Способ лечения воспаления и связанных с ним заболеваний и способ улучшения уровня не менее одного показателя крови, а также способ очистки дрожжевой рибонуклеиновой кислоты














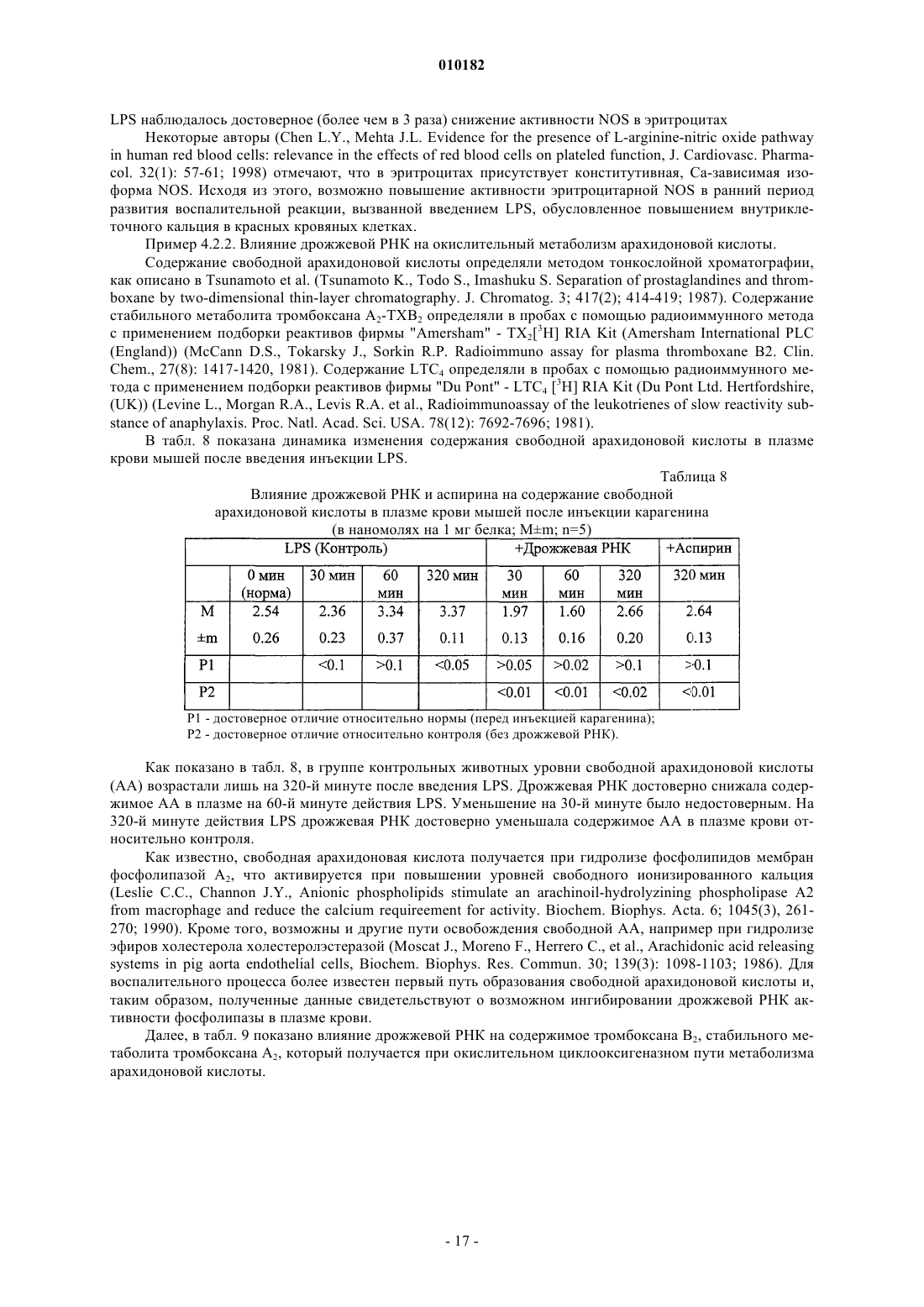
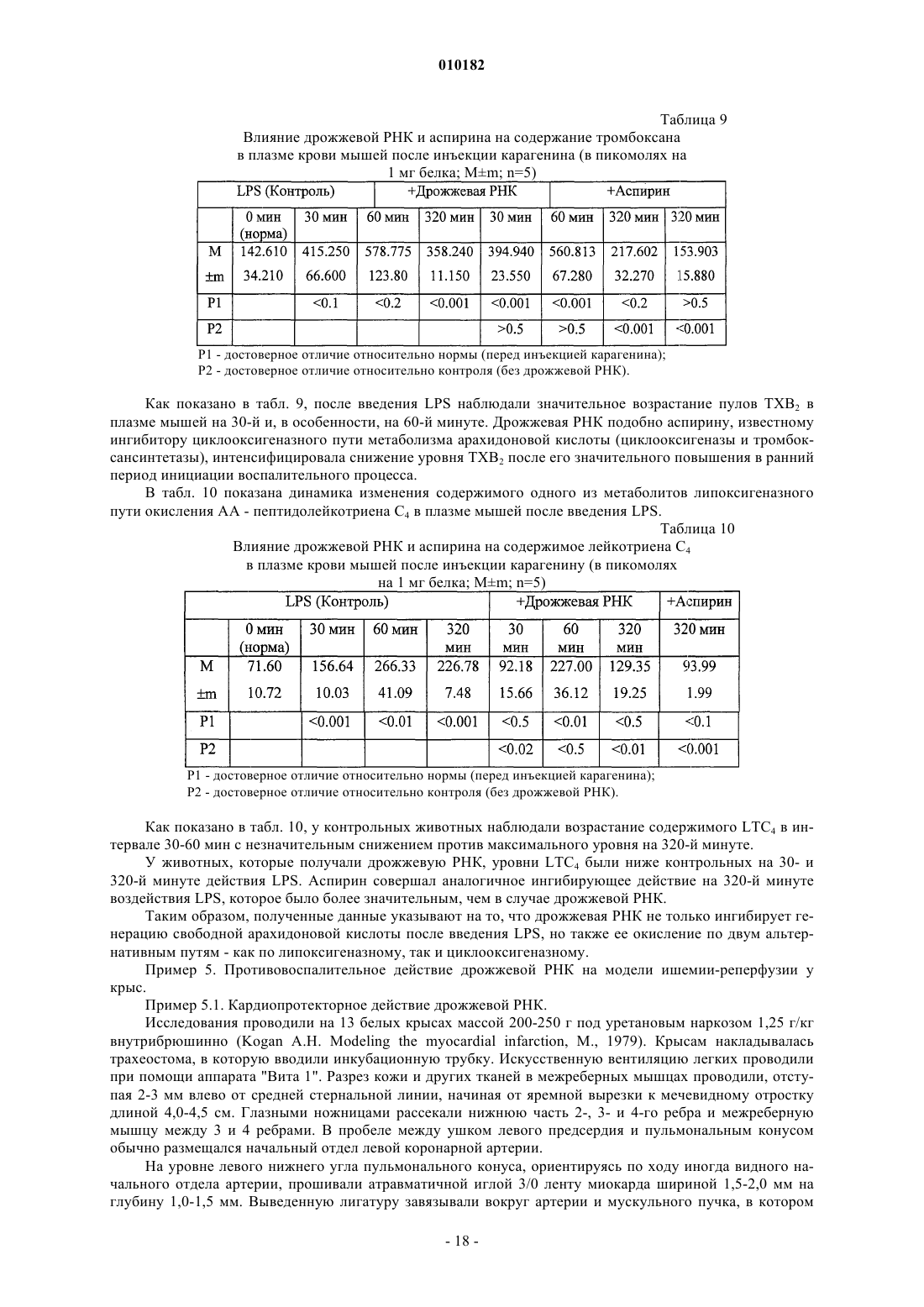

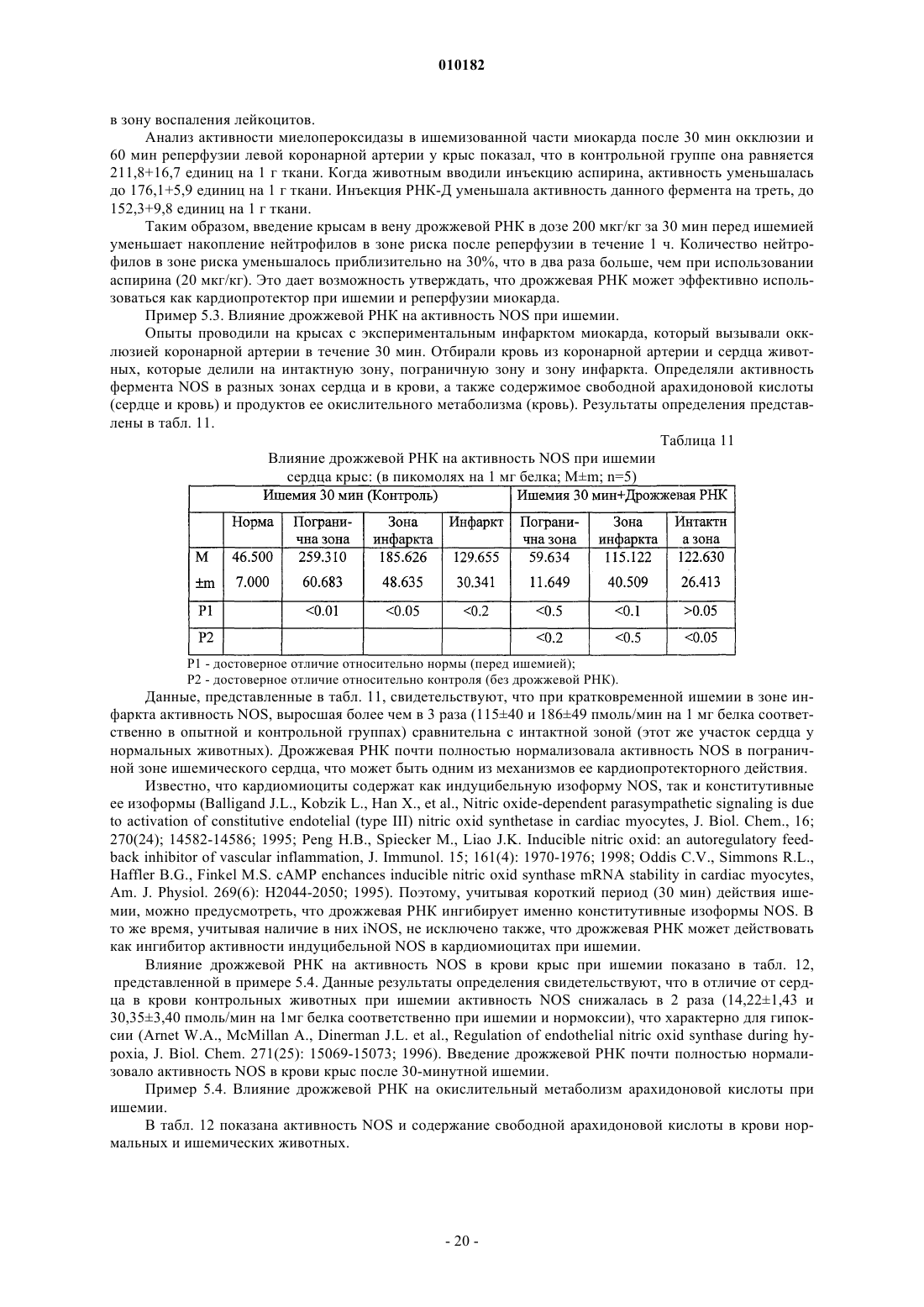
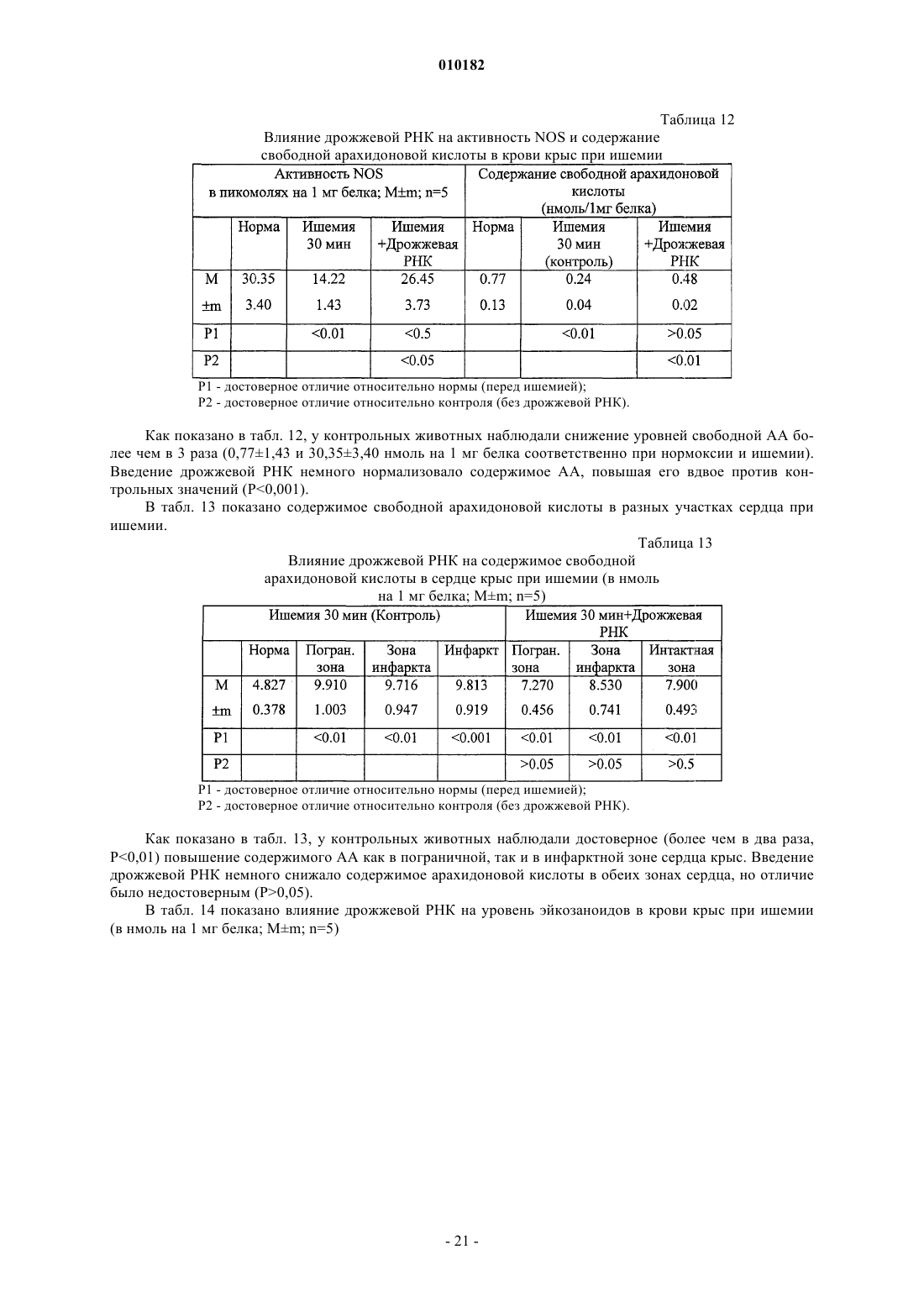



Формула / Реферат
1. Способ лечения воспаления и связанных с ним заболеваний за счет стабилизации клеточных мембран, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном улучшать симптомы воспаления или связанных с ними заболеваний, в фармакологически приемлемых наполнителях, носителях и растворах.
2. Способ по п.1, отличающийся тем, что млекопитающим, у которых имеются повреждения клеточных мембран, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном стабилизировать повреждение клеточной мембраны, в фармакологически приемлемых наполнителях, носителях и растворах.
3. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном угнетать окисление компонентов клеточных мембран млекопитающих, в фармакологически приемлемых наполнителях, носителях и растворах.
4. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном нормализовать активность NO-синтетазы, в фармакологически приемлемых наполнителях, носителях и растворах.
5. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном угнетать агрегацию тромбоцитов, в фармакологически приемлемых наполнителях, носителях и растворах.
6. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном препятствовать или лечить цитопению крови, в фармакологически приемлемых наполнителях, носителях и растворах.
7. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают интрадермально, подкожно, перорально, интраабдоминально, внутримышечно или внутривенно либо назначают в месте воспаления или в месте, связанном с воспалительным заболеванием.
8. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде капсул.
9. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде суппозиториев.
10. Способ по п.1, отличающийся тем, что связанным с ним заболеванием является инфаркт.
11. Способ по п.1, отличающийся тем, что связанным с ним заболеванием является инсульт.
12. Способ по п.1, отличающийся тем, что связанным с ним заболеванием является артрит.
13. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде фармацевтической композиции, которая содержит очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах.
14. Способ улучшения уровня не менее одного показателя крови за счет стабилизации мембран клеток крови, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном улучшать уровень не менее одного показателя крови, в фармакологически приемлемых наполнителях, носителях и растворах.
15. Способ по п.14, отличающийся тем, что такой показатель крови выбирают из соответствующих уровней лейкоцитов, эритроцитов, тромбоцитов, гемоглобина, нейтрофилов и гематокрита.
16. Способ по п.14, отличающийся тем, что млекопитающим, которые требуют профилактики или лечения анемии, назначают очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах.
17. Способ по п.14, отличающийся тем, что млекопитающим, которые требуют профилактики или лечения тромбоцитопении, назначают очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах.
18. Способ по п.14, отличающийся тем, что млекопитающим, которые требуют профилактики или лечения нейтропении, назначают очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах.
19. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают интрадермально, подкожно, перорально, интраабдоминально, внутримышечно или внутривенно либо назначают в месте воспаления или в месте, связанном с воспалительным заболеванием.
20. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде капсул.
21. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде суппозиториев.
22. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде фармацевтической композиции, которая содержит очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах.
23. Способ очистки дрожжевой рибонуклеиновой кислоты, отличающийся тем, что рибонуклеиновую кислоту очищают от белков и ДНК при помощи многократной инкубации с применением ферментов, в частности панкреатина.
24. Способ по п.23, отличающийся тем, что рибонуклеиновую кислоту получают из Saccharomyces cerevisiae.
25. Способ по п.23, отличающийся тем, что рибонуклеиновую кислоту получают из Candida utilis.
26. Способ по п.23, отличающийся тем, что очищенная рибонуклеиновая кислота содержит азот в количестве более 14,5 мас.%.
27. Способ по п.23, отличающийся тем, что очищенная рибонуклеиновая кислота содержит фосфор в количестве более 8,5 мас.%.
Текст
010182 Область изобретения Данное изобретение описывает соединение и фармацевтическую композицию для лечения воспаления и болезней, сопровождающих воспалительный процесс, в частности воспаление, влияющее на клеточную мембрану. Данное изобретение также описывает терапевтические способы для улучшения или предупреждения воспаления. Предпосылки создания изобретения Воспаление. Воспаление всегда сопровождается изменениями в обмене арахидоновой кислоты, обмене оксида азота и образованием свободных радикалов. Противовоспалительные нестероидные препараты (ПВНП),такие как аспирин, могут блокировать определенные этапы воспалительного процесса, но они не могут стабилизировать клеточные мембраны, что ограничивает их влияние на воспалительный процесс и делает это влияние малоэффективным. Воспаление - это локализованная реакция живой ткани на повреждение, происходящее под влиянием различных факторов экзогенного и эндогенного происхождения. Среди экзогенных факторов различают физические, химические и биологические. Среди эндогенных факторов выделяют медиаторы воспаления, антигены и антитела. Эндогенные факторы часто образуются под влиянием экзогенных повреждений. Воспалительная реакция обязательно сопровождается нарушением строения и проницаемости клеточной мембраны. На уровне тканей и органов воспаление проявляется болями, покраснением, повышением температуры, опухлостью, а порой и потерей функции. Воспаление начинается с сублетального повреждения и заканчивается или полным выздоровлением, или долгосрочным разрушением ткани. Без воспаления невозможно выздоровление после ранения. В результате повреждения ткани возникает немедленный ответ, который реализуется через медиаторы, выделяющиеся вследствие экзоцитозиса или лизиса клетки. Главными медиаторами при воспалении являются соединения кининовой и фибринолитической системы, системы комплемента, метаболиты арахидоновой кислоты, сосудистоактивные амины и прочие химические вещества. К химическим медиаторам воспаления относятся гистамин, серотонин, простагландины, белок, относящийся к гену кальцитонина, оксид азота и др. Важную роль в воспалении играют также разнообразные виды реактивного кислорода. Эти соединения синтезируются, когда кислород превращается в очень разрушительные соединения, образовывая свободные радикалы. Это такие атомы или молекулы, которые имеют неспаренные электроны. Разнообразные свободные радикалы имеют разные уровни реактивности. Запуск воспаления происходит под влиянием экзогенных и эндогенных агентов. С участием эндогенных факторов, а именно медиаторов, антигенов и аутоантигенов определяется выраженность и характер воспалительной реакции, особенности ее протекания в месте повреждения. Когда повреждение ограничивается образованием медиаторов, развивается острое воспаление. Когда же в процесс включаются иммунологические реакции, а именно последующие взаимодействия антигенов, антител и аутоантигенов, это приводит к продолжительному хроническому воспалительному процессу. Разные агенты, например инфекция, травма, радиация, повреждая клеточную мембрану, запускают биохимические реакции, которые обеспечивают протекание воспалительного процесса на молекулярном уровне. В основе воспалительных процессов лежит метаболизм арахидоновой кислоты, которая превращается в простагландины (PG), тромбоксаны (TX) и лейкотриены (LT). Простагландины, тромбоксаны и лейкотриены являются главными участниками всех воспалительных процессов. Известны два пути каскада превращений арахидоновой кислоты. Первый путь ведет к образованию простагландинов G2 и H2. Этот процесс катализируется простагландин-циклооксигеназой. Циклооксигеназа катализирует образование PGA2, PGE2, PGD2, PGF2, а тромбоксан-синтетаза с PGH2 образует тромбоксан A2 (TXA2). Лучше всего изученным метаболическим путем является каскад преобразований арахидоновой кислоты, получаемой из фосфолипидов мембран при помощи фосфолипазы A2. Через циклогеназный и липоксигеназные каскады арахидоновая кислота превращается, соответственно, в простагландины и лейкотриены. Циклооксигеназный путь приводит к двум биологически активным продуктам - простациклину (PGI2) и тромбоксану (TXA2). Эти продукты задействованы в различных проявлениях воспаления: сужение бронхов, расширение и сужение сосудов, агрегация тромбоцитов, аналгезия, пироксия и т. д. Другой путь метаболизма арахидоновой кислоты при помощи 5-липоксигеназы ведет к образованию лейкотриенов: LTA4, LTB4, LTC4, LTD4, LTE4, и LTF4, которые имеют сильное противовоспалительное и сужающее сосуды бронхов действие, вместе с тем они играют важную роль в проницаемости сосудов. Лейкотриены важны как потенциально хемоактивные факторы, они увеличивают миграцию лимфоцитов и отвечают за многие действия веществ, медленно выделяемых при анафилаксисе (SRS-A). Простагландины могут играть значительную роль в развитии системных воспалительных реакций. При ревматоидном артрите большие количества простагландинов и лейкотриенов в синовиальной жидкости оказывают содействие развитию воспалительного процесса и деминерализации околосоединительной костной ткани. Лейкотриены считаются основными патофизиологическими медиаторами воспалительных реакций. Они значительно сильнее, чем простагландины, влияют на проницаемость сосудов и адгезию лейкоцитов со стенками сосудов, а также на развитие отека.-1 010182 Простагландины эффективно регулируют агрегацию тромбоцитов. PGE1 - мощный ингибитор агрегации тромбоцитов, тогда как PGE2, который в норме высвобождается из тромбоцитов, - стимулирует этот процесс. Однако главную роль в свертывании крови играет PGI2, или простациклин, синтезируемый в стенках кровеносных сосудов из арахидоновой кислоты. Это самый мощный ингибитор агрегации тромбоцитов, имеющий сосудорасширяющее свойство. Синтезируемый в тромбоцитах тромбоксан имеет противоположное действие. При повреждении эндотелия происходит адгезия тромбоцитов с субэндотелиальной соединительной тканью и агрегация тромбоцитов. Основную роль в этом процессе играет тромбоксан A2. Наоборот,простагландин I2 ингибирует агрегацию тромбоцитов. Поэтому от соотношения простагландина I2 и тромбоксана A2 зависит, будет ли нормально протекать процесс коагуляции. Кроме того, особую роль в процессе выздоровления от воспаления играет оксид азота (NO). Этот газ очень быстро проникает в разные органы и ткани и как свободный радикал он обладает мощной реактивностью. Оксиду азота свойственна активность вазодилатора, нейротрансмиттера и медиатора, поэтому он играет исключительную роль при астматическом воспалении. Оксид азота вырабатывается эндогенно из аминокислоты L-аргинина при помощи NO-синтетазы. Известны три формы NO-синтетазы. Две формы - конститутивные и третья индуцибельная. Индуцибельная NO-синтетаза, которая экспрессируется в эпителиальных клетках, быстро повышает свою активность при выделении противовоспалительных цитокинов, таких как интерлейкин 1 beta (IL-1) и фактор некроза опухолей (TNF-). Оксид азота имеет положительные и отрицательные свойства по отношению к воспалительному процессу. Очень важным потенциально положительным свойством NO является способность расслаблять бронхиальную гладкую мышцу, что приводит к бронходиляции. Среди отрицательных свойств выделяется его способность оказывать содействие воспалительному процессу, увеличивая хемотаксис нейтрофилов, моноцитов и эозинофилов с использованием гуанозин-монофосфат зависимого механизма. Считают, что он ингибирует адгезию лейкоцитов к сосудистому эндотелию и бронхиальному эпителию.NO играет исключительную роль при астматическом воспалении.NO играет особенно важную биологическую роль в определении базального сосудистого тонуса,регулирует сокращаемость миокарда и модулирует взаимодействие между тромбоцитами и сосудистой стенкой (Zhou Q., Hellermann G.R., Solomonson L.P., Nitric oxide release from resting human platelets,Thromb.Res., 1;77(1):87-86; 1995). Известно участие NO в тромбоцитарной активации при патогенезе разнообразных сосудистых тромботических заболеваний человека, а также есть данные, чтоNO-опосредствованные эффекты снижены при гипертонии (Calver A., Collier J., Moncada S., Vallance P.,Effect of local intraarterial NG-monomethyl-L-arginin in patients with hypertension: the nitric oxide dilatorpatient by L-arginine, Lancet., 21-28; 338(8782-8783); 1546-1550; 1991). Эти данные указывают на возможность успешного лечения больных за счет препаратов, которые могут увеличивать активностьNO-синтетазы. Тромбоциты человека способны синтезировать оксид азота. В больших количествах, таких как это могут делать эндотелиальные клетки, он может синтезироваться как интактными, так и стимулированными тромбоцитами. Поэтому оксид азота тромбоцитарного происхождения играет важную роль в поддержке сосудистого гомеостаза и других NO-чувствительных процессах (Zhou et al., 1995). Вместе с общими чертами в каждом конкретном заболевании воспалительный процесс имеет свою специфику, связанную с особенностями функционирования органа, факторами, которые приводят к повреждению: вирусы, микроорганизмы, ранение, отравление и т. д. Например, общим механизмом заболеваний сердца является нарушение структуры и функции мембран клеток сердца. Вследствие этого возрастает синтез лейкотриенов и тромбоксанов, которым свойственно коронароконстрикторное и проагрегатное действие. (Bangham A.D., Hill M.W., Miller N.,Preparation and use of liposom as model of biological membranes, Methods in Membrane Biology, Acad. Press,v. 1, NY, p. 1-68, 1974). Важным фактором патогенеза повреждений сердца является коронароконстрикторное и хемоатрактивное относительно нейтрофилов действие лейкотриенов LTC4, LTD4, LTB4 (Hoshida S., Kuzuya Т.,Nishida M., et al., Amer. J. Cardiol., 7; 63(10): 24 Е-2 Е; 1989; Lam B.K., Gagnon L., Austen K.F. et al., J. Biol.Chem., 15; 265(23): 1338-1341; 1990; Svendsen J.N., Hansen P.R., Ali S. et al., Cardiovasc. Res., 27(7): 12881294; 1993). Вещества, которые смогут блокировать этот процесс, в свою очередь, смогут уменьшать размеры некроза при остром инфаркте и, таким образом, существенно уменьшить летальность при тяжелых случаях заболевания сердца, например при обширных инфарктах. Одновременно такие вещества должны стабилизировать мембраны клеток сердца. Известно, что коронароконстрикторные и хемоатрактивные эффекты при инфаркте сопровождаются возрастанием агрегации тромбоцитов, поэтому блокирование этого процесса тоже будет приводить к уменьшению размеров поражения.-2 010182 Кроме того, нарушения агрегатного состояния крови играют важную роль в патогенезе многих заболеваний, в особенности это проявляется в патогенезе разных сосудистых тромботических заболеваний человека. Установлено, что одним из ключевых факторов разлада регуляции агрегатного состояния крови является нарушение тромбоцитарно-сосудистого звена гомеостаза. Оно приводит к изменению реологичных свойств крови, содействуя образованию сосудистых агрегатов. Тромбоцитарные повреждения приводят к нарушению процессов микроциркуляции, возникновению дефицита притока крови к тканям. Начальным этапом тромбообразования являются активация тромбоцитов с их последующей адгезией на поврежденном эндотелии и агрегация с образованием первичного тромбоцитарного тромба. Сейчас существует достаточно доказательств в пользу тесного взаимодействия между воспалительными нарушениями агрегатного состояния крови и сердечно-сосудистыми нарушениями (Anderson J.L.,Carlqist J.L., et al. Evaluation of C-reactive protein an inflammatory marker, and infectious serology as risk factors for coronary artery disease and myocardial infarction, J. Am. Coll. Card., 32: 35-41; 1998). Повреждение клеточных мембран или воспалительные процессы в организме часто сопровождаются цитопенией. Во многих случаях такие пациенты имеют анемию, тромбоцитопению или нейтропению. Анемия сопровождается уменьшением количества эритроцитов и гемоглобина, сопровождающимся потерей крови, нарушением синтеза эритроцитов, увеличением их разрушения или комбинацией этих факторов. В случае тромбоцитопении количество тромбоцитов в крови уменьшается, что приводит к нарушению тромбогенеза с последующим кровоизлиянием. Нейтропения характеризуется уменьшением количества нейтрофилов в крови, часто приводящим к увеличению чувствительности таких больных к разнообразным инфекциям. Нормальное образование клеток крови, или гемопоэз, начинается из кроветворных стволовых клеток предшественников костного мозга, которые называются CFU-GEMM. Эти клетки у взрослых людей и животных формируются в костном мозге и под влиянием факторов роста превращаются в специализированные клетки крови. Например, эритроциты получаются из CFU-GEMM под влиянием эритропоэтина. Под влиянием тромбопоэтина CFU-GEMM превращаются в тромбоциты. Соответственно, под влиянием гранулоцит-макрофаг-колоний стимулирующего фактора CFU-GEMM превращаются в гранулоциты и моноциты. В то же время лимфоциты происходят из лимфоидных стволовых клеток. Наиболее резко выраженная цитопения с тяжелыми последствиями проходит у больных раком, в особенности после химиотерапии или радиотерапии, у больных СПИДом и инфицированных вирусомUpdate, 2000, Medscape, Inc.). Роль клеточных мембран при воспалении. Функции клеточных мембран и их взаимосвязь с воспалением хорошо изучены. Известно, что плазматическая мембрана клеток занимает особое место среди мембранных структур, выполняя ряд наиболее важных функций: барьерную, транспортную, обеспечение контакта клетки с внешней средой, принимает участие в регуляции гомеостаза клеток, обеспечивает сигнальные механизмы этой регуляции,определяет индивидуальность и целостность клеток. Структурная организация, динамика, функции мембран эритроцитов и закономерности разных типов гемолиза - осмотического, окислительного, иммунного (вызванного гемолитическими вирусами, токсинами, комплементом), гемолиза детергентами,фотогемолиза и др. исследованы достаточно детально (Bashford C.L., Alder G.M., Menestrina G.,et al.peptide toxins, aliphatic alcohols, related glycols and benzylidene derivatives, Biochim. Biophys. Acta., 16; 1029(2): 252-258; 1990). Показано, что варьирование pH внешней среды поднимает баланс действующих на мембране сил,что приводит к изменениям структурного состояния и степени агрегации мембранных белков. При этом выделяют структурную перестройку мембраны в диапазоне pH 7,0-6,0 и при уменьшении pH ниже 4,5(Zavodnik I.B., Pileckaya T.P. Acid lysis of human erythrocytes, Biophizika. V. 42,5, p. 1106-1112, 1997). Вследствие второго перехода происходит дестабилизация мембраны со следующим лизисом эритроцитов. Известно, что при pH 4,7 в эритроцитарной мембране наблюдаются нарушения гликокаликса, которые трансформируются в поры (Arvinte Т., Cudd A., Schulz В., Nicolau C., Biochim. Biophys. Acta., 19; 981(1): 61; 1989). Уменьшение pH среды изменяет конформацию, способ упаковки и подвижность фосфолипидов модельных мембран. Агрегация денатурированных снижением pH мембранных белков является причиной мембранных повреждений и кислотного лизиса эритроцитов. Предложена схема процесса гемолиза эритроцитов HCl, которая сводится к кооперативному протонированию некоторого центра в строме или на мембране эритроцита с последующим образованием пор,достаточных для выхода гемоглобина. Исследование механизмов и закономерностей процесса кислотного гемолиза позволяет получить информацию о структурной организации мембраны и взаимодействиях,которые ее стабилизируют. Наиболее известными эндогенными стабилизаторами гемолиза эритроцитов (в основном исследован осмотический гемолиз) являются альбумин плазмы крови, ионы металлов K+, Na+, Mg2+ и в особен-3 010182 ности Ca2+, которые модулируют каналы плазматической мембраны эритроцитов, в том числе, возможно,и протонный канал (Anderson D.R., Davis J.L., Carraway K.L. Calcium-promoted changes of the human erythrocyte membrane. Involvement of spectrin, transglutaminase, and a membrane-bound protease. J. Biol. Chem.,10; 252(19): 6617-6623, 1977), холестерол, адсорбированный на поверхности эритроцитовerythrocyte membrane, J. Cell. Biol., 85(2): 283-291; 1980), и полиамины, которые связываются с остатками жирных кислот фосфолипидов мембран (Rennert O.M., Shukla J.B. Polyamines in health and diseaseAdvances in Polyamine research, Raven Press, v. 2, NY, p. 195-21, 1978). Наиболее известными активаторами эндогенного гемолиза эритроцитов являются длинноцепочечные жирные кислоты и(Rybszynska M., Csordas A. Chain length-dependent interaction of free fatty acids with the erythrocyte membrane, Life Sci., 44(9): 625-632; 1989), в особенности, свободные радикалы кислорода и азота (Sato Y.,Kamato S., Takahashi T. et al. Mechanism of free radical-induced hemolysis of human erythrocytes: hemolysisbleeding time by acute hemolysis in rats: a role for nitric oxide. Am. J. Physiol. 