Применение антитела, специфичного к рецептору fgf-r4
Номер патента: 21580
Опубликовано: 30.07.2015
Авторы: Реми Элизабет, Борен Николя, Бланш Франсис, Миколь Венсан, Камерон Беатрис, Дабдуби Тарик, Берн Пьер-Франсуа, Эрбер Корантен, Боно Франсуаз



























Формула / Реферат
1. Применение антитела, специфичного к рецептору FGF-R4, для лечения заболеваний, связанных с патологическим ангиогенезом.
2. Применение антитела, специфичного к рецептору FGF-R4, для ингибирования роста опухоли одновременно с ингибированием ангиогенеза.
3. Применение по любому из пп.1 и 2 для лечения гепатокарцином или любых других видов рака печени.
4. Применение по любому из пп.1 и 2 для лечения рака поджелудочной железы.
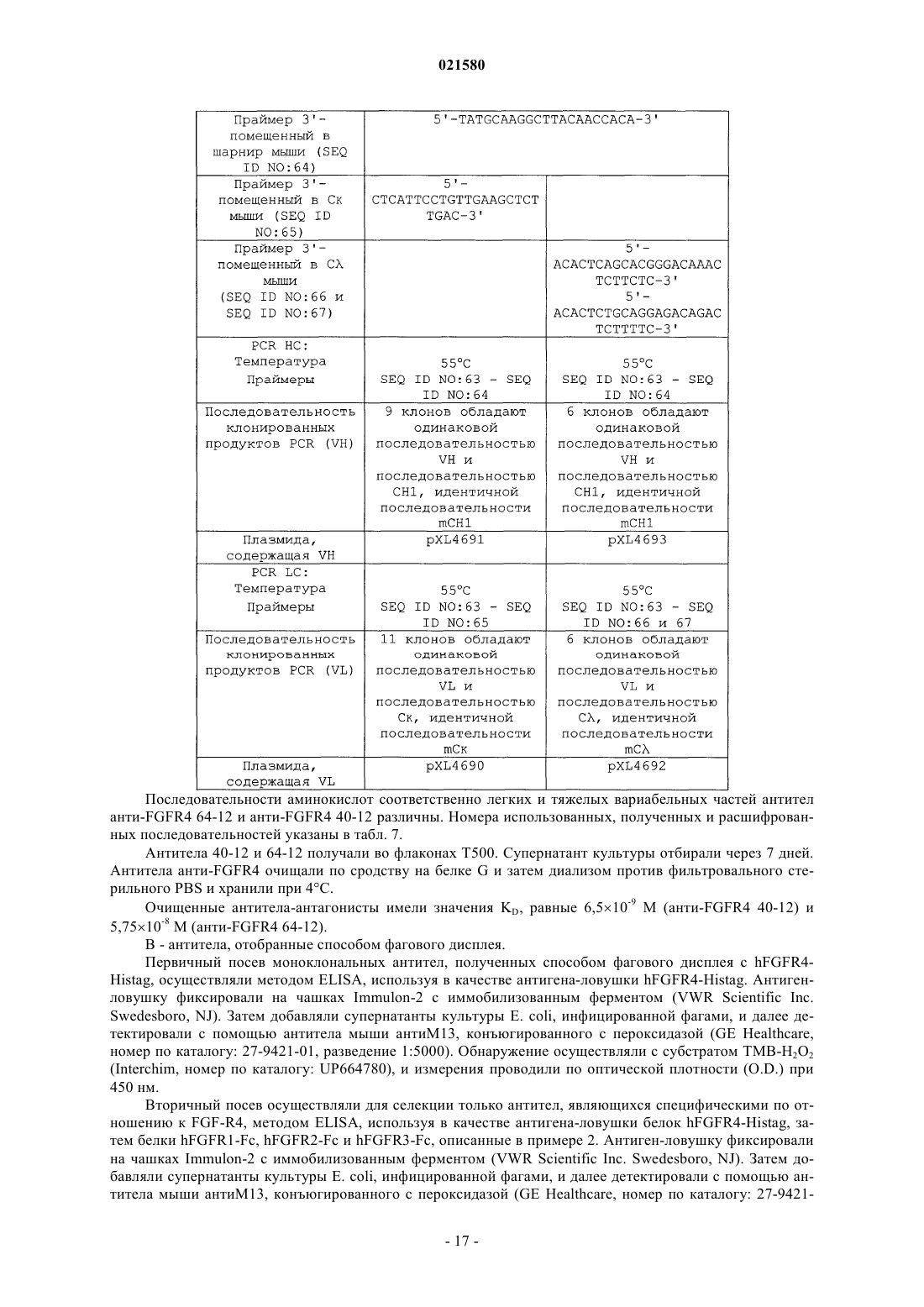
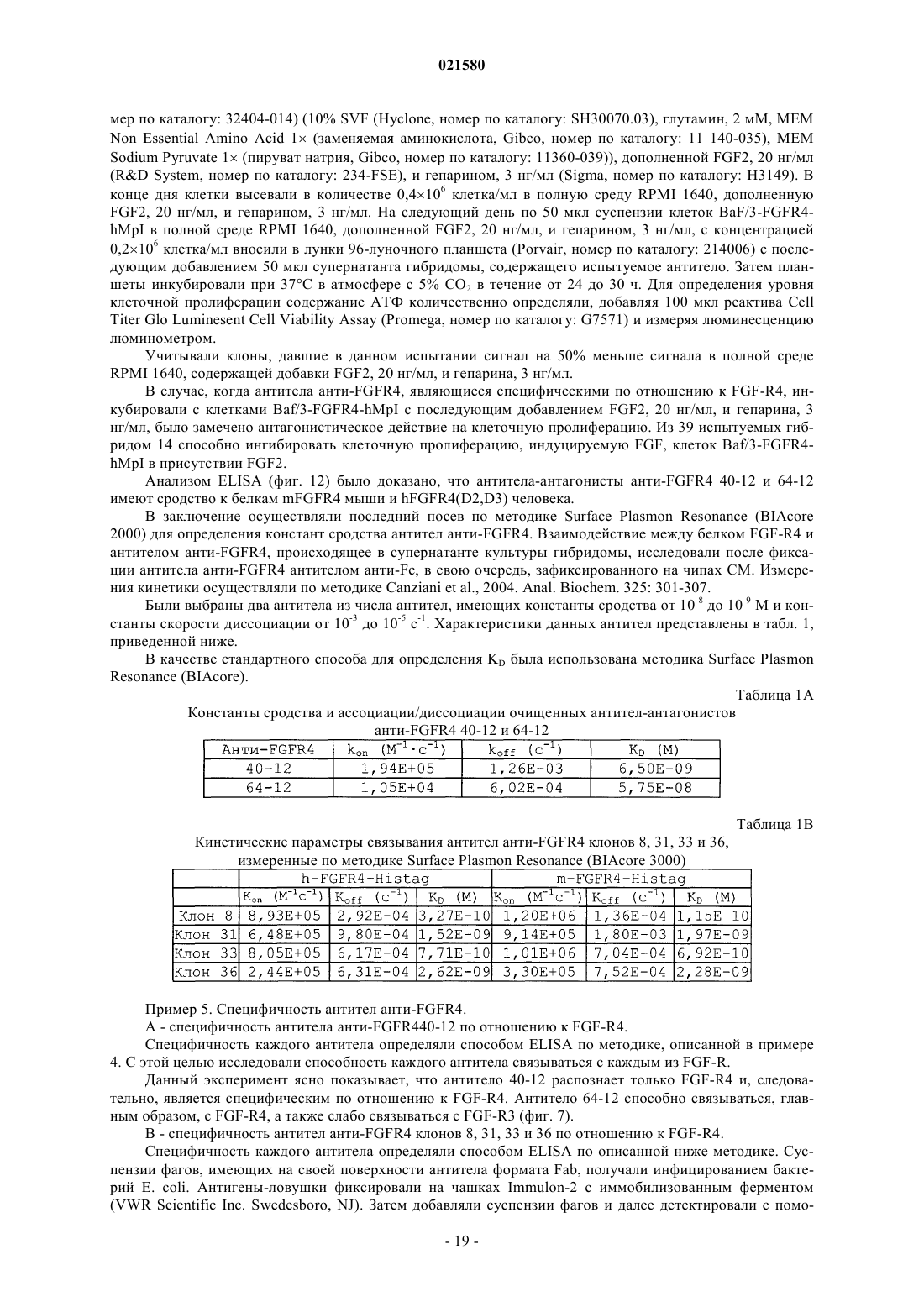
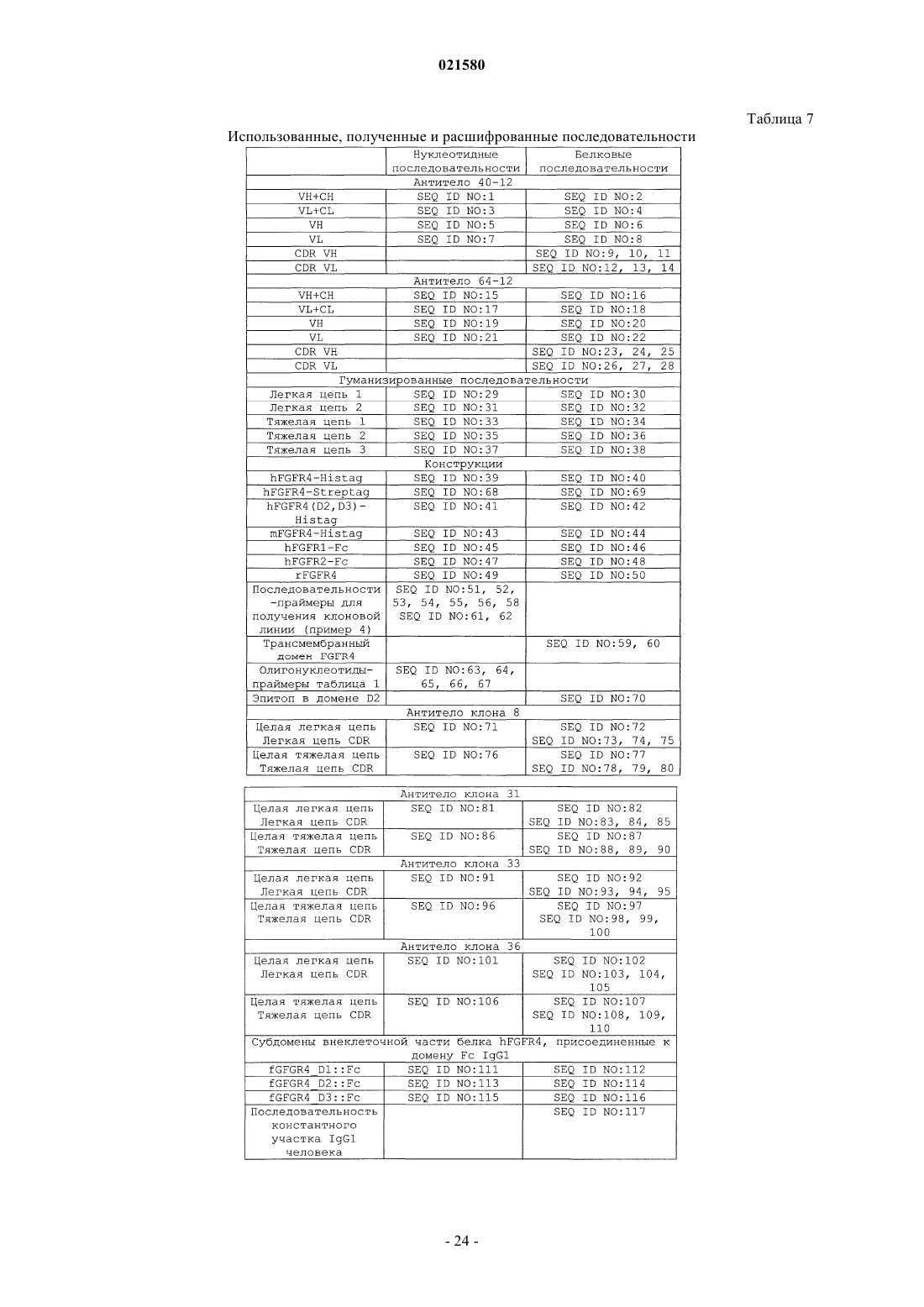
5. Применение по любому из пп.1-4, в котором антитело, специфичное к рецептору FGF-R4, содержит CDR последовательности SEQ ID NO: 9, 10, 11, 12, 13 и 14; или 73, 74, 75, 78, 79 и 80; или 83, 84, 85, 88, 89 и 90; или 93, 94, 95, 98, 99 и 100; или 103, 104, 105, 108, 109 и 110, где по меньшей мере один из CDR может отличаться одной или двумя аминокислотами по сравнению по меньшей мере с одной из указанных выше последовательностей, при том, что антитело сохраняет свою специфичность связывания.
6. Применение по п.5, в котором вариабельная часть тяжелой цепи специфичного антитела содержит нуклеотидную последовательность, идентичную по меньшей мере на 80% последовательности SEQ ID NO: 5, 76, 86, 96 или 106, и в котором вариабельная часть его легкой цепи содержит нуклеотидную последовательность, идентичную по меньшей мере на 80% последовательности SEQ ID NO: 7, 71, 81, 91 или 101, при том, что антитело сохраняет свою специфичность связывания.
7. Применение по п.5, в котором последовательность антитела, специфичного к рецептору FGF-R4, содержит полипептидные последовательности SEQ ID NO: 2 и 4, или 6 и 8, или 72 и 77, или 82 и 87, или 92 и 97, или 102 и 107, или последовательности, соответствующие таким последовательностям и идентичные им по меньшей мере на 80%, при том, что антитело сохраняет свою специфичность связывания.
8. Применение по любому из пп.1-7, отличающееся тем, что указанное антитело представляет собой гуманизированное антитело.
9. Применение по п.8, отличающееся тем, что указанное антитело содержит легкую цепь, вариабельная область которой идентична по меньшей мере на 80% одной из полипептидных последовательностей SEQ ID NO: 30 или 32, и тяжелую цепь, вариабельная область которой идентична по меньшей мере на 80% последовательности SEQ ID NO: 34, 36 или 38.
10. Применение по любому из пп.1-9, отличающееся тем, что антитело конъюгировано с цитотоксическим агентом.
11. Применение по любому из пп.1-10, в котором указанное антитело кодируется полинуклеотидом, по меньшей мере на 80% идентичным одной из последовательностей SEQ ID NO: 1, 3, 5, 7, 29, 31, 33, 35 или 37, 71, 76, 81, 86, 91, 96, 101 или 106, при том, что кодируемый им полипептид сохраняет свою специфичность связывания с рецептором FGF-R4.
