Полипептиды и пути биосинтеза
Номер патента: 9284
Опубликовано: 28.12.2007
Авторы: Хикс Паула М., Даллуге Джозеф, Жао Лишэн, Макфарлан Сара С., Абрахам Тимоти В., Камерон Дуглас С., Вейнер Давид П., Миллис Джим, Ховсон Рассел Дж., Розацца Джон





























Формула / Реферат
1. Способ получения монатина, включающий превращение глюкозы в монатин.
2. Способ получения монатина, включающий превращение триптофана и/или индол-3-молочной кислоты в монатин, причем этот способ включает
контакт триптофана или индол-3-молочной кислоты с первым полипептидом, где первый полипептид превращает триптофан или индол-3-молочную кислоту в индол-3-пируват;
контакт индол-3-пирувата со вторым полипептидом и источником трехатомного (С3) С3-углерода, где второй полипептид превращает индол-3-пируват и источник С3-углерода в 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту, и
контакт 2-гидрокси-2-(индол-3-илметил-4-кетоглутаровой кислоты с третим полипептидом, где третий полипептид превращает 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту в монатин.
3. Способ по п.2, отличающийся тем, что первый полипептид выбирают из группы, включающей триптофан аминотрансферазу (ЕС 2.6.1.27), триптофан дегидрогеназу (ЕС 1.4.1.19), тирозин(ароматическую) аминотрансферазу (ЕС 2.6.1.5), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), аспартат аминотрансферазу (ЕС 2.6.1.1), триптофан оксидазу, оксидазу L-аминокислоты (ЕС 1.4.3.2), дегидрогеназу D-аминокислот (ЕС 1.4.99.1), оксидазу D-аминокислот (ЕС 1.4.3.3.), D-триптофан аминотрансферазу, D-аланин аминотрансферазу (ЕС 2.6.1.21), глутамат дегидрогеназу (ЕС 1.4.1.2-4), фенилаланин дегидрогеназу (ЕС 1.4.1.20) или комбинацию этих полипептидов, и первый полипептид превращает триптофан в индол-3-пируват.
4. Способ по п.2, отличающийся тем, что первый полипептид выбирают из группы, включающей индол-лактат дегидрогеназу (ЕС 1.1.1.110), R-4-гидроксифенил-лактат дегидрогеназу (ЕС 1.1.1.222), 3-(4)-гидроксифенилпируват редуктазу (ЕС 1.1.1.237), лактат дегидрогеназу (ЕС 1.1.1.27, ЕС 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил)лактат дегидрогеназу (ЕС 1.1.1.111) и лактат оксидазу (ЕС 1.1.3-) или комбинацию этих полипептидов, и первый полипептид превращает индол-3-молочную кислоту в индол-3-пируват.
5. Способ по п.2 или 3, отличающийся тем, что первый и/или третий полипептид включает
аминокислотную последовательность, показанную среди SEQ ID ь 2, 4, 6, 8, 10, 12, 14, 32, ь NP-388848.1 по Каталогу банка генов, ь ZP00005082.1 по Каталогу банка генов или ь ААС74014.1 по Каталогу банка генов;
аминокислотную последовательность, не менее чем на 90% идентичную последовательности, показанной среди SEQ ID ь 2, 4, 6, 8, 10, 12, 14, 32, ь NP-388848.1 по Каталогу банка генов, ь ZP00005082.1 по Каталогу банка генов или ь ААС74014.1 по Каталогу банка генов; или аминокислотную последовательность, которая отличается от SEQ ID ь 2, 4, 6, 8, 10, 12, 14, 32, ь NP-388848.1 по Каталогу банка генов, ь ZP00005082.1 по Каталогу банка генов или ь ААС74014.1 по Каталогу банка генов менее чем на 50 консервативных аминокислотных замен, где аминокислотная последовательность превращает триптофан в индол-3-пируват.
6. Способ по в п. 2, отличающийся тем, что второй полипептид представляет собой ЕС 4.1.2.- или ЕС 4.1.3.0.
7. Способ по п.6, отличающийся тем, что фермент ЕС 4.1.3.-представлен 4-гидрокси-2-оксоглутаратглиоксилат-лиазой (ЕС 4.1.3.16), 4-гидрокси-4-метил-2-оксоглутарат пируват-лиазой (ЕС 4.1.3.17) или комбинацией этих ферментов.
8. Способ по п.2, отличающийся тем, что третий полипептид выбирают из группы, включающей тирозин(ароматическую) аминотрансферазу (ЕС 2.6.1.5), триптофан-дегидрогеназу (ЕС 1.4.1.19), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), триптофан-аминотрансферазу (ЕС 2.6.1.27), аспартат аминотрансферазу (ЕС 2.6.1.1), D-аланин аминотрансферазу (ЕС 2.6.1.21), триптофан оксидазу, оксидазу L-аминокислоты (ЕС 1.4.3.2), дегидрогеназу D-аминокислот (ЕС 1.4.99.1), оксидазу D-аминокислот (ЕС 1.4.3.3.), D-триптофан аминотрансферазу, глутамат дегидрогеназу (ЕС 1.4.1.2-4), фенилаланин дегидрогеназу (ЕС 1.4.1.20), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), дегидрогеназу D-аминокислот (ЕС 1.4.99.1) или комбинацию этих полипептидов.
9. Способ по п.2, отличающийся тем, что первый полипептид включает оксидазу аминокислоты (ЕС 1.4.3-) и каталазу (ЕС 1.11.1.6) и что первый полипептид превращает триптофан в индол-3-пируват.
10. Способ по п.9, отличающийся тем, что оксидаза аминокислоты представлена триптофан-оксидазой.
11. Способ по п.2, отличающийся тем, что источник С3-углерода выбирают из группы, которая включает пируват, фосфоенолпируват, аланин, серии, цистеин или комбинацию этих соединений.
12. Способ по п.11, отличающийся тем, что пируват получают из аланина и полипептида, способного осуществлять трансаминирование аланина; серина и полипептида, способного осуществлять бета-элиминацию серина; аспартата и полипептида, способного вступать в бета-лиазные реакции с дикарбоновой аминокислотой; лактата и лактат оксидазы (ЕС 1.1..3-) или лактат дегидрогеназы (ЕС 1.1.1.27, 1.1.1.28, 1.1.1.2.3) или комбинации этих соединений.
13. Способ по п.2, отличающийся тем, что способ далее включает восстановление пероксида водорода в количестве, образовавшемся при превращении триптофана в индол-3-пируват, при контакте между пероксидом водорода и каталазой.
14. Способ по п.2, отличающийся тем, что второй полипептид включает аминокислотную последовательность, которую выбирают из группы, включающей:
- последовательность SEQ ID 66, ь САС46344 по Каталогу банка генов, ь CAB14127.1 по Каталогу банка генов, ь ААС74920.1 по Каталогу банка генов или ь САС47463.1 по Каталогу банка генов;
аминокислотную последовательность, не менее чем на 90% идентичную последовательности SEQ ID 66, ь САС46344 по Каталогу банка генов, ь CAB14127.1 по Каталогу банка генов, ь ААС74920.1 по Каталогу банка генов ь САС47463.1 по Каталогу банка генов или
аминокислотную последовательность, которая отличается от последовательности SEQ ID 66, ь САС46344 по Каталогу банка генов, ь САВ14127.1 по Каталогу банка генов, ь ААС74920.1 по Каталогу банка генов или ь САС47463.1 по Каталогу банка генов, менее чем на 50 консервативных аминокислотных замен, где аминокислотная последовательность превращает индол-3-пирувати пируват в ПМ.
15. Способ по п.8, отличающийся тем, что третий полипептид включает аспартат-аминотрансферазу и 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровая кислота контактирует с аспартатом с образованием оксалацетата и монатина.
16. Способ по п.15, отличающийся тем, что оксалацетат далее контактирует с декарбоксилазой.
17. Способ по п.2, отличающийся тем, что третий полипептид представлен лизин-эпсилон-аминотрансферазой и 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровая кислота далее контактирует с лизином.
18. Способ по п.2, отличающийся тем, что третий полипептид представлен ферментом, осуществляющим восстановительное аминирование и способным к рециклизации НАД(Ф)Н.
19. Способ по п.18, отличающийся тем, что фермент, осуществляющий восстановительное аминирование выбран из группы, включающей глутамат дегидрогеназу, фенилаланин дегидрогеназу, триптофан дегидрогеназу, или представлен комбинацией этих соединений.
20. Способ по п.18, отличающийся тем, что полипептид, способный к рециклизации НАД(Ф)Н, далее продуцирует летучий продукт.
21. Способ по п.20, отличающийся тем, что полипептид, способный к рециклизации и продуцированию летучего продукта, представлен формиат дегидрогеназой и образующийся летучий продукт представляет собой диоксид углерода.
22. Способ по п.2, отличающийся тем, что способ включает превращение индол-3-молочной кислоты в монатин.
23. Способ по п.2, отличающийся тем, что монатин образуется в количестве не менее 10 г/л.
24. Способ получения монатина, включающий превращение индол-3-пирувата в монатин и взаимодействие индол-3-пирувата с полипептидом.
25. Способ получения монатина, включающий превращение триптофана в монатин.
26. Клетка, продуцирующая монатин, где клетка включает по крайней мере одну экзогенную последовательность нуклеиновой кислоты, кодирующей по крайней мере один полипептид, и этот по крайней мере один полипептид осуществляхт превращение первого промежуточного продукта по пути получения монатина с образованием второго промежуточного продукта или монатина.
27. Клетка по п.26, отличающаяся тем, что по крайней мере один полипептид выбран из группы, которая включает триптофан аминотрансферазу (ЕС 2.6.1.27), триптофан дегидрогеназу (ЕС 1.4.1.19), тирозин (ароматическую) аминотрансферазу (ЕС 2.61.5), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), триптофан оксидазу, оксидазу L-аминокислот (ЕС 1.4.3.2), аспартат аминотрансферазу (ЕС 2.6.1.1), дегидрогеназу D-аминокислот (ЕС 1.4.99.1), оксидазу D-аминокислот (ЕС 1.4.3.3), D-аланин аминотрансферазу (ЕС 2.6.1.21), D-триптофан аминотрансферазу, индоллактат дегидрогеназу (ЕС 1.1.1.110), R-4-гидроксифениллактат дегиидрогеназу (ЕС 1.1.1.222), 3-(4)-гидроксифенилпируват редуктазу (ЕС 1.1.1.237), лактат дегидрогеназу (ЕС 1.1.1.27, 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил) лактат дигидрогеназу (ЕС 1.1.1.111), лактат оксидазу (ЕС 1.1.3.-), синтазу/лиазу (4.1.2.-), синтазу/лиазу (4.1.3.-), фенилаланин дегидрогеназу (ЕС 1.4.1.20), глутамат дегидрогеназу (ЕС 1.4.1.2., 1.4.1.3., 1.4.1.4) и комбинацию этих соединений.
28. Клетка по п.27, отличающаяся тем, что по крайней мере один полипептид выбран из группы, которая включает триптофан аминотрансферазу (ЕС 2.6.1.27), триптофан дегидрогеназу (ЕС 1.4.1.19), тирозин (ароматическую) аминотрансферазу (ЕС 2.6.1.5), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), аспартат аминотрансферазу (ЕС 2.6.1.1), триптофан оксидазу, оксидазу L-аминокислот (ЕС 1.4.3.2), дегидрогеназу D-аминокислот (ЕС 1.4.99.1), оксидазу D-аминокислот (ЕС 1.4.3.3), D-аланин аминотрансферазу (ЕС 2.6.1.21), D-триптофан аминотрансферазу, синтазу/лиазу (4.1.3.-), синтазу/лиазу (4.1.2.-), фенилаланин дегидрогеназу (ЕС 1.4.1.20), глутамат дегидрогеназу (ЕС 1.4.1.2., 1.4.1.3., 1.4.1.4) и комбинацию этих соединений.
29. Клетка по п. 27, отличающаяся тем, что по крайней мере один полипептид представлен активностью, выбранной из группы, которая включает индоллактат дегидрогеназу (ЕС 1.1.1.110), R-4-гидроксифениллактат дегидрогеназу (ЕС 1.1.1.222), 3-(4)-гидроксифенилпируват редуктазу (ЕС 1.1.1.237), лактат дегидрогеназу (ЕС 1.1.1.27, 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил) лактат дигидрогеназу (ЕС 1.1.1.111), лактат оксидазу (ЕС 1.1.3.-), синтазу/лиазу (4.1.2.-), синтазу/лиазу (4.1.3.-), триптофан дегидрогеназу (ЕС 1.4.1.19), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), триптофан аминотрансферазу (ЕС 2.6.1.27), тироизин (ароматическую) аминотрансферазу (ЕС 2.6.1.5), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), аспартат аминотрансферазу (ЕС 2.6.1.1), глутамат дегидрогеназу (ЕС 1.4.1.2., 1.4.1.3., 1.4.1.4), фенилаланин дегидрогеназу (ЕС 1.4.1.20), D-триптофан аминотрансферазу, дегидрогеназу D-аминокислот (ЕС 1.4.99.1), D-аланин аминотрансферазу (ЕС 2.6.1.21), и комбинацию этих соединений.
30. Клетка по п.27, отличающаяся тем, что по крайней мере один полипептид является синтаза/лиазой (ЕС 4.1.3.-), включающей 4-гидрокси-2-оксоглутарат альдолазу (ЕС 4.1.3.16) или 4-гидрокси-4-метил-2-оксоглутарат альдолазу (ЕС 4.1.3.17).
31. Клетка по любому из пп.26-30, отличающаяся тем, что клетка представлена бактериальной клеткой.
32. Клетка по любому из пп.26-30, отличающаяся тем, что клетка представлена дрожжевой клеткой.
33. Клетка по п.27, отличающаяся тем, что клетка представлена одним или несколькими полипептидами, кодирующими по крайней мере одну экзогенную последовательность нуклеиновой кислоты и одним или более полипептидом, причем клетка дополнительно включает активности, выбранные из группы, которая включает каталазу, оксалацатат декарбоксилазу, лизин эписилон аминотрансферазу и формиат дегидрогеназу.
34. Полипептид, включающий аминокислотную последовательность, выбранной из группы, которая включает Последовательность ь Р00913 по Каталогу банка генов; ь NP_290344 по Каталогу банка генов; ь CAA34096 по Каталогу банка генов; ь NP_246359 по Каталогу банка генов; ь AAB96579 по Каталогу банка генов; ь NP_232561 по Каталогу банка генов; ь Q59342 по Каталогу банка генов; ь P28796 по Каталогу банка генов; ь 1AX4_4 по Каталогу банка генов; ь ВАА34638 по Каталогу банка генов; ь Р31014 по Каталогу банка генов; Р31015 по Каталогу банка генов; ь NP_280989 по Каталогу банка генов; ь AAC33284 по Каталогу банка генов; ь NP_147838 по Каталогу банка генов; AAF63441 по Каталогу банка генов; ь AAA24679 по Каталогу банка генов, где аминокислотные остатки в положениях 103 и 299 по отношению к Последовательности ь NP_418164 были изменены, для того чтобы расширить специфичность полипептида и допустить субстраты, представленные дикарбоновыми аминокислотами.
35. Полипептид по п.34, отличающийся тем, что замена аминокислоты выбрана из группы, которая включает замену аргинина 103 на треонин; замену аргинина 103 на треонин и валина 299 на аргинин и валина 299 на аргинин.
36. Полипептид, включающий аминокислотную последовательность, выбранную из группы, которая включает Последовательность ь 2TPL_A по Каталогу банка генов; ь Р31012 по Каталогу банка генов; ьP31013, А49493 по Каталогу банка генов; ь Р31011 по Каталогу банка генов; ь ААВ24234 по Каталогу банка генов; ь 1TPL_A по Каталогу банка генов; ь 1TPL_B по Каталогу банка генов;
ь NP245748 по Каталогу банка генов; ь AAB41499 по Каталогу банка генов; ь T45297 по Каталогу банка генов; ь AAA71928, где аминокислотные остатки в положениях 100 и 283 по отношению к Последовательности ь X66978 были изменены для того чтобы расширить специфичность полипептида и допустить субстраты, представленные дикарбоновыми аминокислотами.
37. Полипептид по п.36, отличающийся тем, что замена аминокислоты выбрана из группы, группы, которая включает замену аргинина 100 на треонин; замену аргинина 100 на треонин и валина 283 на аргинин и валина 283 на аргинин.
38. Полипептид по п.34 или 36, отличающийся тем, что субстрат представлен аспартатом.
39. Способ получения монатина, включающий реакцию между по крайней мере одним производным аланина с блокированными карбоксильной и аминной группой и по крайней мере одним производным индол-3-пирувата с блокированной карбоксильной группой.
40. Способ получения монатина по п. 39, отличающийся тем, что производное аланина представлено 3-хлороаланином или 3-бромоаланином и что индол-3-пируват вступает в реакцию Гриньяра или реакцию с литий-органическим катализатором с производным аланина.
41. Способ получения монатина, включающий превращение субстрата, выбранного из группы, которая включает триптофан, индол-3-молочную кислоту, индол-3-пируват и 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту, в один или несколько промежуточных продуктов при получении промежуточных продуктов или монатина.
42. Способ по п.41, отличающийся тем, что по крайней мере один промежуточный продукт подвергается превращению посредством химической реакции.
43. Способ по п. 41, отличающийся тем, что по крайней мере один промежуточный продукт, полученный при получении промежуточного продукта или монатина, превращается в полипептид.
44. Изолированная нуклеиновая кислота, последовательность которой по крайней мере на 90% идентична SEQ ID NO: 11.
45. Изолированная нуклеиновая кислота по п.44, отличающаяся тем, что нуклеиновая кислота включает последовательность SEQ ID NO: 11.
46. Изолированная нуклеиновая кислота по п.44, отличающаяся тем, что нуклеиновая кислота включает по крайней мере 20 последовательных нуклеотидов последовательности SEQ ID NO: 11.
47. Вектор, представляющий собой изолированную нуклеиновую кислоту по п.44.
48. Рекомбинантная клетка, включающая вектор по п.47.
49. Изолированный белок, кодируемый изолированной нуклеиновой кислотой по п.44.
50. Изолированный белок по п.49, отличающийся тем, что белок включает последовательность, которая по крайней мере на 90% идентична SEQ ID NO: 12.
51. Изолированный белок по п.50, отличающийся тем, что белок включает последовательность SEQ ID NO: 12.
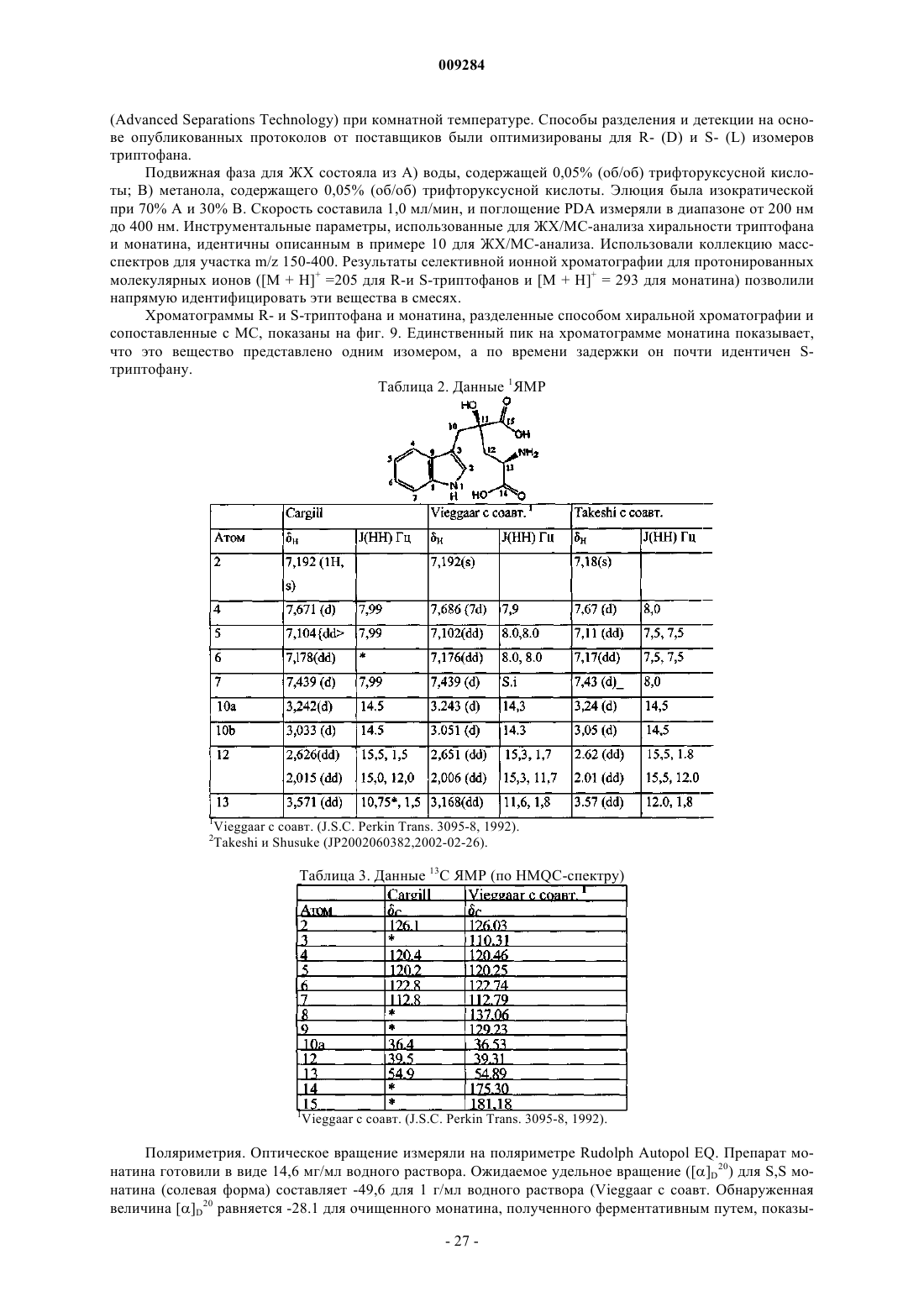
52. Споёюс получения монатина, включающий превращение 2-гидрокси 2-(индол-3-илметил)-4-кетоглутаровой кислоты в монатин.
53. Способ по п.52, отличающийся тем, что далее включает определение 2-гидрокси 2-(индол-3-илметил)-4-кетоглутаровой кислоты способом масс-спектрометрии.
54. Композиция, включающая изолированную или очищенную, или ненатурально возникающую 2-гидрокси 2-(индол-3-илметил)-4-кетоглутаровую кислоту.
55. Способ по п.2, отличающийся тем, что триптофан получают из индола с использованием полипептида из ЕС 4.1.99.1.
56. Способ получения монатина, включающий реакцию триптофана с одним или несколькими полипептидами.