272(6): 2875-2884; 1997). Таким образом, есть достаточно доказательств, которые свидетельствуют, что структура мембраны изменяется в процессе воспаления. Однако модель нарушения мембраны при воспалении не использовалась для скрининга лекарств, способных лечить и предупреждать воспаления или связанные с воспалением заболевания. Неудовлетворительное состояние имеющегося противовоспалительного лекарства. Имеющиеся противовоспалительные препараты не соответствуют современным требованиям, поскольку сложность и разнообразие биохимических реакций при воспалении, отсутствие точных данных о патогенезе воспаления в значительной мере усложняют отбор в эксперименте фармакологических препаратов, способных регулировать воспаление. Имеющиеся препараты могут влиять только на отдельные этапы воспалительного процесса. До сих пор не получен препарат, способный в равной степени регулировать большинство компонентов любой воспалительной реакции. Большинство известных нестероидных противовоспалительных препаратов (NSAIDS) выборочно влияют на те или другие фазы этого патологического процесса. В первую очередь они влияют на нарушения проницаемости сосудов, характерные при остром воспалении, и на разнообразные клеточные реакции, присущие хроническому воспалению. Вдобавок, многие NSAIDS влияют на метаболизм через механизм свободных радикалов. Для первичного скрининга противовоспалительных процессов используют три основные группы методов. Сначала изучают влияние препаратов на признаки воспаления, которые легко определить. Это опухлость, гиперемия, некроз и т. д. Для более углубленного изучения применяют методы экспериментальной терапии, модели артритов, кардитов и т. д., которые напоминают болезни людей. На третьем этапе изучают механизмы влияния препаратов на конкретные метаболические пути. Вследствие детального изучения метаболизма арахидоновой кислоты было предложено много противовоспалительных препаратов, действие которых направлено на регуляцию образования продуктов ее метаболизма. Чаще всего такие вещества обладают активностью ингибиторов ферментов метаболизма арахидоновой кислоты. Примером может служить предложенная как противовоспалительная фармакологическая комбинация ингибитора циклооксигеназы 2 и ингибитора лейкотриена A.суб. 4 гидролазыPatent5,990,148, Nov. 1999). Подобный подход предложен на базе аналогов пиримидина - составной части нуклеиновых кислотagents, United States Patent5,240,929, Aug. 1993). Поскольку эти соединения оказались ингибиторами ключевых ферментов метаболизма арахидоновой кислоты 5-липоксигеназы и циклооксигеназы, авторы предложили использовать их как противовоспалительные препараты для лечения очень широкого спектра болезней: от аллергических заболеваний и ревматоидного артрита до атеросклероза, инфаркта миокарда и инсульта (Haslanger M.F., Prostacyclin analogs and their use in inhibition of arachidonic acid-inducedplatelet aggregation and bronchoconstriction, United States Patent4,192,891, Mar. 1980). Однако, поскольку воспалительный процесс запускает очень большое количество метаболических каскадов, использование ингибиторов или аналогов метаболизма арахидоновой кислоты не дает возможности сбалансировать все реакции и таким образом комплексно регулировать прохождение воспалительного процесса. Более того, аспирин, используемый в практической медицине продолжительное время, тоже был предложен для блокирования ферментов метаболизма арахидоновой кислоты. Ингибиторы простагландинов, такие как аспирин, довольно эффективно влияют на воспалительный процесс. По этой причине они успешно используются в клинике для лечения ревматоидного артрита, остеоартрита и других воспалительных процессов. Аспирин обладает антикоагулятивным действием, поскольку он ингибирует синтез TXA2, тем не менее, хотя и частично, он действует на синтез PGI2. Ежедневный прием аспирина в-4 010182 дозах до 3 г в день очень широко применяют при профилактике стенокардии и в постинсультном периоде, а также у людей с высоким риском развития сердечно-сосудистых заболеваний. Однако изучение синтеза TXA2 и PGI2 in vivo показало, что одно пероральное употребление 0,5-1,0 г аспирина уменьшает выделение PGI2 только на 2-3 ч, тогда как выделение тромбоксана задерживает на 10 дней (Vesterqvist О., Measurements of the in vivo synthesis of thromboxane and prostacyclin inhumans, Scand. J. Clin. Lab. Invest. 48(5): 401-407; 1988). Эта и другие работы (Lorenz R.L., Boehlin В.,Uedelhoven M.W., Weber P.C. Superior antiplatelet action of alternate day pulsed dosing versus split dose administration of aspirin, Am. J. Cardiol. 15; 64(18): 1185-1188; 1989) не только показывают трудность в назначении дозы аспирина, но и экспериментально обосновывают те частые побочные эффекты, которые вызывает аспирин при его продолжительном применении. В частности, аспирин и прочие нестероидные противовоспалительные средства могут быть причиной анафилактоидных реакций у чувствительных к ним людей. В их основе лежит зависимый от дозы токсично-идиосанкратический, а не иммунологический механизм. Аспирин является причиной наиболее распространенных случайных отравлений, большому риску подвергаются дети, которые перед отравлением лечились аспирином. Часто случается передозировка аспирина, трудно поддающаяся коррекции. Эффективные дозы аспирина при многих заболеваниях, например при ревматоидном артрите составляют от 3,0 до 6,5 г в день, что, кроме проявлений токсичности, приводит к раздражению желудочнокишечного тракта. Аспирин тяжело переносится больными желудочно-кишечными заболеваниями, вызывает эрозии и кровотечения язвы желудка, диарею, язву двенадцатиперстной кишки. Аспирин широко применяется благодаря своей антитромбоцитарной активности, но он тяжело переносим вследствие своего свойства вызывать побочные эффекты при продолжительном использовании. Более того, неспецифично ингибируя циклооксигеназу, аспирин влияет на синтез тромбоксана, который является сильным агрегантом и сосудосужающим препаратом и вазоконстриктором, и может приводить к уменьшению уровня простациклина, который имеет одновременно антиагрегантные и сосудорасширяющие свойства. Все эти отрицательные проявления аспирина и других NSAIDS побуждают к поиску новых препаратов, которые, обладая противовоспалительным действием аспирина, были бы нетоксичными в широком спектре концентраций, не вызывали побочных проявлений при продолжительном употреблении,останавливали и предупреждали возникновение очагов воспаления. Для лечения цитопении используют кроветворные факторы роста (J. Crawford, HematopoieticGrowth factor: Current Practice and Future Directions, 42nd Annual Meeting of the American Society of Hematology, 2000, Medscape, Inc.). Однако такое лечение довольно сложное и дорогостоящее. Например, для лечения цитопении у ВИЧ-инфицированных больных в зависимости от имеющихся нарушений используют факторы роста. У таких больных для лечения анемии используют эритропоэтин, для лечения тромбопении используют тромбопоэтин, а для лечения нейтропении - G-CSF. Однако лечение цитопении у ВИЧ-инфицированных больных требует очень дорогих диагностических исследований эндогенного уровня указанных факторов роста и очень дорогостоящих лекарств этих факторов роста, для производства которых широко используют рекомбинантную технологию. Поэтому довольно важным является поиск недорогих лекарств, которые могут нормализовать анемию, тромбопению и нейтропению больных. Фармацевтическое использование нуклеиновых кислот. Нуклеиновые кислоты широко используются как фармакологические препараты. (Rothenberg M.,Jonson G., Laughlin C. et al. Oligodeoxynucleotides as anti-sense inhibitors of gene expression: therapeutic implications, J. Natl. Cancer Inst., 18; 81(20): 1539-1544; 1989; Zon G., Oligonucleotides analogues as potentialchemotherapeutic agents, Pharm. Res., 5; (9): 539-549; 1988). Однако фармакологическое использование нуклеиновых кислот не включает воспаления и болезни, которые сопровождаются воспалением. Например, Anderson et al. предложили метод лечения цитомегаловирусной инфекции у людей при помощи олигонуклеотида, который, связываясь с мРНК цитомегаловируса, модулирует эффект данной инфекции.(Anderson K., Draper K., Baker В. Oligonucleotides for modulating the effects of cytomegalovirus infections,United States Patent5,442,049, August 15, 1995). На основе специфической нуклеиновой кислоты, которая кодирует последовательность 3'нетранслированного региона протеинкиназы C; Boggs et al. предложили метод диагностики и лечения заболеваний, которые ассоциированны с протеинкиназой C-альфа.(Boggs R.T., Dean N.M. Nucleic acid sequences encoding protein kinase C and antisense inhibition of expression thereof, United States Patent5,681,747, Oct. 1997). Так, Yano et al., запатентовали ДНК соединение,полученное из Mycobacterium bovis и Bacillus subtilis для лечения язвы желудка (Yano О., Kitano Т.Method for the treatment of digestive ulcers, United States Patent4,657,896, Apr. 1987). В частности, известно, что рибонуклеиновая кислота (РНК), продукты ее частичного гидролиза и синтетические полинуклеотиды обладают широким спектром биологической активности (Kordyum V.A.,Kirilova V.S., Likhachova L.I. Biological action of exogenous nucleic acids, Visnyk ASC USSR, v. 41,6,p. 67-78, 1977). Они активируют протеиновый синтез в клетках (Sved S.C., The metabolism of exogenousribonucleic acids injected into mice, Canad. J. Biochem.,v. 43,7, p. 949, 1965) и обладают противоопухолевой активностью (Niu M.C. Effect of ribonucleic acid on mouse acids cells, Sciens.,131, p. 1321, 1960). Препараты РНК могут увеличивать образование антител и сокращают индуктивную фазу антителогенезаevents in antibody formation, Proc. Soc. Exper. Biol. Med., 5, v. 119,3, p. 701, 1967). Показано, что некоторые пониженные, или повышенные иммунологические показатели под влиянием дрожжевой РНК нормализовались. В первую очередь, это касалось T-лимфоцитов, кооперации T- и В-лимфоцитов, активации функции макрофагов и т. д. Более того, экзогенная РНК при делении клеток используется для синтеза ДНК, а в метаболизме клетки - для синтеза РНК. Показано, что через 2 ч после введения экзогенная РНК включается в РНК лимфоцитов и макрофагов (Enesco N.E. Fate of 14C-RNA infected into mice, Exper. Cell Res., v. 42,3,p. 640, 1966). Получены доказательства, которые разрешают предполагать, что дрожжевая тРНК может включаться в клетки в форме интактных молекул (Herrera F., Adamson R.H., Gallo R.C. Uptake of transferribonucleic acid by normal and leucemic cells, Proc. Nat. Acad. Sci. USA, 67(4): 1943-1950; 1970). Аналитическими методами показано, что РНК, в частности, присутствует во всех мембранах животных клеток (в мембранах эндоплазматического ретикулума, митохондрий, ядра и плазматических мембранах). В зависимости от типа ткани и метода выделения содержание РНК колеблется от 0,5 до 4% сухого веса мембран. Экспериментальные данные свидетельствуют, что в мембранах имеется особая мембранная РНК (Shapot V.S., Davidova S.Y. Liponucleoprotein as an integral part of animal cell membrans.reticulum of animal cells. Biochim et Biophis Acta, 24; 129(1); 206-209; 1966). Функции такой мембранной РНК до конца не ясны. Функции рибосомальной РНК мембран лучше изучены (Cundliffe E. Intracellular distribution ofribosoms and poliribosomes in Bacillus megaterriium. J. Mol. Biol., 28; 52(3): 467-481; 1970) Рибосомальная РНК содержится в бактериальных мембранах, мембранах ядра, внутренних и внешних мембранах митохондрий, внутренних мембранах аппарата Гольджи, который прилегает к плазматической мембране, в шероховатом эндоплазматическом ретикулуме, в разнообразных тканях животных, людей, растений,микроорганизмов и простейших. Считают, что гликолипиды и гликопротеины мембран, которые содержат N-ацетилнеураминовую кислоту, связывают рибосомальную РНК с рибосомами, поскольку мембраны, обработанные неуронидазой, теряют способность связываться с рибосомами (Scott-Burden Т.,Hawtrey A.O. Preparation of ribosome free membranes from rat liver microsomes by means of lithium chloride.Biochem. J. 115(5): 1063-1069; 1969). Кроме того, возможно, что места связывания рибосом с мембранами активируются половыми гормонами, причем канцерогены повреждают этот физиологический механизм. В пользу этого вывода свидетельствует уменьшение уровня мембраносвязанной РНК в процессе старения (Mainwaring W.J. The effect of age on protein synthesis in mouse liver. Biochem. J. 113(5): 869-878; 1969) и после кастрации животных (Tata J.R. The formation, distribution and function of ribosomes and microsomalmembranes during induced amphibian metamorphosis. Biochem J. 105(2): 783-801, 1967). Удаление из мембраны спермина ведет к обособлению от мембраны части связанной с ней РНК (Khawaja J.A. Interactionmagnesium. Biochim. Biophis. Acta., 29; 254(1): 117-128); 1971). При обработке РНКазой мембран нативных малых рибосом клеток миеломы они откалываются от мембраны, тогда как большие нативные субъединицы рибосом остаются связаными с мембраной (Mechler B.,Vassalli P. Membrane-bound ribosomes ofproperties of ribosomes. J. Cell. Biol. 67(1): 1-15; 1975). Вдобавок, нуклеотидные компоненты различных мембранных ферментов, например полиА-РНК фосфофруктокиназы, составляют возможный пул мембранной РНК (Hofer H.W., Pette D. The complex nature of phosphofructokinase - a nucleic acid countainingenzyme, Life Sci. 4(16): 1591-1596; 1965). Однако нуклеиновые кислоты, и в особенности РНК или их соединения, до сих пор не используются для лечения, или предупреждения воспаления, а также связанных с воспалением заболеваний. Большинство исследований по РНК выполнялось ранее на уровне in vitro. Более того, ни один из этих методов не позволяет непосредственно использовать РНК для лечения или предупреждения воспаления, а также связанных с воспалением заболеваний. Необходимость нового лекарства. Таким образом, сейчас существует необходимость в новых противовоспалительных препаратах, которые будут регулировать расстройства агрегатного состояния крови и будут иметь меньше отрицательных эффектов, чем аспирин и прочие NSAIDS. Более того, поскольку воспалительный процесс в своей начальной стадии сопровождается изменениями в структуре и функции мембран многих клеток, вовлеченных в воспалительный процесс, нужны лекарства, которые бы не только регулировали все компоненты воспалительного метаболического каскада, но также стабилизировали структуру и функцию мембран этих клеток. В частности, поскольку традиционная терапия малоэффективна при обширных инфарктах,осложненных кардиогенным шоком, существует потребность в новом лекарстве, способном остановить разрушения кардиоцитов.-6 010182 Краткое описание изобретения Данное изобретение предлагает способ лечения, предупреждения воспаления и связанных с ним процессов. Также изобретение относится к способу улучшения показателя крови за счет стабилизации мембран клеток крови. Назначают очищенную дрожжевую РНК, способствующую улучшению симптомов воспаления или связанных с ними заболеваний. Дрожжевая РНК - это гетерогенное соединение низкомолекулярной РНК, которое включает разное количество нуклеотидов, в основном от 5 до 25 нуклеотидов, полимеров нуклеотидов. В дрожжевой РНК преобладают олигонуклеотиды и транспортная РНК с большим количеством минорных основ. Поскольку главным свойством всех воспалительных процессов на молекулярном уровне является изменение проницаемости и структуры мембраны, в данном изобретении использован метод отбора лекарств, основанный на их способности стабилизировать клеточную мембрану при воспалении. Таким образом, анализируя разрушительные механизмы, индуцированные в плазматических мембранах разнообразными факторами, и изучая структурные элементы их взаимодействия, которые обеспечивают оптимальную организацию клетки, можно отобрать для практической медицины лекарства, которые имеют мембраностабилизирующее действие. Поскольку, как сейчас установлено, мембраны имеют низкомолекулярную РНК, которая, вероятно, играет мембраностабилизирующую роль, введение в организм экзогенной низкомолекулярной РНК приведет к стабилизации поврежденной мембраны, например тех клеточных мембран, которые находятся в воспалительном процессе. Стабилизация клеточной мембраны соединением данного изобретения приводит к нормализации метаболизма арахидоновой кислоты и обмена оксида азота, который имеет значительное противовоспалительное действие и является главным участником воспалительных процессов, например ревматоидного артрита, остеоартрита, аллергии (такой как астма), и других воспалительных проявлений, таких как боль, отек, жар, псориаз, воспаление кишечника, желудочно-кишечные язвы, сердечно-сосудистые заболевания, включая ишемическое заболевание сердца, атеросклероз, частичное повреждение мозга, вызванное инсультом, заболевания кожи (экзема, солнечная еритема, угри), лейкотриен-опосредствованные воспаления легких, почек, гастрокишечного тракта, кожи, простатиты и парадонтозы. Дрожжевая РНК эффективна при уменьшении активности iNOS во время аутоиммунных процессов как в начале болезни, так и в хронической стадии. Эти свойства разрешают использовать дрожжевую РНК при патологических процессах, которые сопровождаются индукцией iNOS: диабет, рак, гепатит,инфекционные болезни, нейродегенеративные болезни (болезни Паркинсона, Альцгеймера, рассеянный склероз, энцефалит) и пр. Более того, использование природных молекул нуклеиновых кислот, таких как соединение данного изобретения, в высоких концентрациях в качестве фармакологических препаратов не приводит даже к незначительному побочному действию, в особенности принимая во внимание тот факт, что эти соединения постоянно вводятся в человеческие и животные организмы вместе с пищей. Кроме того, данное изобретение предлагает способ лечения воспаления и связанных с воспалением заболеваний, который предусматривает введение животным, которым необходимо такое лечение, очищенной дрожжевой рибонуклеиновой кислоты в эффективном количестве и в фармакологически пригодных наполнителях, носителях и растворах для улучшения симптомов воспаления или связанных с воспалением заболеваний. Более того, данное изобретение предлагает способ улучшения уровня не менее одного показателя крови за счет стабилизации клеточных мембран, который предусматривает введение животным, имеющим повреждение клеточных мембран, очищенной дрожжевой рибонуклеиновой кислоты в эффективном количестве и в фармакологически пригодных наполнителях, носителях и растворах. Более того, данное изобретение предлагает способ улучшения уровня не менее одного показателя крови, который предусматривает введение животным, требующим такого лечения, дрожжевой рибонуклеиновой кислоты в эффективном количестве и в фармакологически пригодных наполнителях, носителях и растворах для улучшения уровня не менее одного показателя крови. Показателем крови является любой, который представляет уровни лейкоцитов, эритроцитов, тромбоцитов, гемоглобина, нейтрофилов и гематокрита. Более того, данное изобретение предлагает способ предупреждения и лечения любой цитопении,анемии, тромбоцитопении, нейтропении, что предусматривает введение животным, которым необходимо такое лечение, дрожжевой рибонуклеиновой кислоты в эффективном количестве и в фармакологически пригодных наполнителях, носителях и растворах. Рибонуклеиновая кислота вводится в количествах около 0,1 мг-1,0 г на 1 кг веса млекопитающих,например в границах от 0,1 до 1,0 г, чаще от 250 до 350 мг. Вместе с тем данное изобретение предлагает соединение рибонуклеиновой кислоты, выделенной из дрожжей, например из Saccharomyces cerevisiae или из Candida utilis. Желательно, чтобы рибонуклеиновая кислота имела содержание азота более 14,5% от массы, количество фосфора более 8,5% от массы,более предпочтительно, чтобы количество азота было больше чем 14,7%, а количество фосфора больше чем 8,6% от массы, еще предпочтительней, чтобы количество азота было больше 15,0%, а количество фосфора больше 9,0% от массы.