Текст
Изобретение относится к применению антитела, специфичного к рецептору FGF-R4, для лечения заболеваний, связанных с патологическим ангиогенезом, а также для ингибирования роста опухоли одновременно с ингибированием ангиогенеза. Настоящее изобретение относится к специфическим антагонистам рецептора 4 FGF (FGF-R4), позволяющим ингибировать активность этого рецептора. Такие антагонисты представляют собой, в частности, антитела, специфически направленные к рецептору 4 FGF (FGF-R4). Настоящее изобретение относится также к терапевтическому применению таких антагонистов предпочтительно в области ангиогенеза и в лечении некоторых видов рака.FGF (факторы роста фибробластов) считаются первыми соединениями, описанными как соединения, способные стимулировать пролиферацию, миграцию и дифференцировку клеток сосудов in vitro и invivo. Во многих научных статьях описана индукция ангиогенеза и образования кровеносных капилляровin vitro и in vivo, обуславливаемая FGF. FGF также участвуют в ангиогенезе опухолей, способствуя образованию кровеносных сосудов, питающих опухоли. Семейство FGF человека включает в себя по меньшей мере 23 члена, имеющих консервативный центральный домен из 120 аминокислот. Они проявляют свою биологическую активность, взаимодействуя со своими рецепторами, обладающими высоким сродством типа тирозинкиназы (FGF-R) и гепарансульфатов протеогликанов, представляющих собой соединения, содержащиеся на поверхности большей части клеток и внеклеточных матриц (сайты связывания с низким сродством), с образованием тройного комплекса. Некоторые FGF обладают высоким сродством к нескольким FGF-R, тогда как другие факторы специфически активируют один рецептор или одну изоформу рецептора. Специфический лиганд FGF-R4 был идентифицирован Xie et al. (Cytokine, 1999, 11:729-35). Этот лиганд, называемый FGF19, представляет собой лиганд, который обладает исключительно высоким сродством к FGF-R4 и связь которого с рецептором зависит от гепарина или гепарансульфатов. FGF19 был идентифицирован у взрослого животного только на уровне гепатоцитов и тонкого кишечника, где он регулирует синтез желчной кислоты печенью. Он может представлять собой фактор роста, действующий во время эмбрионального развития, и может участвовать в развитии зародышевого головного мозга рыб,зебры и человека. Другие лиганды FGF-R4 описаны как FGF1 или FGF2. Эти лиганды сильно активируют FGF-R4, но не являются специфическими по отношению к FGF-R4: они с связываются также с другими FGF-R (Ornitz et al., J. Biol. Chem., 1996, 271:15292-7). Активация рецептора FGF-R4 ведет к нескольким типам клеточной сигнализации. Среди таких типов наиболее традиционная форма соответствует осуществлению пути сигнализации по каскаду фосфорилирования в продолжение стимуляции FGF-R4 посредством FGF. Такая индукция ведет к аутофосфорилированию домена тирозинкиназы FGF-R4 и служит инициации пути внутриклеточной сигнализации,зависящей от фосфорилирования других белков сигнализации, таких как AKT, р 44/42, JNK и т.д. Такая сигнализация фосфорилированием варьирует в зависимости от типа клеток и корецепторов или адгезионно связанных молекул, присутствующих на поверхности данных клеток (Cavallaro et al., Nat. Cell Biol. 3/7, 650-657 (2001); Stadler et al., Cell Signal. 18/6, 783-794 (2006); Lin et al., J. Biol. Chem., 2007, 14:2727784 (2007. Другим вариантом сигнализации, представляющимся важным для FGF-R, является вариант, в котором FGF-R4 представляет собой объект интернализации рецептора после активации в сочетании с его лигандом. Данный механизм зависит не от активности тирозинкиназы рецептора, а от короткой последовательности в С-концевом домене FGF-R4 (Klingenberg et al., J. Cell Sci., 113/Pt10:1827-1838(2000. В литературе описаны четыре различные формы FGF-R4: полноразмерная форма с 2 полиморфными вариантами в положении 388, а именно FGF-R4 Gly388, представляющая собой нормальную форму рецептора, и форма Arg388, описанная в отношении нескольких опухолей (Bange et al., Cancer Res. 62/3,840-847 (2002); Spinola et al., J. Clin. Oncol. 23, 7307-7311 (2005); Stadler et al., Cell Signal. 18/6, 783-794(2006. Также была обнаружена растворимая форма, экспрессируемая в опухолевых клетках молочных желез (Takaishi et al., Biochem. Biophys. Res. Commun., 2000, 267:658-62). Четвертая форма, усеченная во внеклеточной части, была описана в случае некоторых гипофизарных аденом (Ezzat et al., J. Clin. Invest.,2002, 109:69-78).FGF-R4 экспрессируется главным образом в тканях, представляющих собой производные эндодермы, таких как желудочно-кишечный тракт, поджелудочная железа, печень, мышцы и надпочечник. Из литературных источников FGF-R4 известен как рецептор, играющий в клетках несколько ролей,из числа которых три главные роли описаны ниже. Во-первых, этот рецептор участвует в регуляции различных процессов дифференцировки клеток invitro и in vivo, таких как дифференцировка и регенерация скелетных мышц, дифференцировка мезенхиматозных тканей, остеогенез, а также в формировании альвеол в ходе постнатального развития печени. Во-вторых, описана причастность FGF-R4 к регуляции гомеостаза желчной кислоты и холестерина и участие в регуляции ожирения. Кроме того, равновесие между продуцированием желчной кислоты и холестерина регулируется FGF-R4 in vitro и in vivo. В-третьих, FGF-R4 участвует в некоторых опухолевых феноменах, таких как развитие гепатоцеллюлярных карцином или раковых опухолей толстой кишки, в пролиферации клеток фиброаденом молочных желез, эпителиальных клеток раковых опухолей молочных желез, в подвижности клеток карцином молочных желез или колоректальных карцином. Причастность FGF-R4 к опухолям преимуществен-1 021580 но связана с явлением полиморфизма (Gly388Arg), коррелирующего с прогрессированием развития опухолей молочных желез и колоректальных опухолей (Bange et al., Cancer Res. 62/3, 840-847, 2002), опухолей предстательной железы (Wang et al., Clin. Cancer Res. 10/18, 6169-6178, 2004) или опухолей печени(Nicholes et al., Am. J. Pathol. 160/6, 2295-307, 2002). Данный полиморфизм связан также с плохим прогнозом в случае сарком (Morimoto et al., Cancer, 98/10, 2245-2250, 2003), аденокарцином легкого (Spinolaet al., J. Clin. Oncol. 23, 7307-7311, 2005) или сквамозных сарком (da Costa Andrade et al., Exp. Mol. Pathol. 82, 53-7, 2007). Сверхэкспрессия FGF-R4 описана также в случае некоторых линий раковых опухолей поджелудочной железы (Shah et al., Oncogene, 21/54, 8251-61, 2002) и коррелируется со злокачественностью астроцитом (Yamada et al., Neurol. Res. 24/3, 244-8, 2002). Кроме того, использование моноклонального антитела анти-FGF19 в моделях in vivo ксенотрансплантатов опухолей толстой кишки или в моделях гепатоцеллюлярных карцином, показывает, что инактивация FGF19 и, таким образом, блокирование активации FGF-R4 может быть благоприятным в лечении рака толстой кишки или печени. В литературных источниках допускают, что пары FGF/FGF-R1 и FGF/FGF-R2 принимают участие в образовании новых кровеносных сосудов в контексте нормы или патологии. Однако потенциальная причастность FGF-R4 к регуляции этого клеточного феномена до настоящего времени не была исследована. На практике, до настоящего времени предполагалось, что активация ангиогенеза опосредована черезFGF-R1 и/или FGF-R2 (Presta et al., Cytokine Growth Factor Rev., 2005, 16:159-78). Примеры антагонистов FGF-R4 описаны в литературе, в частности, малые молекулы, не нацеленные специально на FGF-R4, но также вызывающие нежелательные эффекты. Так, например, малые химические молекулы, являющиеся ингибиторами домена тирозинкиназы и ингибирующие некоторые рецепторы FGF-R, а также другие рецепторы тирозинкиназы были описаны Thompson et al., (Thompson etal., J. Med. Chem., 2000, 43:4200-11). Малые химические молекулы, ингибирующие FGF-R за счет ассоциации с их внеклеточной частью, также были описаны в WO 2007/080325. Были исследованы также антитела, такие как антитела анти-FGFR1 и/или анти-FGFR4, описанные в международных заявках WO 2005/066211 и WO 2008052796 или Chen et al., (Hybridoma, 24/3, 152-159,2005). В WO 2005/037235 описаны антитела-антагонисты FGF-R для лечения ожирения и диабета. С другой стороны, антитела-агонисты анти-FGF-R4 описаны в WO 03/063893. Объектом настоящего изобретения является антагонист рецептора FGF-R4, отличающийся тем, что он специфически связывается с этим рецептором. Указанный антагонист предпочтительно представляет собой специфическое антитело к рецепторуFGF-R4. В одном из вариантов осуществления антагонист, являющийся объектом настоящего изобретения,связывается с доменом D2-D3 рецептора FGF-R4. В предпочтительном варианте осуществления антагонист, являющийся объектом настоящего изобретения, связывается с доменом D2 рецептора FGF-R4. В более предпочтительном варианте осуществления антагонист связывается с последовательностью SEQID NO:70. Специфический антагонист рецептора FGF-R4 в отношении рецептора FGF-R4 предпочтительно имеет значение KD, определенное по методике Biacore, меньше 10 Е-8 М, меньше 510 Е-9 М, меньше 210 Е-9 М или меньше 110E-9 M. В предпочтительном варианте осуществления специфический антагонист рецептора FGF-R4 одновременно является активным против FGF-R4 человека и FGF-R4 мыши. В другом предпочтительном варианте осуществления специфический антагонист рецептора FGF-R4 одновременно является активным против FGF-R4 человека, FGF-R4 мыши и FGF-R4 крысы. В предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит по меньшей мере один CDR, представляющий собой последовательность,идентичную последовательности SEQ ID NO: 9, 10, 11, 12, 13, 14, 73, 74, 75, 78, 79, 80, 83, 84, 85, 88, 89,90, 93, 94, 95, 98, 99, 100, 103, 104, 105, 108, 109 или 110, или по меньшей мере один CDR, последовательность которого отличается одной или двумя аминокислотами по сравнению с последовательностямиSEQ ID NO: 9, 10, 11, 12, 13, 14, 73, 74, 75, 78, 79, 80, 83, 84, 85, 88, 89, 90, 93, 94, 95, 98, 99, 100, 103, 104,105, 108, 109 или 110, при том, что антитело сохраняет свою специфичность связывания. В особо предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом FGF-R4, содержит CDR последовательности SEQ ID NO: 9, 10, 11, 12, 13, 14, 73, 74, 75, 78, 79, 80,83, 84, 85, 88, 89, 90, 93, 94, 95, 98, 99, 100, 103, 104, 105, 108, 109 или 110, или CDR, последовательности которых отличаются одной или двумя аминокислотами по сравнению соответственно с указанными последовательностями, если только это обстоятельство не изменяет специфичность связывания антитела с рецептором FGF-R4. В предпочтительном варианте осуществления антитела по настоящему изобретению содержат по меньшей мере одну тяжелую цепь и по меньшей мере одну легкую цепь, причем тяжелая цепь содержит три последовательности CDR, имеющие последовательности аминокислот, выбранные из группы, в которую входят SEQ ID NO: 9, 10 и 11, или 73, 74 и 75, 83, 84 и 85, или 93, 94 и 95, или 103, 104 и 105, причем указанная легкая цепь содержит три последовательности CDR, имеющие последовательности ами-2 021580 нокислот, выбранные из группы, в которую входят SEQ ID NO: 12, 13 и 14, или 78, 79 и 80, или 88, 89 и 90, или 98, 99 и 100, или 108, 109 и 110. В предпочтительном варианте осуществления вариабельные части тяжелой цепи антитела, являющегося специфическим антагонистом FGF-R4, содержат последовательность, идентичную по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 6, 77, 87, 97 или 107. В предпочтительном варианте осуществления вариабельные части легкой цепи антитела, являющегося специфическим антагонистом рецептора FGF-R4, содержат последовательность, идентичную по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 8, 72, 82, 92 или 102. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему тяжелую цепь, содержащую вариабельную часть, кодируемую нуклеотидной последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательностиSEQ ID NO: 5, 76, 86, 96 или 106. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему легкую цепь, содержащую вариабельную часть, кодируемую нуклеотидной последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQID NO: 7, 71, 81, 91 или 101. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему тяжелую цепь, содержащую вариабельную часть полипептидной последовательности SEQ ID NO: 6, 77, 87, 97 или 107. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему легкую цепь, содержащую вариабельную часть полипептидной последовательности SEQ ID NO: 8, 72, 82, 92 или 102. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему последовательности, кодируемые нуклеотидными последовательностями SEQ ID NO: 5 и 7, или 71 и 76, или 81 и 86, или 91 и 96, или 101 и 106. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему полипептидные последовательности SEQ ID NO: 6 и 8, или 72 и 77,или 82 и 87, или 92 и 97, или 102 и 107. В предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит последовательности, идентичные по меньшей мере на 80, 90, 95 или 99% последовательностям SEQ ID NO: 2 и/или SEQ ID NO: 4, или SEQ ID NO: 72 и/или SEQ ID NO: 77, илиSEQ ID NO: 82 и/или SEQ ID NO: 87, или SEQ ID NO: 92 и/или SEQ ID NO: 97, или SEQ ID NO: 102 и/или SEQ ID NO: 107. В особо предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит тяжелую цепь, кодируемую нуклеотидной последовательностью,идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 1. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему тяжелую цепь полипептидной последовательности, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 2. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему легкую цепь, кодируемую нуклеотидной последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 3. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему легкую цепь полипептидной последовательности, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 4. Настоящее изобретение относится также антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему последовательности, кодируемые нуклеотидными последовательностями SEQ ID NO: 1 и 3. В более предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит тяжелую цепь, содержащую последовательность SEQ ID NO: 2, и легкую цепь, содержащую последовательность SEQ ID NO: 4. Антитело, состоящее из тяжелой цепи последовательности SEQ ID NO: 2 и легкой цепи последовательности SEQ ID NO: 4, в дальнейшем описании обозначается как 40-12. В одном из вариантов осуществления настоящего изобретения специфические антитела к FGF-R4 активны одновременно в отношении FGF-R4 человека, FGF-R4 мыши и FGF-R4 крысы. В предпочтительном варианте осуществления специфический антагонист рецептора FGF-R4 индуцирует ингибирование клеточных путей AKT/р 38. В предпочтительном варианте осуществления специфический антагонист рецептора FGF-R4 индуцирует ингибирование клеточных путей Erk1/2. В предпочтительном варианте осуществления специфический антагонист рецептора FGF-R4 индуцирует ингибирование путей клеточной сигнализации, регулируемых FGF-R4. В другом предпочтительном варианте осуществления специфический антагонист рецептора FGF-R4 индуцирует ингибирование клеточной пролиферации опухолей. В другом предпочтительном варианте осуществления специфический антагонист рецептора FGF-R4 индуцирует ингибирование ангиогенеза. В другом особо предпочтительном варианте осуществления специфический антагонист рецептораFGF-R4 имеет сродство к FGF-R4, в 10 раз превышающее его сродство к другим рецепторам FGF. В особо предпочтительном варианте осуществления антитело по настоящему изобретению представляет собой гуманизированное антитело, являющееся специфическим антагонистом рецептора FGFR4. В одном из вариантов осуществления гуманизированное антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит легкую цепь, вариабельная часть которой кодируется нуклеотидной последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательностиSEQ ID NO: 29 или последовательности SEQ ID NO: 31. В другом варианте осуществления гуманизированное антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит легкую цепь, вариабельная часть которой идентична по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 30 или последовательности