Применение дефектного по репликации рекомбинантного аденовируса, содержащего гетерологичную нуклеиновую кислоту, кодирующую антиген cs возбудителя малярии, и белкового антигена, содержащего белок cs или его фрагмент, для лечения или профилактики малярии
Номер патента: 16648
Опубликовано: 30.06.2012
Авторы: Хеппнер Доналд, Пау Мария Грация, Гоудсмит Яап, Коэн Джозеф Д., Стюарт В.Энн, Дюбуа Патрис М.





















Формула / Реферат
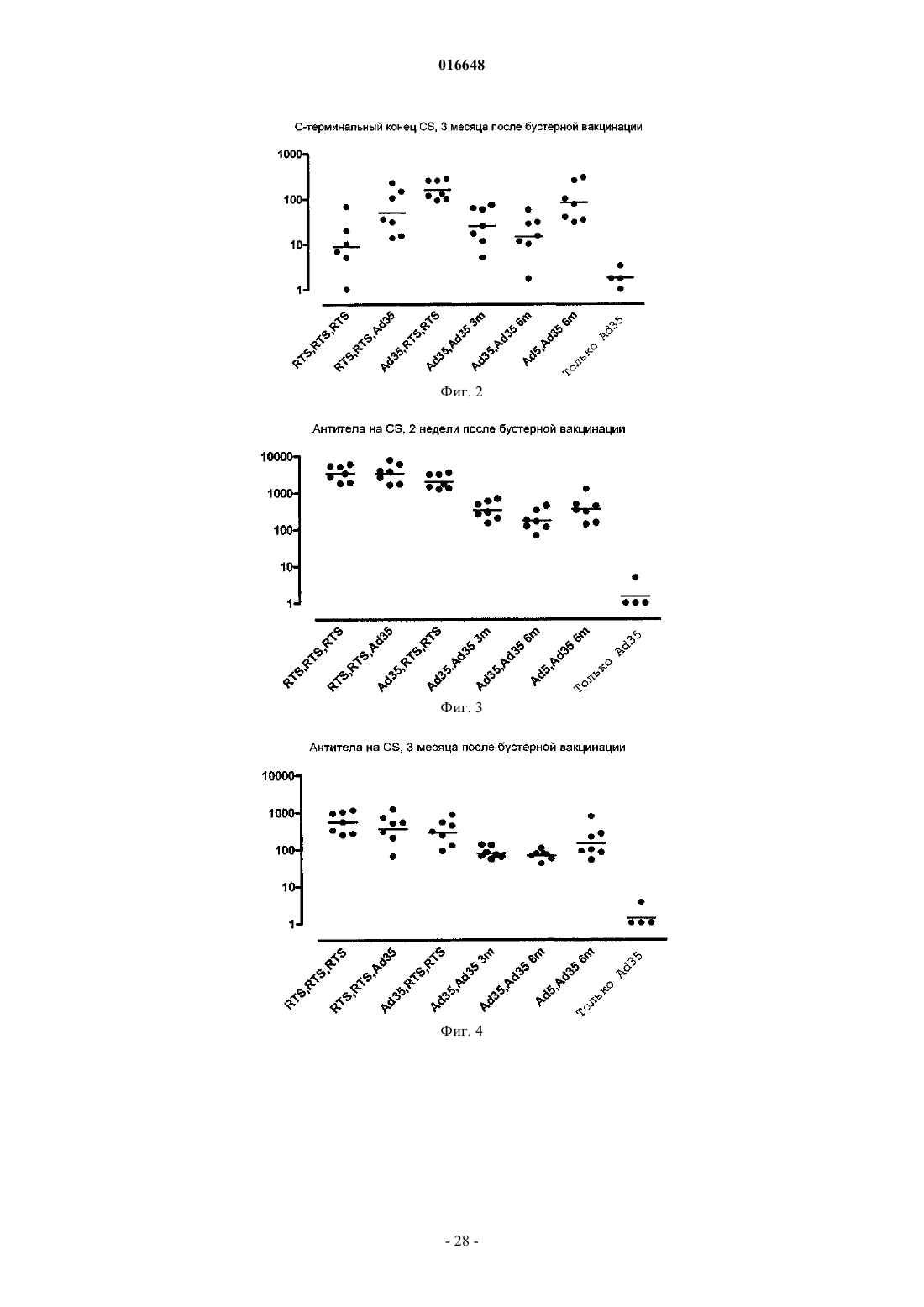
1. Применение дефектного по репликации рекомбинантного аденовируса, содержащего гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии, и смешанного с адъювантом белкового антигена, содержащего белок CS или его иммуногенный фрагмент, для производства лекарственного препарата для лечения или профилактики малярии, где указанный рекомбинантный аденовирус представляет собой аденовирус человека серотипа 11, 24, 26, 34, 35, 48, 49 или 50 и при этом используется для первичной вакцинации, а указанный смешанный с адъювантом белковый антиген используется для бустерной вакцинации, где указанный белковый антиген содержит гибридный белок, состоящий из белка CS или его иммуногенного фрагмента, слитого с поверхностным антигеном вируса гепатита B (HBsAg) в виде липопротеиновых частиц с HBsAg.
2. Применение по п.1, где указанный смешанный с адъювантом белковый антиген содержит RTS,S.
3. Способ вакцинации млекопитающего против малярийной инфекции, в котором предусмотрены этапы
проведения первичной вакцинации указанного млекопитающего дефектным по репликации рекомбинантным аденовирусом, выбранным из группы, состоящей из аденовируса человека серотипа 11, 24, 26, 34, 35, 48, 49 или 50 в фармацевтически приемлемом наполнителе, где указанный аденовирус содержит гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии; и
проведения бустерной вакцинации указанного млекопитающего смешанным с адъювантом белковым антигеном, содержащим гибридный белок, состоящий из белка CS или его иммуногенного фрагмента, слитого с поверхностным антигеном вируса гепатита B (HBsAg) в виде липопротеиновых частиц с HBsAg.
4. Способ по п.3, где указанный белковый антиген содержит RTS,S.
5. Способ по любому из пп.3, 4, где указанный белковый антиген смешан с адъювантом QS21 и 3D-MPL.
6. Способ по любому из пп.3-5, где указанный возбудитель малярии представляет собой Plasmodium falciparum.
7. Способ по любому из пп.3-6, где указанная гетерологичная нуклеиновая кислота кодон-оптимизирована для усиленной продукции кодируемого белка у млекопитающего, предпочтительно у человека.
Текст
ПРИМЕНЕНИЕ ДЕФЕКТНОГО ПО РЕПЛИКАЦИИ РЕКОМБИНАНТНОГО АДЕНОВИРУСА, СОДЕРЖАЩЕГО ГЕТЕРОЛОГИЧНУЮ НУКЛЕИНОВУЮ КИСЛОТУ,КОДИРУЮЩУЮ АНТИГЕН CS ВОЗБУДИТЕЛЯ МАЛЯРИИ, И БЕЛКОВОГО АНТИГЕНА, СОДЕРЖАЩЕГО БЕЛОК CS ИЛИ ЕГО ФРАГМЕНТ, ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ МАЛЯРИИ(71)(73) Заявитель и патентовладелец: КРУСЕЛЛ ХОЛЛАНД Б.В. (NL); ГЛАКСОСМИТКЛАЙН БАЙОЛОДЖИКАЛС,С.А. (BE); ДЗЕ ГОВЕРМЕНТ ОФ ДЗЕ ЮНАЙТЕД СТЕЙТС, ЭС РЕПРЕЗЕНТЕД БАЙ ДЗЕ СЕКРЕТАРИ ОФ ДЗЕ АРМИ,ОН БЕХАФ ОФ ДЗЕ УОЛТЕР РИД АРМИ ИНСТИТЬЮТ ОФ РИСЕРЧ (US) Настоящее изобретение относится к применению дефектного по репликации рекомбинантного аденовируса, содержащего гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии, и смешанного с адъювантом белкового антигена, содержащего белок CS или его иммуногенный фрагмент, для производства лекарственного препарата для лечения или профилактики малярии, где указанный рекомбинантный аденовирус представляет собой аденовирус человека серотипа 11, 24, 26, 34, 35, 48, 49 или 50 и при этом используется для первичной вакцинации, а указанный смешанный с адъювантом белковый антиген используется для бустерной вакцинации, где указанный белковый антиген содержит гибридный белок, состоящий из белка CS или его иммуногенного фрагмента, слитого с поверхностным антигеном вируса гепатита B (HBsAg) в виде липопротеиновых частиц с HBsAg. 016648 Область техники, к которой относится изобретение Настоящее изобретение относится к области медицины. В частности, изобретение относится к новым стратегиям использования первичных/бустерных вакцин, используя полученные рекомбинантным способом аденовирусные векторы и смешанные с адъювантом очищенные белки для профилактики тропической малярии. Уровень техники В настоящее время малярия является одной из наиболее распространенных инфекций во всех тропических и субтропических областях мира. За год в развивающихся странах и странах с формирующейся экономикой в результате малярийной инфекции умирает до 2,7 млн человек. Широкое распространение и рост заболеваемости малярией являются следствием увеличения количества резистентных к лекарственным средствам паразитов и резистентных к инсектицидам паразитарных векторов. Другие факторы включают изменения окружающей среды и климатических условий, гражданские беспорядки и возросшую мобильность популяций. Малярию вызывают переносимые комарами протозойные паразиты, принадлежащие роду Plasmodium. У человека заболевание вызывают четыре вида простейших Plasmodium (P.falciparum, P.vivax,P.ovale и P.malarxae); многие другие вызывают заболевание у животных, например P.yoelii и P.bergheii,P.falciparum является причиной большинства инфекций у людей и является причиной большинства летальных случаев. Жизненный цикл малярийных паразитов состоит из четырех отдельных стадий. Каждая из этих стадий может индуцировать специфические иммунные ответы против паразита и соответственно приводит к выработке антигенов, специфических для этой стадии, тем не менее, малярия, возникшая естественным образом, не защищает от повторной инфекции. Малярийные паразиты передаются человеку несколькими видами самок комаров Anopheles. Инфицированные комары впрыскивают спорозоитную форму малярийного паразита в кровоток млекопитающего. В течение нескольких минут до внедрения в гепатоциты спорозоиты циркулируют в крови. На этой стадии паразиты находятся во внеклеточной среде и подвергаются атаке антител, в основном направленной на белок циркумспорозоита (CS), основной компонент поверхности спорозоита. Затем в печени паразит реплицируется и развивается в шизонт. Во время этой стадии внедрение паразита сопровождается бесполым размножением и продукцией до 20000 дочерних мерозоитов на инфицированную клетку. Во время этой внутриклеточной стадии паразита главными участниками иммунного ответа хозяина являются T-лимфоциты, главным образом CD8+ T-лимфоциты (Romero et al., 1989). Примерно через одну неделю после инфицирования печени тысячи мерозоитов высвобождаются в кровоток и проникают в эритроциты (RBC), которые становятся мишенями для антитело-опосредованного иммунного ответа и цитокинов, секретированных T-клетками. После внедрения в эритроциты мерозоиты подвергаются нескольким стадиям репликации, трансформируясь в трофозоиты и шизонты. Эритроциты, разрываясь, дают новую генерацию мерозоитов, которые впоследствии инфицируют новые RBC. Эритроцитарная стадия связана с явным клиническим проявлением заболевания. Небольшое число тропозоитов может развиться в мужские или женские гаметоциты, которые представляют собой половую стадию паразита. Когда восприимчивые комары засасывают гаметоциты, оплодотворение таких гамет приводит к образованию зигот и последующей их трансформации в оокинеты, затем в ооциты и, наконец, в спорозоиты, которые мигрируют в слюнные железы для завершения цикла. Двумя основными формами патоген-специфического иммунного ответа, который возникает в ответ на внедрение паразита в организм, являются клеточный и гуморальный. С одной стороны - клеточный ответ, относящийся к CD8+ и CD4+ T-клеткам, которые принимают участие в иммунном ответе. Цитотоксические T-лимфоциты (CTL) экспрессируют CD8 и способны специфически убивать инфицированные клетки, которые экспрессируют на своей поверхности патогенные антигены. CD4+ T-клетки или Tхелперные клетки поддерживают развитие CTL, продуцируют различные цитокины, а также стимулируют индуцию B-клеток, которые делятся и продуцируют антиген-специфические антитела. Во время гуморального ответа специфические в отношении конкретного антигена B-клетки активизируются, размножаются, дифференцируются и продуцируют антиген-специфические антитела. Обе формы иммунного ответа относятся к защите против малярийной инфекции. Когда инфекционные спорозоиты мигрируют в печень и проникают в гепатоциты, спорозоиты становятся внутриклеточными патогенами, которые проводят некоторое время вне инфицированных клеток. На этой стадии особенно важны CD8+ T-клетки и CD4+ T-клетки, поскольку эти T-клетки и их продукты цитокины, такие как интерферон- (IFN-), участвуют в уничтожения инфицированных гепатоцитов хозяина. На мышиной модели малярии было обнаружено, что элиминация внутриклеточных паразитов в печени зависит от CD8+ T-клеточного ответа, направленного против пептидов, экспрессируемых паразитами на печеночной стадии (Hoffman and Doolan, 2000). Сокращение CD8+ T-клеток снимает защиту от спорозоитной стимуляции, а адаптивный перенос CD8+ T-клеток "наивным" животным, обеспечивает их защиту. Когда малярийная инфекция достигает эритроцитарной стадии, в которой мерозоиты размножаются в RBC, также обнаруживаются мерозоиты, свободно циркулирующие в кровяном потоке. Поскольку эритроцит не экспрессирует молекулы MHC ни класса I, ни класса II, которые необходимы для аутоиммунных взаимодействий с T-клетками, было сделано предположение, что на этой стадии наиболее важ-1 016648 ным является гуморальный ответ. И, наконец, наиболее эффективным подходом в создании противомалярийной вакцины была бы индуция сильного клеточного иммунного ответа такой вакциной, а также сильного гуморального иммунного ответа на различных стадиях развития паразита в организме человека. Современные подходы к созданию противомалярийной вакцины могут быть классифицированы в соответствии с различными стадиями развития паразита, как описано выше. Можно выделить три типа вакцин: преэритроцитарные вакцины, которые направлены против гепатоцитов, инфицированных спорозоитами и/или шизонтами. Исторически этот подход доминирует в стратегиях, основанных на (CS). Поскольку преэритроцитарная фаза инфекции является бессимптомной, то преэритроцитарная вакцина должна идеально индуцировать стерильный иммунитет, опосредованный гуморальным и клеточным иммунными ответами, и полностью предотвращать латентную малярийную инфекцию; вакцины, направленные на бесполую кровяную стадию, либо на инфицированный RBC, либо на сам мерозоит, созданы для минимизации клинических проявлений. Эти вакцины должны уменьшать заболеваемость и смертность, и предназначены для профилактики проникновения и/или развития паразита в эритроцитах; вакцины, блокирующие трансмиссию, которые разработаны для задержки развития паразита в комаре-хозяине. Этот тип вакцин должен способствовать уменьшению скорости распространения малярийной инфекции. Наконец, возможность разработки комбинированных малярийных вакцин, которые направлены на множество стадий жизненного цикла паразита, достигается благодаря так называемым многокомпонентным и/или многостадийным вакцинам. В настоящее время отсутствуют коммерческие вакцины против малярии, хотя поиск вакцин против малярии начался более 30 лет назад. Иммунизация грызунов, нечеловеческих приматов и людей спорозоитами, ослабленными радиацией, обеспечивала защиту против последующего заражения жизнеспособными спорозоитами (Nussenzweig et al., 1967; Clyde et al., 1973). Однако до сих пор отсутствие поддержки и подходящей крупномасштабной культуральной системы для продукции облученных спорозоитов затрудняют широкое применение таких вакцин (Luke и et al., 2003). На сегодняшний день наиболее перспективные вакцины, протестированные на людях, были основаны на небольших количествах поверхностных антигенов спорозоита. Белок CS является единственным антигеном P.falciparum, в отношении которого была продемонстрирована способность к постоянному предупреждению малярии при его использовании как основы активной иммунизации людей против передаваемой комарами инфекции, хотя и зачастую на недостаточном уровне. Теоретический анализ показал, что масштаб действия вакцины, а также эффективность вакцины должны составлять более 85%, или иначе в эндемические районы могут ускользнуть мутанты, которые являются более вирулентными (Gandon et al., 2001). Одним из способов индукции иммунного ответа у млекопитающего является введение инфекционного вектора, который содержит нуклеиновую кислоту, кодирующую в своем геноме антиген. Одним из таких носителей является рекомбинантный аденовирус, который является дефектным по репликации в результате удаления областей внутри генома, которые, как правило, значимы для репликации, например,области E1. В данной области техники известны примеры рекомбинантных аденовирусов, которые содержат гены, кодирующие антигены (WO 96/39178). Например, было показано, что ВИЧ-производные антигенные компоненты