-7 010182 Более того, данное изобретение предлагает фармацевтическую композицию для лечения и предупреждения воспаления и связанных с воспалением заболеваний, что предусматривает введение животным очищенной дрожжевой рибонуклеиновой кислоты в фармакологически пригодных наполнителях,носителях и растворах. Детальное описание изобретения Проведено комплексное изучение известных нуклеиновых кислот с использованием разнообразных моделей in vitro и in vivo. Отобранные модели соответствовали определенному типу воспалительного процесса обычного или иммунологического происхождения. Изучение влияния рибонуклеиновой кислоты (РНК), а именно очищенной дрожжевой РНК, сравнивали с действием известных противовоспалительных лекарств в широком диапазоне противовоспалительной активности. Резюме экспериментальных моделей и результатов. 1. Модель агрегации тромбоцитов in vitro. Первичный скрининг препаратов экзогенных нуклеиновых кислот проводили in vitro на модели агрегации тромбоцитов человека, индуцированных арахидоновой кислотой (Born L.V.R. The aggregation ofblood platelets by difosfate and its reversal, Nature, v. 94, p. 327, 1962). Изучали экзогенные ДНК и РНК прокариот и эукариот. За стандарт брали широко известный противовоспалительный препарат аспирин. Показано, что аспирин угнетал агрегацию тромбоцитов, индуцированных арахидоновой кислотой до определенного уровня. Дезоксирибонуклеиновая кислота эритроцитов цыплят ДНК-ЭК фирмы"Reanal" (Венгрия) угнетала агрегацию тромбоцитов в границах действия аспирина. В то же время ДНК тимуса большого рогатого скота (ДНК-Т) и транспортная РНК E. coli (тРНК) производства "Serva"(США) угнетали агрегацию индуцированных тромбоцитов почти в два раза. Высочайший угнетающий эффект обнаружила суммарная РНК дрожжей, которая существенно угнетала агрегацию тромбоцитов в широком диапазоне концентраций. РНК-Ф, загрязненная белком, действует на треть слабее. Угнетение агрегации тромбоцитов дрожжевой РНК зависит от того, находится ли она в виде кислоты или в виде натриевой соли, от уровня ее чистоты и наличия белка. Натриевая соль дрожжевой РНК-ПН в высокой концентрации активна только на половину, а в низкой концентрации бездействует. Поскольку модель агрегации тромбоцитов, индуцированных арахидоновой кислотой, используется для отбора противовоспалительных препаратов, можно считать, что препараты нуклеиновых кислот, и в особенности дрожжевая РНК, обладают сильно выраженными противовоспалительными свойствами. 2. Модель кислотной резистентности мембран эритроцитов in vitro. Исходя из определения, что главным проявлением воспалительного процесса является нарушение мембраны клетки, применялась модель кислотной резистентности мембраны эритроцитов in vitro для скрининга мембранопротекторных и, таким образом, противовоспалительных свойств этих препаратов. Для изучения иммуностабилизирующего действия экзогенных нуклеиновых кислот выбрали эритроциты крыс. Изучали реакцию мембран эритроцитов на повреждающее действие нитрит аниона. С помощью определения кислотной резистентности эритроцитов кинетическим методом оценивали мембраностабилизирующее действие экзогенных нуклеиновых кислот и повреждающее действие экзогенного и эндогенного нитрит аниона (Terskov I.A., Hittelzon L.L. Chemical (acid) erythrogram method, Biophizika, 2(2): 259-266; 1957). Суть метода состоит в определении динамики изменения количества клеток, которые гемолизируют под действием слабой кислоты за определенный промежуток времени. Лизис эритроцитов в кислой среде включает три основные стадии: проникновение ионов водорода (протонов, H+) через плазматическую мембрану эритроцитов, протонированние гемоглобина и белка мембраны и, как следствие последнего, осмотическое разрушение эритроцитов. Используя этот метод, оценивали влияние экзогенных нуклеиновых кислот на кинетику прохождения протона через плазматическую мембрану эритроцитов, что зависит от состояния его нативности. Скорость проникновения протона в цитозоль клетки в большей мере зависит от состояния окисления как липидных (Kellogg E.W., Fridovich I. Liposome oxidation and erythrocyte lysis by enzymically generatedsuperoxide and hydrogen peroxide J. Biol. Chem. 10; 252(19): 6721-6728; 1977), так и белковых компонентов, в особенности белка полосы 3 окисления плазматической мембраны, и определяется как активностью [Н+]-АТФаз, так и разного типа обменников (Sato Y., Kamo S., Takahashi Т., Suzuki Y. Mechanismmembrane: properties, regulation, and functional role, Am. J. Physiol., 265(1 Pt. 1): p. 3-14; 1993). Кинетическим методом записывали кислотные эритрограммы. В опытах in vitro записывали кислотные эритрограммы в присутствии нитрита натрия (повреждающий фактор) и разных концентраций экзогенных нуклеиновых кислот. С помощью модели окислительного разрушения эритроцитов нитрит анионом в опытах in vitro показано стабилизирующее и мембранопротекторное действие экзогенных нуклеиновых кислот. На этой модели кислотной резистентности мембраны эритроцитов изучали набор тех же препаратов, что и на модели агрегации тромбоцитов. Показано, что препараты дрожжевой РНК в широком спектре концентраций обладают мембранопротекторными свойствами. Более детальное изучение показало, что мембранопротекторное действие-8 010182 дрожжевой РНК зависит от формы ее пребывания (кислота или соль), ее чистоты, наличия белка. Наиболее эффективной оказалась особо очищенная рибонуклеиновая кислота РНК-П, характер эритрограммы которой в концентрации 10 и 100 мкг был аналогичным норме. Натриевая соль дрожжевой РНК-ПН оказалась менее эффективной, в особенности в концентрации 10 мкг. Загрязнение дрожжевой РНК-Ф белком вообще приводило к потере мембраностабилизирующего действия. Другие препараты тРНК, ДНК-Т,ДНК-ЭК в изученных концентрациях приводили к дестабилизации мембраны эритроцитов, поэтому, несмотря на их некоторые противовоспалительные свойства на других моделях, их нельзя применять как противовоспалительные препараты. 3. Эритроцитная модель аутоиммунной реакции у крыс. Использовалась модель окислительного повреждения плазматических мембран эритроцитов для исследования мембраностабилизирующего действия экзогенных нуклеиновых кислот. Окислительное повреждение белковых и липидных компонентов плазматической мембраны эритроцитов имело место в опытах in vivo в процессе развития аутоиммунной реакции (адъювантный артрит), за которой значительно активировался биосинтез оксида азота, который является активным окислительным агентом, в особенности гемоглобина эритроцитов (Eich R.F., Li Т., Lemon D.D. Mechanism of NO-induced oxidation ofbleomycin-treated rats. Biochem.-Biophys. Res. Commun., 15; 182(1); 151-158; 1992; Kosaka H., Harada N.,Watanabe M. et al. Synergistic stimulation of nitric oxide hemoglobin production in rats by recombinant interleukin 1 and tumor necrosis factor. Biochem. Byophis. Res. Commun. 30; 189(1): 392-398; 1992). Оксид азота,так же как и пероксид водорода, играет критическую роль в повреждении клеток, в том числе клеток крови в процессе развития аутоиммунной реакции. Противовоспалительные цитокины (гамма-интерферон,IL-1) индуцируют экспрессию индуцибельной формы NO-синтетазы (iNOS). Исследовали изменения активности NOS в крови крыс в динамике развития аутоиммунной реакции(адъювантного артрита), для того чтобы оценить иммуномодулирующий эффект препарата, а также получить информацию о возможных уровнях одного из наиболее активных окислительных гемолитиков оксида азота (в виде его стабильного метаболита нитрит-аниона). Определяли активность ферментаNO-синтетазы (NOS), которая вырабатывает эндогенный нитрит-анион. Эти показатели характеризуют защитное действие экзогенных нуклеиновых кислот от разрушительного влияния нитрит аниона относительно мембраны эритроцитов. Особый интерес к изменениям стабильности эритроцитов в процессе развития аутоиммунных реакций с нашей стороны вызван большим количеством данных относительно наличия иммуномодуляторных свойств у эритроцитов (Karalnik B.V. Erythrocytes, their receptors, and immunity, Uspekhi Sovremennoy Biologii., v. 112,1, p. 52-61, 1992; Prokopenko L.H., Siplivaya L.E.,Erythrocytes as modulators of immunologic reactions, Uspekhi Phiziologicheskikh Nauk., v. 23,4, p. 89-106,1992), что дало возможность даже использовать понятие "эритроцитарная иммунная система". Развитие аутоиммунного процесса сопровождалось значительным снижением кислотной резистентности эритроцитов в ранний период и, наоборот, значительным превышением нормальной стабильности в завершающий период. Дрожжевая РНК повышала стабильность мембраны, нормализуя процесс транспорта протонов, который обусловливается состоянием белковых и липидных компонентов плазматической мембраны эритроцитов в ранний период, и поддерживала на стабильном уровне, близком к уровню нормы, в следующий период аутоиммунной реакции. Таким образом, показано, что с развитием аутоиммунного процесса активность NOS в крови крыс изменяется. Доказано увеличение активности NOS в крови крыс и в начальной и в конечной стадиях развития этого процесса. Дрожжевая РНК так уменьшает активность NOS, что в заключительной стадии процесса она практически в норме. Вместе с тем развитие аутоиммунного процесса сопровождается значительным снижением кислотной резистентности эритроцитов на ранней стадии и, наоборот, существенным избытком относительно нормы на завершающей стадии в сравнении со стойкостью нормальных эритроцитов. Дрожжевая РНК увеличивала стабильность мембран на начальной стадии процесса, нормализуя транспорт протонов, который зависит от состояния белковых и липидных компонентов эритроцитной плазматической мембраны, и поддерживая на стабильном уровне, близком к уровню нормы, в следующий период развития аутоиммунной реакции. С учетом вышесказанного защитные свойства дрожжевой РНК, как показано на модели аутоиммунного процесса, доказывают ее способность лечить не только аллергические болезни, но также и другие хронические воспалительные процессы, такие как артрит, атеросклероз, а также другие заболевания,включающие аутоиммунные реакции. 4. Модель отека у мышей, вызванного карагенином. Для отбора препаратов нуклеиновых кислот с противовоспалительным действием использовали известную модель воспалительного отека ножки мышей, вызванного подкожным введением карагенина. Вызванный карагенином отек чувствителен к действию препаратов, которые уменьшают капиллярную проницаемость.-9 010182 В начале карагенинзависимого антивоспалительного процесса важную роль играет кинин, а на поздних стадиях более важными являются протеолитические ферменты и простагландины. Карагениновая модель - это постепенный процесс, который дает достаточно времени для исследования биохимических механизмов противовоспалительного действия лекарства. Поэтому использовали данную модель для изучения влияния дрожжевой РНК на синтез тромбоксана и лейкотриена. Вместе с тем изучали влияние дрожжевой РНК на NO-синтетазную активность. Изучение противовоспалительного действия препаратов нуклеиновых кислот на карагениновой модели показало, что они имеют значительную противовоспалительную активность. Однако только дрожжевая РНК в концентрации 10 мг на мышку дает 50% угнетения отека. На мышах изучали дрожжевую РНК в концентрациях от 1 до 15 мг на мышку. В концентрациях ниже 1 мг на мышку препарат дрожжевой РНК не проявил видимой активности, вместе с тем в концентрации более 15 мг отек уменьшался на 53-55%. Более того, биохимические тесты показали стабилизирующее влияние дрожжевой РНК на активность NO-синтетазы, а также количество тромбоксанов и лейкотриенов, которое изменяется на протяжении образования отека. Аспирин, который изучали в рекомендованной терапевтической дозе 20 мг/кг, наоборот, значительно слабее влиял на увеличение отека и не показал стабилизирующего влияния на уровень биохимического метаболизма. 5. Модель острой ишемии у крыс. Дальнейшее изучение дрожжевой РНК выполняли на модели острой ишемии-реперфузии миокарда у крыс. Данная модель базируется на общих фундаментальных механизмах в развитии нарушений различных сердечных заболеваний, включающих нарушение структуры и функции мембран в эндотелиоцитах, кардиоцитах и других клетках сердца. Эти изменения приводят к деградации фосфолипидов мембраны и образованию очень эффективных биоактивных соединений, таких как лейкотриены или тромбоксаны, которые обладают сосудосужающим, аритмогенным, хемоактивным и проагрегантым действием (Bangham A.D., Hill M.W., Miller N. Preparation and use of liposom as model of biological membranes,Method in Membrane Biology, Acad. Press, v. 1, NY, p. 1-16, 1974). Как показали результаты изучения, дрожжевая РНК, введенная внутривенно крысам в концентрации 40 мг на крысу, нормализует функцию сердца при остром инфаркте. Это было показано в четко выраженном противоаритмическом действии препарата и значительном уменьшении зоны некроза в ишемизованном миокарде сердца. Лекарство также полностью нормализовало NO-синтетазную активность в крови и пограничной зоне ишемизованного сердца. Инъекция дрожжевой РНК нормализовала до обычного уровня количество арахидоновой кислоты в крови и сердце животных при остром инфаркте, а также полностью нормализовала уровень эйкозаноидов в крови крыс при ишемии. У животных, которых лечили дрожжевой РНК, активность миелопероксидазы, маркерного фермента нейтрофилов, который помогает оценить антиоксидантные свойства препарата, уменьшалась в два раза. Изучение активности дрожжевой РНК на модели ишемии-реперфузии позволило определить значительную стабилизирующую активность препарата на разных этапах воспалительного процесса в ишемизованном сердце, которая проявляется в продолжительном антиинфарктном действии и уменьшении размера инфарктной зоны в миокарде. На основе изучения активности дрожжевой РНК в ишемизовано-перфузийном сердце животных можно сделать вывод, что дрожжевая РНК обладает антиинфарктным действием или противовоспалительным действием при инфаркте, благодаря стабилизации структуры и функции мембран в эндотелиоцитах, кардиоцитах и других клетках сердца. 