SEQ ID NO: 32. В другом варианте осуществления гуманизированное антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит легкую цепь, вариабельная часть которой кодируется последовательностью, идентичной нуклеотидной последовательности SEQ ID NO: 29 или последовательности SEQID NO: 31. Настоящее изобретение относится также к гуманизированному антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему тяжелую цепь, вариабельная часть которой кодируется последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательностиSEQ ID NO: 33, последовательности SEQ ID NO: 35 или последовательности SEQ ID NO: 37. Настоящее изобретение относится также к гуманизированному антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему тяжелую цепь, вариабельная часть которой идентична по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 34, последовательностиSEQ ID NO: 36 или последовательности SEQ ID NO: 38. Настоящее изобретение относится также к гуманизированному антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему тяжелую цепь, кодируемую нуклеотидной последовательностью SEQ ID NO: 33 и/или SEQ ID NO: 35 и/или SEQ ID NO: 37. Настоящее изобретение относится также к гуманизированному антителу, которое является специфическим антагонистом рецептора FGF-R4, и последовательности которого, гуманизированы последовательностью SEQ ID NO: 30 или 32, используют в комбинации с последовательностями, гуманизированными последовательностью SEQ ID NO: 34, 36 или 38. В другом варианте осуществления антитело, являющееся специфическим антагонистом рецептораFGF-R4, содержит CDR последовательности SEQ ID NO: 73, 74, 75, 78, 79 и 80, или CDR, последовательности которых отличаются одной или двумя аминокислотами, по сравнению соответственно с указанными последовательностями, если только это обстоятельство не изменяет специфичность связывания антитела с рецептором FGF-R4. В другом варианте осуществления антитело, являющееся специфическим антагонистом рецептораFGF-R4, содержит CDR последовательности SEQ ID NO: 83, 84, 85, 88, 89 и 90, или CDR, последовательности которых отличаются одной или двумя аминокислотами, по сравнению соответственно с указанными последовательностями, если только это обстоятельство не изменяет специфичность связывания антитела с рецептором FGF-R4. В другом варианте осуществления антитело, являющееся специфическим антагонистом рецептораFGF-R4, содержит CDR последовательности SEQ ID NO: 93, 94, 95, 98, 99 и 100, или CDR, последовательности которых отличаются одной или двумя аминокислотами по сравнению соответственно с указанными последовательностями, если только это обстоятельство не изменяет специфичность связывания антитела с рецептором FGF-R4. В другом варианте осуществления антитело, являющееся специфическим антагонистом рецептораFGF-R4, содержит CDR последовательности SEQ ID NO: 103, 104, 105, 108, 109 и 110, или CDR, последовательности которых отличаются одной или двумя аминокислотами, по сравнению соответственно с указанными последовательностями, если только это обстоятельство не изменяет специфичность связывания антитела с рецептором FGF-R4. В предпочтительном варианте осуществления настоящего изобретения антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит CDR последовательности SEQ ID NO: 83, 84, 85,88, 89 и 90. В предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, представляет собой антитело человека, в котором вариабельные части тяжелой цепи содержат нуклеотидную последовательность, идентичную по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 76, 86, 96 или 106. В предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, представляет собой антитело человека, в котором вариабельные части легкой цепи содержат нуклеотидную последовательность, идентичную по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 71, 81, 91 или 101. Настоящее изобретение относится также к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему тяжелую цепь, содержащую вариабельную часть, кодируемую белковой последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательностиSEQ ID NO: 77, 87, 97 или 107. Настоящее изобретение относится также к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему легкую цепь, содержащую вариабельную часть, кодируемую белковой последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательностиSEQ ID NO: 72, 82, 92 или 102. Настоящее изобретение относится также к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему последовательности, кодируемые нуклеотидными последовательностями SEQ ID NO: 71 и 76 или нуклеотидными последовательностями SEQ ID NO: 81 и 86, или нуклеотидными последовательностями SEQ ID NO: 91 и 96, или нуклеотидными последовательностямиSEQ ID NO: 101 и 106. Настоящее изобретение относится также к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему полипептидные последовательности SEQ ID NO: 72 и 77 или полипетидные последовательности SEQ ID NO: 82 и 87, или полипетидные последовательности SEQID NO: 92 и 97, или полипетидные последовательности SEQ ID NO: 102 и 107. В предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, содержит последовательности, идентичные по меньшей мере на 80, 90, 95 или 99% последовательностям SEQ ID NO: 72 и/или SEQ ID NO: 77. В другом предпочтительном варианте осуществления антитело человека, являющееся специфическим антагонистом рецептора FGF-R4, содержит последовательности, идентичные по меньшей мере на 80, 90, 95 или 99% последовательностям SEQ ID NO: 82 и/или SEQ ID NO: 87. В другом предпочтительном варианте осуществления антитело человека, являющееся специфическим антагонистом рецептора FGF-R4, содержит последовательности, идентичные по меньшей мере на 80, 90, 95 или 99% последовательностям SEQ ID NO: 92 и/или SEQ ID NO: 97. В другом предпочтительном варианте осуществления антитело человека, являющееся специфическим антагонистом рецептора FGF-R4, содержит последовательности, идентичные по меньшей мере на 80, 90, 95 или 99% последовательностям SEQ ID NO: 102 и/или SEQ ID NO: 107. В предпочтительном варианте осуществления настоящее изобретение относится к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему легкую цепь, кодируемую нуклеотидной последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 82, и легкую цепь полипептидной последовательности, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 87. Более предпочтительно настоящее изобретение относится к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему последовательности, кодируемые нуклеотидными последовательностями SEQ ID NO: 82 и 87. Антитело, состоящее из тяжелой цепи последовательности SEQ ID NO: 77 илегкой цепи последовательности SEQ ID NO: 72, в дальнейшем описании обозначается как клон 8. Антитело, состоящее из тяжелой цепи последовательности SEQ ID NO: 87 илегкой цепи последовательности SEQ ID NO: 82, в дальнейшем описании обозначается как клон 31. Антитело, состоящее из тяжелой цепи последовательности SEQ ID NO: 97 и легкой цепи последовательности SEQ ID NO: 92, в дальнейшем описании обозначается как клон 33. Антитело, состоящее из тяжелой цепи последовательности SEQ ID NO: 107 илегкой цепи последовательности SEQ ID NO: 102, в дальнейшем описании обозначается как клон 36. Объем охраны настоящего изобретения не ограничивается антителами, содержащими эти последовательности. На практике любые антитела, которые специфически связываются с FGF-R4, оказывая антагонистическое действие на этот рецептор, входят в объем охраны настоящего изобретения. Настоящее изобретение относится также к антителу, являющемуся специфическим антагонистом рецептора FGF-R4 и конъюгированному с цитотоксическим агентом. Настоящее изобретение относится к применению специфического антагониста рецептора FGF-R4 в лечении заболеваний, связанных с ангиогенезом. Настоящее изобретение относится к применению специфического антагониста рецептора FGF-R4 в лечении рака. Настоящее изобретение относится к применению специфического антагониста рецептора FGF-R4 в лечении гепатокарцином или любых других видов рака печени. Настоящее изобретение относится к применению специфического антагониста рецептора FGF-R4 в лечении рака поджелудочной железы. Настоящее изобретение относится к специфическому антителу к рецептору FGF-R4, который может быть использован одновременно в лечении заболеваний, связанных с ангиогенезом, и в лечении гепатокарцином или любых других видов рака печени. Настоящее изобретение относится к специфическому антителу к рецептору FGF-R4, который может быть использован одновременно в лечении заболеваний, связанных с ангиогенезом, в лечении гепатокарцином или любых других видов рака печени, в лечении рака поджелудочной железы, органов желудочно-кишечного тракта или любых других органов, экспрессирующих FGF-R4. Подробное описание изобретения Настоящее изобретение относится к применению антитела, специфически направленного к FGF-R4(без перекрестной реакции с FGF-R1, R2 или R3), для ингибирования ангиогенеза и роста опухолей. Неожиданным образом авторами было обнаружено, что FGF-R4 играет активную и специфическую роль в регуляции ангиогенеза. Такая функция FGF-R4 ранее не была обнаружена или предложена. Таким образом, этот рецептор может быть использован в качестве мишени для лечения патологий, при которых имеет место ангиогенная дисфункция. Лиганды FGF-R4, способные модулировать его активность, представляют собой, таким образом, потенциальные вещества терапевтического действия в случае многих патологий, связанных с ангиогенезом. Таким образом, настоящее изобретение может быть использовано в лечении любых патологий, вызывающих нарушение регуляции ангиогенеза и требующих его ингибирования. Релевантные патологии могут представлять собой рак, требующий применения антагонистов по настоящему изобретению в качестве ингибиторов ангиогенеза опухолей, или патологии, при которых нарушение регуляции ангиогенеза описывается как возрастная дегенерация желтого пятна или DMLA, воспалительные заболевания, такие как ревматоидный артрит, остеоартрит, колиты, язвы или любые воспалительные заболевания кишечника, атеросклероз или ожирение. Применение таких антител также пояснено в случае ингибирования роста опухолей. Таким образом, антагонисты по настоящему изобретению могут быть использованы для лечения некоторых раковых заболеваний, вызывающих нарушение регуляции FGF-R4, и более предпочтительно раковых заболеваний печени, толстой кишки, молочной железы,легкого, предстательной железы, поджелудочной железы, кожи или пищевода. Одним из наибольших преимуществ антагонистов по настоящему изобретению является выбор в качестве специфической мишени рецептора FGF, в данном случае FGF-R4. Такая специфичность позволяет ограничить нежелательные эффекты, которые могут вызывать малые химические молекулы, являющиеся ингибиторами домена тирозинкиназы. Кроме того, поскольку FGF-R4 экспрессируется не повсеместно, а экспрессируется, в частности, на эндотелиальных клетках, таких как клетки печени, желчного пузыря, молочных желез, предстательной железы, яичника, поджелудочной железы, почек, это обстоятельство предоставляет в распоряжение способ лечения заболеваний, связанных с нарушением регуляции активности FGF-R4, с ограничением вторичных явлений. Под "антагонистом" понимают любой лиганд, способный уменьшить или полностью ингибировать активность FGF-R4. Такое соединение-антагонист также называют ингибитором FGF-R4. Такой антагонист может представлять собой любой лиганд FGF-R4, такой как химическая молекула, рекомбинантный белок, олигосахарид, полисахарид, олигонуклеотид или антитело, способные специфически связываться с рецептором FGF-R4, исключая любые другие FGFR. Таким образом, объектом настоящего изобретения является специфический антагонист FGF-R4. Согласно настоящему изобретению под антагонистом, специфически связывающимся с FGF-R4, понимают лиганд, который не связывается с другими рецепторами FGF, а именно с FGF-R1, FGF-R2 или FGFR3. В частности, специфическое антитело к FGF-R4 представляет собой антитело, которое не вступает в перекрестную реакцию с FGF-R1, FGF-R2 или FGF-R3. Под специфической связью понимают по меньшей мере 10-кратное различие между силой связи с одним рецептором по сравнению с другим, в данном случае между связью с FGF-R4 и возможными связями с FGF-R1, FGF-R2 или FGF-R3. В одном из вариантов осуществления настоящего изобретения лиганд FGF-R4 представляет собой антитело. Под "антителом" понимают антитела или производные молекулы любого типа, такие как поликлональные и моноклональные антитела. В число производных молекул моноклональных антител входят гуманизированные антитела, антитела человека, мультиспецифические антитела, химерные антитела,фрагменты антител, нанотела. В одном из вариантов осуществления настоящего изобретения специфический антагонист FGF-R4 представляет собой поликлональное антитело."Поликлональное антитело" представляет собой антитело, полученное в смеси антител, происходящих от нескольких клонов лимфоцитов В и распознающих серию различных эпитопов. В предпочтительном варианте осуществления специфический антагонист FGF-R4 представляет собой моноклональное антитело."Моноклональное антитело" представляет собой антитело, полученное исходя из популяции, по существу, однородных антител, происходящей от одного типа лимфоцитов В и амплифицированной клонированием. Антитела, образующие такую популяцию, являются идентичными, за исключением возможных естественных мутаций, которые могут содержаться в очень малом количестве. Такие антитела направлены против одного эпитопа и, таким образом, являются высокоспецифическими. Под "эпитопом" понимают сайт антигена, с которым связывается антитело. Если антиген представляет собой полимер, такой как белок или полисахарид, то эпитоп может быть образован смежными или несмежными остатками. В предпочтительном варианте осуществления по настоящему изобретению антитело-антагонист анти-FGF-R4 связывается с эпитопом, принадлежащим домену D2-D3 рецептора FGF-R4. В более предпочтительном варианте антитело связывается с эпитопом, входящим в домен, содержащий аминокислоты 144-365 рецептора FGF-R4. В наиболее предпочтительном варианте антитело связывается с эпитопом, входящим в домен D2 рецептора FGF-R4, причем этот эпитоп соответствует аминокислотам 145-242, описанным в последовательности SEQ ID NO:70. Антитело, называемое также иммуноглобулиновым, состоит из двух одинаковых тяжелых цепей("VH") и двух одинаковых легких цепей ("VL"), связанных дисульфидным мостиком. Каждая цепь содержит константную и вариабельную области. Каждая вариабельная область содержит три сегмента, называемых "областями, определяющими комплементарность" ("CDR") или "гипервариабельными областями", которые являются принципиально ответственными за связь антигена с эпитопом. Термин "VH" относится к вариабельным областям тяжелой цепи иммуноглобулина антитела, включающего в себя тяжелые цепи фрагментов Fv, scFv, dsFv, Fab, Fab' или F(ab)'. Термин "VL" относится к вариабельным областям легкой цепи иммуноглобулина антитела, включающего в себя легкие цепи фрагментов Fv, scFv, dsFv, Fab, Fab' или F(ab)'. Под "фрагментом антитела" понимают любую его часть: Fab (антигенсвязывающий фрагмент), Fv,scFv (одиночная цепь Fv), Fc (кристаллизуемый фрагмент). Данные функциональные фрагменты предпочтительно представляют собой фрагменты типа Fv, scFv, Fab, F(ab')2, Fab', scFv-Fc или димерные тела,имеющие, в общем случае, такую же специфичность связывания, что и моноклональное, химерное или гуманизированное антитело, от которого они происходят. Согласно настоящему изобретению фрагменты антител по настоящему изобретению могут быть получены исходя из моноклональных, химерных или гуманизированных антител такими способами, как гидролиз ферментами, такими как пепсин или папаин,и/или расщеплением дисульфидных мостиков химическим восстановлением. Под "областью или областями CDR" понимают гипервариабельные области тяжелых и легких цепей иммуноглобулинов, такие как области, определенные Kabat et al. (Kabat et al., Sequences of proteins ofimmunological interest, 5t" Ed., US Department of Health and Human Services, NIH, 