вызывают иммунный ответ при доставке рекомбинантными аденовирусами(WO 01/02607; WO 02/22080; US 6733993). В случае малярии были разработаны вакцины на основе рекомбинантных аденовирусов. Эти векторы экспрессируют полноразмерный белок CS P.yoelii, одна из мышиных моделей малярии, и было показано, что в ответ на единичную дозу иммунизации эти векторы способны вызывать стерильный иммунитет у мышей (Brua-Romero et al., 2001). Было показано, что такую защиту, индуцированную аденовирусом, главным образом, опосредуют CD8+ T-клетки. Поскольку высокий процент индивидуумов уже имеет иммунитет против обычно используемых аденовирусных векторов, таких как аденовирусный серотип 5 (Ad5), в данной области техники были разработаны новые методики, в которых рекомбинантные аденовирусы, дефектные по репликации, основаны на серотипах, которые уже вызывали иммунный ответ в виде нейтрализующих антител только у небольшого процента здоровых индивидуумов. Такие серотипы обычно называются слабо нейтрализуемыми серотипами или редкими серотипами. Было обнаружено, что Ad11, Ad24, Ad26, Ad34, Ad35, Ad48,Ad49 и Ad50 являются особенно эффективными (WO 00/70071; WO 02/40665; WO 2004/037294; WO 2004/083418; Vogels et al., 2003). Вакцины на основе ДНК, содержащие плазмиды, которые экспрессируют белок CS, P.falciparum,были разработаны Vical, Inc. San Diego, CA, США и Naval Medical Research Center (Horn et al., 1995). Исследования на мышиной модели показали индукцию антиген-специфичного CTL и гуморального ответов после иммунизации плазмидной ДНК (Doolan et al., 1998). Однако на сегодняшний день доказано, что одиночное применение ДНК-вакцин является недостаточным для индукции защитного иммунного ответа у людей. Было обнаружено, что при вакцинации ДНК-вакцины добровольцам антитела против белка CS не вырабатывались, что было определено непрямым анализом с иммунофлуоресцентными антителами(непрямой ИФА) к высушенным на воздухе спорозоитам и ELISA к рекомбинантным и синтетическим пептидам (Wang et al., 2001), хотя их CTL-ответы были значительными. Напротив, подход, основанный на малярийной вакцине с RTS,S (очищенный белок) (Gordon et al.,1995; US 6306625; WO 93/10152), способен индуцировать устойчивый гуморальный ответ на белок CS(Kester et al., 2001; Stoute et al., 1997 и 1998), хотя она, вероятно, также вызывает клеточный иммунитет типа Th1 и гуморальный иммунитет. Наиболее важным является тот факт, что такая вакцина повторно защищает приблизительно половину реципиентов. Однако защита, возникающая под действием RTS,S,имеет короткий период действия (Stoute et al., 1998). Иммунизация RTS,S индуцирует геморальный ответ на CS и CD4+ T-зависимый IFN- ответ, но слабо индуцирует CD8+ Т-зависимые CTL или IFN- ответы(Lalvani et al., 1999). Тем не менее, в клинических тестах на людях было показано, что такие минимальные продуцируемые CD8+ ответы вызывают защиту (Sun et al., 2003). Таким образом, было бы целесообразно сосредоточить попытки на усиление индукции CD8+ T-клеточного ответа на CS, вызванногоRTS,S. Остается нерешенной задача создания вакцины против тропической малярии, которая имеет эффективность защиты по меньшей мере 85%. Эта задача является особенно трудной, поскольку в отличие от других заболеваний со смертельным исходом, таких как корь или оспа, в случае малярии даже после перенесения заболевания и выработки естественного иммунитета отсутствует защита против повторной малярийной инфекции. Из всех вероятных кандидатов и подходов для доставки вакцины, протестированных на настоящий момент, только RTS,S имеет некоторый постоянный уровень защиты. Другие протестированные кандидаты были либо неадекватно иммуногенными, либо иммуногеннными, но не обеспечивающими адекватную защиту. В настоящей заявке описаны стратегии объединения вакцинных композиций, созданные на преимуществе оптимальной серологической иммуногенности белок/адъювантного подхода наряду с прекрасной индукцией клеточного ответа, обеспечиваемого рекомбинантными аденовирусными векторами, дефектными по репликации. Краткое описание чертежей Фиг. 1 - гетерологичные режимы первичной/бустерной вакцинации, за которыми следует измерениеT-клеточного ответа на C-терминальный конец CS при помощи IFN- ELISPOT анализа. Ответ измеряли через две недели после последней бустерной вакцинации. Горизонтальные столбцы представляют среднее геометрическое; фиг. 2 - T-клеточный ответ на C-терминальный конец CS, измеренный при помощи IFN- ELISPOT анализа. Ответ измеряли через три месяца после последней бустерной вакцинации. Горизонтальные столбцы представляют среднее геометрическое; фиг. 3 - гуморальный ответ на область повтора CS, измеренный при помощи ELISA, через две недели после бустерной вакцинации. Горизонтальные столбцы представляют среднее геометрическое; фиг. 4 - гуморальный ответ на область повтора CS, измеренный при помощи ELISA, через три месяца после бустерной вакцинации. Горизонтальные столбцы представляют среднее геометрическое; фиг. 5 - T-клеточный ответ в экспериментах первичной вакцинации рекомбинантным векторомAd35-CS и бустерной вакцинации RTS,S или Ad35-CS, измеренный при помощи IFN- ELISPOT анализа через две недели (слева) или три месяца (справа). В качестве контроля использовали гомологичный режим первичной/бустерной/бустерной вакцинации RTS,S/RTS,S/RTS,S; фиг. 6 - гуморальный ответ в экспериментах первичной вакцинации рекомбинантным векторомAd35-CS и бустерной вакцинации RTS,S или Ad35-CS, измеренный при помощи ELISA анализа через две недели (слева) или три месяца (справа). В качестве контроля использовали гомологичный режим первичной/бустерной/бустерной вакцинации RTS,S/RTS,S/RTS,S; фиг. 7 - T-клеточный ответ в экспериментах бустерной вакцинации рекомбинантным векторомAd35-CS и первичной вакцинации RTS,S или Ad5-CS, измеренный при помощи IFN- ELISPOT анализа через две недели (слева) или три месяца (справа), В качестве контроля использовали гомологичный режим первичной/бустерной/бустерной вакцинации RTS,S/RTS,S/RTS,S; фиг. 8 - гуморальный ответ в экспериментах бустерной вакцинации рекомбинантным векторомAd35-CS и первичной вакцинации RTS,S или Ad5-CS, измеренный при помощи IFN- ELISPOT анализа через две недели (слева) или три месяца (справа). В качестве контроля использовали гомологичный режим первичной/бустерной/бустерной вакцинации RTS,S/RTS,S/RTS,S; фиг. 9 - T-клеточный ответ на N-терминальный конец CS, измеренный при помощи IFN- ELISPOT анализа через две недели после бустерной вакцинации. Горизонтальные линии представляют среднее геометрическое; фиг. 10 - T-клеточный ответ на N-терминальный конец CS, измеренный при помощи IFN- ELISPOT анализа через три месяца после бустерной вакцинации. Горизонтальные линии представляют среднее геометрическое; фиг. 11 - T-клеточный ответ на N-терминальный конец в экспериментах первичной вакцинации рекомбинантным вектором Ad35-CS и бустерной вакцинации RTS,S или Ad35-CS, измеренный через две недели (слева) или три месяца (справа). В качестве контроля использовали гомологичный режим первич-3 016648 ной/бустерной/бустерной вакцинации RTS,S/RTS,S/RTS,S; фиг. 12 - T-клеточный ответ на N-терминальный конец в экспериментах бустерной вакцинации рекомбинантным вектором Ad35-CS и первичной вакцинации RTS,S или Ad5-CS, измеренный через две недели (слева) или три месяца (справа). В качестве контроля использовали гомологичный режим первичной/бустерной/бустерной вакцинации RTS,S/RTS,S/RTS,S. Сущность изобретения Настоящее изобретение относится к составному набору, содержащему дефектный по репликации рекомбинантный аденовирус с подходящим наполнителем, где указанный аденовирус содержит гетерологичную нуклеиновую кислоту, кодирующую антиген циркумспорозоита (CS), возбудителя малярии; и смешанный с адъювантом белковый антиген, предпочтительно также из возбудителя малярии; при этом указанный рекомбинантный аденовирус выбран из группы, состоящей из серотипов 11, 24, 26, 34, 35, 48,49 и 50 аденовируса человека. Предпочтительный белковый антиген содержит RTS,S. Предпочтительным возбудителем малярии является Plasmodium falciparum. В другом варианте осуществления изобретение также относится к применению дефектного по репликации рекомбинантного аденовируса, содержащего гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии; и смешанный с адъювантом белковый антиген предпочтительно возбудителя малярии, такого как Plasmodium falciparum, для производства лекарственного препарата для лечения или профилактики малярии, где указанный рекомбинантный аденовирус представляет собой аденовирус обезьяны или серотип 11, 24, 26, 34, 35, 48, 49 или 50 аденовируса человека. В настоящем изобретении описаны определенные предпочтительные режимы первичной бустерной вакцинации, где предпочтительными являются режимы, использующие дефектный по репликации рекомбинантный аденовирус в качестве композиции для первичной вакцинации, а смешанный с адъювантом белковый антиген используется в качестве композиции для бустерной вакцинации. Настоящее изобретение также относится к способу вакцинации млекопитающего против малярийной инфекции, в котором предусмотрены этапы первичной вакцинации указанного млекопитающего рекомбинантным аденовирусом с нарушенной репликацией в соответствующем наполнителе, причем указанный аденовирус содержит гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии; и бустерной вакцинации указанного млекопитающего смешанным с адъювантом белковым антигеном, предпочтительно RTS,S. Подробное описание настоящего изобретения Настоящее изобретение относится к составному набору, содержащему дефектный по репликации рекомбинантный аденовирус в фармацевтически приемлемом наполнителе, причем указанный аденовирус содержит гетерологичную нуклеиновую кислоту, кодирующую антиген циркумспорозоита (CS) возбудителя малярии; и смешанный с адъювантом белковый антиген; при этом указанный рекомбинантный аденовирус выбран из группы, состоящей из серотипов 11, 24, 26, 34, 35, 48, 49 и 50 аденовируса человека. Предпочтительно указанный рекомбинантный аденовирус представляет собой серотип 35 аденовируса человека. Также предпочтительным является набор по настоящему изобретению, в котором указанный белковый антиген содержит белок CS возбудителя малярии или его иммуногенный фрагмент. Указанный белковый антиген предпочтительно содержит гибридный белок, состоящий из белка CS, или его иммуногенный фрагмент слитого с ним с поверхностного антигена вируса гепатита B (HBsAg) в форме липопротеиновых частиц с HBsAg. В дополнительном предпочтительном варианте осуществления белковый антиген содержит RTS,S. Также предпочтительно, чтобы указанный белковый антиген был смешан с адъювантами QS21 и 3D-MPL предпочтительно в композиции с липосомами, содержащими холестерин. Хотя в данной области техники известно, что у человека малярию вызывают различные паразиты,один из вариантов осуществления настоящего изобретения относится к составному набору по настоящему изобретению, в котором указанный возбудитель малярии представляет собой Plasmodium falciparum. Для надлежащего иммунного ответа предпочтительным является, чтобы указанная гетерологичная нуклеиновая кислота была кодон-оптимизированной для усиленной продукции кодируемого белка у млекопитающего, предпочтительно у человека. Рекомбинантный аденовирус может находиться в смеси с адъювантом. Тот факт, что аденовирусы обезьян могут использоваться в вакцинах или генной терапии людей,хорошо известен специалистам в данной области техники. Кроме того, было обнаружено, что другие нечеловеческие аденовирусы, например аденовирусы собаки или быка, инфицируют клетки человека invitro и, следовательно, также применимы у человека, поскольку доминирование их серотипа в человеческих образцах является низким. Таким образом, изобретение также относится к составному набору, содержащему рекомбинантный аденовирус обезьяны, собаки или быка с нарушенной репликацией в фармацевтически приемлемом наполнителе, причем указанный аденовирус содержит гетерологичную нуклеиновую кислоту, кодирующую кодон-оптимизированный антиген циркумспорозоита (CS) Plasmodiumfalciparum; и смешанный с адъювантом белковый антиген, содержащий PTS,S, при этом указанный белковый антиген смешан с адъювантами QS21 и 3D-MPL предпочтительно в лекарственной форме с липосомами, содержащими холестерин. В настоящем изобретении описано, что определенные режимы первичной бустерной вакцинации-4 016648 давали неожиданный и поразительный результат относительно иммунного ответа, если различные компоненты описанного составного набора вводили в определенном порядке. Таким образом, изобретение также относится к составному набору по настоящему изобретению, в котором указанный дефектный по репликации рекомбинантный аденовирус представляет собой композицию для первичной вакцинации, а белковый смешанный с адъювантом антиген представляет собой композицию для бустерной вакцинации. Иммунный ответ, вызываемый единичным (первичным) введением вакцины, часто не является достаточно сильным и/или устойчивым для обеспечения эффективной защиты. Повторное введение (бустерное) может значительно усилить гуморальный и клеточный ответы на антигены вакцины (например, см. Estcourt et al., 2002). Изобретение также относится к применению дефектного по репликации аденовируса, содержащего гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии, и смешанный с адъювантом белковый антиген, для производства лекарственного препарата для лечения или профилактики малярии, причем указанный рекомбинантный аденовирус представляет собой обезьяний, собачий,бычий аденовирус или серотип 11, 24, 26, 34, 35, 48, 49 или 50 аденовируса человека, где предпочтительно указанный дефектный по репликации аденовирус используется в качестве композиции для первичной вакцинации, а указанный смешанный с адъювантом белковый антиген используется в качестве композиции для бустерной вакцинации. В соответствии с одним из вариантов осуществления изобретение относится к применению по настоящему изобретению, в котором белковый антиген содержит белок CS возбудителя малярии, предпочтительно Plasmodium falciparum, или его иммуногенный фрагмент. Указанный белковый антиген предпочтительно содержит гибридный белок, состоящий из белка CS или его иммуногенного фрагмента, слитого с поверхностным антигеном вируса гепатита B (HBsAg), в виде липопротеиновых частиц с HBsAg. RTS,S представляет собой предпочтительный смешанный с адъювантом белковый антиген, при этом предпочтительный адъювант представляет собой QS21 и 3D-MPL предпочтительно в лекарственной форме с липосомами, содержащими холестерин. Для оптимальной экспрессии, за которой следует оптимальный иммунный ответ у млекопитающих,предпочтительно людей, гетерологичная нуклеиновая кислота, используемая в настоящем изобретении,является кодон-оптимизированной для усиленной продукции кодируемого белка у млекопитающего,предпочтительно человека. В другом варианте осуществления настоящее изобретение относится к способу вакцинации млекопитающего против малярийной инфекции, в котором предусмотрены этапы первичной вакцинации указанного млекопитающего рекомбинантным аденовирусом с нарушенной репликацией, находящимся в фармацевтически приемлемом наполнителе, причем указанный аденовирус содержит гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии; и бустерной вакцинации указанного млекопитающего смешанным с адъювантом белковым антигеном, содержащим гибридный белок,состоящий из белка CS или его иммуногенного фрагмента, слитого с HBsAg, в виде липопротеиновых частиц с HBsAg. Предпочтительно белковый антиген содержит RTS,S, причем предпочтительный адъювант представляет собой QS21 и 3D-MPL предпочтительно в лекарственной форме с липосомами, содержащими холестерин, где предпочтительным возбудителем малярии является Plasmodium falciparum. Предпочтительные аденовирусы, которые используются для получения рекомбинантного аденовируса и которые используются в способе по настоящему изобретению, могут быть аденовирусами человека или нечеловеческими аденовирусами, такими как аденовирусы обезьяны, собаки и быка, при этом особенно предпочтительным является использование аденовирусов, на которые не выработан иммунитет у (человека) хозяина, которому вводят рекомбинантный вирус. Очень подходящими в этом случае являются аденовирусы обезьян и некоторые серотипы аденовирусов человека, как описано в настоящем описании. Предпочтительные аденовирусы человека, которые используются для способа, применения и составного набора по настоящему изобретению, представляют собой серотипы 11, 24, 26, 34, 35, 48, 49 и 50 аденовируса человека. Изобретение также относится к способу вакцинации млекопитающего против малярийной инфекции при помощи составного набора по настоящему изобретению. Если составной набор по настоящему изобретению используется для вакцинации млекопитающего против малярийной инфекции с использованием предпочтительного режима первичной бустерной вакцинации, как описано в настоящем описании, то вслед за бустерной вакцинацией предпочтительно следует одна или несколько последующих бустерных вакцинаций. Настоящее изобретение относится к применению рекомбинантного аденовируса в качестве носителя по меньшей мере одного малярийного антигена, который используется в гетерологичной комбинации с одним из смешанных с адъювантом белков в режиме первичной/бустерной вакцинации. Обнаружено,что комбинация вирусного вектора и смешанного с адъювантом белка в гетерологичном режиме первичной/бустерной вакцинации дает эффективный иммунный ответ у приматов в единицах исходного Tклеточного ответа и продолжительности иммунного ответа. В частности, было обнаружено, что первичная вакцинация млекопитающего вирусным вектором, несущим нуклеиновую кислоту, кодирующую антиген, вслед за которой следует бустерная вакцинация либо одной, либо многократной инъекцией смешанного с адъювантом белкового антигена, дает хорошие результаты в качественных и/или количе-5 016648 ственных показателях иммунного ответа. Предпочтительные вирусные векторы представляют собой аденовирусные векторы, более предпочтительно аденовирусные векторы человека и еще более предпочтительно аденовирусные векторы человека, которые вызывают низкий уровень нейтрализующей активности у млекопитающего-хозяина, которому он введен. Наиболее предпочтительными серотипами являются аденовирусы 11, 24, 26, 34, 35, 48, 49 и 50. Согласно одному из предпочтительных вариантов осуществления белковый антиген и антиген, кодируемый вирусным вектором, представляют собой малярийный антиген, более предпочтительно белок циркумспорозоита (CS) Plasmodium falciparum или его иммуногенные производные/фрагменты. В качестве иллюстрации этой концепции полипептид, кодируемый вирусным вектором, содержит нуклеиновую кислоту, кодирующую белок CS P.falciparum, включая N-терминальную часть, область повтора центральной части и C-терминальную часть (с делецией 14 аминокислот, наиболее близко расположенных кC-терминальной части: GPI якорной последовательности), при этом белковый антиген содержит конструкцию RTS,S, в которой отсутствует N-терминальная часть. Смешанный с адъювантом белковый антиген, использующийся в любом или всех аспектах изобретения, может содержать белок CS P.falciparum или его иммуногенный фрагмент, который может находиться в виде слитого белка. Например, антиген может содержать гибридный белок, состоящий из белкаCS или его иммуногенного фрагмента, слитого с поверхностным антигеном вируса гепатита B (HBsAg),где гибридный белок может экспрессироваться прокариотическими или эукариотическими клеткамихозяевами и может принимать форму липопротеиновых частиц. Слитый белок может содержать, например, по существу, всю C-терминальную часть белка CS, четыре или более тандемных повтора иммунодоминантной области и поверхностный антиген вируса гепатита B (HBsAg). Например, гибридный белок содержит последовательность, которая содержит по меньшей мере 160 аминокислот и которая, по существу, гомологична C-терминальной части белка CS и может быть лишена концевых аминокислот Cтерминальной части белка CS, например последних 10-12 аминокислот. Гибридный белок может находиться в форме смешанных липопротеиновых частиц, например с HBsAg. В частности, таким белком является гибридный белок, описанный в WO 93/10152, обозначенный как "RTS", но в настоящем описании упоминаемый как "RTS", который может находиться в форме смешанных липопротеиновых частиц с HBsAg, в настоящем описании обозначаемых как RTS,S. Отношение гибридный белок:антиген S в этих смешанных частицах составляет, например, 1:4. Гибридный белок, обозначаемый в настоящем описании "RTS", получен используя генную последовательность белка CS P.falciparum NF54 (клон 3D7; Caspers et al., 1989) и содержит, по существу, всю область 207-395 белка CS P.falciparum NF54. Часть последовательности белка CS NF54 (3D7), которая включена в RTS, представляет собой нижеследующую последовательность, состоящую из 189 аминокислот: В частности, RTS представляет собой метиониновый остаток, кодируемый нуклеотидами 1059-1061, полученными из генной последовательности TDH3 Saccharomyces cerevisiae (нуклеотиды 1-1058 в этой рамке считывания составляют промотер собственно TDH3) (Musti et al., 1983); три аминокислоты: Met, Ala, Pro, полученные из нуклеотидной последовательности (1062-1070),созданной процедурой клонирования, которая используется для конструирования гибридного гена; отрезок из 189 аминокислот (приведенны выше, SEQ ID NO:1), кодируемый 1071-1637, соответствующий аминокислотам 207-395 белка CS линии NF54 P.falciparum (клон 3D7; Caspers et al., 1989); аминокислоту (Gly), кодируемую нуклеотидами 1638-1640, созданными процедурой клонирования,которая используется для конструирования гибридного гена; четыре аминокислоты Pro Val Thr Asn, кодируемые нуклеотидами 1641-1652, которые представляют четыре карбоксиконцевых остатка белка preS2 вируса гепатита B (Valenzuela et al., 1979); отрезок из 226 аминокислот, кодируемых нуклеотидами 653-2330, определяющий белок S вируса гепатита B (серотип adw) (Valenzuela et al., 1979).RTS может находиться в форме смешанных частиц, RTS,S, где отношение RTS:S составляет, например, 1:4. Хотя изобретение не ограничивается малярийными антигенами, далее оно будет подробно описано на примере использования вирусных векторов, кодирующих малярийный антиген в комбинации со смешанным с адъювантом белковым малярийным антигеном. Специалисты в данной области техники могут модифицировать общую методику, представленную в настоящем описании, используя различные антигенные вставки и соответствующие белковые антигены других патогенных агентов, включая паразитов,бактерий, вирусы, дрожжи и даже сами антигены, включая, без ограничений, раковые антигены (например, PSA, gp100, CEA, MUC1, Her2/neu и т.п.).-6 016648 Настоящее изобретение относится к дефектному по репликации рекомбинантному аденовирусному вектору, содержащему последовательность гетерологичной нуклеиновой кислоты, кодирующую антигенPlasmodium falciparum. В предпочтительном варианте осуществления указанный вирусный вектор представляет собой аденовирус, полученный из серотипа, выбранного из группы, состоящей из Ad11, Ad24,Ad26, Ad34, Ad35, Ad48, Ad49 и Ad50. Причина такого выбора аденовирусов человека заключается в сложности широкого использования аденовирусов в качестве вакцинных векторов вследствие регулярного инфицирования людей аденовирусом дикого типа, который вызывает легкую или бессимптомную форму заболеваний, таких как простуда. Иммунный ответ, возникающий во время такой инфекции исходным серотипом дикого типа, может негативно повлиять на эффективность серотипа рекомбинантного аденовируса при его использовании в качестве последующего рекомбинантного вакцинного вектора,например вакцины против малярии, в которой используются аденовирусы. Распространенность различных аденовирусных серотипов в мировой человеческой популяции изменяется от одной географической области к другой. Как правило, предпочтительными серотипами являются серотипы, вызывающие низкую нейтрализующую активностью у большинства людей в мире, как отмечается в некоторых сообщениях, относящихся к данной области техники. Авторы настоящего изобретения разработали новую комбинацию рекомбинантного аденовируса с очищенным белком в схеме последовательной вакцинации, названной гетерологичной первичной/бустерной, причем в схеме используются разные иммунные ответы, индуцируемые разными компонентами первичной/бустерной вакцины. Выбор рекомбинантного вектора связан с серотипами, которые вызывают нейтрализующей активностью у небольшого процента человеческой популяции, нуждающейся в вакцинации. Неожиданно было обнаружено, что комбинация антигена, кодируемого аденовирусным вектором, и смешанного с адъювантом белкового антигена обеспечивает значительное усиление иммунного ответа по сравнению с иммунным ответом, наблюдаемым при применении каждой вакцины по отдельности. Усиление иммунного статуса продемонстрировано посредством in vitro обнаружения иммунного ответа in vivo у макак-резус, как описано в настоящем описании. В другом варианте осуществления дефектный по репликации рекомбинантный аденовирус представляет собой аденовирус обезьяны, такой как аденовирус, выделенный из шимпанзе. Примерами таких подходящих аденовирусов являются C68 (также известный, как Pan 9; US 6083716) и Pan 5, 6 и 7 (WO 03/046124). В одном конкретном аспекте изобретения дефектный по репликации рекомбинантный вирусный вектор содержит последовательность нуклеиновой кислоты, кодирующую белок CS или его иммуногенную часть или фрагмент. Предпочтительно указанная последовательность гетерологичной нуклеиновой кислоты является кодон-оптимизированной для усиления экспрессии у млекопитающего, предпочтительно у человека. Оптимизация кодонов основана на требуемом содержании аминокислот, обычном использовании оптимального кодона у представляющего интерес млекопитающего и ряде аспектов, которые следует учитывать для гарантии надлежащей экспрессии. Такие аспекты могут представлять собой сплайсирование донорных или акцепторных сайтов, стоп-кодонов, Chi-сайтов, поли(A) отрезков, GC- иAT-богатых последовательностей, внутренних TATA-боксов и т.