Электропорация микобактерии и сверхэкспрессия антигенов микобактерий
Номер патента: 12182
Опубликовано: 28.08.2009
Авторы: Сунь Ронггай, Хоун Дэвид Майкл, Сэдофф Джералд С.






















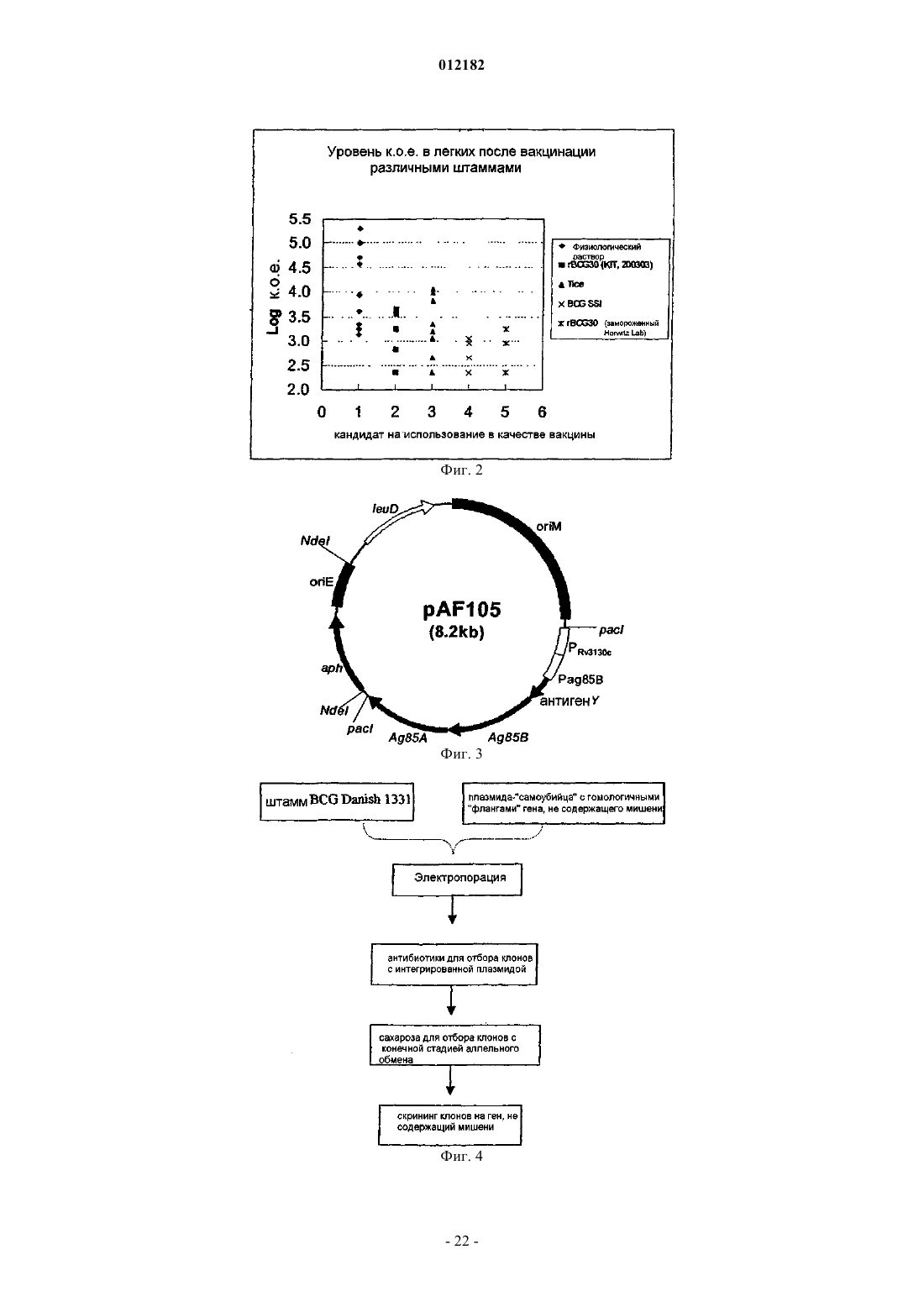
Формула / Реферат
1. Трансформированная бактерия или ее потомство, которые включают нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная нуклеотидная последовательность присутствует в плазмиде, и где указанная нуклеотидная последовательность кодирует белок, необходимый для выживания, и где ген, кодирующий указанный необходимый для выживания белок, делетирован из бактериального генома указанной трансформированной бактерии.
2. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде, и где указанная плазмида содержит ген, кодирующий высвобождение из эндосомы, и где указанный ген, кодирующий высвобождение из эндосомы, может представлять собой чужеродную нуклеотидную последовательность, которая присутствует в указанной плазмиде или может присутствовать в указанной плазмиде в дополнение к чужеродной нуклеотидной последовательности, присутствующей в указанной плазмиде.
3. Трансформированная бактерия или ее потомство по п.2, где указанный ген, кодирующий высвобождение из эндосомы, представляет собой ген перфринголизина О (pfo) из Clostridium perfringens.
4. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде и где указанная чужеродная нуклеотидная последовательность кодирует антиген 85A, антиген 85B или антиген 85A/85B.
5. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде и где указанная плазмида содержит ген, кодирующий белки, которые сохраняют и/или стабилизируют плазмиду.
6. Трансформированная бактерия или ее потомство по п.5, где указанный ген, кодирующий белки, кодирует антиген 85A, антиген 85B или антиген 85A/85B.
7. Трансформированная бактерия или ее потомство по п.2, где указанной бактерией является Mycobacterium.
8. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, где указанная чужеродная нуклеотидная последовательность кодирует апоптоз.
9. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде, и где указанная плазмида содержит ген, кодирующий апоптоз.
10. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером и где указанная чужеродная нуклеотидная последовательность не может реплицироваться в грамотрицательных бактериях.
11. Трансформированная бактерия или ее потомство по п.1, где указанной трансформированной бактерией является ауксотрофная бактерия.
12. Трансформированная бактерия или ее потомство по п.1, где указанная чужеродная нуклеотидная последовательность является по меньшей мере частью однонаправленного челночного вектора.
13. Способ трансформирования бактерии, включающий стадию введения нуклеотидной последовательности, которая реплицируется и экспрессируется в указанной бактерии, где указанная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная нуклеотидная последовательность присутствует в плазмиде, где указанная нуклеотидная последовательность кодирует белок, необходимый для выживания, и где ген, кодирующий указанный необходимый для выживания белок, делетирован из бактериального генома указанной бактерии.
14. Способ по п.13, где указанную стадию введения осуществляют путем электропорации.
15. Способ по п.13, где указанная нуклеотидная последовательность является по меньшей мере частью однонаправленного челночного вектора.
16. Трансформированная Mycobacterium или ее потомство, содержащие чужеродную нуклеотидную последовательность, которая кодирует представляющий интерес белок, и где указанная трансформированная Mycobacterium включает плазмиду, которая не способна реплицироваться в грамотрицательных бактериях.
17. Трансформированная Mycobacterium или ее потомство по п.16, где указанная чужеродная нуклеотидная последовательность является частью плазмиды.
18. Трансформированная Mycobacterium или ее потомство по п.17, где указанная плазмида не содержит селектируемого маркера.
19. Трансформированная Mycobacterium или ее потомство по п.17, где указанный белок является необходимым для выживания и соответствует белку выживания, который кодируется нуклеотидной последовательностью, делетированной из бактериального генома указанной трансформированной микобактерии.
20. Трансформированная Mycobacterium или ее потомство по п.19, где указанным белком является leuD.
21. Трансформированная Mycobacterium или ее потомство по п.19, дополнительно содержащие промоторные последовательности, которые активируются in vivo.
22. Трансформированная Mycobacterium или ее потомство по п.16, где указанная трансформированная Mycobacterium является аттенюированной.
23. Трансформированная Mycobacterium или ее потомство по п.22, где указанной трансформированной Mycobacterium является BCG.
24. Трансформированная Mycobacterium или ее потомство по п.23, где указанный BCG выбран из нижеследующих штаммов: BCG1331, BCG Пастер, BCG Токио и BCG Копенгаген.
25. Вакцина, включающая трансформированную Mycobacterium или ее потомство, содержащие чужеродную нуклеотидную последовательность, которая кодирует представляющий интерес ген, и где указанная трансформированная Mycobacterium включает плазмиду, которая не способна реплицироваться в грамотрицательных бактериях.
26. Трансформированная Mycobacterium по п.16, где указанная трансформированная Mycobacterium не обладает резистентностью к антибиотикам.
27. Трансформированная Mycobacterium по п.16, где указанная трансформированная Mycobacterium является ауксотрофной.
28. Вакцина по п.25, где указанная трансформированная Mycobacterium не обладает резистентностью к антибиотикам.
29. Вакцина по п.25, где указанная трансформированная Mycobacterium является ауксотрофной.
30. Способ трансформирования бактерии, включающий стадию введения чужеродной нуклеотидной последовательности, которая реплицируется и экспрессируется в указанной бактерии, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером и где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде, где указанная чужеродная нуклеотидная последовательность не может реплицироваться в грамотрицательных бактериях.
Текст
012182 Предшествующий уровень техники Область, к которой относится изобретение Настоящее изобретение относится к штаммам Mycobacterium, обладающим улучшенными вакцинными свойствами, позволяющими использовать их в качестве вакцин против туберкулеза. ШтаммыMycobacterium предпочтительно выбирают из родительских штаммов, которые были идентифицированы как штаммы, обладающие высокой иммуногенностью и не обладающие резистентностью к антибиотикам, и гены которых не подвергаются горизонтальному переносу в грамотрицательные бактерии. Настоящее изобретение также относится к Mycobacterium, обладающим улучшенными свойствами, обеспечивающими доставку трансгенов, которые обладают вакцинными свойствами, позволяющими использовать их для вакцинации против других заболеваний и для лечения рака. Предшествующий уровень техники Бактерией Mycobacterium tuberculosis (M.tb) инфицирована одна треть населения всего мира, и эти инфекции вызывают активную форму заболевания у 8 млн чел., из которых каждый год умирает 1,6-2,2 млн чел., причем большинство из них проживает в развивающихся странах. Туберкулез (ТБ) представляет собой эпидемическое заболевание глобального масштаба, причем заболеваемость туберкулезом и смертность от этого заболевания все возрастает, поскольку его распространенность пересекается с распространенностью ВИЧ-инфекции. ТБ является главной причиной смертности больных СПИДом.BCG, т.е. широко используемая в настоящее время вакцина против ТБ, была разработана 80 лет назад и, как показали испытания, уровень ее эффективности против туберкулеза легких широко варьируется, причем эта вакцина оказалась неэффективной в большинстве испытаний в условиях практической работы, проведенных в последнее время в Индии (Fine et al., Vaccine, 16(20): 1923-1928, 1998; Anonymous, Indian J. Med. Res., Aug; 110: 56-69, 1999). Тем не менее, в настоящее время Всемирная Организация Здравоохранения рекомендует BCG-вакцинацию всех детей при рождении или при первом посещении патронажной службы (за исключением детей с симптомами ВИЧ-инфекции/СПИДа) в странах с высоким риском заболеваемости туберкулезом. Такая тактика основана на полученных данных, свидетельствующих о том, что BCG обеспечивает защиту от тяжелых форм детского ТБ (Lanckriet et al., Int. J. Epidemiol, 24(5): 1042-1049, 1995; Rodrigues et al., J. Epidemiol. Community Health 45(1): 78-80, 1991). Иммунизация противотуберкулезной вакциной BCG детей далеко не раннего возраста является предметом обсуждения, поскольку имеется лишь ограниченное количество данных, дающих противоречивые результаты. Однако высокая заболеваемость туберкулезом у детей и взрослых в развивающихся странах,где широко практикуется BCG-иммунизация детей при рождении, указывает на то, что применяемая в настоящее время вакцина BCG не очень эффективна, и уже в течение многих лет люди в этих странах подвержены высокому риску заболевания ТБ. Поэтому очевидно, что BCG не обеспечивает адекватную защиту человека от заболевания ТБ и не является средством борьбы против заражения ТБ. Приблизительно 70% людей, контактирующих с микроорганизмами, вызывающими ТБ, и имеющих нормальную иммунную систему, не подвергаются инфицированию, и лишь у 5% из всех инфицированных индивидуумов развивается заболевание в первые два года. У большинства инфицированных индивидуумов наблюдается подавление инфекции, ассоциированное с продуцированием стойкого клеточного иммунного ответа против антигенов М.tb. А у еще 5% индивидуумов наблюдаются рецидивы этого заболевания при снижении иммунитета. Как первичное заболевание, так и рецидивы наблюдаются, главным образом, у людей, страдающих ВИЧ-инфекциями/СПИДом, что еще раз подтверждает важную роль, которую играет иммунитет в предупреждении и устранении инфекции. Описание сущности изобретения Поскольку организм большинства людей способен бороться с ТБ, то есть все основания надеяться,что индуцирование стойкого иммунитета определенного типа позволит разработать эффективные вакцины, которые способны предупреждать возникновение первичной инфекции после контакта с микроорганизмами, вызывающими эту инфекцию, и раннее прогрессирование заболевания, а также предупреждать реактивацию при латентном состоянии и возникновение рецидивов после лечения. И, наконец, может быть использована комбинация системного введения вакцины и проведения химиотерапии, которые, в конечном счете, будут способствовать уничтожению микроорганизма M.tb, являющегося патогеном для человека. Исходя из того факта, что вакцинация BCG в детстве играет решающую роль для предупреждения развития острой формы ТБ, очевидно, что замена BCG на новую вакцину против ТБ в испытаниях, проводимых для оценки кандидатов на вакцину против ТБ, не может быть осуществлена без получения огромного количества данных, свидетельствующих о том, что такая новая вакцина является в высокой степени эффективным продуктом. Однако проблема заключается в том, что M.tb является, главным образом, возбудителем инфекций у человека, и животные-модели лишь частично имитируют взаимодействие хозяина с патогеном. Таким образом, окончательные данные, подтверждающие повышенную эффективность новой вакцины ТБ, могут быть получены только в контролируемых испытаниях в условиях практической работы с участием человека. Исходя из этих соображений, многие исследователи пришли к выводу, что ключевой стадией в создании улучшенной вакцины против ТБ является получение улучшенных штаммов BCG и моделей-животных, несмотря на их ограниченность, и высказали предположение-1 012182 что рекомбинантные штаммы, сверхэкспрессирующие защитные антигены, будут обладать более высокой эффективностью по сравнению с родительским BCG. Некоторые антигены M.tb обладают вакцинными свойствами и при их введении животным в виде вакцинных препаратов обеспечивают защиту, аналогичную защите, достигаемой с использованием лишь одной вакцины BCG (Anderson, Infect. Immun. 62(6): 2536-2544, 1994). Для улучшения свойств этих вакцин-кандидатов может быть применена стратегия усиления иммуногенности антигенов, присутствующих в BCG. Таким образом, были получены штаммы BCG, которые сверхэкспрессируют выбранные антигены M.tb, и было показано, что эти рекомбинантные штаммы BCG (rBCG) индуцируют более сильный иммунитет по сравнению с родительскими штаммами BCG, из которых был получен штамм rBCG(Horwitz et al., Infect. Immun. 