор плазминогена, в частности, измененный t-PA, который используется, может быть модифицирован посредством делеции крингл-1-домена,таким образом, что t-PA не способен более связывать рецептор. Как результат, активатор тканевого плазминогена более не метаболизируется естественным образом в печени, что приводит к удлинению периода полужизни in vivo (Rijken DC, Otter M, Kuiper J, von Berkel TJC: Receptor-mediated endocytosis oftissue-type plasminogen activator (t-PA) by liver cells. Thromb. Res. 1990; Supel. X: 63-71). Делеция крингл-1 домена из ретеплазы известна и описана, например, Martin U, Bader R, Bhm E, Kohnert U, von MllendorfE, Fischer S, Sponder G (BM 06.022: A Novel recombinant plasminogen activator. Cardiovascular drug reviews 1993; 11:299-311). Для этого применялись мутанты, в которых аминокислоты с 4 по 176 удалены. Увеличение периода полужизни делает возможным болюсное введение активатора плазминогена. В предпочтительном варианте осуществления активатор плазминогена, который может быть использован в соответствии с изобретением, основан на одной из аминокилотных последовательностей,показанных на фиг. 3, 11 или 13. Каждая из этих аминокислотных последовательностей составляет модифицированную t-PA, в которой удален крингл-2 и сделано структурное изменение на участке последовательности сайта расщепления t-PA при активации. Это изменение может либо состоять только в замещении аминокислот R и I в сайте расщепления при активации (например, HS; см. последовательностьSEQ ID3, фиг. 3) или дополнительно воздействовать на близлежащие аминокислоты (например, замещение FRIK на LHST в последовательности SEQ ID4, фиг. 11). Последняя модификация в этом-3 011156 месте соответствует структуре нативного DSPA. В соответствии с изобретением предпочтение отдается получению и использованию для лечения инсульта активатора плазминогена, который особенно подходящим образом сочетает преимущество нативного t-PA, а именно, в частности, низкую иммуногенность при использовании у человека, с преимуществами DSPA, а именно отсутствием нейротоксичности. В одном варианте осуществления изобретения удаление крингл 2-домена из t-PA соответственно выбирается так, что переносимый участок соотносится с соответствующей структурой в DSPA. Переносимый участок между оставшимся крингл 1 доменом измененной t-PA и нижележащий цистеиновый мостик, соответственно, формируются преимущественно последовательностью SKAT. В DSPA последовательность SKAT расположена между кринглдоменом и цистеиновым мостиком. Эта преимущественная структура реализуется, например, в последовательности SEQ ID4, как показано на фиг. 11. Фиг. 12 (множественное выравнивание последовательностей) показывает сравнение между t-PA и двумя описанными вариантами осуществления в соответствии с изобретением. В еще одном варианте осуществления в соответствии с изобретением используется активатор плазминогена как изображено на фиг. 13 (SEQ ID5). Естественно, также возможно, в соответствии с изобретением, использовать белки, обладающие последовательностями, которые гомологичны или частично идентичны аминокислотным последовательностям, изображенным на фиг. 3, 11 и 13. Предпочтение отдается гомологам или последовательностям,идентичным по крайней мере на 70%, предпочтительно на 80-95%. Эти гомологичные или идентичные белки демонстрируют активность плазминоген-активирующего фактора (предпочтительно проявляющуюся в освобождении pNA) и вызывают лизис кровяного сгустка in vitro (см. пример 3). Активатор плазминогена, который может быть использован в соответствии с изобретением, может характеризоваться отсутствием фингер-домена и домена эпидермального фактора роста. Эта делеция существенно снижает связывание с рецепторами печени и снова удлиняет период полужизни (Larson GR,Timony GA, Horgan PG, Barone KM, Henson KS, Angus LB, Stoudemire JB: Protein engineering of novelclinical implications of drug design Am J Cardiol. 1996; 78 (suppl. 