57. Способ по п.56, отличающийся тем, что один или несколько полипепидов выбирают из группы, состоящей из триптофан аминотрансферазы (ЕС 2.6.1.27), триптофан дегидрогеназы (ЕС 1.4.1.19), тироизин (ароматической) аминотрансферазы (ЕС 2.6.1.5), триптофан-фенилпируват трансаминазы (ЕС 2.6.1.28), аминотрансферазы множественных субстратов (ЕС 2.6.1.-), аспартат аминотрансферазы (ЕС 2.6.1.1), триптофан оксидазы, оксидазы L-аминокислот (ЕС 1.4.3.2), дегидрогеназы D-аминокислот (ЕС 1.4.99.1), оксидазы D-аминокислот (ЕС 1.4.3.3), D-аланин аминотрансферазы (ЕС 2.6.1.21), D-триптофан аминотрансферазы, синтазы/лиазы (4.1.3.-), синтазы/лиазы (4.1.2.-), фенилаланин дегидрогеназы (ЕС 1.4.1.20), глутамат дегидрогеназы (ЕС 1.4.1.2, 1.4.1.3, 1.4.1.4) и комбинации этих соединений.
58. Клетка, способная продуцировать монатин, отличающаяся тем, что включает 2-гидрокси 2-(индол-3 илметил)-4 кетоглутаровую кислоту (предшественник монатина), где клетка включает по крайней мере одну экзогенную последовательность нуклеиновой кислоты, кодирующей по крайней мере один полипептид, и этот по крайней мере один полипептид осуществляет превращение первого промежуточного продукта по пути получения монатина с образованием второго промежуточного продукта или монатина.
59. Клетка по п.58, отличающаяся тем, что по крайней мере один полипептид выбран из группы, которая включает триптофан аминотрансферазу (ЕС 2.6.1.27), триптофан дегидрогеназу (ЕС 1.4.1.19), тирозин (ароматическую) аминотрансферазу (ЕС 2.61.5), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), триптофан оксидазу, оксидазу L-аминокислот (ЕС 1.4.3.2), аспартат аминотрансферазу (ЕС 2.6.1.1), дегидрогеназу D-аминокислот (ЕС 1.4.99.1), оксидазу D-аминокислот (ЕС 1.4.3.3), D-аланин аминотрансферазу (ЕС 2.6.1.21), D-триптофан аминотрансферазу, индоллактат дегидрогеназу (ЕС 1.1.1.110), R-4-гидроксифениллактат дегиидрогеназу (ЕС 1.1.1.222), 3-(4)-гидроксифенилпируват редуктазу (ЕС 1.1.1.237), лактат дегидрогеназу (ЕС 1.1.1.27, 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил) лактат дигидрогеназу (ЕС 1.1.1.111), лактат оксидазу (ЕС 1.1.3.-), синтазу/лиазу (4.1.2.-), синтазу/лиазу (4.1.3.-), фенилаланин дегидрогеназу (ЕС 1.4.1.20), глутамат дегидрогеназу (ЕС 1.4.1.2., 1.4.1.3., 1.4.1.4) и комбинацию этих соединений.
60. Клетка по п.59, отличающаяся тем, что по крайней мере один полипептид выбран из группы, которая включает триптофан аминотрансферазу (ЕС 2.6.1.27), триптофан дегидрогеназу (ЕС 1.4.1.19), тирозин (ароматическую) аминотрансферазу (ЕС 2.6.1.5), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), аспартат аминотрансферазу (ЕС 2.6.1.1), триптофан оксидазу, оксидазу L-аминокислот (ЕС 1.4.3.2), дегидрогеназу D-аминокислот (ЕС 1.4.99.1), оксидазу D-аминокислот (ЕС 1.4.3.3), D-аланин аминотрансферазу (ЕС 2.6.1.21), D-триптофан аминотрансферазу, синтазу/лиазу (4.1.3.-), синтазу/лиазу (4.1.2.-), фенилаланин дегидрогеназу (ЕС 1.4.1.20), глутамат дегидрогеназу (ЕС 1.4.1.2., 1.4.1.3., 1.4.1.4) и комбинацию этих соединений.
61. Клетка по п.59, отличающаяся тем, что по крайней мере один полипептид представлен активностью, выбранной из группы, которая включает индоллактат дегидрогеназу (ЕС 1.1.1.110), R-4-гидроксифениллактат дегиидрогеназу (ЕС 1.1.1.222), 3-(4)-гидроксифенилпируват редуктазу (ЕС 1.1.1.237), лактат дегидрогеназу (ЕС 1.1.1.27, 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил) лактат дигидрогеназу (ЕС 1.1.1.111), лактат оксидазу (ЕС 1.1.3.-), синтазу/лиазу (4.1.2.-, синтазу/лиазу (4.1.3.-), триптофан дегидрогеназу (ЕС 1.4.1.19), триптофан-фенилпируват трансаминазу (ЕС 2.6.1.28), триптофан аминотрансферазу (ЕС 2.6.1.27), тироизин (ароматическую) аминотрансферазу (ЕС 2.6.1.5), аминотрансферазу множественных субстратов (ЕС 2.6.1.-), аспартат аминотрансферазу (ЕС 2.6.1.1), глутамат дегидрогеназу (ЕС 1.4.1.2., 1.4.1.3., 1.4.1.4), фенилаланин дегидрогеназу (ЕС 1.4.1.20), D-триптофан аминотрансферазу, дегидрогеназу D-аминокислот (ЕС 1.4.99.1), D-аланин аминотрансферазу (ЕС 2.6.1.21) и комбинацию этих соединений.
62. Клетка по п.59, отличающаяся тем, что по крайней мере один полипептид является синтаза/лиазой (ЕС 4.1.3.-), включающей 4-гидрокси-2-оксоглутарат альдолазу (ЕС 4.1.3.16) или 4-гидрокси-4-метил-2-оксоглутарат альдолазу (ЕС 4.1.3.17).
63. Клетка по п.58, отличающаяся тем, что клетка представлена бактериальной клеткой.
64. Клетка по п.59, отличающаяся тем, что клетка представлена дрожжевой клеткой.
65. Клетка по п.58, отличающаяся тем, что клетка представлена одним или несколькими полипептидами, кодирующими по крайней мере одну экзогенную последовательность нуклеиновой кислоты и одним или более полипептидом, причем клетка дополнительно включает активности, выбранные из группы, которая включает каталазу, оксалацатат декарбоксилазу, лизин эписилон аминотрансферазу и формиат дегидрогеназу.
66. Способ по п.43, отличающийся тем, что один или более субстратов получены химической реакцией.
67. Способ по п.2, отличающийся тем, что производится по крайней мере 1 мг/мл монатина.
Текст
009284 Область применения Содержанием данного изобретения являются полипептиды, а также пути биосинтеза, которые могут найти применение при производстве индол-3-пирувата, 2-гидрокси 2-(индол-3 илметил)-4 кетоглутаровой кислоты (ПМ) и/или монатина. Предпосылки Индол-пируват. Индол-3-пируват является сильным антиоксидантом, который считается хорошей защитой от окислительного стресса в тканях с высокими концентрациями кислорода (Politi с соавт. "Recent advances inTryptophan Research", под ред. G. A. Filippini и др., Plenum Press, Нью-Йорк, 1996, стр. 291-298). Кроме того, индолпируват является промежуточным продуктом при получении индол-уксусной кислоты (ИУК),первичного гормона роста растений ауксина (фактор диффузного роста). ИУК проявляет активность в микрограммовых количествах в широком диапазоне физиологических процессов, таких как верхушечное доминирование, тропизм, удлинение боковых ветвей, индукция деления клеток камбия и инициация укоренения. Синтетические ауксины используются в садоводстве для индукции укоренения, стимулирования посадочного материала и формирования плодов. В высоких концентрациях синтетические ауксины являются эффективными гербицидами в отношении широколиственных растений. Натуральные ауксины,полученные путем ферментации, можно считать с экологической точки зрения более безопасными по сравнению с гербицидами, полученными в результате химического синтеза. Мировой объем продаж регуляторов роста составил в 1999 году 0,4 миллиарда фунтов (1,4 миллиарда долларов США). Среди примеров патентов на индол-уксусную кислоту и ее производные можно назвать следующие: патент США No. 5843782 Микрометод размножения розовых кустов с использованием ауксина в культуральных средах и патент США No. 5952231 Микрометод размножения розовых кустов. Помимо использования в растениеводстве производные индол уксусной кислоты находят применение и в фармацевтике. Например, патент США No. 5173497 "Метод получения альфа-оксопирроло-[2,3 В]-индол-уксусных кислот и их производных" предлагает применять эти соединения при лечении нарушений памяти, таких как при болезни Альцгеймера и сенильной деменции. Механизм действия, изложенный в патенте США No. 5173497, заключается в том, что эти соединения подавляют активность полипептида ацетилхолинэстеразы и повышают содержание ацетилхолина в головном мозге. Индол-3-карбинол получается из индол-3-уксусной кислоты в результате реакции окисления, катализируемой пероксидазой, и может быть легко превращен в дииндолилметан. Имеются сообщения, что оба эти соединения выводят токсины и способствуют синтезу гормонов, полезных для здоровья женщин. Производные триптофана Обнаружено, что хлорпроизводные D-триптофана являются некалорийными заменителями сахара,и в мире сохраняется повышенный интерес к поиску других соединений с подобными свойствами. Монатин является природным заменителем сахара, который по структуре похож на аминокислоту триптофан. Его можно экстрагировать из коры южно-африканского кустарника Sclerochiton ilicifolius, и он имеет многообещающие перспективы на применение в качестве высокоэффективного заменителя сахара в пищевой промышленности, в частности при производстве напитков. В качестве примеров патентов на монатин можно привести: патент США No. 5994559 "Синтез монатина-А - высокоэффективного заменителя сахара", патент США No. 4975298 "Соединения 3-(1-амино-1,3-дикарбокси-3-гидрокси-бут-4-ил)индола", патент США No. 5128164 "Композиция для питания человека, содержащая соединения 3-(1 амино-1,3-дикарбокси-3-гидрокси-бут-4-ил)-индола" и патент США No. 5128482 "Процесс получения 3(1-амино-1,3-дикарбокси-3-гидрокси-бут-4-ил)-индола. Некоторые из описанных предшественников монатина могут оказаться полезными в качестве синтетических заменителей сахара или промежуточных продуктов при синтезе производных монатина. Краткое описание Настоящее изобретение посвящено способам биосинтеза, применяемым для получения монатина из глюкозы, триптофана, индол-3-лактата, и/или через такие предшественники монатина, как индол-3 пируват и 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровая кислота. Кроме того, в состав изобретения входят полипептиды и нуклеотидные последовательности могут быть использованы при получении монатина, индол-3-пирувата и 2-гидрокси-2-(индол-3-илметил)-4- кетоглутаровой кислоты. Монатин можно получить через индол-3-пируват, 2-гидрокси-2-(индол-3-илметил)-4-кетоглутарат(фиг. 1). Раскрываются методы получения или синтеза монатина или его производных, показанные на фиг. 1-3 и 11-13, которые включают превращение субстрата в первый продукт, затем первого продукта во второй продукт и так далее, до получения нужного конечного продукта. На фиг. 1-3 и 11-13 в прямоугольных рамках показаны потенциальные промежуточные и конечные продукты. Так, превращение одного продукта в другой, например глюкозы в триптофан, триптофана в индол-3-пируват, индол-3-пирувата в ПМ, ПМ в монатин или индол-3-лактата (индол-лактата) в индол-3 пируват, может быть осуществлено с применением этих методов. Эти превращения можно упростить химическими или биологическими способами. Термин "превратить" относится к применению или химических средств, или полипептидов в реакции, которая приводит к получению из первого промежуточного-1 009284 соединения второго промежуточного соединения. Термин "химическое превращение" относится к реакциям, в которых полипептиды не принимали активного участия. Термин "биологическое превращение" относится к реакциям, в которых активно участвовали полипептиды. Превращения могут проходить какin vivo, так и in vitro. Если применялись биологические превращения, то полипептиды и/или клетки могут быть иммобилизованы на различных носителях, например, с помощью химической связи с полимерным носителем. Превращение может осуществляться с помощью любого реактора, известного и применяемого в соответствующей области, например в смесителе или реакторе непрерывного действия. Кроме того, представлены методы, которые включают контактирование первого полипептида с субстратом и получение первого продукта, затем контактирование первого продукта со вторым полипептидом и получение второго продукта, а затем контактирование полученного второго продукта с третьим полипептидом с получением третьего продукта, например монатина. Используемые полипептиды и получаемые продукты представлены на фиг. 1-3 и 11-13. Раскрываются полипептиды, которые можно использовать для проведения превращений, представленных фиг. 1-3 и 11-13, а также кодирующие их последовательности. В некоторых случаях для получения монатина используются полипептиды, имеющие одну или несколько точковых мутаций, которые позволяют модифицировать субстратную специфичность и/или активность полипептидов. В формулу изобретения входят выделенные и рекомбинантные клетки, которые продуцируют монатин. Этими клетками могут быть клетки любого происхождения: растительного, животного, бактериального, дрожжевого, из водорослей, архей или грибов. В одном из примеров патентуемые клетки включают одну или несколько следующих активностей, например две или более или три или более следующих активностей: триптофан аминотрансфераза (ЕС 2.6.1.27), тирозин (ароматическая) аминотрансфераза (ЕС 2.6.1.5), аминотрансфераза множественных субстратов (ЕС 2.6.1.-), аспартат аминотрансфераза(ЕС 1.4.1.2, 1.4.1.3, 1.4.1.4). В другом примере клетки содержат одну или несколько, например две или более или три или более,следующих активностей: индоллактат-дегидрогеназа (ЕС 1.1.1.110), R-4-гидроксифениллактатдегидрогеназа (ЕС 1.1.1.222), 3-(4)-гидроксифенилпируват редуктаза (ЕС 1.1.1.237), лактатдегидрогеназа(ЕС 1.1.1.27, 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил)-лактатдегидрогеназа (ЕС 1.1.1.111), лактат-оксидаза (ЕС 1.1.3.-), синтаза/лиаза (4.1.3.-), например, 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (ЕС 4.1.3.17) или 4-гидрокси-2-оксоглутарат-альдолаза (ЕС 4.13.16), синтаза/лиаза (4.12.-), триптофан-дегидрогеназа (ЕС 1.4.1.19), триптофан-фенилпируват трансаминаза (ЕС 2.6.1.28), триптофан аминотрансфераза (ЕС 2.6.127), тирозин (ароматическая) аминотрансфераза (ЕС 2.6.1.5), аминотрансфераза множественных субстратов (ЕС 2.6.1.-), аспартат-аминотрансфераза (ЕС 2.6.1.1) фенилаланин-дегидрогеназа (ЕС 1.4.1.20), глутаматдегидрогеназа (ЕС 1.4.12, 1.4.1.3, 1.4.1.4), дегидрогеназа D-аминокислот (ЕС 1.4.99.1),D-триптофан аминотрансфераза, и/или D-аланин аминотрансфераза (ЕС 2.6.121). Кроме того, патентуемые могут содержать одну или несколько следующих активностей, например две или более или три или более следующих активностей: триптофан аминотрансфераза (ЕС 2.6.127),тирозин (ароматическая) аминотрансфераза (ЕС 2.6.1.5), аминотрансфераза множественных субстратов(ЕС 1.1.1.27, 1.1.1.28, 1.1.23), (3-имидазол-3-ил)-лактатдегидрогеназа (ЕС 1.1.1. 111), лактат-оксидаза (ЕС 1.1.3.-), синтаза/лиаза (4.1.3.-), например, 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (ЕС 4.1.3.17) или 4-гидрокси-2-оксоглутарат-альдолаза (ЕС 4.13.16), синтаза/лиаза (4.1.2.-), глутамат-дегидрогеназа (ЕС 1.4.1.2, 1.4.1.3, 1.4.1.4), фенилаланин-дегидрогеназа (ЕС 1.4.1.20), и/или D-триптофан аминотрансфераза. Монатин можно получить методом, который включает контактирование триптофана и/или индол-3 лактата с первым полипептидом, при этом первый полипептид превращает триптофан и/или индол-3 лактат в индол-3-пируват (можно использовать или D-, или L-форму триптофана или индол-3-лактата в качестве субстрата, превращаемого в индол-3-пируват; требуется особое мастерство, чтобы выбранные на этом этапе полипептиды идеально проявляли соответствующую специфичность), контактирование полученного индол-3-пирувата со вторым полипептидом, при этом второй полипептид превращает индол-3-пируват в 2-гидрокси 2-(индол-3-илметил)-4-кетоглутаровую кислоту (ПМ), и контактирование ПМ с третьим полипептидом, при котором третий полипептид превращает ПМ в монатин. Примеры по-2 009284 липептидов, которые могут использоваться для этих превращений, приведены на фиг. 2 и 3. Еще один аспект изобретения касается таких композиций, как ПМ, клеток, которые содержат не менее двух полипептидов или иногда не менее трех или не менее четырех полипептидов, которые кодируются по крайней мере одной экзогенной последовательностью нуклеотидов. Эти и другие аспекты изобретения разъясняются в приведенных ниже детальном описании и иллюстративных примерах. Краткое описание иллюстраций Фиг. 1 показывает различные пути биосинтеза, используемые для получения монатина и/или индол 3-пирувата. При одном из них получают индол-3-пируват через триптофан, тогда как при другом индол 3-пируват получают через индол-3-лактат. Монатин впоследствии получают через промежуточное соединение 2-гидрокси-2-(индол-3-илметил)-4-кетоглутарат (ПМ). Соединения, указанные в прямоугольных рамках, являются субстратами и продуктами, получаемыми в результате биосинтетических реакций. Вещества, указанные рядом со стрелками, это кофакторы, или реагенты, которые могут быть использованы в процессе превращения субстрата в продукт. Использование того или иного кофактора или реагента зависит от полипептида, применяемого для проведения конкретного этапа в последовательности биосинтетических реакций. Кофактор ПФ (пиридоксаль-5'-фосфат) может катализировать реакции независимо от полипептида, и поэтому наличие одного ПФ может обеспечить переход субстрата в продукт. Фиг. 2 представляет более подробную диаграмму пути биосинтеза, при котором в качестве промежуточного продукта используется ПМ. Субстраты для каждого этапа показаны в прямоугольных рамках. Полипептиды, осуществляющие превращения между субстратами, указаны рядом со стрелками между субстратами. Для каждого полипептида указано его название и номер по Международной классификации ферментов (ЕС). Фиг. 3 представляет более подробную диаграмму пути биосинтеза, при котором происходит превращение индол-3-лактата в индол-3-пируват. Субстраты для каждого этапа представлены в прямоугольных рамках, а полипептиды, осуществляющие превращения между субстратами, указаны рядом со стрелками между субстратами. Для каждого полипептида указано его название и номер по Международной классификации ферментов (ЕС). На фиг. 4 представлена одна из возможных реакций получения ПМ химическим способом. На фиг. 5 А и 5 В представлены хроматограммы, показывающие результаты качественного определения монатина, полученного с применением ферментов, методами ЖХ/МС. На фиг. 6 представлен полученный с помощью электрораспылителя масс-спектр монатина, синтезированного с помощью ферментов. На фиг. 7 А и 7 В представлены хроматограммы, показывающие результаты ЖХ/МС/МС-дочернего ионного анализа монатина, полученного с применением смеси ферментов. На фиг. 8 представлена хроматограмма, показывающая результаты высокоточного измерения массы монатина, полученного с применением ферментов. На фиг. 9 А-9 С представлены хроматограммы, показывающие результаты хирального разделения(А) R-триптофана, (В) S-триптофана и (С) монатина, полученного с применением ферментов. На фиг. 10 представлена гистограмма, показывающая относительное содержание монатина, продуцируемого бактериальными клетками после индукции IPTG. Знаком (-) обозначено отсутствие добавления субстрата (не было добавлено ни триптофана, ни пирувата). На фиг. 11-12 показаны схематические диаграммы, демонстрирующие пути, используемые для увеличения выхода монатина, синтезируемого из триптофана или индол-3-пирувата. На фиг. 13 представлена схематическая диаграмма, показывающая способ увеличения выхода монатина, получаемого из триптофана или индол-3-пирувата. Список последовательностей Нуклеотидные и аминокислотные последовательности, перечисленные в прилагаемом списке последовательностей, представлены с помощью стандартных однобуквенных сокращений для оснований нуклеотидов и трехбуквенных сокращений для аминокислот. Представлена только одна цепочка для каждой нуклеиновой кислоты, но подразумевается, что комплементарная цепочка включается при ссылке на представленную последовательность.SEQ ID : 1 и 2 представляют нуклеотидную и аминокислотную последовательности аминотрансферазы из Sinorhisobium meliloti, соответственно (ген tatA, называемый в литературе тирозин аминотрансфераза или ароматическая аминотрансфераза).SEQ ID : 3 и 4 представляют нуклеотидную и аминокислотную последовательности тирозин аминотрансферазы из Rhodobacter sphaeroides (2.4.1), соответственно (по гомологии с геном tatA (SEQ ID: 1 и 2), предсказанным как "аспартат аминотрансфераза" с помощью компьютерных программ для геномного анализа).SEQ ID : 5 и 6 представляют нуклеотидную и аминокислотную последовательности аминотрансферазы из Rhodobacter sphaeroides (35053), соответственно (новый ген, клонированный на основе после-3 009284 довательности для 2.4.1 SEQ ID3 и 4).SEQ ID : 7 и 8 представляют нуклеотидную и аминокислотную последовательности аминотрансферазы широкого спектра действия (bsat) из Leishmania major, соответственно.SEQ ID : 9 и 10 представляют нуклеотидную и аминокислотную последовательности ароматической аминотрансферазы (oral) из Bacillus subtilis, соответственно.SEQ ID : 11 и 12 представляют новые нуклеотидную и аминокислотную последовательности ароматической аминотрансферазы (oral) из Lactobacillus amylovorus, соответственно (идентифицированной по гомологии как ароматическая аминотрансфераза).SEQ ID : 13 и 14 представляют нуклеотидную и аминокислотную последовательности аминотрансферазы множественных субстратов (mar) из R. sphaeroides (35053), соответственно (идентифицированной как аминотрансфераза множественных субстратов по гомологии с AC No. AAAE01000093.1,н.