1991, и последующие издания). Существуют 3 CDR тяжелой цепи и 3 CDR легкой цепи. Термин "область или области CDR" используется в настоящем описании для обозначения в соответствующих случаях одной или нескольких из таких областей или совокупности таких областей, содержащих большинство остатков аминокислот,ответственных за аффинную связь антитела с антигеном или эпитопом, распознаваемым им. Наиболее консервативные области вариабельных доменов называют областями или последовательностями FR("framework" (каркас, или "каркасными областями". В одном из вариантов осуществления настоящего изобретения специфический антагонист FGF-R4 представляет собой химерное антитело. Под "химерным антителом" понимают антитело, в котором константная область или ее часть изменена, заменена или замещена, так что вариабельная область оказывается связанной с константной областью, которая отличается видом или принадлежит к другому классу или подклассу антител. Под "химерным антителом" понимают также антитело, в котором вариабельная область или ее часть изменена, заменена или замещена так, что константная область оказывается связанной с вариабельной областью, которая отличается видом или принадлежит к другому классу или подклассу антител. Способы получения химерных антител известны специалистам в данной области техники, см., например, Morrison, 1985, Science, 229:1202; Oi et al., 1986, Bio Techniques, 4:214; Gillies et al., 1989, J. Immunol. Methods, 125:191-202; патенты US 5807715, US 4816567 и US 4816397. Такие химерные версии антител могут представлять собой слияние вариабельных частей VL и VH с константными частями Скарра и СН (IgG1) человеческой природы для получения моноклонального химерного антитела. Часть СН (IgG1) также может быть модифицирована точечными мутациями для повышения сродства фрагмента Fc к рецептору FcRIIIa и, тем самым, повышения эффекторных функций антитела (Lazar etal., 2006, Proc. Natl. Acad. Sci. USA, 103: 4005-4010; Stavenhagen et al., 2007, Cancer Res. 67: 8882-8890). Настоящее изобретение относится к гуманизированным версиям антител. Под "гуманизированным антителом" понимают антитело, которое содержит, главным образом, иммуноглобулиновые последовательности человека. Этот термин в общем случае относится к отличному от человеческого иммуноглобулину, модифицированному введением последовательностей человека или остатков, содержащихся в последовательностях человека. В общем случае гуманизированные антитела содержат один или, как правило, два вариабельных домена, в которых все или часть областей CDR соответствуют исходным частям родственной последовательности, отличной от последовательности человека, и в которых все или часть областей FR представляют собой области, происходящие от иммуноглобулиновой последовательности человека. При этом гуманизированное антитело может содержать по меньшей мере часть константного участка иммуноглобулина (Fc), в частности константного участка выбранного референтного иммуноглобулина человека. Таким образом, пытаются получить антитело, которое является наименее иммуногенным в отношении человека. Также можно заменять одну или две аминокислоты одного или нескольких CDR аминокислотами, являющимися менее иммуногенными в отношении человека, существенно не уменьшая специфичность связывания антитела с FGF-R4. Остатки каркасных областей также могут не представлять собой человеческие, и их можно не модифицировать, если они не способствуют иммуногенному потенциалу антитела. Существует несколько известных специалистам в данной области техники способов гуманизации для модификации родственного антитела, отличающегося от антитела человека, до антитела, являющегося менее иммуногенным в отношении человека. Глобальная идентичность последовательностей антителам человека не является обязательно необходимой. На практике глобальная идентичность последовательности необязательно является предсказуемым индикатором пониженной иммуногенности, а модификация ограниченного числа остатков может приводить к гуманизированным антителам, обладающим иммуногенным потенциалом, являющимся сильно ослабленным в отношении человека (Molecular Immunology (2007) 44, 1986-1998). К различным способам относятся, например, внедрение CDR (трансплантация) (ЕРО 0239400, WO 91/09967, US 5530101 и US 5585089), ресурфейсинг (ЕРО 0592106, ЕРО 0519596; Padlan, 1991, Molec.Imm. 28 (4/5): 489-498; Studnicka et al., 1994, Prot. Eng. 7 (6): 805-814; Roguska et al., 1994, PNAS 91:969973) или также смешивание цепей (US 5565332). Настоящее изобретение предпочтительно относится к гуманизированным антителам, вариабельные части которых модифицированы согласно описанным ниже технологиям. Легкие и тяжелые цепи, являющиеся наиболее сходными с соответствующими цепями антитела мыши анти-FGF-R4 40-12, идентифицируют сравнением с базой Protein Data Bank (H.M. Berman, J. Westbrook, Z. Feng, G. Gilliland, T.N. Bhat, H. Weissig, I.N. Shindyalov, P.E. Bourne. Nucleic Acids Research,2000, 28:235-242). Выравнивание последовательностей осуществляют, используя алгоритм BLAST (J.Mol. Biol. 1990, Oct. 215:403-410). Речь идет о трехмерных структурах, соответствующих кодам PDB 1NDM1ETZ и соответственно используемых для конструирования моделей по гомологии легких и тяжелых цепей вариабельных доменов. Такие трехмерные модели затем уточняют, минимизируя их потенциальную энергию по стандартной методике, реализованной в программе МОЕ (Molecular OperatingEnvironment, Chemical Computing Group, Квебек, Канада). Затем посредством программы Amber осуществляют молекулярно-динамическое моделирование (DM), исходя из таких минимизированных трехмерных моделей антител (D.A. Case, Т.Е. Cheatham, III, Т. Darden, H. Gohlke, R. Luo, K.M. Merz, Jr., A. Onufriev, C. Simmerling, B. Wang and R. Woods. J. Computat. Chem. 2005, 26:1668-1688). Такое моделирование осуществляют с гармоническими напряжениями, приложенными к атомам белкового каркаса, при температуре 500 К в течение 1,1 нс в модели неявного растворителя, такого как растворитель, реализованный в обобщенном способе Борна (GallicchioLevy, J. Comput. Chem. 2004, 25:479-499). Таким образом,таким первичным моделированием получают десять различных конформаций из расчета одной трехмерной конформаций на каждые сто пикосекунд в течение последней наносекунды моделирования. Затем с каждой из полученных десяти различных конформаций осуществляют молекулярно-динамическое моделирование без приложения напряжений к белковому каркасу при температуре 27 С в течение 2,3 нс в модели неявного растворителя, такого как растворитель, реализованный в обобщенном способе Борна. К связям, вовлекающим атомы водорода, прикладывают напряжения по алгоритму SHAKE (Barth et al., J.Comp. Chem., 1995, 16:1192-1209), один шаг длится 1 фемтосекунду, и моделирование осуществляют согласно уравнению Ланжевена при постоянном объеме и температуре 27 С. Затем в каждой из десяти молекулярно-динамических моделей последние две тысячи конформаций, полученных с частотой в одну пикосекунду, используют для количественного определения смещения положений атомов для каждой аминокислоты гуманизируемого антитела по сравнению со средней конформацией (называемой медоидом) аминокислоты. Совокупность анализа, описанного ниже, осуществляют на скриптовом языке Scientific Vector Language (SVL) программы МОЕ. Конформация-медоид аминокислоты представляет собой исходную молекулярно-динамическую конформацию, являющуюся наиболее близкой к средней конформаций, рассчитанной, исходя из положений атомов всех конформаций аминокислоты. Расстояние,используемое для детектирования конформации-медоида, равно среднеквадратичному отклонению скалярных расстояний между атомами двух конформаций аминокислоты. Аналогичным образом, смещение положений атомов конформаций аминокислоты по сравнению с конформацией-медоидом количественно определяют, рассчитывая среднеквадратичное отклонение скалярных расстояний между атомами аминокислоты смоделированной конформаций и теми же атомами конформации-медоида. Далее сравнивая среднеквадратичное отклонение положений атомов аминокислоты (i), являющейся средней в совокупности десяти молекулярно-динамических моделей (Fi), со среднеквадратичным отклонением положений всех аминокислот антитела, являющегося средним для совокупности десяти молекулярно-динамических моделей (Fm), принимают решение, является ли аминокислота достаточно гибкой, чтобы рассматривать ее в качестве кислоты, которая может потенциально взаимодействовать с рецепторами Т-клеток и провоцировать активацию иммунной системы. Аминокислоту (i) рассматривают в качестве гибкой, если ее показатель гибкости Zi превышает 0,15 при расчете по формуле: Zi=(Fi-Fm)/Fm. Таким образом, были идентифицированы как гибкие 45 аминокислот в вариабельном домене антитела, за исключением участка, определяющего комплементарность к антигену (RDC), и его непосредственного окружения. Непосредственное окружение RDC определяют как любую аминокислоту, в которой атом углерода в положении располагается на расстоянии 5 или меньше от атома углерода в -положении RDC. Затем перемещения 60 наиболее гибких аминокислот антитела в течение 20 нс (102 нс) моделирования сравнивают с перемещениями аминокислот, соответствующих 49 моделям по гомологии терминальных последовательностей антитела человека, для каждой из которых были реализованы десять молекулярно-динамических моделей (102 нс) по той же методике. 60 наиболее гибких аминокислот не входят в область, определяющую комплементарность к антигену (RDC), и ее непосредственное окружение. 49 моделей терминальных последовательностей антитела человека были построены систематическим комбинированием 7 наиболее часто встречающихся легких цепей человека (vk1, vk2, vk3, vk4,vlambda1, vlambda2, vlambda3) и 7 наиболее часто встречающихся тяжелых цепей человека (vh1a, vh1b,vh2, vh3, vh4, vh5, vh6) (Nucleic Acids Research, 2005, Vol. 33, Database issue D593-D597). Сходство гуманизируемого антитела с 49 моделями терминальных последовательностей человека количественно определяют, калибруя положения некоторых атомов 60 гибких аминокислот антитела, в ходе десяти вариантов молекулярно-динамического моделирования посредством одной и той же трехмерной кубической решетки с разрешением 1 . В данном случае, имеется в виду четырехмерное сходство. Используемая трехмерная решетка содержит 445740 точек, и ее инициализируют, исходя из трехмерной структуры антитела человека, соответствующего коду PDB 8FAB. Структуру 8FAB используют также для позиционирования всех конформаций антитела, калибруемых в трехмерной решетке. С этой целью молекулярно-динамическую конформацию-медоид антитела накладывают на структуру 8FAB. Такое наложение представляет собой выравнивание моментов инерции двух конформаций с последующей оптимизацией скалярных расстояний между атомами углерода в -положении обеих конформаций. Все другие молекулярно-динамические конформаций антитела накладывают на конформацию-медоид таким же способом. Осуществляют два типа калибровки с получением двух вариантов сходства (электростатическое сходство и сходство по липофильности) для пары сравниваемых антител, которые суммируют для получения тотального сходства. При первой, электростатической, калибровке рассматривают все атомы боковой цепи аминокислоты. Значение в точке х решетки рассчитывают, используя в отношении атомов боковой цепи аминокислоты трехмерную функцию Гаусса f(x), которую уравновешивают с электростатическим зарядом атома, согласно описанию в силовом поле Amber99 (Cieplak J. et al.,J. Comp. Chem. 2001, 22: 1048-1057). Функция f(x) ориентирована по 3 осям декартовых координат и соответствует формуле где х и u представляют собой декартовы координаты точки х решетки и эталонного атома соответственно, s=r/1,6 (r означает ковалентный радиус атома). Калибровку повторяют для всех конформаций аминокислоты и полученные результаты усредняют по всем точкам х трехмерной решетки. При второй калибровке по лиофильности рассматривают только липофильные атомы боковой цепи аминокислоты. Значение в точке х решетки рассчитывают с помощью такой же функции Гаусса f(х) без уравновешивания. Таким образом, оба множества молекулярно-динамических конформаций обоих сравниваемых антител калибруют по одной и той же трехмерной решетке. Электростатическое сходство (sim-elec) 2 антител а и Сходство по липофильности рассчитывают по такой же формуле по конечным данным калибровки по липофильности, описанной выше. Таким образом, модель терминальных последовательностей человека vlambda2-vh2 показывает наиболее сильное четырехмерное сходство (тотальное сходство составляет 58%) 60 гибких аминокислот по сравнению с гибкими аминокислотами антитела мыши 40-12. Таким образом, для гуманизации гуманизируемого антитела была использована модель терминальной последовательности человека vlambda2vh2 с фокусированием на 45 гибких аминокислотах. Для определения мутаций, подлежащих исполне-9 021580 нию, трехмерную структуру модели антитела мыши 40-12 накладывают на модель, полученную для терминальных последовательностей, имеющих наиболее сильное сходство, оптимизируя положения атомов углерода в -положении аминокислот. Аминокислоты, идентифицированные как гибкие, заменяют соответствующими аминокислотами в модельной последовательности, имеющей наиболее сильное сходство. К рассмотренным нежелательным мотивам последовательностей относятся: аспартат-пролин (пептидная связь, являющаяся неустойчивой в кислой среде), аспарагин-Х-серин/треонин (сайт гликозилирования, X означает любую аминокислоту, за исключением пролина), аспартат-глицин/серин/треонин (потенциальное образование звена сукцинимид/изоаспартат в гибких зонах), аспарагин-глицин/гистидин/серин/аланин/цистеин (сайты, подверженные деамидированию), метионин (окисление в экспонируемых зонах). Полученные таким образом гуманизированные последовательности в итоге сравнивают по алгоритму BLAST сравнения последовательностей с последовательностями базы данных IEDBPLoS Biol. 2005, Mar., 3(3):e91) для проверки того, что последовательности не содержат известные эпитопы, обеспечивающие распознавание клетками лимфоцитов В и Т. Если последовательность содержит остатки, которые представляют собой нежелательные последовательности, то их также модифицируют. Если комбинированная последовательность содержит известный эпитоп, описанный в IEDB, то в качестве модели используют другую референтную терминальную структуру, обладающую сильным сходством. Более предпочтительно антитело по настоящему изобретению содержит последовательности, идентичные по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 30 или SEQ ID NO: 32, в комбинации с последовательностями, идентичными по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 34, последовательности SEQ ID NO: 36 или последовательности SEQ ID NO: 38. В одном из вариантов осуществления антитело по настоящему изобретению содержит вариабельные легкие цепи последовательности SEQ ID NO: 30 и вариабельные тяжелые цепи последовательностиSEQ ID NO: 34. В другом варианте осуществления антитело по настоящему изобретению содержит вариабельные легкие цепи последовательности SEQ ID NO: 32 и вариабельные тяжелые цепи последовательности SEQID NO: 38. В другом варианте осуществления антитело по настоящему изобретению содержит вариабельные легкие цепи последовательности SEQ ID NO: 30 и вариабельные тяжелые цепи последовательности SEQID NO: 36. В другом варианте осуществления антитело по настоящему изобретению содержит вариабельные легкие цепи последовательности SEQ ID NO: 32 и вариабельные тяжелые цепи последовательности SEQID NO: 34. В другом варианте осуществления антитело по настоящему изобретению содержит вариабельные легкие цепи последовательности SEQ ID NO: 32 и вариабельные тяжелые цепи последовательности SEQID NO: 36. В другом варианте осуществления антитело по настоящему изобретению содержит вариабельные легкие цепи последовательности SEQ ID NO: 30 и вариабельные тяжелые цепи последовательности SEQID NO: 38. Настоящее изобретение относится также к антителам человека, являющимся специфическими антагонистами FGF-R4. Такие антитела могут быть получены способами фагового