72(4): 1672-1679, 2003). В одном исследовании было подтверждено, что штамм rBCG, который экспрессирует антиген 85 В (обозначаемый далее "Ag85 В"), является более эффективным, чем штамм BCG, смешанный с тем же самым антигеном (Horwitz et al., см. выше, 2003). Эти данные дают основание предположить, что такой метод является в высшей степени перспективным. В некоторых случаях штаммы BCG, которые сверхэкспрессируют антигены, могут быть использованы для вырабатывания надежных и эффективных иммунных ответов, обеспечивающих защиту против ТБ-инфекций. Настоящее изобретение относится к генетически сконструированным (рекомбинантным) штаммамMycobacterium, обладающим улучшенными вакцинными свойствами, которые позволяют использовать их для вакцинации против туберкулеза. Эти штаммы обладают различными отличительными свойствами, каждое из которых способствует повышению их иммуногенности. Рекомбинантные штаммы Mycobacterium согласно изобретению получают из родительских штаммов, специально отобранных по их высокой иммуногенности. Другими словами, штамм Mycobacterium,который был выбран в качестве родительского штамма для проведения генетических манипуляций (например, для сверхэкспрессии туберкулезного антигена), был способен, даже до проведения генетических манипуляций, вырабатывать сильный иммунный ответ у вакцинированного хозяина. Таким штаммом является, например, штамм BCG Дания 1331. Затем эти штаммы предпочтительно модифицируют, например, так, чтобы они сверхэкспрессировали представляющий интерес туберкулезный антиген. При получении генетически рекомбинантной микобактерии предпочтительно используют промотор, который активируется in vivo. Кроме того, рекомбинантные штаммы Mycobacterium согласно изобретению были генетически модифицированы так, чтобы для их отбора не требовалось введения генов резистентности к антибиотикам, либо эти штаммы были сконструированы так, чтобы вообще не требовалось использования селектируемых маркеров, что позволяет, по существу, обеспечивать их безопасность для вакцинации человека. Так, например, ген, необходимый для репликации микобактерий, удаляют и помещают в экспрессионную плазмиду. Кроме того, гены рекомбинантных штаммов Mycobacterium согласно изобретению не подвергаются горизонтальному переносу в грамотрицательные бактерии, а поэтому они не способны "ускользать" из организма хозяина. Это также гарантирует их безопасность в качестве вакцинных агентов при введении человеку. В качестве другого примера в настоящем изобретении описана экспрессия антигена 85B в плазмидах, применяемого в качестве фактора, стабилизирующего эту плазмиду, что позволяет избежать необходимости использования отбора по резистентности к антибиотикам для сохранения этих плазмид. Прямая трансформация штаммов микробактерий высокими концентрациями минимальных плазмид, экспрессирующих Ag85 В и другие антигены, с использованием ПЦР-позитивного отбора для их идентификации позволяет получить штаммы микобактерий, сверхэкспрессирующие антигены, обладающие стабильностью в данной плазмиде, без применения антибиотиков или ауксотрофного отбора. Кроме того,рекомбинантные штаммы Mycobacterium согласно изобретению представляют собой превосходные средства, которые могут быть использованы при получении вакцин против туберкулеза, при этом они могут быть также генетически сконструированы так, чтобы они экспрессировали или сверхэкспрессировали антигены, не являющиеся возбудителями туберкулеза, а поэтому они могут быть также использованы в качестве вакцинных агентов против других заболеваний. Более того, rBCG, сверхэкспрессирующие антигены ТБ или антигены, играющие важную роль в патогенезе других заболеваний, могут быть использованы в схемах комбинированной иммунизации вместе с рекомбинантными белками, адъювантами, рекомбинантными вирусными векторами либо с ДНК- или РНК-вакцинами, используемыми для бустериммунизации. Настоящее изобретение относится к трансформированной бактерии или к ее потомству, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в трансформированной бактерии (или в ее потомстве), где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером. В одном из вариантов изобретения чужеродная нуклеотидная последовательность присутствует в плазмиде, а в некоторых вариантах изобретения плазмида кодирует ген, необходимый для выживания бактерии, где указанный ген, необходимый для выживания бактерии, делетирован из бактериального генома трансформированной бактерии. В еще одном варианте изобретения плазмида содержит ген, кодирующий высвобождение из эндосомы, например pfo. В других вариантах изобретения чужеродная нуклеотидная последовательность кодирует высвобождение из эндо-2 012182 сомы например ген pfo. В других вариантах настоящего изобретения чужеродная нуклеотидная последовательность кодирует антиген 85A, антиген 85B или антиген 85A/85B. В других вариантах изобретения плазмида содержит ген, кодирующий белки, которые сохраняют и/или стабилизируют плазмиду. В некоторых вариантах изобретения ген, кодирующий белки, кодирует антиген 85A, антиген 85B или антиген 85A/85B. В одном из вариантов изобретения указанной бактерией является Mycobacterium. В другом варианте изобретения чужеродная нуклеотидная последовательность кодирует апоптоз. В других вариантах изобретения плазмида содержит ген, кодирующий апоптоз. В еще одном варианте изобретения чужеродная нуклеотидная последовательность не может реплицироваться в грамотрицательных бактериях. В некоторых вариантах изобретения трансформированной бактерией является ауксотрофная бактерия. В других вариантах изобретения чужеродной нуклеотидной последовательностью является по меньшей мере часть однонаправленного челночного вектора. Настоящее изобретение также относится к способу трансформации бактерии. Способ включает стадию введения чужеродной нуклеотидной последовательности, которая реплицируется и экспрессируется в бактерии, и чужеродной нуклеотидной последовательности, не связанной с селектируемым маркером. В одном из вариантов изобретения стадию введения осуществляют путем электропорации. В другом варианте изобретения чужеродная нуклеотидная последовательность присутствует в плазмиде, а электропорацию осуществляют в условиях, при которых отношение плазмидной ДНК к бактериальным клеткам составляет от 1 до 5 мкг плазмидной ДНК к 1,25108 бактериальных клеток. В одном из вариантов изобретения это отношение составляет приблизительно 1,6 мкг плазмиды к 1,25108 бактериальных клеток. В некоторых вариантах изобретения чужеродная нуклеотидная последовательность не может реплицироваться в грамотрицательных бактериях. В других вариантах изобретения чужеродной нуклеотидной последовательностью является по меньшей мере часть однонаправленного челночного вектора. В других вариантах изобретения чужеродная нуклеотидная последовательность присутствует в плазмиде и кодирует ген, необходимый для выживания бактерии и делетированный из бактериального генома бактерии. Настоящее изобретение также относится к трансформированной Mycobacterium или ее потомству,содержащим чужеродную нуклеотидную последовательность, кодирующую представляющий интерес ген, в том случае, если выполняются одно или несколько из следующих условий: а) трансформированная Mycobacterium включает плазмиду, которая не способна реплицироваться в грамотрицательных бактериях;b) трансформированная Mycobacterium не обладает резистентностью к антибиотикам; с) трансформированная Mycobacterium является ауксотрофной иd) трансформированная Mycobacterium содержит однонаправленный челночный вектор. В одном из вариантов изобретения чужеродной нуклеотидной последовательностью является часть плазмиды. В другом варианте изобретения плазмида не содержит селектируемого маркера. В еще одном варианте изобретения чужеродная нуклеотидная последовательность кодирует ген, необходимый для выживания бактерии, где указанный ген, необходимый для выживания бактерии, делетирован из бактериального генома трансформированной микобактерии. В некоторых вариантах изобретения геномом,необходимым для выживания, является leuD. Трансформированная Mycobacterium или ее потомство могут дополнительно содержать промоторные последовательности, которые активируются in vivo. Трансформированная Mycobacterium или ее потомство могут быть аттенюированными. ТрансформированнаяMycobacterium или ее потомство могут представлять собой штамм BCG, которым может быть, например,BCG1331, BCG Пастер, BCG Токио или BCG Копенгаген. Настоящее изобретение также относится к вакцине, включающей трансформированную Mycobacterium или ее потомство, содержащие чужеродную нуклеотидную последовательность, кодирующую представляющий интерес ген, в том случае, если выполняются одно или несколько из следующих условий: а) трансформированная Mycobacterium включает плазмиду, которая не способна реплицироваться в грамотрицательных бактериях;b) трансформированная Mycobacterium не обладает резистентностью к антибиотикам; с) трансформированная Mycobacterium является ауксотрофной иd) трансформированная Mycobacterium содержит однонаправленный челночный вектор. Описание графического материала Фиг. 1. Карта вектора-"самоубийцы" pAF103. Каждый из сегментов ДНК имеет следующие обозначения:aph означает ген аминогликозид-фосфотрансферазы (GenBank accessionХ 06402), который сообщает плазмиде резистентность к канамицину;oriE означает ориджин репликации pUC (GenBank accessionAY234331);Ble означает ген (GenBank accessionL36850), который сообщает плазмиде резистентность к зеоцину;SacB означает ген (GenBank accessionY489048), кодирующий левансахаразу, которая сообщает бактерии восприимчивость к сахарозе;Phsp60 означает промоторную последовательность гена белка теплового шока (т.е. Rv0440);MCS означает сайты множественного клонирования для указанных ферментов рестрикции. При этом следует отметить, что кластер, расположенный между двумя Pad-сайтами, может быть заменен, если это необходимо, другими генами эндосомалитических ферментов. Фиг. 2. Уровень иммунной защиты, измеряемый по числу КОЕ в легких после иммунизации различными современными вакцинными штаммами. Фиг. 3. Схематическое представление экспрессионного вектора, не содержащего гена резистентности к антибиотику и используемого для введения в рекомбинантную Mycobacterium, т.е. rBCG. Ген, экспрессируемый в rBCG, клонируют в плазмиду посредством pacI-сайта. Перед электропорацией в rBCG плазмиду расщепляют указанными ферментами рестрикции для удаления областей oriE иKan, в результате чего получают однонаправленный челночный экспрессионный вектор. Каждый из сегментов ДНК имеет следующие обозначения:PRv3130 означает промоторную последовательность антигена Rv3130c;PAg85B означает промоторную последовательность антигена 85 В (т.е. Rv1886c); антиген Y представляет собой микобактериальный антиген ТВ 10.4 (т.е. Rv0288);Ag85B представляет собой последовательность ДНК, кодирующую антиген 85 В (т.е. Rv1886c);aph означает ген аминогликозид-фосфотрансферазы (GenBank accessionХ 06402), который сообщает плазмиде резистентность к канамицину;oriE означает ориджин репликации pUC (GenBank accessionAY234331);oriM представляет собой ориджин репликации в Mycobacterium (GenBank accessionМ 23557). Фиг. 4. Блок-схема основных стадий аллельного обмена. Фиг. 5. ПЦР-анализ отобранных колоний на присутствие экспрессионной плазмиды. ПЦР проводили, как описано в разделе "Материалы и методы". ПЦР-продукты анализировали с помощью гель-электрофореза в 1,0% агарозном геле. Дорожка 1: ДНК-леддер (Invitrogen) использовали как 1 т.п.н. плюс ДНК-стандарт. Дорожка 2: в качестве негативного контроля использовали ПЦР-матрицу. Дорожка 3: ПЦР для штамма BCG Дания 1331. Дорожки 4-7: ПЦР для колоний, обозначенных 59, 61, 69 и 84 соответственно. Дорожка 8: контрольная лунка. Дорожка 9: ПЦР для исходной плазмиды. Подробное описание изобретения Варианты осуществления настоящего изобретения Настоящее изобретение относится к генетически сконструированным (рекомбинантным) штаммамMycobacterium, обладающим улучшенными вакцинными свойствами, которые позволяют использовать их для вакцинации против туберкулеза. Эти штаммы обладают рядом свойств, каждое из которых способствует повышению иммуногенности этих штаммов. Рекомбинантные штаммы Mycobacterium согласно изобретению были получены из родительских штаммов, специально отобранных по их высокой иммуногенности. Другими словами, штамм Mycobacterium, отобранный в качестве родительского штамма для проведения генетических манипуляций (например, для осуществления сверхэкспрессии туберкулезного антигена), был выбран, потому что этот штамм, даже до проведения генетических манипуляций,был способен вырабатывать сильный иммунный ответ у вакцинированного хозяина. Таким штаммом является, например, штамм BCG Дания 1331. Затем эти штаммы предпочтительно модифицируют, например, для сверхэкспрессии представляющего интерес туберкулезного антигена. В такой генетически рекомбинантной микобактерии предпочтительно используют промотор, который активируется in vivo. Кроме того, рекомбинантные штаммы Mycobacterium согласно изобретению являются генетически модифицированными, так что для их отбора не требуется использования генов резистентности к антибиотикам, что, тем самым, обеспечивает их безопасность при вакцинации человека. Так, например, ген, необходимый для репликации микобактерий, удаляют и помещают в экспрессионную плазмиду. Кроме того, рекомбинантные штаммы Mycobacterium согласно изобретению не подвергаются горизонтальному переносу в грамотрицательные бактерии, а поэтому они не способны "ускользать" из организма хозяина(т.е. они представляют собой "однонаправленные челночные векторы"). Это также гарантирует их безопасность в качестве вакцинных агентов при введении человеку. Кроме того, рекомбинантные штаммыMycobacterium согласно изобретению представляют собой превосходные средства, которые могут быть использованы в вакцинах против туберкулеза, и при этом они могут быть также генетически сконструированы так, чтобы они экспрессировали или сверхэкспрессировали антигены, не имеющие отношения к туберкулезу, а поэтому они могут быть также использованы в качестве вакцинных агентов против других заболеваний. В предпочтительных вариантах изобретения штаммами Mycobacterium являются аттенюированные штаммы, например BCG. Однако, как очевидно для специалистов в данной области, могут быть также использованы и другие аттенюировнные и неаттенюированные штаммы Mycobacterium. Примерами та-4 012182 ких других штаммов Mycobacteria являются, но не ограничиваются ими, Mycobacterium microti,Mycobacterium H37Ra, Mycobacterium vaccae и т.