п.14743-16155 и АС No. ZP00005082.1).SEQ ID : 15 и 16 представляют праймеры, использованные для клонирования последовательностиSEQ ID : 17 и 18 представляют праймеры, использованные для клонирования последовательностиSEQ ID : 19 и 20 представляют праймеры, использованные для клонирования последовательностиSEQ ID : 21 и 22 представляют праймеры, использованные для клонирования последовательностей (2.4.1 и 35053) аминотрансферазы множественных субстратов из Rhodobacter sphaeroides.SEQ ID : 23 и 24 представляют праймеры, использованные для клонирования последовательностиSEQ ID : 25 и 26 представляют праймеры, использованные для клонирования последовательностиSEQ ID : 27 и 28 представляют праймеры, использованные для клонирования последовательностей tatA из R. sphaeroides (и 2.4.1, и 35053).SEQ ID : 29 и 30 представляют праймеры, использованные для клонирования последовательностиaspC из E.coli (нуклеотидная последовательность зарегистрирована в Genbank под AC No.: AE000195.1; белковая последовательность зарегистрирована в Genbank под AC No.: AAC74014.1).SEQ ID : 31 и 32 представляют нуклеотидную и аминокислотную последовательности ароматической аминотрансферазы (tyrB) из E.coli, соответственно.SEQ ID : 33 и 34 представляют праймеры для клонирования последовательности tyrB из Е. coli.SEQ ID : 35-40 представляют праймеры, использованные для клонирования полипептидов с активностью 4-гидрокси-2-оксоглутарат-альдолазы (KHG) (ЕС 4.1.3.16).SEQ ID : 41 и 42 представляют нуклеотидные последовательности триптофаназы (tna) из E.coli и тирозин фенол-лиазы (tpl) из Citrobocter freundii, кодирующих белки Р 00913 (GI:401195) и Р 31013(GI:401201), соответственно,SEQ ID : 43-46 представляют праймеры, использованные для клонирования последовательности полипептидов триптофаназы и полипептидов р-тирозиназы (тирозин фенол-лиазы).SEQ ID : 47-54 представляют праймеры, использованные для мутационной модификации полипептидов триптофаназы и полипептидов р-тирозиназы.SEQ ID : 55-64 представляют праймеры, использованные для клонирования полипептидов с активностью 4-гидрокси-4-метил-2-оксоглутарат-альдолазы (ЕС 4.13.17).SEQ ID : 67-68 представляют праймеры, использованные для клонирования последовательности 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (proA) из С. testosterone в опероне с aspC из Е. coli в рЕТ 30 Ха/LIC.SEQ ID : 69-72 представляют праймеры, использованные для клонирования последовательностейSEQ ID : 73-74 представляют последовательности, добавленные к 5'-концу праймеров, использованных для клонирования генов, о которых идет речь в настоящем документе. Подробное описание некоторых примеров осуществления Сокращения и термины Представленные ниже объяснения терминов и методов приведены для того, чтобы лучше описать данное изобретение и ознакомить квалифицированных специалистов с приложениями данного изобретения. В настоящем документе термин содержащий означает включающий, а существительные в единственном числе включают множественные объекты, если их в контексте явно не указывается другое. Например, выражение содержащий белок подразумевает один или множество таких белков, а выражение содержащий клетку подразумевает наличие одной или многих клеток и соответствующие эквиваленты, известные квалифицированным специалистам, и т.д. Если не объясняется иное, все используемые в настоящем документе технические и научные тер-4 009284 мины имеют то же значение, которое подразумевается специалистами, имеющими достаточную квалификацию в области, к которой относится данное изобретение. Хотя методы и материалы, аналогичные или эквивалентные описанным в настоящем документе, можно применять в практике или при тестировании данного изобретения, ниже описываются соответствующие методы и материалы. Примеры, методы и материалы приведены исключительно в иллюстративных целях и не предусматривают каких бы то ни было ограничений. Из дальнейшего подробного описания и пунктов заявки станут ясными другие особенности и преимущества данного изобретения. кДНК (комплементарная ДНК): фрагмент ДНК, лишенный внутренних, некодирующих участков(интронов) и регуляторных последовательностей, которые определяют транскрипцию. кДНК можно синтезировать и в лабораторных условиях методом обратной транскрипции с матричной РНК, выделенной из клеток. Консервативная замена: одна или более аминокислотных замен (например, 2, 5, или 10 остатков) на аминокислотные остатки со сходными бихимическими свойствами. Обычно консервативные замены мало влияют на активность полученного полипептида. Например, в идеале, полипептид триптофанаминотрансфераза, содержащий одну или несколько консервативных замен, сохраняет триптофанаминотрансферазную активность. В результате модификации нуклеотидной последовательности, например, с помощью таких стандартных методов, как направленный мутагенез или ПЦР, можно получить полипептид, содержащий одну или несколько консервативных замен. Замещенные варианты полипептидов - это те, у которых по крайней мере один остаток в аминокислотной последовательности удален и на его место вставлен другой остаток. Ниже приведены примеры аминокислот, которые могут заместить оригинальную аминокислоту в белковой последовательности, и считаются консервативными заменами: Ala замещается на ser или thr; arg замещается на gln, his или lys;asn, glu, gln, his, arg; met замещается на ile, ten, val, phe; phe замещается на trp, tyr, met, ile или leu; ser замещается на thr, ala; thr замещается на ser или ala; trp замещается на phe, tyr; tyr замещается на his, phe или trp; и val замещается на met, ile, leu. Дополнительная информация о консервативных заменах может быть найдена в других источниках,в частности Ben-Bassat с соавт., (J. Bacteriol 169:751-7,1987), O'Regan с соавт., (Gene 77:237-51,1989), Sahin-Toth с соавт., (Protein Ssi. 3:240-7,1994), Hochuli et al., (Bio/Technology 6:1321-5,1988), WO 00/67796(Curd с соавт.) и в стандартных учебниках генетики и молекулярной биологии. Экзогенный: термин экзогенный, используемый в настоящем документе в отношении нуклеиновой кислоты и конкретной клетки, указывает на то, что нуклеиновая кислота происходит не из данной клетки в результате естественных процессов. Так, нуклеиновая кислота неприродного происхождения считается экзогенной по отношению к клетке, как только она введена в клетку. Нуклеиновая кислота естественного происхождения также может быть экзогенной для данной клетки. Например, целая хромосома, полученная из клетки человека X, является экзогенной для клетки из организма человека Y, как только эта хромосома окажется в клетке Y. Функциональный эквивалент: имеющий эквивалентную функцию. Так, в число функционально эквивалентных молекул какого-либо фермента входят разные молекулы, которые сохраняют функцию этого фермента. Например, функциональные эквиваленты можно получить путем изменения последовательности фермента, если пептид с одной или несколькими модификациями последовательности сохраняет функцию неизмененного пептида. В частности, функциональный эквивалент триптофанаминотрансферазы сохраняет способность превращать триптофан в индол-3-пируват. В число примеров изменения последовательности входят (но без ограничений) консервативные замены, делеции, мутации, сдвиги рамки считывания и инсерции. Например, какой-либо полипептид связывает антитела, тогда его функциональный эквивалент - полипептид, который связывается с теми же антителами. То есть понятие "функциональный эквивалент" включает пептиды, которые имеют ту же специфичность связывания, что и основной полипептид, и которые могут использоваться в качестве реагента вместо этого полипептида. Так, в одном из примеров функциональным эквивалентом является полипептид, у которого связывающая последовательность прерывается, при этом антитела связываются с линейным эпитопом. То есть, если последовательность пептида - MPELANDLGL (аминокислоты 1-10 последовательности SEQ ID : 12), то функциональный эквивалент содержит эпитопы, расположенные прерывисто, и его последовательность выглядит следующим образом ( - вставка из любого числа аминокислот): NH2MPELANDLGL-COOH. В этом примере полипептид является функционально эквивалентным аминокислотам 1-10 последовательности SEQ ID : 12, если трехмерная структура этого полипептида позволяет ему связывать моноклональные антитела, которые связываются с-5 009284 аминокислотами 1-10 of последовательности SEQ ID : 12. Гибридизация: термин "гибридизация", используемый в настоящем документе, относится к методу тестирования комплементарности по нуклеотидной последовательности двух молекул нуклеиновой кислоты, основанный на способности комплементарной одноцепочечной ДНК и/или РНК образовывать двойную молекулу (дуплекс). Метод гибридизации нуклеиновых кислот можно использовать для получения изолированной нуклеиновой кислоты в рамках данного изобретения. Практически любую нуклеиновую кислоту, имеющую определенную гомологию с последовательностью, представленную под SEQID :11, можно использовать для выявления похожей нуклеиновой кислоты с помощью гибридизации в условиях умеренной или высокой жесткости. После выявления нуклеиновую кислоту можно очистить, секвенировать и проанализировать, чтобы определить, входит ли она в рамки данного изобретения. Гибридизацию можно проводить по Саузерну или методом нозерн-гибридизации для выявления последовательности ДНК или РНК, соответственно, которая гибридизуется с выбранным зондом. Зонд можно пометить биотином, диоксигенином, полипептидом или радиоизотопом, например Р 32. Анализируемая ДНК или РНК может быть выделена методом электрофореза на агарозе или в полиакриламидном геле, перенесена на нитроцеллюлозу, нейлон или другую подходящую мембрану, а затем гибридизована с соответствующим зондом с помощью стандартного метода, хорошо известного специалисту, например,одного из описанных в разделах 7.39-7.52 книги Sambrook с соавт., (1989) Molecular Cloning, второе издание, Cold Spring Harbor Laboratory, Plainview, NY. Как правило, длина зонда составляет около 20 нуклеотидов. Например, зонд, соответствующий непрерывной 20-нуклеотидной последовательности, представленной под SEQ ID : 11, можно использовать для идентификации идентичной или похожей нуклеиновой кислоты. Кроме того, можно использовать зонды длиннее или короче 20 нуклеотидов. Изобретение включает также выделенные нуклеотидные последовательности, которые имеют не менее 12 нуклеотидов в длину (то есть, по крайней мере, около 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 40, 50,60, 100, 250, 500, 750, 1000, 1500, 2000, 3000, 4000 или 5000 оснований в длину) и гибридизуются при соответствующих условиях гибридизации со смысловой или антисмысловой цепью нуклеиновой кислоты, представленной в SEQ ID : 11. Условия гибридизации могут быть умеренно или высоко жесткими. Для целей данного изобретения умеренно жесткие условия гибридизации означают, что гибридизация проводится при температуре около 42 С в гибридизационном растворе, содержащем 25 мМ KPO4(pH 7,4), 5 Х SSC, 5X раствор Денхарта, 50 мкг/мл денатурированной обработанной ультразвуком ДНК спермы лосося, 50% формамид, 10% декстран-сульфат и 1-15 нг/мл зонда (около 5107 квм/мкг), при этом отмывание проводится при температуре около 50 С промывающим раствором, содержащим 2 Х SSC и 0,1% додецилсульфат натрия. Гибридизация в жестких условиях означает, что гибридизация проводится при температуре около 42 С в гибридизационном растворе, содержащем 25 мМ KPO4 (pH 7,4), 5 Х SSC, 5X раствор Денхарта, 50 мкг/мл денатурированной обработанной ультразвуком ДНК спермы лосося, 50% формамид, 10% декстран-сульфат и 1-15 нг/мл зонда (около 5 х 107 квм/мкг), при этом отмывание проводится при температуре около 65 С промывающим раствором, содержащим 0,2 Х SSC и 0,1% додецилсульфат натрия. Изолированная: термин "изолированная", применяемый в настоящем документе по отношению к нуклеиновой кислоте, означает, что имеющая природное происхождение нуклеиновая кислота не примыкает непосредственно к обеим последовательностям, с которыми она непосредственно граничила (с одной на 5'-конце и с другой на 3'-конце) в природных условиях генома того организма, из которого она происходит. Например, изолированная нуклеиновая кислота может быть, без ограничений, рекомбинантной молекулой ДНК любой длины при условии, что одна из фланкирующих нуклеотидных последовательностей такой рекомбинантной молекулы ДНК удалена или отсутствует. Таким образом, под изолированной нуклеиновой кислотой понимается, без ограничений, рекомбинантная ДНК, которая представлена в виде отдельной молекулы (например, кДНК или фрагмент геномной ДНК, полученный с помощью ПЦР или обработки рестрикционной эндонуклеазой), независимой от других последовательностей, а также рекомбинантная ДНК, которая встроена в вектор, автономно реплицирующуюся плазмиду,вирус (например, ретровирус, аденовирус или вирус герпеса) или в геномную ДНК прокариот или эукариот. Кроме того, изолированная нуклеиновая кислота может включать рекомбинантную молекулу ДНК,которая является частью гибридной или сшитой нуклеотидной последовательности. Термин "изолированная", применяемый в настоящем документе по отношению к нуклеиновой кислоте, включает также любую нуклеиновую кислоту неприродного происхождения, так как нуклеотидные последовательности неприродного происхождения не встречаются в природе и не ограничены другими последовательностями в составе природного генома. Так, нуклеиновая кислота неприродного происхождения, например, полученная методами генной инженерии, считается изолированной нуклеиновой кислотой. Сконструированная нуклеиновая кислота может быть получена обычными методами молекулярного клонирования или химическими методами синтеза нуклеиновых кислот. Нуклеиновая кислота неприродного происхождения может быть независимой от других последовательностей, или встроена в вектор, автономно реплицирующуюся плазмиду, вирус (например, ретровирус, аденовирус или вирус герпеса) или в геномную ДНК прокариот или эукариот. Кроме того, нуклеи-6 009284 новая кислота неприродного происхождения может включать молекулу нуклеиновой кислоты, которая является частью гибридной или сшитой нуклеотидной последовательности. Квалифицированным специалистам понятно, что нуклеиновая кислота, существующая среди сотен или миллионов других молекул нуклеиновых кислот, например в составе библиотек кДНК, или геномных ДНК, или гелевых пластин, содержащих набор рестрикционных фрагментов геномной ДНК, не считается изолированной нуклеиновой кислотой. Нуклеиновая кислота: термин " нуклеиновая кислота", применяемый в настоящем документе, относится и к РНК, и к ДНК, включая, без ограничений, кДНК, геномной ДНК и синтетической (т.е. химически синтезированной) ДНК. Нуклеиновая кислота может быть двухцепочечной или одноцепочечной. В случае одноцепочечной, нуклеиновая кислота может быть смысловой или антисмысловой. Кроме того,нуклеиновая кислота может быть кольцевой или линейной. Оперативно сцеплена: первая последовательность нуклеотидов считается "оперативно сцепленной" со второй последовательностью нуклеотидов, если первая последовательность нуклеотидов находится в функциональной связи со второй последовательностью нуклеотидов. Например, промотор считается оперативно сцепленным с кодирующей последовательностью, если этот промотор влияет на ее транскрипцию. Как правило, оперативно сцепленные последовательности ДНК непосредственно граничат друг с другом и если необходимо соединить два полипептид-кодирующих региона в пределах одной рамки считывания. Пептидные модификации: в формулу настоящего изобретения входят ферменты, а также синтетические гомологи. Кроме того, аналоги (органические молекулы непептидной природы), производные(химически функциональные пептидные молекулы, полученные на основе раскрываемых пептидных последовательностей), и варианты (гомологи), обладающие нужной ферментной активностью, могут быть использованы в методах, описанных в настоящем документе. Представленные в настоящем документе пептиды включают последовательность аминокислот, которые могут быть L- и/или Dаминокислотами, естественного или иного происхождения. Пептиды могут быть модифицированы разными химическими методами с получением производных, которые обладают в сущности той же активностью, что и немодифицированные пептиды, а иногда и дополнительно приобретают другие полезные свойства. Например, карбоксильные группы белков, как терминальные, так и боковых цепей, могут быть представлены в виде соли или фармацевтически приемлемого катиона или этерифицированы с образованием С 1-С 16 эфира или превращены в амид с формулойNR1R2, где R1 и R2 независимо друг от друга Н- или С 1-С 16-алкильная группа, или соединены с образованием гетероциклического кольца, например, 5- или 6-звенного кольца. Аминогруппы этого пептида,как терминальная, так и аминогруппы боковых цепей, могут быть в форме фармацевтически приемлемой соли таких кислот, как HCl, HBr, уксусная, бензойная, толуол-сульфоновая, малеиновая, винная и другие органические кислоты, или могут быть модифицированы С 1-С 16-алкильными или диалкиламиногруппами или превращены в амид. Гидроксильные группы боковых цепей пептидов могут быть превращены в С 1-С 16-алкоксигруппы или в С 1-С 16-эфирные группы с помощью хорошо известных методов. Фенильные и фенольные кольца боковых цепей пептидов могут быть замещены одним или несколькими атомами галогенов, такими какF, Cl, Br или I, или С 1-С 16-алкильными, С 1-С 16-алкоксигруппами, карбоксильными кислотами и их эфирами или амидами таких карбоксильных кислот. Метиленовые группы боковых цепей пептидов могут быть увеличены до гомологичных С 2-С 4-алкиленов. Тиоловые группы можно защитить любой из числа хорошо известных защитных групп, например ацетамидными группами. Кроме того, специалисты узнают методы введения циклических структур в пептиды, входящие в формулу этого изобретения, чтобы выбирать и создавать конформационные связки в структуре, что повышает ее стабильность. Например, С- или Т-терминальный цистеин можно присоединить к пептиду таким образом, чтобы при окислении пептид содержал пептидную связь и превращался в циклический пептид. Циклизацию пептидов можно провести и другими способами, например через образование тиоэфиров и карбоксильных и амино-терминальных амидов и эфиров. Пептидомиметические и органомиметические аналоги также входят в формулу настоящего изобретения, в связи с чем трехмерное расположение химических компонентов таких пептидо- и органомиметиков повторяет трехмерное расположение пептида боковых цепей аминокислот, что приводит к наличию у таких пептидо- и органомиметиков белков, входящих в формулу данного изобретения, значимой ферментативной активности. При компьютерном моделировании фармакофор является идеальным трехмерным определением структурных требований к биологической активности. Пептидо- и органомиметики можно сконструировать таким образом, чтобы каждый фармакофор соответствовал текущему программному обеспечению для компьютерного моделирования (используя систему конструирования лекарств с помощью компьютера, или CADD). См. описания подходов, используемых в CADD, в статьях:of Pharmacology, 1995, ChapmanHall. Кроме того, в формулу данного изобретения включаются миметики, полученные с использованием таких подходов. В одном из примеров миметик имитирует фермент-7 009284 ную активность, которую демонстрирует фермент или его вариант, фрагмент или фузионный продукт. Зонды и праймеры: Нуклеотидные зонды и праймеры можно легко приготовить на основе аминокислотных и нуклеотидных последовательностей, представленных в данном документе. Понятие "зонд" включает изолированную нуклеиновую кислоту, содержащую определяемую меченую или репортерную молекулу. Примером метки могут служить, без ограничений, радиоактивные изотопы, лиганды, хемилюминесцентные агенты и полипептиды. Способы мечения и рекомендации по выбору меток, соответствующих различным целям, обсуждаются, например, в следующих источниках: Sambrook с соавт. (ed.),Molecular Cloning: A Laboratory Manual 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring"Праймеры" - это типичные молекулы нуклеиновых кислот, содержащие 10 и более нуклеотидов(например, молекулы нуклеиновой кислоты длиной примерно от 10 до 100 нуклеотидов). Праймер может быть получен в результате отжига с комплементарной цепью нужной нуклеиновой кислоты путем гибридизации с образованием гибрида между праймером и цепочкой нужной нуклеиновой кислоты, а затем удлинен вдоль нужной нуклеиновой кислотой, например, с помощью полипептида ДНК-полимеразы. Можно использовать пары праймеров для амплификации последовательности нуклеотидов, например,способом полимеразной цепной реакции (ПЦР) или другим способом амплификации нуклеиновых кислот, известных специалистам. Способы получения и применения зондов и праймеров описаны, например, в таких источниках, как:Publishing % Wiley-Interscience, Нью-Йорк (с периодическими обновлениями), 1987; и Innis и др. (ред.),ПЦР Protocols: A Guide to Methods и Applications, Academic Press: Сан-Диего, 1990. Пары праймеров для ПЦР могут быть получены на основе известной последовательности, например, с помощью компьютерных программ, предназначенных для таких целей, таких как Primer (версия 0,5, 1991, Whitehead Institutefor Biomedical Research, Кембридж, штат Массачусетс). Квалифицированный специалист согласится с тем, что специфичность конкретного зонда или праймера увеличивается с длиной, и в то же время длина зонда или праймера колеблется в диапазоне от полноразмерной последовательности до всего лишь пяти последовательно расположенных нуклеотидов. Так, например, праймер из 20 последовательно расположенных нуклеотидов можно отжечь с более высокой специфичностью, чем аналогичный праймер длиной только 15 нуклеотидов. Поэтому чтобы получить более высокую специфичность, можно выбрать зонды и праймеры, которые содержат, например, 10, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300,350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300,1350, 1400, 1450, 1500, 1550, 1600, 1650, 1700, 1750, 1800, 1850, 1900, 2000, 2050, 2100, 2150, 2200, 2250,2300, 2350, 2400, 2450, 2500, 2550, 2600, 2650, 2700, 2750, 2800, 2850, 2900, 3000, 3050, 3100, 3150, 3200,3250, 3300, 3350, 3400, 3450, 3500, 3550, 3600, 3650, 3700, 3750, 3800, 3850, 3900, 4000, 4050, 4100, 4150,4200, 4250, 4300, 4350, 4400, 4450, 4500,4550, 4600, 4650, 4700,4750, 4800, 4850, 4900, 5000, 5050, 5100,5150, 5200, 5250, 5300, 5350, 5400, 5450 или более последовательно расположенных нуклеотидов. Промотор: ряд управляющих нуклеотидных последовательностей, которые направляют транскрипцию нуклеиновой кислоты. Промотор содержит обязательные нуклеотидные последовательности, расположенные вблизи сайта инициации (начала) транскрипции, таких как, в случае промотора полимеразы II типа, ТАТА-элемент. В состав промотора может входить дистальный энхансер или репрессорные элементы, которые могут располагаться на расстоянии в несколько тысяч пар нуклеотидов от сайта инициации транскрипции. Очищенный: термин "очищенный", применяемый в настоящем документе, не подразумевает абсолютной чистоты, это скорее относительное понятие. Так, например, очищенный полипептид или очищенный препарат нуклеиновой кислоты может быть таким препаратом, в котором данный полипептид или нуклеиновая кислота, соответственно, находится в более высокой концентрации, чем полипептид или нуклеиновая кислота в естественном окружении в организме. Например, препарат полипептида может считаться очищенным, если содержание полипептида в препарате составляет не менее 50, 60, 70, 80,85, 90, 92, 95, 98 или 99% от общего содержания белка в этом препарате. Рекомбинатный: "рекомбинантная" нуклеиновая кислота - нуклеиновая кислота, которая (1) состоит из последовательности, которой нет в организме в естественных условиях, или (2) получена искусственной комбинацией двух не соединявшихся ранее более коротких последовательностей. Эта искусственная комбинация часто осуществляется путем химического синтеза или, чаще, с помощью искусственных манипуляций с изолированными фрагментами нуклеиновых кислот, например, способами генной инженерии. Термин "рекомбинатный" используется также для описания молекул нуклеиновых кислот, с которыми проводились искусственные манипуляции, но они содержат те же регуляторные последовательности и кодирующие участки, которые встречаются и в том организме, из которого выделена данная нуклеиновая кислота. Идентичность последовательностей: сходство между аминокислотными последовательностями выражается в терминах сходства между последовательностями, иначе называемым идентичностью после-8 009284 довательностей. Идентичность последовательностей часто измеряется в процентной идентичности (или сходстве или гомологии); чем выше процент, тем более похожи этим последовательности. Гомологи или варианты пептида, такого как SEQ ID :12, обладают относительно высокой степенью идентичности при выравнивании стандартными способами. Способы выравнивания последовательностей для сравнения хорошо известны квалифицированным специалистам. Различные программы для сравнения хорошо известны специалистам. Различные программы и алгоритмы выравнивания описаны в следующих источниках: Smith и Waterman, Adv. Appl.Corpet с соавт., Nucleic Acids Research 16:10881-10890,1988; и Altschul с соавт., Nature Genet. 