отображения, известными специалистам в данной области техники (McCafferty J. et al., 1990; Hoogenboom H.R. et al., 2005). Могут быть использованы другие технологии получения антител человека, такие как технология XenoMouse,описанная в US 5939598. В предпочтительном варианте осуществления антитело, являющееся специфическим антагонистом рецептора FGF-R4, представляет собой антитело человека, в котором вариабельные части тяжелой цепи содержат последовательность, идентичную по меньшей мере на 80, 90, 95 или 99% последовательностиSEQ ID NO: 76, 86, 96 или 106. В другом варианте осуществления антитело, являющееся специфическим антагонистом рецептораFGF-R4, представляет собой антитело человека, в котором вариабельные части легкой цепи содержат последовательность, идентичную по меньшей мере на 80, 90, 95 или 99% последовательности SEQ IDNO: 71, 81, 91 или 101. Настоящее изобретение относится также к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему тяжелую цепь, содержащую вариабельную часть, кодируемую нуклеотидной последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 77, 87, 97 или 107. Настоящее изобретение относится также к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и имеющему легкую цепь, содержащую вариабельную часть, кодируемую нуклеотидной последовательностью, идентичной по меньшей мере на 80, 90, 95 или 99% последовательности SEQ ID NO: 72, 82, 92 или 102. Настоящее изобретение относится также антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему последовательности, кодируемые нуклеотидными последова- 10021580 тельностями SEQ ID NO: 71 и 76 или нуклеотидными последовательностями SEQ ID NO: 81 и 86 или нуклеотидными последовательностями SEQ ID NO: 91 и 96, или нуклеотидными последовательностямиSEQ ID NO: 101 и 106. Настоящее изобретение относится также к антителу человека, являющемуся специфическим антагонистом рецептора FGF-R4 и содержащему полипептидные последовательности SEQ ID NO: 72 и 77 или полипетидные последовательности SEQ ID NO: 82 и 87, или полипетидные последовательности SEQID NO: 92 и 97, или полипетидные последовательности SEQ ID NO: 102 и 107. Более предпочтительно антитело человека, являющееся специфическим антагонистом рецептораFGF-R4, содержит полипептидные последовательности SEQ ID NO: 82 и 87. Модифицированные таким образом последовательности аминокислот могут быть модифицированы также посредством посттрансляционных изменений в ходе продуцирования в клетке млекопитающего. В частности, использование неполных стабильных линий в биосинтезе фукозы может обеспечивать получение моноклональных антител, в которых N-гликан Fc (положение 297) частично или полностью не содержит фукозу и обеспечивает усиление действия эффектора ADCC (Kanda et al., 2006, Biotechnol. Bioeng. 94:680-688 и Ripka et al., 1986, Arch. Biochem. Bioph. 249: 533-545). В другом варианте осуществления настоящего изобретения специфический антагонист FGF-R4 представляет собой конъюгированное антитело. Антитела могут быть конъюгированы с цитотоксическим агентом. Термин "цитотоксический агент", в данном случае, означает вещество, которое затрудняет или блокирует функционирование или рост клеток или вызывает разрушение клеток. В одном из вариантов осуществления антитело или его связывающий фрагмент может быть конъюгировано с лекарственным средством, таким как маитансиноид, для образования "пролекарства", обладающего цитотоксичностью по отношению к клеткам, экспрессирующим антиген. Цитотоксический агент по настоящему изобретению может представлять собой любое соединение,следствием действия которого является гибель клетки, или которое индуцирует гибель клетки, или которое различными способами снижает жизнеспособность клеток. В число предпочтительных цитотоксических агентов входят, например, маитансиноиды и аналоги маитансиноидов, таксоиды, СС-1065 и аналоги СС-1065, доластатин и аналоги доластатина, определенные выше. Такие цитотоксические агенты конъюгированы с антителами, фрагментами антител, функциональными эквивалентами, улучшенными антителами и их аналогами, как описано в настоящем описании. Конъюгированные антитела могут быть получены способами in vitro. Для связывания лекарственного средства или пролекарства с антителом используют связующую группу. Подходящие связующие группы хорошо известны специалистам в данной области техники и включают, в частности, дисульфидные группы, тиоэфирные группы, лабильные кислотные группы, фотонестабильные группы, лабильные пептидазные группы и лабильные эстеразные группы.Предпочтительные связующие группы представляют собой дисульфидные и тиоэфирные группы. Например, конъюгированное соединение может быть получено по реакции дисульфидного обмена или за счет образования тиоэфирного мостика между антителом и лекарственным средством или пролекарством. Такие соединения, как метотраксетат, даунорубицин, винкристин, винбластин, мелфалан, митомицин С, хлорамбуцил, калихеамицин, тубулизин и аналоги тубулизина, дуокармицин и аналоги дуокармицина, доластатин и аналоги доластатина, являются также подходящими для получения конъюгированных соединений по настоящему изобретению. Молекулы также могут быть связаны с молекулами антитела посредством промежуточной молекулы, такой как сывороточный альбумин. Доксорубицин и соединения доксорубицина, как описано, например, в US 09/740991, также могут представлять собой подходящие цитотоксические агенты. Антитела, представляющие собой объект настоящего изобретения, могут быть связаны с цитотоксической молекулой или соединением. Они также могут быть связаны с антиангиогенным соединением,действующим на другие пути ангиогенеза. Под "клетками, экспрессирующими FGF-R4" понимают любую эукариотную клетку, в частности,млекопитающих и предпочтительно человека, которая экспрессирует рецептор FGF-R4 в его нативной или мутантной форме. FGF-R4 также может находиться в своей полноразмерной или усеченной форме,содержащей, например, внеклеточный домен FGR-R4, в частности домены D2-D3. FGF-R4 также может быть рекомбинирован в химерную форму. Соединения по настоящему изобретению могут быть введены в фармацевтические композиции для введения топическим, пероральным, парентеральным, интраназальным, внутривенным, внутримышечным, подкожным, интраокулярным и т.д. путями. Фармацевтические композиции предпочтительно содержат фармацевтически приемлемые носители для состава, предназначенного для инъекций. Речь может идти, в частности, о солевых растворах (мононатрийфосфат, динатрийфосфат, хлориды натрия, калия, кальция или магния и т.д. или смеси таких солей), стерильных растворах, изотонических растворах или сухих композициях, в частности лиофилизованных композициях, которые при соответствующем добавлении стерилизованной воды или физиологического раствора дают возможность получения растворов для инъекций. Релевантные патологии могут представлять собой любые заболевания, связанные с ангиогенезом опухолевой или неопухолевой природы. Релевантные патологии могут представлять собой рак (требующий использования антагонистов по настоящему изобретению в качестве ингибиторов ангиогенеза опухолей), в частности рак печени, толстой кишки, молочной железы или легкого, предстательной железы, поджелудочной железы или кожи,или такие патологии, при которых нарушение регуляции ангиогенеза описывается как возрастная дегенерация желтого пятна или DMLA, воспалительные заболевания, такие как ревматоидный артрит, остеоартрит, колиты, язвы или любые воспалительные заболевания кишечника, атеросклероз или ожирение. Антитела анти-FGF-R4 также могут быть использованы для лечения рака, в частности гепатокарциномы и других раковых заболеваний печени, и рака поджелудочной железы в качестве ингибитора, оказывающего действие непосредственно на рост опухоли. Настоящее изобретение поясняется приведенными далее примерами без ограничения объема охраны. Описание фигур Фиг. 1 А и 1 В. Эндотелиальные клетки человека типа HUVEC способны in vitro (фиг. 1 А) образовывать сеть псевдотубул, называемую ангиогенной. Эту сеть стимулируют добавлением 1 нг/мл FGF2. Такая индукция может быть получена также добавлением 10 нг/мл FGF19, представляющего собой специфический лиганд FGF-R4, тогда как добавление 10 нг/мл FGF4 (неактивирующий лиганд FGF-R4) не способно стимулировать ангиогенез. Аналогично in vivo (фиг. 1B), на мышиной модели индукции ангиогенеза в имплантате губки на спине мыши, FGF2 способен индуцировать увеличение числа новых сосудов, являющихся функциональными по отношению к губке, что характеризуется увеличением уровня гемоглобина в таких губках, по сравнению с контролем. FGF19 также способен индуцировать такой ангиогенез в губке. Фиг. 2 А. Карта плазмиды pXL4 614, обеспечивающей экспрессию hFGFR4-Histag (SEQ ID NO: 40). Фиг. 2 В. Карта плазмиды pXL4 613, обеспечивающей экспрессию hFGFR4-Streptag (SEQ ID NO: 69). Фиг. 3. Карта плазмиды pXL4615, обеспечивающей экспрессию hFGFR4(D2,D3)-Histag (SEQ IDSEQ ID NO: 48). Фиг. 7. Пластины ELISA, покрытые 4 рецепторами человека FGF. Способность антител антиFGFR4 40-12 и 64-12 распознавать различные FGF-R определяют анализом ELISA. Клон 40-12 (гистограмма черного цвета) является специфическим к FGF-R4. Кроме того, клон 64-12 (гистограмма серого цвета) распознает в очень малой степени FGF-R3. Фиг. 8 А и 8 В. Активное антитело-антагонист 40-12 и его инактивный контроль 64-12 не оказывают собственного действия на базальный ангиогенез. Напротив, при дозе 30 мкг/мл (приблизительно 200 нМ) моноклональное антитело анти-FGFR4 40-12 способно ингибировать ангиогенез клеток HUVEC, индуцированный FGF2, тогда как контрольное антитело 64-12 не способно на такое действие (фиг. 8 А). FGF2,меченный реактивом AlexaFluo r 488 нм, способен связываться с FGF-R4, экспрессируемым клетками 300-19 (гистограмма белого цвета). Это взаимодействие может быть разрушено немеченым FGF2 (гистограммы черного цвета) или антителом анти-FGFR4 40-12 (гистограммы темно-серого цвета), тогда как антитело-свидетель 64-12 не способно на такое действие (гистограммы светло-серого цвета) (фиг. 8 В). Фиг. 8 С и 8D. Действие антител анти-FGFR4, происходящих от клонов 8, 31, 33 и 36, при 10 мкг/мл на ангиогенез in vitro, индуцированный 3 нг/мл FGF-2. Клоны 8 (фиг. 8 С) и 31, 33 и 36 (фиг. 80) ингибируют ангиогенез клеток HUVEC, индуцированный FGF-2. Фиг. 9 А и 9 В. Клетки гепатокарциномы человека Нер 3b стимулируют посредством FGF19 при 30 нг/мл. Такая стимуляция индуцирует специфическую клеточную сигнализацию FGF-R4, приводящую к синтезу белков cFos и JunB и к фосфорилированию Erk1/2, наблюдаемому способом вестерн-блоттинга(фиг. 9 А). При этом для каждой полосы осуществляют количественное определение. Такое количественное определение представлено в виде гистограммы (фиг. 9 В). Антитело 40-12 при 100 мкг/мл полностью ингибирует индукцию специфической клеточной сигнализации посредством FGF-R4, тогда как контрольное антитело не оказывает какого-либо действия. На практике антитело 40-12 полностью блокирует синтез JunB и cFos, а также фосфорилирование Erk1/2, индуцированное посредством FGF19 после стимуляции в течение 3 ч. Фиг. 9 С. Ингибирующее действие антитела анти-FGFR4 40-12 на фосфорилирование Erk1/2, индуцированное в клетках Нер 3b посредством FGF-19 (30 нг/мл), подтверждено по методике ELISA с антифосфоERk1/2. Антитело 40-12 также способно ингибировать фосфорилирование Erk1/2 в клетках Нер 3b,стимулированное посредством FGF-2 (1 нг/мл) или 10%-й сывороткой (SVF). Фиг. 9D. Процентная степень ингибирования фосфорилирования Erk1/2, индуцированного посредством FGF-19 (30 нг/мл), по методике ELISA в клетках Нер 3b с использованием антител, происходящих от клонов 8, 31, 33 и 36. Фиг. 10 А и 10 В. Пролиферация клеток Нер 3b может быть стимулирована добавлением сыворотки(фиг. 10 А) или FGF19 (фиг. 10 В). Индукция сывороткой частично ингибирована антителом 40-12 при 100 мкг/мл, тогда как контрольное антитело не оказывает такого действия (фиг. 10 А). Пролиферация,индуцированная посредством FGF19, полностью блокирована антителом анти-FGFR4 40-12 при 10 мкг/мл, тогда как контрольное антитело при такой же дозе не способно на такое действие (фиг. 10 В). Фиг. 11 А-11D. Антитело анти-FGFR4 40-12 способно уменьшить развитие опухолей поджелудочной железы на мышиной модели RipTag, ингибируя ангиогенез опухоли. Модель Rip1-Tag2 представляет собой мышиную модель, в которой трансгенные мыши экспрессируют антиген Т SV40 в -клетках островков поджелудочной железы, продуцирующих инсулин (Hanahan D. Nature, 1985, 9-15:115-22.). Такой антиген Т экспрессируется в ходе эмбрионального развития поджелудочной железы до 4-5 недель жизни без очевидного действия. При этом часть островков поджелудочной железы, экспрессирующих антиген Т, прогрессирует в течение следующих 5 недель с образованием ангиогенных островков, связанных с активацией сосудистой сети, и затем с развитием небольших опухолей типа аденомы. Спустя несколько недель некоторые аденомы развиваются с образованием инвазивных карцином (фиг. 11 А). Мышам Rip1Tag2 подкожно вводили один раз в неделю в промежутке от 10 до 13 недель дозы 25 мг/кг антитела 40-12 или контрольного антитела. Через 13 недель мышей умерщвляли. Определяли объем опухолей, а также число опухолей поджелудочной железы и численную плотность сосудов. Обработка антителом антиFGFR4 40-12 позволяет значительно уменьшить объем опухолей на 55%, тогда как антитело-свидетель не оказывает какого-либо действия (фиг. 11 В). Антитело 40-12 позволяет также уменьшить на 34% число опухолей поджелудочной железы по сравнению со свидетелем (фиг. 11 С). Такое уменьшение объема опухолей сопровождается уменьшением численной плотности сосудов как в отношении числа мелких сосудов, так и числа средних или крупных сосудов (фиг. 11D). Фиг. 12. Способность антител 40-12 и 64-12 распознавать полноразмерный внеклеточный доменFGF-R4 человека или мыши, а также внеклеточный домен FGF-R4, усеченный в области D1, определяют по методике ELISA. Клон 40-12 способен также хорошо связываться с 3 конструкциями FGF-R4, тогда как клон 64-12 плохо распознает мышиную форму FGF-R4. Фиг. 13 А-13 С. Антагонистическое действие антитела анти-FGFR4 клона 40-12 на связь FGF2/FGFR4 определяют в сравнительных испытаниях на связывание с FGF2, меченным реактивом AlexaFluor 488 нм. Клон 40-12 способен блокировать связывание с FGF2 человека (фиг. 13 А), мыши (фиг. 13 В) и крысы (фиг. 13 С) на клетках 300-19 мыши, трансфицированных посредством кДНК, кодирующей человеческие, мышиные или крысиные формы рецептора FGF-R4, причем с одинаковой эффективностью(3500, 4110 и 3940 нг/мл или 23, 27 и 26 нМ для комплексов человека, мыши или крысы соответственно). Фиг. 14 А. Карта плазмиды pXL4794, обеспечивающей экспрессию hFGFR4D1:Fc. Фиг. 14 В. Последовательность аминокислот белка hFGFR4D1:Fc, секретируемого линией HEK293,трансфицированной плазмидой pXL4794. Фиг. 15 А. Карта плазмиды pXL4796, обеспечивающей экспрессию hFGFR4D2:Fc. Фиг. 15 В. Последовательность аминокислот белка hFGFR4D2:Fc, секретируемого линией HEK293,трансфицированной плазмидой pXL4796. Фиг. 16 А. Карта плазмиды pXL4799, обеспечивающей экспрессию hFGFR4D3:Fc. Фиг. 16 В. Последовательность аминокислот белка hFGFR4D3:Fc, секретируемого линией HEK293,трансфицированной плазмидой pXL4799. Примеры Пример 1. Выяснение роли FGF-R4 в регуляции ангиогенеза. Для демонстрации роли FGF-R4 в регуляции ангиогенеза проводили испытания по ангиогенезу invitro с эндотелиальными клетками человека типа HUVEC, стимулированными некоторыми FGF. FGF2 представляет собой лиганд, активирующий значительную часть рецепторов FGF. FGF19 представляет собой специфический лиганд, активирующий FGF-R4, и FGF4 представляет собой лиганд, не активирующий FGF-R4. С этой целью получали гели, внося в каждую из лунок слайд-камеры (коллаген Biocoat Cellware,тип I, 8-луночные слайд-камеры для культивирования: Becton Dickinson 354630), 160 мкл матригеля, разведенного в соотношении 1/6 (матригель без фактора роста: Becton Dickinson 356230) в коллагене (коллаген из хвоста крысы, тип I: Becton Dickinson 354236). Гель выдерживали в течение 1 ч при 37 С для обеспечения его полимеризации. Затем