Белок, связывающийся с &beta-амилоидным пептидом, кодирующий его полинуклеотид и способы их использования
Номер патента: 4256
Опубликовано: 26.02.2004
Авторы: Волкер Стефен Гленн, Джекобсен Джек Стивен, Озенбергер Бредли Элтон, Кажковски Эйлин Мари, Бард Джонатан Адам, Софиа Хейди







Формула / Реферат
1. Фрагмент молекулы кДНК, включающий нуклеотиды 202-807 последовательности SEQ ID NO:1, приведенной в Списке последовательностей, кодирующий фрагмент белка, связывающегося с b -амилоидным пептидом (BBP), включающим аминокислоты 68-269 последовательности SEQ ID NO:2, приведенной в данном Списке, а также вариант указанного фрагмента кДНК, полученный на основании вырожденности генетического кода.
2. Фрагмент молекулы кДНК по п.1, представляющий собой вставку клона pEK 196 ATCC 98399.
3. Молекула кДНК, имеющая последовательность нуклеотидов SEQ ID NO:1, приведенную в Списке последовательностей, кодирующая белок, связывающийся с b -амилоидным пептидом (BBP), имеющим аминокислотную последовательность SEQ ID NO: 2, приведенную в данном Списке, а также вариант указанной молекулы кДНК, полученный на основании вырожденности генетического кода.
4. Молекула кДНК по п.3, представляющая собой вставку клона BBP1-fl ATCC 98617.
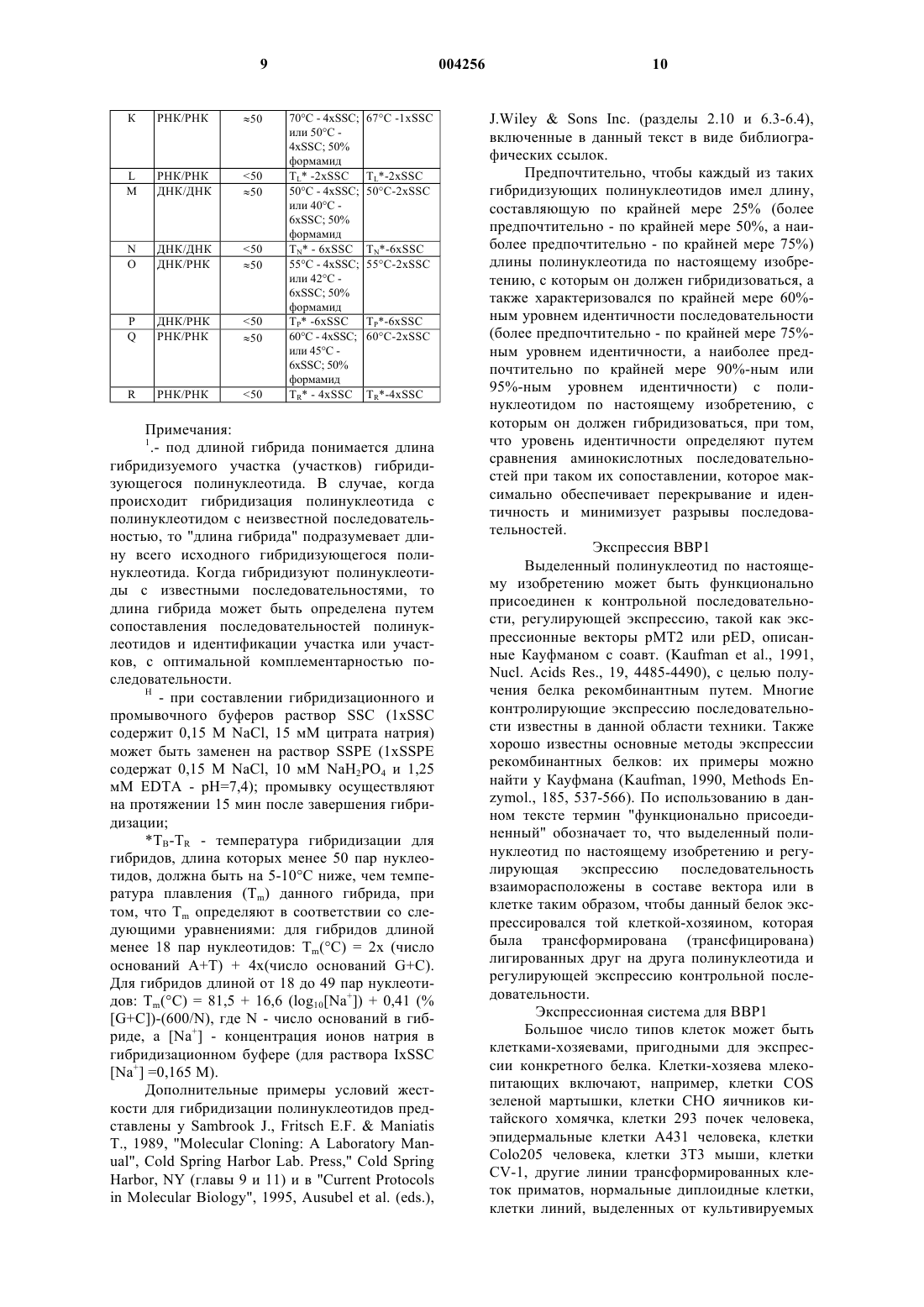
5. Полинуклеотид, который гибридизуется в жестких условиях с молекулой кДНК по любому из пп.1-4 или с комплементарной ей молекулой, при том, что указанный полинуклеотид имеет длину, составляющую по крайней мере 75% длины указанной молекулы кДНК, и кодирует b -амилоидный пептид.
6. Зонд или праймер, включающий нуклеотидную последовательность, способную гибридизоваться с нуклеотидами 172-194 молекулы кДНК по п.3.
7. Полинуклеотид, включающий по крайней мере одну последовательность, регулирующую экспрессию, функционально связанную по крайней мере с одним полинуклеотидом, выбранным из группы, состоящей из молекулы кДНК по любому из пп.1-4.
8. Штамм клетки-хозяина, трансформированный полинуклеотидом по п.7, при том, что указанная клетка представляет собой прокариотическую клетку.
9. Штамм клетки-хозяина, трансформированный полинуклеотидом по п.7, при том, что указанная клетка представляет собой эукариотическую клетку.
10. Фрагмент белка, связывающегося с b -амилоидным пептидом (BBP), включающий аминокислоты 68-269 последовательности SEQ ID NO:2, приведенной в Списке последовательностей, кодируемый фрагментом молекулы кДНК, включающим нуклеотиды 202-807 последовательности SEQ ID NO:1, приведенной в данном Списке, или кодируемый вариантом указанного фрагмента кДНК, полученного на основании вырожденности генетического кода.
11. Фрагмент белка, связывающегося с b -амилоидным пептидом (BBP), кодируемый фрагментом молекулы кДНК по п.1, представляющим собой вставку клона pEK 196 ATCC 98399.
12. Белок, связывающийся с b -амилоидным пептидом (BBP), имеющий аминокислотную последовательность SEQ ID NO: 2, приведенную в Списке последовательностей, кодируемый молекулой кДНК, имеющей последовательность нуклеотидов SEQ ID NO:1, приведенную в данном Списке, или кодируемый вариантом указанной молекулы кДНК, полученным на основании вырожденности генетического кода.
13. Белок BBP, включающий аминокислотную последовательность, кодируемую молекулой кДНК по п.3, представляющей собой вставку клона BBP1-fl ATCC 98617.
14. Способ получения белка по любому из пп.12-13 или фрагмента данного белка по любому из пп.10-11, заключающийся в том, что выращивают штамм клетки-хозяина по любому из пп.8-9 в подходящей культуральной среде и очищают продуцируемый указанным штаммом белок или его фрагмент.
15. Белок, полученный в соответствии со способом по п.14.
16. Слитый белок BBP, включающий белок по любому из пп.12-13, 15 или фрагмент данного белка по любому из пп.10-11, связанный с гетерологичной белковой или пептидной последовательностью.
17. Олигонуклеотид, представляющий собой ингибитор экспрессии гена BBP1, включающий нуклеотидную последовательность, состоящую из 15-35 нуклеотидов, которая гибридизуется в жестких условиях с молекулой кДНК по любому из пп.1-4.