п. Отбор штамма BCG. В предшествующих работах было высказано предположение, что BCG не является гомогенным штаммом, и вместо этого штамма был получен массив из различных генетических линий (Oettinger et al.,Tuber Lung Dis. 79(4): 243-250, 1999). До настоящего времени не было точно известно, имеются ли какиелибо различия в иммуногенности и эффективности между членами семейства BCG. Однако в настоящее время, как описано в настоящем изобретении, было обнаружено, что конкретный штамм, из которого был получен рекомбинантный BCG (далее обозначаемый rBCG), по своей иммуногенности значительно отличается от rBCG. В примере 1, приведенном ниже, показано, что BCG штамм Дания 1331 (далее обозначаемый "BCG1331") представляет собой вакцину, обладающую значительно лучшими свойствами по сравнению с BCGTice. Таким образом, хотя сверхэкспрессия антигена 85 В в штамме rBCG30 способствует повышению иммуногенности этого штамма по сравнению с родительским штаммом BCGTice, из которого был получен указанный rBCG30, однако штамм rBCG30, сверхэкспрессирующий антиген 85 В, присутствующий в штамме BCGtice, не обнаруживал эффективности, аналогичной эффективности BCG1331. Таким образом, некоторые преимущества, получаемые в результате сверхэкспрессии антигена в BCG, могут быть достигнуты путем отбора эффективного родительского штамма BCG в начале процесса конструирования вакцины. Хотя задним числом такое решение может показаться очевидным, однако специалистами в данной области не было проведено такой коррекции (Horwitz et al., Proc. Natl. Acad. Sci. USA,97(25): 13853-13858, 2000), что свидетельствует о том, что ранее не было известно или не предполагалось, что перед началом конструирования rBCG-вакцин важно определить, обладает ли родительский штамм BCG адекватной эффективностью. Такие штаммы являются в высокой степени подходящими для сверхэкспрессии BCG и антигенов ТБ или чужеродных антигенов. Эффективные родительские штаммы BCG могут быть отобраны из группы, включающей, но не ограничивающейся ими, BCG1331, BCG Пастер, BCG Токио и BCG Копенгаген. При инокуляции морских свинок, используемых в качестве модели заражения, родительский штамм BCG, по сравнению со штаммом BCGTice, должен снижать уровень жизнеспособных инфекционных микроорганизмов Mycobacteriumtuberculosis по меньшей мере более чем на 0,41010 при введении в низкой дозе в форме аэрозоля, как показано ниже в примере 1. Усиление иммуногенности BCG. Как обсуждалось выше, иммуногенность штамма BCG не является постоянной. Более того, в примере 1 показано, что, хотя иммуногенность антигенов BCG может быть усилена путем генетической модификации BCG, однако едва ли такая модификация будет эффективной, если родительский штаммBCG, в котором были сделаны рекомбинантные изменения, не является эффективным в модели иммунизации морских свинок. Отсюда следует, что модификации, которые усиливают иммуногенность BCG,будут также усиливать иммуногенность рекомбинантных штаммов, полученных из указанных родительских штаммов. При применении подхода, подробно описанного выше, для отбора соответствующего штамма BCG,используемого в качестве родительского штамма для получения вакцины rBCG и вакцинных векторов, в указанный штамм вносят генетические модификации для генерирования нужных вакцин rBCG и вакцинных векторов. Методы, используемые для конструирования отдельных штаммов rBCG, не имеют важного значения для осуществления настоящего изобретения и в этих целях могут быть применены любые методы или их комбинации, известные специалистам в данной области (Horwitz et al., PNAS 97(25): 13853-13858, 2000; Hess et al., Proc. Natl. Acad. Sci. USA, 95: 5299-5304, 1998). Кроме того, в вакцинах rBCG предпочтительно использовать промотор, который активируется invivo после инфицирования. Так, например, использование конститутивно активного промотора, такого как промоторы, происходящие от антигена 85 В, антигена 85 А, Hsp60 или Rv1908c (KatG), обеспечивает конститутивную экспрессию данного антигена in vivo после иммунизации. Поэтому в каждой стадии в процессе инфицирования вырабатывается стойкий иммунный ответ против такой инфекции. Выбор активных промоторов на латентной стадии, таких как промоторы генов Rv2032, Rv3127, Rv2031c илиRv3030c и т.п., обеспечивает экспрессию выбранных антигенов в штамме rBCG, а особенно специфических антигенов на латентной стадии, когда вакцины rBCG вступают в латентную стадию in vivo после иммунизации. Экспрессионные векторы. Разработка системы отбора без использования антибиотиков. Как было указано выше, плазмиды, применяемые в настоящее время для сверхэкспрессии защитных антигенов в штаммах rBCG, в данном случае не могут быть использованы из-за того, что их сохранение зависит от присутствия в них генов резистентности к антибиотикам и из-за присущей этим плазмидам способности к горизонтальному переносу в микробные хозяева широкого ряда, что создает угрозу передачи генов резистентности к антибиотикам и экспрессии антигенных кластеров окружающим микроорганизмам. Для устранения этих важных ограничений в настоящем изобретении предлагаются новые,не основанные на антибиотиках системы отбора и однонаправленная система челночных векторов для-5 012182 введения и сохранения экспрессионных векторов в штаммах-хозяевах Mycobacterium, таких как rBCG. Отбор плазмид без применения антибиотиков может быть достигнут путем селективной делеции гена-хозяина, играющего важную роль в репликации, и последующей комплементации этой делеции путем введения функциональной копии гена в экспрессионную плазмиду. Таким образом, выживание бактериальных хозяев зависит от экспрессионной плазмиды, а поэтому необходим механизм сохранения плазмиды в микобактериальном хозяине без применения отбора на антибиотики. Предпочтительным методом является инактивация генов для создания ауксотрофного фенотипа. Так, например, в M.tb иBCG инактивация гена leuD (Genome SEQ ID NO Mb3011C) приводит к продуцированию лейцинзависимого фенотипа, и выживание штаммов, которые имеют инактивированный ген leuD, зависит от присутствия лейцина (Hondalus et al., Infect. Immun. 68(5): 2888-98. 2000). Кроме того, штаммы MycobacteriumleuD не способны реплицироваться in vivo (Hondalus et al., см. выше, 2000), и, таким образом, в leuDмутантах M.tb и rBCG будут сохраняться плазмиды leuD+ in vitro и in vivo. Выбор какого-либо конкретного метода введения ауксотрофной мутации в целевые штаммы Mycobacterium не имеет важного значения для осуществления настоящего изобретения и такой метод может быть выбран из любых методов аллельного обмена, хорошо известных специалистам в данной области(Parish et al., Microbiology, 145: 3497-3503, 1999). Аналогичным образом, комплементация ауксотрофной мутации может быть достигнута путем введения функциональной копии инактивированного гена (например, leuD+) в экспрессионный вектор. Такой экспрессионный вектор также требует присутствия ориджина репликации Mycobacterium (например, oriM; Labidi et al., Plasmid, 27(2): 130-140, 1992) для обеспечения репликации этого вектора в целевых штаммах M.tb и rBCG. Выживание штаммов Mycobacterium, содержащих такую плазмиду, зависит от экспрессии кодируемого плазмидой гена leuD после удаления лейцина из среды. Получение новых однонаправленных челночных векторов. Выше описан способ создания селективной системы для сохранения экспрессионных векторов вM.tb и rBCG. Однако для эффективного внесения модификаций в плазмидную структуру, которая затем вводит эти модификации в Mycobacterium, такая векторная система должна обладать способностью к репликации в Escherichia coli. Кроме того, для расширения круга потенциальных рекомбинантных штаммов-хозяев E.coli, которые могут быть использованы в процессе конструирования плазмид, исследователями был использован хозяин E.coli, который облегчает конструирование плазмиды, а в экспрессионный вектор предпочтительно были включены селектируемый маркер резистентности к антибиотикам (например, резистентности к канамицину) и ориджин репликации в широком круге хозяев (например, oriE;Halpern et al., Proc. Natl. Acad. Sci., USA, 76(12): 6137-6141, 1979; Mosig et al., New Biol. 1(2): 171-179,1989). Эти элементы были фланкированы уникальными сайтами расщепления рестрикционных эндонуклеаз (например, NdeI) для последующего удаления маркера резистентности к антибиотикам и ориджина репликации E.coli перед введением данной плазмиды в целевые штаммы Mycobacterium. Кроме того,могут быть включены уникальные рестрикционные сайты эндонуклеаз (например, PacI), в которые могут быть введены антигенные экспрессирующие кластеры. После осуществления этих модификаций в E.coli и после идентификации и характеризации нужной плазмиды рекомбинантную плазмидную ДНК выделяют и расщепляют рестрикционной эндонуклеазой, которая высвобождает селектируемый маркер резистентности к антибиотикам и oriE. Затем расщепленную плазмидную ДНК лигируют с использованием ДНК-лигазы Т 4. Полученная таким образом плазмида содержит ген, комплементирующий ауксотрофию хозяина Mycobacterium; не обладает резистентностью к антибиотику и не способна реплицироваться в грамотрицательных бактериях. Затем плазмиду, которая может также включать антигенный экспрессионный кластер, вводят в целевой ауксотрофный мутант Mycobacterium в соответствии со стандартными процедурами электропорации. Рекомбинантные штаммы, содержащие указанную плазмиду, выделяют путем культивирования в среде, не содержащей метаболита, необходимого для ее роста (например, лейцина). Уникальное преимущество этой системы состоит в том, что такая конечная экспрессионная плазмида больше не содержит ген резистентности к антибиотику. А поэтому ген резистентности к антибиотику не может передаваться другим обычно используемым экспрессионным плазмидам. Кроме того, экспрессионная плазмида согласно изобретению больше не способна реплицироваться в широком круге хозяев, поскольку из нее были удалены генетические элементы, обеспечивающие такую репликацию. Таким образом, указанные векторы будут далее называться "однонаправленными" челночными векторами. Сверхэкспрессия антигенов ТБ. В соответствии с настоящим изобретением ген, введенный в экспрессионный кластер в однонаправленном челночном векторе, а затем в rBCG, может кодировать иммуноген M.tb. Иммуногеном M.tb может быть, например, полноразмерный нативный белок, химерный гибридный белок, состоящий из двух или более иммуногенов или миметиков M.tb, или фрагмент или фрагменты иммуногена M.tb, происходящие от Mycobacterium tuberculosis. Антигены M.tb экспрессируются в однонаправленном челночном векторе под контролем промотора, который является активным по меньшей мере на одной стадии инфицирования микобактерией invivo. Выбор какого-либо конкретного промотора не имеет важного значения для осуществления настоящего изобретения, и такой промотор может быть выбран из конститутивно активных промоторов, таких-6 012182 как антиген 85 В, Hsp60, антиген 85 А, Rv1908c (KatG), и/или промоторов, которые являются активными на латентной стадии инфицирования, таких как промоторы генов Rv3130C (Florczyk et al., Infect. Immun. 71(9): 5332-5343, 2003; Voskuil et al., J. Exr. Med. 198(5): 705-713, 2003), Rv2032, Rv3127 и/или Rv2031c. Для увеличения уровня экспрессии антигена может быть использована миниклетка, продуцирующая производное векторов штаммов Mycobacterium. Миниклетку, продуцирующую штаммы бактерии видаMycobacterium, получают путем сверхэкспрессии FtsZ (Genome databaseMb2174 с) или путем сайтнаправленной инактивации whiB3. Изменение уровня экспрессии FtsZ или инактивация whiB3 могут быть осуществлены стандартными генетическими методами, хорошо известными специалистам в данной области. Так, например, сверхэкспрессию FtsZ осуществляют путем введения гена ftsZ в однонаправленный челночный вектор под контролем сильного промотора, такого как промоторы антигена 85 В, антигена 85 А, Hsp60 или Rv1908c (KatG), которые являются конститутивно активными, и/или промоторы, которые являются активными на латентной стадии инфицирования, такие как промоторы генов Rv2032,Rv3127, Rv2031c, and Rv3130C (Florczyk et al., см. выше, 2003; Voskuil et al., см. выше, 2003). Сайтнаправленную инактивацию whiB3 осуществляют путем аллельной замены в соответствии с процедурами, описанными ниже. Примеры чужеродных антигенов, которые могут быть введены в рекомбинантную Mycobacterium. В соответствии с настоящим изобретением экспрессионный кластер в однонаправленном челночном векторе, присутствующий в векторе Mycobacterium, может кодировать иммуноген, которым может быть либо чужеродный иммуноген, происходящий от вирусных, бактериальных или паразитарных патогенов, либо эндогенный иммуноген, такой как, но не ограничивающийся им, аутоиммунный антиген или опухолевый антиген. Такими иммуногенами могут быть, например, полноразмерный нативный белок,химерные гибриды, состоящие из чужеродного иммуногена и эндогенного белка или миметика, или фрагмент или фрагменты иммуногена, происходящего от вирусных, бактериальных и паразитарных патогенов. Используемый здесь термин "чужеродный иммуноген" означает белок или его фрагмент, которые обычно не экспрессируются в клетках или тканях животного-реципиента, такие как, но не ограничивающиеся ими, вирусные белки, бактериальные белки, паразитарные белки, цитокины, хемокины, иммунорегуляторные агенты или терапевтические средства. Термин "эндогенный иммуноген" означает белок или его часть, которые обычно присутствуют в клетках или тканях животного реципиента, такие как, но не ограничивающиеся ими, эндогенный клеточный белок, иммунорегуляторный агент или терапевтическое средство. Альтернативно или дополнительно такой иммуноген может кодироваться синтетическим геном и может быть сконструирован стандартными методами рекомбинантных ДНК, известными специалистам в данной области. Чужеродным иммуногеном может быть любая молекула, которая экспрессируется любым вирусным, бактериальным или паразитарным патогеном, до или в процессе его инвазии или колонизации организма животного-хозяина или репликации в этом организме. rBCG может экспрессировать иммуногены или их части, происходящие от вирусных, бактериальных и паразитарных патогенов. Такие патогены могут быть инфекционными для человека, домашних животных или диких животных. Вирусными патогенами, от которых происходят вирусные антигены, являются, но не ограничиваются ими, ортомиксовирусы, такие как вирус гриппа (Taxonomy ID: 59771); ретровирусы, такие как RSV,HTLV-1 (Taxonomy ID: 39015) и HTLV-II (Taxonomy ID: 11909); герпесвирусы, такие как EBV (Taxonomy ID: 10295); CMV (Taxonomy ID: 10358) или вирус простого герпеса (АТССVR-1487); лентивирусы, такие как ВИЧ-1 (Taxonomy ID: 12721) и ВИЧ-2 (Taxonomy ID: 11709); рабдовирусы, такие как вирус бешенства; пикорновирусы, такие как полиовирус (Taxonomy ID: 12080); поксвирусы, такие как вирус коровьей оспы (Taxonomy ID: 10245); ротавирус (Taxonomy ID: 10912) и парвовирусы, такие как аденоассоциированный вирус 1 (Taxonomy ID: 85106). Примерами вирусных антигенов могут служить антигены группы, включающей, но не ограничивающейся ими, антигены вируса иммунодефицита человека Nef (National Institute of Allergy and Infectious(National Institute of Allergy and Infectious Disease HIV Repository Cat.238; GenBank accessionAJ237568) и T- и В-клеточные эпитопы gp120 (Hanke and McMichael, AIDS Immunol. Lett., 66: 177,1999; Hanke, et al., Vaccine, 17: 589, 1999; Palker et al., J. Immunol., 142: 3612-3619, 1989), химерные производные Env и gp120 ВИЧ-1, такие как, но не ограничивающиеся ими, гибриды белков gp120 и CD4(Fouts et al., J. Virol. 