клетки венозного эндотелия человека (HUVEC, номер по каталогу: С-12200 - Promocell) высевали из расчета 15103 клетка/лунка в 400 мкл среды ЕВМ (Clonetics С 3121)+2% FBS+hEGF, 10 мкг/мл. Данная методика может быть адаптирована к 96-луночным планшетам: 60 мкл на лунку 96-луночных планшетов (планшеты Biocoat collagen cellware, Becton Dickinson 354407). Матрицу готовили, смешивая 1/3 матригеля, коллаген до конечной концентрации 1 мг/мл,NaOH (0,026 коллагена в мкл), PBS 1, затем объем смеси доводили водой. Эндотелиальные клетки стимулируют посредством 1 нг/мл FGF2 (RD, 133-FB-025), или 10 нг/млFGF4 (RD, 235-F4-025), или FGF19 (RD, 969-FG-025) в течение 24 ч при 37 С в атмосфере с 5% СО 2. Через 24 ч измеряют длину сети образовавшихся микротубул посредством компьютерной системы ана- 13021580 лиза изображений (Imagenia Biocom, Куртабеф, Франция) и определяют общую длину псевдотубул в каждой из лунок. Среднее значение общей длины сети микрокапилляров рассчитывают в мкм для каждого режима, что соответствует среднему из 6 повторов. Стимуляция посредством FGF2 или FGF19 обеспечивает индукцию образования новых тубул, тогда как FGF4 не оказывает такого действия (фиг. 1 А). Полученные результаты показывают, что специфическая активация FGF-R4 обеспечивает индукцию формирования новых сосудов, и, таким образом, позволяют сделать вывод, что FGF-R4 регулирует ангиогенез in vitro. Корреляция данных in vivo с данными in vitro была получена посредством модели индукции ангиогенеза in vivo в имплантатах целлюлозы на мышах. Данная модель представляет собой адаптацию модели, описанной Andrade et al. (Microvascular Research, 1997, 54, 253-61). Животных, близкородственных белых мышей BALB/cJ, анестезировали введением внутрибрюшинно смеси "ксилазин (Rompun, 10 мг/кг)/кетамин (Imalgene 1000, 100 мг/кг)". Спину животного брили и дезинфицировали средством Hexomedine. На спине мыши создавали воздушный мешок инъекцией подкожно 5 мл стерильного воздуха. На спине животного выполняли разрез приблизительно 2 см длиной для введения имплантата из стерильной целлюлозы (диск диаметром 1 см, толщиной 2 мм, Cellspon,номер по каталогу: 0501), пропитанной 50 мкл стерильного раствора, содержащего белок или испытуемое вещество. Затем разрез зашивали и дезинфицировали средством Hexomedine. В последующие после внедрения имплантата дни мышам вводили в имплантат белок или испытуемое вещество инъекцией чрескожно (50 мкл/имплантат/день) после ингаляционной анестезии (5%-й изофлюран (Aerrane,Baxter. Через семь дней после внедрения губки мышей умерщвляли введением внутрибрюшинно летальной дозы пентобарбитала натрия (CEVA sante animale). Затем кожу разрезали на расстоянии приблизительно 1 см вокруг губки, избегая шва, для изъятия кожи и губки. Затем губку разрезали на несколько кусков и помещали в пробирки Ribolyser, содержащие 1 мл лизирующего буферного раствора (CellDeath Detection ELISA, Roche). Пробирки перемешивали последовательно 4 раза в течение 20 с в режиме 4 на измельчителе клеток (FastPrep FP 120). Затем пробирки центрифугировали в течение 10 мин при 2000 g и 20 С и супернатанты замораживали при -20 С до проведения количественного определения гемоглобина. В день анализа после размораживания пробирки снова центрифугировали и определяли концентрацию гемоглобина с реактивом Драбкина (Sigma, об./об.) на спектрофотометре при 405 нм по сравнению с эталонной серией бычьего гемоглобина (Sigma). Концентрацию гемоглобина в каждом образце выражали в мг/мл в соответствии с полиномиальной регрессией, полученной исходя из эталонной серии. Результаты выражали как среднее значение (СКО) для каждой группы. Различия между группами проверяли методом ANOVA по критерию Даннетта относительно квадратного корня значений. В данной модели FGF19 в количестве 50 нг на одно место при 5 повторных инъекциях способен индуцировать значительную колонизацию губки зрелыми сосудами, вновь образованными с такой же эффективностью, как и в случае FGF2, в количестве 5 нг на одно место. Наличие функциональных кровеносных сосудов в губке было выявлено по присутствию гемоглобина (фиг. 1 В). Полученные результаты показывают, что специфическая активация FGF-R4 обеспечивает увеличение числа функциональных кровеносных сосудов, указывающее на то, что FGF-R4 регулирует ангиогенез также in vivo. Пример 2. Описание белков FGFR, использованных в качестве иммуногенов и антигенов. Рецепторы факторов роста FGFR и, в частности, их внеклеточные домены FGF-R1, FGF-R2, FGFR3, FGF-R4 соединяли с меткой (Histag) или с доменом Fc иммуноглобулина. кДНК, кодирующая внеклеточный домен FGF-R4 человека, представляет собой белок, описанный вSwissProt FGF-R4HUMAN, положения 1-365 с мутацией L136P. Ее клонировали в экспрессионном эукариотическом векторе pXL4 614, показанном на фиг. 2, для экспрессии белка, содержащего Histag в Сконцевом положении во внеклеточном домене. Белки, называемые hFGFR4-Histag, последовательности SEQ ID NO: 40 получали транзиторной трансфекцией в линии HEK293 EBNA (Invitrogen) посредством плазмиды pXL4614 и вспомогательных плазмид pXL4544 и pXL4551, обеспечивающих экспрессию двух ферментов гликозилирования Nгликанов, т.е. -2,3-сиалилтрансферазы и -1,4-галактозилтрансферазы, как описано в WO 2008/065543. Белок hFGFR4-Histag, экспрессированный в супернатант культуры клеток HEK293 EBNA, очищали хроматографией на колонке с Ni-хелатирующей сефарозой (Amersham Biosciences, номер по каталогу: 17-0575-01) при элюировании буферным раствором имидазола и затем смешивали с буферным раствором PBS (Invitrogen, номер по каталогу: 14190-094). Анализ композиции моносахаридов и количественное определение сиаловых кислот N-гликанов, как описано Saddic et al. (2002, Methods Mol. Biol. 194:2336) и Anumula et al. (1998, Glycobiology, 8:685-694), позволили показать, что белок был в значительной степени сиалилирован (91%). Следовательно, белок имел все характеристики для того, чтобы обладать достаточными фармакокинетическими свойствами. Для сравнения белок hFGFR4-Streptag (SEQ ID NO: 69) очищали хроматографией на колонке StrepTactin Superflow (IBA, номер по каталогу: 2-0575-01) при элюировании буферным раствором дезтиобиотина и затем смешивали с буферным раствором PBS. Белок hFGFR4(D2,D3)-Histag соответствует последовательности SEQ ID NO: 42. кДНК клонировали в экспрессионной эукариотической плазмиде pXL4615, показанной на фиг. 3, и белок получали и очищали в условиях, сравнимых с условиями получения белка hFGFR4-Histag. кДНК, кодирующую белок mFGFR4-Histag (SEQ ID NO: 43), клонировали в экспрессионной эукариотической плазмиде pXL4621, показанной на фиг. 4, и белок получали и очищали в условиях, сравнимых с условиями получения белка hFGFR4-Histag. Белок hFGFR1-Fc (SEQ ID NO: 46) содержит внеклеточный домен FGF-R1 IIIc человека, присоединенный С-концевым участком к домену Fc IgG1 человека. кДНК клонировали в экспрессионной эукариотической плазмиде pXL4728, показанной на фиг. 5, и белок получали в условиях, сравнимых с условиями получения белка hFGFR4-Histag, затем очищали аффинной хроматографией на колонке с протеинG-сефарозой (Amersham Biosciences) при элюировании буферным раствором 100 мМ глицин/HCl, рН 2,7 и затем смешивали с буферным раствором PBS. Белок hFGFR2-Fc последовательности SEQ ID NO: 48 содержит внеклеточный домен FGF-R2 IIIc человека, присоединенный С-концевым участком к домену Fc IgG1 человека. кДНК клонировали в экспрессионной эукариотической плазмиде pXL4327, показанной на фиг. 6, и белок получали и затем очищали в условиях, сравнимых с условиями получения белка hFGFR1-Fc. Белок hFGFR3-Fc, представляющий собой белок, присоединяющий внеклеточный домен FGF-R3IIIc человека к домену Fc IgG1 человека, получали в системе RD (номер по каталогу: 760-FR). Белок hFGFR4-Fc, представляющий собой белок, присоединяющий внеклеточный домен FGF-R4 человека к домену Fc IgG1 человека, получали в системе RD (номер по каталогу: 685-FR). Различные субдомены внеклеточной части белка hFGFR4 присоединяли к домену Fc IgG1 человека. Субдомен D1 содержится в конструкции SABVA4794 (SEQ ID NO: 112, фиг. 14 А и 14 В). Субдомен D2 содержится в конструкции SABVA4796 (SEQ ID NO: 114, фиг. 15 А и 15 В). Субдомен D3 содержится в конструкции SABVA4799 (SEQ ID NO: 116, фиг. 16 А и 16 В). Указанные три субдомена распространяются соответственно на положения 1-179 для SABVA4794, 1-32 и 145-242 для SABVA4796 и 1-32 и 228-360 для SABVA4799 (положения, описанные в SwissProt FGF-R4 HUMAN). Данные белки получали исходя из плазмиды pXL4794 (кодирующая последовательность SEQ IDNO: 111), pXL4796 (кодирующая последовательность SEQ ID NO: 113) и pXL4799 (кодирующая последовательность SEQ ID NO: 115) в условиях, сравнимых с условиями получения белка FGFR1-Fc. Пример 3. Генерирование и посев моноклональных антител анти-FGF-R4. А - антитела, полученные иммунизацией. Моноклональные антитела получали иммунизацией иммуногеном hFGFR4-Histag пяти мышейBALB/cJ (Charles River) в возрасте от 6 до 8 недель, каждая из которых была иммунизирована общей дозой 24 мкг hFGFR4-Histag способом RIMMS, описанным Kilpatrick et al. (1997, Hybridoma, 16:381389), и по методике слияния, описанной в наборе ClonaCell-HY Hybridoma Cloning (StemCell Technologies,номер по каталогу: 03800). Через два дня после последней инъекции мышей умерщвляли и лимфатические узлы смешивали с клетками миеломы P3X63-AG8.653 (АТСС, CRL-1580) в соотношении 5:1 в присутствии полиэтиленгликоля (ClonaCell-HY, номер по каталогу: 03806). Суспензию клеток распределяли в стерильных условиях по чашкам Петри, инкубируемых при 37 С в атмосфере с 5% СО 2. Колонии, появившиеся через 12 дней инкубирования, выделяли и культивировали в среде Е (ClonaCell-HY, номер по каталогу: 03805) в 96-луночных планшетах. Первичный посев моноклональных антител, полученных иммунизацией посредством hFGFR4Histag, осуществляли методом ELISA, используя в качестве антигена-ловушки hFGFR4-Streptag. Антиген-ловушку фиксировали на чашках Immulon-4 с иммобилизованным ферментом (VWR Scientific Inc.Swedesboro, NJ). Затем добавляли супернатанты культуры гибридом и детектировали с помощью антимышиного антитела кролика IgG, конъюгированного с пероксидазой (Sigma, номер по каталогу: А 9044 разведение 1:50000). Обнаружение осуществляли с субстратом ТМВ-Н 2 О 2 (Interchim, номер по каталогу:UP664780) и измерения проводили с помощью микропланшетного фотометра при 450 нм. Из 444 испытуемых гибридом 129 были позитивными при анализе ELISA в случае антигена hFGFR4-Streptag, причем из этого числа 120 гибридом были позитивными также в случае димерного белка hFGFR4-Fc. Вторичный посев осуществляли для селекции только антител, являющихся специфическими по отношению к FGF-R4, методом ELISA, используя в качестве антигена-ловушки белок hFGFR4-Streptag,затем белки hFGFR1-Fc, hFGFR2-Fc и hFGFR3-Fc, описанные в примере 2. Антиген-ловушку фиксировали на чашках Immulon-4 с иммобилизованным ферментом (VWR Scientific Inc. Swedesboro, NJ). Затем добавляли супернатанты культуры гибридом и детектировали с помощью антимышиного антитела кролика IgG, конъюгированного с пероксидазой (Sigma, номер по каталогу: А 9044 - разведение 1:50000). Обнаружение осуществляли с субстратом ТМВ-H2O2 (Interchim, номер по каталогу: UP664780) и измерения проводили с помощью микропланшетного фотометра при 450 нм. Из 129 испытуемых гибридом, бывших позитивными при анализе ELISA в случае антигена, 84 гибридомы были позитивными в случае hFGFR4-Streptag и не имели сродства ни к hFGFR1-Fc, ни кhFGFR2-Fc, ни к hFGFR3-Fc. В зависимости от способности к росту и морфологии 39 гибридом были законсервированы. Их изотип определяли с помощью набора SEROTEC (номер по каталогу: ММТ 1); 95% относились к IgG1. Третичный посев осуществляли в испытании на пролиферацию модифицированных клеток Baf/3,индуцируемую FGF2, для характеризования ингибирования антителами анти-FGFR4. Гибридомы мыши, экспрессирующие антитела-антагонисты анти-FGFR4, клонировали предельным разведением. Исходя из культивированных клеток гибридом в фазе экспоненциального роста, кодирующую последовательность (кДНК) определяли после экстракции мРНК посредством набора Oligotex (Qiagen, номер по каталогу: 72022); получение и амплификация кДНК способом RACE-RT с набором GeneRacer, обратная транскриптаза Superscript III (Invitrogen, номер по каталогу: L1500) и инициаторы (праймеры) описаны в табл. 2, приведенной ниже; амплификация фрагментов кДНК посредством полимеразыPhusion (Finnzymes, номер по каталогу: F-530S), инициаторы (праймеры) и условия температуры описаны в табл. 2. Амплифицированные фрагменты, содержащие части, кодирующие VH (вариабельную часть тяжелой цепи НС) или VL (вариабельную часть легкой цепи LC), клонировали в векторе pGEM-T Easy компании Promega, номер по каталогу: А 137 А, вставки полученных плазмид секвенировали так, чтобы кодирующая последовательность каждого вариабельного домена была исследована в направлении 5'-3' и 3'-5' по меньшей мере для 6 плазмид, соответствующих антителам анти-FGFR440-12 и анти-FGFR4 6412. Анализ последовательностей, смежностей и выравниваний осуществляли посредством программы,доступной в Vector NTI (Invitrogen). Плазмиды, содержащие консенсусные последовательности, кодирующие вариабельные части антител анти-FGFR4, консервировали. Плазмида pXL4691 содержит последовательность, кодирующую VH последовательности SEQ ID NO: 5 антитела анти-FGFR4 40-12, и плазмида pXL4693 содержит последовательность, кодирующую VH последовательности SEQ ID NO: 19 антитела анти-FGFR4 64-12, соответственно, представленные в приведенной ниже табл. 2. Плазмида pXL4690 содержит нуклеотидную последовательность SEQ ID NO: 7, кодирующую VL последовательности SEQ ID NO: 8 антитела анти-FGFR4 40-12, и плазмида pXL4692 содержит нуклеотидную последовательность SEQ ID NO: 21, кодирующую VL последовательности SEQ ID NO: 22 антитела анти-FGFR4 64-12, соответственно, представленные в приведенной ниже табл. 2. Таблица 2 Рабочие условия и анализ реакций обратной транскриптазы и PCR-идентификация плазмид pXL4 690-pXL4693 Последовательности аминокислот соответственно легких и тяжелых вариабельных частей антител анти-FGFR4 64-12 и анти-FGFR4 40-12 различны. Номера использованных, полученных и расшифрованных последовательностей указаны в табл. 7. Антитела 40-12 и 64-12 получали во флаконах Т 500. Супернатант культуры отбирали через 7 дней. Антитела анти-FGFR4 очищали по сродству на белке G и затем диализом против фильтровального стерильного PBS и хранили при 4 С. Очищенные антитела-антагонисты имели значения KD, равные 6,510-9 М (анти-FGFR4 40-12) и 5,7510-8 М (анти-FGFR4 64-12). В - антитела, отобранные способом фагового дисплея. Первичный посев моноклональных антител, полученных способом фагового дисплея с hFGFR4Histag, осуществляли методом ELISA, используя в качестве антигена-ловушки hFGFR4-Histag. Антигенловушку фиксировали на чашках Immulon-2 с иммобилизованным ферментом (VWR Scientific Inc.Swedesboro, NJ). Затем добавляли супернатанты культуры Е. coli, инфицированной фагами, и далее детектировали с помощью антитела мыши антиМ 13, конъюгированного с пероксидазой (GE Healthcare,номер по каталогу: 27-9421-01, разведение 1:5000). Обнаружение осуществляли с субстратом ТМВ-Н 2 О 2(Interchim, номер по каталогу: UP664780), и измерения проводили по оптической плотности (O.D.) при 450 нм. Вторичный посев осуществляли для селекции только антител, являющихся специфическими по отношению к FGF-R4, методом ELISA, используя в качестве антигена-ловушки белок hFGFR4-Histag, затем белки hFGFR1-Fc, hFGFR2-Fc и hFGFR3-Fc, описанные в примере 2. Антиген-ловушку фиксировали на чашках Immulon-2 с иммобилизованным ферментом (VWR Scientific Inc. Swedesboro, NJ). Затем добавляли супернатанты культуры Е. coli, инфицированной фагами, и далее детектировали с помощью антитела мыши антиМ 13, конъюгированного с пероксидазой (GE Healthcare, номер по каталогу: 27-9421- 17021580 01, разведение 1:5000). Обнаружение осуществляли с субстратом ТМВ-Н 2 О 2 (Interchim, номер по каталогу: UP664780) и измерения проводили по оптической плотности (O.D.) при 450 нм. Отобранные клоны, являющиеся