18. Способ определения количества полинуклеотида, кодирующего белок, связывающегося с b -амилоидным пептидом (BBP), в образце, включающий следующие стадии:
(a) гибридизацию зонда по п.6 с образцом в жестких условиях и
(b) определение количества указанного зонда, связавшегося с полинуклеотидами в образце.
19. Антитело, которое специфически связывается с внеклеточной областью белка по любому из пп.12-13, 15-16 или фрагмента данного белка по любому из пп.10-11.
20. Способ определения в образце количества белка по любому из пп.12-13, 15-16 или фрагмента данного белка по любому из пп.10-11, включающий следующие стадии:
a) инкубацию антитела по п.19 с образцом и контролем, при том, что указанный контроль включает различные количества известного белка ВВР или фрагмента белка BBP и где указанное антитело связано с репортерной молекулой;
b) определение количества указанного антитела, связанного с указанным белком BBP или фрагментом белка в образце и
c) количественное определение указанного белка BBP или фрагмента белка BBP путем сравнения количества антитела, определенного в b), с указанными контролями.
21. Способ лечения пациента, нуждающегося в ингибировании накопления b -амилоидного пептида в головном мозге, включающий введение пациенту терапевтически эффективного количества белка по любому из пп.12-13, 15-16 или фрагмента данного белка по любому из пп.10-11.
Текст
1 Данная заявка раскрывает преимущества предварительной заявки США 60/064583,поданной 16 апреля 1997 г, содержание которой включено здесь в качестве ссылки. Область техники, к которой относится изобретение Настоящее изобретение касается новых полинуклеотидов и белков, кодируемых такими полинуклеотидами, наряду с терапевтическим,диагностическим и исследовательским применением этих полинуклеотидов и белков. В частности, настоящее изобретение касается полинуклеотидов и белков, кодируемых такими полинуклеотидами, которые связываются с амилоидным пептидом, являющимся одним из первичных компонентов амилоидных отложений, связанных с патогенезом болезни Альцгеймера. Предпосылки для изобретения Болезнь Альцгеймера (AD) - нарушение,связанное с прогрессирующей деменцией у пожилых людей, характеризующееся рядом структурных аномалий головного мозга. В множественных участках центральной нервной системы(ЦНС) нейроны становятся дисфункциональными и погибают, в результате чего происходит изменение синаптических путей. Тела и проксимальные дендриты этих уязвимых нейронов содержат нейрофибриллярные клубки,состоящие из парных спиральных нитей, основным компонентом которых является фосфорилированный, связывающийся с микротрубочками белок, обозначенный как "-белок" (белок"тау"). Одним из существенных признаков данного заболевания является накопление амилоидных отложений в пределах головного мозга,называемых старческими (или нейритными) бляшками. Принципиальным компонентом амилоидных бляшек является -амилоидный пептид(здесь и далее обозначается как "ВАР", а в литературе также известный как "А", "АР" и др.),который образует плотные скопления в процессе развития AD. ВАР - состоящий из 39-43 аминокислот белок, образующийся в результате протеолитического расщепления из амилоидного полипептида-предшественника (здесь и далее обозначаемого как "АРР") и состоящий из части трансмембранного домена и домена "просвета эндоплазматического ретикулюма" АРР. Считается, что пептид ВАР, состоящий из 42 аминокислот (ВАР 42), у человека является потенциально более токсичной агрегированной формой. АРР встречается в виде нескольких ВАРвключающих изоформ. Основные изоформы состоят из 695, 751 и 770 аминокислот, при том,что последние две АРР включают домен, обладающий структурной и функциональной гомологией с ингибиторами сериновой протеазы Кюнитца. У здоровых индивидуумов ВАР не накапливается и быстро удаляется из циркули 004256 2 рующих жидкостей организма. Однако пептид способен образовывать бляшки на поверхности аномальных дендритов и аксонов, микроглии и реактивных астроцитов. Агрегирование и отложение ВАР в виде нейритных бляшек рассматривается как один из запускных механизмов развития болезни Альцгеймера. Изучение процессов, приводящих к экспрессии и функций ВАР и их собственной роли в патогенезе AD,занимает существенное внимание в нейрологических исследованиях. В частности, открытие белков, которые могут связываться с белком ВАР, является принципиальным в связи с пониманием патогенетических путей данного заболевания и приводит к появлению новых потенциальных терапевтических мишеней. До настоящего изобретения белки и их фрагменты, которые бы связывались с белком ВАР человека и которые бы могли быть вовлечены в биологические эффекты ВАР в ходе развития AD, идентифицированы не были. Резюме изобретения Настоящее изобретение представляет новые выделенные полинуклеотиды, которые кодируют генные продукты, избирательно связывающиеся с аминокислотными последовательностями -амилоидного пептида человека(ВАР). В одном из вариантов настоящее изобретение представляет композицию, включающую выделенный полинуклеотид, выбираемый из группы, включающей:(a) полинуклеотид, включающий нуклеотидную последовательность по SEQ ID NO 1;(b) полинуклеотид, включающий нуклеотидную последовательность связывающегося сBBP1-fl, внесенного под депозитарным номером АТСС 98617;(ВВР), кодируемый вставкой кДНК клонаBBP1-fl, внесенного под депозитарным номером АТСС 98617;(d) полинуклеотид, включающий нуклеотидную последовательность по SEQ ID NO 1 от 202-го нуклеотида до 807-го нуклеотида;(e) полинуклеотид, включающий нуклеотидную последовательность связывающегося с-амилоидным пептидом белка (ВВР) клона рЕК 196, внесенного под депозитарным номером АТСС 98399;(ВВР), кодируемый вставкой кДНК клона рЕК 196, внесенного под депозитарным номером АТСС 98399;(g) полинуклеотид, кодирующий белок,включающий аминокислотную последовательность по SEQ ID NO 2;(h) полинуклеотид, кодирующий белок,включающий фрагмент аминокислотной последовательности по SEQ ID NO 2, обладающий активностью по связыванию с -амилоидным пептидом, т.е. фрагмент, включающий аминокислотную последовательность от 68-й аминокислоты до 269-й аминокислоты в соответствии с SEQ ID NO 2;(j) полинуклеотид, являющийся аллельным вариантом полинуклеотида по пп.(a)-(f), описанным выше;(k) полинуклеотид, который кодирует вариант, гомологичный белку по пп.(g)-(i), описанный выше; и(l) полинуклеотид, способный гибридизоваться в жестких условиях с любым полинуклеотидом по пп.(a)-(h). Предпочтительно такой полинуклеотид включает нуклеотидную последовательность поSEQ ID NO 1, нуклеотидную последовательность связывающегося с -амилоидным пептидом белка (ВВР) клона BBP1-fl, внесенного под депозитарным номером АТСС 98617, или полинуклеотид, кодирующий связывающийся с амилоидным пептидом белок (ВВР), кодируемый вставкой кДНК клона BBP1-fl, внесенного под депозитарным номером АТСС 98617. Другой вариант представляет ген, соответствующий последовательности кДНК по SEQ ID NO 1. В других вариантах настоящее изобретение представляет композицию, включающую белок, при том, что упомянутый белок включает аминокислотную последовательность, выбираемую из группы, которая включает:(a) аминокислотную последовательность по SEQ ID NO 2;(b) аминокислотную последовательность по SEQ ID NO 2 от 68-й аминокислоты до 269-й аминокислоты;(c) аминокислотную последовательность,кодируемую вставкой кДНК клона BBP1-fl, внесенного по депозитарным номером АТСС 96817; и(d) фрагменты аминокислотной последовательности по SEQ ID NO 2, включающие аминокислотную последовательность от 185-й аминокислоты до 217-й аминокислоты по SEQ IDNO 2. Предпочтительно такой белок включает аминокислотную последовательность по SEQ IDNO 2 или аминокислотную последовательность по SEQ ID NO 2 от 68-й аминокислоты до 269-й аминокислоты. Слитые белки также формулируются настоящим изобретением. В ряде предпочтительных вариантов полинуклеотид функционально присоединен к