6. Влияние дрожжевой РНК на показатели крови. Изучали образцы крови, взятые у групп пациентов перед и после лечения препаратом дрожжевой РНК, определяя количество лейкоцитов [WBC], эритроцитов [RBC] и тромбоцитов [PLT] в 1 мкл крови,количество гемоглобина [HGB] в г/дл, нейтрофилов (NTP) и гематокрит [HCT] в процентах. Препарат дрожжевой РНК давали в виде капсул, в концентрации 250 мг дрожжевой РНК на капсулу или в супозитариях, в концентрации 1 г дрожжевой РНК на супозитарий. Для исследования влияния дрожжевой РНК отбирали группы относительно здоровых пациентов,спортсменов, больных раком и инфицированных HIV пациентов. Результаты изучения показали, что лечение дрожжевой РНК приводило к стабилизации или улучшению показателей крови. В частности, лечение больных раком и ВИЧ-инфицированных пациентов препаратом дрожжевой РНК приводило к стабильной нормализации цитопении. Экспериментальные методы и результаты исследований. Пример 1. Метод выделения дрожжевой РНК. Пример 1.1. Выделение РНК. Из Saccharomyces cerevisiae была получена РНК-Д, а из Candida utilis выделили РНК-П, РНК-ПН и РНК-Ф. Экстракцию дрожжевой РНК проводили при помощи 10-12% раствора хлористого натрия при температуре 100-110C. РНК отделяли от дрожжевых остатков, охлаждали до 0C и подкисляли соляной кислотой до pH 1-2. После выпадения в осадок РНК промывали этиловым спиртом, высушивали и растворяли в воде. Гидроокисью натрия раствор доводили до pH 8,0-8,2. К раствору добавляли панкреатин и- 10010182 выдерживали около 1 ч при температуре 37-40 С. Фермент инактивировали кипячением, после чего раствор фильтровали. РНК осаждали охлажденным этиловым спиртом, подкисленным соляной кислотой доpH 1-2 и высушивали. Таким образом была получена РНК-Ф. Далее осадок РНК фильтровали, промывали этиловым спиртом и растворяли в воде, прибавляя гидроокись натрия до pH 6,2-6,5. РНК-ПН осаждали этиловым спиртом, осадок фильтровали и сушили. РНК-П получали из РНК-Ф путем дополнительной очистки от белков при помощи повторной инкубации с панкреатином при 37-40C. Потом фермент инактивировали кипячением на протяжении 10 мин. Раствор РНК-П фильтровали и осаждали подкисленным до pH 1-2 спиртом. Осадок РНК-П фильтровали, промывали этиловым спиртом и сушили. Конечный продукт имел желто-серый цвет. Таблица 1 Химический анализ препаратов дрожжевой РНКN14,7%, P (тотальный) 8,6%, белок (биуретовая реакция) - отрицательно, ДНК (колометрично) - 2,0%,сахар (хроматографично) - отрицательно, полисахариды (биологическое определение) - отрицательно. Пример 1.2. Отсутствие токсичности. Установлено, что РНК-П и РНК-Д являются нетоксичными. Одинарное или многоразовое введение дрожжевой РНК внутрибрюшинно в биоактивных концентрациях (от 250 до 500 мг на 1 кг веса тела) не приводит к существенным изменениям количества периферийных лимфоцитов у мышей. Существенные изменения этого показателя свидетельствуют о наличии эндотоксинов. Аналогичные результаты получены при внутривенном введении нуклеиновых кислот. Определяли изменения количества периферических лейкоцитов у кроликов через 1-3 ч после внутривенной инъекции 100 мг раствора дрожжевых РНК-П или РНК-Д. В качестве контроля нетоксичности использовали внутривенную инъекцию 0,85% раствора NaCl. Показано, что аналогично стандарту инъекция дрожжевых РНК-П или РНК-Д через 3 ч после введения не вызывала изменения в количестве лейкоцитов. У животных, которые получили 0,85% раствор NaCl, количество лейкоцитов отвечало 13000980, тогда как животные, получившие РНК-П или РНК-Д, показали соответственно 12700850 и 12900980 лейкоцитов,что является нормой. Если кролики получали инъекцию 10 мг полисахаридов протея, количество лейкоцитов через 1 ч уменьшалось с 130501100 до 29001210 и оставалось на этом уровне на протяжении 3 ч. Полученные результаты свидетельствуют о нетоксичности дрожжевой РНК. Более того, при введении кроликам внутривенно 100 мг дрожжевой РНК-П или РНК-Д на 1 кг веса тела у животных не определялся острофазный С-реактивный белок, являющийся свидетельством эндотоксичного действия. В частности, дрожжевая РНК является непирогенным соединением, как это было показано на кроликах. В группе кроликов на протяжении двух дней 4 раза в день с 2-часовым интервалом изучали температуру тела. На третий день кроликам ввели инъекцию 0,85% раствор NaCl и продолжали определять температуру тела через 1, 2 и 3 ч после инъекции. На шестой день кроликов разделили на 3 группы,две из которых получили внутривенно 100 мг РНК-П и РНК-Д соответственно. Снова у животных определяли температуру тела. Контрольные животные показали температурные колебания от 0,1 до 0,4C. Подопытные животные, которые получали препараты РНК, имели изменения темпаратуры в тех же границах, что и контрольные: от 0,1 до 0,4C. Эти результаты подтверждают непирогенность дрожжевой РНК. Пример 2. Изучение противовоспалительного действия препаратов нуклеиновых кислот на модели агрегации тромбоцитов in vitro. Противовоспалительное действие препаратов нуклеиновых кислот изучали на модели агрегации тромбоцитов in vitro методом Born (Born L.V.R. The aggregation of blood platelets by diphosphate and itsreversal, Nature, v. 94, p. 327, 1962). Венозную кровь людей собирали в силиконовые пробирки фирмы(Becton Dickson), которые содержали 3,8% раствор цитрата натрия. Для того чтобы получить плазму,богатую тромбоцитами, цитратную кровь центрифугировали при 1500 об/мин в течение 7 мин. Безтромбоцитную плазму получали путем центрифугированния в течение 15 мин при 3000 об/мин отобранных 2,0 мл из средних пластов плазмы. В тромбоцитарной плазме подсчитывали количество тромбоцитов,потом они разводились бестромбоцитарной плазмой к конечной концентрации 200,0-300,0109/л. Агрегацию тромбоцитов проводили при помощи агрегометра фирмы "Tromlite" (Польша). Для индукции агрегации использовали раствор арахидоновой кислоты фирмы ICN (США), которую растворяли в буфере Михаэлиса из расчета 1 мг/мл. В агрегометр помещали две пробирки: одну с 0,2 мл тромбоцитарной плазмы и другую с 0,2 мл бестромбоцитарной плазмы и 0,1 мл изотонического раствора хлорида- 11010182 натрия. После включения прибора в пробирку с тромбоцитарной плазмой добавляли 0,1 мл арахидоновой кислоты. Потом в течение 5 мин регистрировали светопропускающую способность тромбоцитарной плазмы, что отвечало степени агрегации тромбоцитов. В исследовательских вариантах при изучении действия препаратов нуклеиновых кислот на агрегацию тромбоцитов перед началом измерения раствор тромбоцитарной сыворотки подвергали предварительной инкубации 5 мин при 37C с 0,1 мл раствора соответствующей концентрации препарата. В параллельную пробирку с бестромбоцитарной плазмой, соответственно, добавляли 0,2 мл изотонического раствора хлорида натрия. После окончания инкубации включали прибор и в пробирку с тромбоцитарной плазмой и препаратом, добавляли 0,1 мл арахидоновой кислоты и через 5 мин заканчивали измерение,определяя конечную степень агрегации тромбоцитов. Параметром агрегации был выбран индекс агрегации клеток IA, который равнялся где D1 - оптическая плотность обогащенной тромбоцитами плазмы с арахидоновой кислотой как индуктором агрегации тромбоцитов;D2 - оптическая плотность проинкубированной с препаратом нуклеиновой кислоты, обогащенной тромбоцитами плазмы с арахидоновой кислотой как индуктором агрегации тромбоцитов. Статистическую обработку результатов проводили, используя критерий Стьюдента и программное обеспечение, описанное в примере 4.1. Исследовали следующие препараты нуклеиновых кислот: ДНК-Т, ДНК-ЭК, тРНК и суммарную РНК-Д дрожжей в конечной концентрации 110-2%. За стандарт противовоспалительного препарата использовали аспирин в концентрации 0,06 мг на пробирку с обогащенной тромбоцитами плазмой. Результаты исследования представлены в табл. 2. Таблица 2 Влияние нуклеиновых кислот и аспирина на агрегацию тромбоцитов,индуцированную арахидоновой кислотой Результаты опытов показали, что препараты нуклеиновых кислот в концентрации 110-2% угнетают агрегацию тромбоцитов, индуцированных арахидоновой кислотой. Показано, что дрожжевая РНК-Д в концентрации 110-2% почти в два раза сильнее угнетает агрегацию индуцированных тромбоцитов, чем аспирин (38,6%), соответственно дрожжевая РНК 59,7%, а транспортная РНК E. coli 52,23%. Препарат ДНК-ЭК эритроцитов цыплят (36,93%) действовал на уровне аспирина, а препарат ДНК-Т тимуса большого рогатого скота угнетал агрегацию тромбоцитов на 54,45%, что почти на уровне дрожжевой РНК. Поскольку в препаратах ДНК всегда есть большой процент РНК, можно предположить, что выявленный угнетающий эффект ДНК присутствует за счет имеющейся в ней РНК. Кроме того, изучение влияния разных концентраций дрожжевой РНК на агрегацию индуцированных тромбоцитов, представленное в табл. 3, показало, что она действует в широком диапазоне концентраций от 0,1 до 110-5%, соответственно угнетая агрегацию от 78,5 до 14,2%. Таблица 3 Зависимое от концентрации влияние дрожжевой РНК-Д на агрегацию тромбоцитов,индуцированную арахидоновой кислотой Далее в табл. 4 показано, что эффект угнетения агрегации зависит от чистоты препарата РНК и ее натриевой соли. В табл. 4 показано, что РНК-Ф с примесями белка и меньшим содержанием азота и фосфора действовала значительно слабее в диапазоне концентраций от 110-1 до 110-3%. Например, в наивысшей концентрации препарат РНК-Ф угнетал агрегацию тромбоцитов на 57%, а в наиболее низкой концентрации на 22,7%. В то же время хорошо очищенная РНК-П угнетала агрегацию тормбоцитов на треть сильнее,соответственно на 84 и 29,7%. Но наиболее радикально антиагрегантные свойства РНК теряет при ее переведении в натриевую соль. Этот препарат РНК-ФН в наивысшей концентрации действовал в два раза слабее (44,4%), а в наиболее низкой концентрации вообще не обладал антиагрегантными свойствами. Ведь на модели агрегации тромбоцитов, индуцированных арахидоновой кислотой показано, что препараты РНК, и в особенности очищенные РНК дрожжей, владеют хорошо выраженными антиагрегантными свойствами в широком спектре концентраций, что свидетельствует об их антивоспалительном действии. Пример 3. Противовоспалительное действие нуклеиновых кислот, изученное на модели стабилизации мембран эритроцитов in vitro. Мембраностабилизирующее и антирадикальное действие препаратов нуклеиновых кислот оценивали на эритроцитах крыс в опытах in vitro. Инициировали повреждения плазматической мембраны эритроцитов нитрит-анионом - стабильным метаболитом оксида азота, который вызывает окислительное повреждение белковых (особенно гемоглобина) и липидных компонентов мембран. С целью оценки эффективности мембраностабилизирующего от повреждения свободными радикалами действия препаратов нуклеиновых кислот использовали определение кислотной резистентности отмытых от плазмы эритроцитов крови нормальных крыс. Эритроциты трижды отмывали холодным(4C) раствором 0,15 М NaCl и удаляли пласт лейкоцитов и тромбоцитов. Кислотный лизис отмытых эритроцитов в суспензии (10 мкл) индуцировали нитритом натрия в постоянной концентрации (250 мкг на 1 мл) для инициации окислительного повреждения эритроцитов. Суспензия состояла из разбавленных к определенной плотности эритроцитов (0,7106 клеток на 1 мл изоосмотической среды) с 0,14 М NaCl и 0,01 М цитратно-фосфатного буфера pH 2,5 и из разных концентраций нуклеиновых кислот(10 или 100 мкг). Лизис эритроцитов инициировали внесением 1 мл 0,004N HCl и записывали изменение экстинции при 750 нМ. Расчеты делали, как показано в примере 6.3. Показано, что дрожжевая РНК-Д в дозах 10 и 100 мкг увеличивала значения суммарной стойкости эритроцитов от 288 единиц (контрольное значение в присутствии NaNO2 без препарата) до 449 (концентрация дрожжевой РНК 10 мкг) и 437 единиц (концентрация дрожжевой РНК 100 мкг), что было близко к норме (475 единиц). РНК-ПН повышала суммарную стойкость до 328 единиц в дозе 10 мкг и до 415 единиц в дозе 100 мкг. РНК-П повышала суммарную стойкость до 315 единиц в дозе 10 мкг и до 462 единицы в дозе 100 мкг (максимально приближенная к нормальным значениям величина этого параметра). Препарат РНК-Ф повышал суммарную стойкость до 338 единиц в дозе 10 мкг и, наоборот, несколько снижал (до 271 единиц) в дозе 100 мкг. Препарат ДНК-Т повышал суммарную стойкость до 338 единиц в дозе 10 мкг и до 654 единиц в дозе 50 мкг, что более чем в 2 раза выше контрольных значений и даже выше уровня нормы (без повреждения NaNO2). Однако в дозе 100 мкг эффект его был обратным - мембранодестабилизирующим, о чем свидетельствует снижение суммарной стойкости до 158 единиц, что почти вдвое ниже уровня контроля. Препарат ДНК-ЭК в дозе 100 мкг не изменял кислотную резистентность эритроцитов в рассматриваемой модели окислительного повреждения. Тогда как в дозе 10 мкг он повышал ее до 408 единиц, что лишь незначительно ниже выявленного протекторного действия для РНК-Д (449 единиц для дозы 10 мкг).- 13010182 Таким образом, экзогенная ДНК, независимо от происхождения, имеет значительное антистабилизирующее действие на клеточную мембрану. Поскольку она разрушает клеточную мембрану, она не может использоваться как лекарство или как пищевая добавка. Препарат тРНК в обоих исследованных дозах (10 мкг до 279 единиц и 100 мкг до 296 единиц) не менял кислотную резистентность эритроцитов. Проведенные исследования показали, что дрожжевая РНК, в зависимости от формы, происхождения и чистоты, в опытах in vitro, показала мембраностабилизирующие и антирадикальные свойства. Наиболее эффективным препаратом оказалась хорошо очищенная РНК-Ф, противовоспалительные свойства которой были изучены более детально. Пример 4. Изучение противовоспалительного действия нуклеиновых кислот на модели локального воспалительного процесса, вызванного карагенином (LPS). Пример 4.1. Влияние дрожжевой РНК на отек в модели локального воспаления, вызванного карагенином (LPS) in vivo. Для изучения противовоспалительного действия препаратов использовали модель местного воспаления у мышей. Воспаление у мышей линии BALB моделировали при помощи классического флогогенного агента - карагенина. За 30 мин к инъекции карагенина мышам внутрибрюшинно вводили препарат в 2 мл физиологического раствора (PS). Карагенин (LPS) производства фирмы Serva Fein Biochemica (Германия) готовили в виде 1% раствора в PS. Полученный вяжущий раствор вводили мышам субплантарно в заднюю левую лапку в объеме 40 мкл. Правая лапка (интактна) служила контролем. Через 4 ч после введения карагенина мышей забивали путем декапитации и отрезали обе задние лапки на одном уровне,немного выше голеностопного сустава. После этого их каждую отдельно взвешивали с точностью до 1 мг. Полученные результаты были обработаны на компьютере при помощи статистического анализа в программе MultiFac2.2. SPSS 8/0 Противовоспалительное действие препарата рассчитывали по формуле где Vk - среднее увеличение объема (массы) отека лапки в контроле;Vo - среднее увеличение объема (массы) отека лапки мышей, которых лечили. В первом опыте животных разделили на восемь групп. Первая группа состояла из контрольных животных, которым внутрибрюшинно вводили по 2 мл физиологического раствора, а в левую лапку инъекцией вводили также 40 мкл физиологического раствора. Эта группа представляла по сути интактных мышей, на которых определяли влияние самой травмы (инъекция без флогогенного агента) на воспалительный процесс в левой лапке. Вторая группа мышей - это контроль с карагенином, которым внутрибрюшинно по 2 мл вводили физиологический раствор, а в левую лапку - карагенин. Третья группа мышей получала внутрибрюшинно по 0,4 мг аспирина в 2 мл физиологического раствора на животное, а в левую лапку - карагенин. Четвертая, пятая и шестая группы мышей получали внутрибрюшинно растворенную в физиологическом растворе дрожжевую РНК в концентрации по 5, 10, и 15 мг в 2 мл физиологического раствора на животное. Карагенин вводили в левую лапку для провоцирования отека. Седьмая и восьмая группы мышей лечились, соответственно, ДНК-Т и ДНК-ЭК, которые вводили инъекцией в концентрации по 15 мг на мышку, как это описано ниже для групп, которых лечили РНК. Правые лапки мышей были интактны. Обе правые и левые лапки были отделены от тела и потом в каждой группе мышей определяли их массу. Результаты опытов по противовоспалительному действию нуклеиновых кислот представлены в табл. 5.- 14010182 Таблица 5 Влияние нуклеиновых кислот на локальное воспаление лапок мышей Как показано в табл. 5, аспирин во введенных концентрациях уменьшает развитие отека на лапках мышей на 18,02%. Эти данные согласовываются с результатами, полученными в других роботах на данной модели, и свидетельствуют о соответствии данной модели воспалительного процесса. Концентрации аспирина также отвечают дозе 20 мг/кг, которая теперь рекомендуется для клинического использования. Кроме того, данная доза имеет некоторые отрицательные последствия при продолжительном использовании в разных формах воспалительных процессов. Кроме того, препарат дрожжевой РНК-Д обнаружил значительную противовоспалительную активность, которая имеет концентрационную зависимость. В концентрации 5, 10, и 15 мг на мышку препарат угнетает образование отека на 36,74; 47,17 и 53,13% соответственно. Препарат ДНК-Т и ДНК-ЭК также обнаружил противовоспалительную активность, хотя в довольно высоких концентрациях (15 мг на мышку), тем не менее, показатели противовоспалительного действия были в два раза ниже (соответственно 26,58 и 33,27%). На основе полученных результатов можно сделать вывод, что нуклеиновые кислоты имеют довольно выраженные противовоспалительные свойства сравнительно с аспирином. Однако дрожжевая РНК обнаружила наиболее значительную противовоспалительную активность. Пример 4.2. Влияние дрожжевой РНК на биохимические показатели на модели локального воспаления, вызванного карагенином (LPS) in vitro. Противовоспалительное действие дрожжевой РНК сравнивали с действием аспирина в процессе развития воспалительной реакции мышей (на 0-, 30-, 60-, 320-й мин) после инъекции LPS. Определяли влияние дрожжевой РНК на активность фермента NO-синтетазы (NOS) в плазме крови и эритроцитах, а также на количество в плазме крови арахидоновой кислоты и продуктов ее оксидного метаболизма, которые образуются вследствие липоксигеназного (лейкотриен C4 (LTC4 и циклогеназного (тромбоксан 2(TX2 путей. Пример 4.2.1. Действие дрожжевой РНК на активность NO-синтетазы. Активность фермента NO-синтетазы в плазме крови и эритроцитах определяли колориметричным методом по продукту реакции - нитрит-аниону (Yan L., Vandivier R.W., Suffredini A.F., Danner R.L.,Human polymorphonuclear leukocytes lack detectable nitric oxide synthetase activity. J. Immunol., 15; 153(4): 1825-1834; 1994). Инкубационная смесь (1 мл) состояла из 50 мМ HEPES (pH 7,4),1,25 мМ CaCl2, 1 мМ NADPH, 80 мкМ ФАД, 20 мкМ тетрагидробиоптерина, 13 мкг/мл кальмодулина, 1 мМ L-аргинина, 60 мМ L-валина, 100 единиц/мл супероксиддисмутазы.HEPES - это N(2 гидроксиэтил)-1-пиперазинэтансульфоновая кислота от Sigma Chemical Co. (USA),NADPH - это бета-никотинамидадениндинуклеотид фосфат в восстановленной форме от SigmaChemical Co. (USA). Реакцию запускали, добавляя 0,1 мл пробы, которая содержала около 500 мкг общего белка, который предварительно определяли по Бредфорду. Инкубацию проводили при 37C в течение 60 мин. Реакцию останавливали добавлением 0,2 мл 2N HCl4. Смесь центрифугировали при 10000 g в течение 10 мин и в надосадочной жидкости определяли содержание нитрит-аниона (стабильного метаболита оксида азота). Нитрит-анион определяли с помощью реактива Гриса в колориметрической реакции, которая описана Green L.C., Waagner D.A., Glogowski J. et al., Analysis of nitrate, nitrite and [15N] nitrate in biologicalfluids, Anal. Biochem., 126(1): 131-138; 1982. Реактив Гриса готовили, смешивая равные части 0,1% водного раствора нафтилэтилендиамингидрохлорида с 1% раствором сульфаниламина в 5% Н 3 РО 4 непосредственно перед определением. Определение проводили в безбелковых аликвотах проб, добавляя реактив Гриса в соотношении 1:1. Определяли величину экстинции при 543 нм через 5 мин после смешивания. Количество NO2- определяли по калибровочной кривой, построенной для NaNO2. Результаты определения представлены в табл. 6.