6:119-29,1994. Разработанная в Национальном Центре биотехнологической информации США (National Center forBiotechnology Information, NCBI) программа BLAST(Basic Local Alignment Search Tool) (Altschul с соавт., J. Mol. Biol. 215:403-10,1990) может быть получена из разных источников, включая NCBI (Bethesda,MD) и Интернет, для применения в связи с программами анализа последовательностей blastp, blastn,blast, tblastn и tblastx. Варианты пептида обычно характеризуются наличием по крайней мере 50%-ного сходства последовательностей в расчете на полноразмерное выравнивание аминокислотной последовательности с помощью программы NCBI Blast 2.0, используя параметры по умолчанию для сравнения с разрывами. Для сравнения аминокислотных последовательностей длиной более 30 аминокислот применяется функцияBlast 2 с использованием набора матриц BLOSUM62 по умолчанию с параметрами по умолчанию (стоимость разрыва 11, стоимость одного остатка в разрыве 1). При выравнивании коротких пептидов (менее 30 аминокислот) выравнивание проводится с функцией последовательностей Blast 2 с использованием набора матриц РАМ 30 по умолчанию с параметрами по умолчанию (открытый разрыв 9, дополнительный штраф за разрыв 1). Белки даже с большим сходством с последовательностями сравнения будут показывать возрастание процентной идентичности при оценке этим способом, например, не менее 80%, не менее 90%, не менее 95%, не менее 98% или даже не менее 99% идентичности. Если сравнивается неполная последовательность на предмет идентичности последовательности, гомологи и варианты будут показывать не менее 80% идентичности последовательности на коротких окнах из 10-20 аминокислот, и могут демонстрировать идентичность на уровне не менее 85%, не менее 90%, не менее 95% или 98% в зависимости от их сходства с основной последовательностью. Способы определения идентичности последовательностей при таких коротких окнах описаны на web-сайте, поддерживаемом в Национальном Центре биотехнологической информации (г. Бетезда, штат Мэриленд). Квалифицированный специалист согласится, что такие диапазоны идентичности последовательности представлены только для примера и что можно получить значительно совпадающие гомологи, которые окажутся вне представленных диапазонов. Аналогичные способы можно применить для определения процента идентичности нуклеотидной последовательности. В одном из примеров гомологичная последовательность выравнивается против нативной последовательности, и количество совпадений определяется путем подсчета числа позиций, в которых идентичный нуклеотид или аминокислотный остаток представлен в обеих последовательностях. Процент идентичности последовательностей определяется делением числа совпадений на длину последовательности, определенной в идентифицированной последовательности (например, SEQ ID 11), или по заявленной длине (например, 100 последовательных нуклеотидов или аминокислотных остатков из последовательности, определенной по идентифицированной последовательности) с последующим умножением результата на 100. Например, нуклеотидная последовательность, которая имеет 1166 совпадений при выравнивании против последовательности в SEQ ID11, на 75,0% идентична последовательности в SEQ ID11 (т.е. 1166-1554100=75,0). Следует отметить, что процент идентичности последовательности округляется до ближайшей десятой. Например, 75,11, 75,12, 75,13 и 75,14 округляются до 75,1, а 75,15, 75,16, 75,17, 75,18 и 75,19 округляются до 75,2. Кроме того, следует отметить, что длина всегда является целым числом. В другом примере целевая последовательность, содержащая фрагмент длиной 20 нуклеотидов, который выравнивается с 20 последовательными нуклеотидами идентифицированной последовательности, содержит участок, который показывает 75% идентичности с этой идентифицированной последовательностью (т.е. 15-20100=75). Специфически связывающийся агент: агент, который способен специфически связываться с одним из полипептидов, описанных в настоящем документе. Среди примеров можно назвать, без ограничений,поликлональные антитела, моноклональные антитела (включая гуманизированные моноклональные ан-9 009284 титела) и фрагменты моноклональных антител, такие как Fab, F(ab') и Fv-фрагменты, а также любые другие агенты, которые могут специфически связываться с эпитопом таких полипептидов. Антитела к представленным в настоящем документе полипептидам (или фрагментам, вариантам или фузионным продуктам) можно использовать для очистки и идентификации таких полипептидов. Аминокислотные и нуклеотидные последовательности, представленные в настоящем документе, позволяют получить связывающие агенты на основе специфических антител, которые распознают полипептиды, представленные в настоящем документе. Можно получить моноклональные или поликлональные антитела к полипептидам, частям полипептидов или их вариантам. Считается оптимальным, если антитела против одного или нескольких эпитопов на полипептидном антигене специфически выявляют такой полипептид. То есть антитела, полученные против того или иного полипептида, будут распознавать и связываться с этим полипептидом, но практически не будут реагировать и связываться с другими полипептидами. Определение того, что антитела специфически связываются с данным полипептидом, проводится одним из многочисленных стандартных способов иммуноопределения; например вестерн-блоттинг (см., например, Sambrook и др. (ред.), Molecular Cloning: A Laboratory Manual, 2-е изд., тт. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor,N.Y., 1989). Чтобы определить, что препарат данного антитела (такой как препарат, полученный у мышей против полипептида с аминокислотной последовательностью, представленной в SEQ ID12) специфически определяет соответствующий полипептид (например, полипептид с аминокислотной последовательностью, представленной в SEQ ID 12) способом вестерн-блот-анализа, можно экстрагировать общий клеточный белок из клеток и разделить его способом электрофореза в полиакриламидном геле сSDS. Затем разделенный клеточный белок можно перенести на мембрану (например, нитроцеллюлозу) и проинкубировать ее с препаратом антител. После промывания мембраны для удаления неспецифически связавшихся антител наличие специфически связанных антител можно определить с помощью соответствующих вторичных антител (например, антимышиных антител), конъюгированных с таким полипептидом, как щелочная фосфатаза, поскольку применение 5-бром-4-хлор-3-индолилфосфат/нитросинего тетразолиевого приведет к образованию темно-синего вещества благодаря иммуно-связанной щелочной фосфатазе. Практически чистые полипептиды, пригодные для применения в качестве иммуногенов, можно получить из трансфицированных клеток, трансформированных клеток или интактных клеток. Концентрация полипептида в конечном препарате может быть отрегулирована, например, путем концентрирования на фильтре Amicon, до уровня в несколько микрограммов на миллилитр. Кроме того, полипептиды размером от полноразмерных полипептидов до полипептидов всего в несколько аминокислотных остатков можно использовать в качестве иммуногенов. Такие полипептиды можно получать в клеточной культуре,можно синтезировать химическими способами или получать путем расщепления крупных полипептидов на полипептиды меньшего размера, которые можно затем очистить. Полипептиды длиной всего девять аминокислотных остатков могут быть иммуногенными, если они представлены иммунной системе в контексте молекул главного комплекса гистосовместимости (МНС), таких как молекула МНС класса I или МНС класса II. Соответственно, полипептиды длиной не менее 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40,45, 50, 55, 60, 76, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 900, 1000,1050, 1100, 1150, 1200, 1250, 1300, 1350 или более последовательных аминокислотных остатков любой аминокислотной последовательности, представленной в настоящем документе, могут быть использованы в качестве иммуногенов для получения антител. Моноклональные антитела к любому из полипептидов, представленных в настоящем документе,можно получить из гибридомы мыши классическим способом KohlerMilstein (Nature 256:495-497,1975) или другим подобным способом. Поликлональная антисыворотка, содержащая антитела к гетерогенным эпитопам любого полипептида из представленных в настоящем документе, может быть приготовлена путем иммунизации соответствующих животных полипептидом (или его фрагментом), который может быть немодифицированным или модифицированным для улучшения иммуногенности. Протокол эффективной иммунизации кроликов можно найти в работе Vaitukaitis с соавт. (J. Clin. Endocrinol. Metab. 33:988-91,1971). Вместо целых антител можно использовать фрагменты антител, которые проще экспрессировать в прокариотических клетках. Способы получения и применения иммунологически активных фрагментов моноклональных антител, называемых также "фрагменты антител", хорошо известны и описаны, в частности, в работах: BetterHorowitz (Methods Enzymol. 178:476-96,1989), Glockshuber с соавт. (Biochemistry 29:1362-7,1990), патент США No. 5648237 ("Экспрессия функциональных фрагментов антител"), патент США No. 4946778 ("Молекулы, связывающие единичную полипептидную цепь"), патент США No. 5455030 ("Иммунотерапия с использованием молекул, связывающих единичную полипептидную цепь "),и в процитированной в этих работах литературе. Трансформированный: "трансформированная" клетка - это клетка, в которую введена молекула нуклеиновой кислоты с помощью, например, молекулярно-биологических способов. Трансформация происходит при всех способах, при которых нуклеиновая кислота может быть введена в такую клетку,- 10009284 включая, но не ограничиваясь, трансфекцию вирусным вектором, конъюгацию, трансформацию плазмидным вектором и введение чистой ДНК с помощью электропорации, липофекции и пушки для ускоренного введения частиц. Варианты, фрагменты и фузионные белки: представленные в документе белки включают варианты,фрагменты и фузионные белки. Последовательности ДНК (например, SEQ ID 11), которые кодируют белок (например, SEQ ID12), фузионный белок или фрагмент или вариант белка, могут быть сконструированы таким образом, чтобы позволить экспрессию белка в эукариотических клетках, бактериях,насекомых и/или растениях. Чтобы получить экспрессию, последовательность ДНК можно изменить и опреративно сцепить с другими регуляторными последовательностями. Конечный продукт, который содержит регуляторные последовательности и белок, называется вектором. Этот вектор можно ввести в клетки эукариот, бактерий, насекомых и/или растений. Как только вектор оказывается внутри клетки,начинается продукция выбранного белка. Фузионный белок включает белок, например триптофан-аминотрансферазу (или вариант, полиморфизм, мутантный вариант или фрагмент), например SEQ ID12, сцепленный с другими аминокислотными последовательностями, которые не подавляют полезную активность данного белка, например способность превращать триптофан в индол-3-пируват. В одном из примеров, прочие аминокислотные последовательности имеют длину не более 10, 12, 15, 20, 25, 30 или 50 аминокислот. Любой специалист согласится, что последовательность ДНК можно изменить разнообразными способами, не изменяя при этом биологическую активность кодируемого белка. Например, для получения вариаций последовательности ДНК, которая кодирует белок, можно применить ПЦР. Такие варианты могут быть вариантами, оптимизирующими предпочтительность кодонов в клетке хозяина, использованной для экспрессии белка, или другие изменения последовательности, которые облегчают экспрессию. Вектор: молекула нуклеиновой кислоты, введенная в клетку, в результате чего получается трансформированная клетка. Вектор может содержать последовательности нуклеотидов, которые позволяют ему реплицироваться в клетке, такие как точка начала репликации. Вектор может также содержать один или несколько селекционируемых маркерных генов и другие генетические элементы, известные специалистам. Обзор биосинтетических путей Как показано на фиг. 1-3 и 11-13, для получения монатина или промежуточных продуктов его биосинтеза, таких как индол-3-пируват или ПМ, можно использовать разные пути биосинтеза. Для превращения каждого из субстратов (глюкоза, триптофан, индол-3-лактат, индол-3-пируват и ПМ) в каждый продукт (триптофан, индол-3-пируват, ПМ и монатин) можно использовать несколько разных полипептидов. Кроме того, эти реакции можно проводить in vivo, in vitro, или комбинируя реакции in vivo с реакциями in vitro, например, реакциями in vitro, которые включают неферментативные химические реакции. Таким образом, фиг. 1-3 и 11-13 являются примерными и показывают разнообразные множественные пути, которые можно использовать для получения желаемых продуктов. Превращение глюкозы в триптофан Многие организмы способны синтезировать триптофан из глюкозы. Конструкт(ы), содержащий ген(ы), необходимые для получения монатина, ПМ и/или индол-3-пирувата из глюкозы и/или триптофана, можно клонировать в таких организмах. В настоящем документе показано, что триптофан можно превратить в монатин. В других примерах организм конструируется с помощью известных полипептидов для продукции триптофана или гиперпродукции триптофана. Например, патент США No. 4371614 (в настоящем документе цитируется по ссылке) описывает штамм Е. coli, трансформированный плазмидой, содержащий оперон триптофана дикого типа. Максимальные титры триптофана, указанные в патенте США No 4371614 составляют примерно 230E.coli, который генетически сконструирован для получения триптофана, и обсуждает увеличение ферментативной продукции L-триптофана. Специалисты узнают, что организмы, способные продуцировать триптофан из глюкозы также способны утилизовать другие углеродные и энергетические ресурсы, которые могут превращать глюкозу или фруктозо-6-фосфат с такими же результатами. Примерами углеродных и энергетических ресурсов могут служить сахароза, фруктоза, крахмал, целлюлоза, глицерин и др. Превращение триптофана в индол-3-пируват Некоторые полипептиды можно использовать для превращения триптофан в индол-3-пируват. Примерами таких полипептидов могут служить ферменты, относящиеся по Международной классификации (ЕС) к классам 2.6.1.27, 1.4.1.19, 1.4.99.1, 2.6.1.28, 1.4.3.2, 1.4.3.3, 2.6.1.5, 2.6.1.-, 2.6.1.1, и 2.6.1.21. Эти классы включают полипептиды, называемые триптофан-аминотрансфераза (называемая также Lфенилаланин-2-оксоглутарат-аминотрансфераза,триптофантрансаминаза,5-гидрокситриптофанкетоглутарат-трансаминаза, гидрокситриптофан-аминотрансфераза, L-триптофан-аминотрансфераза, Lтриптофан-трансаминаза и L-триптофан-2-оксоглутарат-аминотрансфераза), которые превращают Lтриптофан и 2-оксоглутарат в индол-3-пируват и L-глутамат; D-триптофан-аминотрансфераза, которая превращает D-триптофан и 2-оксо-кислоту в индол-3-пируват и аминокислоту; триптофан-дегидрогеназаL-аминокислота:кислород оксидоредуктаза (деаминирующая, которая превращает L-аминокислоту,Н 2 О и О 2 в 2-оксокислоту и NH3 и Н 2 О 2; оксидаза D-аминокислот (также называемая оксидаза офиоаминокислот и D-аминокислота:кислород оксидоредуктаза (деаминирующая, которая превращает Dаминокислоту, Н 2 О и О 2 в 2-оксокислоту и NH3 и Н 2 О 2; и триптофан-оксидаза, которая превращает Lтриптофан, Н 2 О и О 2 в индол-3-пируват, NH3 и Н 2 О 2. Эти классы включают также тирозин (ароматическую) аминотрансферазу, аспартат-аминотрансферазу, D-аминокислот (или D-аланин) аминотрансферазу, и аминотрансферазу широкого спектра (множественных субстратов), которая проявляет множественную аминотрансферазную активность, и, в частности, может превращать триптофан и 2-оксокислоту в индол-3-пируват и аминокислоту. Одиннадцать ферментов класса аминотрансфераз, которые имеют такую же активность, описаны ниже в примере 1, в том числе новая аминотрансфераза, представленная в SEQ ID : 11 и 12. Поэтому данное изобретение представляет изолированные нуклеотидные и белковые последовательности, которые имеют не менее 80%, не менее 85%, не менее 90%, не менее 95%, не менее 98%, или даже не менее 99% идентичность с последовательностями SEQ ID : 11 и 12. Кроме того, входящие в состав этого изобретения фрагменты и продукты слияния SEQ ID : 11 и 12, которые сохраняют активность аминотрансферазы или кодируют белок с аминотрансферазной активностью. Примерами фрагментов могут служить, в частности, непрерывные нуклеотидные фрагменты длиной не менее 10, 12, 15, 20, 25, 50, 100,200, 500 или 1000 нуклеотидов из SEQ ID : 11 или непрерывные цепочки длиной не менее 6, 10, 15, 20,25, 50, 75, 100, 200, 300 или 350 аминокислот из SEQ ID12. Раскрываемые последовательности (и соответствующие варианты, фрагменты и фузионные белки) могут быть частью вектора. Вектор можно использовать для трансформации клеток хозяина, продуцируя тем самым рекомбинантные клетки, которые могут синтезировать индол-3-пируват из триптофана, а в некоторых примерах продолжать синтез дальше до ПМ и/или монатина. Известны оксидазы L-аминокислот (1.4.3.2), и их последовательности можно получить из разных источников, таких как Vipera lebetine (sp P81375), Ophiophagus hannah (sp P81383), Agkistrodoncresentus, Chlamydomonas reinhardtii, Mus musculus, Pseudomonas syringae и Rhodococcus str. Кроме того, в литературе описаны и могут быть выделены триптофан-оксидазы, например, из Coprinuis sp. SF-1, китайской капусты с болезнью клубней, Arabidopsis thaliana, и печени млекопитающих. Ниже в примере 3 обсуждается один фермент из класса оксидаз L-аминокислот, который превращает триптофан в индол-3 пируват, а также альтернативные источники для молекулярного клонирования. Многие гены оксидаз Dаминокислот помещены в доступные базы данных для молекулярного клонирования. Известны триптофан-дегидрогеназы, и их можно выделить, например, из шпината, Pisum sativum,Prosopis juliflora, гороха, пшеницы, маиса, томата, табака, Chromobacterium violaceum и Lactobacilli. Известны последовательности многих дегидрогеназ D-аминокислот. Как показано на фиг. 11-13, если для превращения триптофана в индол-3-пируват используется оксидаза аминокислот, например триптофан-оксидаза, то для снижения или даже устранения перекиси водорода можно добавить каталазу. Превращение индол-3-лактата в индол-3-пируват Реакция, которая превращает индол-3-лактат в индол-3-пируват, может катализироваться различными полипептидами, такими как полипептиды классов 1.1.1.110, 1.1.1.27, 1.1.128, 1.1.2.3, 1.1.1.222,1.1.1.237, 1.1.3.- или 1.1.1.111. Класс полипептидов 1.1.1.110 включает индоллактат-дегидрогеназы (также называемые индоллактат:NAD+-оксидоредуктаза). Классы 1.1.1.27, 1.1.1.28 и 1.1.2.3 включают лактатдегидрогеназы (также называемые лактат-дегидрогеназы, лактат:NAD+-оксидоредуктаза). Класс 1.1.1.222 содержит (R)-гидроксифениллактат-дегидрогеназу (также называемую D-ароматическая лактат дегидрогеназа,R-ароматическая лактат-дегидрогеназа иR-3-(4-гидроксифенил)-лактат:NAD(Р)+-2 оксидоредуктаза), а класс 1.1.1.237 содержит 3-(4-гидроксифенилпируват)-редуктазу (также называемую гидроксифенилпируват-редуктаза и 4-гидроксифениллактат:NAD+-оксидоредуктаза). Класс 1.1.3.- содержит лактат-оксидазы, а класс 1.1.1.111 содержит (3-имидазол-5-ил)-лактат-дегидрогеназы (также называемые (S)-3-имидазол-5-ил)-лактат:NAD(Р)+-оксидоредуктаза). Вероятно, некоторые полипептиды в этих классах способны осуществлять синтез индол-3-пирувата из индол-3-лактата. Примеры такого превращения представлены в примере 2. Для превращения индол-3-лактата в индол-3-пируват сложно применять и химические реакции. Такие химические реакции включают этап окисления, который можно проводить несколькими способами,например, окисление воздухом с применением катализатора В 2 (China Chemical Reporter, v.13, n28, p.18(1), 2002), разбавленного перманганата и перхлората или перекиси водорода в присутствии металлических катализаторов. Превращение индол-3-пирувата в 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту (ПМ) Для превращения индол-3-пирувата в ПМ можно использовать несколько известных полипептидов. Такие полипептиды могут принадлежать классам 4.1.3.-, 4.1.3.16, 4.1.3.17 и 4.1.2.-. В эти классы входят углерод-углерод синтазы/лиазы, такие как альдолазы, которые катализируют реакцию конденсации двух субстратов - карбоксиловых кислот. Класс пептидов ЕС 4.1.3.