специфическими по отношению к FGF-R4, секвенировали и реклонировали в экспрессионном векторе для транзиторной трансфекции клеток HEK293. С этой целью на первой стадии области, кодирующие Fab, то есть легкую цепь антитела, сайт связывания бактериальной рибосомы и вариабельную область тяжелой цепи антитела, извлекали из фагмиды рестрикцией и вставляли в плазмиду экспрессии IgG клеток млекопитающих в выходной части сигнальной последовательности эукариотического антитела и во входной части константной области тяжелой цепи IgG1 человека. На второй стадии область, содержащую сайт связывания бактериальной рибосомы и бактериальный сигнальный пептид тяжелой цепи, заменяли последовательностью IRES и эукариотической сигнальной последовательностью. IgG экспрессировали транзиторной трансфекцией клеток HEK293. Этот процесс подробно описан Т. Jostock et al., Journal of Immunological Methods, 289 (2004), 65-80. Пример последовательности константной области IgG1 человека, которая может быть использована в настоящем изобретении, представляет собой последовательность SEQ ID NO: 117. Третичный посев осуществляли в испытании на пролиферацию модифицированных клеток Baf/3,индуцируемую FGF2 и описанную в примере 4, для характеризования ингибирования антителами антиFGFR4, используя супернатанты культур клеток HEK293, транзиторно трансфицированных экспрессионными векторами антител. Данный посев позволил идентифицировать антитела клонов 8, 31, 33 и 36. Соответствующие последовательности описаны в табл. 7. Пример 4. Получение клоновой линии мышиных клеток BaF/3 FGF-R4-hMpI и методика клеточной пролиферации. Внеклеточную и трансмембранную часть FGF-R4 клонировали транслирующим слиянием с внутриклеточным доменом hMpI в мутантном векторе pEF6/V5-His А для получения в положении 5' метки НА перед химерным рецептором FGF-R4-hMpI. Конструкция мутантного pEF6A. Вектор pEF6/V5-His A (Invitrogen, номер по каталогу: V961-20) улучшали для интеграции полилинкера MCS (сайт множественного клонирования), связанного с меткой НА, размещенной в сигнальном пептиде IgGk pDisplay (Invitrogen, номер по каталогу: V660-20). С этой целью MCS, связанные с меткой НА и ее сигнальным пептидом, амплифицировали посредством PCR между прямым праймером последовательности SEQ ID NO: 51 и обратным праймером последовательности SEQ ID NO: 52, что позволяет вставлять сайты рестрикции KpnI в положение 5' и XbaI в положение 3'. Фрагмент PCR гидролизовали ферментами KpnI и XbaI и затем клонировали в векторе pEF6/V5-His А, раскрытом теми же ферментами. Наконец, первый сайт BamHI MCS заменяли сайтом BsrGI, гидролизуя вновь образованный вектор ферментами KpnI и SpeI и вставляя между данными сайтами праймеры последовательностей SEQ ID NO: 53 и SEQ ID NO: 54, гибридизованные между собой и содержащие ферментативный сайт BsrGI. Полученный вектор был назван pEF6mut-HA. Конструкция вектора pEF6mut-HA-FGF-R4-hMpI. Внутриклеточный домен MpI амплифицировали в векторе pEF6/V5-His TOPO (Invitrogen, номер по каталогу: К 9610-20) между прямым праймером последовательности SEQ ID NO: 55, позволяющим встраивать гидролизующий сайт SacI, и обратным праймером последовательности SEQ ID NO: 56, называемой традиционно revBGH. Затем полученные фрагменты PCR гидролизовали ферментами SacI и NotI. Внеклеточный и трансмембранный домен FGF-R4 амплифицировали посредством пары праймеровNO: 58). Данные праймеры обеспечивают вставку ферментативных сайтов BamHI в положение 5' и SacI в положение 3'. Затем полученный фрагмент PCR гидролизовали ферментами BamHI и SacI. Затем амплификации ДНК, кодирующей hMpI и FGF-R4, клонировали одновременно в вектореpEF6mut-HA, раскрытом BamHI-NotI. Полученная в результате конструкция была названа "pEF6mut-HAFGF-R4aIIIc-hMpI2". Затем эту конструкцию улучшали направленным мутагенезом для замены трансмембранного домена FGF-R4 (белковая последовательность SEQ ID NO: 59) последовательностью SEQID NO: 60. С этой целью использовали набор "QuickChange Site-directed mutagenesis" (Clontech, номер по каталогу: 200518) с прямым праймером последовательности SEQ ID NO: 61 и обратным праймером последовательности SEQ ID NO: 62. Полученная новая химерная конструкция была названа pEF6mut-HA-FGF-R4IIIcmut-hMpI2. Создание стабильной линии трансфекцией линии мышиных клеток BaF/3 посредством конструкцииFGF-R4IIIcmut-hMpI2. Конструкцию "pEF6mut-HA-FGF-R4IIIcmut-hMpI2" стабильно вставляли электропорацией в геном мышиных клеток BaF/3. Полученную линию отбирали в присутствии FGF2 при 20 нг/мл (RD, номер по каталогу: 234-FSE-025) и гепарина при 100 нг/мл (Sigma, номер по каталогу: Н 3149). При этом трансфицированная и отобранная линия имеет клональный тип. Методика клеточной пролиферации клеток линии BaF/3-FGFR4-hMpI. Клетки BaF/3-FGFR4-hMpI культивировали и сохраняли в полной среде RPMI 1640 (Invitrogen, но- 18021580(RD System, номер по каталогу: 234-FSE), и гепарином, 3 нг/мл (Sigma, номер по каталогу: Н 3149). В конце дня клетки высевали в количестве 0,4106 клетка/мл в полную среду RPMI 1640, дополненнуюFGF2, 20 нг/мл, и гепарином, 3 нг/мл. На следующий день по 50 мкл суспензии клеток BaF/3-FGFR4hMpI в полной среде RPMI 1640, дополненной FGF2, 20 нг/мл, и гепарином, 3 нг/мл, с концентрацией 0,2106 клетка/мл вносили в лунки 96-луночного планшета (Porvair, номер по каталогу: 214006) с последующим добавлением 50 мкл супернатанта гибридомы, содержащего испытуемое антитело. Затем планшеты инкубировали при 37 С в атмосфере с 5% СО 2 в течение от 24 до 30 ч. Для определения уровня клеточной пролиферации содержание АТФ количественно определяли, добавляя 100 мкл реактива CellTiter Glo Luminesent Cell Viability Assay (Promega, номер по каталогу: G7571) и измеряя люминесценцию люминометром. Учитывали клоны, давшие в данном испытании сигнал на 50% меньше сигнала в полной средеRPMI 1640, содержащей добавки FGF2, 20 нг/мл, и гепарина, 3 нг/мл. В случае, когда антитела анти-FGFR4, являющиеся специфическими по отношению к FGF-R4, инкубировали с клетками Baf/3-FGFR4-hMpI с последующим добавлением FGF2, 20 нг/мл, и гепарина, 3 нг/мл, было замечено антагонистическое действие на клеточную пролиферацию. Из 39 испытуемых гибридом 14 способно ингибировать клеточную пролиферацию, индуцируемую FGF, клеток Baf/3-FGFR4hMpI в присутствии FGF2. Анализом ELISA (фиг. 12) было доказано, что антитела-антагонисты анти-FGFR4 40-12 и 64-12 имеют сродство к белкам mFGFR4 мыши и hFGFR4(D2,D3) человека. В заключение осуществляли последний посев по методике Surface Plasmon Resonance (BIAcore 2000) для определения констант сродства антител анти-FGFR4. Взаимодействие между белком FGF-R4 и антителом анти-FGFR4, происходящее в супернатанте культуры гибридомы, исследовали после фиксации антитела анти-FGFR4 антителом анти-Fc, в свою очередь, зафиксированного на чипах СМ. Измерения кинетики осуществляли по методике Canziani et al., 2004. Anal. Biochem. 325: 301-307. Были выбраны два антитела из числа антител, имеющих константы сродства от 10-8 до 10-9 М и константы скорости диссоциации от 10-3 до 10-5 с-1. Характеристики данных антител представлены в табл. 1,приведенной ниже. В качестве стандартного способа для определения KD была использована методика Surface Plasmon Таблица 1B Кинетические параметры связывания антител анти-FGFR4 клонов 8, 31, 33 и 36,измеренные по методике Surface Plasmon Resonance (BIAcore 3000) Пример 5. Специфичность антител анти-FGFR4. А - специфичность антитела анти-FGFR440-12 по отношению к FGF-R4. Специфичность каждого антитела определяли способом ELISA по методике, описанной в примере 4. С этой целью исследовали способность каждого антитела связываться с каждым из FGF-R. Данный эксперимент ясно показывает, что антитело 40-12 распознает только FGF-R4 и, следовательно, является специфическим по отношению к FGF-R4. Антитело 64-12 способно связываться, главным образом, с FGF-R4, а также слабо связываться с FGF-R3 (фиг. 7). В - специфичность антител анти-FGFR4 клонов 8, 31, 33 и 36 по отношению к FGF-R4. Специфичность каждого антитела определяли способом ELISA по описанной ниже методике. Суспензии фагов, имеющих на своей поверхности антитела формата Fab, получали инфицированием бактерий Е. coli. Антигены-ловушки фиксировали на чашках Immulon-2 с иммобилизованным ферментом(VWR Scientific Inc. Swedesboro, NJ). Затем добавляли суспензии фагов и далее детектировали с помо- 19021580 щью антифагового антитела мыши М 13, конъюгированного с пероксидазой (GE Healthcare, номер по каталогу: 27-9421-01, разведение 1:5000). Обнаружение осуществляли с субстратом ТМВ-H2O2(Interchim, номер по каталогу: UP664780), и измерения проводили по оптической плотности (O.D.) при 450 нм. В табл. 2 кратко представлены полученные результаты. С этой целью исследовали способность каждого антитела связываться с каждым из FGF-R. Данный эксперимент ясно показывает, что антитела клонов 8, 31, 33, 36 распознают только FGF-R4 и, следовательно, являются специфическими по отношению к FGF-R4. Таблица 2 Специфичность антител анти-FGF-R4, происходящих от клонов 8, 31, 33 и 36,при определении по способу ELISA (сигнале=O.D. при 450 нм) Пример 6. Антагонистическое действие антитела 40-12 и антител, происходящих от клонов 8, 31,33, 36, на ангиогенез (in vitro). Для определения биологической активности моноклонального антитела 40-12 анти-FGFR4 в ходе ангиогенеза эндотелиальных клеток человека проводили испытания по ангиогенезу in vitro с клеткамиHUVEC, стимулированными посредством FGF2, в присутствии антитела 40-12 или антитела-свидетеля в дозах, увеличивающихся от 1 до 30 мкг/мл (фиг. 8 А и 8 В). В итоге показано, что активное моноклональное антитело-антагонист анти-FGFR4 40-12 способно ингибировать ангиогенез клеток HUVEC, индуцируемый FGF2, причем при дозе 30 мкг/мл или 200 нМ,тогда как антитело-свидетель 64-12 не оказывает какого-либо действия. Кроме того, антитело 40-12 не оказывает какого-либо собственного действия на базальный ангиогенез. Полученные результаты указывают на то, что антитело, являющееся специфическим антагонистомFGF-R4, способно ингибировать ангиогенез. Как и в случае антитела 40-12, антитела анти-FGFR4 клонов 8, 31, 33 и 36, полученных способом фагового дисплея, оценивали относительно их способности ингибировать ангиогенез эндотелиальных клеток человека типа HUVEC, индуцируемый FGF-2. Эти 4 антитела блокируют стимуляцию ангиогенеза in vitro, вызванную посредством FGF-2, причем при дозе 10 мкг/мл (фиг. 8 С и 8D) . Пример 7. Антагонистическое действие антитела 40-12 и антител, происходящих от клонов 8, 31,33, 36, на клетки гепатокарциномы человека (in vitro). Для определения противоопухолевого действия моноклонального антитела-антагониста антиFGFR4 40-12 проводили испытания на клетках гепатокарциномы человека Нер 3b, в случае которой пролиферация и приводящие к ней пути сигнализации зависят от пары лиганд-рецептор FGF19/FGF-R4. Во-первых, исследование путей сигнализации, зависящих от FGF-R4 и приводящих к пролиферации клеток Нер 3b, осуществляли способом вестерн-блоттинга. Такая клеточная сигнализация происходит посредством неосинтеза белков cFos и JunB, а также фосфорилирования Erk1/2 (Lin et al., J. Biol.Chem., 2007, 14:27277-84). С этой целью 5105 клеток высевали в чашку диаметром 35 мм в 2 мл полной среды (DMEM, 10% SVF, 2 мМ глутамина). Через 24 ч клетки помещали в условия дефицита питания на 24 ч в 1,8 мл бессывороточной среды. Затем клетки стимулировали в течение 3 ч посредством 200 мклFGF19, сконцентрированного в 10 раз, в отсутствие или в присутствии контрольного антитела антиFGFR4 или антитела 40-12. Далее среду удаляли, клетки промывали один раз холодным PBS и лизировали в чашке в течение 30 мин при 4 С посредством 75 мкл буферного раствора RIPA, дополненного ингибитором протеазы. Затем общий белковый экстракт центрифугировали в течение 10 мин при 4 С и 13000 об/мин и супернатант анализировали способом вестерн-блоттинга. Далее мембраны инкубировали в течение 2 ч при комнатной температуре в смеси TBS, 0,05% твин, 5% молока и затем добавляли первичное антитело анти-cFos (Cell Signaling Technology, номер по каталогу: 2250), анти-JunB (Cell Signaling Technology, номер по каталогу: 3746) и анти-фосфоErk1/2 (Cell Signaling Technology, номер по каталогу: 4377) с соотношением 1/1000 и инкубировали в течение ночи при 4 С и медленном перемешивании. Мембрану промывали три раза смесью TBS и 0,05% Твин и вторичное антитело, конъюгированное сHRP, инкубировали в течение 4 ч при 4 С при соотношении 1/2000 в смеси TBS, 0,05% Твин, 5% молока. Затем результаты вестерн-блоттинга выражали количественно посредством программы Chemigenius(Syngene). Интенсивность полос, полученных с различными антителами, сравнивали с интенсивностью полос, полученных с антиактиновым антителом, непосредственно связанным с HRP и используемым при соотношении 1/3000 (Santa Cruz Biotechnology, номер по каталогу: Sc-8432-HRP).FGF19 при 30 нг/мл индуцирует синтез белков JunB и cFos, a также фосфорилирование Erk1/2 в клетках Нер 3b. Неосинтез белка и фосфорилирование Erk полностью ингибируется антителом антиFGFR4 40-12 при 100 мкг/мл, тогда как контрольное антитело не оказывает какого-либо ингибирующего действия. Эффекты, проявившиеся на мембранах вестерн-блоттинга (фиг. 9 А), выражали количественно и интенсивность полос каждой мембраны отображали графически (фиг. 9 В). Во-вторых, проводили испытания в отношении собственно клеточной пролиферации. 5000 клеток высевали в лунки 96-луночного планшета в 100 мкл среды DMEM, 10% SVF, 2 мМ глутамина. Через 24 ч клетки помещали в условия дефицита питания на 24 ч в культуральную бессывороточную среду. Затем клетки Нер 3b стимулировали в течение 72 ч посредством 100 мкл бессывороточной среды, дополненнойFGF19, 10 нг/мл (продукция для собственных нужд Sanofi-Aventis RD), или 10% сыворотки, в отсутствие или в присутствии антитела-свидетеля или моноклонального антитела-антагониста анти-FGFR4 4012. Через 3 дня клеточную пролиферацию количественно определяли с помощью набора CellTiter Glo(Promega, Франция). Из данных испытаний следует, что сыворотка, а также FGF19 способны стимулировать пролиферацию Нер 3b. Антитело-антагонист анти-FGFR4 40-12 ингибирует частично при 100 мкг/мл такую пролиферацию, индуцированную сывороткой, тогда как антитело-свидетель не проявляет ингибирующей активности (фиг. 10 А). Кроме того, антитело 40-12 при 10 мкг/мл полностью блокирует пролиферацию,индуцированную FGF19 (фиг. 10 В). Антитело-свидетель не оказывает такого действия. Эти обстоятельства доказывают, что антагонист анти-FGFR4, представляющий собой объект настоящего изобретения, может быть использован в качестве противоопухолевого терапевтического средства в случае опухолей, зависящих от FGF19 или FGF-R4, и это антитело может быть особенно эффективным в лечении гепатокарцином. Для упрощения исследования действия антител анти-FGFR4 на клетки Нер 3b способ ELISA был доработан для детектирования фосфорилирования Erk1/2 с последующей стимуляцией этих клеток посредством FGF-19 (в корреляции с испытаниями, описанными в примере 8), FGF-2 или фетальной телячьей сыворотки. С этой целью 50000 клеток Нер 3b высевали в лунки 96-луночного планшета с черным прозрачным дном (COSTAR, номер по каталогу: 3603) в 100 мкл среды DMEM, 10% SVF, 2 мМ глутамина. Через 24 ч клетки помещали в условия дефицита питания на 24 ч в среду DMEM, 10% SVF, 2 мМ глутамина. Затем среду удаляли аспирацией и заменяли 100 мкл предварительно кондиционированной при 37 С дефицитной среды, содержащей FGF или SVF, а также испытуемые антитела в различных дозах. Клетки инкубировали в течение 3 ч при 37 С в атмосфере с 5% СО 2. Затем стимулирующую среду удаляли аспирацией,лунки промывали PBS при 4 С и клетки фиксировали добавлением 200 мкл 4%-го раствора PFA (параформальдегид) в PBS в течение 15 мин при комнатной температуре. PFA удаляли и клетки промывалиPBS три раза по 200 мкл. Мечение антителами для детектирования фосфо-Erk1/2 непосредственно на клетках Нер 3b начинали насыщением неспецифических сайтов 100 мкл насыщающего буферного раствора (21,25 мл PBS, 1,25 мл 10%-й неиммунной козьей сыворотки (Zymed, номер по