- 15010182 Таблица 6 Влияние дрожжевой РНК и аспирина на активность NO-синтетазы (NOS) в плазме крови мышей после инъекции карагенина (LPS) (в пикомолях через 1 мин на 1 мг белка; Mm; n=5)P1 - достоверное отличие относительно нормы (перед LPS инъекцией); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Из табл. 6 видно, что после введения LPS без предшествующего введения дрожжевой РНК (контроль) наблюдали резкое (большее чем в 10 раз) возрастание активности NOS в плазме крови за 30 мин. После чего происходило снижение активности фермента с последующим незначительным повышением(но на уровне, значительно большем, чем нормальный уровень) его активности. Предшествующее инъекционное введение мышам дрожжевой РНК существенно уменьшало повышение активности NOS в плазме крови в ранний период (30 и 60 мин) развития воспалительной реакции. Этот протекторный эффект дрожжевой РНК уже не наблюдался на 320-й минуте развития воспалительной реакции, в то время как введение аспирина угнетало активность NOS именно в этот период. Таким образом, дрожжевая РНК совершает выраженное ингибиторное действие на активацию окислительного пути метаболизма L-аргинина при введении LPS, что выражается в ингибировании активности NOS в плазме крови. Поскольку существуют разные изоформы NOS, как конститутивные, так и индуцибельные, в разных ядерных клетках плазмы крови - нейтрофилах, тромбоцитах, лимфоцитах, макрофагах (Hibbs J.B.,Taintor R.R., Vavrin Z., Rachlin Е.М., Nitric oxide: a cytotoxic activated macrophage effector molecule.messenger RNA-expression and nitric oxid production by lipopolysaccharide in vivo: the role of macrophage,endogenous IFN-gamma and TNF receptor-1-mediated signaling. J. Immunol. 15; 158(2): 905-912; 1997), можно предположить, что в ранний период после введения LPS (30-60 мин) имеет место активация конститутивных изоформ (нейрональной или эндотелиальной NOS), а в более поздний период (320 мин), возможно, активируется также и индуцибельная изоформа NOS (iNOS) макрофагов крови. Дальше в табл. 7 показана динамика изменения активности NOS в эритроцитах крови мышей после введения LPS. У контрольных животных наблюдали незначительное повышение активности фермента на 30-й мин, после которого наблюдали значительное (почти в 2 раза) уменьшение NOS в эритроцитах. Таблица 7 Влияние дрожжевой РНК и аспирина на активность NOS в эритроцитах мышей после инъекции карагенина (в пикомолях за 1 мин на 1 мг белка;P1 - достоверное отличие относительно нормы (перед инъекцией карагенина); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Такая же, но значительно более выраженная динамика изменения активности NOS наблюдалась в эритроцитах мышей, которым предварительно вводили дрожжевую РНК. Так, на 30- и 60-й минутах наблюдали повышение активности NOS (соответственно более чем в 3 и 2 раза). На 320-й минуте действияLPS наблюдалось достоверное (более чем в 3 раза) снижение активности NOS в эритроцитах Некоторые авторы (Chen L.Y., Mehta J.L. Evidence for the presence of L-arginine-nitric oxide pathwayin human red blood cells: relevance in the effects of red blood cells on plateled function, J. Cardiovasc. Pharmacol. 32(1): 57-61; 1998) отмечают, что в эритроцитах присутствует конститутивная, Ca-зависимая изоформа NOS. Исходя из этого, возможно повышение активности эритроцитарной NOS в ранний период развития воспалительной реакции, вызванной введением LPS, обусловленное повышением внутриклеточного кальция в красных кровяных клетках. Пример 4.2.2. Влияние дрожжевой РНК на окислительный метаболизм арахидоновой кислоты. Содержание свободной арахидоновой кислоты определяли методом тонкослойной хроматографии,как описано в Tsunamoto et al. (Tsunamoto K., Todo S., Imashuku S. Separation of prostaglandines and thromboxane by two-dimensional thin-layer chromatography. J. Chromatog. 3; 417(2); 414-419; 1987). Содержание стабильного метаболита тромбоксана A2-TXB2 определяли в пробах с помощью радиоиммунного метода с применением подборки реактивов фирмы "Amersham" - TX2[3H] RIA Kit (Amersham International PLCChem., 27(8): 1417-1420, 1981). Содержание LTC4 определяли в пробах с помощью радиоиммунного метода с применением подборки реактивов фирмы "Du Pont" - LTC4 [3H] RIA Kit (Du Pont Ltd. Hertfordshire,(UK (Levine L., Morgan R.A., Levis R.A. et al., Radioimmunoassay of the leukotrienes of slow reactivity substance of anaphylaxis. Proc. Natl. Acad. Sci. USA. 78(12): 7692-7696; 1981). В табл. 8 показана динамика изменения содержания свободной арахидоновой кислоты в плазме крови мышей после введения инъекции LPS. Таблица 8 Влияние дрожжевой РНК и аспирина на содержание свободной арахидоновой кислоты в плазме крови мышей после инъекции карагенинаP1 - достоверное отличие относительно нормы (перед инъекцией карагенина); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Как показано в табл. 8, в группе контрольных животных уровни свободной арахидоновой кислоты(AA) возрастали лишь на 320-й минуте после введения LPS. Дрожжевая РНК достоверно снижала содержимое AA в плазме на 60-й минуте действия LPS. Уменьшение на 30-й минуте было недостоверным. На 320-й минуте действия LPS дрожжевая РНК достоверно уменьшала содержимое AA в плазме крови относительно контроля. Как известно, свободная арахидоновая кислота получается при гидролизе фосфолипидов мембран фосфолипазой A2, что активируется при повышении уровней свободного ионизированного кальцияfrom macrophage and reduce the calcium requireement for activity. Biochem. Biophys. Acta. 6; 1045(3), 261270; 1990). Кроме того, возможны и другие пути освобождения свободной AA, например при гидролизе эфиров холестерола холестеролэстеразой (Moscat J., Moreno F., Herrero C., et al., Arachidonic acid releasingsystems in pig aorta endothelial cells, Biochem. Biophys. Res. Commun. 30; 139(3): 1098-1103; 1986). Для воспалительного процесса более известен первый путь образования свободной арахидоновой кислоты и,таким образом, полученные данные свидетельствуют о возможном ингибировании дрожжевой РНК активности фосфолипазы в плазме крови. Далее, в табл. 9 показано влияние дрожжевой РНК на содержимое тромбоксана В 2, стабильного метаболита тромбоксана A2, который получается при окислительном циклооксигеназном пути метаболизма арахидоновой кислоты.- 17010182 Таблица 9 Влияние дрожжевой РНК и аспирина на содержание тромбоксана в плазме крови мышей после инъекции карагенина (в пикомолях на 1 мг белка; Mm; n=5)P1 - достоверное отличие относительно нормы (перед инъекцией карагенина); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Как показано в табл. 9, после введения LPS наблюдали значительное возрастание пулов ТХВ 2 в плазме мышей на 30-й и, в особенности, на 60-й минуте. Дрожжевая РНК подобно аспирину, известному ингибитору циклооксигеназного пути метаболизма арахидоновой кислоты (циклооксигеназы и тромбоксансинтетазы), интенсифицировала снижение уровня ТХВ 2 после его значительного повышения в ранний период инициации воспалительного процесса. В табл. 10 показана динамика изменения содержимого одного из метаболитов липоксигеназного пути окисления AA - пептидолейкотриена C4 в плазме мышей после введения LPS. Таблица 10 Влияние дрожжевой РНК и аспирина на содержимое лейкотриена C4 в плазме крови мышей после инъекции карагенину (в пикомолях на 1 мг белка; Mm; n=5)P1 - достоверное отличие относительно нормы (перед инъекцией карагенина); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Как показано в табл. 10, у контрольных животных наблюдали возрастание содержимого LTC4 в интервале 30-60 мин с незначительным снижением против максимального уровня на 320-й минуте. У животных, которые получали дрожжевую РНК, уровни LTC4 были ниже контрольных на 30- и 320-й минуте действия LPS. Аспирин совершал аналогичное ингибирующее действие на 320-й минуте воздействия LPS, которое было более значительным, чем в случае дрожжевой РНК. Таким образом, полученные данные указывают на то, что дрожжевая РНК не только ингибирует генерацию свободной арахидоновой кислоты после введения LPS, но также ее окисление по двум альтернативным путям - как по липоксигеназному, так и циклооксигеназному. Пример 5. Противовоспалительное действие дрожжевой РНК на модели ишемии-реперфузии у крыс. Пример 5.1. Кардиопротекторное действие дрожжевой РНК. Исследования проводили на 13 белых крысах массой 200-250 г под уретановым наркозом 1,25 г/кг внутрибрюшинно (Kogan A.H. Modeling the myocardial infarction, М., 1979). Крысам накладывалась трахеостома, в которую вводили инкубационную трубку. Искусственную вентиляцию легких проводили при помощи аппарата "Вита 1". Разрез кожи и других тканей в межреберных мышцах проводили, отступая 2-3 мм влево от средней стернальной линии, начиная от яремной вырезки к мечевидному отростку длиной 4,0-4,5 см. Глазными ножницами рассекали нижнюю часть 2-, 3- и 4-го ребра и межреберную мышцу между 3 и 4 ребрами. В пробеле между ушком левого предсердия и пульмональным конусом обычно размещался начальный отдел левой коронарной артерии. На уровне левого нижнего угла пульмонального конуса, ориентируясь по ходу иногда видного начального отдела артерии, прошивали атравматичной иглой 3/0 ленту миокарда шириной 1,5-2,0 мм на глубину 1,0-1,5 мм. Выведенную лигатуру завязывали вокруг артерии и мускульного пучка, в котором- 18010182 она находилась. После чего наблюдали появление макропризнаков ишемии и начало развития инфаркта. Они проявлялись в выраженном осветлении ткани в течение 20-30 с начала ишемии (в особенности возле верхушки сердца), которое местами изменялось тотальным посинением (цианозом) и ослабленном сокращении зоны окклюзии и ее дилятацией. Регистрация ЭКГ в одном стандартном определенном месте от конечностей велась беспрерывно в течение 30 мин ишемии и 60 мин реперфузии. Инъекции дрожжевой РНК и аспирина вводили за 30 мин до начала ишемии в дозах 200 и 20 мкг/кг соответственно. Для определения площади и объема постинфарктного рубца парафиновые срезы миокарда крыс окрашивали р-нитроголубым тетразолиевым азокарминовым методом (Mueller В., Maass В., Krause W.,Witt W. Limitation of myocardial unperfused area and necrotic zone 24 hours and 7 days after coronary arteryligation in rats by the stable prostacyclin analogue iloprost, Prostaglandins Leucot. Med. 21(3): 331-340; 1986). После реперфузии животные были гепаринизованы (150 IU/кг i.v.), сердце подвергали глубокой эфирной анестезии и ретроградной реперфузии раствором 0,05% р-нитроголубого тетразолия в фосфатном буфере(30 мин; 100 мм Hg; 37C). После 24-часовой фиксации в растворе формальдегида желудочки взвешивали, рассекали накрест на 5 кусочков каждый, неокрашенную часть миокарда отделяли от окрашенной и взвешивали. Таким образом определяли зону некроза. Изучение зоны некроза через 60 мин после ишемии свидетельствует, что зона риска в левом желудочке сердца равнялась 33,33,4% массы левого желудочка. В контрольной группе зона инфаркта составляла 60,33,8% от зоны риска. Инъекция дрожжевой РНК за 30 мин перед началом инфаркта уменьшала пропорцию между зоной инфаркта и зоной риска до 41,02,5%. Таким образом, инъекция дрожжевой РНК на 32% уменьшала отрицательное влияние инфаркта на зону некроза левого желудочка сердца. При анализе ЭКГ было установлено, что предшествующее введение крысам дрожжевой РНК уменьшает количество экстрасистол. Только у одного из пяти животных этой группы было выявлено 4 экстрасистолы. В контрольной группе, которую составляли животные, не получавшие дрожжевую РНК, у трех крыс были зарегистрированы экстрасистолы, что составляло в среднем 8,71,7 экстрасистол на группу. Эпизоды пароксимальной тахикардии в контрольной серии были более продолжительными: у двух из пяти животных они продолжались в среднем 4,21,3 с. У пяти животных, которым вводили дрожжевую РНК, только в одном случае был выявлен участок пароксимальной тахикардии протяжностью 1,5 с. Анализ данных ЭКГ показал, что дрожжевая РНК содействует улучшению работы сердца при ишемии-реперфузии миокарда, стабилизируя работу ведущей системы и проявляя выраженное антиритмическое действие сердца, уменьшая количество экстрасистол и интервалы пароксимальной тахикардии. Таким образом, полученные результаты показывают, что дрожжевая РНК имеет выраженное кардиопротекторное действие при инфаркте миокарда крыс. Пример 5.2. Влияние дрожжевой РНК на активность миелопероксидазы в ишемизированом участке миокарда крыс. Миелопероксидазную активность (МПО) в миокарде определяли по методу Bradley et al.inflammatory cell infiltration in rat cardial tissue, J. Pharmacol. Methods, 20(3): 225-235, 1988). Для этого сердце быстро вынимали и промывали в охлажденном до 0C физиологическом растворе. После промывания в центральной зоне ишемии вырезали кусочек миокарда (0,1 г ткани) и замораживали его при-30C. Суммарную фракцию готовили как 10% гомогенат с экстрагирующим буфером, содержащим 0,5% гексадецилтриметил аммония бромид, растворенный в 50 ммоль/л калий-фосфатном буфере (pH 6,0) при комнатной температуре. После этого центрифугировали 20 мин при температуре 4C и 12000 g. Верхнюю фракцию (30 мкл) отбирали для реакции с 0,167 мг/мл O-дианизидина в 50 мМ/л калий-фосфатного буфера (pH 6,0). Реакцию запускали добавлением 0,005% раствора Н 2 О 2. Реакцию беспрерывно тестировали при длине волны 460 нм в течение 5 мин, снимая показатели каждую минуту. По полученным результатам строили графики. Единица МПО определялась количеством фермента, который разрушал 1 мкмоль/мин Н 2 О 2 при 25C. Данные выражались значением МПО на 1 г ткани. Ишемия и реперфузия миокарда вызывают острый воспалительный ответ, в котором центральное место занимают нейтрофилы (Entman M.L., Smith C.W. Postreperfusion inflammation: a model for reactionto injury in cardiovascular disease, Cardiovasc. Res. 28(9): 1301-1311, 1994). Установлено, что реперфузия ишемизированного миокарда сопровождается интенсивным накоплением нейтрофилов в границах зоны риска (Hearse D.J., Bolli R. Reperfusion induced injury: manifestation, mechanisms and clinical relevance. Cardiovasc. Res. 26(2): 101-108; 1992), нейтрофилы высвобождают разные медиаторы воспаления, такие как радикалы кислорода, цитокины и хемокины, и увеличивают ишемически-реперфузное повреждение миокарда (Entman M.L., Michael L., Rossen R.D., et al. Inflammation in the course of early myocardial ischemia,FASEB J., 5(11): 2529-2537; 1991). Установлена прямая зависимость между интенсивностью накопления нейтрофилов в ишемизированном миокарде и активностью миелопероксидазы - специфического фермента, который есть в нейтрофилах. Повышение МПА прямо коррелирует с количеством мигрирующих- 19010182 в зону воспаления лейкоцитов. Анализ активности миелопероксидазы в ишемизованной части миокарда после 30 мин окклюзии и 60 мин реперфузии левой коронарной артерии у крыс показал, что в контрольной группе она равняется 211,8+16,7 единиц на 1 г ткани. Когда животным вводили инъекцию аспирина, активность уменьшалась до 176,1+5,9 единиц на 1 г ткани. Инъекция РНК-Д уменьшала активность данного фермента на треть, до 152,3+9,8 единиц на 1 г ткани. Таким образом, введение крысам в вену дрожжевой РНК в дозе 200 мкг/кг за 30 мин перед ишемией уменьшает накопление нейтрофилов в зоне риска после реперфузии в течение 1 ч. Количество нейтрофилов в зоне риска уменьшалось приблизительно на 30%, что в два раза больше, чем при использовании аспирина (20 мкг/кг). Это дает возможность утверждать, что дрожжевая РНК может эффективно использоваться как кардиопротектор при ишемии и реперфузии миокарда. Пример 5.3. Влияние дрожжевой РНК на активность NOS при ишемии. Опыты проводили на крысах с экспериментальным инфарктом миокарда, который вызывали окклюзией коронарной артерии в течение 30 мин. Отбирали кровь из коронарной артерии и сердца животных, которые делили на интактную зону, пограничную зону и зону инфаркта. Определяли активность фермента NOS в разных зонах сердца и в крови, а также содержимое свободной арахидоновой кислоты(сердце и кровь) и продуктов ее окислительного метаболизма (кровь). Результаты определения представлены в табл. 11. Таблица 11 Влияние дрожжевой РНК на активность NOS при ишемии сердца крыс: (в пикомолях на 1 мг белка; Mm; n=5)P1 - достоверное отличие относительно нормы (перед ишемией); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Данные, представленные в табл. 11, свидетельствуют, что при кратковременной ишемии в зоне инфаркта активность NOS, выросшая более чем в 3 раза (11540 и 18649 пмоль/мин на 1 мг белка соответственно в опытной и контрольной группах) сравнительна с интактной зоной (этот же участок сердца у нормальных животных). Дрожжевая РНК почти полностью нормализовала активность NOS в пограничной зоне ишемического сердца, что может быть одним из механизмов ее кардиопротекторного действия. Известно, что кардиомиоциты содержат как индуцибельную изоформу NOS, так и конститутивные ее изоформы (Balligand J.L., Kobzik L., Han X., et al., Nitric oxide-dependent parasympathetic signaling is dueto activation of constitutive endotelial (type III) nitric oxid synthetase in cardiac myocytes, J. Biol. Chem., 16; 270(24); 14582-14586; 1995; Peng H.B., Spiecker M., Liao J.K. Inducible nitric oxid: an autoregulatory feedback inhibitor of vascular inflammation, J. Immunol. 