12): 2-7). Помимо делеционных мутантов, активаторы плазминогена, которые используются в соответствии с изобретением, могут также быть точечно изменены посредством сайт-направленного мутагенеза. Так,известно, например, для rt-PA-TNK (тенектеплазы), что три мутации в сайте связывания ингибитора активатора плазминогена, с одной стороны, и в сайте связывания, посредством которого t-PA связывается с клетками печени, с другой стороны, приводит к возрастанию каталитической активности, в то время как способность инактивироваться PAI снижается. Это, таким образом, обеспечивает тенектеплазе константу каталитической конверсии Kкат/Km, которая повышена в 100 раз (в этой связи см. также Paoni NF, Keyttrial. Circulation 1995; 92 (suppl.): 1-415). Пример 1. 1. Сравнительный анализ эффективности бета-амилоида (от 1 до 42), фибриногена и фибрина как кофакторов активации плазминогена DSPAalpha1 и рекомбинантного человеческого t-PA. Материалы и способы: Были использованы следующие вещества:DSPAalphal (приготовленный Paion, серия No. 2DSA01, 12 декабря 2002, образец 10). 10 мг вещества были растворены в 10 мл стерильной воды, для получения конечной концентрации 1 мг/мл; рекомбинантный t-PA (Actilyse, приготовленный Paion, серия No. 102572). 10 мг были растворены в 10 мл стерильной воды для получения конечной концентрации 1 мг/мл;Glu-плазминоген человека, очищенный из плазмы крови человека (приготовленной Paion) и растворенный в буфере PCLA в концентрации 25 мкМ; фибриноген человека, по существу, свободный от плазминогена, был получен от Sigma (каталожный No. F4883, серия 12K7620). 25 мг были растворены в 25 мл буфера PCLA для получения конечной концентрации 1 мг/мл; тромбин человека (Sigma; каталожный No. T7009. Серия 61K7603). 100 единиц были растворены в 10 мл буфера PCLA для получения конечной концентрации 10 единиц /мл.Flavigen.pli цветной реагент (D-but-CHT-Lys-p-nitroaniline-DHCL) от Biolool (каталожный No. 101353, серия 1512016). 100 мкМ были растворены в 50 мл буфера PLCR для получения 2 миллимолярной конечной концентрации; белок бета-Амилоид (1-42) от Bachem (каталожный No. Н-1368, серия 0535120). 4 мг были растворены в 4 мл 0,1-процентного гидроксида аммония для получения конечной концентрации 1 мг/мл.-4 011156 Все реагенты были разделены на порции и хранились при -20 С в течение не более двух недель.No. 207780); 1 мМ Na2HPO4 2H2O (Fluka, каталожный No. 71638); 0,3 мМ MgCl2 6H2O (Acros, каталожный No. 197530); 0,4 мМ MgSO4 7H2O (Fluka, каталожный No. 63140); 20 мМ HEPES (Applichem, каталожный No. A1969); 0,01% Polysorbate 80 (Fluka, каталожный No. 93781). Исследование активации плазминогена Исследование активации плазминогена проводилось на микропланшетах для титрования в суммарном объеме 0,15 мл, как описано у Bringmann et al., (loc. cit.). Реагенты добавлялись в следующем порядке: 50 мкл плазминогена (0-24 мкМ); 15 мкл кофактора (1 мг/мл); 10 мкл активатора плазминогена (7,5 нМ) и 75 мкл Flavigen (2 мМ). Конечные концентрации были таковы: активатор плазминогена (t-PA или DSPA1): 0,5 нМ; Gluплазминоген: 0,0625, 0,125, 0,25, 0,5, 1, 2, 4 и 8 мкМ; FlavigenPli: 1 мМ и кофакторы: 100 мкг/мл. Кофакторы бета-амилоид (1-42), фибриноген и фибрин сравнивались с контролем без кофакторов. В контроле реакционная смесь содержала добавочные 0,13 единиц/мл тромбина человека. Сразу после заполнения планшет для титрования помещался при 25 С в molecular device ThermoMax microtiter reader. Перемешивание производилось, начиная с t=0. Оптическая плотность при 405 нм и 490 нм измерялась с регулярными интервалами. Оптические плотности при 490 нм вычитались из оптических плотностей при 405 нм для того, чтобы исключить различия, обусловленные током жидкости. Все эксперименты выполнялись в трех повторах для каждой концентрации плазминогена и активатора плазминогена. Определялись значения Km и ккат (фиг. 4-6). Фиг. 4a) определение молярной экстинкции pNA;b) конверсия Flavigen.pli 10 нМ плазмином как функция концентрации Flavigen.pli (в мкМ);c) график Михаэлиса-Ментен для конверсии Flavigen 10 нМ плазмином. Начальные скорости из фиг. 4b были переведены в pNA/c. Фиг. 5 Кривые изменения оптической плотности во времени для четырех различных комбинаций активаторов плазминогена и кофакторов. Для этой цели были использованы концентрации плазминогена: 8; 4; 2; 1; 0,5; 0,25; 0,125 и 0,0625 мкМ.h) DSPAalpha1 с фибрином как кофактором. Фиг. 6. График значений =ks. ккат.