2000, 74: 11427-11436, 2000); усеченные или модифицированные производные env ВИЧ-1, такие как, но не ограничивающиеся ими, gp140 (Stamatos et al. J. Virol., 72: 9656-9667, 1998), или производные Env и/или gp140 ВИЧ-1 (Binley, et al. J. Virol., 76: 2606-2616, 2002; Sanders, et al. J. Virol.,74: 5091-5100, 2000; Binley, et al. J. Virol., 74: 627-643, 2000), поверхностный антиген вируса гепатита В(GenBank accessionAF043578; Wu et al., Proc. Natl. Acad. Sci., USA, 86: 4726-4730, 1989); ротавирус-7 012182 ные антигены, такие как VP4 (GenBank accessionAJ293721; Mackow et al., Proc. Natl. Acad. Sci., USA,87: 518-522, 1990) и VP7 (GenBank accessionAY003871; Green et al., J. Virol., 62: 1819-1823, 1988), антигены вируса гриппа, такие как гемагглютинин (GenBank accessionAJ404 627; Pertmer and Robinson,Virology, 257: 406, 1999); нуклеопротеин (GenBank accessionAJ289872; Lin et al., Proc. Natl. Acad. Sci.,97: 9654-9658, 2000), антигены вируса простого герпеса, такие как тимидинкиназа (GenBank accessionAB047378; Whitley et al., In: New Generation Vaccines, p. 825-854, 2004). Бактериальными патогенами, от которых происходят бактериальные антигены, являются, но не ограничиваются ими, Mycobacterium spp., Helicobacter pylori, Salmonella spp., Shigella spp., E.coli, Rickettsiaspp., Listeria spp., Legionella pneumoniae, Pseudomonas spp., Vibrio spp. и Borellia burgdorferi. Примерами защитных антигенов бактериальных патогенов являются соматические антигены энтеротоксигенных бактерий E.coli, такие как антиген фимбрий CFA/I (Yamamoto et al., Infect. Immun., 50: 925-928, 1985) и нетоксическая В-субъединица термолабильного токсина (Klipstein et al., Infect. Immun.,40: 888-893, 1983); пертактин Bordetella pertussis (Roberts et al., Vacc., 10: 43-48, 1992), аденилатциклазагемолизин В. pertussis (Guiso et al., Micro. Path., 11: 423-431, 1991), фрагмент С столбнячного токсинаClostridium tetani (Fairweather et al., Infect. Immun., 58: 1323-1326, 1990), OspA Borellia burgdorferi (Sikand,et al., Pediatrics, 108: 123-128, 2001; Wallich, et al., Infect Immun., 69: 2130-2136, 2001), защитные паракристаллические поверхностные белки Rickettsia prowazekii и Rickettsia typhi (Carl, et al., Proc. Natl. Acad.Sci. USA, 87: 8237-8241, 1990), листериолизин (также известный как "Llo" и "Hly") и/или супероксиддисмутаза (также известная как "SOD" and "p60") Listeria monocytogenes (Hess, J., et al., Infect. Immun. 65: 1286-92, 1997; Hess, J., et al., Proc. Natl. Acad. Sci. 93: 1458-1463, 1996; Bouwer, et al., J. Exp. Med. 175: 1467-71, 1992), уреаза Helicobacter pylori (Gomez-Duarte, et al., Vaccine 16, 460-71, 1998; Corthesy-Theulaz,et al., InfectionImmunity 66, 581-6, 1998) и связывающийся с рецептором домен летального токсина и/или защитный антиген Bacillus anthrax (Price, et al., Infect. Immun. 69, 4509-4515, 2001). Паразитарными патогенами, от которых происходят паразитарные антигены, являются, но не ограничиваются ими, Plasmodium spp., такие как Plasmodium falciparum (ATCC30145); Trypanosome spp.,такие как Trypanosoma cruzi (ATCC50797); Giardia spp., такие как Giardia intestinalis (ATCC30888D); Boophilus spp., Babesia spp., такие как Babesia microti (ATCC30221); Entamoeba spp., такие как Entamoeba histolytica (ATCC30015); Eimeria spp., такие как Eimeria maxima (ATCC40357);Leishmania spp. (Taxonomy ID: 38568); Schistosome spp., Brugia spp., Fascida spp., Dirofilaria spp., Wuchereria spp. и Onchocerea spp. Примерами защитных антигенов паразитарных патогенов являются антигены круглого спорозоитаPlasmodium spp. (Sadoff et al., Science, 240: 336-337, 1988), такие как антиген круглого спорозоитаP.bergerii или антиген круглого спорозоита P.falciparum; поверхностный антиген мерозоита Plasmodium(Handman et al., Vaccine, 18: 3011-3017, 2000), парамиозин Brugia malayi (Li et al., Mol. Biochem. Parasitol.,49: 315-323, 1991), триозо-фосфат-изомераза Schistosoma mansoni (Shoemaker et al., Proc. Natl. Acad. Sci.,USA, 89: 1842-1846, 1992); секретируемый глобинподобный белок Trichostrongylus colubriformis (Frenkelet al., Mol. Biochem. Parasitol., 50: 27-36, 1992); глутатион-S-трансфераза Frasciola hepatica (Hillyer et al.,Exp. Parasitol., 75: 176-186, 1992), Schistosoma bovis и S. japonicum (Bashir et al., Trop. Geog. Med., 46: 255258, 1994) и KLH Schistosoma bovis и S. japonicum (Bashir et al., см. выше, 1994). Как было упомянуто выше, вакцины rBCG могут кодировать эндогенный иммуноген, которым может быть любой клеточный белок, иммунорегуляторный агент или терапевтическое средство, или их части, которые могут экспрессироваться в клетке реципиента, включая, но не ограничиваясь ими, опухолевые иммуногены, иммуногены трансплантатов и аутоиммунные иммуногены или их фрагменты и производные. Таким образом, в соответствии с настоящим изобретением rBCG могут кодировать опухолевые иммуногены, иммуногены трансплантатов или аутоиммунные иммуногены или их части или производные. Альтернативно, rBCG может содержать синтетические гены (описанные выше), кодирующие опухолеспецифические антигены, антигены трансплантантов или аутоиммунные антигены или их части. Примерами опухолеспецифических антигенов являются специфический для предстательной железы антиген (Gattuso et al., Human Pathol., 26: 123-126, 1995), TAG-72 и СЕА (Guadagni et al., Int. J. Biol.Markers, 9: 53-60, 1994), MAGE-1 и тирозиназа (Coulie et al., J. Immunothera., 14: 104-109, 1993). Недавно было обнаружено, что у мышей, которые были иммунизованы незлокачественными клетками, экспрессирующими опухолевый антиген, наблюдается вакцинный эффект, а также вырабатывается иммунный ответ, способствующий выведению злокачественных опухолевых клеток, несущих тот же самый антиген(Koeppen et al., Anal. N.Y. Acad. Sci., 690: 244-255, 1993). Примером антигенов трансплантатов является молекула CD3 на Т-клетках (Alegre et al., Digest. Dis.Sci., 40: 58-64, 1995). Было показано, что обработка антителом против рецептора CD3 способствует быстрому выведению циркулирующих Т-клеток и предотвращению клеточно-опосредуемого отторжения трансплантата (Alegre et al., см. выше, 1995).-8 012182 Примером аутоиммунноых антигенов является -цепь IAS (Topham et al., Proc. Natl. Acad. Sci.,USA, 91: 8005-8009, 1994). Было продемонстрировано, что вакцинация мышей пептидом -цепи IAS,состоящим из 18 аминокислот, вызывает у мышей с экспериментальным аутоиммунным энцефаломиелитом выработку иммунитета к указанному заболеванию и подавление такого заболевания (Topham et al.,см. выше, 1994). Получение штамма rBCG, экспрессирующего адъювант. Может быть сконструирован rBCG, кодирующий иммуноген и адъювант, и такой штамм может быть использован для усиления иммунного ответа у хозяина на rBCG. Альтернативно, для усиления ответа у хозяина на иммуногены, кодируемые партнером rBCG, может быть сконструирован адъюванткодирующий rBCG в комбинации с другими rBCG. Выбор адъюванта, кодируемого rBCG, не имеет решающего значения для осуществления изобретения и такими адъювантами могут быть субъединица А холерного токсина (например, CtxA; GenBankaccessionX00171, AF175708, D30053, D30052) или ее части и/или мутантные производные (например,домен А 1 субъединицы A Ctx (например, CtxA1; GenBank accession . K02679), происходящие от любого классического штамма Vibrio cholerae (например, штамма V. cholerae 395, АТСС 39541) или штамма El Tor V. cholerae (например, штамма V. cholerae 2125, АТСС 39050). Альтернативно, вместо CtxA может быть использован любой бактериальный токсин, который является членом семейства бактериальных аденозин-дифосфат-рибозилирующих экзотоксинов (Krueger and Barbier, Clin. Microbiol. Rev., 8: 34,1995), например субъединица А термолабильного токсина (обозначаемого далее EltA) энтеротоксигенной бактерии Escherichia coli (GenBank accessionM35581), субъединица S1 коклюшного токсина (например, ptxS1, GenBank accessionAJ007364, AJ007363, AJ006159, AJ006157, и т.п.); а в качестве другой альтернативы таким адъювантом может быть один из аденилат-циклазогемолизинов бактерий(АТСС 15237), например гены суаА В. pertussis (GenBank accessionX14199), В.parapertussis(GenBank accessionAJ24 9835) или В.bronchiseptica (GenBank accessionZ37112). Получение штамма rBCG, экспрессирущего иммунорегуляторный агент. Может быть сконструирован rBCG, кодирующий иммуноген и цитокин, и такой штамм может быть использован для усиления иммунного ответа у хозяина на rBCG. Альтернативно, для усиления ответа у хозяина на иммуногены, кодируемые партнером rBCG, может быть сконструирован rBCG, кодирующий указанный цитокин, либо отдельно, либо в комбинации с другими rBCG. Выбор конкретного цитокина, кодируемого rBCG, не имеет решающего значения для осуществления изобретения и такими цитокинами могут быть, но не ограничиваются ими, интерлейкин-4 (обозначаемый далее "IL-4"; GenBank accessionAF352783 (мышиный IL-4) или NM000589 (человеческий IL4; IL-5 (GenBank accessionNM010558 (мышиный IL-5) или NM000879 (человеческий IL-5; IL-6TNF. Апоптоз представляет собой запрограммированную клеточную гибель, которая, по характеру и по последствиям ее индуцирования, значительно отличается от некротической клеточной гибели. Апоптоз клеток, содержащих чужеродные антигены, представляет собой известный мощный стимулятор развития клеточного иммунитета против таких антигенов. Механизм, посредством которого апоптоз антигенсодержащих клеток приводит к вырабатыванию клеточного иммунитета, иногда называют перекрестным примированием (W.R. Heath, G.T. Belz, G.M. Behrens, С.M.Smith, S.P. Forehan, I.A., Parish, G.M. Davey,N.S. Wilson, F.R. Carbone and J.A. Villandangos. 2004. Cross-presentaion, dentritic cell subsets, and the generation of immunity to cellular antigens. Immunol. Rev. 199: 9; S. Gallucci, M. Lolkema, and P. Matzinger. 1999. Natural adjuvants: Endogenous activators of dendritic cells. Nature Biotechnology. 5: 1249; M.L. Albert,B. Sauter, and N. Bhadrdwaj. 1998. Dendtritic cells acquire antigen from apoptotic cells and induce classI-restricted CTLs. Nature 392: 86). Существует несколько механизмов индуцирования апоптоза, которые приводят к усилению антигенспецифического клеточно-опосредуемого иммунитета. Апоптоз, опосредуемый каспазой 8, приводит к вырабатыванию антигенспецифического клеточного иммунитета. Продуцирование каспазы 8 штаммом rBCG и ее секреция в цитоплазму эукариотических клеток в присутствии чужеродных антигенов, экспрессируемых штаммом rBCG, антигенов против BCG и других туберкулезных антигенов, сверхэкспрессируемых штаммом rBCG, а также антигенов против самих антигенов BCG приводит к вырабатыванию высокого уровня антигенспецифического клеточного иммунитета. Рецептор 5 гибели клеток (DR-5), также известный как TRAIL-R2 (рецептор 2 TRAIL) или TNFR-SF-10B (член суперсемейства факторов некроза опухоли 10 В), также индуцирует апоптоз, опосредуемый каспазой 8by a family of signaling and decoy receptors. Science, 277: 818). Индуцируемый реовирусом апопотоз опосредуется TRAIL-DR5 и приводит к последующему выведению вируса (P. Clarke, S.M. Meintzer,S. Gibson, С. Widmann, Т.P. Garrington, G.L. Johnson, and K.L. Tyler. 2000. Reovirus-induced apoptosis ismediated by TRAIL. J. Virol. 74: 8135). Экспрессия DR-5 рекомбинантным штаммом BCG обеспечивает сильное стимулирующее действие, направленное на индуцирование антигенспецифического клеточного иммунитета против rBCG-экспрессируемых антигенов. Апоптоз антигенэкспрессирующих клеток также может быть индуцирован посредством присоединения Fas, который является сильным стимулятором индуцирования антигенспецифических клеточных иммунных ответов (M.A. Chattergoon, J.J. Kim,J.S. Yang, Т.M. Robbinson, D.J. Lee, T. Dentchev, D.M. Wilson, V. Ayyavoo, and D.B. Weiner. 2000. TargetedBiotechnology, 18: 974). Рекомбинантный BCG, экспрессирующий Fas или гибридный белок, содержащий цитоплазматический домен Fas/эктодомен CD4, индуцирует апопотоз и антигенспецифические клеточные иммунные ответы. Усиление клеточного иммунитета под действием штамма rBCG, который продуцирует стимуляторы апоптоза, описанные выше, не ограничивается только действием антигенов BCG или антигенов, специфически кодируемых благодаря их сверхэкспресси в rBCG, a также может быть вызвано действием любого антигена, присутствующего в эукариотической клетке, которая может быть инфицирована вышеупомянутым штаммом rBCG. Так, например, если такой штамм rBCG ввести в опухолевые клетки, апоптоз которых он индуцирует, то это будет приводить к вырабатыванию клеточного иммунитета против важных опухолевых антигенов с последующей элиминацией, подавлением или предотвращением роста опухоли и/или возникновения метастазов. Такой противоопухолевый эффект может быть продуцирован в дополнение к общему противоопухолевому эффекту, который вырабатывается штаммом BCG при его местном введении, например, при раке мочевого пузыря. В другом варианте настоящего изобретения rBCG, стимулируемый посредством продуцирования специфических медиаторов апоптоза, доставляемых внутрь опухоли или других клеток, где указанныйrBCG также продуцирует чужеродные антигены, против которых будут вырабатываться сильные клеточные иммунные ответы, будет индуцировать вырабатывание сильных клеточных ответов против клеток,содержащих указанные чужеродные антигены. Такие клеточные ответы будут приводить к иммуноопосредуемой деструкции опухолевых клеток, а также к перекрестному примированию и индуцированию клеточного иммунного ответа против опухолевых или других важных антигенов с последующим уничтожением опухоли и/или метастазов или подавлением или профилактики развития опухоли и/или метастазов. Примером такого чужеродного антигена является антиген HLA, отличающийся от HLA клетокхозяев, против которого вырабатывается сильный гетерологичный клеточный ответ. Штамм rBCG, способность которого к идуцированию апоптоза усиливается путем экспрессии специфических медиаторов апоптоза, также экспрессирующих специфические опухолевые антигены, будет индуцировать сильные антигенспецифические клеточные ответы против этих опухолевых антигенов,включая подавление, до некоторой степени, толерантности к этим антигенам, что будет приводить к уничтожению опухоли и/или метастазов или к подавлению или профилактики развития опухоли и/или метастазов, и не требует прямой доставки rBCG в саму опухоль. Апоптоз, вызываемый разрушением ДНК или действием каспазы 9, индуцирует толерантность к некоторым антигенам (S. Hugues, Е. Mougneau, W. Ferlin, D. Jeske, P. Hofman, D. Homann, L. Beaudoin,D. Schrike, M. Von Herrath, A. Lehuen, and N. Glaichenenhaus, 2002, Tolerance to islet antigens and prevention from diabetes induced by limited apoptosis of pancreatic beta cells. Immunity, 16: 169). Индуцирование толерантности играет важную роль в лечении или профилактике аутоиммунных заболеваний, таких как,но не ограничивающихся ими, диабет, ревматоидный артрит, болезнь Крона, воспалительное заболевание кишечника и рассеянный склероз. Продуцирование каспазы 9 или других белков, индуцирующих опосредуемую апоптозом толерантность, штаммом rBCG в клетках, таких как, но не ограничивающихся ими, панкреатические В-клетки, клетки ободочной кишки и нервные клетки, будет приводить