последовательности, регулирующей экспрессию. Также настоящее изобретение представляет клетку-хозяин, включая бактериальные, дрожжевые клетки, клетки насекомых и млекопи 004256 4 тающих, трансформированные такими полинуклеотидными композициями. Также представляются способы выработки ВВР, которые включают (а) выращивание культуры клеток-хозяев по п.3 формулы изобретения в подходящей культуральной среде; и (b) очистку белка из культуральной среды. Также настоящим изобретением представляются композиции, включающие антитело,специфичное в отношении таких ВВР. Способы и диагностические приемы также представляются в связи с выявлением заболевания, характеризующегося аномальной экспрессией человеческого белка ВАР, равно как и способы идентификации соединений, участвующих в регуляции активности ВВР. Другой вариант настоящего изобретения включает трансгенных животных, несущих полинуклеотид, кодирующий ВВР, функционально соединенный с последовательностью, регулирующей экспрессию. Краткое описание чертежей Следующие чертежи характеризуют некоторые варианты настоящего изобретения. Они являются исключительно иллюстративными и ни в чем не ограничивают настоящее изобретение. Фиг. 1. Тест на дигибридный дрожжевой скрининг. Хозяинный штамм Y2H, экспрессирующий ДНК-связывающий домен Gal4, слитый с BAF42 (ВАРBD; плазмида, несущая маркерY2 Н-библиотекой кДНК клеток плодного головного мозг человека (плазмида, несущая маркер LEU2), экспрессирующей слитые белки("unknownAD"), включающие активаторный домен Gal4, в соответствии с описанным в данном тексте. Соответственно, штаммы, несущие три внехромосомные плазмиды, обозначенные окружностями, экспрессировали обозначенный белок. Положительные межбелковые взаимодействия восстанавливают активность Gal4 по расположенной выше активаторной последовательности (GALUAS), в результате чего индуцируется транскрипция репортерного гена HIS3. Фиг. 2. Выявление связывания ВВРl/ВАР. Штаммы Y2H были тестированы на прототрофию по гистидину путем получения 10-кратных серийных разведении и раскапыванием по 5 мкл на синтетическую агаровую среду, лишенную триптофана, лейцина, гистидина и содержащую 25 мМ 3-аминотриазола, в соответствии с описанным в данном тексте. Все штаммы несут плазмиду рЕК 162, экспрессирующую слитый белок ВАР, что отмечено меткой "ВАР". Первые столбцы ("вектор") соответствуют независимо сформированным штаммам, несущим плазмиду рЕК 162 и вектор рАСТ 2, экспрессирующий посторонний слитый белок. Этот вариант служит в качестве фона для сравнения со штаммами, которые экспрессируют интересующие белки. Ко 5 лонки, обозначенные "BBP1tm", соответствуют экспрессии укороченного ВВР 1 с плазмиды рЕК 198 так, как это описано в настоящем тексте. Взаимодействие между слитыми белками ВАР и BBP1tm восстанавливает активностьGal4, в результате чего происходит индуцирование репортерного гена HIS3 (см. фиг. 1), что выявляется по усилению прототрофного роста в сравнении с контрольными штаммами. Фиг. 3. Биотесты, показывающие взаимодействие ВВР 1 с G-белками. Предполагаемый внутриклеточный домен белка ВВР 1 экспрессировали в качестве ДНК-связывающего домена Gal4, соединенного с частями факторовGs, Go или Gi2 крысы, экспрессированными в качестве слитых белков, включающих активаторный домен Gal4. Ответы Y2H,представляющих два независимо образованных клона каждого штамма, сравнивали с ответами клеток, лишенных G-белкового компонента (колонки "вектор"). Методика описана в легенде к фиг. 2. Фиг. 4. Картирование взаимодействий между ВВР 1 и ВАР. BBP1tm был разделен на два перекрывающихся сегмента в соответствии с описанным в данном тексте. Эти белки, обозначенные ВВР 1 С и BBP1N, были протестированы по их взаимодействию с ВАР. Протокол тестирования и штаммы, обозначенные как"BBP1N", экспрессируют обозначенный сегмент ВВР 1 в виде слитого белка. Фиг. 5. Экспрессия мРНК ВВР 1 в тканях человека (А) и участках головного мозга (В). Нейлоновые мембраны, на которые было нанесено по 2 мкг фракционированной по размеру полиаденилированной РНК, выделенной из обозначенных тканей, были получены от Clontech. Их гибридизовали с радиоактивно помеченным зондом кДНК-ВВР 1 так, как это описано в данном тексте. Основной бэнд, соответствующий 1250 пар нуклеотидов (определено по маркерам молекулярной массы: данные здесь не включены) был выявлен во всех линиях. Более высокие значение молекулярной массы, по-видимому,соответствуют гетероядерным РНК; ген ВВР 1 включает несколько интронов. Блоты были освобождены и повторно зондированы -актином с целью контроля загрузки и интегрированности РНК: все линии проявили одинаковый сигнал(данные здесь не включены). Фиг. 6. Экспрессия ВВР 1 и АРР в клетках гиппокампа. Представлены изображения авторентгенограмм, полученных в ходе гибридизации in situ и показывающих параметры экспрессии ВВР 1 (А) и АРР (В) в гиппокампе и в энторинальной области коры головного мозга человека. Срезы тканей, использовавшиеся для получения данных изображений, были взяты при проведении вскрытия двух различных пациен 004256 6 тов. Сокращения: DG - зубчатая извилина коры головного мозга; СА 1 - нижняя область гиппокампа; ЕС - эн-торинальная область коры головного мозга. Фиг. 7. Сравнение взаимодействия ВВР 1 с ВАР человека и грызуна. ВАР грызуна был сконструирован и экспрессирован в виде слитого белка в соответствии с описанием в данном тексте. Штаммы, помеченные "ВАР человека",идентичны тем, которые были показаны на фиг. 2. Штаммы, помеченные "ВАР грызуна", экспрессируют ВАР грызуна в виде слитого белка,включающего ДНК-связывающийся домен Gal4."Вектор" определяет контрольные штаммы, несущие только вектор в противоположность слитым белкам ВАР; "ВВР 1" определяет штаммы,экспрессирующие слитый белок BBP1tm. Подробное описание изобретения Настоящее изобретение касается выделения и клонирования белка (ВВР 1), связывающегося с -амилоидным пептидом человека. ВВР 1 был охарактеризован как слитый белок в дигибридном дрожжевом тесте по связыванию с 42 аминокислотным фрагментом ВАР (ВАР 42). Экспрессия ВВР 1 была продемонстрирована в тканях человека и в конкретных отделах головного мозга (фиг. 5). Важно, что для ВВР 1 показана избирательность в связывании с ВАР человека в системе дигибридного дрожжевого теста в сравнении с ВАР грызуна. Эти данные подтверждают предположение о том, что ВВР 1 по настоящему изобретению могут быть использованы в диагностике и лечении болезни Альцгеймера, равно как и для оценки и скрининга лекарственных средств, предназначенных для регуляции накопления амилоидных бляшек в головном мозге. Кодирующая последовательность ВВР 1 Исходный клон ВВР 1 человека (обозначенный как "клон 14") был получен с использованием дигибридного дрожжевого генетического теста (Y2H), разработанного для целей идентификации белков, которые взаимодействуют с ВАР 42 человека, являющимся потенциально более токсичной формой ВАР. ВАР 42 был экспрессирован при слиянии с ДНК-связывающимся доменом Gal4 дрожжей и также был экспрессирован в виде свободного пептида (фиг. 1). Этот штамм был трансформирован Y2 Нбиблиотекой кДНК плодного головного мозга человека. Единственный клон, обозначенный как " 14", из примерно миллиона независимых трансформантов, обусловливал отчетливую активацию репортерного гена и включают открытую кодирующую рамку, переходящую в доменGAL4. кДНК-вставка включала 984 пары оснований и заканчивалась полиадениловым трактом. Эта последовательность кодировала 201 аминокислоту (от 68-й аминокислоты до 269-й аминокислоты в соответствии с SEQ ID NO 2) и имела два участка, характеризующиеся длиной 7 и уровнем гидрофобности, достаточными для проникновения через клеточную мембрану. Также в ее составе имеются потенциальные сайты гликозилирования по остаткам аспарагина(N-гликозилирования). "Клон 14" был обозначен как клон рЕК 196 и внесен в Американскую коллекцию типовых культур под депозитарным номером АТСС 98399. Производная от библиотеки плазмида была выделена из клона 