15; 161(4): 1970-1976; 1998; Oddis C.V., Simmons R.L.,Haffler B.G., Finkel M.S. cAMP enchances inducible nitric oxid synthase mRNA stability in cardiac myocytes,Am. J. Physiol. 269(6): H2044-2050; 1995). Поэтому, учитывая короткий период (30 мин) действия ишемии, можно предусмотреть, что дрожжевая РНК ингибирует именно конститутивные изоформы NOS. В то же время, учитывая наличие в них iNOS, не исключено также, что дрожжевая РНК может действовать как ингибитор активности индуцибельной NOS в кардиомиоцитах при ишемии. Влияние дрожжевой РНК на активность NOS в крови крыс при ишемии показано в табл. 12,представленной в примере 5.4. Данные результаты определения свидетельствуют, что в отличие от сердца в крови контрольных животных при ишемии активность NOS снижалась в 2 раза (14,221,43 и 30,353,40 пмоль/мин на 1 мг белка соответственно при ишемии и нормоксии), что характерно для гипоксии (Arnet W.A., McMillan A., Dinerman J.L. et al., Regulation of endothelial nitric oxid synthase during hypoxia, J. Biol. Chem. 271(25): 15069-15073; 1996). Введение дрожжевой РНК почти полностью нормализовало активность NOS в крови крыс после 30-минутной ишемии. Пример 5.4. Влияние дрожжевой РНК на окислительный метаболизм арахидоновой кислоты при ишемии. В табл. 12 показана активность NOS и содержание свободной арахидоновой кислоты в крови нормальных и ишемических животных.- 20010182 Таблица 12 Влияние дрожжевой РНК на активность NOS и содержание свободной арахидоновой кислоты в крови крыс при ишемииP1 - достоверное отличие относительно нормы (перед ишемией); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Как показано в табл. 12, у контрольных животных наблюдали снижение уровней свободной AA более чем в 3 раза (0,771,43 и 30,353,40 нмоль на 1 мг белка соответственно при нормоксии и ишемии). Введение дрожжевой РНК немного нормализовало содержимое AA, повышая его вдвое против контрольных значений (Р 0,001). В табл. 13 показано содержимое свободной арахидоновой кислоты в разных участках сердца при ишемии. Таблица 13 Влияние дрожжевой РНК на содержимое свободной арахидоновой кислоты в сердце крыс при ишемии (в нмоль на 1 мг белка; Mm; n=5)P1 - достоверное отличие относительно нормы (перед ишемией); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Как показано в табл. 13, у контрольных животных наблюдали достоверное (более чем в два раза,Р 0,01) повышение содержимого AA как в пограничной, так и в инфарктной зоне сердца крыс. Введение дрожжевой РНК немного снижало содержимое арахидоновой кислоты в обеих зонах сердца, но отличие было недостоверным (Р 0,05). В табл. 14 показано влияние дрожжевой РНК на уровень эйкозаноидов в крови крыс при ишемии- 21010182 Таблица 14 Влияние дрожжевой РНК на соединения эйкозаноидов в крови крыс при ишемии (в пикомоль на 1 мг белка; Mm; n=5)P1 - достоверное отличие относительно нормы (перед ишемией); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). В табл. 14 показано влияние дрожжевой РНК на содержимое эйкозаноидов в крови крыс при ишемии. У контрольных животных наблюдали более выраженное повышение содержимого продукта циклооксигеназной реакции ТВХ 2 (более чем в 2 раза, но отличие недостоверно, Р 0,05), чем продукта липоксигеназной реакции LTC4 (Р 0,2). Введение дрожжевой РНК почти полностью нормализовало содержимое эйкозаноидов в крови крыс при ишемии. Таким образом, кроме модулирующего действия на активность NOS при ишемии (ингибирование в кардиомиоцитах и, наоборот, повышение в крови), кардиопротекторный эффект дрожжевой РНК может быть опосредованный также модуляцией окислительного метаболизма арахидоновой кислоты. Пример 6. Изучение противовоспалительного действия дрожжевой РНК на модели аутоиммунной патологии (адъювантного артрита) in vivo. Адъювантный артрит развивается у крыс после инъекции им адъюванта Фрейда и является частью генерализированных процессов, которые сопровождаются повреждением костных и соединительных тканей. Морфологические исследования показывают, что в процессе развития адъювантных артритов в тканях, которые окружают сустав, а также в середине суставной сумки и суставного хряща возникают воспалительно-дегенеративные изменения. Считают, что эта воспалительная реакция имеет все свойства иммунологических процессов и выражает приторможенную иммунную реакцию на микробный антиген. Патологические процессы, которые проходят при адъювантных артритах, очень похожи на артриты у людей. Пример 6.1. Влияние дрожжевой РНК на модели аутоиммунной патологии (адъювантный артрит). Адъювантный артрит смоделирован на самцах крыс соответственно методу Courtright et al.(Courtright L.J., Kuzell W.C., Sparing effect of neurological deficit and trauma on the course of adjuvant arthritis in the rat, Ann. Reum. Dis. 24(4): 360-368; 1965). Контрольные животные подкожно получали одинарную дозу 0,1 мл стандартного адъюванта Фрейда в дистальную часть хвоста. Адъювантный артрит развивался на 14-20-й день после инъекции. Симптомы артрита определяли рентгенологично: места затемнения и тени вокруг сустава задних конечностей означали начало повреждения сустава и хрящевых тканей. За день до инъекции адъюванта Фрейда опытной группе вводили дрожжевую РНК, растворенную в 0,9% растворе NaCl, из расчета 100 мг препарата на крысу. После инъекции адъюванта Фрейда животным также вводили дрожжевую РНК тремя сериями в течение 4 дней с трехдневным интервалом. Результаты изучения показали, что артрит в контрольной группе начинал развиваться на 14-й день и выражался эксудативно-пролиферативным ростом синовиальной капсулы и повреждением хряща. На 20-й день было свидетельство отвердения тканей вокруг сустава и начало фиброза синовиальной капсулы. На 30-й день стало явным разрушение хряща. В опытной группе, которая получала дрожжевую РНК до 20 дня, не было свидетельства проявлений артрита. Проявления артрита, подобные тем, которые проявлялись в контрольной группе на 14-й день, появлялись только на 30-й день. В контрольной группе в процессе развития адъювантного артрита задние ножки стали увеличиваться. Так, на 30-й день в контрольной группе размер задних ножек достоверно увеличился на 1,04 мм(4,90,13 в сравнении с 3,860,1 в начале опыта). В подопытной группе ножки увеличились только на 0,24 мм (4,10,11 на 30-й день опыта до 3,960,08 в начале опыта). Таким образом, дрожжевая РНК задерживает развитие адъювантного артрита, что также содействует уменьшению уровня увеличения задних ножек. Пример 6.2. Влияние дрожжевой РНК на активность NOS в крови крыс при аутоиммунной патологии (адъювантный артрит). Активность NOS изучали в крови нормальных крыс, у контрольных животных на 3-й день (I), 8-й день (II) и 14-й день (III) развития аутоиммунной патологии (без введения дрожжевой РНК) и у опытных- 22010182 животных, которым вводили дрожжевую РНК. Результаты изучения представлены в табл. 15. Таблица 15 Влияние препарата дрожжевой РНК на активность NOS в крови крыс в процессе развития адъювантного артритаP1 - достоверное отличие относительно нормы (при адъювантном артрите); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК). Как показано в табл. 15, в группе контрольных животных наблюдали значительное повышение активности NOS на 3- и 14-й день развития аутоиммунной патологии сравнительно с нормой (соответственно 30,657,35 пмоль за 1 мин на 1 мг белка в норме и 236,7676,42 пмоль за 1 мин на 1 мг белка на 3-й день и 111,5415,78 пмоль за 1 мин на 1 мг белка на 14-й день). Такое значительное повышение активности NOS указывает на то, что основным компонентом измеренной активности NOS является активность индуцибельной изоформы NOS (iNOS), синтез которой запускается противоспалительными цитокинами INF-, IL-I, TNF- и др. В промежутке между 3- (инициация аутоиммунного процесса) и 14-м днем (развитие патологии) наблюдали нормализацию уровня активности NOS в крови (24,348,60 пмоль за 1 мин на 1 мг белка),что, очевидно, обусловлено активацией защитной реакции организма и может быть вызвано как ингибированием экспрессии iNOS, так и модуляцией стабильности ее мРНК, или ингибированием процесса ее трансляции. В группе животных, которые получали дрожжевую РНК, инициация аутоиммунного процесса(на 3-й день (I) сопровождалась значительно меньшим, сравнительно с контролем, повышением активности NOS в крови (70,009,24 пмоль за 1 мин на 1 мг белка против 236,7676,42 пмоль за 1 мин на 1 мг белка соответственно), причем активность NOS прогрессивно снижалась в течение всего следующего периода развития аутоиммунного процесса (40,665,05 пмоль за 1 мин на 1 мг белка на 8-й день (II) и 33,966,04 пмоль за 1 мин на 1 мг белка на 14-й день (III. Таким образом, проведенные исследования изменения активности NOS в крови крыс в динамике развития аутоиммунного процесса позволяют сделать вывод, что дрожжевая РНК является эффективным агентом в снижении активности iNOS при развитии аутоиммунного процесса (как в период его инициации, так и на стадии его хронизации). Эти свойства дают возможность использовать дрожжевую РНК при патологических состояниях, которые сопровождаются индукцией iNOS, таких как воспалительные процессы, диабет, атеросклероз, гепатиты, инфекции, рак, нейродегенеративные заболевания (паркинсонизм, болезнь Альцгеймера, рассеянный склероз, энцефалит) и пр. Пример 6.3. Мембранопротекторное действие дрожжевой РНК. Опыты проводили in vivo на модели развития хронического аутоиммунного процесса, который сопровождается генерацией большого количества свободных радикалов (в особенности оксида азота) в ранний период инициации процесса. Мембранопротекторное действие дрожжевой РНК оценивали, определяя кислотную резистентность эритроцитов в динамике развития аутоиммунного процесса. Кислотная резистентность характеризует целостность мембраны эритроцитов. Она возрастает при хроническом периоде развития многих патологий и, наоборот, снижается в период острого развития патологического состояния (процесс его инициации). Например, в ранний период развития воспалительного процесса происходит значительная активация свободнорадикальных процессов, вызванных генерацией свободных радикалов кислорода и азота, в том числе оксида азота, который генерируется индуцибельной изоформой NOS (iNOS). Степень повреждения эритроцитов при действии различных факторов в процессе развития аутоиммунного процесса оценивали по кинетическим параметрам гемолиза, который индуцировали снижениемpH среды. Кинетические параметры гемолиза регистрировали, спектрофотометрично, определяя количество разрушенных клеток через соответствующие промежутки времени (30 с) по изменению величины интегрального светорассеяния суспензии эритроцитов (=750 нмоль). Спектры поглощения регистрировали с помощью спектрофотометра СФ-26 (Россия). Кислотный лизис эритроцитов инициировали, внося 10 мкл разбавленной в 20 раз крови в изотонической среде: 0,14 М NaCl+0,01 М цитратно-фосфатный бу- 23010182 фер со значением pH 2,0-3,5 (объем: 1 мл; плотность эритроцитов в суспензии: 0,7106 клеток на 1 мл). При такой плотности суспензии эритроцитов величина интегрального светорассеяния, которая зависит от числа, размеров и формы клеток, пропорциональна числу клеток в суспензии. Результаты представлены в виде диаграмм кислотного гемолиза эритроцитов в табл. 16 и в виде интегрального параметра этого процесса: суммарной кислотной стойкости эритроцитов, которую определяли, суммируя произведения количества клеток, которые гемолизировали в данный период времени (ai) на ti (суммарная стойкость (I) = ai, ti). Снижение экстинции на диаграммах гемолиза отображает очередность включения в гемолиз эритроцитов возрастающей стойкости. Снижение экстинции начинается, как правило, через 1,5-2 мин со времени введения гемолитика (1 мл 0,004N HCl, которые готовили из фиксанала HCl и проверяли титрованием), а лаг-период гемолиза обусловлен предгемолизным изменением формы эритроцитов (сферуляция). Продолжительность гемолиза отдельного эритроцита не превышает 10 с, поэтому 30-секундный интервал между измерениями экстинции гарантирует невозможность двукратного учета лизирующего эритроцита. Отсюда вытекает, что при фотометрической регистрации кинетики гемолиза из полученного ряда экстинций с промежутком в 30 с можно высчитать процент распределения эритроцитов по группам стойкости. Определение изменения экстинции (E) с момента начала гемолиза (Eп, tп) к его полному завершению (Ек, tк) пропорционально числу всех клеток, которые принимают участие в процессе гемолиза, которое принимается за 100% и тогда Е=Ек-Еп=100%. Это суммарное количество эритроцитов, которые гемолизируют (100%), состоит из суммы гемолизирующих эритроцитов за каждые 30 с (Ei+1-Ei) в интервале tк-tп = продолжительность гемолиза:E=Ei+1-Ei=100%. Результаты представлены в табл. 16 Таблица 16 Влияние препарата дрожжевой РНК на кислотную резистентность эритроцитов в динамике развития адъювантного артритаP1 - достоверное отличие относительно нормы (при адъювантном артрите); Р 2 - достоверное отличие относительно контроля (без дрожжевой РНК.) В табл. 16 показана суммарная стойкость неотмытых эритроцитов крыс в динамике развития аутоиммунного процесса. Для нормальных эритроцитов значение этого параметра составляет 71285 единиц. В период инициации аутоиммунного процесса суммарная стойкость эритроцитов уменьшалась более чем в 7 раз и составляла 9538 единиц, постепенно повышаясь в процессе развития патологии, так что на 14-й день (III) она составляла 1114290 единиц. Такое существенное снижение кислотной резистентности эритроцитов характеризует значительные изменения как в самой плазматической мембране клеток (очевидно за счет окисления белкового и липидного компонентов свободными радикалами, которые активно генерируются в этот период, в том числе оксидом азота), так и в плазме (можно предусмотреть модуляцию содержимого свободного холестерола, полиаминов и других стабилизаторов, а также возрастание содержимого дестабилизаторов), таких как, например, полиненасыщенные свободные жирные кислоты. У животных, которые получали дрожжевую РНК, период инициации аутоиммунного процесса не сопровождался таким значительным, как в контроле, снижением кислотной резистентности эритроцитов,о чем свидетельствует величина суммарной стойкости 37373 единиц, которые хотя и достоверно ниже уровня нормы (Р 0,05), в то же время достоверно более высокое значение в контроле (Р 0,02). В следующие периоды развития аутоиммунной патологии суммарная стойкость эритроцитов у животных, которые получали дрожжевую РНК, была на уровне нормы. Таким образом, дрожжевая РНК обнаружила мембраностабилизирующее действие, учитывая основные механизмы повреждения при данной патологии - окислительный стресс, свободнорадикальное повреждение компонентов плазматической мембраны - также и антирадикальное действие.- 24010182 Пример 7. Влияние дрожжевой РНК на показатели крови. Образцы крови людей до и после употребления препарата дрожжевой РНК изучали на автоматическом гемоцитометре фирмы "Serono1900", Austria, соответственно инструкциям, рекомендованным фирмой. Фиксировали значения количества лейкоцитов [WBC], эритроцитов [RBC] и тромбоцитов [PLT] в 1 мкл крови, количество гемоглобина [HGB] в г/дл, нейтрофилов (NTP) и гематокрита [HGB] в процентах. Здоровых и больных добровольцев отбирали после предшествующего изучения по вышеупомянутым показателям. Для дальнейшего изучения отбирали группы от 4 до 6 добровольцев, у которых обнаружили стойкое уменьшение ниже нормы одного или более вышеупомянутых показателей. В зависимости от типа заболевания лечение проводили от 1 до 18 недель. Препарат дрожжевой РНК давали или в виде капсул в концентрации по 250 мг дрожжевой РНК на одну капсулу, или в виде супозиториев в концентрации 1,0 г дрожжевой РНК на супозиторий. Пример 7.1. Влияние дрожжевой РНК на цитопению крови относительно здоровых людей и атлетов. Курс лечения проходил от 10 дней до 6 недель. В капсулах препарат давали по 1 г каждый день, а в супозитории давали по 1 г на три дня. Анализы проводили 1-2 раза в неделю, результаты представлены в табл. 17 и 18. В табл. 17 представлены результаты лечения группы добровольцев с симптомами анемии, которые принимали препарат дрожжевой РНК в капсулах по 1 г в день в течение 3 недель. Как видно из таблицы,за время лечения обнаружено стойкое увеличение концентрации гемоглобина, которое повысилось с 12 до 14 г/дл, а гематокрит повысился с 30 до 37%. В этой же группе обнаружено повышение количества эритроцитов на 17,7% вместе с повышением количества лимфоцитов и тромбоцитов соответственно на 26,5 и 59,9%. Таблица 17 Влияние препарата дрожжевой РНК (в капсулах 1 г/дл) на показатели цитопении крови относительно здоровых пациентов Поскольку употребление препарата per os приводит к его быстрому гидролизу, то в кровь попадает значительно меньшая его концентрация. Поэтому было изучено действие препарата дрожжевой РНК в виде супозиториев в концентрации 1 г/день в группе относительно здоровых людей. Супозитории вводили на 1, 3 и 6 дни. Результаты исследований показали, что гематокрит возрос при этих условиях с 33,2 до 41,5%, в то же время гемоглобин возрос с 13,5 до 14,5 г/дл. Количество лейкоцитов возросло на 46,5%, а эритроцитов на 10,6%, количество тромбоцитов осталось стабильным. Итак, употребление препарата дрожжевой РНК в виде супозиториев, которое приравнивается к внутривенному введению препарата, разрешает в три раза быстрее достичь результата (7 дней в виде супозиториев вместо 21 дня в виде капсул) при семикратном уменьшении с 21 до 3 г общего количества препарата дрожжевой РНК на курс лечения. Такая ускоренная нормализация показателей крови важна при кровотечениях, если требуется переливание крови. В табл. 18 показаны результаты лечения заниженных показателей крови у атлетов. Известно, что во время интенсивных тренировок у этой группы людей часто уменьшаются показатели гематокрита, гемоглобина и пр. Препарат дрожжевой РНК давали по 1,5 г/день на протяжении 3 недель. Определяли только показатели гематокрита и гемоглобина.