к ограниченному апоптозу, который будет индуцировать толерантность к антигенам-мишеням, продуцирующим аутоиммунитет в этих клетках, и, тем самым, способствовать лечению или профилактике аутоиммунных заболеваний. Идентификация специфических антигенов, участвующих в аутоиммунных реакциях, позволяет индуцировать толерантность к этим аутоиммунным антигенам-мишеням посредством продуцирования штаммом rBCG этих антигенов и каспазы 9 или других молекул, способных индуцировать опосредуемую апоптозом толерантность. Такой rBCG может быть использован для лечения и/или профилактики этих аутоиммунных заболеваний. Методы рекомбинантных ДНК и РНК, применяемые для введения функциональных экспрессионных кластеров в целях создания rBCG, способного экспрессировать иммунорегуляторный агент в эукариотических клетках или тканях, описаны ниже. Нижеследующие примеры приводятся в целях иллюстрации различных аспектов настоящего изобретения и не должны рассматриваться как ограничение осуществления изобретения. При этом следует- 10012182 отметить, что в настоящем изобретении могут быть использованы различные альтернативные материалы, условия и процедуры, которые являются очевидными для среднего специалиста в данной области, но которые не должны выходить за рамки общего объема изобретения, представленного в его описании. Примеры Методы. Культивирование штаммов Mycobacterium. Выбранные штаммы BCG культивировали в жидкой среде, такой как среда Миддлбрука 7 Н 9 или синтетическая среда Saulton, предпочтительно при 37 С. Эти штаммы могут храниться в виде стационарной или перемешиваемой культуры. Кроме того, скорость роста BCG может быть увеличена путем добавления олеиновой кислоты (0,06% об./об.; Research Diagnostics Cat.01257) и детергентов, таких как тилоксапол (0,05% об./об.; Research Diagnostics Cat.70400). Чистота культур BCG может быть оценена путем равномерного посева 100 мкл аликвот культуры BCG, серийно разведенной (например,10-кратно разведенной чистой культуры (Neat) - 10-8) в забуференном фосфатом физиологическом растворе (далее обозначаемом PBS), на 3,5-дюймовых чашках, содержащих 25-30 мл твердой среды, такой как среда Миддлбрука 7 Н 10. Кроме того, чистота указанной культуры может быть дополнительно оценена с использованием коммерчески доступных наборов, таких как наборы, содержащие тиогликолят(Science Lab, Cat.1891) и соевую среду с казеином (BD, Cat.211768). Посевные лоты BCG хранили при -80 С при плотности 0,1-2107 КОЕ/мл. Жидкие культуры обычно собирали при оптической плотности (600 нм), составляющей 0,2-4,0 по отношению к стерильному контролю и эти культуры высевали в центрифужные пробирки соответствующего размера, а затем микроорганизмы подвергали центрифугированию при 8,000g в течение 5-10 мин. Супернатант отбрасывали и микроорганизмы ресуспендировали в растворе для хранения, состоящем из среды Миддлбрука 7 Н 9,содержащей 10-30% (об./об.) глицерина при плотности 0,1-2107 КОЕ/мл. Эти суспензии в аликвотах по 1 мл распределяли по стерильным 1,5-миллилитровым боросиликатным сосудам для глубокого замораживания, а затем хранили при -80 С. Общие методы молекулярной биологии. Рестриктирующие эндонуклеазы (обозначаемые здесь "RE"); (New England Biolabs Beverly, MA),ДНК-лигазу Т 4 (New England Biolabs, Beverly, MA) и полимеразу Taq (Life Technologies, Gaithersburg,MD) использовали в соответствии с протоколами производителей. Плазмидную ДНК получали с использованием наборов для лабораторной очистки (Qiagen Miniprep kit, Santa Clarita, CA) или для крупномасштабной очистки (Qiagen Maxiprep kit, Santa Clarita, CA) плазмидных ДНК в соответствии с протоколами производителей (Qiagen, Santa Clarita, CA). Воду milli-Q, не содержащую нуклеазы и очищенную до чистоты, требуемой для проведения молекулярно-биологических экспериментов; трис-HCl (рН 7,5),EDTA, рН 8,0, 1 М MgCl2, 100% (об./об.) этанол, сверхчистую агарозу и буфер для электрофореза в агарозном геле закупали у фирмы Life Technologies, Gaithersburg, MD. Расщепление ферментами рестрикции, ПЦР, реакции лигирования ДНК и электрофорез в агарозном геле проводили в соответствии с хорошо известными процедурами (Sambrook, et al., Molecular Cloning: A Laboratory Manual. 1, 2, 3, 1989);Straus, et al., Proc. Natl. Acad. Sci. USA. Mar; 87(5): 1889-93, 1990). Секвенирование нуклеотидов для подтверждения последовательности ДНК каждой рекомбинантной плазмиды, описанной в нижеследующих разделах, проводили в соответствии со стандартными автоматизированными методами секвенирования ДНК с использованием автоматического секвенатора Applied Biosystems, модели 373 А. ПЦР-праймеры закупали у коммерческих фирм-поставщиков, таких как Sigma (St. Louis, МО), и синтезировали с использованием ДНК-синтезатора Applied Biosystems (модель 373 А). ПЦР-праймеры использовали при концентрации 150-250 мкМ, а температуру отжига для ПЦР-реакций определяли с помощью компьютерной программы Clone manager software version 4.1 (Scientific and Educational SoftwareInc., Durham NC). ПЦР проводили на роботизированной термоячейке Strategene Robocycler, модель 400880 (Strategene, La Jolla, CA). ПЦР-праймеры для амплификации конструировали с помощью компьютерной программы Clone Manager software version 4.1 (Scientific and Educational Software Inc., DurhamNC). Эта программа позволяет конструировать ПЦР-праймеры и идентифицировать рестрикционные сайты, совместимые со специфическими ДНК-фрагментами, подвергаемыми модификации. ПЦР проводили в термоячейке, такой как роботизированная термоячейка Strategene Robocycler, модель 400880(Strategene), а время отжига, удлинения и денатурации праймеров в ПЦР устанавливали в соответствии с протоколами стандартных процедур (Straus et al., см. выше, 1990). Затем расщепление ферментами рестрикции и ПЦР анализировали с помощью электрофореза в агарозном геле стандартными методами(Straus et al., см. выше, 1990, и Sambrook et al., см. выше, 1989). Положительный клон определяли как клон, который обнаруживал соответствующую рестрикционную карту и/или картину ПЦР. Плазмиды,идентифицированные в соответствии с указанной процедурой, могут быть дополнительно оценены в соответствии со стандартными процедурами секвенирования ДНК, описанными выше. Штаммы Escherichia coli, такие как DH5 и Sable2, закупали у фирмы Life Technologies (Gaithersburg, MD), и эти штаммы служили в качестве исходного хозяина для рекомбинантных плазмид, описанных ниже в примерах. Рекомбинантные плазмиды вводили в штаммы E.coli путем электропорации с- 11012182 применением электроимпульсного устройства для подачи импульсов высокого напряжения, такого какGene Pulser (BioRad Laboratories, Hercules, CA), с установленными параметрами 100-200 Ом, 15-25 мкФ и 1,0-2,5 кВ, как описано в литературе (Straus et al., см. выше, 1990). Оптимальные условия электропорации идентифицировали путем определения параметров, которые дают максимальную скорость трансформации на 1 мкг ДНК/бактерию. Бактериальные штаммы культивировали на трипсинизированном соевом агаре (Difco, Detroit, MI) или в трипсинизированном соевом бульоне (Difco, Detroit, MI), которые приготавливали в соответствии с инструкциями производителя. Если это не указано особо, то все бактерии культивировали при 37 С в 5%CO2 (об./об.) при легком помешивании. Если необходимо, то в среды добавляли антибиотики (Sigma, St.Louis, МО). Бактериальные штаммы хранили при -80 С и суспендировали в среде Difco, содержащей 30% (об./об.) глицерина (Sigma, St. Louis, МО) при плотности примерно 109 колониеобразующих единиц(далее обозначаемых "КОЕ") на 1 мл. Аллельный обмен в BCG. Ранее были описаны методы введения модифицированных аллелей в штаммы Mycobacterium, и эти методы являются известными и могут быть применены специалистами в данной области (Parish et al.,Microbiology, 146: 1969-1975, 2000). Новый метод генерирования плазмиды с заменой аллелей предусматривает использование синтезированной ДНК. Преимущество такого метода заключается в том, что о данном плазмидном продукте, полученном таким методом, может быть дана полная информация, а поэтому на его применение может быть получено разрешение Регуляторных органов, тогда как применяемые ранее методы, хотя и являются эффективными, очень плохо задокументированы с точки зрения протоколов проведения лабораторного культивирования, а поэтому на осуществление этих методов может быть не получено разрешения. Если данный продукт используется для введения человеку, то в соответствии с существующими правовыми нормами на его применение должно быть получено разрешение Регуляторных органов США и Европейских стран. Вектором-"самоубийцей" для замены аллелей в Mycobacterium является плазмида, которая способна реплицироваться в штаммах E.coli, но не способна реплицироваться в штаммах Mycobacterium spp.,таких как M.tb и BCG. Выбор того или иного конкретного вектора-"самоубийцы" для его использования в процедурах аллельного обмена не имеет решающего значения для осуществления настоящего изобретения, и такой вектор может быть выбран из векторов, полученных в научно-исследовательских лабораториях (Parish et al., см. выше, 2000) или являющихся коммерчески доступными. Предпочтительная конструкция плазмиды-"самоубийцы" для аллельного обмена представлена на фиг. 1. Эта плазмида состоит из нижеследующих ДНК-сегментов: последовательности oriE для репликации данной плазмиды в E.coli(GenBank accessionL09137); последовательности гена резистентности к канамицину для отбора вE.coli и в Mycobacterium (GenBank accessionААМ 97345) и вспомогательного маркера для отбора на резистентность к антибиотику (например, гена резистентности к зеоцину (GenBank accessionAAU06610, который находится под контролем промотора Mycobacterium (например, промотораhsp60). Указанный второй маркер для отбора на резистентность к антибиотику не является обязательным, и он был включен для осуществления двойного отбора в целях предотвращения чрезмерного роста резистентных к канамицину изолятов, спонтанно образующихся в процессе аллельного обмена. (Garbe etal., Microbiology, 140: 133-138, 1994). Конструирование таких векторов-"самоубийц" может быть осуществлено стандартными методами рекомбинантных ДНК, описанными в настоящем изобретении. Однако современные стандарты, разработанные Регуляторными органами, требуют обратить внимание на угрозу попадания прионовых частиц,образующихся в продуктах, обработанных коровьими продуктами, содержащими BSE-инфицированный материал. Поэтому во избежание попадания этих материалов (например, последовательностей ДНК) в целевой штамм неизвестного происхождения предпочтительно, чтобы все ДНК в векторе-"самоубийце" были синтезированы с использованием коммерческих источников (например, Picoscript, Inc.). В соответствии с этим предпочтительным методом конструирования векторов-"самоубийц" является разработка протокола получения последовательностей ДНК с помощью компьютерной программы для конструирования ДНК (например, Clone Manager) с последующим синтезом ДНК коммерческой фирмойпоставщиком (например, Picoscript Inc.), предоставляющей такие услуги за соответствующую плату. Этот метод был использован для продуцирования вектора-"самоубийцы" pA100 (не приводится), который был затем модифицирован для его применения в конкретных целях (pAF103 схематически представлен на фиг. 1, а также описан в таблице). Вектор-"самоубийца" Такой вектор-"самоубийца" имеет те преимущества, что он содержит два маркера для отбора на резистентность к антибиотикам, что позволяет минимизировать отбор спонтанных мутантов, обнаруживающих резистентность к одному антибиотику и присутствующих в отношении примерно 1/108 на генерацию. Спонтанная резистентность к двум антибиотикам встречается крайне редко и наблюдается только- 12012182 примерно в 1 случае на 1016-генерацию. Таким образом, вероятность появления штаммов, обладающих такой двойной резистентностью, в культурах, используемых для проведения процедуры аллельного обмена, составляет менее чем 1/106. Для негативного отбора в процессе аллельного обмена был включен ген sacB (геномная последовательность SEQ IDNT01BS4354), который сообщает фенотип чувствительности к сахарозе, в целях обогащения культуры штаммами, которые были подвергнуты конечной стадии рекомбинации ДНК и в которых была завершена процедура аллельного обмена. Стратегия изготовления вакцины и вакцинации. Стратегия изготовления вакцины основана на исследованиях, проводимых в целях определения максимальной жизнеспособности и стабильности этой вакцины в процессе ее изготовления. Эта стратегия включает оценку максимальной жизнеспособности данного микроорганизма (времени жизни) в процессе культивирования с использованием различных стандартных сред, обычно применяемых для культивирования Mycobacteria, с добавлением глицерина, сахаров, аминокислот, детергентов или солей. После культивирования клетки собирают путем центрифугирования или фильтрации в тангенциальном потоке и ресуспендируют в стабилизирующей среде для защиты клеток во время замораживания или лиофилизации. Обычно используемыми стабилизирующими агентами являются глутамат натрия или аминокислоты или их производные, глицерин, сахара или обычно используемые соли. Конечная композиция должна содержать микроорганизм, который является достаточно жизнеспособным для его доставки путем внутрикожного введения, чрескожного введения, вливания или перорального введения и достаточно стабильным при его хранении, кроме того, она должна иметь адекватный срок годности для ее доставки и применения. Преклиническая оценка вакцин против туберкулеза. Общая процедура испытания на безопасность. Группы мышей BALB/c, состоящие из шести животных, внутрибрюшинно инфицировали 2106 КОЕ представляющего(их) интерес штамма(ов) rBCG и аналогичных родительских штаммов. Мониторинг общего состояния животных и их массы тела проводили в течение 14 дней после инфицирования. Животные, которым вводили штаммы BCG и rBCG, оставались здоровыми, и у них не наблюдалось снижения массы тела и не обнаруживалось каких-либо явных признаков заболевания за время их наблюдения. Вирулентность новых штаммов rBCG у иммунокомпетентных мышей. Группы, состоящие из 15 иммунокомпетентных мышей BALB/c, внутривенно инфицировали 2106 КОЕ штамма rBCG и родительского штамма BCG соответственно. На 1-й день после инфицирования трех мышей в каждой группе умерщвляли и оценивали КОЕ в селезенке, легких и печени для подтверждения того, что каждое животное получило одну и ту же дозу заражение. Через 