д. Способы оптимизации кодонов у млекопитающих-хозяев хорошо известны специалистам в данной области техники, и их можно найти в публикациях по молекулярной биологии. В предпочтительном варианте осуществления изобретение относится к дефектному по репликации рекомбинантному аденовирусному вектору по настоящему изобретению, в котором содержание аденина плюс тимина в указанной гетерологичной нуклеиновой кислоте по сравнению с содержанием цитозина плюс гуанина ниже 87%, предпочтительно ниже 80%, более предпочтительно ниже 59% и наиболее предпочтительно равно приблизительно 45%. В одном из вариантов осуществления изобретение предоставляет дефектный по репликации рекомбинантный аденовирусный вектор, в котором белок CS представляет собой любой белок CS, как раскрыто в WO 2004/055187, более предпочтительно белок CSP.falciparum или его иммуногенный фрагмент. Создание рекомбинантных аденовирусных векторов, содержащих гетерологичные гены, хорошо известно в данной области техники и обычно включает использование пакующей клеточной линии,адапторных конструкций и космид и делеции по меньшей мере части генома (см. также ниже пакующие системы и предпочтительные клеточные линии). Изобретение также относится к наборам, содержащим в качестве компонентов, с одной стороны,рекомбинантный аденовирусный вектор, который вызывает низкую нейтрализующую активность у хозяина, а с другой стороны, очищенный белок, причем предпочтительным является случай, когда очищенный белок находится в смеси с адъювантом. Предпочтительным адъювантом является QS21 и 3DMPL предпочтительно в лекарственной форме с липосомами, содержащими холестерин. Компоненты используются в стратегии доставки гетерологичной первичной/бустерной вакцины, в которой предпочтительным является введение первым рекомбинантного вирусного вектора в качестве первичного агента,а затем очищенного белка в качестве бустерного агента, причем бустерная вакцинация может повторяться более одного раза. Компоненты обычно находятся в фармацевтически приемлемых носителях. Фармацевтически приемлемые носители хорошо известны в данной области техники и имеют достаточно ши-7 016648 рокую область применений в терапевтических продуктах. Предпочтительно используются носители, которые эффективны для вакцин. Более предпочтительными являются вакцины, дополнительно содержащие адъювант. Адъюванты хорошо известны в данной области техники как дополнительно усиливающие иммунный ответ на используемый антиген. Изобретение также относится к использованию набора по настоящему изобретению в терапевтическом, профилактическом или диагностическом лечении малярии. Настоящее изобретение относится к способу лечения млекопитающего от малярийной инфекции или защиты млекопитающего от малярийной инфекции, причем в указанном способе (в любом порядке или одновременно) предусмотрены этапы введения рекомбинантного аденовируса, несущего антиген P.falciparum; и введение по меньшей мере одного очищенного белка P.falciparum, при этом указанный белок смешан с адъювантом. Предпочтительно рекомбинантный аденовирус выбран из группы, состоящей из Ad11, Ad24, Ad26, Ad34, Ad35, Ad48, Ad49 и Ad50, при этом предпочтительным случаем также является случай, когда рекомбинантный аденовирус содержит ген, кодирующий белок CS или его иммуногенный фрагмент. Предпочтительный очищенный белок, который используется в комбинации с рекомбинантным аденовирусом, представляет собой RTS,S, при этом предпочтительным адъювантом являетсяQS21 и 3D-MPL предпочтительно в лекарственной форме с липосомами, содержащими холестерин. Движущей силой после развития иммунного ответа являются цитокины, некоторые идентифицированные белки-мессенджеры, которые оказывают содействие клеткам иммунной системы и направляют возможное развитие иммунного ответа либо по типу Th1, либо по типу Th2. Таким образом, высокие уровни цитокинов типа Th1 способствуют индукции клеточно-опосредованного иммунного ответа на данный антиген, в то время как уровни цитокинов типа Th2 способствуют индукции гуморального иммунного ответа на указанный антиген. Интересно отметить, что разница между иммунным ответом типаTh1 и типа Th2 не является абсолютной. В действительности, у индивидуума может быть иммунный ответ, который описывается, иммунный ответ с преобладанием компонента Th1 или компонента Th2. Однако обычно семейство цитокинов рассматривается в терминах, как это описано у Mosmann и Coffman(1989) для мышиных CD4+ T-клеточных клонов. Обычно, ответ типа Th1 связан с продукцией Тлимфоцитами цитокинов INF- и IL-2. Другие цитокины, которые часто связаны непосредственно с индукцией иммунного ответа типа Th1, не продуцируются T-клетками, например IL-12. Напротив, ответ типа Th2 связан с секрецией IL-4, IL-5, IL-6, IL-10 и фактора некроза опухоли (TNF-ss). Подходящие адъюванты для использования по изобретению включают соль алюминия, например гель гидроксида алюминия (квасцы) или фосфат алюминия, но также могут представлять собой соль кальция, железа или цинка или могут представлять собой нерастворимую суспензию ацилированного тирозина или ацилированные сахара, катионные или анионные полисахариды, полифосфазены или липосомы с монтанидами. В лекарственной форме вакцин для использования по изобретению в контексте аденовирусного вектора адъювант может вводиться или нет. В случае белкового компонента комбинации композицию адъюванта можно выбирать так, чтобы индуцировать предпочтительно Th1 ответ. Однако также могут быть индуцированы другие типы ответа, включая гуморальный ответ других типов. Определенные адъюванты, использующиеся в вакцинах, являются особенно подходящими для стимуляции ответа посредством цитокинов либо типа Th1, либо типа Th2. Обычно, лучшие индикаторы баланса Th1:Th2 иммунного ответа после вакцинации или инфекции включают непосредственное измерение уровня продуцирования in vitro T-лимфоцитами цитокинов Th1 или Th2 после повторной стимуляции антигеном и/или измерение отношения IgG1:IgG2a антиген-специфического гуморального ответа. Таким образом, адъювант типа Th1 представляет собой адъювант, который стимулирует изолированныеT-клеточные популяции продуцировать высокие уровни цитокинов типа Th1 при повторной стимуляцииin vitro антигеном и индуцирует антиген-специфический иммуноглобулиновый ответ, связанный с изотипом типа Th1. Например, иммуностимуляторы типа Th1, которые могут быть включены в лекарственную форму для продукции адъювантов, подходящих для применения в настоящем изобретении, могут включать монофосфориллипид A, в частности 3-де-O-ацилированный монофосфориллипид A (3D-MPL). 3D-MPL представляет собой хорошо известный адъювант, который производится Ribi Immunochem,Montana. С точки зрения химического состава он часто используется в виде смеси 3-де-Oацилированного монофосфориллипида A либо с 4, 5 или 6 ацилированными цепями. Он может быть очищен и получен способами, описанными в GB 2122204 B, который в качестве ссылки также раскрывает получение дифосфориллипида A и его 3-O-деацилированные варианты. Описаны другие очищенные и синтетические липополисахариды (патент США 6005099, EP 0729473 B1, EP 0549074 B1). В одном из вариантов осуществления 3D-MPL находится в виде мелкодисперсной лекарственной формы, содержащей небольшие частицы, диаметр которых меньше чем 0,2 мкм, и этот способ производства раскрыт вEP 0689454. Сапонины представляют собой другой пример иммуностимуляторов Th1, которые могут использоваться в изобретении. Сапонины являются хорошо известными адъювантами. Например, Quil A (получаемый из коры хинного дерева Quillaja Saponaria Molina, произрастающего в Южной Америке) и его фракции описаны в патенте США 5057540 и EP 0362279 B1. Гемолитические сапонины QS21 и QS17(ВЭЖХ-очищенные фракции Quil A) описаны как потенциальные системные адъюванты, и способ их-8 016648 получения раскрыт в патенте США 5057540 и EP 0362279 B1. Также в этих ссылках описывается применение QS7 (негемолитическая фракция Quil-A), который действует как потенциальный адъювант для системных вакцин. Также хорошо известны комбинации QS21 и полисорбата или циклодекстрина (WO 99/10008). Мелкодисперсные адъювантные системы, содержащие фракции QuilA, такие как QS21 и QS7,описаны в WO 96/33739 и WO 96/11711. Другим примером иммуностимулятора является иммуностимуляторный олигонуклеотид, содержащий неметилированные динуклеотиды CpG. CpG является аббревиатурой мотива динуклеотида цитозингуанозин, присутствующего в ДНК. CpG известен в данной области техники как адъювант как для системного, так и мукозального способа введения (WO 96/02555, EP 0468520). Было обнаружено, что фракция ДНК бациллы Calmette-Guerin (BCG) может оказывать противораковый эффект. В дальнейших исследованиях было показано, что синтетические олигонуклеотиды, полученные из генных последовательностей BCG, способны индуцировать иммуностимуляторные эффекты (как in vitro, так и in vivo). Авторы этих исследований пришли к заключению, что определенные палиндромные последовательности, включая центральный мотив CG, обладают такой активностью. Подробный анализ показал, что мотив CG должен находиться в контексте определенной последовательности и что такие последовательности обычно встречаются в бактериальных ДНК, но являются редкими для ДНК позвоночных. Иммуностимуляторная последовательность часто представляет собой пурин, пурин, C, G, пиримидин, пиримидин; где мотив CG не является метилированным, но известно, что другие неметилированные CpG последовательности являются иммуностимуляторными и могут использоваться в настоящем изобретении. В определенных комбинациях шести нуклеотидов может присутствовать палиндромная последовательность. В одном и том же нуклеотиде могут присутствовать некоторые из таких мотивов либо в виде повторов одного мотива, либо в виде комбинации разных мотивов. Наличие одного или нескольких из таких олигонуклеотидов, содержащих иммуностимуляторные последовательности, может активировать различные иммунные поднаборы, включая естественные клетки-киллеры (которые продуцируют интерферон и имеют цитолитическую активность) и макрофаги. Показано, что другие неметилированныеCpG, содержащие последовательности, не имеющие этой консенсусной последовательности, также могут быть иммуномодуляторными. При составлении лекарственной формы вакцины CpG обычно вводят в виде свободного раствора вместе со свободным антигеном (WO 96/02555, 68) или ковалентно конъюгированным с антигеном (WO 98/16247), или в составе лекарственной формы с носителем, таким как гидроксид алюминия (поверхностный антиген гепатита). Иммуностимуляторы, такие как описано выше, могут находиться в лекарственной форме совместно с носителями, такими как, например, липосомы, эмульсии "масло-в-воде", и/или солями металлов, включая соли алюминия (такие как гидроксид алюминия). Например, 3D-MPL может находиться в лекарственной форме с гидроксидом алюминия (EP 0689454) или эмульсиями масло-в-воде (WO 95/17210);QS21 преимущественно может находиться в лекарственной форме с липосомами, содержащими холестерин (WO 96/33739), с эмульсией масло-в-воде (WO 95/17210) или с квасцами (WO 98/15287); CpG может находиться в лекарственной форме с квасцами или с другими катионными носителями. Также могут использоваться комбинации иммуностимуляторов, такие как комбинация монофосфориллипида A и производного сапонина (WO 94/00153; WO 95/17210; WO 96/33739; WO 98/56414; WO 98/05355; WO 99/12565; WO 99/11241) или комбинация QS21 и 3D-MPL, как раскрыто в WO 94/00153. В качестве альтернативы в настоящем изобретении также может быть использована комбинация CpG плюс сапонин, такой как QS21. Таким образом, подходящие адъювантные системы включают, например, комбинацию монофосфориллипида A, такого как 3D-MPL, с солью алюминия. В другом варианте осуществления объединены монофосфориллипид А и производное сапонина, например комбинация QS21 и 3DMPL, как раскрыто в WO 94/00153, или менее реактивная композиция, в которой QS21 находится внутри липосом, содержащих холестерин (DQ), как раскрыто в WO 96/33739. Другая лекарственная форма адъюванта, включающая QS21, 3D-MPL и токоферол в эмульсии "масло-в-воде", описана в WO 95/17210. В другом варианте осуществления олигонуклеотиды CpG используются отдельно или совместно с солью алюминия. Подходящий адъювант для использования в настоящем изобретении предпочтительно представляет собой адъювант, стимулирующий Th1, например адъювант, содержащий сапонин, такой как QS21, или производное монофосфориллипида A, такое как 3D-MPL, или адъювант, содержащий оба компонента необязательно вместе с липосомами, содержащими холестерин. Комбинация QS21 и 