каталогу: 50-062Z) ,75 мкл тритона Х 100) в течение 2 ч. Насыщающий буферный раствор заменяли 50 мкл первичного антитела анти-фосфо-Erk1/2 (Cell Signaling Technology, номер по каталогу: 4377), разведенного при соотношении 1/100 в смеси буферного раствора PBS, 1% BSA, 0,3% тритона Х 100. Первичное антитело инкубировали с клетками в течение ночи при 4 С. Затем промывали 3 порциями по 200 мкл PBS и обнаруживали с помощью вторичного антикроличьего антитела, конъюгированного с реактивом AlexaFluor 488(Molecular Probes, номер по каталогу: А 11008), разведенного при соотношении 1/5000 в смеси буферного раствора PBS, 1% BSA, 0,3% тритона Х 100, в течение 4 ч. Затем вторичное антитело промывали 3 порциями по 200 мкл PBS и далее в каждую лунку добавляли по 100 мкл PBS. Флюоресценцию измеряли в положении сверху микропланшетным фотометром типа Envision 2103 Multilabel Reader (Perkin Elmer),используя фильтр FITC. Такая методика позволяет подтвердить, что FGF-19 при 30 нг/мл индуцирует фосфорилированиеErk1/2 в клетках Нер 3b, и антитело 40-12 способно блокировать эту стимуляцию при дозе 3 мкг/мл (фиг. 9 С). Как FGF-2, так и сыворотка также способны индуцировать систему. В случае 2 последних веществ антитело 40-12 ингибирует эффект FGF-2 и сыворотки при дозах более высоких (30-100 мкг/мл; фиг. 9 С). Детектирование фосфо-Erk1/2 способом ELISA также позволило показать, что активные антитела анти-FGFR4 на модели ангиогенеза in vitro также способны на 70-95% блокировать фосфорилированиеErk1/2, индуцируемое FGF-19 с дозой 3 мкг/мл (фиг. 9D). Антитела по настоящему изобретению предпочтительно обладают антагонистическим действием одновременно в отношении патологического ангиогенеза, связанного с развитием опухоли, и в отношении собственно роста опухоли печени, в частности, на модели гепатокарциномы. Пример 8. Антагонистическое действие антитела 40-12 на мышиной модели рака поджелудочной железы. В данной фармакологической модели использовали самок мышей Rip1-Tag2 (Charles River Laboratory, Франция) генетического фонда C57BI/6J. Животным, начиная с 9-й недели после рождения, давали для питья воду, содержащую 5% сахарозы. Мышей обрабатывали в период с 10-й недели до 12,5 недель согласно методике вмешательства, выполняя один раз в неделю подкожно инъекцию антитела антиFGFR4 40-12 или контрольного антитела с дозой 25 мг/кг (фиг. 14 А). Данная методика утверждена Комитетом по экспериментам на животных (Animal Care and Use Committee) подразделения Sanofi-AventisRecherche. Зоотехнический комплекс авторов, уход за животными, а также методики обработки соответствуют обязательным принципам Европейской конвенции по защите позвоночных животных и эвтаназии после периода обработки или в случае тяжести опухолей и/или вторичных явлений, обязывающих исключать животных из исследования. По мере тяжести опухолей животных подвергали эвтаназии в конце эксперимента, и опухоли микротомировали, используя поджелудочную железу непосредственно после удаления. Объем опухоли в мм 3 определяли с помощью штангенциркуля по формуле [объем=0,52(ширина)2(длина)] для сопоставления с объемом сфероида. Тяжесть опухолей мышей рассчитывали по совокупности объемов опухолей каждой из мышей. Для гистохимического анализа численной плотности сосудов животных анестезировали, отбирали поджелудочную железу, фиксировали в течение ночи в средстве accustain (Sigma) и затем покрывали парафином. Для каждого образца готовили срезы толщиной 5 мкм. Эндотелиальные клетки детектировали, выдерживая срезы с трипсином (Zymed, номер по каталогу: 00-3003) при 37 С в течение 10 мин, затем с антителом мыши анти-CD31, полученным от крысы и разведенным при соотношении 1/50 (BDPharmingen). Для обнаружения областей, меченных антителом, срезы выдерживали с антикрысиным антителом, конъюгированным с биотином, в течение 30 мин, затем со стрептавидином, конъюгированным с HRP, в течение также 30 мин (Vectastain ABC kit, Vector) и, наконец, с DAB в течение 5 мин (Vector,номер по каталогу: SK4100). Затем срезы метили гематоксилином, разведенным при соотношении 1/10(Dako, S-3309). Снимки делали с помощью камеры, установленной на микроскоп (Nikon, E-800) с 200 кратным общим увеличением. Изображения анализировали с помощью компьютерной программы (Visiolab, Biocom). Число кровеносных сосудов в опухоли подсчитывали и классифицировали в зависимости от их площади поверхности: мелкие сосуды - от 5 до 20 мкм 2, средние сосуды - от 21 до 100 мкм 2 и крупные сосуды - более 101 мкм 2. Два препарата поджелудочной железы канализовали для определения численной плотности сосудов, соответствующей общему числу меченых элементов в поле зрения. В данной модели введение подкожно антитела анти-FGFR4 40-12 с дозой 25 мг/кг один раз в неделю в промежутке от десятой по двенадцатую неделю позволяет уменьшить тяжесть опухолей на 55%, т.е. значительным образом (фиг. 11 В), и с тенденцией уменьшения на 34% числа опухолей поджелудочной железы (фиг. 11 С), тогда как контрольная обработка не оказывает какого-либо действия. Ингибирование развития опухолей с помощью антитела анти-FGFR4 40-12 сопровождается значительным уменьшением на 31% общей численной плотности сосудов (фиг. 11D), соответствующим наблюдаемому уменьшению числа кровеносных сосудов в совокупности групп по размерам сосудов, меченных антителом анти-CD31(фиг. 11D). Полученные результаты in vivo ясно показывают, что антитело-антагонист анти-FGFR4 способно ингибировать увеличение числа и образование кровеносных сосудов в опухоли. Такое ингибирование васкуляризации опухолей сопровождается уменьшением числа опухолей поджелудочной железы и общего объема опухолей. Антитела по настоящему изобретению предпочтительно обладают антагонистическим действием одновременно в отношении патологического ангиогенеза и в отношении роста опухоли печени (на модели гепатокарциномы) и поджелудочной железы. Пример 9. Перекрестная реактивность антитела по отношению к FGF-R4 человека, мыши и крысы. На первой стадии проводили сравнительные испытания на связывание. С этой целью, FGF2 метили на уровне 2 свободных цистеиновых остатков реактивом AlexaFluor 488 нм, С-5 малеинимид (Molecular Probes, A10254) , согласно рекомендациям поставщика. Полученный FGF2-AF488 при 10 нг/мл способен связываться с FGF-R4 человека, экспрессируемым на поверхности трансфицированных клеток 30019 (фиг. 8). Такое связывание является специфическим, так как добавление избытка немеченого FGF2 позволяет заместить взаимодействие FGF2/FGF-R4 (фиг. 8 А). Такое же испытание проводили, добавляя увеличивающиеся дозы антитела 40-12 или антитела-свидетеля 64-12. Только антитело-антагонист 40-12 способно заменять связь FGF2-AF488/FGF-R4, причем с IC50, равной 3500 нг/мл или 23 нМ, свидетельствуя, что антагонистическое действие этого антитела, возможно, обязано его способности заменять связьFGF/FGF-R4. На второй стадии для определения приемлемости клона 40-12 для блокирования связи FGF/FGF-R4 у других видов, отличающихся от человека, проводили испытания на диссоциацию связей FGF2/FGF-R4 мыши и крысы по сравнению с человеком. С этой целью FGF2 мыши (RD, номер по каталогу: 3139-FB025) или крысы (RD, номер по каталогу: 3339-FB-025) метили аналогично FGF2 человека реактивомAlexaFluor 488. Испытания на диссоциацию осуществляли, используя линии 300-19, трансфицированные рецепторами FGF-R4 мыши или крысы. Результаты показывают, что антитело-антагонист анти-FGFR4 40-12 способно вызывать диссоциацию связи FGF2/FGF-R4 в системе мыши или крысы с такой же эффективностью, что и в системе человека. На практике IC50 равно 3500, 4110 и 3940 нг/мл или 23, 27 и 26 нМ для комплексов FGF2/FGF-R4 человека, мыши или крысы соответственно (фиг. 13 А, 13 В и 13 С соответственно). Такая способность связываться с FGF-R4 грызунов была проверена способом ELISA. Антитело 40-12 связывается также хорошо с FGF-R4 человека и мыши (фиг. 12). Полученные результаты указывают на то, что антитело анти-FGFR440-12 может быть использовано в фармакологических моделях с грызунами (по меньшей мере, в случае мышей и крыс), и полученные результаты, возможно, должны предсказывать эффективность в отношении человека. Исследования, проведенные с антителами, происходящими от клонов 8, 31, 33 и 36, также показывают, что данные антитела распознают одновременно FGF-R4 человека и FGF-R4 мыши (см. табл. 4). Таблица 4 Кинетические параметры связывания антител анти-FGFR4 клонов 8, 31, 33 и 36,измеренные по методике Surface Plasmon Resonance (BIAcore 3000) Пример 10. Определение эпитопа, распознаваемого антителами анти-FGFR4. А - определение эпитопа, распознаваемого антителом анти-FGFR4 40-12. Посев осуществляли способом ELISA для определения специфического домена FGFR4, распознаваемого антителом 40-12. Благодаря использованию в способе ELISA делетированной формы домена D1FGF-R4, было установлено, что антитело 40-12 взаимодействовало на уровне доменов D2-D3 FGF-R4(фиг. 12). Второй посев осуществляли способом ELISA, используя в качестве антигена-ловушки конструкты,содержащие домен D1 (SABVA4794), или домен D2 (SABVA4796), или домен D3 (SABVA4799) белкаhFGFR4, как описано в примере 2. Антиген-ловушку фиксировали на чашках Immulon-4 с иммобилизованным ферментом (VWR Scientific Inc. Swedesboro, NJ). Затем добавляли гибридому 40-12 и далее детектировали с помощью антимышиного антитела кролика IgG, конъюгированного с пероксидазой(Sigma, номер по каталогу: А 9044 - разведение 1:50000). Обнаружение осуществляли с субстратом ТМВН 2 О 2 (Interchim, номер по каталогу: UP664780) и измерения проводили по оптической плотности (O.D.) при 450 нм. В табл. 5 кратко представлены полученные результаты. Таблица 5 Определение связывания антитела 40-12 с различными субдоменами FGF-R4 Таким образом, антитело анти-FGFR4 распознает домен D2 внеклеточной части белка FGFR4. С другой стороны, белок FGFR4-FC, денатурированный посредством SDS, не распознается антителом 40-12 при анализе способом вестерн-блоттинга, что указывает на то, что эпитоп, выбранный в качестве мишени для антитела 40-12 в домене D2 FGFR4, имеет конформационный тип. В - определение эпитопа, распознаваемого антителами анти-FGFR4 клонов 8, 31, 33, 36. Посев осуществляли способом ELISA для определения специфического домена FGFR4, распознаваемого антителами клонов 8, 31, 33, 36, используя в качестве антигена-ловушки конструкты, содержащие как домены D2 и D3 (SEQ ID NO: 42), так и белок hFGFR4-Histag (SABVA4614, SEQ ID NO: 40). Антиген-ловушку фиксировали на чашках Immulon-4 с иммобилизованным ферментом (VWR ScientificInc. Swedesboro, NJ). Затем добавляли супернатанты культур HEK293, транзиторно трансфицированных плазмидами, обеспечивающими секрецию антител клонов 8, 31, 33, 36, и далее детектировали с помощью античеловеческого антитела кролика IgG, конъюгированного с пероксидазой (DakoCytomation, номер по каталогу: Р 0214 - разведение 1:5000). Обнаружение осуществляли с субстратом TMB-H2O2 (Interchim, номер по каталогу: UP664780), и измерения проводили по оптической плотности (O.D.) при 450 нм. В приведенной ниже табл. 6 кратко представлены полученные результаты. Таблица 6 Связывание антител, происходящих от клонов 8, 31, 33 и 36, с целым FGF-R4 или субдоменом D2-D3 (сигнал O.D. при 450 нм)
МПК / Метки
МПК: C12N 15/13, A61K 39/395, C07K 16/28, A61P 35/00, C12N 5/18
Метки: специфичного, fgf-r4, антитела, применение, рецептору
Код ссылки
<a href="https://eas.patents.su/30-21580-primenenie-antitela-specifichnogo-k-receptoru-fgf-r4.html" rel="bookmark" title="База патентов Евразийского Союза">Применение антитела, специфичного к рецептору fgf-r4</a>
Антитела к рецептору эфрина epha2 и их применение

Номер патента: 20324
Опубликовано: 30.10.2014
Авторы: Паркер Фабьенн, Стрели Мишель, Чжан Чунхой, Таварес Даниел, Блан Вероник, Чжоу Сяо-Май, Фромон Клодья, Хань Цзявэнь, Ли Минь
МПК: C07K 16/28, A61P 35/00, A61K 39/395...
Метки: применение, epha2, рецептору, антитела, эфрина
Формула / Реферат:
1. Антитело или его эпитопсвязывающий фрагмент, которое специфически связывается с рецептором эфрина EphA2 и является антагонистом указанного рецептора, отличающееся тем, что указанное антитело или его эпитопсвязывающий фрагмент содержит одну или несколько гипервариабельных областей с аминокислотными последовательностями, выбранными из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 61, 62, 63, 64,...
Моноклональные антитела человека к киназе-1, подобной рецептору активина, и их применение

Номер патента: 18453
Опубликовано: 30.08.2013
Авторы: Цзян Синь, Белуски Шелли Симс, Норт Майкл Айдан, Ху-Леуе Дана Дан, Ван Цзяньин, Карличек Шеннон Мари, Келлерманн Зирид-Аймее, Уикман Грант Рэймонд, Томсон Джеймс Артур, Бедиан Вахе, Амундсон Карин Кристина, Чжан Цзинчуань
МПК: C07K 16/40, A61K 39/395
Метки: человека, применение, активина, киназе-1, рецептору, антитела, моноклональные, подобной
Формула / Реферат:
1. Моноклональное антитело или его антигенсвязывающий участок, которые связываются с киназой-1, подобной рецептору активина (ALK-1), где антитело выбрано из группы, состоящей из(a) антитела, которое содержит аминокислотные последовательности CDR1, CDR2 и CDR3 последовательности VH-домена, имеющей любую из последовательностей SEQ ID NO: 6, 14, 18, 26, 30, 38, 46, 50, 54, 58, 62, 66, 70 или 104, или последовательность, кодируемую нуклеотидной...
Моноклональные антитела к растворимому рецептору лпнп человека, способы их получения и применения и гибридомы, продуцирующие указанные антитела

Номер патента: 7494
Опубликовано: 27.10.2006
Авторы: Йонах Начум, Бельзер Илана, Суисса Дани, Дреано Мишель, Смоларски Моше, Антонетти Франческо
МПК: C12N 5/20, A61P 31/20, C07K 16/28...
Метки: указанные, гибридомы, рецептору, продуцирующие, применения, способы, растворимому, человека, лпнп, получения, моноклональные, антитела
Формула / Реферат:
1. Моноклональное антитело, химерное антитело, полностью гуманизированное антитело, антитело к антиидиотипическому антителу или его фрагмент, которое специфически распознает и связывает растворимый рецептор ЛПНП человека и его фрагменты, которое получают путем иммунизации животных формой +291 растворимого рецептора ЛПНП человека, включающей аминокислотную последовательность от Asp +4 до Glu +291 последовательности ЛПНП человека, и которое...
Антитело к рецептору интерлейкина-1 типа 1 (il-1r1) и его применение

Номер патента: 18072
Опубликовано: 30.05.2013
Авторы: Кван Хьюминг, Витт Алисон, Эллиот Гари, Мартин Фрэнк, Хуан Хайчунь, Варнум Брайан, Везина Крис
МПК: A61K 39/395, C07K 16/00, G01N 33/53...
Метки: рецептору, il-1r1, интерлейкина-1, антитело, применение, типа
Формула / Реферат:
1. Изолированное человеческое антитело, которое специфично связывается с рецептором интерлейкина-1 типа 1 (IL-1R1), содержащее тяжелую цепь и легкую цепь, где тяжелая цепь содержит вариабельный участок, имеющий аминокислотную последовательность, представленную любой из SEQ ID NO: 10, SEQ ID NO: 14 или SEQ ID NO: 16, или содержащий их антигенсвязывающий фрагмент.2. Изолированное человеческое антитело, которое специфично связывается с рецептором...
Соединения, специфические к аденозиновому а1, а2a и а3 рецептору, и их применение

Номер патента: 7254
Опубликовано: 25.08.2006
Авторы: Кастелано Арлиндо Л., Уиттер Дэвид Дж., Маккиббен Брайан
МПК: A61P 5/46, A61K 31/519, A61P 11/06...
Метки: аденозиновому, рецептору, специфические, применение, соединения
Формула / Реферат:
1. Соединение, имеющее структуру где R1NR2 вместе образуют кольцо, имеющее структуру или R1 представляет собой Н и R2 представляет собой где, если R1NR2 вместе являются то R5 представляет собой Н, -СН2(NC5H8)(ОН)(С6Н5), -СН2ОСН3, -(СН2)2С(O)ОН, -CH2OCH2C(O)NH2, -С(О)ОН, -СН3, -СH2O(СН2)2ОН, -СН2ОСН2С(O)ОСН3 или -СН2ОСН2С(О)ОН; или если R1 представляет собой Н и R2 представляет собой то R5 представляет собой -СН2(N2C3H3), -С(О)ОН,...
Предыдущий патент: Клапанная система для отвода жидкой среды и способ управления клапанной системой
Следующий патент: Производные изохинолинона в качестве антагонистов nk3
Случайный патент: Способ изготовления огнеупорных изделий и огнеупорное изделие для футеровки металлургических тепловых агрегатов