РА.Е.Р./(Km+Р) в зависимости от концентрации плазминогенаa) графики для t-PA без кофактора или с бета-амилоидом, фибриногеном или фибрином как кофактором; Табл. 1: значения ккат/Km для активации плазминогена рекомбинантными t-PA и DSPAalpha1 человека с использованием бета-амилоида (1-42), фибриногена и фибрина как кофакторов. Результаты даны как среднеестандартное отклонение для трех независимых экспериментов. Определить точные значения ккат и Km было невозможно, потому что значения Km были выше максимальной концентрации (8 мкМ) плазминогена, которую можно было использовать. Вывод кинетических констант сам по себе недостаточен для сравнения эффективностей различных кофакторов, поскольку скорости реакции при концентрации плазминогена, близкой или меньшей чемKm, зависят от концентрации плазминогена. По этой причине скорости реакции вычисляются с использованием кинетики Михаэлиса-Ментен при характерной для плазмы крови физиологической концентрации плазминогена - 2 мкМ. Относительные эффективности различных кофакторов в активации плазминогена со стороны t-PA и DSPAalpha1 (табл. 2) определены на этой основе. Таблица 2 Относительные скорости активации плазминогена при его физиологической концентрации - 2 мкМ,вычисленные с использованием кинетических параметров табл. 1. Сочетание DSPA контроля составляет 1. Пример 2. Прионные болезни, например губчатые энцефалопатии, представляют собой фатальные нейродегенеративные явления, которые характеризуются накоплением неправильной изоформы прионного белка(PrPsc) в амилоидных отложениях. Нормальный прионный белок (PrPc) экспрессируется во многих тканях, особенно, в мозге, где он концентрируется в синапсах. Механизм перехода конформации PrPc в PrPsc до сих пор недостаточно понятен, и непонятен механизм, который ведет к нейродегенерации посредством образования PrPsc. Возможный механизм мог бы заключаться в действии на PrPsc в связи с активацией плазминогена t-PA. Авторы уже показали, используя рекомбинантный белок, что PrP может повышать катализируемуюt-PA активацию плазминогена в 300 раз (Ellis V, Daniels M, Misra R, Brown DR (2000): Plasminogen activation is stimulated by prion protein and regulated in a copper-dependent manner, Biochemistry 41: 6891-6896). Способность PrP стимулировать активацию плазминогена зависит от конформации PrP, причем на конформацию влияет наличие или отсутствие Cu2+. Последняя форма содержит большое количество бета-структур, которые соответствуют изоформе PrPsc и стимулируют активацию плазминогена. Эти данные согласуются с более ранними наблюдениями в том, что, когда плазминоген, связанный с PrPsc, который выделен из мозга мышей, инфицированных scrapie, он не связывал PrPc, как в нормальных клетках мозга (Fisher M, Roeckl С, Rarizek P, Schwarz HP, Aguzzi A (2000): Binding of disease-associated prion protein to plasminogen. Nature 408: 479-483). Более того, эксперименты авторов показали, что механизм активации плазминогена, стимулированной PrP, также заключает в себе связывание t-PA с apoPrP (то есть PrP без Cu2+). Структуры, которые-6 011156 необходимы для этого связывания, детально еще не определены. Однако связывание специфично, так как оно предотвращается DFP-инактивированным t-PA. Другие способы: Препараты PrP, которые использовались в этих исследованиях, были в каждом случае получены из рекомбинантной мышиной PrP, выделенной из E.coli. После очистки His-меченой PrP, белок снова сворачивается, в отсутствие или в присутствии хлорида меди, для того, чтобы сформировать либо holo-PrP,или аро-PrP. Агрегированные формы этих белков в каждом случае получают быстрым растворением в воде. Белок был собран при центрифугировании. Вещество устойчиво к обработке протеиназой K. Мутантную форму 51-90, которая не содержит Cu2+-связывающий сайт, получают таким же образом. Действие этих разных форм PrP на активацию плазминогена DSPA было исследовано путем инкубации PrP(0-100 мкг/м) с Lys-плазминогеном (25 нм) и DSPA (0,25 нм). Активация плазминогена определяется по