4, 8, 12 и 16 недель после инфицирования трех мышей в каждой группе умерщвляли и оценивали КОЕ в селезенке, печени и легких для анализа роста штаммов rBCG in vivo по сравнению с ростом родительского штамма BCG. Штаммы rBCG, как и ожидалось, обнаруживали вирулентность, аналогичную вирулентности родительского штамма BCG. Строгий тест на безопасность, проводимый на мышах с иммунодефицитом. Мышей с иммунодефицитом SCID (с тяжелым комбинированным иммунодефицитом), распределенных на группы по 10 животных в каждой, внутривенно инфицировали 2106 КОЕ штамма rBCG и родительского штамма BCG соответственно. На 1-й день после инфицирования трех мышей в каждой группе умерщвляли и оценивали КОЕ в селезенке, печени и легких для подтверждения того, что каждое животное получило одну и ту же дозу заражения. Остальных семь мышей в каждой группе подвергали мониторингу на их общее состояние и массу тела. Затем оценивали выживаемость этих мышей и результаты этого теста считались положительными, если в течение всего периода наблюдения показатели выживаемости rBCG-инфицированных мышей были лучше, чем показатели выживаемости животных, которые были инфицированы родительским штаммом. Тест на безопасность для морских свинок. Безопасность штаммов rBCG по сравнению с вакциной BCG, полученной на основе родительского штамма и имеющей хорошо установленный профиль безопасности для человека, также оценивали на морских свинках, используемых в качестве модели. Сначала оценивали влияние вакцины на общее состояние здоровья животных, включая их массу тела. Морских свинок внутримышечно иммунизировали 107 (100 дозой вакцинации) КОЕ рекомбинантного и родительского штаммов и проводили мониторинг общего состояния здоровья животных и их массы тела в течение шести недель. Животных, которые погибали до завершения этого шестинедельного периода времени, подвергали паталогоанатомическому исследованию. По истечении 6 недель после инфицирования всех животных умерщвляли и проводили макроскопический патологический анализ. При этом у животных не наблюдалось какой-либо потери массы и какого-либо аномального поведения, и аутопсия, проводимая через 6 недель, показала, что все органы были нормальными. Результат теста считался положительным, если после введения rBCG-Pfoвакцины не наблюдалось какого-либо ухудшения состояния животных и если прирост массы животных- 13012182 оставался на нормальном уровне по сравнению с животными, инокулированными родительским штаммом. Одновременно с этим проводили мониторинг уровней бактерий в органах животных. Морских свинок, иммунизованных вакциной, полученной на основе либо родительского, либо рекомбинантного штамма, подвергали эвтаназии через различные промежутки времени после инокуляции, а затем легкие,селезенку и региональные (паховые) лимфоузлы анализировали на уровень КОЕ BCG или rBCG. Тест на токсичность. Для оценки токсичности штаммов rBCG морских свинок, по 12 животных в каждой группе, внутрикожно вакцинировали одной дозой, которая была в 4 раза выше или в 4 раза ниже разовой дозы штаммовrBCG, родительского штамма BCG или физиологического раствора, вводимых человеку, соответственно. На 3-й день после вакцинации шесть животных умерщвляли для оценки кратковременного действия данной вакцины на этих животных. Через 28 дней после вакцинации шесть оставшихся животных умерщвляли для оценки продолжительности действия этих вакцин на животных. В обоих случаях одновременно регистрировали массу тела каждого животного и осуществляли исследование на наличие макропатологий, а также проводили осмотр участков тела в месте инъекции. Затем брали кровь для ее биохимического анализа и проводили гистопатологический анализ внутренних органов и участков тела в месте инъекции. Исследования для определения иммунитета Исследование для определения иммунитета у мышей. Мышей С 57 В 1/6 (самок в возрасте 5-6 недель), по 13 животных в каждой группе, подкожно иммунизировали 106 КОЕ rBCG, родительского BCG или физиологического раствора. Другую группу мышей использовали в качестве нормального контроля. Через 8 недель после иммунизации мышей инокулировали штаммом M.tb Эрдмана (или резистентным к канамицину штаммом H37Rv) путем аэрозольной ингаляции из 10-миллилитровой моноклеточной суспензии, содержащей всего 107 КОЕ штамма для инокуляции, т.е. в дозе, доставляющей 100 живых бактерий в легкие каждого животного, как описано выше. Мониторинг выживаемости экспериментальных животных проводили одновременно с мониторингом выживаемости неинокулированных животных. После инокуляции животных также обследовали на потерю массы и на общее состояние. На 1-й день после инокуляции трех мышей в каждой группе умерщвляли и проводили анализ на уровни КОЕ в легких для подтверждения дозы инокуляции, а одну мышь умерщвляли для проведения гистопатологического анализа селезенки и легких. Через 5 недель после инокуляции девять животных в каждой группе умерщвляли и проводили гистопатологический и микробиологический анализ каждого животного. Ткани легких и селезенки, взятые у шести мышей, оценивали на уровни КОЕ (для идентификации вакцинного штамма от инокулирующего штамма использовали планшеты, в которые добавляли селектируемые маркеры). Если инокуляцию проводили резистентным к канамицину штаммом H37Rv, то для идентификации инокулирующего штамма от вакцинного штамма использовали Kan или ТСН. Если инокуляцию проводили штаммом M.tb Эрдмана, то для идентификации вакцинного штамма от инокулирующего штамма использовали ТСН (BCG является восприимчивым штаммом, а M.tb обычно является резистентным). Индуцирование кожной гиперчувствительности замедленного типа (ГЗТ). Морских свинок, не имеющих каких-либо обнаружимых патогенов (SPF), внутрикожно иммунизировали 103 штамма rBCG или родительского штамма BCG. Через 9 недель после иммунизации спинку животных выбривали и чрескожно инъецировали 10 мкг ППД в 100 мкл забуференного фосфатом физиологического раствора. Через 24 ч измеряли диаметр твердого уплотнения. Штаммы rBCG должны индуцировать ГЗТ, равную или превышающую ГЗТ, индуцируемую родительскими штаммами BCG. Исследование путем инокуляции морских свинок. Для определения эффективности rBCG-вакцин против инокулирующего патогена M.tb морских свинок (молодых взрослых животных SPF Hartley, массой 250-300 граммов, самцов), по 12 животных в группе, иммунизировали (каждым) rBCG, родительским штаммом BCG или физиологическим раствором. Вакцины и контроль вводили внутрикожно в дозе 106 КОЕ. Через 10 недель после иммунизации животных, иммунизованных штаммом rBCG и штаммом BCG, и ложноиммунизованных животных заражали путем ингаляции аэрозоля, содержащего патоген M.tb и генерируемого из 10-миллилитровой моноклеточной суспензии, содержащей всего 107 КОЕ M.tb; и эта процедура позволяет вводить 100 живых бактерий в легкие каждого животного, как описано ранее (Brodin et al., J. Infect Dis. 190(1), 2004). После инокуляции проводили мониторинг выживаемости животных одновременно с мониторингом здоровых животных в группах, которые не были подвергнуты вакцинации и заражению. После инокуляции животных также обследовали на потерю массы и на общее состояние. Через 10 недель после инокуляции шесть животных в каждой группе умерщвляли, а остальные шесть животных в каждой группе через 70 недель после инокуляции подвергали продолжительному обследованию. Одновременно проводили гистопатологический и микробиологический анализ каждого животного. Ткани легких и селезенки подвергали гистопатологическому анализу и анализу на уровни КОЕ (для идентификации вакцинного штамма от инокулирующего штамма использовали планшеты, в которые были добавлены селектируемые маркеры). Если инокуляцию проводили резистентным к канамицину штаммом H37Rv, то для идентификации инокулирующего штамма от вакцинного штамма использовали Kan или ТСН. Если инокуляцию проводили- 14012182 штаммом M.tb Эрдмана, то для идентификации вакцинного штамма от инокулирующего штамма использовали ТСН (BCG является восприимчивым, а M.tb обычно является резистентным). Результаты экспериментов можно считать удовлетворительными в том случае, если ложноиммунизованные животные погибали вскоре после заражения, а rBCG-иммунизованные животные жили дольше, чем животные, иммунизованные родительским штаммом BCG. Исследования безопасности вакцин и картины заражения у приматов. Совсем недавно оценку эффективности вакцины против M.tb. проводили на собакоподобных обезьянах. Эволюционное родство между человеком и другими приматами и аналогичные клинические и патологические симптомы туберкулеза у этих видов делают указанных приматов, не являющихся человеком, привлекательной моделью для экспериментальных исследований заболеваний ТБ и эффективности вакцин против ТБ. Эта модель, характеризующаяся образованием каверн в легких, может быть, очевидно, использована для исследования развития ТБ у человека. За процессом инфицирования и развития заболевания наблюдали путем проведения рентгеновского анализа и оценки потери массы тела, а также путем проведения различных гематологических анализов, включая анализ на скорость оседания эритроцитов (СОЭ),анализ на пролиферацию мононуклеарных клеток периферической крови (МКПК) и продуцирование цитокинов, анализ на активность цитотоксических Т-лимфоцитов (ЦТЛ) и анализ на гуморальные ответы. После инфицирования у собакоподобных обезьян развивалась патология легких с характерными поражениями, и в течение 4-6 месяцев после инфицирования, в зависимости от дозы заражения, животные погибали от острой инфекции дыхательных путей. Более низкие дозы инфицирования могут приводить к возникновению хронических инфекций без развития заболевания, как это часто наблюдается у людей. Протокол исследования. В этом исследовании проводили прямое сравнение различных доз родительского штамма BCG с дозами рекомбинантного BCG, вводимых отдельно, или с последующим введением двух бустер-доз вакцины, содержащей последовательности, сверхэкспрессирующиеся в rBCG-конструкциях. Последние дозы могут быть введены в виде рекомбинантного белка в подходящей адъювантной композиции, в виде ДНК-конструкции или в виде Ad35-конструкций. В первом исследовании оценивали протективную эффективность родительского штамма BCG по сравнению с конструкциями rBCG, вводимыми без последующего применения бустер-инъекций. Это исследование, в котором участвовали три группы животных (по 10 животных в каждой группе), проводили в соответствии со следующим протоколом: одному животному из каждой группы вводили BCG,rBCG и физиологический раствор. Двух животных из каждой группы подвергали кожному тесту на сверхэкспрессию антигенов в rBCG-конструкциях, а также с использованием стандартного ППД и физиологического раствора в качестве контроля. Явные и более крупные уплотнения у мышей rBCGгруппы, наблюдаемые по сравнению с мышами BCG-группы, указывали на то, что вакцина функционировала in vivo и вырабатывала иммунный ответ. Остальных восемь животных из каждой группы инокулировали аэрозолем, содержащим низкую дозу штамма M.tb Эрдмана, а уровень иммунного ответа определяли по снижению бактериальной нагрузки через 16 недель после заражения или по выживаемости как конечной цели исследования. Протокол мониторинга BCG-примирования был, по существу, аналогичен описанному выше, за исключением того, что животных сначала вакцинировали штаммом BCG, штаммом rBCG и/или физиологическим раствором, а затем вводили две бустер-дозы со сверхэкспрессированными антигенами. Исследования иммуногенности и иммунитета у приматов-моделей, не являющихся человеком, проводили в целях изучения иммунобиологических и иммунопатологических аспектов туберкулеза и оценки эффективности rBCG-конструкций у макак. Перед началом эксперимента неполовозрелых животных или молодых взрослых животных, содержащихся в неволе и имеющих среднюю массу 2-3 кг, подвергали длительной адаптации к условиям эксперимента. Исследования путем предварительной инокуляции предусматривали проведение основных анализов крови, которые включают рутинные гематологические анализы, анализы на скорость оседания эритроцитов, а также анализы на пролиферацию лимфоцитов. В качестве предварительной части исследования, т.е. перед инфицированием, проводили кожные тесты с использованием ППД для подтверждения отсутствия у животного чувствительности к туберкулину и рентгеновский анализ грудной клетки. Период иммунизации продолжался всего 21 неделю, в течение которых проводили первичную вакцинацию штаммом BCG или rBCG на неделю 0 и бустер-инъекцию антигенов на недели 12 и 16. Антигенспецифический иммунитет оценивали путем определения уровня пролиферации и секреции интерферона(IFN-) в тестах на стимуляцию лимфоцитов. Уровень продуцирования IFN- лимфоцитами определяли с помощью твердофазного иммуноферментного анализа(ELISPOT) или клеточного сортинга с активацией флуоресценции (FACS). Для этой цели брали пробы крови на недели 0, 4, 8, 12, 16 и 20 после первичной вакцинации. Через 4-6 недель после последней иммунизации животных инокулировали путем интратрахеального введения 3 мл (1000 КОЕ) штамма М. tuberculosis Эрдмана в один и тот же день и одного и того же препарата. Процесс инфицирования оценивали по потере массы, температуре, повышенной СОЭ, РГЗТ в- 15012182 ответ на ППД, пролиферативному ответу in vitro МКПК, стимулированных ППД, и по уровню антигенов,сверхэкспрессируемых в rBCG, с последующим измерением уровней продуцирования IFN-. Для детекции патологий, ассоциированных с туберкулезом легких, проводили рентгеновский анализ грудной клетки и, наконец, через 12-16 недель после инокуляции животных подвергали аутопсии. Клиническая оценка ТБ-векторов и вакцин против ТБ Исследования на безопасность и токсичность. Преклинические исследования на безопасность и токсичность, разрешенные Регуляторными органами, проводили аналогично преклиническим исследованиям на безопасность и токсичность, описанным выше. После этих исследований проводили исследования вакцин на безопасность для человека. Эти исследования сначала проводили с участием здоровых взрослых индивидуумов с отрицательным результатом теста на квантиферон, а затем аналогичные исследования проводили с участием детей и новорожденных. Исследования на иммуногенность. Исследования на иммуногенность у мышей и приматов могут быть проведены стандартными методами оценки клеточного иммунитета, такими как, но не ограничивающимися ими, методы оценки уровней IFN-, ELISPOT и/или проточная цитометрия с кратковременной и длительной