14 и использована для реконструкции штаммов, включенных в тест Y2H. Тестирование этих штаммов показало, что слитый белок ВАР специфическим образом взаимодействует с белком, кодируемым клоном 14,хотя ответ оказался достаточно слабым. С учетом того, что обладающие высоким уровнем гидрофобности белковые домены, такие как трансмембранные участки, подавляют ответы в тесте Y2H (Ozenberger, неопубликованные данные), то клон 14 был укорочен (с получением варианта BBP1tm: см. ниже табл. 2 в связи с дальнейшим описанием) с целью удаления участка жесткой гидрофобности с последующим повторным тестированием по взаимодействию с ВАР. Существенно более мощный ответ в тестеY2H был отмечен с вариантом BBP1tm: это подтверждает замечание о том, что делегированные последовательности кодировали вероятный трансмембранный "якорь" ("tm"). Клон 14 идентифицирует новый ВАР-связывающийся белок в форме слитого белка. Последовательности кДНК ВВР 1, находящиеся в клоне 14, были идентифицированы как утратившие 5'-кодирующий участок, на что указывает отсутствие потенциального инициирующего кодона метионина. Многочисленные попытки применения стандартного метода 5'RACE ("быстрая амплификация концов кДНК") с применением обычной обратной транскриптазы позволили добавить всего 27 нуклеотидов. Таким образом, методика геномного клонирования, описанная ниже в примере 2, была применена для выделения 5'-концевого участка. Поскольку последовательность, кодирующая 5'-концевой участок, является производной от геномной библиотеки, существует опасность присутствия в этой последовательности интрона. Такая вероятность была оценена с применением двух методов, описанных ниже в примере 2. Полученные в результате данные подтверждают, что точность выделения лежащих выше последовательностей (как из геномных, так и из кДНК-источников) и отсутствие интронов в этом участке. Плазмида BBP1-f1, включающая вставку кДНК, кодирующую полноразмерный белок ВВР 1, была заключена в Американскую коллекцию типовых культур под депозитарным номером АТСС 98617. Полный кодирующий участок и расшифрованная последовательность полипептида показаны в SEQ ID NO 1 и SEQ ID 8 14 (рЕК 196) включаются 3'-нетранслируемые нуклеотидные последовательности. В соответствии с настоящим изобретением нуклеотидные последовательности, которые кодируют ВВР 1, его фрагменты, слитые белки или функциональные эквиваленты, могут быть использованы для формирования рекомбинантных молекул ДНК, которые бы направляли экспрессию ВВР 1, или функционально активного пептида в подходящей клетке-хозяине. С другой стороны, нуклеотидные последовательности,которые гибридизуются с фрагментами последовательности ВВР 1, могут быть использованы в тестах по гибридизации нуклеиновых кислот,в Саузерн-блоттинге, в Нозерн-блоттинге и т.п. Настоящее изобретение также представляет полинуклеотиды, последовательности которых комплементарны описанным в данном тексте полинуклеотидам. Также настоящее изобретение представляет полинуклеотиды, способные гибридизовать в условиях сниженной жесткости, более предпочтительно при условиях обычной жесткости и наиболее предпочтительно в условиях высокой жесткости с описанными в данном тексте полинуклеотидами. Примеры условий жесткости гибридизации показаны ниже в таблице: условиями высокой жесткости являются такие условия, которые сопоставимы с теми, которые приведены, например, в графах A-F; условия обычной жесткости являются по крайней мере настолько жесткими, насколько это показано, например, в графах G-L; и условия сниженной жесткости по крайней мере таковы, как это показано, например, в графах M-R. Условия жесткости УслоТемпература Длина Полинуквия и буфер гибрида леотидный жест(пар нук- гибридизагибрид кости ции леотидов)1 А ДНК/ДНК 65 С - 1xSSC; 50 или 42 С 1xSSC; 50% формамид В ДНК/ДНК 50 ТB-1xSSC С ДНК/РНК 67 С - 1xSSC; 50 или 45 С 1xSSC; 50% формамид.- под длиной гибрида понимается длина гибридизуемого участка (участков) гибридизующегося полинуклеотида. В случае, когда происходит гибридизация полинуклеотида с полинуклеотидом с неизвестной последовательностью, то "длина гибрида" подразумевает длину всего исходного гибридизующегося полинуклеотида. Когда гибридизуют полинуклеотиды с известными последовательностями, то длина гибрида может быть определена путем сопоставления последовательностей полинуклеотидов и идентификации участка или участков, с оптимальной комплементарностью последовательности.- при составлении гибридизационного и промывочного буферов раствор SSC (1xSSC содержит 0,15 М NaCl, 15 мМ цитрата натрия) может быть заменен на раствор SSPE (1xSSPE содержат 0,15 М NaCl, 10 мМ NaH2PO4 и 1,25 мМ EDTA - рН=7,4); промывку осуществляют на протяжении 15 мин после завершения гибридизации; ТB-ТR - температура гибридизации для гибридов, длина которых менее 50 пар нуклеотидов, должна быть на 5-10 С ниже, чем температура плавления (Тm) данного гибрида, при том, что Тm определяют в соответствии со следующими уравнениями: для гибридов длиной менее 18 пар нуклеотидов: Tm(C) = 2x (число оснований А+Т) + 4 х(число оснований G+C). Для гибридов длиной от 18 до 49 пар нуклеотидов: Tm(C) = 81,5 + 16,6 (log10[Na+]) + 0,41 (%[G+C])-(600/N), где N - число оснований в гибриде, a [Na+] - концентрация ионов натрия в гибридизационном буфере (для раствора IxSSC[Na+] =0,165 М). Дополнительные примеры условий жесткости для гибридизации полинуклеотидов представлены у Sambrook J., Fritsch E.F.Маniatis Т., 1989, "Molecular Cloning: A Laboratory Manual", Cold Spring Harbor Lab. Press," Cold SpringJ.WileySons Inc. (разделы 2.10 и 6.3-6.4),включенные в данный текст в виде библиографических ссылок. Предпочтительно, чтобы каждый из таких гибридизующих полинуклеотидов имел длину,составляющую по крайней мере 25% (более предпочтительно - по крайней мере 50%, а наиболее предпочтительно - по крайней мере 75%) длины полинуклеотида по настоящему изобретению, с которым он должен гибридизоваться, а также характеризовался по крайней мере 60%ным уровнем идентичности последовательности(более предпочтительно - по крайней мере 75%ным уровнем идентичности, а наиболее предпочтительно по крайней мере 90%-ным или 95%-ным уровнем идентичности) с полинуклеотидом по настоящему изобретению, с которым он должен гибридизоваться, при том,что уровень идентичности определяют путем сравнения аминокислотных последовательностей при таком их сопоставлении, которое максимально обеспечивает перекрывание и идентичность и минимизует разрывы последовательностей. Экспрессия ВВР 1 Выделенный полинуклеотид по настоящему изобретению может быть функционально присоединен к контрольной последовательности, регулирующей экспрессию, такой как экспрессионные векторы рМТ 2 или pED, описанные Кауфманом с соавт. (Kaufman et al., 1991,Nucl. Acids Res., 19, 4485-4490), с целью получения белка рекомбинантным путем. Многие контролирующие экспрессию последовательности известны в данной области техники. Также хорошо известны основные методы экспрессии рекомбинантных белков: их примеры можно найти у Кауфмана (Kaufman, 1990, Methods Enzymol., 185, 537-566). По использованию в данном тексте термин "функционально присоединенный" обозначает то, что выделенный полинуклеотид по настоящему изобретению и регулирующая экспрессию последовательность взаиморасположены в составе вектора или в клетке таким образом, чтобы данный белок экспрессировался той клеткой-хозяином, которая была трансформирована (трансфицирована) лигированных друг на друга полинуклеотида и регулирующей экспрессию контрольной последовательности. Экспрессионная система для ВВР 1 Большое число типов клеток может быть клетками-хозяевами, пригодными для экспрессии конкретного белка. Клетки-хозяева млекопитающих включают, например, клетки COS зеленой мартышки, клетки СНО яичников китайского хомячка, клетки 293 почек человека,эпидермальные клетки А 431 человека, клетки Соlо 205 человека, клетки 3 Т 3 мыши, клеткиCV-1, другие линии трансформированных