- - это синтазы/лиазы, которые образуют углерод-углеродные связи с использованием в качестве электрофильного агента оксо-кислотных субстратов (таких как индол-3-пируват), тогда как класс ЕС 4.1.2.- - это синтазы/лиазы, которые образуют углерод-углеродные связи с использованием в качестве электрофильного агента альдегидных субстратов(таких как бензальдегид). Например, полипептид, описанный в ЕР 1045-029 (ЕС 4.1.3.16, 4-гидрокси-2-оксоглутаратглиоксилат-лиаза, также называемая 4-гидрокси-2-оксоглутарат-альдолаза, 2-оксо-4-гидроксиглутаратальдолаза или КГГ-альдолаза), превращает глиоксиловую кислоту и пируват в 4-гидрокси-2 кетоглутаровую кислоту и полипептид 4-гирокси-4-метил-2-оксоглутарат-альдолаза (ЕС 4.13.17, также называемая 4-гидрокси-4-метил-2-оксоглутарат пируват-лиаза или ProA-альдолаза), конденсирует две кето-кислоты, такие как два пирувата в 4-гидрокси-4-метил-2-оксоглутарат. Реакции с использованием таких лиаз описаны в настоящем документе. На фиг. 1-2 и 11-13 показаны схематические изображения этих реакций, при которых 3-углеродная молекула (С 3) соединяется с индол-3-пируватом. Многие представители классов ЕС 4.1.2.- и 4.1.3.-, в частности, ПФ-утилизирующие полипептиды, могут использовать С 3-молекулы, которые являются аминокислотами, такими как серин, цистеин и аланин или их производные. Реакции альдольной конденсации, катализируемые представителями ЕС 4.1.2.- и 4.1.3.-, требуют, чтобы трехуглеродной молекулой был пируват или его производное. Однако и другие вещества могут служить источником трехуглеродной цепочки и превращаться в пируват. Аланин может трансаминироваться многими ПФ-утилизирующими трансаминазами, включая многие из тех, что были упомянуты выше, и превращаться в пируват. Пируват и аммоний могут быть получены в результате реакции бета-элиминации (например, катализируемой,триптофаназой или -тирозиназой) L-серина, L-цистеина, и производных серина и цистеина с соответсвующими группами, таких как О-метил-L-серин, О-бензил-L-серин, S-метилцистеин, S-бензилцистеин,S-алкил-L-цистеин, О-ацил-L-серин, и 3-хлор-L-аланин. Аспартат может служить источником пирувата в ПФ-опосредованных бета-лиазных реакциях, таких как катализируемые триптофаназой (ЕС 4.1.99.1) и/или -тирозиназой (ЕС 4.1.99.2. также называемой тирозин-фенол-лиаза). Скорость бета-лиазных реакций можно увеличить с помощью сайт-направленного мутагенеза полипептидов классов 4.1.99.1-2 как описано в работе Mouratou с соавт. (J. Biol. Chem. 274:1320-1325, 1999) и в примере 8. Эти модификации позволяют полипептидам воздействовать на дикарбоксильные аминокислоты в качестве субстратов. Лактат также может служить источником пирувата, и он окисляется до пирувата при добавлении лактат-дегидрогеназы и окисляющего кофактора или лактат-оксидазы и кислорода. Примеры подобных реакций приведены ниже. Например, как показано на фиг. 2 и фиг. 11-13, ProA альдолаза может контактировать с индол-3-пируватом, если в качестве С 3-молекулы используется пируват. ПМ также может образоваться в результате химических реакций, например, альдольной конденсации, представленной в примере 5. Превращение ПМ в монатин Превращение ПМ в монатин может катализироваться одним или несколькими из следующих ферментов: триптофан-аминотрансферазы (2.6.1.27), триптофан-дегидрогеназы (1.4.1.19), дегидрогеназы Dаминокислот (1.4.99.1), глутамат-дегидрогеназы (1.4.1.2-4), фенилаланин-дегидрогеназа (ЕС 1.4.1.20),триптофан-фенилпируват-трансаминазы (2.6.1.28), или чаще представителями семейства аминотрансфераз (2.6.1.-), такими как аспартат-аминотрансфераза (ЕС 2.6.1.1), тирозин (ароматическая) аминотрансфераза (2.6.1.5), D-триптофан-аминотрансфераза или D-аланин (2.6.1.21) аминотрансфераза (фиг. 2). Ниже описаны одиннадцать представителей класса аминотрансфераз (пример 1), включая новый фермент этого класса, описанный в SEQ ID : 11 и 12, а реакции, демонстрирующие активность ферментов аминотрансферазы и дегидрогеназы, представлены в примере 7. Эта реакция может быть проведена и химическими способами. Аминирование кетокислоты (ПМ) проводится восстановительным аминированием с помощью аммония и цианоборгидрида натрия. На фиг. 11-13 показаны другие полипептиды, которые можно использовать для превращения ПМ в монатина также для увеличения выхода монатина при получении его из индол-3-пирувата или триптофана. Например, если в качестве донора аминогруппы используется аспартат, то для превращения аспартата в оксалоацетат можно использовать аспартат-аминотрансферазу (фиг. 11). Оксалоацетат превращается в пируват и двуокись углерода под действием декарбоксилазы, например оксалоацетат-декарбоксилазы(фиг. 11). Кроме того, если в качестве донора аминогруппы используется лизин, то для превращения лизина в аллизин можно использовать лизин-эпсилон-аминотрансферазу (фиг. 12). Аллизин спонтанно пре- 13009284 вращается в 1-пиперидин-6-карбоксилат (фиг. 12). Если для превращения ПМ в монатин используется полипептид, способный катализировать реакции восстановительного аминирования (например, глутаматдегидрогеназа), то можно использовать полипептид, который способен осуществлять рециклизациюNAD(P)H и/или продуцировать летучий продукт (фиг. 13), например формиат-дегидрогеназу. Дополнительные замечания по устройству биосинтетических путей В зависимости от того, какие полипептиды используются для образования индол-3-пирувата, ПМ и/или монатина, для повышения выхода продукта реакции продуцирующей клетке можно предоставить кофакторы, субстраты и/или дополнительные полипептиды. Удаление перекиси водорода Перекись водорода (Н 2 О 2) - это продукт, который в случае его образования является токсичным для продуцирующих клеток и может повредить полипептиды или получающиеся промежуточные продукты. Оксидаза L-аминокислот, описанная выше, образует Н 2 О 2 в качестве одного из продуктов. Поэтому при применении оксидазы L-аминокислот образующуюся Н 2 О 2 можно удалять или снижать ее концентрацию, чтобы уменьшить уровень повреждения клеток или продуктов. Для снижения уровней Н 2 О 2 в клетке можно использовать каталазы (фиг. 11-13). Продуцирующая клетка может экспрессировать ген или последовательность кДНК, которая кодирует каталазу (ЕС 1.11.1.6), катализирующую разложение перекиси водорода на воду и газооразный кислород. Например,каталазу может экспрессировать вектор, трансфицированный в продуцирующую клетку. В качестве примера каталаз, которые используются для этого, можно назвать, но не ограничиваясь, следующие:trQ9EV50 (Staphyhcoccus xylosus), trQ9KBE8 (Bacillus halodurans), trQ9URJ7 (Candida albicans),trP77948 (Streptomyces coelicolor), trQ9RBJ5 (Xanthomonas campestris) (приведены номера Accessionпо базе данных SwissProt). Биокаталитические реакторы, исолпьзующие оксидазу L-аминокислот, оксидазу D-аминокислот или триптофан оксидазу также могут содержать полипептид каталазу. Модулирование доступности ПФ (пиридоксаль-5'-фосфата) Как показано на фиг. 1, ПФ может применяться на одном или нескольких этапах биосинтеза, описанных в настоящем документе. Концентрация ПФ может быть увеличена, чтобы ПФ не стал лимитирующим фактором проведения реакции. Биосинтетический путь для витамина В 6 (предшественника ПФ) хорошо изучен на примере Е. coli и некоторые участвующие в нем белки выделены в кристаллическом виде (Labor с соавт., FEBS Letters,449:45-48, 1999). В других метаболических путях участвуют два гена (epd или gapB и serC), в то время как только для биосинтеза пиридоксаль-фосфата требуется три гена (pdxA, pdxB mpdxJ). Одним из стартовых материалов для пути Е. coli является 1-дезокси-D-ксилозо-3-фосфат (DXP). Синтез этого предшественника из общих 2- и 3-углеродных центральных метаболитов катализируется полипептидом 1 дезокси-D-ксилозо-5-фосфат синтазой (DSX). Другим предшественником является производное треонина, полученное из 4-углеродного сахара, D-эритрозо 4-фосфата. Гены, необходимые для превращения в фосфо-4-гидроксил-L-треонин (НТР), - epd, paxB и serC. Последняя реакция для образования ПФ - сложная внутримолекулярная конденсация и реакция замыкания кольца между DXP и НТР, катализируемая продуктами генов pdxA и pdxJ. Если ПФ становится лимитирующим фактором в процессе ферментации при продукции монатина,повышенная экспрессия одного или нескольких генов, отвечающих за синтез в продуцирующей клетке хозяина, может быть использована для увеличения выхода монатина. Организм хозяина может содержать множественные копии генов его нативных путей, или копии генов ненативных путей можно включить в геном хозяина. Кроме того, множественные копии генов усовершенствованных метаболических путей можно встроить в организм хозяина. Один из усовершенствованных путей, который сохраняется во всех организмах, повторно использует различные производные витамина В 6 и переводит их в активную форму ПФ. Полипептиды, участвующие в этом пути, -pdxK киназа, pdxH оксидаза, pdxY киназа. Гиперэкспрессия одного или нескольких из этих генов может увеличить доступность ПФ. Уровни витамина В 6 можно увеличить путем удаления или подавления метаболической регуляции генов, отвечающих за нативные биосинтетические пути в организме хозяина. ПФ подавляет полипептиды, вовлеченные в биосинтез предшественника - производного треонина в бактериях Flavobacterium sp.,штамм 238-7. Этот штамм, освобожденный от метаболического контроля, осуществляет гиперпродукцию пиридоксальпроизводных и может выделять до 20 мг/л ПФ. Сходные генетические манипуляции с организмом хозяина, продуцирующего монатин, позволят повысить продукцию ПФ без гиперэкспрессии генов биосинтетических путей. Утилизация аммония Реакции с участием триптофаназы можно проводить в направлении синтеза (продукция триптофана из индола), если сделать аммоний более доступным, или путем удаления воды. Реакции восстановительного аминирования, например, катализируемые глутамат-дегидрогеназой, также могут быть направлены в одну сторону избытком аммония. Доступность аммония можно повысить путем добавления солей карбоната аммония или фосфата аммония в карбонат- или фосфат-буферной системе. Аммоний также можно ввести в виде пирувата ам- 14009284 мония или формиата аммония. Другим способом аммоний можно доставить, если реакция проводится в сопряжении с реакцией образования аммония, например, с участием глутамат-дегидрогеназы или триптофан-дегидрогеназы. Аммоний может образоваться при добавлении натуральных субстратов для ЕС 4.1.99.-(тирозин или триптофан), которые будут гидролизованы до фенола или индола, пирувата и NH3. Это также даст возможность повысить выход продуктов синтеза по сравнению с нормальным равновесным количеством за счет предоставления возможности ферменту гидролизовать свой предпочтительный субстрат. Удаление продуктов и побочных продуктов Превращение триптофана в индол-3-пируват через триптофан-аминотрансферазу может оказать побочный эффект в виде влияния на скорость продукции индол-3-пирувата, так как реакция приводит к образованию глутамата и требует наличия косубстрата 2-оксоглутарата (-кетоглутарата). Глутамат может вызвать подавление аминотрансферазы, и в результате реакции будет уничтожено большое количество косубстрата. Кроме того, высокие концентрации глутамата мешают дальнейшим процессам разделения. Полипептид глутамат-дегидрогеназа (GLDH) превращает глутамат в 2-оксоглутарат, приводя к повторному использованию этого косубстрата в реакции, катализируемой триптофан-аминотрансферазой. Кроме того, GLDH образует восстановленных эквивалентов (NADH или NADPH), которые могут использоваться в качестве источников энергии в клетке в аэробных условиях. Утилизация глутамата ферментом GLDH также уменьшает образование побочных продуктов. К тому же в результате реакции образуется аммоний, который служит источником азота для клетки или субстратом для восстановительного аминирования на заключительном этапе, показанном на фиг. 1. Поэтому клетка-продуцент, которая производит гиперэкспрессию GLDH-полипептида, может использоваться для увеличения выхода и снижения стоимости сред и/или процессов разделения. На пути превращения триптофана в монатин донор аминогруппы на этапе 3 (например, глутамат или аспартат) может быть вновь превращен в аминоакцептор для этапа 1 (например, 2-оксоглутарат или оксалоацетат), если используется аминотрансфераза из соответствующего класса ферментов. Использование двух разных трансаминаз для этого пути, где субстрат одной трансаминазы неконкурентно подавляет активность другой трансаминазы, может увеличить эффективность этого пути. Многие из реакций описанных путей обратимы и поэтому достигнуто равновесие между субстратами и продуктами. Выход продукта на этом пути может быть увеличен путем постоянного удаления продуктов от полипептидов. Например, секреция монатина в ферментационный бульон с помощью пермеазы или другого транспортного белка или избирательной кристаллизации монатин из потока биокаталитического реактора с сопутствующим повторным использованием субстратов увеличит выход реакции. Удаление побочных продуктов с помощью добавочных каталитических реакций или путем замещения донорских аминогрупп - еще один способ увеличения выхода реакции. Несколько примеров на эту тему обсуждаются в примере 13 и показаны на фиг. 11-13. В идеале в ходе реакции образуется побочный продукт, который не доступен для реакции в обратном направлении, либо из-за фазового перехода (испарение) или благодаря спонтанному превращению в нереагирующий конечный продукт, например, двуокись углерода. Модулирование субстратных пулов Пул индола можно модулировать путем увеличения продукции предшественников триптофана и/или изменения катаболических путей, использующих индол-3-пируват и/или триптофан. Например,продукция индол-3-уксусной кислоты из индол-3-пирувата может быть снижена или прекращена при функциональном отключении гена, кодирующего фермент класса ЕС 4.1.1.74 в клетке хозяина. Продукция индола из триптофана может быть снижена или прекращена при функциональном отключении гена,кодирующего фермент класса ЕС 4.1.99.1 в клетке хозяина. Альтернативно, избыток индола можно утилизовать в качестве субстрата в процессах in vitro или in vivo в комбинации с повышенным количеством гена, кодирующего фермент класса ЕС 4.1.99.1 (Kawasaki с соавт., J. Ferm. и Bioeng. 82:604-6, 1996). Можно провести генетические модификации для увеличения уровня промежуточных продуктов, таких как D-эритрозо-4-фосфата и хоризмата. Синтез триптофана регулируется у большинства организмов. Один из механизмов осуществляется через подавление обратной связи для некоторых ферментов этого метаболического пути; поскольку уровень триптофана возрастает, скорость его синтеза снижается. Так, при использовании клеток хозяина,сконструированных для продукции монатина через промежуточный продукт триптофан, можно использовать организм, который нечувствителен к концентрациям триптофана. Например, штамм Catharanthusroseu, который устойчив к подавлению роста разными аналогами триптофана, был выведен путем повторного воздействия высокими концентрациями 5-метилтриптофана (Schallenberg, Berlin,Z.Naturforsch 34:541-5,1979). На активность полученной триптофан-синтазы их этого штамма мало влияет подавление продуктом, благодаря наличию мутаций в гене. Аналогичным образом можно оптимизировать клетку хозяина, используемую для продукции монатина. Продукцию триптофана можно оптимизировать с помощью эволюции, направленной на выявление- 15009284 полипептидов, которые менее чувствительны к подавлению их продуктами реакций. Например, можно провести скрининг на плашках, среда в которых не содержит триптофана, но обогащена неметаболизируемыми аналогами триптофана в высоких концентрациях. Патенты США 5756345, 4742007 и 4371614 описывают способы повышения продуктивности ферментационных организмов по триптофану. Последним этапом биосинтеза триптофана является присоединение серина к индолу; потому для увеличения продукции триптофана можно увеличить доступность серина. Количество монатина, продуцируемого ферментационным организмом, может быть увеличено путем повышения количества пирувата, синтезируемого в организме хозяина. Некоторые дрожжи, такие как Trichosporon cutaneum (Wang с соавт., Lett. Appl Microbiol 35:338-42,2002) и Torulopsis glabrata (Li с соавт., Appl Microbiol. Biotechnol. 57:451-9,2001) осуществляют гиперпродукцию пирувата и могут быть использованы для реализации способов, изложенных в настоящем документе. Кроме того, можно провести генетические модификации организмов, способствующие продукции пировиноградной кислоты,такие как в случае с штаммом E.coli W1485lip2 (Kawasaki с соавт. J. Ferm. и Bioeng. 82:604-6,1996). Контроль хиральности Вкусовой профиль монатина можно изменить, управляя стереохимическими параметрами (хиральностью) этой молекулы. Например, разные изомеры монатина могут требоваться в разных концентрациях для разных пищевых систем. Хиральность можно контролировать с помощью комбинации pH и полипептидов. Рацемизация в положении С-4 монатина (см. выше на рисунке) может возникнуть в результате депротонизации и репротонизации альфа-углерода, что может произойти при сдвиге pH или в результате реакции с кофактором ПФ. Маловероятно, что у микроорганизма произойдет смещение pH, достаточное для осуществления рацемизации, но ПФ находится в избытке. Способы контроля хиральности полипептидами зависят от биосинтетического пути, использующего продукцию монатина. Если монатин образуется в результате реализации пути, представленного на фиг. 2, можно рассмотреть следующий вариант. При биокаталитической реакции хиральность углерода-2 определяется ферментом, который превращает индол-3-пируват в ПМ. Многие ферменты (например, из классов ЕС 4.1.2.-,4.1.3.-) могут превращать индол-3-пируват в ПМ. Поэтому можно выбрать фермент, который образует нужный изомер. Альтернативно, энантиоспецифичность фермента, который превращает индол-3-пируват в ПМ можно модифицировать путем направленной эволюции или с помощью каталитических антител катализирующих нужную реакцию. Как только получен ПМ (ферментативным путем или в результате химической конденсации), можно стереоспецифически добавить аминогруппу с помощью одной из трансаминаз, описанных в настоящем документе. R-или S-конфигурация углерода-4 может быть создана в зависимости от того, используется для проведения реакции аминотрансфераза D- или L-ароматических аминокислот. Большинство аминотрансфераз специфически действуют на L-изомер, однако в некоторых растениях имеются D-триптофан аминотрансферазы (Kohiba, Mito, Proceedings of the 8th InternationalD-аланин аминотрансфераза (2.6.1.21), D-метионин-пируват-аминотрансфераза (2.6.1.41), а также (R)-3 амино-2-метилпропаноат-аминотрансфераза (2.6.1.61) и (S)-3-амино-2-метилпропаноат-аминотрансфераза (2.6.1.22). Некоторые аминотрансферазы могут воспринимать субстрат для этой реакции только в конкретной конфигурации С 2-углерода. Поэтому, даже если превращение в ПМ не является стереоспцифическим, стереохимия конечного продукта может контролироваться благодаря правильному выбору трансаминазы. Поскольку эти реакции обратимы, то непрореагировавший ПМ (нежелательный изомер) может разлагаться на исходные составные части и рацемическую смесь ПМ можно приготовить заново. Активация субстратов Фосфорилированные субстраты, такие как фосфоенолпируват (PEP), можно использовать в реакциях, описанных в настоящем документе. Фосфорилированные субстраты могут оказаться энергетически более предпочтительными, и поэтому могут использоваться для увеличения скорости и/или выхода реакции. В реакциях альдольной конденсации добавление фосфатной группы стабилизирует нуклеофильный субстрат, делая его более реакционноспособным. В других реакциях, фосфорилированный субстрат часто обеспечивает более удобное удаление группы. Аналогичным образом субстраты можно активировать путем превращения их в производные CoA или пирофосфатов.- 16009284 Пример 1. Клонирование и экспрессия триптофан-аминотрансферазы. В этом примере описаны способы, которые используются для клонирования триптофанаминотрансфераз, которые можно применять для превращения триптофана в индол-3-пируват. Обзор экспериментальных данных Одиннадцать генов, кодирующих аминотрансферазы, клонированы в E.coli. Этими генами были Dаланин-аминотрансфераза (dat, Genbank Accession No. Y14082.1 п.н. 28622-29470 и Genbank AccessionNo. NP 388848.1, нуклеотидная и аминокислотная последовательности, соответственно) из Bacillus subtilis, тирозин-аминотрансфераза (tatA, SEQ ID : 1 и 2, нуклеотидная и аминокислотная последовательности, соответственно) из Sinorhizobium meliloti (также называемого Rhisobitan meliloti), тирозинаминотрансфераза (talA заявлена по гомологии, SEQ ID : 3 и 4, нуклеотидная и аминокислотная последовательности, соответственно), из Rhodobacter sphaeroides штамм 2.4.1, тирозин аминотрансфераза (заявлена по гомологии, SEQ ID : 5 и 6, нуклеотидная и аминокислотная последовательности соответственно) из R. sphaeroides 35053, аминотрансфераза широкого спектра действия (bsat, заявлена по гомологии с фрагментами пептида из L.mexicana, SEQ ID : 7 и 8, нуклеотидная и аминокислотная последовательности, соответственно) из Leishmania major, ароматическая аминотрансфераза (araT, заявлена по гомологии, SEQ ID : 9 и 10, нуклеотидная и аминокислотная последовательности, соответственно) изBacillus subtilis; ароматическая аминотрансфераза (araT заявлена по гомологии, SEQ ID : 11 и 12, нуклеотидная и аминокислотная последовательности, соответственно) из Lactobacillus amylovorus, аминотрансфераза множественных субстратов (заявлена по гомологии, SEQ ID : 13 и 14, нуклеотидная и аминокислотная последовательности, соответственно) из R. sphaeroides 35053, аминотрансфераза множественных субстратов (msa заявлена по гомологии, Genbank Accession No. AAAE01000093.1, п.н. 1474316155 и Genbank Accession No. ZP00005082.1, нуклеотидная и аминокислотная последовательности, соответственно) из Rhodobacter sphaeroides штамм 2.4.1, аспартат-аминотрансфераза (aspC, Genbank Accession No. AE000195.1 п.