стимуляцией антигеном или пептидом. Аналогичные методы были применены для оценки ответов у человека. Для оценкиCD4- и CD8-ответов после вакцинации человека проводили исследования с использованием тетрамеров. Оптимизация стратегий комбинированной иммунизации "прайм-буст". Штамм rBCG является достаточно эффективным при его использовании в качестве отдельно взятой вакцины против ТБ или других заболеваний, против которых он был сконструирован, причем этот штамм был сконструирован так, чтобы он экспрессировал соответствующие трансгены. Используемый здесь термин "трансген" означает ДНК-сегмент, функционально присоединенный к микобактериальному промотору и экспрессирующий нужный белок. Описанный здесь rBCG, служащий в качестве вакцины против ТБ или экспрессирующий трансгены для вырабатывания иммунитета против других заболеваний,также является исключительно эффективным для примирования иммунной системы в целях бустер-иммунизации рекомбинантными белками, смешанными с адъювантами или вирусными или бактериальными векторными антигенами. В преклинических исследованиях на животных и в исследованиях с участием человека первичное введение BCG с последующей бустер-инъекцией рекомбинантного белка/адъюванта или вектора оптимизировали в отношении схемы и дозы введения. Эти"прайм-буст"-стратегии являются наиболее сильным средством для индуцирования иммунитета у человека, что обусловлено эффективностью первичного введения BCG, как, в частности, описано в настоящем изобретении, а также локализацией и усилением бустер-ответа иммунной системы на введение рекомбинантного белка или вектора. Исследование действия терапевтической вакцины на животных после их заражения. Для выявления латентной инфекции использовали мышей C57BL/6, которым вводили терапевтические вакцины на той стадии, когда M.tb-специфический иммунитет, индуцированный низкой дозой инфекции, вырабатывался лишь на очень незначительном уровне и на той стадии, когдаM.tb-специфический иммунитет был пониженным и основную его часть составляли лишь Т-клетки памяти. Затем через 2 и 5 месяцев после введения последней терапевтической вакцины эту терапевтическую вакцину оценивали на терапевтическую эффективность путем подсчета числа КОЕ в легких и селезенках отдельных мышей. Число КОЕ в группах мышей анализировали стандартными статистическими методами и полученные результаты использовали для того, чтобы определить, приводит ли терапевтическая вакцинация к значительному снижению латентной M.tb-инфекции у мышей. Аналогичные методы, если необходимо, могут быть применены для оценки иммунных ответов у других животных. Клиническая оценка BCG-векторов: пероральное введение rBCG-вакцин. Пероральная вакцинация нужного животного штаммом rBCG согласно изобретению может быть проведена описанными ранее методами (Miller et al., Can. Med. Assoc. J. 121(1): 45-54, 1979). Количество перорально вводимого rBCG может варьироваться в зависимости от вида индивидуума, а также от заболевания или состояния, подвергаемого лечению. В общих чертах, используемая доза может содержать примерно 103-1011 жизнеспособных микроорганизмов, а предпочтительно примерно 105-109 жизнеспособных микроорганизмов.rBCG обычно вводят вместе с фармацевтически приемлемым носителем или разбавителем. Выбор какого-либо конкретного фармацевтически приемлемого носителя или разбавителя не имеет решающего значения для осуществления настоящего изобретения. Примерами разбавителей являются забуференный фосфатом физиологический раствор, буфер для забуферивания против действия кислоты желудочного сока, продуцируемого в желудке, такой как цитратный буфер (рН 7,0), содержащий сахарозу, отдельно взятый бикарбонатный буфер (рН 7,0) (Levine et al., J. Clin. Invest., 79: 888-902, 1987; and Black et al., J.Infect. Dis., 155: 1260-1265, 1987), или бикарбонатный буфер (рН 7,0), содержащий аскорбиновую кислоту, лактозу и, необязательно, аспартам (Levine et al., Lancet, II: 467-470, 1988). Примерами носителей яв- 16012182 ляются белки, например, содержащиеся в сепарированном молоке, сахара, например сахароза или поливинилпирролидон. Обычно эти носители используются в концентрации, составляющей примерно 0,1-90% (мас./об.), а предпочтительно, в пределах 1-10% (мас./об.). Пример 1. Эффективность родительских штаммов BCG у морских свинок: исследование иммунитета, вырабатываемого у морских свинок штаммом rBCG 30. В качестве примера исследований, проводимых с использованием имеющейся вакцины rBCG30,могут служить крупномасштабные исследования на морских свинках, целью которых является сравнение защитного действия двух лотов rBCG30 с защитным действие самого родительского штамма BCG Tice и других коммерчески доступных вакцин BCG (штамма SSI-1331), используемых во всем мире для вакцинации людей. Указанные два лота rBCG30, используемые для введения человеку, были получены либо в лабораторных условиях (rBCG30-UCLA), либо в условиях, соответствующих стандартизированным международным требованиям (GMP), (rBCG30-Kit). Морских свинок (10 животных на группу) иммунизировали подкожным введением одной дозы 103 КОЕ каждой из BCG-вакцин. В данное исследование была включена группа негативного контроля(иммунизованная физиологическим раствором). Через 8 недель после вакцинации животных инокулировали вирулентным штаммом Эрдмана с помощью аэрозольного ингалятора после калибровки распылительного клапана аппарата Миддлбрука для воздушно-капельного введения в целях доставки в легкие приблизительно 10-15 бактерий. Животных умерщвляли на 10-ю неделю после заражения. При аутопсии у животных удаляли легкие и селезенку и определяли число жизнеспособных бактерий путем посева серийного 10-кратного разведения гомогенатов доли легких и селезенки на питательную среду Миддлбрука 7 Н 11. Через 21 день после инкубирования при 37 С в атмосфере 5% (об./об.) CO2 подсчитывали образовавшиеся бактериальные колонии. Полученные данные выражали в виде log10 среднего числа выделенных бактерий. Полученные результаты (фиг. 2) показали, что по сравнению с негативным контролем, т.е. контролем с использованием физиологического раствора, все вакцины индуцировали иммунитет против M.tbинфекции, при этом различия между этими вакцинами можно разделить на две группы: 1) rBCG30 (UCLA) и BCG Дания 1331 имели более эффективное защитное действие и 2) BCG (родительский штамм Tice) и rBCG30 (kit) имели менее эффективное защитное действие. Отсюда можно сделать вывод, что вакцина rBCG30 (UCLA), полученная в лабораторных условиях,но не имеющая GMP-чистоту, в лучшем случае индуцировала более высокий уровень иммунитета, чем родительский штамм Tice, либо она продуцировала иммунитет, лишь сравнимый с иммунитетом, вырабатываемым коммерчески доступной вакциной BCG SSI. Поэтому для достижения эффективности, превышающей эффективность вакцины BCG Дания 1331, необходимо получить новую rBCG-вакцину. Пример 2. Конструирование хозяев для их использования в качестве носителей экспрессионных векторов, не содержащих маркеров резистентности к антибиотикам. Конструирование плазмиды для удаления гена leuD в BCG штамме Дания 1331. Левую и правую фланкирующие 1 т.п.н.-области гена leuD объединяли путем ДНК-синтеза (DNA 2,0, СА) с получением 2 т.п.н.-сегмента ДНК с pacI-сайтами на обоих концах. Этот ДНК-фрагмент клонировали в вышеупомянутую плазмиду для аллельного обмена путем расщепления ферментом рестрикции PacI с последующим лигированием и получением плазмиды, не содержащей гена leuD. Инактивация гена leuD путем аллельного обмена. Инактивацию гена leuD осуществляли, как описано выше, за исключением того, что для культивирования штамма, не содержащего гена leuD, в среду добавляли 50 мкг/мл лейцина. Протокол главных стадий этой процедуры приводится на фиг. 4. Подтверждение элиминации гена leuD. Фенотипический тест. Полученный штамм тестировали на зависимость его роста от присутствия лейцина. В частности,бактерии культивировали в среде 7 Н 9 с 10% 0ADC и 0,05% (об./об.) тилоксапола в присутствии или в отсутствии 50 мкг/мл лейцина и мониторинг роста бактерий проводили путем измерения величины оптической плотности OD600. Анализ региональной геномной последовательности. Геномную ДНК сконструированого штамма получали, как описано выше. Для ПЦР-амплификации использовали пары праймеров, комплементарных левому и правому 1 т.п.н.-флангам гена-мишени, и получали фрагмент хромосомы размером приблизительно 2 т.п.н. Этот ПЦР-продукт секвенировали для подтверждения отсутствия гена leuD в указанной области. Пример 3. Сверхэкспрессия антигенов M.tb в штаммах rBCG Модификации ДНК. Антигены M.tb ТВ 10.4 (Rv0288), Ag85B (Rv1886c) и Ag85A (Rv3804c) подвергали полицистронной экспрессии в указанном порядке с использованием промоторов Ag85B плюс Rv3130 (Florczyk et al., см. выше, 2003). ДНК-последовательности, кодирующие пептид, имеющий последовательность KDEL, помещали в конец каждого антигена в качестве сигнала удерживания в эндоплазматическом ретикулуме в целях улучшения презентации каждого антигена. Кроме того, для гарантии стабильности транскриби- 17012182 руемой полицистронной мРНК вводили 5'-петлевую структуру и 3'-последовательности терминации транскрипции. И, наконец, для фланкирования обоих концов в целях облегчения клонирования экспрессионного кластера в экспрессионный вектор использовали последовательности фермента рестрикцииPacI. Все ДНК в экспрессионном кластере получали путем генного синтеза (Picoscript Inc., TX). Экспрессионный кластер клонировали в экспрессионный вектор с использованием PacI-сайтов. После амплификации в E.coli плазмиду расщепляли ферментом NdeI для элиминации oriE и гена резистентности к канамицину, а затем лигировали с получением однонаправленной челночной системы, которую затем вводили в ауксотрофный по leuD мутант Mycobacterium в соответствии со стандартными процедурами электропорации (Parish et al., Microbiology, 145: 3497-3503, 1999). На фиг. 3 схематически представлен пример экспрессионного вектора, не содержащего гена резистентности к антибиотикам. Экспрессия антигенов M.tb с использованием системы отбора без применения антибиотиков. Аутотрофный по лейцину штамм BCG Дания 1331, используемый в качестве хозяина для системы отбора без применения антибиотиков, культивировали в среде 7 Н 9, в которую были добавлены 10%OAD (олеиновая кислота-альбумин-декстроза-каталаза), 0,05% (об./об.) тилаксопола и 50 мкг/мл лейцина. После электропорации рекомбинантные штаммы, содержащие антигенэкспрессирующую плазмиду,выделяли путем культивирования на планшетах со средой 7 Н 10 (BD Difco), не содержащей лейцин. Выживаемость бактерий зависит от присутствия антигенэкспрессирующего гена leuD в данной плазмиде,который, в свою очередь, функционирует как механизм сохранения плазмиды в клетках. Полученные отдельные колонии выделяли и культивировали в среде 7 Н 9, как описано выше, за исключением того,что в эту среду не добавляли лейцин. Подтверждение экспрессии. Экспрессию каждого антигена детектировали с помощью вестерн-блот-анализа. В частности, супернатант культуры собирали и обрабатывали, как описано ранее (Harth et al., Infect. Immun. 65(6): 23212328, 1997). Затем экспрессируемые антигены выделяли с помощью электрофореза в ДСН-ПААГ и осуществляли блоттинг с помощью антител против антигенов 85 А, 85 В и 10.4. Уровень экспрессии каждого антигена определяли путем количественной оценки интенсивности каждой специфической полосы по сравнению с полосой, продуцируемой бактериальным хозяином, не содержащим экспрессионной плазмиды. Пример 4. Экспрессия выбранных антигенов в микобактериях без применения отбора на резистентность к антибиотикам. Материалы и методы. Плазмидный и микобактериальный препарат для электропорации. Рекомбинантную плазмидную ДНК выделяли и расщепляли рестриктирующей эндонуклеазой NdeI(New England Biolabs) в целях высвобождения маркера для отбора на резистентность к антибиотикам(например, резистентность к канамицину) и области ориджина репликации (oriE) E.coli. Затем расщепленный фрагмент плазмидной ДНК подвергали циркуляризации с использованием ДНК-лигазы Т 4 (NewEngland Biolabs) в соответствии с инструкциями производителя. Полученную плазмиду, содержащую ориджин репликации Mycobacterium и выбранные антигены, но не содержащую гена резистентности к антибиотиками и не способную реплицироваться в грамотрицательных бактериях, вводили в выбраннуюMycobacterium. В целях приготовления Mycobacterium для электропорации бактерии штамма BCG Дания 1331 культивировали при 37 С до достижения экспоненциальной фазы роста в среде Миддлбрука 7 Н 9(BD Biosciences), в которую добавляли 10% OADC. Для диспергирования бактерий использовали тилоксапол (0,05% об./об., Research Diagnostics Cat.70400). Клетки три раза промывали 10% глицерином + 0,05% тилоксаполом для удаления культуральной среды, а затем проводили электропорацию. Для каждой процедуры электропорации использовали 1,6 мкг плазмиды на 1,25108 бактериальных клеток. Электропорацию проводили при 2,2 кВ, емкости 25 мкФ и сопротивлении 1,0 кОм. После электропорации клетки непосредственно высевали на планшеты с агаром Миддлбрука 7 Н 10 (BD Biosciences) в 10-кратных серийных разведениях и инкубировали при 37 С. ПЦР-скрининг бактериальных колоний, содержащих экспрессионную плазмиду. Рекомбинантные штаммы сначала скринировали с помощью ПЦР с использованием прямого праймера GTTAAGCGACTCGGCTATCG (SEQ ID NO: 1) и обратного праймера ATGCCACCACAAGCACTACA (SEQ ID NO: 2) для амплификации ДНК-последовательности области oriM в экспрессионной плазмиде. ПЦР осуществляли в следующем режиме: стадия 1: при 95 С, 4 мин один цикл; стадия 2: при 95 С, 1 мин, при 60 С, 1 мин, а затем при 72 С, 1 мин, всего 30 циклов; стадия 3: при 72 С, 10 мин, один цикл; стадия 4: хранение при 4 С. Полученные ПЦР-продукты анализировали с помощью электрофореза в агарозном геле для подтверждения присутствия плазмиды в клетках.