клеток приматов, нормальные диплоидные клетки,клетки линий, выделенных от культивируемыхin vitro первичных тканей, первичных эксплантатов, клетки HeLa, клетки L мыши, клетки линий ВНК, HL-60, U937 НаК или Jurkat. С другой стороны, возможным является синтезировать белок в клетках низших эукариот, таких как дрожжи, или в прокариотах, таких как бактерии. Потенциально подходящими дрожжевыми штаммами являются штаммы видовcerevisiae,Schizosaccharomyces ponibe, штаммы Kluyveromyces,Candida или любые другие дрожжевые штаммы,способные вырабатывать чужеродные белки. Потенциально пригодными бактериальными штаммами являются штаммы Escherichia coli,Bacillus subtilis, Salmonella typhimurium или любой другой бактериальный штамм, способный экспрессировать гетерологичные белки. Если белок был синтезирован в клетках дрожжей или бактерий, то его необходимо модифицировать,например, по подходящим сайтам фосфорилирования или гликозилирования с целью получения функционально активного белка. Такое ковалентное присоединение может быть осуществлено с использованием известных химических или ферментных методов. Белок также может быть получен путем функционального соединения выделенного полинуклеотида по настоящему изобретению с подходящими контрольными последовательностями в составе одного или большего числа экспрессионных векторов для клеток насекомых с использованием экспрессионной системы клеток насекомых. Материалы и методы для осуществления экспрессионной системы "бакуловирус/клетки насекомого" доступны на коммерческой основе в виде стандартных наборов,производимых, например, фирмой In-vitrogen(San Diego, CA, США) - набор МахВас 7: такие методы хорошо известны в данной области техники и описаны, в частности, Саммерсом и Смитом (SummersSmith, 1987, Texas Agricult.Exper. Stat. Bull., N 1555: включено в данный текст в виде библиографической ссылки). По использованию в данном тексте клетка насекомого, способная экспрессировать полинуклеотид по настоящему изобретению, называется"трансформированной". Белок по настоящему изобретению может быть получен путем культивирования трансформированных клеток-хозяев в таких условиях, которые являются подходящими для экспрессии рекомбинантного белка. Полученный в результате экспрессированный белок затем может быть очищен из такой культуры (например,из культуральной среды или из клеточного экстракта) с использованием известных процедур очистки, таких как гель-фильтрация и ионобменная хроматография. Очистка белка также может включать хроматографию на аффинных колонках, содержащих агенты, которые бы связывались с данным белком: один или большее число этапов очистки на колонках с такими аф 004256 12 финными смолами, как конканавалин-А-агароза,гепарин-tоуореаrl-7 или сефароза-7 с цибахромом голубым 3GA; один или большее число этапов хроматографии с гидрофобным взаимодействием с использованием таких смол, как фениловый эфир, бутиловый эфир или пропиловый эфир,или иммуноаффинная хроматография. С другой стороны, белок по настоящему изобретению также может быть экспрессирован в такой форме, которая бы облегчила его очистку. Например, он может быть экспрессирован в виде слитого белка, такого как мальтозосвязывающегося белка (МBР), глутатион-Sтрансферазы (GST) или тиреодоксина (TRX). Стандартные наборы для экспрессии и очистки таких слитых белков доступны в продаже, как продукция фирм, соответственно, New EnglandBioLab (Beverly, MA), Pharmacia (Piscataway,NJ) и Invitrogen. Также белок может быть помечен эпитопом и последовательно очищен с использованием специфичного антитела, направленного на этот эпитоп. Один из таких эпитопов("Flag") доступен на коммерческой основе на фирме Kodak (New Haven, CT). Наконец, для дальнейшей очистки белка могут быть использованы один или большее число этапов высокоразрешающей жидкостной хроматографии (ВЭЖХ) в обратной фазе с использованием гидрофобной среды для ОФВЭЖХ - такой как силикагель, включающий метильную или другую алифатическую боковую цепь. Все или некоторые из названных выше этапов очистки в различных сочетаниях также могут быть применены для получения в существенной степени гомогенного выделенного рекомбинантного белка. Очищенный таким образом белок, по сути свободен от других белков млекопитающих и в соответствии с настоящим изобретением определяется как "выделенный белок". Белок по настоящему изобретению также может быть экспрессирован в виде продукта,вырабатываемого трансгенным животным: например, в виде компонента молока у трансгенных коров, коз, свиней или овец, которые бы характеризовались соматическими или половыми клетками, несущими нуклеотидную последовательность, кодирующую белок. Также белок может быть получен с помощью стандартного химического синтеза. Методы конструирования белков по настоящему изобретению синтетическим путем хорошо известны специалистам в данной области техники. Созданные синтетическим путем последовательности белков за счет гомологии первичной,вторичной или четвертичной структурных и(или) конформационных параметров может обеспечить общие биологические свойства,включая активность белка. Таким образом, они могут быть использованы в качестве биологически активных или иммунологических замести 13 телей естественных очищенных белков при проведении скрининга лекарственных соединений и иммунологических процессов, связанных с формированием антител. Представляемые здесь белки также включают белки, характеризующиеся аминокислотными последовательностями, сходными с таковыми у очищенных белков, но при этом имеющими естественно возникшие или искусственно созданные модификации. Например, модификации в пептидных последовательностях или последовательностях ДНК могут быть выполнены с применением методов, хорошо известных специалистам в данной области техники. Представляющие интерес модификации белковых последовательностей могут включать замещения, замены, вставки или делеции выбранного аминокислотного остатка в составе кодирующей его последовательности. Например, один или большее число остатков цистеина могут быть делегированы или замещены другой аминокислотой с целью изменения конформации данной молекулы. Методики таких замещений, замен, вставок или делеции хорошо известны специалистам в данной области техники (см., например,патент США 4518584). Предпочтительно,чтобы такие замены, замещения, вставки и делеции сохраняли желательную активность данного белка. Другие фрагменты и производные последовательностей белков, которые предположительно будут сохранять активность белка полностью или отчасти и таким образом могут быть использованы для скрининга или иных иммунологических методик, также могут быть легко сформированы специалистами в данной области техники на основе описанного в данном тексте. Такие модификации также охватываются масштабом настоящего изобретения. Дигибридные дрожжевые тесты Тесты Y2H показали, что связывание ВАР со слитым белком ВВР 1 является специфичным. Связывание ВВР 1 с ВАР подтверждает, что активность ВВР 1 может играть определенную роль в патогенезе болезни Альцгеймера. Последовательности ВВР 1 сравнивали с данными GenBank, используя программу для локального сопоставления-поиска (BLAST:Altschul et al., 1990). Были сопоставлены белок ВВР 1 и трансляты включенных в базу данных экспрессируемых маркеров с последующим поиском консервативных участков и оценкой их с помощью программы для поиска специфических полипептидых мотивов (MoST: Tatusov etal., 1994). Такой анализ позволил установить возможность эволюционного родства с семейством G-связанных белковых рецепторов (GPCR). Конкретно этот анализ показал, что ВВР 1 включает два потенциальных трансмембранных домена, эквивалентных трансмембранным доменам 3 и 4, имеющимся в составе G-связанных белковых рецепторов. Расположенная между 14 ними гидрофильная петля включает хорошо известный мотив, состоящий из трех аминокислот - остаток аспарагиновой (D) или глутаминовой кислоты, далее аргинин (R) и затем ароматический