гидролизу плазмин-специфичного красителя Н-d-Val-Leu-Lys-7-амидо-4-метилкумарина при 37 С, при использовании считывающего устройства для микропланшетов SPECTRAmax Gemini-lourescence. Контроли для этих экспериментов содержат sc-t-PA в качестве положительного контроля эффекта PrP на активацию плазминогена, и фрагменты фибрина, в качестве положительного контроля для стимуляции активностей t-PA и DSPA. Специфичность связывания плазминогена определяется по конкуренции с t-PA, инактивированнойDFP. Лизиновый аналог EAGA был также использован в качестве конкурентного ингибитора. Результаты Результаты экспериментов представлены в табл. 3 и фиг. 7-10. Фиг. 7, в частности, предоставляет информационное обеспечение. Эта фигура объясняет, что PrP специфично активирует только t-PA, а не DSPA. Кроме того, она объясняет, что должен быть задействован лизин-связывающий сайт крингл-2, в качестве существенного различия между DSPA и t-PA. Таблица 3. Скорости реакций, М-1 с-1 Пример 3. а) Получение и очистка гуманизированного DSPA (humDSPA) (аминокислотная последовательность, как показано на фиг. 11). ДНК бакуловируса, кодирующая рекомбинантный человеческий DSPA, была получена с использованием системы Bac-to-Bac (Invitrogen) и очищенная вирусная ДНК была трансфецирована в Sf9 клетки насекомых. Рекомбинантный бакуловирус, который был продуцирован, был размножен с использованием Sf9 клеток. HumDSPA экспрессируется с применением High Five клеток насекомых в среде, не содержащей сыворотки, в суспензионной культуре. Супернатант клеточной культуры, содержащий рекомбинантный белок, был заморожен при 20 С до того, как он подвергнется дальнейшей обработке. Супернатант клеточной культуры размораживали медленно при комнатной температуре на шейкере и затем центрифугировали при 4000g в течение 1 ч. рН раствора белка (1,5 л) был уравновешен до 6,0 при помощи ацетата аммония, нанесен на ионообменную колонку (с объемом слоя 350 мл) SP-sepharoseXL (Amersham) и промыт 5 колоночными объемами 50 мМ раствора ацетата аммония. Связанный белок элюировали 0-100% градиентом 1 М ацетата аммония, рН 6,0, в 10 объемах колонки. Фракции, содержавшие humDSPA, были идентифицированы Вестерн-блоттингом; после определения белков эти фракции подвергались тесту на активность, замораживались при -80 С и впоследствии лиофилизировались. В отличие от стандартного определения активности, в этом эксперименте использовали 50 мкл-7 011156 элюата (=humDSPA-содержащая фракция), 50 мкл 0,2 М Tris, рН 8,0, 100 мкл 2 мМ S-2288 в PBS. б) Исследование активности DSPA. Применяемое исследование анализа активности представляет собой определение скорости, с которой активатор плазминогена превращает бесцветный субстрат S-2288 в окрашенный продукт. Это стандартный способ определения протеолитической активности активирующих плазминоген факторов. Активность визуализируется посредством определения высвобождения хромогена (п-нитроанилина, pNA) тестируемым веществом. Компания Chromogenix предлагает для этой цели хромоген S-2288. В таком исследовании скорость реакции не определяется непосредственно (например, в каталитических единицах); вместо этого делается сравнение со стандартом, который принимается за 100% активность. Скорость реакции измерялась в буфере, имеющем следующий состав: 25 мМ Tris-HCl, рН 8/0,1% альбумин/100 мМ NaCl /1,1 мМ глицин/1,2 мМ маннит/2,5 мМ S-2288. Измерение проводилось в 96-луночном планшете с использованием контролей, стандартов и образцов, содержавших различные концентрации белка. Изменение поглощения при 405 нм во времени регистрировалось с использованием фотометра. Наклон линейного участка кривой определялся с помощью Excel. Этот параметр давал значения активности. Фиг. 14 показывает активности фракций (выраженные через активность S-2288). Гели, окрашенные серебром, можно видеть на фиг. 15 а и 15b. Фиг. 16 а и 16b демонстрируют иммуноблоты. Белок, выявленный на иммуноблоте, соответствует белку, отмеченному стрелкой на геле, окрашенном серебром.