н. 2755-1565 и Genbank Accession No. AAC74014.1, нуклеотидная и аминокислотная последовательности (соответственно) из Escherichia coli и тирозин аминотрансфераза (tyrB, SEQ ID: 31 и 32, нуклеотидная и аминокислотная последовательности, соответственно) из Е. coli. Гены клонировали, экспрессировали и тестировали на способность превращать триптофан в индол 3-пируват параллельно с коммерчески доступными ферментами. Все одиннадцать клонов показали ферментную активность. Идентификация бактериальных штаммов, которые могут содержать полипептиды с нужной активностью В базе данных NCBI (Национального Центра биотехнологической информации США) нет ни одного гена, продукт которого носит название триптофан-аминотрансферазы. Однако идентифицированы организмы, обладающие этой ферментативной активностью. Активность L-триптофан-аминотрансферазы (ТАТ) измеряли в клеточных экстрактах или в препаратах очищенного белка из следующих источников: ризобактериальный изолят из Festuca octoflora, митохондрии и цитоплазма гороха, клетки подсолнечника, Rhizobium leguminosarum сорт trifoli, Envinia herbicoia pv gypsophilae, Pseudomonas syringae, Agrobacterium tumefaciens, Azospirillum lipferumbrazilense, Enterobacter cloacae, Enterobacter agglomerans, Bradirhizobium elkanii, Candida maltosa, Azotobacter vinelandii, головной мозг крыс, печень крыс, Sinorhizobium meliloti, Pseudomonas fluorescens CHAO, Lactococcus lactis, Lactobacillus casei, Lactobacillus helveticus, проросшие зерна пшеницы, ячмень, Phaseolus aureus (бобы), Saccharomyces uvarumStreptomyces griseus. Выделение геномной ДНК для клонированияS. meliloti (ATCC номер 9930) выращивали на среде TY при 25 С, рН 7,2. Клетки растили до оптической плотности при 600 нм(OD600) 1,85 и 2% инокулят использовали для приготовления геномной ДНК. Для выделения геномной ДНК использовали набор Qiagen genomic tip 20/G (Валенсия, Калифорния).Bacillus subtilis 6051 (ATCC) выращивали при 30 С на питательном бульоне Bereto (Difco; Детройт,Мичиган). Протокол Qiagen genomic tip 20/G использовали для приготовления геномной ДНК со следующими изменениями: концентрации протеиназы K и лизоцима удваивали, а время инкубации увеличивали в 2-3 раза. Геномную ДНК из Leishmania major ATCC 50122 получали от IDI, Inc. (Квебек, Канада) в буфере ТЕ рН 8,0, концентрация 17 нг/мкл.Rhodobacter sphaeroides 2.4.1 (предоставленный профессором Сэмом Капланом, Университет Техаса, г. Хьюстон), R. sphaeroides 35053 (ATCC) и L. amylovorus-геномную ДНК готовили стандартным способом экстракции фенолом. Клетки собирали на поздней log-фазе, ресуспендировали в TEN-буфере (10 мМ трис-HCl, рН 7,5, 1 мМ ЭДТА, 100 мМ NaCl) и лизировали добавлением 0,024 мл раствора лаурилсаркозина натрия на мл клеточной суспензии. После экстракции не менее трех раз равным объемом фенола, насыщенного буфером ТЕ (10 мМ трис-HCl, рН 7,5, 1 мМ ЭДТА), раствор ДНК экстрагировали однократно смесью хлороформ-фенол (9:1) и три раза хлороформом. ДНК осаждали добавлением 0,1 объема 3 М ацетата натрия, рН 6,8 и 2 объемами этанола. Осадок собирали центрифугированием и про- 17009284 мывали 1 раз 70% этанолом. В заключение ДНК растворяли в 0,10 мл дистиллированной воды. Геномную ДНК из Escherichia coli выделяли из штамма DH10B (Invitrogen) и готовили с помощью набора Qiagen Genomic-tip (500/G). Из 30 мл этого штамма, выросшего в LB до OD650 1,87, получили 0,3 мг очищенной ДНК. Очищенную ДНК растворяли в элюционном буфере Qiagen в концентрации 0,37 мкг/мкл. Протокол полимеразной цепной реакции Конструировали праймеры с соответствующим выступом для вектора рЕТ 30 Ха/LIC вектор (Novagen, Мэдисон, Висконсин). Вектор рЕТ имеет 12-нуклеотидный одноцепочечный выступ на 5'-стороне от сайта Ха/LIC и 15-нуклеотидный одноцепочечный выступ на 3'-стороне от сайта Ха/LIC. Конструировали плазмиду для клонирования независимо от сшивки, с N-терминальным His и S-тэгами и дополнительным С-терминальным His-тэгом. Сайт узнавания протеазы Ха (IEGR) находится непосредственнно перед стартовым кодоном интересующего гена, так что тэги фузионных белков можно удалить. Добавляли следующие последовательности к 5'-концам организм-специфических последовательностей при конструировании праймеров: прямой праймер, 5' GGTATTGAGGGTCGC (SEQ ID 73); обратный праймер 5' AGAGGAGAGTTAGAGCC(SEQ ID :74).Escherichia coli tyrB: N-конец: 5'-GGTATGAGGGTCGCGTGTTTCAAAAAGTTGACGC и С-конец: 5'-AGAGGAGAGTTAGAGCCTTACATCACCGCAGCAAACG-31 (SEQ ID : 33 и 34). Ген из S. meliloti (tatA) амплифицировали с помощью следующего протокола ПЦР: использовали в 50 мкл реакц. 0,1-0,5 мкг матрицы, 1,5 мкМ каждого праймера, 0,4 мМ каждого dNTP, 3,5 U высокоточной полимеразы Expand (Roche, Indianapolis, IN), и 1X Expand буфер с Mg. Использованная программа термоциклера включала горячий старт при 96 С в течение 5 мин с последующими 29 повторами следующих этапов: 94 С в течение 30 с, 55 С в течение 2 мин и 72 С в течение 2,5 мин. После 29 повторов образец выдерживали при 72 С в течение 10 мин и затем хранили при 4 С. Этот ПЦР-протокол давал продукт размером 1199 п.н. Последовательности генов из R.sphaeroides (msa и tatA), L. amylovorus araT, и Bacillus araT амплицифировали с помощью следующего протокола ПЦР. Добавляли в 50 мкл реакц. 0,1-0,5 мкг матрицы, 1,5 мкМ каждого праймера, 0,4 мМ каждого dNTP, 3,5 ед. полимеразы Expand High Fidelity и 1X Expand буфер с Mg. Термоциклер включали на горячий старт при 96 С в течение 5 мин с последующими 29 повторами следующих этапов: 94 С в течение 30 с, 40-60 С в течение 1 мин 45 с (градиентный термоциклер) и 72 С в течение 2 мин 15 с. После 29 повторов образец выдерживали при 72 С в течение 10 мин и затем хранили при 4 С. Для каждого гена R. sphaeroides msa при температурах отжига 42 и 48 С получали множественные продукты, но четкую полосу примерно в районе 1464 п.н. Для L. amylovorus araT, температуры отжига 42, 48 и 56 С давали единичные продукты с интенсивными полосами в районе 1173 п.н. Для В. subtilisaraT, температуры отжига 40, 45, 50, 55 С давали единичные продукты с интенсивными полосами в 1173 п.н., от обеих геномных ДНК и колоний. Для L.major bsat температура отжига 55 С дала чистейший продукт (1239 п.н.). Для генов Rhodobacter sphaeroides tatA температуры отжига 50-55 С давали чистый продукт с правильным размером (1260 п.н.). Для обоих генов Е. coli и гена В.subtilis dat использовали температуры отжига 55-60 С, а время отжига сокращали до 45 с. Получены чистые продукты верного размера(около 1,3 кб для генов Е. coli, и 850 п.н. для гена dat). Клонирование Продукты ПЦР очищали в агарозном геле с 0,8-1% ТАЕ с помощью набора Qiagen gel extraction(Валенсия, Калифорния). Продукты ПЦР определяли количественно путем сравнения со стандартами на агарозном геле, а затем обрабатывали Т 4 ДНК-полимеразой и рекомендованными протоколами производителя для независимого клонирования Ligation Independent Cloning (Novagen, Мэдисон, Висконсин). Около 0,2 пмоль очищенного продукта ПЦР обрабатывали 1 ед. Т 4 ДНК-полимеразы в присутствииdGTP в течение 30 мин при 22 С. Полимераза удаляет основания последовательно с 3'-концов ПЦРпродукта. Если полимераза встречает гуаниновый остаток, 5'-3'-полимеразная активность фермента нейтрализует экзонуклеазую активность и эффективно препятствует дальнейшему расщеплению. В результате получается одноцепочечные липкие концы, которые совмещаются с вектором рЕТ Ха/LIC. Полимераза инактивировали путем инкубации при 75 С в течение 20 мин. Вектор и обработанную вставку отжигали в соответствии с рекомендациями Novagen. Около 0,02 пмоль обработанной вставки и 0,01 пмоль вектора инкубировали в течение 5 мин при 22 С, добавляли 625 мМ ЭДТА (конечная концентрация) и повторяли инкубацию при 22 С. Реакцию отжига (1 мкл) добавляли к NovaBlue единичным компетентным клеткам (Novagen, Мэдисон, Висконсин) и инкубировали на льду в течение 5 мин. После перемешивания клетки трансформировали тепловым шоком в течение 30 с при 42 С. Клетки помещали на лед на 2 мин и давали восстановиться в 250 мкл при комнатной температуре SOC в течение 30 мин при 37 С с встряхиванием при 225 об./мин. Клетки помещали на плашкиLB с канамицином (25-50 мкг/мл). Плазмидную ДНК очищали с помощью набора Qiagen spin miniprep и скринировали на наличие правильных вставок, обрабатывая рестриктазами XhoI и XbaI. Последовательности плазмид с правильными вставками проверяли секвенированием терминирующей ДНК.SEQ ID : 1-14 и 31-32 представляют нуклеотидную и соответствующую аминокислотную последовательность рекомбинантных аминотрансфераз, любые отличия от последовательностей Genbank были либо молчащими, либо проявлялись консервативными заменами в белковой последовательности. SEQ ID: 11 и 12 - это новые последовательности. Экспрессия генов и способы определения Плазмидную ДНК, верифицированную с помощью анализа последовательности, субклонировали в экспрессирующие клетки Е. coli BLR(DE3) или BL21(DE3) (Novagen, Мэдисон, Висконсин). Выращивали культуры и выделяли плазмиды с помощью набора Qiagen miniprep, а затем проводили рестрикционный анализ для подтверждения идентичности. Индукцию проводили сначала с L. amylovorus araT, В. subtilis araT, и R.meliloti tatA в клеткахBLR(DE3) и BL21(DE3). Временное исследование проводили на культурах, растущих на LB с канамицином (30 мг/л) до OD600 от 0,5-0,8, индуцированных 1 мМ IPTG (изопропилфлуогалактозид) и отобраны через 0, 1, 2 и 4 ч после индукции. Клетки от 2,0 мл ресуспендировали в 0,10 мл 120 мМ трис-HCl, pH 6,8, содержащем 10% додецилсульфат натрия, 10% бета-меркаптоэтанол и 20% глицерин, грели при 95 С в течение 10 мин, охлаждали и разводили 0,10 мл Н 2 О. Аликвоты этих образцов с общим клеточным белком анализировали способом электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) в 4-15% градиентном геле. Не обнаружено существенных различий по количеству белка, экспрессирующегося через 2 и через 4 ч после индукции, а также между клетками BLR(DE3) и BL21(DE3). Кроме того, готовили клеточные экстракты из четырехчасовых образцов путем суспендирования клеточных осадков из 2 мл культуры в 0,25 мл реагента Novagen BugBuste, содержащего 0,25 мкл бензоназы нуклеазы, инкубации при комнатной температуре в течение 20 мин с осторожным встряхиванием и центрифугированием при 16,000 х g для удаления клеточных остатков. Супернатанты (клеточные экстракты) вводили на 4-15% градиентные гели для анализа клеточных растворимых белков. Три клона (L. amylovorus araT (SEQ ID : 11 и 12), В. subtilis araT (SEQ ID : 9 и 10) и R.melilotitatA (SEQ ID : 1 и 2) показали наличие растворимого белка, который соответствует заданному размеру(около 45 кДа). Продукт гена В. subtilis araT был гиперэкспрессирован на самом высоком уровне и/или был более растворим, чем два других белка. Способами последовательной экспрессии, плазмидную ДНК из положительных клонов субклонировали в клетки BL21(DE3) в связи с повышенными ростовыми характеристиками этих клеток. Повторили индукцию, используя 1 мМ EPTG с культурами, растущими на LB с канамицином 50 мг/л, в тот момент, когда OD600 достигла около 0,8. Клетки собирали через 4 роста при 37 С, центрифугировали при 3000 об/мин, в течение 10 мин при 4 С, промывали буфером с TEGGP (50 мМ трис-HCl (рН 7,0), 0,5 мМ ЭДТА, 100 мг/л глутатион, 5% глицерин, с полным коктейлем Roche с ингибитором протеазы) и замораживали в этаноле при -80 С. Образцы ресуспендировали в реагенте BugBuster (Novagen) (5 мл/г сырого веса клетки), содержащем 5 мкл/мл коктейля с ингибитором протеазы 3 (Calbiochem-Novabiochem Corp., Сан-Диего, Калифорния) и 1 мкг/мл бензоназы нуклеазы. Образцы инкубировали при комнатной температуре в течение 20 мин на вращающемся шейкере. Нерастворимую клеточную взвесь удаляли центрифугированием при 16,000 X g в течение 20 мин при 4 С. Клеточные экстракты анализировали способом SDS-PAGE и определяли активность триптофан- 19009284 аминотрансферазы по продукции индол-пирувата по следующему протоколу. Одномиллилитровые реакции проводили с 50 мМ тетрабората натрия (рН 8,5), 0,5 мМ ЭДТА, 0,5 мМ арсената натрия, 50 мкМ пиридоксальфосфата, 5 мМ альфа-кетоглутарата и 5 мМ L-триптофана. Реакции инициировали добавлением бесклеточных экстрактов или очищенного фермента и инкубировали 30 мин при 30 С. 20% ТСА (200 мкл) добавляли, чтобы остановить реакцию и осажденный белок удаляли центрифугированием. Измеряли поглощение при 327 нм и сравнивали со стандартной кривой свежеприготовленного раствора индол 3-пирувата в буфере определения. Кроме того, проводились контрольные реакции без субстрата триптофана или с использованием бесклеточных экстрактов из клонов, трансформированных одним рЕТ 30 а. Из-за наличия фоновой активности нативных аминотрансфераз Е. coli в клеточных экстрактах рекомбинантные белки очищали с помощью картриджей His-Bind в соответствии с протоколами производителя (Novagen, Мэдисон, Висконсин). Фракции элюента обессоливали на колонках PD-10 (AmershamBiosciences, г.Пискатавей, Нью-Джерси) и элюировали в 50 мМ трис-буфере, рН 7,0. Очищенные белки анализировали способом SDS-PAGE и определяли аминотрансферазную активность. Результаты индукции при 37 С с 1 мМ IPTG (4 ч) демонстрируют, что L. major bsat, S. meliloti tatA,E.coli aspC, и оба клона из R. sphaeroides tatA показывают значительные уровни активности триптофанаминотрансферазы. Белок araT из В. subtilis был переэкспрессирован и растворим, но показал низкую ферментативную активность. Продукт гена L. amylovorus araT оказался растворимым в клеточном экстракте, но очистка с помощью картриджа His-Bind привела к выходу малого количества белка с нужным молекулярным весом. Продукты гена msa окзались нерастворимыми, и последующие эксперименты с экспрессией проведены при 24 С для минимизации образования инклюзионных телец. Несколько концентраций IPTG от 10 мкМ до 1 мМ использованы для повышения количества растворимого белка. В табл. 1 перечислены величины активностей, измеренные в микрограммах образовавшегося индол 3-пирувата (ИЗП) на миллиграмм белка в минуту. В некоторых случаях очень небольшое количество рекомбинантного белка показывало высокие уровни активности, которые выходили за пределы эффективного линейного диапазона определения. В этих случаях знакпредшествует величине активности. Таблица 1. Специфическая активность клонов в клеточных экстрактах (СЕ),очищенного (Р) и коммерческого ферментов Выравнивание для сравнения всех клонированных рекомбинатных белков показывает, что между последовательностями генов araT, tatA, bsat и msa немного высококонсервативных участков. Выравнивание рекомбинантных белков с более высокой активностью: гомологи продуктов гена R. sphaeroides tatA,аминотрансферазы широкого диапазона из L.major и тирозин-аминотрансферазы Sinorhizobium meliloti показало наличие нескольких высококонсервативных участков, однако они всего на 30-43% идентичны по белку. Доступность аминотрансферазы широкого диапазона и D-специфической (D-аланин) аминотрансферазы может быть полезна при получении других стереоизомеров монатина. Пример 2. Превращение индол-3-лактат в индол-3-пируват. Как показано на фиг. 1 и 3, индол-3-лактат можно использовать для получения индол-3-пируват. Превращение лактат в пируват - обратимая реакция, это верно и в отношении превращений между индол-3-пируватом и индол-3-лактатом. Окисление индол-лактата обычно происходит благодаря высокому количеству фона при 340 нм от индол-3-пирувата. Смесь для стандартного определения содержала 100 мМ фосфат калия, рН 8,0, 0,3 мМ NAD+, 7 единиц лактат-дегидрогеназы (ЛДГ) (Sigma-L2395, Сент-Луис, МО) и 2 мМ субстрат в 0,1 мл. Определение проводили дважды на УФ-проницаемой микротитровочной плашке с помощью регистратора MolecularDevices SpectraMax Plus. Полипептид и буфер смешивали и с помощью пипетки переносили в ячейки плашки, содержащие индол-3-лактат и NAD+, после чего поглощение при 340 нм в каждой ячейке считывали через интервалы в 9 с после короткого перемешивания. Реакцию проводили при 25 С в течение 5 мин. Увеличение поглощения при 340 нм сопровождает образование NADH из NAD+. Отдельно проводили реакцию для 20 отрицательных контролей без NAD+ и без субстрата. Оказалось, что D-ЛДГ из Leuconostoc mesenteroides (Sigma, каталожный номер L2395) показывает более высокую активность в отношении производных индола в качестве субстратов, чем L-ЛДГ из Bacillus stearothermophilus (Sigma каталожный номер L5275). Аналогичные способы применялись в отношении D-лактата и NAD+ или NADH и пирувата, естественных субстратов полипептидов D-ЛДГ. Величина Vмакс для восстановления пирувата оказалась в 1001000 раз выше, чем Vмакс для окисления лактата. Величина Vмакс для реакции окисления индол-3-лактата с помощью D-ЛДГ была примерно в пять раз меньше, чем для лактата. Наличие индол-3-пирувата также измеряли по изменению поглощения при 327 нм (енол-боратные производные) с использованием 50 мМ натрий-боратного буфера, содержащего 0,5 мМ ЭДТА и 0,5 мМ арсената натрия. Обнаружены небольшие, но воспроизводимые изменения поглощения по сравнению с отрицательным контролем и для L, и для D-ЛДГ полипептидов. Кроме того, лактат-дегидрогеназы широкой специфичности (ферменты с активностью, относящейся к классам ЕС 1.1.1.27, ЕС 1.1.1.28, и/или ЕС 1.1.2.3), можно клонировать и использовать для получения индол-3-пирувата из индол-3-лактата. Источниками лактат-дегидрогеназ широкой специфичности являются, в частности, Е. coli, Neisseria gonorrhoeae и Lactobacillus plantarum. Альтернативно, индол-3-пируват можно получить в результате контакта индол-3-лактата с клеточными экстрактами из Clostridium sporogenes, которые содержат индоллактат-дегидрогеназу (ЕС 1.1.1.110); или клеточными экстрактами из Trypanosoma crusi epimastigotes, которые содержат ргидроксифениллактат-дегидрогеназу (ЕС 1.1.1 .222), которая, как известно, обладает активностью в отношении индол-3-пирувата; или клеточными экстрактами из Pseudomonas acidovorans или Е. coli, которые содержат имидазол-5-иллактат-дегидрогеназу (ЕС 1.1.1.111); или из Coleta blumei, которые содержат гидроксифенилпируват-редуктазу (ЕС 1.1.1.237); или Candida maltosa, которые содержат Dароматическую лактат-дегидрогеназу (ЕС 1.1.1.222). Среди источников, описывающих эти виды активности, можно назвать Nowicki с соавт. (FEMS Microbiol Lett 71:119-24,1992), Jean и DeMoss (Canadian J.(Ou et al. I. Mol Catalysis B: Enzymatic: 18:299-305, 2002), можно использовать для окисления индол-3 лактата в индол-3-пируват. Пример 3. Превращение L-триптофана в индол-3-пируват с использованием оксидазы Lаминокислот. В этом примере описываются способы, которые используются для превращения индол-3-пирувата с помощью оксидазы (ЕС 1.4.32), как альтернативы применению триптофан-аминотрансферазы как описано в примере 1. Оксидазу L-аминокислот очищали из Crotalus durissus (Sigma, St Louis, МО, каталожный номер А-2805). Номера последовательностей оксидаз L-аминокислот для молекулярного клонирования включают: CAD21325.1, AAL14831, NP490275, ВАВ 78253, А 38314, САВ 71136, JE0266, Т 08202, S48644,САС 00499, Р 56742, Р 81383, O93364, Р 81382, Р 81375, S62692, Р 23623, AAD45200, ААС 32267, САА 88452,АР 003600 и Z48565. Реакции проводили в микроцентрифужных пробирках общим объемом 1 мл, инкубировали в течение 10 мин, встряхивая при 37 С. Реакционная смесь содержала 5 мМ L-триптофан, 100 мМ натрийфосфатный буфер, рН 6,6, 0,5 мМ арсенат натрия, 0,5 мМ ЭДТА, 25 мМ тетраборат натрия, 0,016 мг каталазы (83 ед., Sigma C-3515), 0,008 мг FAD (Sigma) и 0,005-0,125 единиц оксидазы L-аминокислот. Отрицательные контроли содержали все компоненты, за исключением триптофана, а слепые пробы содержали все компоненты, за исключением оксидазы. Каталазу использовали для удаления перекиси водорода, образующейся в процессе окислительного деаминирования. Тетраборат и арсенат натрия применены для стабилизации енол-боратной форму индол-3-пирувата, которая показывает максимальное поглощение при 327 нм. Стандарты индол-3-пирувата готовили в концентрациях от 0,1 до 1 мМ в реакционной смеси. Коммерческий препарат оксидазы L-аминокислот имеет специфическую активность 540 мкг образовавшегося индол-3-пирувата в минуту на мг белка. Это тот же порядок величин, что обнаруживается и для специфической активности триптофан-аминотрансфераз. Пример 4. Превращение индол-3-пирувата в 2-гидрокси-2-индол-3-илметил)-4-кетоглутарат с помощью альдолазы. В этом примере описываются способы, которые можно использовать для превращения индол-3 пирувата в 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту - предшественник монатина (ПМ) с помощью альдолазы (лиазы) (фиг. 2). Реакции альдольной конденсации - это реакции, которые образуют углерод-углеродные связи между -углеродом альдегида или кетона и карбонильным углеродом дру- 21009284 гого альдегида или кетона. На углероде, находящемся рядом с карбонильной группой одного субстрата,образуется карбанион, который служит в качестве нуклеофила, атакующего карбонильный углерод другого субстрата (электрофильный углерод). Чаще всего электрофильный субстрат является альдегидом,поэтому большинство альдолаз попадает в категорию ЕС 4.1.2.-. Довольно часто нуклеофильный субстрат - это пируват. Реже альдолазы катализируют реакции конденсации между двумя кето-кислотами или двумя альдегидами. Однако идентифицированы альдолазы, которые могут катализировать конденсацию двух карбоксильных кислот. Например, ЕР 1045-029 описывает получение L-4-гидрокси-2-кетоглутарата из глиоксиловой кислоты и пирувата с помощью культуры Pseudomonas (ЕС 4.1.3.16). Кроме того, 4-гидрокси-4 метил-2-оксоглутарат-альдолаза (4-гидрокси-4-метил-2-оксоглутарат-пируват-лиаза, ЕС 4.1.3.17) может катализировать конденсацию двух кетокислот. Поэтому сходные полипептиды альдолаз используются для катализа конденсации индол-3-пирувата с пируватом. 