3D-MPL в лекарственной форме с липосомами, содержащими холестерин, раскрыта, например, в WO 96/33739. Преимущества настоящего изобретения многочисленны. Рекомбинантные вирусы, такие как рекомбинатные аденовирусы, могут вырабатываться в очень высоких титрах путем использования клеток, которые считаются безопасными и которые могут расти в суспензии до больших объемов, используя среду,которая не содержит никаких компонентов животного или человеческого происхождения. Также известно, что рекомбинантные аденовирусы вызывают сильный иммунный ответ против белка, кодируемого последовательностью гетерологичной нуклеиновой кислоты в аденовирусном геноме. Настоящее изобретение объединяет эти особенности путем использования вектора, содержащего ген циркумспорозоитаP.falciparum, и использования смешанного с адъювантом белка для бустерных ответов. Более того, ген-9 016648 является кодон-оптимизированным, обеспечивая уровень экспрессии, который является подходящим для того, чтобы вызвать присущий людям иммунный ответ. Настоящее изобретение относится к вакцине против малярийных инфекций с использованием аденовирусов, которые не встречаются с высокими титрами нейтрализующих антител. Более предпочтительные антивирусы для этой цели представляют собой серотипы 11 и 35 (Ad11 и Ad35, см. WO 00/70071 и WO 02/40665). Содержание нуклеиновой кислоты у патогена возбудителя малярии, такого как P.falciparum, сильно отличается от ее содержания у представляющего интерес хозяина, такого как Homo sapiens. Изобретение предоставляет кодон-оптимизированные нуклеиновые кислоты, обеспечивающие более высокие уровни экспрессии у млекопитающих, таких как человек. Использование различных сущностей для режимов первичной/бустерной вакцинации, как раскрыто в настоящем описании, предоставляет способ вакцинации, который обеспечивает надлежащий иммунный ответ, как клеточной, так и гуморальной формы ответов иммунной системы. Он включает CD8+ Tклетки, CD4+ T-клетки и антитела. Ни одна из этих вакцин при использовании по отдельности не дает устойчивого иммунного ответа, который вызывает оптимальные уровни антиген-специфических CD8+T-клеток, CD4+ T-клеток и антител. Более того, порядок, в котором вводят различные компоненты, может изменять иммунный ответ и может давать в результате различные периоды возможной защиты против будущих инфекций. Способы и наборы настоящего изобретения позволяют вызывать иммунный ответ, который воздействует на все различные стадии жизненного цикла паразита в организме человека, от свободных спорозоитов и мерозоитов до инфицированных гепатоцитов и RBC. Кроме того, изобретение обеспечивает устойчивую защиту против малярийных инфекций в течение более длительного периода времени. В предпочтительном варианте осуществления изобретение относится к применению рекомбинантных аденовирусов, которые дефектны по репликации в результате удаления в аденовирусном геноме по меньшей мере части области E1, поскольку область E1 необходима для процессов репликации, транскрипции, трансляции и упаковки вновь образованного аденовируса. Векторы с удаленной E1 обычно получают на клеточных линиях, которые комплементарны функциям удаленной E1. Такие клеточные линии и их использование для продукции рекомбинатных вирусов достаточно широко описаны и хорошо известны в данной области техники. Предпочтительно клетки PER.C6, представленные клетками, хранящимися под депозитным номером ECACC 96022940 в европейской коллекции животных клеточных культур (ECACC) в Centre for Applied Microbiology and Research (CAMR, UK) или их производные используются для того, чтобы предотвратить продуцирование аденовирусов с компетентной репликацией(rca). В другом предпочтительном варианте осуществления используются клетки, которые поддерживают рост рекомбинантных аденовирусов, отличных от аденовирусов, полученных для серотипа 5 аденовируса (Ad5). В публикациях WO 97/00326, WO 01/05945, WO 01/07571, WO 00/70071, WO 02/40665 иWO 99/55132 раскрыты способы и средства получения штаммов аденовирусов без rca для Ad5, а также другие серотипы аденовирусов, которые могут продуцироваться в клетках HER, иммортализованных E1 из Ad35, или клетках C6, которые дополнительно содержат гены Е 1 из Ad35 для обеспечения надлежащей комплементации аденовирусов B-типа. Необходимо отметить, что в опубликованных документах WO 00/03029, WO 02/24730, WO 00/70071 и WO 02/40665 Ad50 ошибочно назван Ad51. Серотип Ad51, на который ссылаются в вышеуказанных публикациях, является таким же, как серотип Ad50 в статье De Jong et al. (1999), в которой он обозначен как аденовирус группы В. Для ясности, Ad50, как используется в настоящем описании, представляет собой серотип Ad50 группы B, как упоминается у De Jong et al. (1999). Вакцины настоящего изобретения обычно используются в первичных/бустерных наборах, напримерAd/белок; белок/Ad; белок/Ad/Ad; Ad/белок/Ad; Ad/Ad/белок, Ad/белок/белок/белок, Ad/белок/вирусный вектор/белок и т.д. Можно предположить, что использование комбинации с другим видом вакцины (такой как голая ДНК или рекомбинантный вирусный вектор, отличный от аденовируса) может использоваться в сочетании с первичными/бустерными агентами настоящего изобретения. Также могут использоваться дополнительные малярийные антигены или (поли)пептиды. Последовательность "получена", как используется в настоящем описании, если нуклеиновая кислота может быть получена путем прямого клонирования из последовательностей дикого типа, полученных из вирусов дикого типа, хотя они, например, также могут быть получены при помощи ПЦР с использованием различных участков ДНК, в качестве матрицы. Это также означает, что такие последовательности могут находиться в виде дикого типа, а также в измененном виде. Другим вариантом получения такого же результата является комбинирование синтетической ДНК. Также следует отметить, что "полученная" не означает исключительно прямое клонирование ДНК дикого типа. Специалистам в данной области техники также известны возможности молекулярной биологии для получения мутантных форм определенных участков нуклеиновой кислоты. Термины "функциональная часть, ее производное и/или аналог" следует рассматривать как эквиваленты нуклеиновой последовательности, к которой они относятся. Специалистам в данной области техники известен тот факт, что определенные делеции, замены,(точечные) мутации, вставки и т.п., тем не менее, могут приводить к последовательности нуклеиновой- 10016648 кислоты, которая имеет функцию, аналогичную последовательности исходной нуклеиновой кислоты, и будут продуцировать аналогичные или даже идентичные полипептиды после трансляции. Следовательно, также является очевидным, что такие изменения, которые незначительно изменяют функциональность последовательностей нуклеиновых кислот, входят в объем настоящего изобретения. Если определенный аденовирусный вектор получен из определенного предпочтительного аденовирусного серотипа,также является очевидным, что конечный продукт может быть получен опосредованными способами,такими как прямое клонирование или синтез определенных участков геномной ДНК, используя методики, известные в данной области техники. Определенные делеции, мутации и другие изменения содержимого генома, которые не изменяют специфические аспекты изобретения, тем не менее, рассматриваются как часть изобретения. Примерами таких замен являются, например, делеции в каркасе вируса для получения возможности клонировать большие участки гетерологичных нуклеиновых кислот. Примерами таких мутаций являются, например, делеции E3 или делеции и/или замены в областях, кодирующих белкиE2 и/или E4 аденовируса. Такие изменения, применяемые в отношении каркаса аденовируса, известны в данной области техники и часто используются, поскольку длина представляет собой лимитирующий фактор для упаковки аденовируса; это является основной причиной удаления определенных частей аденовирусного генома. Другие причины изменения областей E2, E3 и/или E4 генома могут относиться к стабилизации или целостности аденовирусного вектора, как, например, описано в WO 03/104467 и WO 2004/001032. Такие применения относятся, помимо прочего, к использованию гена E4orf6 из серотипа из одной подгруппы в каркасе аденовируса из другой подгруппы для того, чтобы гарантировать совместимость активности E4orf6 с E1B-55K активностью во время репликации и упаковки в пакующей клеточной линии. Они также относятся к использованию надлежащего функционирующего промотора pIX для получения более высоких уровней экспрессии pIX и более стабильного рекомбинантного аденовирусного вектора."Дефектный по репликации", как используется в настоящем описании, означает, что вирусные векторы не реплицируются в некомплементарных клетках. В комплементарных клетках функции, необходимые для репликации, и, следовательно, продукции вирусного вектора, обеспечиваются комплементарной клеткой. Дефектные по репликации вирусные векторы настоящего изобретения не содержат все элементы, предоставляющие возможность для репликации в любой клетке-хозяине, отличной от комплементарной клетки."Гетерологичный", как используется в настоящем описании в отношении нуклеиновых кислот, означает, что последовательность нуклеиновой кислоты получена из исходного источника, отличного от варианта дикого типа вирусных векторов, в которые клонирована гетерологичная нуклеиновая кислота. Например, в случае аденовирусов гетерологичная нуклеиновая кислота, которая клонирована в аденовирусный вектор с нарушенной репликацией, не является последовательностью нуклеиновой кислоты аденовируса, а происходит из другого представляющего интерес патогенного агента."Гетерологичный", как используется в настоящем описании в отношении стратегий первичнойбустерной вакцины, означает, что два или более отдельных компонента, представленные, например, одним рекомбинантным нереплицирующимся аденовирусным вектором и одним смешанным с адъювантом белком, используемые в заданной комбинации, а не в виде одного компонента, вводятся многократно,как в настоящее время обычно происходит в данной области."Антиген", как используется в настоящем описании, означает любой антиген, полученный из источника, который вызывает иммунный ответ у хозяина, в который доставлен (введен) детерминант. Антиген может быть антигеном внешнего источника, например патогеном, паразитом или даже может представлять собой аутоантиген. Примеры антигенов Plasmodium, которые могут быть доставлены при помощи рекомбинантных вирусов с нарушенной репликацией, настоящего изобретения представляют собой белок циркумспорозоита (CS), полипептид SE36, C-терминальный полипептид поверхностного белка-1 мерозоита 19 kDa (MSP-1p19), MSP-1, MSP-1p42, антиген-1 апикального мерозоита (АМА-1),антиген 1 печеночной фазы (LSA-1), или антиген 3 печеночной фазы (LSA-3), или фрагмент любого из вышеуказанных. В предпочтительном аспекте изобретение относится к белку циркумспорозоита (CS)"Кодон-оптимизированный", как используется в настоящем описании, означает, что содержимое последовательности нуклеиновой кислоты изменено для поддержания достаточно высоких уровней экспрессии представляющего интерес белка в организме-хозяине, которому доставлен ген, кодирующий указанный белок. Достаточно высокие уровни экспрессии в этом контексте означают, что уровни белка должны быть достаточно высокими для того, чтобы вызвать иммунный ответ у хозяина для защиты от инфекции или болезни. В данной области техники известно, что некоторые вакцины вызывают у людей иммунный ответ, благодаря которому приблизительно 60% вакцинированных индивидуумов защищены от заболеваний, индуцируемых последующей стимуляцией патогеном (например, спорозоитами). Следовательно, считается, что уровни экспрессии являются достаточными, если 60% или более подвергнутых обработке индивидуумов защищены от последующих инфекций. Предполагается, что такой процент можно получить путем комбинации аденовирусных аспектов, которые могут быть применены, и выбора антигена, как раскрыто в настоящем описании. Предпочтительно защищенными являются 85% индиви- 11016648 дуумов, хотя наиболее предпочтительна защита от последующего заражения более 90% вакцинированных хозяев. Нуклеиновые кислоты, раскрытые в настоящем описании, являются кодоноптимизированными для экспрессии в организме человека. Согласно Narum et al. (2001) содержание аденина плюс тимина (A+T) в ДНК у Homo sapiens составляет приблизительно 59% по сравнению с процентом цитозина плюс гуанина (C+G). Содержание аденина плюс тимина в P.falciparum составляет примерно 80%. Содержание аденина плюс тимина в CS гене P.falciparum составляет примерно 87%. Для получения достаточной защиты считается, что необходимо улучшить уровни продукции у хозяина. Одним из способов достижения этого является настройка использования кодона, сохраняя при этом ту же самую окончательную аминокислотную последовательность, но используя последовательности кодонов, более типичные для экспрессии