[L = образец, нанесенный на колонку; Т = проскок (материал, не задержанный колонкой); W фракция, полученная промыванием колонки; 0-40% В, А 2, A3 фракции. Пример 4. Лизис сгустка (тромболитическая активность). Остаток содержащей humDSPA фракции В 6 был растворен в 15 мл PBS и раствор был центрифугирован при 4000g в течение 15 мин. Эта фракция была выбрана потому, что она демонстрировала наибольшую чистоту. Из нее была отобрана порция для определения активности. Каталитическая активность этой фракции была приблизительно сравнима с каталитической активностью 5 мкг/мл активазы. Кровяные сгустки, которые были использованы, были получены из образцов нормальной крови,взятых за 24 ч, и в каждом случае формировались из 2 мл крови, которая была перенесена в полипропиленовые пробирки и коагулировала в естественных условиях. Лизис проводился в PBS. Содержавшая humDSPA фракция В 6 (связанная SPXL-сефарозой) была прежде всего лиофилизирована. Остаток был растворен в 15 мл PBS и центрифугирован при 4000 g в течение 15 мин. Образец этого раствора был взят для определения активности с S-2288 до того, как был добавлен сгусток. Каталитическая активность этого образца в большой степени соответствовала каталитической активности 5 мкг/мл активазы. Фиг. 17 а и 17d показывают результаты хронологического хода лизиса сгустка в 0, 3, 4 и 24 ч после добавления humDSPA. Слева в каждом случае показан контрольный эксперимент. Эксперимент с PBS,содержавшим humDSPA в указанном выше количестве, представлен справа. Клетки крови медленно оседают из распадающегося сгустка. Сеть, которую можно различить уже через 4 ч, сохраняется до 24 ч. В исследовании отделившиеся клетки крови удалялись через 4 ч при помощи шприца для того, чтобы оценить структуру оставшегося сгустка. Следует уделить внимание тому факту, что фибринолиз замедляется, когда сгусток больше не подвешен в своем собственном лизате. По этой причине любой сгусток все еще сохраняется через 24 ч. Количественно оценить эту реакцию невозможно. Однако humDSPA проявляет явное повышение активности в сравнении с нативным DSPA, так, что по крайней мере в 4 раза большее количество DSPA должно быть использовано для того, чтобы растворить сгусток такого же размера. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Активатор плазминогена, характеризующийся идентичностью по крайней мере на 70% с последовательностями, приведенными на одной из фиг. 3, 11 и 13. 2. Активатор плазминогена по п.1, отличающийся тем, что в области перехода между кринглдоменом и цистеиновым мостиком активатор плазминогена имеет сегмент аминокислотной последовательности, содержащий SKAT. 3. Активатор плазминогена по п.1 или 2, отличающийся тем, что сайт активации активатора плазминогена имеет последовательность LHST. 4. Активатор плазминогена по любому из пп.1-3, отличающийся замещением D236N или аминокислот, которые гомологичны ему. 5. Применение активатора плазминогена по любому из пп.1-4 для лечения тромботических заболеваний, в частности инсульта. 6. Активатор плазминогена по любому из пп.1-5, характеризующийся тромболитической активностью, которая является повышенной в сравнении с активностью нативного t-PA.
МПК / Метки
МПК: C12N 9/72, A61K 38/49
Метки: активаторы, плазминогена, имеющие, лизин, способность, связывать, сниженную
Код ссылки
<a href="https://eas.patents.su/30-11156-aktivatory-plazminogena-imeyushhie-snizhennuyu-sposobnost-svyazyvat-lizin.html" rel="bookmark" title="База патентов Евразийского Союза">Активаторы плазминогена, имеющие сниженную способность связывать лизин</a>
Способ ингибирования ингибитора 1 активатора плазминогена.

Номер патента: 1003
Опубликовано: 28.08.2000
Авторы: Берг Дэвид Т., Гриннелл Брайан В., Ричардсон Марк А.
МПК: A61K 31/4025
Метки: плазминогена, активатора, ингибирования, способ, ингибитора
Формула / Реферат:
1. Способ ингибирования ингибитора 1 активатора плазминогена, включающий назначение нуждающемуся в этом человеку эффективного количества соединения где R1 и R3 представляют независимо друг от друга водород, -СН3, , или где Аr - необязательно замещенный фенил; R2 выбирают из группы, состоящей из пирролидино, гексаметиленимино и пиперидино; и его фармацевтически приемлемой соли и сольвата. 2. Способ по п.1, отличающийся тем, что...