- 25010182 Таблица 18 Влияние дрожжевой РНК (в капсулах по 1,5 г/день) на показатели крови у атлетов Показано, что уровень гематокрита и гемоглобина в процессе интенсивных тренировок без препарата за два дня упал соответственно с 42,7% и 13,9 г/дл до 41,0% и 13,5 г/дл. Через 10 дней после приема дрожжевой РНК при интенсивных тренировках они возросли до 43,7% и 14,1 г/дл соответственно, после 16 дней - до 45,7% и 14,3 г/дл, а после 21 дня - до 46,5% и 14,8 г/дл соответственно. Итак, в этой группе за три недели интенсивных тренировок обнаружено увеличение гематокрита на 13,4%, а гемоглобина на 6%. Пример 7.2. Влияние дрожжевой РНК на цитопению крови у больных раком. Анемия играет очень негативную роль у больных раком, в особенности после химиотерапии и радиотерапии. Больным с уровнем гемоглобина ниже 80 г/л химиотерапия или радиотерапия вообще не назначаются. Целый ряд исследований указывают на то, что лечение анемии и повышение уровня гемоглобина играют важную роль у больных раком, которые подвергались лечению химиотерапией(Epoetin Alfa) in the Treatment of the Anemia of Cancer, Oncology, vol. 56, p. 46-53 (1999. Применяя эритропоэтин для лечения больных после химиотерапии, ряд исследований показал, что увеличение количества гемоглобина от 80 до 100 г/л улучшает самочувствие больных раком. Более существенное улучшение степени самочувствия возрастает с нарастанием уровня гемоглобина от 100 до 120 г/л. Итак, уровень гемоглобина 120 г/л является оптимальным для больных раком как по степени их самочувствия, так и по результатам лечения (J. Crawford, Anemia, Fatigue, and Erythropoietin, 42nd AnnualMeeting of the American Society of Hematology, 2000, Medscape, Inc.). Изучалась группа больных раком, которым давали препарат дрожжевой РНК по 1 г в день в виде капсул в течение 8 дней до химиотерапии, во время химиотерапии и в перерыве между сеансами повторной химиотерапии в течение 2-3 месяцев. Лабораторные исследования крови проводились соответственно методу, который описан выше. Результаты этих анализов представлены в табл. 19. Таблица 19 Влияние дрожжевой РНК (в капсулах 1-2 г/день) на цитопению крови у больных раком. Как показано в табл. 19, с момента приема препарата дрожжевой РНК у больных неуклонно возрастает количество гемоглобина, процент гематокрита, количество эритроцитов и тромбоцитов. В течение 6 недель лечения количество гемоглобина выросло почти вдвое с 67 до 121 г/л, количество эритроцитов выросло на 86%, а тромбоцитов на 130%. У больных не обнаружены ухудшения других показателей крови и отмечено увеличение степени самочувствия с возрастанием количества гемоглобина до 120 г/л. Пример 7.3. Влияние дрожжевой РНК на цитопению крови у ВИЧ-инфицированных больных. У ВИЧ-инфицированных больных обнаруживается комплексное нарушение гематопоэза, которое приводит к денормализации всех трех клеточных линий, которые происходят из предшественников клеток гематопоэза. Таким образом, не менее 80% инфицированных вирусом HIV в процессе развития инфекции болеют анемией, более 50% таких пациентов болеют нейтропенией и 40% - тромбопенией. Итак,цитопения ВИЧ-инфицированных больных является одним из первых признаков болезни СПИДHIV-infected Patients, HIV Clinical Management, vol. 10, 1999, Medscape, Inc.). Группа отобранных ВИЧ-инфицированных пациентов была под медицинским наблюдением в течение 6 месяцев. После изучения их гемопоэза начали лечение капсулами с дрожжевой РНК в концентрации от 1 до 6 г в день в течение 18 месяцев. Гематологические и биохимические анализы проводили каждые 1-2 недели. Результаты лечения представлены в табл. 20. Показано, что ВИЧ-инфицированные больные в течение 6 месяцев до лечения имели четко выраженную цитопению с падением уровня гемоглобина, количества тромбоцитов, нейтрофилов и процента гематокрита. Ряд больных были инфицированы вирусами гепатита. Между 12-14 неделями лечения во время эпидемии пациенты переболели гриппом,который перерос в воспаление легких. Поэтому в настоящее время они дополнительно лечились антибиотиками. Таблица 20 Влияние дрожжевой РНК (в капсулах 1-2 г/день) на цитопению крови у ВИЧ-инфицированных пациентов Как показано в табл. 20, в результате лечения ВИЧ-инфицированных через 4-6 недель обнаружена стойкая нормализация всех показателей крови. Через 4 недели выявлена нормализация показателей эритроцитов, нейтрофилов и тромбоцитов, что свидетельствует о комплексной нормализации дифференциации всех трех линий клетки предшественника CFU-GEMM. Не обнаружено ненормальной стимуляции количества лимфоцитов. Гемоглобин в течение 4 недель возрос с 99,43 до 125 г/л и поддерживался на этом уровне до конца наблюдения. Нейтрофилы возросли за 4 недели с 36,75 до 49% и в этих границах поддерживались с некоторыми колебаниями в течение 18 недель. Итак, лечение дрожжевой РНК инфицированных HIV пациентов показало стойкую нормализацию у них цитопении. Это свидетельствует о целесообразности использования данного препарата для предупреждения и лечения воспалительных процессов и для поддержания у них высокого уровня самочувствия и трудоспособности. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ лечения воспаления и связанных с ним заболеваний за счет стабилизации клеточных мембран, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном улучшать симптомы воспаления или связанных с ними заболеваний, в фармакологически приемлемых наполнителях, носителях и растворах. 2. Способ по п.1, отличающийся тем, что млекопитающим, у которых имеются повреждения клеточных мембран, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном стабилизировать повреждение клеточной мембраны, в фармакологически приемлемых наполнителях, носителях и растворах. 3. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении,назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном угнетать окисление компонентов клеточных мембран млекопитающих, в фармакологически приемлемых наполнителях, носителях и растворах. 4. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении,назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном нормализовать активность NO-синтетазы, в фармакологически приемлемых наполнителях, носителях и растворах.- 27010182 5. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении,назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном угнетать агрегацию тромбоцитов, в фармакологически приемлемых наполнителях, носителях и растворах. 6. Способ по п.1, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении,назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном препятствовать или лечить цитопению крови, в фармакологически приемлемых наполнителях, носителях и растворах. 7. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают интрадермально, подкожно, перорально, интраабдоминально, внутримышечно или внутривенно либо назначают в месте воспаления или в месте, связанном с воспалительным заболеванием. 8. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде капсул. 9. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде суппозиториев. 10. Способ по п.1, отличающийся тем, что связанным с ним заболеванием является инфаркт. 11. Способ по п.1, отличающийся тем, что связанным с ним заболеванием является инсульт. 12. Способ по п.1, отличающийся тем, что связанным с ним заболеванием является артрит. 13. Способ в соответствии с любым из пп.1-6, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде фармацевтической композиции, которая содержит очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах. 14. Способ улучшения уровня не менее одного показателя крови за счет стабилизации мембран клеток крови, отличающийся тем, что млекопитающим, которые нуждаются в таком лечении, назначают очищенную дрожжевую рибонуклеиновую кислоту в эффективном количестве, способном улучшать уровень не менее одного показателя крови, в фармакологически приемлемых наполнителях, носителях и растворах. 15. Способ по п.14, отличающийся тем, что такой показатель крови выбирают из соответствующих уровней лейкоцитов, эритроцитов, тромбоцитов, гемоглобина, нейтрофилов и гематокрита. 16. Способ по п.14, отличающийся тем, что млекопитающим, которые требуют профилактики или лечения анемии, назначают очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах. 17. Способ по п.14, отличающийся тем, что млекопитающим, которые требуют профилактики или лечения тромбоцитопении, назначают очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах. 18. Способ по п.14, отличающийся тем, что млекопитающим, которые требуют профилактики или лечения нейтропении, назначают очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах. 19. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают интрадермально, подкожно, перорально, интраабдоминально, внутримышечно или внутривенно либо назначают в месте воспаления или в месте, связанном с воспалительным заболеванием. 20. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде капсул. 21. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде суппозиториев. 22. Способ в соответствии с любым из пп.14-18, отличающийся тем, что очищенную рибонуклеиновую кислоту назначают в виде фармацевтической композиции, которая содержит очищенную дрожжевую рибонуклеиновую кислоту в фармакологически приемлемых наполнителях, носителях и растворах. 23. Способ очистки дрожжевой рибонуклеиновой кислоты, отличающийся тем, что рибонуклеиновую кислоту очищают от белков и ДНК при помощи многократной инкубации с применением ферментов, в частности панкреатина. 24. Способ по п.23, отличающийся тем, что рибонуклеиновую кислоту получают из Saccharomycescerevisiae. 25. Способ по п.23, отличающийся тем, что рибонуклеиновую кислоту получают из Candida utilis. 26. Способ по п.23, отличающийся тем, что очищенная рибонуклеиновая кислота содержит азот в количестве более 14,5 мас.%. 27. Способ по п.23, отличающийся тем, что очищенная рибонуклеиновая кислота содержит фосфор в количестве более 8,5 мас.%. Евразийская патентная организация, ЕАПВ Россия, 109012, Москва, Малый Черкасский пер., 2/6
МПК / Метки
МПК: A61P 29/00, C12N 15/11, A61K 36/06, A61K 31/7105
Метки: уровня, дрожжевой, рибонуклеиновой, улучшения, одного, связанных, способ, очистки, также, крови, воспаления, показателя, ним, заболеваний, лечения, кислоты, менее
Код ссылки
<a href="https://eas.patents.su/30-10182-sposob-lecheniya-vospaleniya-i-svyazannyh-s-nim-zabolevanijj-i-sposob-uluchsheniya-urovnya-ne-menee-odnogo-pokazatelya-krovi-a-takzhe-sposob-ochistki-drozhzhevojj-ribonukleinovojj.html" rel="bookmark" title="База патентов Евразийского Союза">Способ лечения воспаления и связанных с ним заболеваний и способ улучшения уровня не менее одного показателя крови, а также способ очистки дрожжевой рибонуклеиновой кислоты</a>
Применение производных 7-(2-окса-5,8-диазабицикло[4,3,0]- нон-8-ил)-хинолонкарбоновой кислоты и -нафтиридонкарбоновой кислоты для лечения инфекций, вызванных helicobacter pylori, и связанных с ними гастродуоденальных заболеваний

Номер патента: 2477
Опубликовано: 27.06.2002
Авторы: Шенке Томас, Химмлер Томас, Баазнер Бернд, Верлинг Ханс-Отто, Матцке Михаэль, Лабишински Харальд, Петерсен Уве, Йетш Томас, Бартель Штефан, Шаллер Клаус
МПК: A61K 31/4709, A61P 1/04
Метки: производных, pylori, helicobacter, лечения, кислоты, 7-(2-окса-5,8-диазабицикло[4,3,0, ними, связанных, гастродуоденальных, нон-8-ил)-хинолонкарбоновой, нафтиридонкарбоновой, применение, инфекций, вызванных, заболеваний
Формула / Реферат:
1. Применение композиции, содержащей в качестве действующего вещества производные хинолинкарбоновой кислоты общей формулы (I) в которой R1 означает алкил с числом атомов углерода от 1 до 4, при необходимости замещенный однократно или двухкратно галогеном, фенил, при необходимости замещенный одним или двумя атомами фтора, или циклопропил, при необходимости замещенный одним или двумя атомами фтора, R2 означает водород, алкил с числом атомов...
Вакцина для лечения воспаления и доброкачественной гиперплазии предстательной железы на основе бактерий lactobacilli и способ лечения указанных заболеваний

Номер патента: 5198
Опубликовано: 30.12.2004
Авторы: Лазар Эрика Агнеш, Ийхейи Карой, Варга Дьюла Арпад, Бартуш Йожеф
МПК: A61K 39/02, A61P 13/08
Метки: железы, бактерий, указанных, вакцина, лечения, гиперплазии, воспаления, lactobacilli, заболеваний, основе, способ, предстательной, доброкачественной
Формула / Реферат:
1. Вакцина для лечения воспаления предстательной железы и доброкачественной гиперплазии предстательной железы (I и II стадии), содержащая штаммы Lactobacillus, входящие в состав вакцин GynevacR, GynatrenR и SolcoTrichovacR, в инактивированной форме, и носители и/или эксципиенты, обычно используемые в вакцинных препаратах. 2. Применение штаммов Lactobacillus, входящих в состав вакцин GynevacR, GynatrenR и SolcoTrichovacR, в инактивированной форме...
Производные аминогуанидина и алкоксигуанидина (варианты) и способ их получения (варианты), фармацевтическая композиция и способ ингибирования протеолиза у млекопитающего, способ лечения различных заболеваний млекопитающего, способы ингибирования индуцированной тромбином агрегации тромбоцитов и свертывания фибриногена в плазме, тромбина, агрегации тромбоцитов и образования тромбов в крови, устройство для сбора крови, искусственного кровообращения и хранения крови

Номер патента: 3461
Опубликовано: 26.06.2003
Авторы: Сайедем Коллин, Лу Тианбао, Маркоутэн Томас П., Солл Ричард М., Томкзук Брюс Э.
МПК: A61K 31/445, A61P 7/02, C07C 279/00...
Метки: лечения, индуцированной, кровообращения, фармацевтическая, производные, млекопитающего, хранения, алкоксигуанидина, протеолиза, свертывания, аминогуанидина, способы, композиция, сбора, варианты, плазме, тромбином, тромбина, способ, фибриногена, получения, крови, тромбоцитов, заболеваний, искусственного, образования, ингибирования, различных, агрегации, тромбов, устройство
Формула / Реферат:
1. Соединение формулы I или его сольват, гидрат или фармацевтически приемлемая соль, где L - означает -C(O)- или -SO2-; R1 означает группу R2 означает группу или R1 и R2 могут быть объединены с атомом азота, к которому они присоединены, с образованием цикла, содержащего от трех до семи атомов, который необязательно содержит дополнительный атом азота или кислорода, и который по выбору является бензо- или пиридоконденсированным циклом, причем...
Способ лечения заболеваний, связанных с нарушениями в эндокринной системе

Номер патента: 5224
Опубликовано: 30.12.2004
Авторы: Соломатова Галина Дмитриевна, Соломатов Геннадий Сергеевич, Соломатова Галина Владимировна, Соломатова Мария Викторовна, Соломатов Виктор Геннадьевич, Соломатова Анна Викторовна
МПК: A61H 39/00
Метки: связанных, эндокринной, лечения, системе, способ, нарушениями, заболеваний
Формула / Реферат:
1. Способ лечения заболеваний, связанных с нарушениями в эндокринной системе, включающий воздействие на биологически активные точки и/или зоны энергорефлексотерапии, отвечающие за нормализацию нейрогуморальной регуляции нервной и/или эндокринной и/или нейроэндокринной систем и сопряженных органов, отличающийся тем, что в качестве биологически активных точек и/или зон энергорефлексотерапии органов, отвечающих за нормализацию регуляции элементов...
Способ лечения заболеваний, связанных с образованием конкрементов

Номер патента: 1750
Опубликовано: 27.08.2001
Авторы: Санников Юрий Петрович, Брижан Мария Валерьевна, Бобренко Андрей Владимирович, Соломатов Виктор Геннадьевич
МПК: A61H 39/08, A61H 39/00
Метки: заболеваний, конкрементов, способ, образованием, связанных, лечения
Формула / Реферат:
1. Способ лечения заболеваний, связанных с образованием конкрементов, включающий воздействие на биологически активные точки и/или рефлексогенные зоны, отвечающие за физико-химическое состояние литогенного субстрата и/или усиление деятельности гладкой мускулатуры полостного органа, включая сфинктеры, отличающийся тем, что в качестве биологически активных точек и/или рефлексогенных зон, ответственных за физико-химическое состояние литогенного...
Предыдущий патент: Фосфолипидные смазывающие вещества в жидкостях для бурения на водной основе
Следующий патент: Сложный моноэфир янтарной кислоты и пробукола для лечения сердечно-сосудистых и воспалительных заболеваний
Случайный патент: Устройство для остеосинтеза около- и внутрисуставных переломов длинных костей