у млекопитающих. Для этого рекомбинантные вирусные векторы, дефектные по репликации, по изобретению имеют содержание аденина плюс тимина в гетерологичных нуклеиновых кислотах настоящего изобретения менее 87%, предпочтительно менее 80% и более предпочтительно менее или равное приблизительно 59%. Основываясь на использовании кодона у человека и содержимом аминокислотной последовательности генов CS P.falciparum и yoelii, указанное процентное содержание в кодон-оптимизированных генах было даже ниже и достигало приблизительно 45% содержания данных аминокислот, как описано в настоящем изобретении. Следовательно, если рассматриваются гены CS,предпочтительно, чтобы содержание аденина плюс тимина составляло приблизительно 45%. Необходимо отметить, что если обрабатываются другие виды, отличные от человека, которые могут иметь другую концентрацию аденина плюс тимина (меньше или больше 59%) и/или другое использование кодона, то такие гены, кодирующие белки CS настоящего изобретения, могут быть откорректированы таким образом, чтобы они соответствовали требуемому содержанию и давали увеличение до подходящих уровней экспрессии для данного конкретного хозяина. Конечно, нельзя исключить то, что небольшие изменения в содержании могут привести к слабым изменениям в уровнях экспрессии в различных географических областях мира. Также следует отметить, что слабые изменения в количестве повторов в аминокислотной последовательности белков могут изменить соответственно процентное содержание. Другие представляющие интерес антигены могут быть модифицированы аналогичным способом. Все такие откорректированные содержания представляют собой часть настоящего изобретения. Белок, обозначенный RTS,S, представляет собой слитый белок, состоящий из C-терминальной половины белка CS P.falciparum (17 из центральных 41 NANP-повторов плюс большая часть Cтерминального участка), экспрессируемого в виде слитого белка с поверхностным антигеном гепатита B. Одно из преимуществ, обеспечиваемых дефектными по репликации аденовирусными векторами,заключается в слабой патогенности родительских вирусов, и документально подтверждено отсутствие характерного заболевания, вызываемого этими векторами у любых индивидуумов, включая тех, кто находится в иммунодепрессивном состоянии. Работа с мышиной моделью малярии P.yeolii показала, что рекомбинантные аденовирусные конструкции, экспрессирующие белок CS, не только вызывают заметный клеточный иммунный ответ, но и обеспечивают хорошую защиту от инфекции. Следовательно, пытаясь улучшить интенсивность T-клеточного ответа и продолжительность иммунного ответа на CS, авторы настоящего изобретения решили объединить аденовирусный подход с подходом рекомбинантных белков в новой гетерологичной первичной-бустерной стратегии. К сожалению, мышь не является идеальной моделью для прогнозирования ответа у людей. В частности, это относится и к аденовирусу 35 (Ad35). Стандартный вектор с нарушенной репликацией представляет собой аденовирус 5 (Ad5), с которым возникли некоторые проблемы, связанные с оптимизацией производительности его вектора, вследствие широкой эндемичности этого вируса и того факта, что существенная часть глобальной популяции человека имеет уже существующий иммунитет к родительскому вирусу. Ad35 обладает потенциалом для увеличения эффективности в качестве вакцинного вектора. Пригодность CSP-несущих конструкций как Ad5, так и Ad35 позволило оценить две последовательные иммунизации гетерологичными аденовирусами с разными конструкциями особенно относительно вопроса иммунитета к CS. Дендритные клетки (ДК) представляют собой наиболее эффективные антиген-представляющие клетки в организме, и тот факт, что как Ad5, так и Ad35 нацелены на ДК человека и резуса, является одним из аспектов их биологии, делает их очень хорошими вакцинными векторами. Однако только Ad5 эффективно инфицирует мышиные ДК; Ad35 надежно инфицирует только ДК приматов. Таким образом,хотя ответы на основные вопросы эффективности, касающиеся конструкций Ad35, можно получить, используя модели небольших животных, ответы на актуальные вопросы, касающиеся иммуногенности,включая Ad35, можно получить, только используя нечеловеческих приматов. Авторы настоящего изобретения решили проверить первичные-бустерные комбинации RTS,S с аденовирусными векторами, содержащими ген CS, для определения, будут ли улучшены противомалярийные клеточный и/или гуморальный ответы по отношению к ответам, наблюдаемым только для RTS,S. Кроме того, был оптимизирован режим введения двух доз только аденовирусной вакцины. Примеры Гетерологичная первичная/бустерная вакцинация макак резус с использованием рекомбинантных аденовирусных векторов и смешанного с адъювантом очищенного белка.- 12016648 Целью эксперимента была оценка RTS,S с последующим Ad35, и Ad35 с последующим RTS,S при сравнении со стандартным режимом иммунизации тремя дозами RTS,S и стандартным режимом иммунизации двумя дозами Ad35. Второй целью была оптимизация режима, содержащего две дозы аденовирусов. Изучали серологический и клеточный иммунные ответы во время и после нескольких различных режимов введения этих конструкций в комбинации. Макак-резус (Масаса mulatta) является хорошей моделью для человеческого иммунного ответа,благодаря очень близким филогенетическим связям. В частности, аллели MHC класса II хорошо сохранились; образование некоторых общих аллелей произошло по оценкам 25 миллионов лет назад до образования видов человека и резус. Таким образом, существует аналогичное использование эпитопа при наличии антигена к Th-клеткам, что сильно увеличивает прогностическую ценность модели. Более важным является то, что модель макака-резуса, как ранее было доказано, является высокопрогностической в отношении ответа как против малярии, так и ВИЧ, другого человеческого заболевания, разработка вакцины против которого была затруднена из-за сложности иммунного ответа. Предварительные эксперименты уже выполняли на мышах с аденовирусными-CS конструкциями мышиной малярии P.yeolii, которые продемонстрировали хорошую иммуногенность и защитное действие. Однако долгая история безуспешных попыток прямой экстраполяции мышиной модели малярии на людей в вопросе разработки вакцин для малярии определяет необходимость промежуточного этапа в виде модели нечеловеческих приматов. Макак-резус представляет собой наилучший выбор из видов благодаря наличию обширной базы данных ранее полученной информации на эти вакцины у этих видов,благодаря филогенетической близости с человеком, благодаря тому, что их размер позволяет отбирать образцы крови в достаточном объеме для гарантии адекватной оценки иммунного ответа, и благодаря наличию реагентов и проб, которые для этого вида уже оптимизированы, и таким образом, не требуются дополнительные протоколы и многие годы для разработки. Кроме того, только у нечеловеческих приматов конструкции аденовируса 35 могут быть протестированы должным образом из-за невозможности эффективного внедрения этого вируса в дендритные клетки других млекопитающих. Конструкции и продуцирование рекомбинантных аденовирусов с нарушенной репликацией, содержащих ген, кодирующий CS P.falciparum (Ad5CS и Ad35CS), использованные в этих исследованиях, более подробно описаны в примерах WO 2004/055187 (клон 02-659; см. фиг. 2). Кратко, эти аденовекторы содержат гетерологичный ген, кодирующий белок CS с аминокислотной последовательностью, которая аналогична белку CS штамма NF54 клона 3D7, имеющего среди прочих N-терминальную сигнальную последовательность 27 повторов NANP, кластер из 3 повторов NANP и один отдельный повтор NANP,универсальный эпитоп (Lockyer et al., 1989; Zevering et al., 1994; Nardin et al., 2001) и делецию последних 14 аминокислот (на C-конце). Отличие от белка RTS,S заключается в том, что у RTS,S отсутствуют Nтерминальная сигнальная последовательность и большой участок области повторов, а также большая часть C-терминально расположенной якорной сигнальной последовательности GPI, которая также отсутствует в аденовирусных конструкциях. Эксперимент представлял собой рандомизированное, слепое исследование иммуногенности различных комбинаций и временных стратегий для оптимизации первичных-бустерных стратегий для конструкций Ad5 и Ad35, несущих CS с RTS,S (Ad5CS и Ad35CS) и для оптимизации только Ad5CS иAd35CS. Существовавший наилучший режим, с которым сравнивали новые стратегии, представлял собой три внутримышечные дозы по 50 мкг смешанного с адъювантом RTS,S, который вводили на 0, 1 и 3 месяц. Это была группа 1, группа положительного контроля. Все группы представлены в табл. 1A. Во всех случаях адъювант представлял собой 50 мкг 3D-MPL, 50 мкг QS21 в лекарственной форме с липосомами, содержащими холестерин, как описано в WO 96/33739. Группа 2 получала две дозы RTS,S/адъювант на 0 и 1 месяц, за которыми следовала одна дозаAd35CS на 3 месяц. Группа 3 получала одну дозу Ad35CS на 0 месяц, за которой следовали две дозыRTS, S/адъювант на 1 и 3 месяц. Группы 4, 5 и 6 получали только аденовирусные конструкции. Предыдущий эксперимент с двумя дозами конструкций аденовируса 5 при различных заболеваниях показал,что оптимальный серологический и клеточный иммунные ответы получали, если интервал между иммунизациями составлял по меньшей мере 6 месяцев. Поскольку существует необходимость в оценке конструкции Ad35 в отношении людей или нечеловеческих приматов, оптимальное время между дозами для этого вектора еще не было установлено. Таким образом, группа 4 получала две дозы Ad35CS по графику на 0, 3 месяц (для прямого контроля белковых групп), а группа 5 получала две дозы по графику на 0, 6 месяц. Для изучения вопроса, зависят ли результаты введения двух доз одной и той же аденовирусной конструкции от изменения конструкции белка CS, группу 5 сравнивали с группой 6, которая получалаAd5CS, a затем Ad35CS по графику на 0, 6 месяц. Наконец, группа 7 получала две дозы обычного (без вставки малярийного гена) Ad35 на 0 и 3 месяц и служила в качестве контрольной группы иммунизации для оценки иммуногенности. В участках инъекции выстригали шерсть, а участок метили, чтобы упростить наблюдение за реактогенностью вакцины. Дополнительно на животных воздействовали седативным средством, и участок инъекции осматривали на появление признаков уплотнения, набухания, увеличения температуры, покраснения или других отклонений от нормы на 24, 48 и 72 ч и на 7 и 14 дни после инъекции. Хотя при- 13016648 знаки системной токсичности не предполагались, животным также вводили седативное средство и осматривали в те же моменты времени, отслеживая появление лимфаденопатии, целлюлита, нагноений,артрита, потери аппетита и потери веса и изменения их гематологических и клинических показателей химии. Кровь отбирали во время инъекции, а также на 24, 48, 72 ч и на 7 и 14 день после каждой инъекции для проведения полного анализа крови (CBC) и для панели клинического химического анализа, который включал (без ограничений) определение BUN, креатинина, AST, ALT, GGT и CK. Чтобы подтвердить отсутствие выделения нерепликативного вектора, ежедневно с 0 по 10 дни для каждой аденовирусной инъекции собирали образцы фекалий и хранили при -70C для тестирования на отсутствие аденовируса. 