Фармацевтическая композиция, имеющая сниженную тенденцию кристаллизации лекарственного ингредиента

Номер патента: 6383
Опубликовано: 29.12.2005
Авторы: Морозович Уолтер, Далга Роберт Дж., Хейджман Майкл Дж., Цао Пин, Форбес Джеймс С., Хассан Фред, Стефански Кевин Дж., Карим Азиз, Хуан Техуа
МПК: A61K 47/38, A61P 29/00
Метки: ингредиента, кристаллизации, сниженную, композиция, тенденцию, фармацевтическая, имеющая, лекарственного
Формула / Реферат:
1. Перорально доставляемая фармацевтическая композиция, включающая (а) лекарственный ингредиент с низкой растворимостью в воде; (б) фармацевтически приемлемую растворяющую жидкость и (в) полимер, понижающий помутнение; где, по крайней мере, существенная часть лекарственного ингредиента находится в растворенной или солюбилизированной форме и где полимер присутствует в количестве, достаточном для того, чтобы в значительной степени подавить...
Замещенные нафтилиндольные производные в качестве ингибиторов ингибитора активатора плазминогена типа 1 (pai-1)

Номер патента: 9696
Опубликовано: 28.02.2008
Авторы: Гандерсен Эрик Гаулд, Элокдах Хассан Махмуд, Крэндэлл Дэвид Лерой, Мэйер Скотт Кристиан
МПК: C07D 209/12, A61K 31/40, A61P 7/02...
Метки: замещенные, активатора, ингибиторов, типа, плазминогена, ингибитора, нафтилиндольные, производные, качестве, pai-1
Формула / Реферат:
1. Соединение формулы I где R1, R2, каждый независимо, представляют собой водород, алкил, содержащий 1-3 атома углерода; R3 и R4, каждый независимо, представляют собой водород, алкил, содержащий 1-3 атома углерода, или галоген; R5 представляет собой водород, алкил, содержащий 1-6 атомов углерода; R6 представляет собой водород, алкил, содержащий 1-6 атомов углерода, алканоил, содержащий 1-6 атомов углерода, или бензил, необязательно замещенный...
Применение цитокина, способного связывать il-18bp и ингибировать активность второго цитокина

Номер патента: 8667
Опубликовано: 29.06.2007
Авторы: Ким Соо Хиун, Динарелло Чарльз А., Буфлер Филип
МПК: A61K 38/20, A61P 29/00
Метки: il-18bp, цитокина, способного, активность, связывать, второго, применение, ингибировать
Формула / Реферат:
1. Применение цитокина-1 или его изоформы, мутеина, слитого белка, функционального производного или фрагмента, способного связываться с IL-18BP или его мутеином, слитым белком, функциональным производным или фрагментом и способного ингибировать рецептор цитокина-2, причем цитокин-2 является членом семейства IL-1, для производства лекарственного средства для лечения или профилактики заболевания, которое вызывается или утяжеляется индуцированием...
Активаторы рецепторов, активируемых пролифератором пероксисом (ppar )

Номер патента: 8928
Опубликовано: 31.08.2007
Авторы: Хэйуард Черилл Майерс, Даггер Роберт Уэйн, Лию Жэнгью, Бэгли Скотт Уильям, Брандт Томас Эндрю, Хэйда Уильям Эндрю
МПК: A61K 31/4545, A61P 3/00, C07D 211/22...
Метки: рецепторов, активируемых, активаторы, пролифератором, пероксисом, ppar
Формула / Реферат:
1. Соединение формулы I его изомер, пролекарство указанного соединения или изомера либо фармацевтически приемлемая соль указанного соединения, изомера или пролекарства; где каждый F и G независимо представляет собой а) водород, б) галогено, в) (С1-С4)алкил, г) (С3-С6)циклоалкил, д) гидрокси или е) (С1-С4)алкокси; X представляет собой a) -Z или б) -B-C(R1)(R2)-Z; В представляет собой а) окси, б) метилен или в) -N(H)-; Z представляет собой а)...
Предыдущий патент: Соединения фенилпиридилпиперазина, способ их получения и фармацевтические композиции, содержащие их
Следующий патент: Способ получения кристаллического циклесонида, обладающего определенным размером частиц
Случайный патент: Биологически активная добавка