остаток (Y или F) (обычно обозначаемый как последовательность "DRY"), который консервативен у почти всех членов данного рецепторного семейства, и для него ранее было показано участие в молекулярном механизме опосредования активации G-белка (AcharyaKarnik, 1996). Данные тестов Y2H (см. фиг. 2-4) показывают, что белок ВВР 1 представляет новый белок, потенциально включающий функциональный модуль, общий с представителями суперсемейства G-связанных белковых рецепторов. Конкретно предполагается, что белок ВВР 1 сохраняет важную последовательность DRF (от 199-й аминокислоты до 201-й аминокислоты в составе SEQ ID NO 2) между двумя предсказываемыми трансмембранными доменами и может обладать потенциалом по связыванию с Gбелком, участвующим в регуляции соответствующего сигнального механизма. Для АРР было показано функциональное связывание с Go (Nishimoto et аl., 1993; Yamatsuji et al., 1996), a BBP1 включает структурный мотив, для которого известно то, что он является активаторной последовательностью для G-белка, находясь в составе родственныхG-связанных белковых рецепторов. Кроме того,гипотеза, основанная на предполагаемом расположении и ориентации трансмембранных доменов в составе BBP1, предполагает, что участок белка, который взаимодействует с ВАР, должен быть топологически соответствующим тому же положению, что и ВАР в АРР. Штаммы для теста Y2H были сконструированы с целью оценки связывания внутриклеточного сегмента BBP1 с G-белками. Предполагаемые внутриклеточные последовательности в составе BBP1 были экспрессированы в виде слитого белка и оценены по их взаимодействию с С-концевыми участками трех G-белков. Сегменты белка, которые были использованы в этих экспериментах, перечислены ниже в табл. 2. Внутриклеточная петля в составе BBP1 взаимодействовала со всеми тремя G-белками(фиг. 3), подтверждая тем самым предположение о том, что BBP1 может функционировать в качестве модулятора активности G-белков. Эти различные Y2 Н-тесты предполагают представляющую интерес модель множественного белкового комплекса, по минимуму состоящего из взаимодействующих мембранных белков BBP1 и АРР, связанных с гетеротримерным G-белком. Таблица 2. Плазмиды, использовавшиеся в дигибридных дрожжевых тестах Экспрессирующая плазмида рЕК 162 рЕК 240 рЕК 196 рЕК 198 рЕК 219 рЕК 216 Дальнейший анализ ВВР 1 был осуществлен с помощью тестов Y2H. Две перекрывающиеся части последовательностей ВВР 1, содержащиеся в составе клона BBP1tm, были амплифицированы и клонированы в состав Y2 Нвектора рАСТ 2 (экспрессирующие плазмиды рЕК 216 и рЕК 219 [табл. 2] и соответствующие им белки BBP1N и ВВР 1 С [фиг. 4]). Конструкция С характеризуется отсутствием обоих трансмембранных доменов, а конструкция N кодирует первый трансмембранный домен плюс предшествующие ему 52 аминокислоты. Эти слитые белки были оценены с использованием ВАР-слитых белков, а полученные ответы сравнивали с ответами штаммов, экспрессирующих более длинный вариант белка BBP1tm. БелокBBP1tm индуцирует слабый ответ в тесте Y2H(сравнивая ВВР 1 С с вектором: фиг. 4), однако,белок BBP1N, включающий первый трансмембранный домен и прилежащие N-концевые последовательности, обусловливает ответ, который лишь немного слабее, чем тот, который получен для белка BBP1tm (фиг. 4). Эти данные подтверждают, что основной детерминант связывания с ВАР находится в пределах того участка ВВР 1, который предположительно топологически сходен с ВАР в составе белка АРР дикого типа. Система Y2H была использована для того,чтобы показать избирательность и специфичность связывания ВВР 1 с ВАР человека в сравнении с ВАР грызуна. В составе аминокислотной последовательности ВАР грызуна имеется три аминокислотные замены (G5R, F10Y иR13H) по сравнению с последовательностью ВАР человека. В тесте Y2H, описанном в примере 6, белок грызуна проявляет сниженную нейротоксичность и отсутствие связывания с гомогенатами ткани головного мозга человека(Maggio et al., 1992). Интересно, следовательно,было бы оценить связываемость ВАР грызуна с ВВР 1 в системе Y2H. Последовательность ВАР человека в составе рЕК 162 была изменена таким образом, чтобы кодировать пептид грызуна, с применением опосредованного олигонуклеоти 16 дами мутагенеза с помощью ПЦР. Полученная в результате плазмида рЕК 240 была идентична плазмиде, экспрессирующей слитый белок ВАР человека, использованной в данном эксперименте, за исключением трех кодонов, продуцирующих три аминокислотные замены, соответствующие пептидной последовательности грызуна. Взаимодействия между слитым белком ВВР 1 и слитыми белками ВАР человека и грызуна сопоставляли с помощью биотеста Y2H. Штаммы, экспрессирующие ВВР 1 и ВАР грызуна, не проявляли способности к ростовому ответу (фиг. 7). Эти данные подтверждают предположение о том, что ВВР 1 может являться специфическим медиатором нейротоксического действия ВАР, а также указывает на механизм,объясняющий сниженную нейротоксичность ВАР грызуна. Важно также, что полученные данные демонстрируют высокую степень специфичности взаимодействия ВВР 1/ВАР в тестахY2H при том, что замены трех аминокислот были достаточны для полного нарушения такой связи (фиг. 7). Выделенные полипептиды ВВР 1 Белки и фрагменты белков по настоящему изобретению включают белки, аминокислотные последовательности которых по длине по крайней мере на 25% (более предпочтительно по крайней мере на 50%, а наиболее предпочтительно по крайней мере на 75%) соответствуют длине представляемого белка, и которые характеризуются по крайней мере 60%-ным уровнем идентичности последовательностей(более предпочтительно по крайней мере 75%-ным уровнем идентичности, а наиболее предпочтительно по крайней мере 90%-ным или 95%-ным уровнем идентичности) с последовательностью представляемого белка, при том, что уровень идентичности определяют путем сравнения аминокислотных последовательностей при таком их сопоставлении, которое максимально обеспечивает перекрывание и идентичность и минимизует разрывы последовательностей. Также в настоящее изобретение включены белки и фрагменты белков, которые включают сегмент, предпочтительно включающий 8 или больше (более предпочтительно 20 или больше,а наиболее предпочтительно - 30 или больше) расположенных подряд аминокислот, который проявляют по крайней мере 75%-ный уровень идентичности последовательностей(более предпочтительно по крайней мере 85%-ный уровень идентичности, а наиболее предпочтительно по крайней мере 95%-ный уровень идентичности) с любым таким сегментом любого представляемого белка. Также настоящим изобретением представляются видовые варианты, гомологичные заявляемым полинуклеотидам и белкам. По использованию в данном тексте видовой гомологичный вариант - это белок или полинуклеотид,происходящий от другого вида в сравнении с 17 данным белком или полинуклеотидом, но характеризующийся существенным уровнем сходства последовательности по сравнению с данным белком или полинуклеотидом. Предпочтительно, чтобы видовой гомологичный вариант полинуклеотида характеризовался по крайней мере 60%-ным уровнем идентичности последовательностей (более предпочтительно по крайней мере 75%-ным уровнем идентичности, а наиболее предпочтительно по крайней мере 90%-ным уровнем идентичности) по сравнению с данным полинуклеотидом, а видовой гомологичный вариант белка характеризовался по крайней мере 30%-ным уровнем идентичности(более предпочтительно по крайней мере 45%ным уровнем идентичности, а наиболее предпочтительно по крайней мере 60%-ным уровнем идентичности) по сравнению с данным белком,при том, что уровень идентичности последовательностей определяют путем сравнения нуклеотидных последовательностей полинуклеотидов или аминокислотных последовательностей белков при таком их сопоставлении, которое максимально обеспечивает перекрывание и идентичность и минимизирует разрывы последовательностей. Видовые гомологичные варианты могут быть выделены и идентифицированы путем формирования подходящих зондов или затравок