4-гидрокси-4-метил-2-оксоглутарат-пируват-лиазы (ProA альдолазы, ЕС 4.1.3.17) и 4-гидрокси-2 оксоглутарат-глиоксилат-лиаза (KHG-альдолаза, ЕС 4.1.3.16) катализируют реакции очень похожие на альдолазную реакцию, представленную на фиг. 2. Сконструированы праймеры с концами, совместимыми с вектором рЕТ 30 Ха/LIC (Novagen, Мэдисон, Висконсин). Конструкция этих праймеров описана выше в примере 1. Для клонирования в рЕТ 30 Xa/LIC сконструированы следующие праймеры: 1. Pseudomonas straminea ген proA (Genbank Accession No.: 12964663 Версия: 12964663) и Comamonas testosteroni ген proA (SEQ ID NOS: 65-66, нуклеотидная и аминокислотная последовательности,соответственно) прямой 5'-GGTATTGAGGGTCGGCATGTACGAACTGGGAGTTGT-3' и обратный 5'AGAGGAGAGTTAGAGCTTAGTCAATATATTTCAGGC-3' (SEQ ID NOS: 55 и 56). 2. Sinorhizobium meliloti 1021 ген SMc00502 (гомологичен гену proA, Genbank AccessionAGCGCGAAG-3' (SEQ ID NOS: 39 и 40). Геномную ДНК из организмов, описанных выше в пунктах 1-2 и 6-8, очищали с помощью протокола Qiagen genomic-tip. Аналогичными способами можно очистить геномную ДНК из организмов, описанных в 3-5.Pseudomonas straminea (ATCC 33636) выращивали на питательном бульоне и среде с гидроксибензоатом при 30 С. Comamonas testosteroni (ATCC 49249) выращивали на питательном бульоне и среде с гидроксибензоатом при 26 С. Sphingomonas sp. LB126 (Flemish Institute for Technological Research, VITO,B-2400 Mol, Бельгия) выращивали согласно способу, описанному Wattiau с соавт. (Research in Microbiol. 152:861-72, 2001). Arthrobacter keyseri (Gulf Ecology Division, National Health and Environmental Effects(ATCC 51124) выращивали при 26 С на среде АТСС TY и среде с гидроксибензоатом. Yersinia pestis штамм СО 92 (АТСС) выращивали при 26 С в среде АТСС medium 739 Hone blood agar. Bacillus subtilis 6051 (АТСС выращивали при 30C в питательном бульоне Bereto (Difco; Детройт, Мичиган). Геномную ДНК Е. coli выделяли из штамма DH10B (Invitrogen), как описано в примере 1. Протоколы для ПЦР, клонирования и скринирования, представленные в примере 1, использовали- 22009284 для клонирования последовательностей генов proA C.testosteroni и the S.meliloti, а также последовательностей khgE. coli, В. subtilis, и S. meliloti. Эти же способы можно использовать для клонирования других последовательностей, описанных выше. Положительные клоны секвенировали с помощью терминационного секвенирования дидеокси-цепи(Seqwright, Хьюстон, Техас) с S-тэгом и Т 7-терминаторными праймерами (Novagen), и внутренними праймерами, полученными от Integrated DNA Technologies,Inc. (Коралвилль, Индиана). Экспрессия и определение активности Плазмидную ДНК (проверенную аналитическим секвенированием) субклонировали в экспрессионную клетку хозяина BL21(DE3) (Novagen). Культуры выращивали в среде LB с 50 мг/л канамицина. Плазмиды выделяли с помощью набора Qiagen spin plasmid miniprep и впоследствии подвергали рестрикционому анализу для подтверждения идентичности. Проводили эксперименты с индукцией конструктов BL21(DE3), выращенных в среде LB, содержащей 50 мг/л канамицина, при 37 С. Экспрессию белка индуцировали с помощью 0,1 мМ IPTG после того, как величина OD600 достигала величины около 0,6. Клетки росли в течение 4 ч при 30 С, после чего их отделяли центрифугированием. Клетки затем лизировали реагентом Bugbuster (Novagen) и рекомбинатные белки с His-тэгами очищали с помощью HisBind картриджей, как описано выше (пример 1). Очищенные белки обессоливали на колонках PD-10 и элюировали в 50 мМ трис-HCl буфере, рН 7,3 с 2 мМ MgCl2. Белки анализировали способом SDS-PAGE на 4-15% градиентных гелях, чтобы определить уровни растворимых белков по предсказанной величине MW рекомбинантного фузионного белка. Определяли активность белков с помощью индол-3-пирувата и пирувата натрия в качестве субстратов. Смесь для определения содержала 100 мМ трис-HCl (рН 7-рН 8.9), 0-8 мМ MgCl2, 3 мМ фосфат калия (рН 8) и 6 мМ каждого субстрата в 1 мл. Реакцию запускали добавлением разных количеств полипептида (например, от 10 до 100 мкг) и инкубировали при 25-37 С в течение 30 мин, фильтровали и замораживали при -80 С. Результаты измерения активности у продуктов гена pro A Генные конструкты С. testosteroni proA и S. meliloti SMc00502 показывают высокий уровень экспрессии при индукции IPTG. Рекомбинантные белки хорошо растворимы, как показывает анализ общего белка и образцов с клеточными экстрактами способом SDS-PAGE. Продукт гена С. testosteroni был очищен до уровня 95%. Поскольку выход генного продукта S. meliloti был очень низким после аффинной очистки с помощью картриджа His-Bind, для определения ферментной активности использовали клеточный экстракт. Обе рекомбинантные альдолазы катализировали образование ПМ из индол-3-пирувата и пирувата. Для проявления ферментативной активности необходимо присутствие двухвалентного магния и фосфата калия. Продукт не выявлялся, если в смеси не присутствовали индол-3-пируват, пируват или фосфат калия. Небольшое количество продукта образовывалось в отсутствие фермента (как правило, на один порядок меньше, при наличии фермента). Пик продукта элюировали с колонки с обратной фазой С 18 несколько позже, чем стандарт индол-3 пирувата, масс-спектр этого пика показал коллизионно-индуцированный родительский ион ([М+Н]+) при 292,1, родительский ион, предполагаемый для продукта ПМ. Основные дочерние фрагменты, присутствующие на масс-спектре, показали, в частности, m/z =158 (1H-индол-3-карбальдегид карбонийион), 168 (3-бута-1,3-диенил-1H-индол карбоний-ион), 274 (292 - Н 2 О), 256 (292 - 2 Н 2 О), 238 (292 3 Н 2 О), 228 (292 - СН 4 О 3) и 204 (затухание пирувата). Продукт также продемонстрировал УФ-спектр, характерный для других индолсодержащих соединений, например триптофана, с макс 279-280 и небольшим плечом примерно при 290 нм. Количество ПМ, продуцируемое альдолазами C.testosteroni, увеличивалось с повышением температуры реакции от комнатной до 37 С, концентрации субстрата и количества магния. Синтетическая активность фермента уменьшалась с повышением рН, при этом максимальное количество продукта получалось при рН 7. При сопоставлении со стандартами триптофана показано, что количество ПМ, полученного в стандартном определении с 20 мкг очищенного белка, составило примерно 10-40 мкг на 1 мл реакции. Благодаря высокой степени гомологии кодирующих последовательностей ProA альдолаз из S.meliloti и С. testosterone с другими описанными выше генами, предполагается, что все продукты рекомбинантных генов могут катализировать эту реакцию. Более того, ожидается, что альдолазы, имеющие треонин (Т) в положениях 59 и 87, аргинин (R) в 119, аспартат (D) в 120 и гистидин (Н) в 31 и 71 (на основе нумерации у С. testosteroni), будут иметь сходную активность. Результаты измерения активности у продуктов гена khs Конструкты на основе генов khg, полученных у В. subtilis и Е. coli, покрывают высокий уровень экспрессии при индукции IPTG, тогда как ген S. meliloti продемонстрировал низкий уровень экспрессии. Рекомбинантные белки хорошо растворимы, как показывает анализ общего белка и образцов с клеточными экстрактами способом SDS-PAGE. Продукты khg-генов В. subtilis и Е. coli были очищены до уровня 95%; выход генного продукта S. meliloti был не столь высоким после аффинной очистки с помощью картриджа His-Bind.- 23009284 Не удалось найти доказательства того, что магний и фосфат необходимы для активности этого фермента. Однако в литературе есть сообщения о проведении определения в натрий-фосфатном буфере, и этот фермент, по мнению этих авторов, является бифункциональным и обладает активностью в отношении фосфорилированных субстратов, таких как 2-кето-3-дезокси-6-фосфоглюконат (KDPG). Определение активности ферментов проводили как описано выше, и в некоторых случаях фосфат был исключен. Результаты показывают, что рекомбинантные KHG-альдолазы продуцируют ПМ, но не так активно какProA альдолазы. В некоторых случаях уровень ПМ, синтезируемого с помощью KHG, почти идентичен количеству, синтезируемому при наличии одних магния и фосфата. Фосфат, очевидно, не повышает активность KHG. Фермент из Bacillus по данным SRM (см. пример 10) обладает самой высокой активностью, приблизительно на 20-25% выше активности, наблюдаемой при наличии только магния и фосфата. Фермент из Sinorhizobium обладает самой низкой активностью, что может быть связано с особенностями фолдинга и проблемами с растворимостью, отмеченными в экспериментах по экспрессии. Все три фермента имеют в активном сайте глутамат (положение 43 по нумерации В. subtilis), а также лизин, необходимый для образования шиффова основания с пируватом (позиция 130); однако фермент из В. subtilis содержит треонин в положении 47 - остаток активного сайта, - а не аргинин. KHG из В. subtilis меньше и,очевидно, принадлежит подсемейству, отличному от ферментов из S. meliloti и Е. coli, но содержащему другие ферменты с треонином в активном сайте. Расхождения в активном сайте могут быть причиной повышенной активности фермента из В. subtilis. Повышение активности альдолаз Каталитические антитела могут быть такими же активными, как природные альдолазы, воспринимать широкий диапазон субстратов и могут использоваться для катализа реакций, представленных на фиг. 2. Альдолазы можно также усовершенствовать способами направленной эволюции, например, как описано ранее для KDPG-альдолазы (высокогомологичной KHG, описанной выше), полученной при перестановках в ДНК и неточной ПЦР, чтобы устранить потребность в фосфатах фосфат и обратить энантиоспецифичность. KDPG-альдолазные полипептиды применимы в биохимических реакциях, поскольку они высокоспецифичны в отношении субстрата-донора (в данном случае, пирувата), но относительно неприхотливы по отношению к субстрату-акцептору (т.е. индол-3-пирувату) (KoellerWong, Nature 409:232-9, 2001). KHG-альдолаза катализирует реакцию конденсации пируват с рядом карбокси-кислот. Считается, что KHG альдолазы млекопитающих имеют более широкую субстратную специфичность, чем бактериальные ферменты, в том числе, проявляя более высокую активность в отношении 4-гицрокси 4 метил-2-оксоглутарата и способность воздействовать на оба стереоизомера 4-гидрокси-2-кетоглутарата. Оказалось, что бактериальные ферменты отдают почти 10-кратное предпочтение R-изомеру. В геномных базах данных открыта информация почти о 100 гомологах KHG, и эта активность продемонстрирована уPseudomonas, Paracoccus, Providencia, Sinorhizobium, Morganella, E.coli и в тканях млекопитающих. Эти ферменты можно использовать в начале работы над энантиоспецифичностью, которая необходима при синтезе монатина. Альдолазы, которые утилизуют пируват и другой субстрат, который может быть кетокислотой и/или молекулой с большой гидрофобной группой, как у индола, можно развивать в направлении специфичности, скорости и избирательности. В добавление к KHG- и ProA-альдолазам, показанным здесь,можно привести в качестве примеров еще несколько ферментов, но без ограничений: KDPG-альдолазы и родственные полипептиды (KDPH); транскарбоксибензальпируват гидратаза-альдолаза из Nocardioidesst; 4-(2-карбоксифенил)-2-оксобут-3-еноат-альдолаза(2'-карбоксибензальпируват-альдолаза), которая конденсирует пируват и 2-карбоксибензальдегид (субстрат, содержащий ароматическое кольцо); трансО-гидроксибензилиден-пируват-гидратаза-альдолаза из Pseudomonas putida и Sphingomonas aromaticivorans, которая утилизует также пируват и альдегид, содержащий ароматическое кольцо, в качестве субстратов; 3-гидроксиаспартат-альдолаза (эритро-3-гидрокси-L-аспартат глиоксилат-лиаза), которая использует в качестве субстратов 2-оксокислоты и, по-видимому, экспрессируется в организме Micrococcus denitrificans; бензоин-альдолаза (бензальдегид-лиаза), которая утилизует субстраты, содержащие бензиловые группы; дигидронеоптерин-альдолаза; L-трео-3-фенилсерин-бензальдегид-лиаза (фенилсеринальдолаза), которая конденсирует глицин с бензальдегидом; 4-гидрокси-2-оксовалерат-альдолаза; 1,2 дигидроксибензилпируват альдолаза и 2-гидрокси бензальпируват альдолаза. Полипептид с нужной активностью можно отобрать при скринировании интересующих клонов с помощью следующих способов. Триптофан-ауксотрофы трансформируют векторами, несущими интересующие клоны, на экспрессионной кассете и выращивают на среде, содержащей небольшие количества монатина или ПМ. Поскольку аминотрансферазные и альдолазные реакции обратимы, клетки способны синтезировать из рацемической смеси монатина. Аналогичным образом организмы (как рекомбинантные, так и интактные) можно скринировать на способность утилизовать ПМ или монатин в качестве источника углерода и энергии. Одним из источников нужной альдолазы может стать экспрессионная библиотека разных штаммов Pseudomonas и ризобактерий. У псевдомонад есть много необычных катаболических путей для деградации ароматических молекул, и они к тому же содержат много альдолаз; тогда как ризобактерий содержат альдолазы, растут в растительной ризосфере и имеют много генов, хорошо- 24009284 описанных для конструирования биосинтетического пути монатина. Пример 5. Химический синтез предшественника монатина. В примере 4 описаны способы превращения индол-3-пирувата в 2-гидрокси-2-(индол-3-илметил)-4 кето-глутаровую кислоту - предшественник монатина (ПМ) с помощью альдолазы. А в этом примере описывается альтернативный способ химического синтеза ПМ. ПМ образуется в результате типичной реакции альдольной конденсации (фиг. 4). Типичная реакция альдольной конденсации происходит через образование карбаниона пируват-эфира с помощью сильного основания, например LDA (диизопропиламид лития), гексаметилдисилазана лития или бутил-лития, образующийся карбанион реагирует с индол-пируватом с образованием конденсированного продукта. Защитные группы, которые можно использовать для защиты атома азота в индольной группе, включают, но не ограничиваются, следующие: t-бутилоксикарбонил (Вос) и бензилоксикарбонил (Cbz). Блокирующие группы для карбоксикислот включают, но не ограничиваются, следующие: алкиловые эфиры(например, метиловые, этиловые, бензиловые эфиры). Если используются такие защитные группы, то становится невозможно контролировать стереохимию образующегося продукта. Однако если R2 и/илиR3 - хиральные защитные группы (фиг. 4), такие как (S)-2-бутанол, ментол или хиральный амин, то можно добиться предпочтительного образования одного энантиомера ПМ перед другим. Пример 6. Превращение триптофана или индол-3-пирувата в монатин. В результате проведения in vitro процесса с использованием двух ферментов, аминотрансферазы и альдолазы, образуется монатин из триптофана и пирувата. На первом этапе альфа-кетоглутарат является акцептором аминогруппы триптофана в реакции трансаминирования с образованием индол-3-пирувата и глутамата. Альдолаза, катализирующая вторую реакцию, в которой пируват реагирует с индол-3 пируватом в присутствии Mg2+ и фосфата с образованием альфа-кето-производного монатина (ПМ), 2 гидрокси-2-(индол-3-илметил)-4-кето-кетоглутарата. Перенос аминогруппы от глутамата, образовавшегося на первом этапе реакции, приводит к образованию конечного продукта монатина. В результате очистки и характеристики этого показано, что полученный изомер является S,S-монатином. Описываются альтернативные субстраты, ферменты и условия, а также усовершенствования, которые можно внести в ход процесса. Ферменты Альдолазу, 4-гидрокси-4-метил-2-оксоглутарат-пируват-лиазу (ProA альдолаза, ген proA) (ЕС 4.1.3.17) из Comamonas testosteroni клонировали, экспрессировали и очищали, как описано в примере 4. 4-Гидрокси-2-окосоглутарат-глиоксилат-лиазы (KHG-альдолазы) (ЕС 4.13.16) из B.subtilis, E.coli, иS.meliloti клонировали, экспрессировали и очищали, как описано в примере 4. Аминотрансферазы, используемые вместе с альдолазами при продукции монатина - это L-аспартатаминотрансфераза, кодируемая геном aspC E.coli, тирозин-аминотрансфераза, кодируемая геном tyrB изE.coli, фермент TatA из S. meliloti, аминотрансфераза широкой субстратной специфичности, кодируемая геном L. major bsat, или глутамат-оксалоацетат-трансаминаза из сердца свиньи (тип IIa). Белки, полученные не у млекопитающих, клонировали, экспрессировали и очищали, как описано в примере 1. А глутамат-оксалоацетат-трансаминазу из сердца свиньи (тип IIа) получали от Sigma ( G7005). Способ с использованием альдолазы proA и L-аспартат-аминотрансфераза Реакционная смесь содержала 50 мМ ацетата аммония, рН 8,0, 4 мМ MgCl2, 3 мМ фосфата калия,0,05 мМ пиридоксаль фосфата, 100 мМ пирувата аммония, 50 мМ триптофана, 10 мМ альфакетоглутарата, 160 мг рекомбинантной ProA-альдолазы из C.testosteroni (неочищенный клеточный экстракт, -30% альдолазы), 233 мг рекомбинантной L-аспартат-аминотрансферазы из Е. coli (неочищенный клеточный экстракт, -40% аминотрансферазы) в одном литре. Все компоненты, кроме ферментов, смешивали вместе и инкубировали при 30 С до полного растворения триптофана. Затем добавляли ферменты и раствор реакционной смеси инкубировали при 30 С, осторожно встряхивая (100 об/мин) в течение 3,5 ч. Через 0,5 и 1 ч после добавления ферментов аликвоты твердого триптофана (по 50 ммоль каждая) добавляли в реакционную смесь. Добавленный триптофан не растворяется полностью, но концентрация его поддерживается на уровне 50 мМ или выше. Через 3,5 ч твердый триптофан отфильтровывают. Анализ реакционной смеси способом ЖХ/МС с определенным количеством триптофана в качестве стандарта показал, что концентрация триптофана в растворе составляет 60,5 мМ, а концентрация монатина -5,81 мМ (1,05 г). Для очистки конечного продукта используют следующие способы. 90% прозрачного раствора наносят на колонку со смолой BioRad AG50W-X8 (225 мл; связывающая способность 1,7 мэкв/мл). Колонку промывают водой, собирают фракции по 300 мл до тех пор, пока поглощение при 280 нм не упадет до уровня 5% от первого прохождения фракции. Затем колонку элюировали 1 М ацетатом аммония, рН 8,4, собирая 4 фракции по 300 мл. Все 4 фракции содержали монатин и были упарены 105 мл с помощью вращающегося упаривателя на водяной бане. Осадок, образующийся по мере уменьшения объема, отделяли фильтрованием в процессе упаривания. Анализ колоночных фракций способами ЛЖ/МС показал, что 99% триптофана и монатина связывается на колонке. Осадок, который формировался в процессе упаривания, содержал 97% триптофана и 2% монатина. Отношение триптофана к продукту в супернатанте составило приблизительно 2:1.- 25009284 Супернатант (7 мл) наносили на 100 мл колонку Fast Flow с DEAE-сефарозой (Amersham Biosciences), предварительно переведенную в ацетатную форму промыванием 0,5 л 1 М NaOH, 0,2 л воды, 1,.0 л 1,0 М ацетата аммония, рН 8,4, и 0,5 л воды. Супернатант загружали со скоростью 2 мл/мин и промывали колонку водой со скоростью 3-4 мл/мин до тех пор, пока поглощение при 280 нм не достигнет значения 0. Монатин элюировали 100 мМ раствором ацетата аммония, рН 8,4, и собирали 4 фракции по 100 мл. Анализ фракций показал, что соотношение триптофана к монатину в протоке через фракции было 85:15, а соотношение в элюируемых фракециях составило 7:93. Считая коэффициент экстинкции при 280 нм равным для монатина и триптофана, фракции элюента содержали 0,146 ммоль продукта. При экстраполировании на общий объем реакционной смеси 1 л будет получено -2,4 ммоль (-710 мг) монатина, с выходом 68%. Фракции элюента, снимаемые с колонки с DEAE-сефарозой, упаривали до объема 20 мл. Затем аликвоту продукта очищали путем нанесения на препаративную колонку C8 для проведения обращннофазовой хроматографии в условииях, описанных в примере 10 для аналитического определения монатина. Для запуска автоматизированного сбора фракций монатина использовали программное обеспечениеWaters Fnctionlynx на основе определения ионов m/z - 293. Фракции с колонки C8 с соответствующим протежированным молекулярным ионом монатина собирали, упаривали досуха, а затем растворяли в небольшом объеме воды. Эту фракцию использовали для снятия характеристик продукта. Полученный продукт характеризовали следующими способами. Спектроскопия в УФ/видимом спектрах. Спектроскопические измерения в УФ/видимом спектрах монатина, полученного ферментативным путем, проводили с помощью спектрофотометра для измерений в УФ/видимом спектрах Gary 100 Bio. Очищенный продукт, растворенный в воде, показал максимум поглощения при 280 нм с плечом при 288 нм, характерный для индол-содержащих веществ. ЖХ/МС-анализ. Анализ смеси монатина, полученного в результате биохимической реакции in vitro,проводили как описано в примере 10. Типичный ЖХ/МС-анализ монатина, полученного в результате биохимической реакции in vitro, показан на фиг. 5. В нижней части фиг. 5 показаны результаты селективной ионной хроматографии для протонированного молекулярного иона монатина при m/z = 293. Это определение характеристик монатина в смеси подтверждается масс-спектром, показанным на фиг. 6. ЖХ/МС-анализ очищенного продукта показал единственный пик с молекулярным ионом 293 и поглощением при 280 нм. Масс-спектр идентичен спектру, показанному на фиг. 6. МС/МС-анализ. ЖХ/МС/МС-анализ дочерних ионов монатина выполнен как описано в примере 10. Масс-спектр дочерних ионов показан на фиг. 7. Проведено экспериментальное структурное распределение для всех фрагментов ионов, отмеченных на фиг. 7. Включены фрагмент-ионы с m/z = 275 (293 Н 2 О), 257 (293-(2 х Н 2 О, 230 (275-СООН), 212 (257-СООН), 168 (3-бута-1,3-диенил-1H-индолкарбоний-ион), 158 (1H-индол-3-карбальдегид-карбоний-ион), 144 (3-этил-1H-индол-карбоний-ион), 130(3-метилен-1H-индол-карбоний-ион) и 118 (индол-карбоний-ион). Многие из этих ионов образуются и при анализе ПМ (пример 4), что можно было предполагать, особенно для индольной части молекулы. Некоторые оказались на 1 единицу массы выше, чем полученные для ПМ, благодаря наличию аминогруппы вместо кетона. МС/МС-анализ высокого разрешения. На фиг. 8 показан масс-спектр, полученный для очищенного монатина с помощью масс-спектрометра Applied Biosystems-Perkin Elmer Q-Star. По результатам измерения масса протонированного монатина с использованием триптофана как внутреннего стандарта для калибровки массы составила 293,1144. Расчетная масса протонированного монатина на основе химического состава C14H17N2O5 равняется 293,1137. Ошибка в измерении массы составила меньше двух миллионных долей, что подтверждает химический состав монатина, полученного ферментативным путем. ЯМР-спектроскопия. Эксперименты ЯМР выполнены на аппарате Varian Inova 500 МГц. Образец монатина (3 мг) растворяли