- 18012182 Результаты. ПЦР, которая была разработана для амплификации области репликации (oriM) указанной плазмиды, проводили в целях скрининга полученных колоний на присутствие сверхэкспрессирующей плазмиды. Поскольку эта область отсутствовала на бактериальной хромосоме, то присутствие этой области в клетках со всей очевидностью указывает на то, что такая плазмида была включена в клетки. Из всех скринированных колоний rBCG некоторые колонии продуцировали ПЦР-продукт, который по своему размеру был аналогичен положительному по данной плазмиде контрольному продукту, как было проанализировано с помощью гель-электрофореза. В противоположность этому родительские бактерии BCG не продуцировали какого-либо ПЦР-продукта, как показано на фиг. 5. Этот эксперимент показал, что данная плазмида была успешно введена в Mycobacterium и что бактериальный клон, содержащий указанную плазмиду, был выделен без отбора на резистентность к антибиотикам. Обсуждение. Стандартные плазмиды, используемые в рекомбинантных штаммах Mycobacterium, содержат область репликации и селектируемый маркер (обычно ген резистентности к антибиотикам, например ген резистентности к канамицину или ген, который комплементирует метаболический дефект, напримерleuD или asd (Galan et al., Gene, 94: 29, 1990) в качестве главных плазмидных элементов, которые используются в экспериментах с применением техники рекомбинантных ДНК. Обычно для отбора клонов, содержащих рекомбинантные плазмиды, используются антибиотики. Однако это может приводить к риску нежелательноого распространения генов резистентности к антибиотикам в тех случаях, когда резистентный к антибиотику генетически модифицированный микроорганизм предназначен для использования за пределами лабораторий, где не могут быть предприняты адекватные меры предосторожности. В вышеописанном исследовании в бактерии согласно изобретению была введена рекомбинантная плазмида, которая обладала способностью экспрессировать антиген, но которая не содержала областиoriE и селектируемого маркера резистентности к антибиотику, и были успешно выделены клоны, содержащие указанную плазмиду, без применения отбора. Хотя в настоящем эксперименте в качестве области репликации плазмиды используется oriM, однако в этой связи следует отметить, что в качестве замены могут служить и другие области репликации плазмиды, такие как область репликации pMF1 (Bachrach etal., Microbiol., 146: 297, 2000). Уникальное преимущество этой системы заключается в том, что указанная рекомбинантная плазмида больше не содержит гена резистентности к антибиотикам. Таким образом, это позволяет избежать случайного распространения генов резистентности к антибиотикам в окружающую среду, как это обычно происходит в случае использования стандартных экспрессионных плазмид. Кроме того, экспрессионная плазмида, представляющая собой однонаправленный челночный вектор согласно изобретению, больше не способна реплицироваться в широком круге хозяев, поскольку из нее были удалены генетические элементы, ответственные за такую репликацию. Такое ограничение сообщает указанной рекомбинантной плазмиде второй уровень защиты, что значительно снижает риск, связанный с высвобождением генетически модифицированного (ГМО) микроорганизма в окружающую среду. Хотя имеющиеся результаты показали, что рекомбинантные плазмиды можно вводить в аттенюированные штаммы Mycobacterium без проведения отбора, однако следует отметить, что в стабильности плазмиды, не содержащей селектируемого маркера, в Mycobacterium, определенную роль могут играть и другие факторы. Так, например, область репликации содержит гены, облегчающие репликацию плазмиды и опосредующие сегрегацию плазмиды в клетках-сибсах, что, тем самым, дает возможность идентифицировать клоны, содержащие указанную плазмиду, без проведения отбора. Возможным фактором, позволяющим осуществлять выделение клонов, содержащих указанную плазмиду, без проведения отбора, является то, что в настоящем подходе используется более высокое отношение плазмиды к клетке. Использование более высокого отношения плазмиды к клетке приводит к увеличению вероятности того, что клетка будет поглощать данную плазмиду, и к снижению числа клеток, не содержащих эту плазмиду. В этом исследовании отношение плазмиды к клетке примерно в 10 раз превышает отношение, которое обычно используется в стандартных методах с применением системы отбора. Теоретически, чем больше отношение плазмиды к клетке, тем больше клонов будет содержать данную плазмиду, но это имеет место до тех пор, пока не будет достигнуто насыщение клетки данной плазмидой, после чего может происходить ингибирование поглощения плазмидной ДНК клетками, подвергнутыми электропорации. В предпочтительных вариантах осуществления изобретения отношение плазмиды к бактериям составляет примерно от 0,5 до 10 мкг плазмидной ДНК к 1,25108 бактериальных клеток, а предпочтительно примерно от 1 до 5 мкг плазмидной ДНК к 1,25108 бактериальных клеток. Кроме того, антигены ТБ, которые сверхэкспрессируются плазмидой pAF105, могут играть важную роль в стабилизации плазмиды. Эта плазмида сверхэкспрессирует два белка антигенного комплекса 85(Ag85A (Rv3804c) и Ag85 В (Rv1886c, которые обладают миколилтрансферазной активностью, необходимой для биосинтеза димиколата трегалозы, доминантной структуры, необходимой для сохранения целостности клеточной стенки. Поэтому возможно, что сверхэкспрессия по меньшей мере одного из этих антигенов может обеспечивать стабильность плазмиды, если она не содержит селектируемого маркера,при создании условий, благоприятствующих преимущественному продуцированию этих антигенов в- 19012182 клетках, содержащих такую плазмиду, что будет облегчать идентификацию клонов, содержащих данную плазмиду, без проведения отбора. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Трансформированная бактерия или ее потомство, которые включают нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная нуклеотидная последовательность присутствует в плазмиде, и где указанная нуклеотидная последовательность кодирует белок, необходимый для выживания, и где ген, кодирующий указанный необходимый для выживания белок, делетирован из бактериального генома указанной трансформированной бактерии. 2. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде, и где указанная плазмида содержит ген, кодирующий высвобождение из эндосомы, и где указанный ген, кодирующий высвобождение из эндосомы, может представлять собой чужеродную нуклеотидную последовательность, которая присутствует в указанной плазмиде или может присутствовать в указанной плазмиде в дополнение к чужеродной нуклеотидной последовательности, присутствующей в указанной плазмиде. 3. Трансформированная бактерия или ее потомство по п.2, где указанный ген, кодирующий высвобождение из эндосомы, представляет собой ген перфринголизина О (pfo) из Clostridium perfringens. 4. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде и где указанная чужеродная нуклеотидная последовательность кодирует антиген 85A, антиген 85B или антиген 85A/85B. 5. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде и где указанная плазмида содержит ген, кодирующий белки, которые сохраняют и/или стабилизируют плазмиду. 6. Трансформированная бактерия или ее потомство по п.5, где указанный ген, кодирующий белки,кодирует антиген 85A, антиген 85B или антиген 85A/85B. 7. Трансформированная бактерия или ее потомство по п.2, где указанной бактерией является Mycobacterium. 8. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, где указанная чужеродная нуклеотидная последовательность кодирует апоптоз. 9. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде, и где указанная плазмида содержит ген, кодирующий апоптоз. 10. Трансформированная бактерия или ее потомство, которые включают чужеродную нуклеотидную последовательность, реплицирующуюся и экспрессирующуюся в указанной трансформированной бактерии или в ее потомстве, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером и где указанная чужеродная нуклеотидная последовательность не может реплицироваться в грамотрицательных бактериях. 11. Трансформированная бактерия или ее потомство по п.1, где указанной трансформированной бактерией является ауксотрофная бактерия. 12. Трансформированная бактерия или ее потомство по п.1, где указанная чужеродная нуклеотидная последовательность является по меньшеймере частью однонаправленного челночного вектора. 13. Способ трансформирования бактерии, включающий стадию введения нуклеотидной последовательности, которая реплицируется и экспрессируется в указанной бактерии, где указанная нуклеотидная последовательность не связана с селектируемым маркером, и где указанная нуклеотидная последовательность присутствует в плазмиде, где указанная нуклеотидная последовательность кодирует белок,необходимый для выживания, и где ген, кодирующий указанный необходимый для выживания белок,- 20012182 делетирован из бактериального генома указанной бактерии. 14. Способ по п.13, где указанную стадию введения осуществляют путем электропорации. 15. Способ по п.13, где указанная нуклеотидная последовательность является по меньшей мере частью однонаправленного челночного вектора. 16. Трансформированная Mycobacterium или ее потомство, содержащие чужеродную нуклеотидную последовательность, которая кодирует представляющий интерес белок, и где указанная трансформированная Mycobacterium включает плазмиду, которая не способна реплицироваться в грамотрицательных бактериях. 17. Трансформированная Mycobacterium или ее потомство по п.16, где указанная чужеродная нуклеотидная последовательность является частью плазмиды. 18. Трансформированная Mycobacterium или ее потомство по п.17, где указанная плазмида не содержит селектируемого маркера. 19. Трансформированная Mycobacterium или ее потомство по п.17, где указанный белок является необходимым для выживания и соответствует белку выживания, который кодируется нуклеотидной последовательностью, делетированной из бактериального генома указанной трансформированной микобактерии. 20. Трансформированная Mycobacterium или ее потомство по п.19, где указанным белком являетсяleuD. 21. Трансформированная Mycobacterium или ее потомство по п.19, дополнительно содержащие промоторные последовательности, которые активируются in vivo. 22. Трансформированная Mycobacterium или ее потомство по п.16, где указанная трансформированная Mycobacterium является аттенюированной. 23. Трансформированная Mycobacterium или ее потомство по п.22, где указанной трансформированной Mycobacterium является BCG. 24. Трансформированная Mycobacterium или ее потомство по п.23, где указанный BCG выбран из нижеследующих штаммов: BCG1331, BCG Пастер, BCG Токио и BCG Копенгаген. 25. Вакцина, включающая трансформированную Mycobacterium или ее потомство, содержащие чужеродную нуклеотидную последовательность, которая кодирует представляющий интерес ген, и где указанная трансформированная Mycobacterium включает плазмиду, которая не способна реплицироваться в грамотрицательных бактериях. 26. Трансформированная Mycobacterium по п.16, где указанная трансформированная Mycobacterium не обладает резистентностью к антибиотикам. 27. Трансформированная Mycobacterium по п.16, где указанная трансформированная Mycobacterium является ауксотрофной. 28. Вакцина по п.25, где указанная трансформированная Mycobacterium не обладает резистентностью к антибиотикам. 29. Вакцина по п.25, где указанная трансформированная Mycobacterium является ауксотрофной. 30. Способ трансформирования бактерии, включающий стадию введения чужеродной нуклеотидной последовательности, которая реплицируется и экспрессируется в указанной бактерии, где указанная чужеродная нуклеотидная последовательность не связана с селектируемым маркером и где указанная чужеродная нуклеотидная последовательность присутствует в плазмиде, где указанная чужеродная нуклеотидная последовательность не может реплицироваться в грамотрицательных бактериях.
МПК / Метки
МПК: A61K 49/00, C12N 1/00, C07K 1/00, A61K 39/00, A61K 39/04, C12N 1/12, C12N 15/74, A61K 39/38, C07H 21/02, A61K 39/02, C12N 15/00
Метки: микобактерий, антигенов, сверхэкспрессия, микобактерии, электропорация
Код ссылки
<a href="https://eas.patents.su/24-12182-elektroporaciya-mikobakterii-i-sverhekspressiya-antigenov-mikobakterijj.html" rel="bookmark" title="База патентов Евразийского Союза">Электропорация микобактерии и сверхэкспрессия антигенов микобактерий</a>
Производные хинолина и их применение в качестве ингибиторов микобактерий

Номер патента: 11277
Опубликовано: 27.02.2009
Авторы: Ланкуа Давид Франсис Ален, Паскье Элизабет Тереза Жанна, Гийемон Жером Эмиль Жорж
МПК: C07D 215/14, A61K 31/4709, A61K 31/47...
Метки: качестве, производные, ингибиторов, применение, хинолина, микобактерий
Формула / Реферат:
1. Соединение общей формулы (Ia) или общей формулы (Ib) его фармацевтически приемлемые кислотно- или основно-аддитивные соли, четвертичные амины, стереохимически изомерные формы, таутомерные формы и N-оксиды, где R1 означает водород, галоген, Ar, Het, С1-6алкил; p означает целое число, равное 1, 2 или 3; R2 означает водород; алкил; гидрокси; тио; алкилокси, необязательно замещенный аминогруппой, моно- или ди(алкил)аминогруппой или радикалом...
Фармацевтические композиции, усиливающие иммуногенность антигенов со слабой иммуногенностью

Номер патента: 5138
Опубликовано: 30.12.2004
Авторы: Санчес Рамирес Белинда, Де Леон Дельгадо Хоэль, Де Ла Баррера Айра Анабель, Перес Родригес Роландо, Диас Родригес Йильдиан, Меса Пардильо Сирсе, Суарес Пестана Эдуардо Рауль, Фернандес Молина Луис Энрике
МПК: A61P 37/04, A61K 39/385
Метки: фармацевтические, композиции, антигенов, усиливающие, иммуногенностью, слабой, иммуногенность
Формула / Реферат:
1. Фармацевтическая композиция, усиливающая иммуногенность антигенов со слабой иммуногенностью, содержащая (A) один или более антигенов со слабой иммуногенностью; (B) один вакцинный носитель, заключающий в себе протеолипосомы, полученные из белкового комплекса наружной мембраны штамма грамотрицательных бактерий (Neisseria meningitidis), который содержит включенные в него ганглиозиды; и (C) возможно, один или более адъювантов. 2. Композиция по...
Способ получения противоопухолевой вакцины на основе поверхностных антигенов эндотелиальных клеток

Номер патента: 9327
Опубликовано: 28.12.2007
Автор: Лохов Петр Генриевич
МПК: A61K 39/00, A61K 38/39
Метки: клеток, способ, поверхностных, эндотелиальных, антигенов, вакцины, основе, получения, противоопухолевой
Формула / Реферат:
1. Способ получения противоопухолевой вакцины с использованием эндотелиальных клеток, отличающийся тем, что живые эндотелиальные клетки подвергают витальному для клеток воздействию протеазы, отбирают высвободившиеся поверхностные антигены, при этом обработку живых эндотелиальных клеток протеазой повторяют с интервалами, необходимыми для восстановления клетками поверхностных антигенов, аккумулируют поверхностные антигены до достижения их дозы,...
Предыдущий патент: Соединения тиадиазола и их применение