1-3 мл сыворотки собирали во время инъекций, а также на 1, 2 и 4 неделях после каждой инъекции и, по меньшей мере, через месяц после этого для определения природы и величины гуморального ответа на CS R32 (область повтора белка CS, используемая для разработки стандартного анализа ELISA на белок CS, см. ниже) при помощи ELISA. Образцы сыворотки хранили при -70C до момента использования и образцы обрабатывали партиями ближе к концу эксперимента для минимизации внутритестовых изменений. Собранные объемы сыворотки были адекватны так, чтобы для каждой аденовирусной инъекции можно было использовать по 0,5-1,0 мл сыворотки, полученной на 0, 1, 7 и 14 день и по меньшей мере каждые 4 недели после этого для определения титра антител против аденовируса. Большие объемы (2040 мл) антикоагулированной ЭДТА или гепарином крови отбирали для урожая клеток до первой иммунизации, спустя 4 недели после второй иммунизации (если позволяли требуемые объемы), спустя 4 недели после третьей иммунизации и спустя 6 месяцев после третьей иммунизации. Мононуклеарные клетки периферийной крови (PBMC) концентрировали из этих образцов, используя стандартные способы разделения при помощи центрифугирования в градиенте плотности. Хотя количество клеток могло сильно меняться у разных животных, обычно больший объем образца соответствовал большему количеству восстановленных клеток. Из-за невозможности предсказания точного количества восстановленных клеток предпочтительно брать больший объем образца, где это возможно, для получения достаточного количества клеток для повторного анализа для обеспечения статистической достоверности. Клетки замораживали для "обеспечения возможности пакетной обработки в более позднее время и таким образом улучшали контроль качества. Клетки замораживали в аутологической сыворотке с 10% ДМСО при управляемой скорости снижения температуры и хранили в парах жидкого азота в течение последней недели перед использованием. Из более крупных животных, чьи данные CBC указывали на их высокую толерантность, отбирали дополнительные образцы для сбора клеток после первичной вакцинации, но перед бустерной вакцинацией. Поскольку было 14 обезьян (две группы), которые получали две дозы RTS,S/адъювант, и 14 обезьян,которые получали одну дозу Ad35CS до наступления 8-й недели, предполагалось отбирать пробы, по меньшей мере, половины и, таким образом, поддерживать статистическую значимость. Урожай клеток собирали не ранее чем через 4 недели после инъекции. Поскольку группы, получавшие только аденовирусные конструкции, получали только две инъекции и, таким образом, имели менее напряженный график забора крови, чем обезьяны, получавшие три инъекции, предполагалось, что промежуточный сбор урожая клеток между двумя инъекциями не вызовет никаких затруднений. Анализы клеточного иммунного ответа включали быстрые ELISPOT анализы для количественного определения клеток, продуцирующих антиген-специфический IFN-. Поточный цитометрический анализ антиген-стимулирующих клеток не смог подтвердить только данные, полученные в ELISPOT анализах,но предоставил дополнительную информацию о фенотипе антиген-специфических клеток, которые отреагировали. Таким образом, также проводили определение поднабора антиген-специфических CD8+,секретирующих IFN-, при помощи внутриклеточного окрашивания и поточной цитометрии. Дополнительные анализы, которые проводили, включали общие ELISPOT анализы для дополнительных цитокинов, внутриклеточное окрашивание для подсчета поднабора T-клеток, секретирующих дополнительные цитокины, другие анализы на основе поточной цитометрии для количественной оценки продуцирования цитокинов поднабором антиген-специфических T-клеток и количественного ОТ-ПЦР для определения корреляции с другими способами. Обезьян делили на две равные группы, отбирая по возрасту, полу, весу и географическому происхождению и затем группы подвергали рандомизации. Все виды клинического анализа и защитные конечные определения устанавливали без информации о том, к какой группе принадлежали обезьяны. Аналогично, все виды иммунологического анализа проводили без предварительной информации о группах,которым принадлежат отдельные образцы. Исключения политики "работы вслепую" касались животных,которые получали иммунизацию по графику на 0 и 6 месяцы, по сравнению с животными, которые получали иммунизацию по графику на 0, 1 и 3 месяцы; однако метод "работы вслепую" сохранялся в отношении введения конкретной инъекции. Размер группы из 7 животных на тестовую группу (и четыре в контрольной группе) является идеальным с точки зрения минимизации размера группы, при котором все еще точно детектируются различия между группами на основе предварительных данных из аналогичных, но имеющих отдаленное от- 14016648 ношение экспериментов. Геометрические средние результатов анализа ELISA сравнивали параметрически, используя стандартный анализ, такой как t-критерий Стьюдента, предполагая равные изменения и два "хвоста", иANOVA. Результаты анализов ELISPOT, выраженные в пятнах на 200000 клеток, обрабатывали аналогичным образом и также оценивали, используя непараметрический анализ, такой как критерий КрускалаУоллиса. Там, где требовалось межгрупповое сравнение, использовали t-критерий Стьюдента для необработанных или логарифмически преобразованных данных для определения различий между любыми парами групп. Перед проведением инъекции шерсть срезали и кожу очищали, протирая 70% спиртом и на коже очерчивали круг диаметром 2,5-3 см несмываемыми чернилами, чтобы упростить определение участка инъекции для последующей пальпации и оценки реактогенности. Инъекции RTS,S смешивали с адъювантом сразу перед введением. Объем последней инъекции составлял 0,5 мл и вводился через иглу размером 25-29 в мышцу передней поверхности бедра. Аденовирусные конструкции получали так, как описано в WO 2004/055187, в забуференном физиологическом растворе и также вводили в том же самом месте внутримышечно с конечным объемом 0,5 мл. Первичный биообразец представлял собой кровь для получения либо сыворотки, либо клеток. График отбора крови представлен в табл. 1B. Проводили мониторинг гематологического статуса животного; что указывало на то, способно ли конкретное животное выдержать повторное взятие образца, или на то,что запланированный график отбора крови следует сократить. Каждый раз при взятии крови проводили общий анализ крови (CBC) при помощи автоматизированного счетчика клеток крови Coulter (требующего 50 мкл некоагулированной крови). При эксплуатации и техническом обслуживании выполняли GLPподобные рекомендации производителя. Значения гематокрита, гемоглобина, среднего объема эритроцита (MCV), число эритроцитов (RBC) и процент ретикулоцитов тщательно отслеживались, чтобы гарантировать то, что животные не становятся анемичными. Венозную кровь отбирали из бедренной вены, подкожной вены ноги или головной вены, используя иглы 20-24 размера, либо шприцом, либо вакуумной трубкой. В общем случае, подкожная вена ноги или головная вена предпочтительно использовались для отбора менее 10 мл крови, а бедренную вену использовали, чтобы избежать гемолизиса и сократить общее время венопункции, если объемы отобранной крови превышали 10 мл. Мононуклеарные клетки периферической крови (PBMC) отбирали. У животных перед иммунизацией, через две недели после последней иммунизации и через три месяца после последней иммунизации. В этом протоколе PMBC выделяли стандартными способами при помощи центрифугирования в градиенте плотности и хранили в замороженном виде в 45% аутологической сыворотке (45% физиологический раствор и 10% ДМСО). Кратко, цельную кровь расслаивали в среде разделения клеток фикол-гепак Lymphoprep (Axis-Shield, Oslo, Норвегия) и центрифугировали при 650g в течение 20 мин. Слой клеток снимали и промывали два раза раствором dPBS (BioWhittaker, Walkersville, MD) при 400g в течение 15 мин. Жизнеспособные клетки подсчитывали, используя гемоцитометр Coulter ACT10. Осадки ресуспендировали в 1107/мл в 50% dPBS, 50% физиологическом растворе. В конечный 10% объем по каплям добавляли ДМСО. Клетки замораживали в аликвотах по 0,55 мл, каждая из которых содержала точно по 5 млн клеток, в изопропаноловой бане с управляемой скоростью снижения температуры в -70C морозильнике в течение ночи и до использования хранили в парах жидкого азота. После последней вакцинации T-клетки, секретирующие интерферон-гамма (INF-), в образцах крови от разных обезьян идентифицировали ферментно-опосредованным иммуноспот анализом (ELISPOT) после стимуляции целыми антигенами и пулами C- и N-терминальных специфических пептидов (как более подробно описано ниже). Результаты приведены в виде диаграмм на фиг. 1 (2 недели после последней вакцинации) и на фиг. 2 (3 месяца после последней вакцинации) и представлены в табл. 2 и 4 соответственно в виде средних значений размеров пятен, образующих единицы на миллион клеток(SFU); статистическое сравнение выполняли, используя дисперсионный анализ (ANOVA) данных, подвергнутых логарифмическому преобразованию. Сравнивали все группы. В случае выявления статистически значимых различий мог выполнятьсяpost-hoc анализ для погруппового сравнения (результаты не показаны). Для сравнения результатов ELISPOT между различными стратегиями обработки вычисляли отношение средних геометрических титров для стратегий с Ad35 в качестве первичной обработки. В этих отношениях средний геометрический титр, полученный обработкой только RTS,S (на 0, 2, 3 месяцы),использовали в качестве контрольной обработки (фиг. 5 и 6 и табл. 10). Аналогично, также вычисляли отношения для стратегий с Ad35 в качестве бустерной обработки (фиг. 7 и 8 и табл. 11). Аналогичные оценки проводили для результатов анализа ELISPOT для специфической стимуляцииT-клеток N-терминальным концом. Результаты показаны в виде диаграмм на фиг. 9 и 10 и в табл. 12 и 14 соответственно. Наконец, вычислили отношения и представили их на фиг. 11 (Ad35 первичная стратегия) и на фиг. 12 (Ad35 бустерная стратегия) и в табл. 16 и 17 соответственно.ELISPOT выполняли на размороженных PBMC в планшетах MultiScreen-IP ELISPOT, дно которых- 15016648 обрабатывали ПВДФ, используя стандартную методику. Стерильность строго соблюдалась до момента удаления клеток на 2-й день для последнего спот-окрашивания. Используемая среда: полная среда (cRPMI) была свежеприготовленной из RPMI-1640 (BioWhittaker,Walkersvile, MD) с добавлением 1:100 смеси пенициллин/стрептомицин, 1:100 L-глутамина, 1:200NaHCO3 (Sigma, St.Louis, MO), 1:100 второстепенных аминокислот, 1:100 пирувата и 1:300 2-ME (Gibco). Фетальную телячью сыворотку (FCS, HyClone, Logan, UT) из партии, ранее охарактеризованной при помощи анализа неспецифической пролиферации, для поддержания хорошего роста обезьяньих клеток при обеспечении небольшой фоновой стимуляции добавляли до 10% конечного объема для cRPMI-10, 20% для cRPMI-20 и т.п. Среду Media-Plus (M+) дополнительно дополняли антителами к CD28 и CD49D обезьян в отношении 1:500 (BD Pharmingen, San Jose, CA). Нижеследующие стимуляторы были свежеприготовленными в двух заданных конечных концентрациях в M+ без добавления сыворотки.Con A: конканавалин A (Sigma) по 2,5 мкг/мл (конечная 1,25 мкг/мл) в качестве положительного контроля для всех пробирок.RTS,S: очищенный полный белковый комплекс RTS,S антигена, подходящий для клеточной культуры (GSK), по 2 мкг/мл (конечная 1 мкг/мл).HEF: полный очищенный белок поверхностного антигена гепатита B (HBS) (компонент S дляRTS,S), также подходящий для клеточной культуры (GSK), по 23,2 мкг/мл (конечная 11,6 мкг/мл).HBS-P: пул 15-мерных пептидов HBS (GSK) по 2,5 мкг/мл каждого пептида (конечная 1,25 мкг/мл). Отрицательный контроль представляет собой M+ без дополнительных добавок. Планшеты подготавливали, как описано ниже. Планшеты покрывали 50 мкл/лунку 1:100 раствора стерильного dPBS первичного моноклонального антитела к IFN- обезьян (UcyTech 21-43-09, Utrecht,Нидерланды) и инкубировали в пластиковом контейнере при 4C в течение 5-6 ч. За 1 ч до использования покрывающие антитела удаляли и планшету блокировали cRPMI-10 при 37C в инкубаторе для клеточных культур с контролируемой влажностью в 5% CO2. Непосредственно перед использованием блокирующую среду удаляли. Оттаивание замороженных РВМС. Замороженные пробирки вращали под струей теплой воды из водопроводного крана (37-40C до начала их оттаивания) и 0,55 мл содержимого сразу переносили в 8 млRPMI-20. Клетки промывали при 350g в течение 13 мин и осадок осторожно ресуспендировали в 2,0 млcRPMI-20. Затем отбирали 40 мкл стерильной аликвоты для определения количества жизнеспособных клеток и доводили объем до необходимого для получения суспензии из единичных 2106 клеток/мл. Предварительная стимуляция: равные объемы клеточной суспензии в cRPMI-20 и стимуляторы вM+ смешивали в полипропиленовых пробирках для клеточных культур для получения конечной желаемой концентрации всех реагентов. Затем клетки помещали в инкубатор по меньшей мере на 5 ч со свободно надетыми крышками и в наклонном положении для облегчения газообмена. Конечная стимуляция: после 5-6-часовой инкубации клетки центрифугировали при 400g в течение 10 мин и сливали супернатанты. Затем клетки сразу ресуспендировали в смеси, наполовину состоящей изpH. Затем клетки перемешивали в течение непродолжительного времени и осторожно отбирали пипеткой 200 мкл (200000 клеток) и переносили в соответствующие лунки в блокированных и пустых планшетах. На всех этапах следили за тем, чтобы лунки не высохли. Затем планшеты инкубировали в течение ночи(16 ч) без какого-либо воздействия. Окрашивание: 1:100 раствора вторичных поликлональных антител к INF- обезьян (UCyTech) приготовили в dPBS с 2% FCS. Клетки и среду быстро извлекали из планшеты; лунки промывали 8 разdPBS-0,5% Твин 20 (Sigma) и заполняли 50 мкл разбавленных вторичных антител. Планшеты инкубировали на качающейся платформе в течение 3 ч при комнатной температуре в пластиковом контейнере. Планшеты снова промывали 8 раз dPBS-0,5% Твин 20 и заполняли 50 мкл/лунку раствора 1:1000 коньюгата стрептовидин-щелочная фосфатаза (Southern Biotech 7100-04, Birminham, AL). Затем планшеты инкубировали в течение 2 ч при комнатной температуре в пластиковом контейнере на качающейся платформе. Наконец, планшеты промывали 8 раз, как описано выше, затем один раз промывали дистиллированной водой и добавляли по 100 мкл/лунку хромогенного субстрата NBT-BCIP (Pierce Biotech, Rockford, IL). Окрашивание проявлялось в течение 10-20 мин, в то время как фон оставался темным. Затем планшеты промывали по меньшей мере два раза 300 мкл дистиллированной воды и сушили на воздухе в течение ночи перед считыванием. Считывание планшеты: планшету считывали ELISPOT-ридером AID ELHR01, используя AID ELISPOT Reader версии 3.1.1. Все лунки были оценены визуально и неподходящие подсчеты пятен (волокна- 16016648 или другие инородные вещества) исключали вручную. Данные сохраняли на рабочей странице программы Excel. Значения лунок с двойными или тройными повторами усредняли и это количество умножали на 5 для получения конечных необработанных данных в пятна/миллион клеток. Контроль качества: среднее количество жизнеспособных восстановленных клеток после процесса заморозка/оттаивание превышало 95%. Эксперименты повторяли, если средние значения для лунок с контрольной средой превышали 20 пятен/миллион или если значения для лунок с ConA составляли менее 500 пятен/миллион. Также жизнеспособность как CD4+, так и CD8+, определенная при помощи поточной цитометрии с освобождением красителя 7-AAD и поверхностного окрашивания, должна была превышать 90%, иначе эксперимент повторяли (данные не включены). Таблица 1A Экспериментальный режим для режима первичной/бустерной вакцинации макака-резус с использованием рекомбинантных аденовирусных векторов на основе серотипов 5 и 35,содержащих ген, кодирующей белок CS P.falciparum, и с использованием смешанного с адъювантом RTS,S в качестве белкового антигенного компонента Таблица 1B График отбора крови. Для урожаев CBC и клеток требуется цельная кровь; для химического иELISA анализа требуется сыворотка. Предполагается, что 1 мл сыворотки представляет 2 мл цельной крови. Указаны только объемы цельной крови. Области значений указывают на случаи, при которых у более крупных обезьян могли быть отобраны образцы в большем объеме. "0,5" в колонке CBC/хим. указывает только CBC, полученные в эти дни.