в 0,5 мл D2O. Сначала в качестве внутреннего стандарта использовали растворитель (D2O) при 4.78 ppm. Поскольку пик для воды был большим, то провели 1 Н-ЯМР с подавлением пика воды. Впоследствии, из-за ширины пика воды, в качестве пика сравнения использовали С-2 часть монатина, установив на опубликованной величине 7,192 ppm. При 13 С-ЯМР первоначальный прогон нескольких сотен сканов показал, что образец был слишком разбавленным, чтобы получить адекватный 13 С-спектр в заданное время. Поэтому были проведены эксперименты с гетероядерной когерентностью с множественным квантованием (HMQC), которые дали возможность провести корреляцию атомов водрода и связанных с ними атомов углерода, а также получить информацию о химических сдвигах атомов углерода. Сводные данные по 1 Н и HMQC представлены в табл. 2 и 3. При сравнении с опубликованными данными данные ЯМР показывают, что монатин, полученный ферментативным путем, является либо(S,S), (RJ1), либо смесью этих двух изомеров. Хиральный ЖХ/МС-анализ. Чтобы установить, что монатин, полученный in vitro, является одним изомером, а не смесью (R,R) и (S,S) энантиомеров, проведен ЖХ/МС-анализ хиральности с использованием инструментария, описанного в примере 10. ЖХ-разделение по хиральности проведено на колонке для хиральной хроматографии Chiiobiotic T(Advanced Separations Technology) при комнатной температуре. Способы разделения и детекции на основе опубликованных протоколов от поставщиков были оптимизированы для R- (D) и S- (L) изомеров триптофана. Подвижная фаза для ЖХ состояла из А) воды, содержащей 0,05% (об/об) трифторуксусной кислоты; В) метанола, содержащего 0,05% (об/об) трифторуксусной кислоты. Элюция была изократической при 70% А и 30% В. Скорость составила 1,0 мл/мин, и поглощение PDA измеряли в диапазоне от 200 нм до 400 нм. Инструментальные параметры, использованные для ЖХ/МС-анализа хиральности триптофана и монатина, идентичны описанным в примере 10 для ЖХ/МС-анализа. Использовали коллекцию массспектров для участка m/z 150-400. Результаты селективной ионной хроматографии для протонированных молекулярных ионов ([М + Н]+ =205 для R-и S-триптофанов и [М + Н]+ = 293 для монатина) позволили напрямую идентифицировать эти вещества в смесях. Хроматограммы R- и S-триптофана и монатина, разделенные способом хиральной хроматографии и сопоставленные с МС, показаны на фиг. 9. Единственный пик на хроматограмме монатина показывает,что это вещество представлено одним изомером, а по времени задержки он почти идентичен Sтриптофану. Таблица 2. Данные 1 ЯМР Поляриметрия. Оптическое вращение измеряли на поляриметре Rudolph Autopol EQ. Препарат монатина готовили в виде 14,6 мг/мл водного раствора. Ожидаемое удельное вращение ([]D20) для S,S монатина (солевая форма) составляет -49,6 для 1 г/мл водного раствора (Vieggaar с соавт. Обнаруженная величина []D20 равняется -28.1 для очищенного монатина, полученного ферментативным путем, показы- 27009284 вает, что он является S,S-изомером. Усовершенствования Условия реакции, включая концентрации реагента и фермента, были оптимизированы и дали на выходе 10 мг/мл при использовании реакционной смеси следующего состава: 50 мМ ацетата аммония(рН 8) 3,2 мМ MgCl2, 200 мМ пирувата (в виде солей натрия или аммония), 5 мМ альфа-кетоглутарата(натрия), 0,05 мМ пиридоксаль-фосфата, деаэрированная вода до конечного объема 1 мл после добавления ферментов, 3 мМ фосфат калия, 50 мкг/мл рекомбинантной ProA альдолазы (клеточный экстракт; общая концентрация белка 167 мкг/мл X 1000 мкг/мл L-аспартат-аминотрансферазы, кодируемой геномaspC из E.coli (клеточный экстракт; общая концентрация белка 2500 мг/мл), и твердый триптофан для поддержания концентрации на уровне клеточный экстракт; общая концентрация белка 60 мМ (насыщенный; частично нераствореннный в процессе реакции). Смесь инкубировали при 30 С в течение 4 ч с осторожным встряхиванием или перемешиванием. Замены Концентрацию альфа-кетоглутарата можно снизить до 1 мМ и дополнить 9 мМ аспартатом с эквивалентным выходом монатина. На первом этапе можно использовать альтернативные акцепторы аминокислот, например оксалоацетат. Если вместо L-аспартат-аминотрансферазы из Е. coli используется рекомбинантная аминотрансфераза широкой субстратной специфичности из L. major, достигается аналогичный выход монатина. Однако способом ЖХ/МС-анализа выявляется второй неидентифицированный продукт (3-10% от основного продукта) с молекулярной массой 292. Монатин в концентрации 0,1-0,5 мг/мл получается, если в качестве аминотрансферазы добавляется фермент, кодируемый геном Е. coli tyrB, или фермент, кодируемый геном S. meliloti tat А, или глутамат-оксалоацетат-трансаминаза из сердца свиньи (type IIa). Если реакция начинается с индол-3-пирувата, то на последнем этапе можно провести восстановительное аминирование с применнеием глутамат-дегидрогеназы и NADH (как в примере 7). Кроме того, для получения монатина ферментативным путем используются KHG-альдолазы из В.subtilis, E. coli, и S. meliloti вместе с L-аспартат-аминотрансферазы из Е. coli. Используются следующие условия реакции: 50 мМ NH4-ОАц (рН 8,3), 2 мМ MgCl2, 200 мМ пируват, 5 мМ глутамат, 0,05 мМ пиридоксаль-фосфат, деаэрированная вода до конечного объема 0,5 мл после добавления ферментов, 3 мМ фосфат калия, 20 мкг/мл рекомбинантной В. subtilis KHG-альдолазы (очищенной), против 400 мкг/мл Lаспартат-аминотрансферазы (AspC) из неочищенного клеточного экстракта Е. coli, и 12 мМ индол-3 пируват. Реакционную смесь инкубировали при 30 С в течение 30 мин со встряхиванием. Количество монатина, полученного с помощью фермента из В. subtilis, составило 80 нг/мл и повышалось с увеличением количества альдолаз. Если индол-3-пируват и глутамат замещали насыщенным триптофаном и 5 мМ альфа-кетоглутаратом, выход монатина возрастал до 360 нг/мл. Реакции повторяли с 30 мкг/мл каждого из трех мышиных KHG-ферментов в 50 мМ трис (рН 8,3) с насыщенным триптофаном и оставляли для завершения на час, чтобы увеличить определение. Фермент из Bacillus обладает самой высокой активностью, как указывалось в примере 4, и продуцирует приблизительно 4000 нг/мл монатина. KHG из Е. coli продуцирует 3000 нг/мл монатина, а фермент из S. meliloti -2300 нг/мл. Пример 7. Взаимопревращение между ПМ и монатином. Аминирование ПМ с образованием монатина может катализироваться аминотрансферазами, например, указанными в примерах 1 и 6, или дегидрогеназами, которые требуют восстанавливающего кофактора: NADH или NADPH. Эти реакции обратимы и могут идти в обоих направлениях. Направленность при применении дегидрогеназы можно контролировать концентрациями солей аммония. Активность дегидрогеназы. Окислительное деаминирование монатина отслеживается с помощью последующего увеличения поглощения при 340 нм, так как NAD(P)+ превращается в более хромофорныйNAD(P)H. Монатин получают ферментативным путем и очищают как как описано в примере 6. Типичная смесь для определения содержит 50 мМ трис-HCl, рН от 8.0 до 8,9, 0,33 мМ NAD+ илиNADP+. От 2 до 22 единиц глутамат-дегидрогеназы (Sigma), и 10-15 мМ субстрата в 0,2 мл. Определение проводится дважды в УФ-проницаемой микротитровочной плашке на детекторе для плашек MolecularDevices SpectraMax Phis. Смесь фермента, буфера, и NAD(P)+ наливается пипеткой в ячейки плашки, содержащей субстрат, и увеличение поглощения при 340 нм фиксируется с 10-секундными интервалами после краткого перемешивания. Реакицонную смесь инкубируют при 25 С в течение 10 мин. Отрицательные контроли проводят без добавления субстрата, а в качестве положительного контроля используют глутамат. Глутамат-дегидрогеназа (тип III) из печени быка (SigmaG-7882) катализирует превращение монатина в предшественник монатина со скоростью, составляющей приблизительно одну сотую от скорости превращения глутамата в альфа-кетоглутарат. Активность трансаминирования. Определение аминотрансферазы проводили с аспартатаминотрансферазой (AspC) из Е. coli, тирозин-аминотрансферазой (TyrB) из Е. coli, аминотранферазой широкой специфичности (BSAT) из L. major и двумя коммерческими препаратами глутаматоксалоацетат-аминотрансферазы свиньи, описанными в примере 1. И оксалоацетат, и альфа-кетоглутарат тетисровали в качестве амино-акцепторов. Смесь для определения содержала (в 0,5 мл) 50 мМ трис-HCl- 28009284 определения инкубировали при 30 С в течение 30 мин и останавливали реакции добавлением 0,5 мл изопропилового спирта. Потери монатина отслеживали способом ЖХ/МС (пример 10). Самая высокая величина активности отмечена у фермента из BSAT из L. major с оксалоацетатом в качестве аминоакцептора, затем следует тот же фермент с альфа-кетоглутаратом в качестве амино-акцептора. Относительная активность с оксалоацетатом оказалась следующей: BSATAspCсвиная, тип IIaсвиная, типI = TyrB. Относительная активность с альфа-кетоглутаратом оказалась следующей: BSATAspCсвиная, тип Iсвиная, тип IIaTyrB. Пример 8. Получение монатина из триптофана и С 3-молекул, не являющихся пируватом. Как описано выше в примере 6, индол-3-пируват или триптофан можно превратить в монатин, используя пируват в качестве С 3-молекулы. Однако в некоторых случаях пируват может быть нежелательным исходным материалом. Например, пируват может оказаться более дорогим, чем другие С 3 источники углерода, или могут при добавлении в среду оказать побочные эффекты на ферментационные процессы. Многие ПФ-ферменты могут трансаминировать аланин с образованием пирувата. Триптофаназо-подобные ферменты проводят реакцию бета-элиминации намного быстрее, чем другие ПФ-ферменты, например, аминотрансферазы. Ферменты этого класса (4.1.99.-) могут производить аммоний и пируват из таких аминокислот, как L-серин, L-цистеин, и производных серина и цистеина с легко отщепляемыми группами, например, О-метил-L-серин, О-бензил-L-серин, S-метилцистеин, Sбензилцистеин, S-алкил-L-цистеин, О-ацил-L-серин, 3-хлор-L-аланин. Процессы получения монатина с помощью полипептидов класса ЕС 4.1.99.-можно усовершенствовать с помощью мутационных изменений -тирозиназы (TPL) или триптофаназы по способу Mouratou с соавт. (J. Biol Chem 274:1320-5,1999). Mouratou с соавт. описывают возможность превратить тирозиназы в -лиазу дикарбоновых аминокислот, которой нет в природе, судя по имеющимся ссообщениям. Изменение специфичности проводится путем превращения валина (V) 283 в аргинин (R) и аргинина (R) 100 в треонин (Т). Эти аминокислотные замены позволяют лиазе соединяться с дикарбоновыми аминокислотами для проведения реакции гидролитического деаминирования (например, аспартата). Аспартат, следовательно, можно также исолпьзовать и в качестве источника пирувата для последующей реакции альдольной конденсации. Кроме того, в клеточные или ферментативные реакторы можно добавлять лактат и фермент, который превращает лактат в пируват. В качестве примеров таких ферментов можно назвать лактатдегидрогеназу и лактат-оксидазу. Выделение геномной ДНК Триптофаназа и ее полипептиды описаны в разных источниках, например, Mouratou с соавт. (JBC 274:1320-5,1999). Чтобы получить гены, которые кодируют полипептиды триптофаназы, использовали геномную ДНК из Е. coli DH10B в качестве матрицы для ПЦР, как описано в примере 1. Ген тирозин-фенол-лиазы выделяли из C.freundii (ATCC, каталожный номер 8090, обозначение АТСС 13316; NCTC 9750) и выращивали на агаре Nutrient (Difco 0001) и питательном бульоне (Difco 0003) при 37 С до OD = 2,0. Геномную ДНК очищали с помощью набора Qiagen Genomic-tip 100/G. Амплификация кодирующих последовательностей с помощью ПЦР Праймеры конструировали с концами, совместимыми с вектором pET 30 Xa/LIC (Novagen, Мэдисон, WI) как описано выше в примере 1. Ген tna из Е. coli (SEQ ID : 41). N-терминальный праймер для клонирования рЕТ 30 Xa/LJC cloning: S'-GGT ATT GAG GGT CGC ATG GAA AAC TTT AAA CAT CT-3' (SEQ ID NO: 43). Стерминальный праймер для клонирования рЕТ 30 Xa/LIC: 5'-AGA GGA GAG TTA GAG CCT TAA ACTTCT TTA AGT TTT G-3' (SEQ ID NO:44). Ген tpl из C.freundii (SEQ ID NO:42). N-терминальный праймер для клонирования рЕТ 30 Xa/LIC: 5'GGT ATT GAG GGT CGC ATGAATTATCCGGCAGAACC-3 ' (SEQ ID NO: 45). С-терминальный праймер для клонирования рЕТ 30 Xa/LIC: 5'-AGA GGA GAG TTA GAG CCTTAGATGTAATCAAAGCGTG3' (SEQ ID NO: 46). Для проведения всех ПЦР использовали термоциклер Eppendorf Mastercycler Gradient 5331. К 50uL мкл добавляли 0,5 мкг матрицы (геномной ДНК), 1,0 мкМ каждого праймера, 0,4 мМ каждого dNTP,3,5 ед. полимеразы Expand High Fidelity Polymerase (Roche), 1 X Expand буфер с Mg и 5% ДМСО (конечная концентрация). Программу термоциклера для ПЦР устанавливали со следующими параметрами: горячий старт 96 С (5 мин), 94 С - 30 с, 40-60 С -1 мин 45 с, 72 С - 2 мин 15 с; 30 повторов. Конечный этап полимеризации продолжался 7 мин, и образцы хранили при 4 С. Клонирование Клонирование и процедуры идентификации положительных клонов, описанные выше в примере 1,использовали для идентификации соответствующих клонов. Экспрессия генов и способы определения активности. Плазмидную ДНК (проверенную секвенированием) субклонировали в экспрессирующих клеткахBL21(DE3) (Novagen). Культуры выращивали в среде LB с 30 мг/л канамицином, и выделяли плазмиды с помощью набора Qiagen miniprep, а затем проводили рестрикционный анализ для подтверждения иден- 29009284 тичности. Индукцию проводили в экспрессирующих клетках BL21(DE3), конструкты выращивали на средеLB с канамицином (50 мг/л) при 37 С. Экспрессию белка индуцировали с помощью 1 мМ IPTG (изопропилфлуогалактозид) после того, как величина OD600 культуры достигала приблизительно 0,6. Клетки росли в течение 4 ч при 30 С, после чего их отделяли центрифугированием. Клетки затем лизировали реагентом Bugbuster (Novagen) (5 мл/г сырого веса клетки), содержащем 5 мкл/мл коктейля с ингибитором протеазы III (Calbiochem) и 1 мкл/мл бензоназы-нуклеазы (Novagen),и рекомбинатные белки с His-тэгами очищали с помощью His-Bind картриджей, как описано выше в примере 1. Очищенные белки обессоливали на колонке PD-10 (G25 Sephadex, Amersham Biosciences) и элюировали 100 мМ трис-HCl буфером, рН 8,0. Белки анализировали способом SDS-PAGE на 4-15% градиентных гелях, чтобы определить уровни растворимых белков по предсказанной величине MW рекомбинантного фузионного белка. Мутагенез Некоторые ферменты, входящие в класс 4.1.99.- (триптофаназа и бета-тирозиназа), катализируют бета-лиазную реакцию с аспартатом или подобными ааминокислотами без какой-либо модификации. Однако некоторым ферментам этого класса могут потребоваться мутационные изменения, чтобы сделать возможным применение субстратов и/или для получения нужного продукта. Кроме того, в некоторых случаях полипептиды, способные производить это превращение, могут быть в дальнейшем оптимизированы с помощью мутагенеза. Сайт-направленный мутагенез проводили на основе анализа трехмерной структуры ПФ(PLP)связывающих полипептидов. Ниже приведены два примера изменения субстратной специфичности полипептидов. Мутагенез триптофаназы - Пример 1. Представленный ниже протокол проведения мутагенеза описывает введение двух точковых мутаций в аминокислотную последовательность. Первая точковая мутация приводит к замене аргинина (R) в положении 103 на треонин (Т), а вторая точковая мутация - к замене валина (V) в положении 299 на аргинин (R) (система отсчета - по последовательности зрелого белка из Е. coli. Эксперименты по мутагенезу проведены Лабораториями ATG (г. Иден-Прери, штат Миннесота). Мутации водили последовательно способом ПЦР на фрагментах гена с последующей сборкой гена, также выполненной способом ПЦР. Праймеры для превращения аргинина (R)103 на треонин (Т): 5'-CCAGGGCACCGGCGCAGA: 48). Праймеры для превращения валина (V)299 на аргинин (R): 5'-TCCTGCACGCGGCAAAID : 50). Полученные мутантные последовательности скринировали способом рестрикционного анализа с помощью Xba I/HindIII и SphI, а затем проверяли секвенированием. Мутагенез тирозин-фенол-лиазы (-тирозиназы) - Пример 2. Произведены две точковые мутации в аминокислотной последовательности тирозин-фенол-лиазы. Эти мутации превращали аргинин (R) в положении 100 в треонин (Т) и валин (V) в положении 283 в аргинин (R) (по последовательности зрелого белка из C.freundii). Праймеры для превращения R100T: 5'-AGGGGACCGGCGCAGAAAACCTGTTATCG-31 (SEQ IDNO: 51) и 5'-AGGGGACCGGCGCAGAAAACCTGTTATCG-3' (SEQ ID NO: 52). Праймеры для превращения V283R: 5'-GTTAGTCCGCGTCTACGAAGGGATGCCAT-3' (SEQ ID NO: 53) и 5'GTAGACGCGGAC TAACTCTTTGGCAGAAG-3' (SEQ ID NO: 54). Применялись способы, описанные выше, и полученные клоны скринировали с помощью рестриктазKpnI/SacI и BstX I. Последовательности верифицировали способом терминального секвенирования двойной цепи. Рекомбинатный белок получали как описано выше для ферментов дикого типа. Реакционная смесь содержала 50 мМ трис-Cl (рН 8,3), 2 мМ MgCl2, 200 мМ С 3-молекулы, 5 мМ альфа-кетоглутарата натрия, 0,05 мМ пиридоксаль-фосфат, деаэрированную воду до конечного объема 0,5 мл после добавления ферментов, 3 мМ фосфат калия (рН 7,5), 25 мкг неочищенное рекомбинантнойProA-альдолазы из C.testosteroni, приготовленной как описано в примере 4, 500 мкг неочищенной Lаспартат-аминотрансферазы (AspC) как описано в примере 1, и твердый триптофан для поддержания концентрации на уровне 60 мМ (насыщенный; не полностью растворяется во время реакции). Реакционную смесь инкубировали при 30 С в течение 30 мин при перемешивании. Серин, аланин, и аспартат служили источниками 3-углеродной цепи. Определение проводили с вторичными ПФферментами (очищенными), способными выполнять бета-элиминацию и бета-лиазные реакции (триптофаназа (TNA), триптофаназа с двойной мутацией, бета-тирозиназа (TPL и без этих ферментов. Результаты представлены в табл. 4.
МПК / Метки
МПК: C12P 17/10, C07D 209/18
Метки: полипептиды, биосинтеза, пути
Код ссылки
<a href="https://eas.patents.su/30-9284-polipeptidy-i-puti-biosinteza.html" rel="bookmark" title="База патентов Евразийского Союза">Полипептиды и пути биосинтеза</a>
Способ биосинтеза кобаламинов, штаммы бактерий, продуцирующие кобаламины, и способ их получения

Номер патента: 2588
Опубликовано: 27.06.2002
Авторы: Крузе Жоэль, Реми Элизабет, Дебюсш Лоран, Камерон Беатрис, Бланш Франсис, Тибо Дени
МПК: C12P 19/42, C12N 15/52
Метки: продуцирующие, получения, способ, биосинтеза, кобаламинов, штаммы, кобаламины, бактерий
Формула / Реферат:
1. Способ биосинтеза кобаламинов путем ферментации прокариотического микроорганизма, продуцирующего кобаламины, отличающийся тем, что в качестве указанного микроорганизма используют рекомбинантный штамм микроорганизма, трансформированный фрагментом ДНК, содержащим гены bluЕ и bluF Rhodobacter capsulatus, кодирующие фермент, вовлеченный в биосинтез O-фосфо-L-треонина, или гомологичным фрагментом и/или фрагментом, который гибридизуется с...
Способ непрерывной укладки рельса на жесткое полотно пути, а также жесткое полотно пути

Номер патента: 5054
Опубликовано: 28.10.2004
Авторы: Райхель Дитер, Шрайнер Ульрике, Линднер Эрих
МПК: E01B 1/00
Метки: также, пути, непрерывной, рельса, укладки, жесткое, полотно, способ
Формула / Реферат:
1. Способ непрерывной укладки рельса (4, 4') на жесткое полотно рельсового пути из бетонных плит, в частности состоящее из готовых бетонных элементов, при котором рельс (4, 4') размещают в желобе (3, 3') бетонной плиты (1) и затем фиксируют путем заливки в желобе (3, 3'), отличающийся тем, что по бокам вдоль рельса (4, 4') располагают эластичные уплотнительные блоки (30) внутри желоба (3, 3'), при этом рельсы ориентируют через эластичные...
Стабилизированные водные композиции, содержащие ингибитор пути метаболизма, тканевого фактора (tfpi) или вариант ингибитора пути метаболизма тканевого фактора

Номер патента: 8038
Опубликовано: 27.02.2007
Автор: Чэнь Бао-Лу
МПК: A61K 47/18, A61K 47/12, A61K 38/57...
Метки: пути, ингибитор, композиции, водные, тканевого, вариант, содержащие, tfpi, метаболизма, стабилизированные, ингибитора, фактора
Формула / Реферат:
1. Водная композиция содержащая от приблизительно 0,05 до приблизительно 15 мг/мл TFPI или варианта TFPI; от приблизительно 50 до приблизительно 600 мМ солюбилизирующего агента, выбранного из группы, состоящей из (i) аргинина или его аналога, (ii) лизина или его аналога и (iii) смесей (i) и (ii); и антиоксидант, выбранный из группы, состоящей из (i) газа, замещающего кислород, (ii) поглотителя кислорода или свободных радикалов, (iii)...
Полипептиды lag-3v1, lag-3v2 и lag-3v3 и кодирующие их кднк

Номер патента: 3086
Опубликовано: 26.12.2002
Авторы: Триебель Фредерик, Романьяни Серджо, Мастранджели Ренато
МПК: C07K 14/725, C12N 15/12, A61K 38/17...
Метки: lag-3v3, кднк, lag-3v1, кодирующие, полипептиды, lag-3v2
Формула / Реферат:
1. кДНК SEQ ID NO:1, приведенная в перечне последовательностей, кодирующая полипептид LAG-3V1 с аминокислотной последовательностью SEQ ID NO:2, приведенной в указанном перечне, а также варианты данной кДНК, полученные на основе вырожденности генетического кода. 2. Последовательность нуклеотидов, полностью комплементарная кДНК по п.1. 3. Полипептид LAG-3V1 с аминокислотной последовательностью SEQ ID NO:2, кодируемый кДНК по п.1. 4. кДНК SEQ ID...
Полипептиды tacis и br3 и их применение

Номер патента: 7984
Опубликовано: 27.02.2007
Авторы: Диксит Вишва, Риджуэй Джон, Грювэл Икбэл, Ян Минхонг
МПК: A61K 38/17, C07K 14/705, A61P 37/00...
Метки: полипептиды, применение, tacis
Формула / Реферат:
1. Выделенная нуклеиновая кислота, представляющая собой (а) ДНК, кодирующую полипептид TACIs, содержащий последовательность аминокислотных остатков 1-246 SEQ ID NO:14, или (б) комплемент молекулы ДНК по п.а). 2. Нуклеиновая кислота по п.1, содержащая кодирующую нуклеотидную последовательность SEQ ID NO:13. 3. Нуклеиновая кислота по п.1, состоящая из кодирующей нуклеотидной последовательности SEQ ID N0:13. 4. Выделенная нуклеиновая кислота,...
Предыдущий патент: Предупреждение и лечение амилоидогенного заболевания
Следующий патент: Композиция конъюгированного лекарственного средства
Случайный патент: Фармацевтическая композиция, стимулирующая биосинтез s-аденозилметионина, и пероральное лекарственное средство