на основе представленных здесь последовательностей и скрининга подходящего источника нуклеиновых кислот, соответствующих желательному виду. Предпочтительно,чтобы видовые гомологичные варианты были выделены у видов млекопитающих. Более предпочтительно, чтобы видовые гомологичные варианты были выделены у таких видов млекопитающих, как, например, шимпанзе Pan troglodytes, горилла Gorilla gorilla, орангутанг PongoMus musculus, крыса Rattus norvegicus, китайский хомячок Cricetulus griseus, домашняя кошка Felis catus, американская норка Mustela vison,домашняя собака Canis familiaris, кролик Oryctolagus cuniculus, крупный рогатый скот Bostaurus, домашняя овца Ovis aries, домашняя свинья Sus scrofa и лошадь Equus caballus, для которых уже были созданы генетические карты,позволяющие оценивать отношения синтении между геномной организацией одного вида и геномной организацией соответствующих генов у других видов (O'BrienSeuanez, 1988, Ann.Genome Res., 7, 1123-1137: все статьи включены здесь в виде библиографических ссылок). Также настоящее изобретение представляет аллельные варианты заявляемых полинуклеотидов или белков: они являются естественно встречающимися альтернативными формами выделенных полинуклеотидов, которые также кодируют белки, идентичные или обладающие существенно сходными последовательностями по сравнению с теми, которые кодируются заявляемыми полинуклеотидами. Предпочтительно,чтобы аллельные варианты характеризовались по крайней мере 60%-ным уровнем идентичности последовательностей (более предпочтительно по крайней мере 75%-ным уровнем идентичности, а наиболее предпочтительно по крайней мере 90%-ным уровнем идентичности) по сравнению с данным полинуклеотидом, при том, что уровень идентичности определяют путем сравнения нуклеотидных последовательностей полинуклеотидов при таком их сопоставлении,которое максимально обеспечивает перекрывание и идентичность и минимизирует разрывы последовательностей. Аллельные варианты могут быть выделены и идентифицированы путем использования подходящих зондов или затравок на основе представленных здесь последовательностей и скрининга подходящего источника нуклеиновых кислот от особей соответствующего вида. Также настоящее изобретение представляет полинуклеотиды, последовательности которых комплементарны последовательностям заявляемых здесь полинуклеотидов. Применения Белки ВВР 1 по настоящему изобретению могут быть применены способами, являющимися стандартными для специалистов в данной области техники исходя из изложенного в настоящем тексте. Конкретно белки ВВР могут быть использованы в качестве иммуногенов для повышения количества вырабатываемых антител, которые специфичны в отношении клонированных полипептидов. Различные процедуры,известные в данной области техники, могут быть использованы для получения антител к белкам ВВР 1. Такие антитела включают, тем самым не ограничиваясь, поликлональные, моноклональные, слитые, одноцепочечные антитела, фрагменты Fab и Fab-экспрессионную библиотеку. Для получения антител животныхреципиентов различных видов, включая, но не ограничиваясь этим, кроликов, мышей и крыс,инъецируют белком ВВР. В одном из вариантов полипептид или фрагмент полипептида, обладающий специфической иммунной активностью, соединяют с иммуногенным носителем. Также в сочетании с полипептидом могут быть введены адъюванты с целью усиления иммунного ответа животного-реципиента. Примерами адъювантов, которые могут быть использованы,являются, не ограничиваясь ими, полный и не 19 полный адъюванты Фройнда, минеральные гели, такие как гидроокись алюминия, поверхностно-активные вещества, такие как лизолецитин, плюроновые высокомолекулярные спирты,полианионы, пептиды, масляные эмульсии, гемоцианин слизня и динитрофенол. Моноклональные антитела к белкам ВВР 1 по настоящему изобретению могут быть получены с использованием любой методики, которая бы обеспечивала выработку антител непрерывно культивируемой клеточной линией. Такие методики хорошо известны специалистам в данной области техники и включают, тем самым не ограничиваясь, гибридомную технологию,первоначально описанную Клером и Мильштейном (KohlerMilstein, 1975, Nature, 256,4202-4207), гибридомную методику с использованием В-клеток человека, описанную Косбором с соавт. (Kosbor et al., 1983, Immunol. Today,4, 72), и основанную на трансформации вирусом Эпштейна-Барр гибридомную методику, описанную Коулом с соавт. (Cole et al., "MonoclonalAntibodies and Cancer Therapy", A.R.Liss Inc., pp. 77-96). Антитела, реагирующие с полипептидами по настоящему изобретению, могут быть затем использованы для скрининга на присутствие и субклеточное распределение сходных полипептидов в биологическом образце. Кроме того,моноклональные антитела, специфичные в отношении белков ВВР 1 по настоящему изобретению, могут быть использованы и как лекарственные средства. Белки ВВР 1 также могут быть использованы в качестве антигенов, применимых в твердофазных тестах, нацеленных на измерение присутствия антител, которые являются иммунореактивными в отношении заявленных пептидов. Тесты на конкуренцию в твердой фазе могут быть использованы для имуннологического количественного анализа родственных клону 14 антигенов в биологических образцах. Такой анализ применим не только для облегчения получения полной характеристики клеточной функции или функций полипептидов по настоящему изобретению, но и также могут быть использованы для выявления пациентов,характеризующихся аномальным количеством этих белков. Белки ВВР 1 по настоящему изобретению также могут быть использованы в качестве реагентов-"уловителей" в аффинной хроматографии для выявления собственно ВАР и агрегаций ВАР, являющихся симптомами болезни Альцгеймера. Кроме того, эти белки ВВР 1 применимы в качестве реагентов в тесте, предназначенном для идентификации молекул "кандидатов", которые влияют на взаимодействие ВАР и клонированного белка. Соединения, которые специфически блокируют такое взаимодействие, мо 004256 20 гут быть использованы для лечения или профилактики болезни Альцгеймера. Также эти белки ВВР 1 применимы в тестах на бесклеточное связывание in vitro, в котором определяется изменение под влиянием соединения связывания данных белков, ассоциированных с -амилоидными пептидами, с ВАР или агрегациями ВАР. Бесклеточные тесты особенно применимы для скрининга большого числа соединений, поскольку такие тесты относительно экономичны и просты в осуществлении в сравнении тестами, основанными на использовании живых клеток. С учетом использования полипептидов по настоящему изобретению разработка таких тестов была бы рутинной процедурой для специалиста в данной области техники. В таких тестах либо ВВР 1, либо ВАР снабжают меткой. Такие метки включают, тем самым не ограничиваясь, радиоактивные метки,антитела или флуоресцентные или ультрафиолетовые метки. Связывание ВВР 1 с ВАР или агрегациями ВАР сначала выявляют в отсутствие любого из тестируемых соединений. Затем в тестовую среду добавляют предназначенные к тестированию соединения с целью определения того, влияют ли эти соединения на данное взаимодействие. Примеры Далее настоящее изобретение описывается в форме примеров. Примеры представлены исключительно для того, чтобы проиллюстрировать изобретение путем отсылки к конкретным вариантам. Эти примеры, иллюстрирующие некоторые конкретные варианты настоящего изобретения, не вносят тем самым каких-либо ограничений или пределов масштаба настоящего изобретения. Дигибридная дрожжевая система (здесь и далее обозначаемая как "Y2H"). Экспрессионные плазмиды для Y2H были сконструированы на векторах pAS2 и рАСТ 2 (описаны WadeHarper et al., 1993) и pCUP (описаны OzenbergerY2H. Генетический скрининг. Метод полимеразной цепной реакции (ПЦР) был применен для амплификации и модифицирования последовательностей, кодирующих белок ВАР. Олигонуклеотиды 1 (5'-CC ATG GAT GCA GAACGC TATGAC AAC ACC GC) были использованы для амплификации ВАР с