означает, что в этой временной точке кровь отбирали не у всех обезьян В таблицах, приведенных ниже, RTS,S для простоты обозначен как RTS.- 18016648 Таблица 2 Специфический T-клеточный иммунитет на C-терминальный PfCS через две недели после бустерной вакцинации: средние значения и средние геометрические IFN- ELISPOT (в SFU/миллион клеток) и сравнение по ANOVA. Даны различные режимы первичной/бустерной вакцинации (слева) Таблица 3 Т-критерий Стьюдента. р-значения для сравнения специфического IFN- ELISPOT на C-терминальныйPfCS, как показано в табл. 2, через две недели после бустерной вакцинации (последняя вакцинация) Таблица 4 Специфический T-клеточный иммунитет посредством IFN- на C-терминальный конец через три месяца после бустерной вакцинации: средние значения и средние геометрические ELISPOT (в SFU/миллион клеток) и сравнение по ANOVA. Даны различные режимы первичной/бустерной вакцинации (слева) Таблица 5 Т-критерий Стьюдента. p-значения для сравнения ELISPOT, как показано в табл. 4,через три месяца после бустерной вакцинации (последняя вакцинация)B-клеточный иммунитет (титр антител к области повтора) через две недели после последней бустерной вакцинации: средние значения и средние геометрические титра ELISA и сравнение по ANOVA. Даны различные режимы первичной/бустерной вакцинации (слева)T-критерий Стьюдента. p-значения для сравнения антител, как показано в табл. 6,через две недели после последней бустерной вакцинации (последняя вакцинация)B-клеточный иммунитет (титр антител) на CS P.falciparum через три месяца после бустерной вакцинации: средние значения и средние геометрические ELISA (в SFU) и сравнение по ANOVA. Даны различные режимы первичной/бустерной вакцинации (слева)T-критерий Стьюдента. р-значения для сравнения антител, как показано в табл. 8,через три месяца после бустерной вакцинации (последняя вакцинация)- 21016648 Таблица 10 Отношение значений средних геометрических. T- и B-клеточные ответы. В качестве первичной вакцины использовали Ad35RTS,RTS,RTS - в качестве контроля. Таблица 11 Отношение значений средних геометрических. T- и B-клеточные ответы. В качестве бустерной вакцины использовали Ad35RTS,RTS,RTS - в качестве контроля. Таблица 12 Специфический T-клеточный иммунитет посредством IFN- на N-терминальный PfCS через две недели после последней вакцинации: средние значения и средние геометрические ELISPOT (в SFU/миллион клеток) и сравнение по ANOVA. Даны различные режимы первичной/бустерной вакцинации (слева)- 22016648 Таблица 13 Т-критерий Стьюдента. p-значения для сравнения ELISPOT, как показано в табл. 12,через две недели после последней бустерной вакцинации (последняя вакцинация) Таблица 14 Специфический T-клеточный иммунитет посредством IFN- на N-терминальный PfCS через три месяца после бустерной вакцинации: средние значения и средние геометрические ELISPOT (в SFU/миллион клеток) и сравнение по ANOVA. Даны различные режимы первичной/бустерной вакцинации (слева) Таблица 15 Т-критерий Стьюдента. p-значения для сравнения ELISPOT, как показано в табл. 14,через три месяца после бустерной вакцинации (последняя вакцинация)- 23016648 Таблица 16 Отношение средних геометрических T-клеточного ответа на N-терминальный PfCS. В качестве первичной вакцины использовали Ad35RTS,RTS,RTS - в качестве контроля. Таблица 17 Отношение средних геометрических T-клеточного ответа на N-терминальный CS. В качестве бустерной вакцины использовали Ad35- 26016648 ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Применение дефектного по репликации рекомбинантного аденовируса, содержащего гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии, и смешанного с адъювантом белкового антигена, содержащего белок CS или его иммуногенный фрагмент, для производства лекарственного препарата для лечения или профилактики малярии, где указанный рекомбинантный аденовирус представляет собой аденовирус человека серотипа 11, 24, 26, 34, 35, 48, 49 или 50 и при этом используется для первичной вакцинации, а указанный смешанный с адъювантом белковый антиген используется для бустерной вакцинации, где указанный белковый антиген содержит гибридный белок, состоящий из белка CS или его иммуногенного фрагмента, слитого с поверхностным антигеном вируса гепатита B (HBsAg) в виде липопротеиновых частиц с HBsAg. 2. Применение по п.1, где указанный смешанный с адъювантом белковый антиген содержит RTS,S. 3. Способ вакцинации млекопитающего против малярийной инфекции, в котором предусмотрены этапы проведения первичной вакцинации указанного млекопитающего дефектным по репликации рекомбинантным аденовирусом, выбранным из группы, состоящей из аденовируса человека серотипа 11, 24,26, 34, 35, 48, 49 или 50 в фармацевтически приемлемом наполнителе, где указанный аденовирус содержит гетерологичную нуклеиновую кислоту, кодирующую антиген CS возбудителя малярии; и проведения бустерной вакцинации указанного млекопитающего смешанным с адъювантом белковым антигеном, содержащим гибридный белок, состоящий из белка CS или его иммуногенного фрагмента, слитого с поверхностным антигеном вируса гепатита B (HBsAg) в виде липопротеиновых частиц сHBsAg. 4. Способ по п.3, где указанный белковый антиген содержит RTS,S. 5. Способ по любому из пп.3, 4, где указанный белковый антиген смешан с адъювантом QS21 и 3DMPL. 6. Способ по любому из пп.3-5, где указанный возбудитель малярии представляет собой Plasmodiumfalciparum. 7. Способ по любому из пп.3-6, где указанная гетерологичная нуклеиновая кислота кодоноптимизирована для усиленной продукции кодируемого белка у млекопитающего, предпочтительно у человека.
МПК / Метки
МПК: A61K 39/015
Метки: кислоту, фрагмент, возбудителя, содержащего, аденовируса, репликации, гетерологичную, профилактики, малярии, дефектного, лечения, нуклеиновую, белок, белкового, рекомбинантного, кодирующую, применение, антигена, антиген
Код ссылки
<a href="https://eas.patents.su/30-16648-primenenie-defektnogo-po-replikacii-rekombinantnogo-adenovirusa-soderzhashhego-geterologichnuyu-nukleinovuyu-kislotu-kodiruyushhuyu-antigen-cs-vozbuditelya-malyarii-i-belkovogo-ant.html" rel="bookmark" title="База патентов Евразийского Союза">Применение дефектного по репликации рекомбинантного аденовируса, содержащего гетерологичную нуклеиновую кислоту, кодирующую антиген cs возбудителя малярии, и белкового антигена, содержащего белок cs или его фрагмент, для лечения или профилактики малярии</a>
Способ получения партий рекомбинантного аденовируса и используемые для этого пакующие клетки

Номер патента: 10924
Опубликовано: 30.12.2008
Авторы: Вогелс Роналд, Зюэйдгест Давид А.Т.М., Хавенга Мензо Янс Эмко
МПК: C12N 15/86, C12N 5/10
Метки: получения, аденовируса, этого, партий, способ, пакующие, используемые, клетки, рекомбинантного
Формула / Реферат:
1. Способ получения клетки с интегрированным в ее геном кодирующими последовательностями Е1А и Е1В-19K и Е1В-55K аденовируса, где способ включает стадию введения в клетку-предшественник: a) единичной молекулы нуклеиновой кислоты, содержащей кодирующие последовательности Е1А и Е1В-19K и Е1В-55K, где по меньшей мере две из этих трех кодирующих последовательностей разделены по меньшей мере 4 т.п.н.; или b) двух или более молекул нуклеиновой...
Гибридный белок проинсулина и способ получения из него рекомбинантного инсулина

Номер патента: 5586
Опубликовано: 28.04.2005
Авторы: Кондо Масааки, Ока Сусаку, Ваки Йосихиро, Юки Хиротака, Ватанабе Сигеаки, Кудо Тосиюки, Хигасикуни Наохико, Сато Сейдзи
МПК: C07K 19/00, C12N 15/17
Метки: получения, рекомбинантного, гибридный, него, инсулина, проинсулина, способ, белок
Формула / Реферат:
1. Гибридный белок проинсулина формулы (I) [Y]-[X1]-[B-цепь]-[X2]-[линкер]-[X3]-[A-цепь] (I) где Y представляет собой лидерную пептидную последовательность для экспрессии и секреции белка, содержащую 9 N-концевых аминокислотных остатков белка MWP, который представляет собой один из белков клеточной стенки (CWP) бактерии, принадлежащей к роду Bacillus; X1 представляет собой аминокислотную последовательность, расщепляемую тромбином; B-цепь...
Конъюгат гаптен-носитель (варианты), антитело и способ его получения, функциональный фрагмент антитела, набор для определения присутствия никотина в образце, вакцинная композиция (варианты) и способ профилактики или лечения никотиновой аддикции (варианты)

Номер патента: 4956
Опубликовано: 28.10.2004
Авторы: Эннифар Софиан, Фаттом Али Ибрагим, Нэйсо Роберт Б.
МПК: A61P 25/34, A61K 47/48
Метки: присутствия, образце, лечения, функциональный, получения, варианты, фрагмент, вакцинная, определения, профилактики, гаптен-носитель, композиция, антитела, никотиновой, антитело, способ, набор, аддикции, конъюгат, никотина
Формула / Реферат:
1. Конъюгат гаптен-носитель формулы где m составляет 1 - 2500, n составляет 0-12, y составляет 1-12, X выбран из группы, состоящей из NH-CO, CO-NH, CO-NH-NH, NH-NH-CO, NH-CO-NH, CO-NH-NH-CO и S-S; Y выбран из группы, состоящей из NH-CO, CO-NH, CO-NH-NH, NH-NH-CO, NH-CO-NH, CO-NH-NH-CO и S-S, а структура -(CH2)n-X-(CH2)y-Y- связана по положению 3', 4' или 51 никотина. 2. Конъюгат по п.1, отличающийся тем, что m составляет 11-17, n равно 1,...
Стимулятор роста возбудителя туберкулеза, питательная среда для выделения возбудителя туберкулеза, способ получения питательной среды, способ выделения возбудителя туберкулеза

Номер патента: 5527
Опубликовано: 28.04.2005
Авторы: Багрий Петр Иванович, Власенко Владимир Васильевич
Метки: стимулятор, получения, среды, возбудителя, роста, питательная, выделения, способ, питательной, туберкулеза, среда
Формула / Реферат:
1. Стимулятор роста возбудителя туберкулеза, содержащий химические соединения, в состав которых входят элементы натрий, кислород, водород, углерод, отличающийся тем, что характеризуется следующим составом ингредиентов, мас.%: Сахароза 2,0-5,0 Спирт ректифицированный 0,1-0,9 Натрий двууглекислый 0,05-0,15 Натрий пироцитрат двузамещенный 0,005-0,9 Растворитель До 1000 мл pH 7,2+0,2 2. Стимулятор роста по п.1, отличающийся...
Применение бактериальной комбинированной композиции для получения средства для профилактики и/или лечения гипероксалурии у людей и животных и способ профилактики и/или лечения указанного заболевания

Номер патента: 4257
Опубликовано: 26.02.2004
Автор: Де Симоне Клаудио
МПК: A61K 39/02, A61P 13/04, A23L 1/03...
Метки: способ, гипероксалурии, указанного, лечения, композиции, получения, комбинированной, применение, заболевания, бактериальной, животных, профилактики, людей, средства
Формула / Реферат:
1. Применение бактериальной комбинированной композиции, включающей в качестве штаммов бактерий (a) Streptococcus thermophilus в смеси с (b) по меньшей мере одним штаммом бактерий, выбранным из группы, состоящей из Lactobacillus brevis, Lactobacillus acidophilus, Lactobacillus plantarum, Bifidobacterium infantis, Bifidobacterium longum и Bifidobacterium breve или их смесей, для получения диетического, фармацевтического или ветеринарного средства...
Предыдущий патент: Теплообменник электрической машины обращенного вида
Следующий патент: Комбинация производных триазина и стимуляторов секреции инсулина
Случайный патент: Удаление магнитных частиц из текучей среды