использованием в качестве матрицы pCLL621, являющимся модифицированным АРР-клоном человека (Jacobsen et al., 1994). Амплифицированная ДНК включает кодоны 389-430 (которые кодируют ВАР 42) белка-предшественника АРР с учетом следующих модификаций. Смысловая затравка добавила рестрикционный 5'-сайт NcoI в состав той же транслируемой открытой рамки, в кото 21 рой имеется NcoI-сайт в составе pAS2. Антисмысловая затравка добавила стоп-кодон и сайты HindIII и SalI, необходимые для проведения клонирования. Продукты данной амплификации были лигированы в состав клонирующей системы ТА (Invitrogen Corp., Carlsbad, CA) с последующим удалением с использованием рестриктаз NcoI и SalI. Полученный фрагмент клонировали в состав плазмиды pAS2, расщепленной рестриктазами NcoI+SalI. Полученную в результате плазмиду -рЕК 162 - подтверждали путем секвенирования ДНК по участку соединенияGAL4/BAP. Белок (ВАР 80: фиг. 1), экспрессируемый с плазмиды рЕК 162, включает слитый белок,содержащий ДНК-связывающийся домен дрожжевого белка-активатора транскрипции Gal4 (лишенного последовательностей функциональной активации) с добавлением с С-конца 42 аминокислот ВАР. Экспрессирующую плазмиду формируют таким образом, чтобы она опосредовала экспрессию немодифицированного ВАР 42. Затравку 2 (5'AAGCTTAAG ATG GAT GCA GAA TTC CGA С) объединяли с затравкой 3 в описанном выше методе ПЦР. Продукт, полученный в результате данной амплификации, включает 5'расположенный HindIII-сайт и сигналы инициации трансляции, оптимизованные для экспрессии в клетках Saccharomyces cerevisiae. Опятьтаки фрагмент ДНК был клонирован в состав системы ТА. Затем он был выделен в видеHindIII-фрагмента, который клонировали в состав плазмиды pCUP по HindIII-сайту. Ориентация гена ВАР в составе полученной в результате плазмиды рЕК 149 (ВАР: фиг. 1) была подтверждена с помощью ДНК-секвенирования. Экспрессирующие ВАР плазмиды рК 149 (использующая ген URA3 в качестве селективного маркера) и рЕК 162 (использующая ген TRP1 в качестве селективного маркера) были использованы для трансформации ими дрожжевых клетокхозяев CY770 (OzenbergerYoung, 1995). Штамм, несущий обе эти плазмиды, был обозначен CY2091. Плазмидную библиотеку,включающую фрагменты кДНК, выделенные из клеток плодного головного мозга человека, клонированную в вектор рАСТ 2 (использующий ген LEU2 в качестве селективного маркера) для дигибридной дрожжевой экспрессии, была приобретена в Clontech Lab. Inc. (Palo Alto, CA). Полученный из библиотеки белок на фиг. 1 обозначен как "unknownAD". Эта библиотека была использована для трансформации CY2091. Образцы были распределены в полной синтетической среде (SC) для культивирования дрожжей,лишенной урацила, триптофана и лейцина с целью отбора клеток, несущих все три плазмиды. В среде также отсутствовал гистидин и присутствовал в концентрации 25 мМ 3-аминотриазол,являющийся ингибитором продукта гена HIS3 дрожжей. 3-Аминотризол был использован для снижения низкого уровня конститутивной ак 004256 22 тивности репортерного гена HIS3. Планшеты инкубировали при 30 С в течение 12 дней. Были выделены 24 колонии, проявляющие усиленную прототрофию по гистидину. Трансформационные контроли показали, что в проведенном скрининге были тестированы 106 отдельных клонов. Для оперативного определения состава позитивных клонов был применен метод ПЦР. Общую ДНК выделяли из каждого позитивного штамма с помощью стандартных приемов. Полученный материал был использован в качестве матрицы для ПЦР с затравками 4 (5' TTTAATACCA CTACAAT GGA Т) +5 (5'TTTTCAGTAT CTACGATTCA Т), которые фланкируют клонирующий сегмент библиотечного вектора рАСТ 2. Фрагменты ДНК были лигированы в состав системы ТА и протестированы путем секвенирования ДНК. Содержащуюся в клоне 14 библиотечную плазмиду(так, как это было описано выше) выделяли по бифункциональному пути в E.coli. Была определена нуклеотидная последовательность кДНК человека, что подтвердило состав последовательности исходного ПЦР-продукта. Биотесты. Штаммы выращивали на протяжении ночи в 2 мл среды SC, лишенной лейцина и триптофана до достижения плотности приблизительно 7 х 107 клеток на 1 мл. Клетки подсчитывали, а затем готовили 10-кратные серийные разведения из расчета от 104 до 108 клеток на 1 мл стерильной воды. Эти образцы раскапывали в виде аликвот по 5 мкл на среду SC, лишенную лейцина, триптофана и гистидина и содержащую 25 мМ 3-аминотриазола. Планшеты инкубировали при 30 С в течение 2-3 дней. Позитивные межбелковые взаимодействия были выявлены по увеличению прототрофного роста в сравнении с контрольными штаммами, экспрессирующими слитный белок с ДНК-связывающимся доменом Gal4 плюс посторонний слитный белок с доменом активации транскрипции(или просто включающий вектор рАСТ, в котором отсутствовали какие-либо встроенные последовательности). Эти контрольные штаммы были определены в описанных выше фигурах как пометка "вектор". Данный тест характеризуется высоким уровнем воспроизводимости результатов и пригоден для выявления даже слабой индукции роста, опосредованной специфическим взаимодействием между белкамимишенями. Исходный клон ВВР 1, обозначенный рЕК 196 и внесенный под депозитарным номером АТСС 98399 (обозначенный здесь как"клон 14"), был использован в качестве ПЦРматрицы для укорочения белкового продукта,экспрессируемого BBP1tm. Смысловую затравку 6 (5'-TTTAATACCA CTACAATGGA Т) отжигали на последовательности GAL4 в составе рАСТ 2. Антисмысловая затравка 7(5'-CTCGAG TTA ААА TCG АТС TGC ТСС САА СС) вносила 3'-стоп-кодон и XhoI-сайт сразу с 3'-стороны последовательности, коди 23 рующей DRF-мотив в составе ВВР 1. ПЦРпродукт был лигирован в состав клонирующей векторной системы ТА с последующим вырезанием рестриктазами EcoRI+XhoI и клонировали в плазмиду рАТС 2. Экспрессируемый этой плазмидой (рЕК 198) гибридный продукт был обозначен BBP1tm. Сходным образом затравка 7 была объединена с затравкой 8 (5'GAATT CCA ААА АТА AT GAC GCT ACG) с целью конструирования плазмиды рЕК 216, экспрессирующей BBP1AN. Опять же ПЦРпродукт был лигирован в состав системы ТА, а полученную в результате плазмиду, обработанную рестриктазами EcoRI+XhoI, несущую фрагмент ВВР 1 (кодоны 123-202), в конечном счете лигировали в состав рАТС 2, расщепленную теми же рестриктазами. Вариант ВВР 1 С был сформирован с использованием специфичной для рАТС затравки 6 и антисмысловой затравки 9 (5'-CTCGAG TCA AGA TAT GGGCTT GAA ААА AC). После ТА-клонирования,выделения EcoRI-XhoI-фрагмента и клонирования в состав рАСТ 2 полученная в результате плазмида рЕК 219 экспрессировала ВВР 1 от 68 го по 175-й аминокислотный остаток. Последовательности, кодирующие внутриклеточную петлю BBP1, амплифицировали с использованием олигонуклеотидных зондов 10 (5'ССТТСС ATG GAA GTG GCA GTC GCA TTG ТСТ) +11 (5'-AACACTC GAG TCA ААА ССС ТАС AGT GCA ААА С). Этот продукт,включающий кодоны BBP1 от 185-го по 217-й,расщепляли рестриктазами NcoI+XhoI и клонировали в состав pAS2, расщепляемую NcoI+SalI с получением плазмиды pOZ339. Конструкции плазмид, экспрессирующих все варианты Gaбелков, использовали BamHI-сайт, расположенный вблизи центра каждой последовательности кДНК крысы (Каng et aL, 1980), в качестве сайта для соединения с рАСТ 2. Смысловые затравки отжигали на последовательности, расположенные с 5'-стороны от BamHI-сайта, а антисмысловые затравки отжигали на последовательности 3'-стоп-кодона, включающие рестрикционный сайт SalI. Использованы такие затравки:TGCTTTGAGG GT, антисмысловая ( 22) - 5'GTCGACGGTC TTCTTGCCCC CATCTTCC. ПЦР-продукты были клонированы в состав ТАвектора. Последовательности генов G были выделены в виде BamHI-SalI-фрагментов, которые клонировали в состав рАСТ 2, расщепляемой рестриктазами BamHI+SalI (см. табл. 2 с обозначением плазмид). Наконец, олигонуклеотид 23 был синтезирован для конверсии ВАР 24 человека в последовательность грызуна. Эта затравка имела такую последовательность: 5'ATATGGCCATG GAT GCA GAA TTC GGACAT GAC TCA GGA TTT GAA GTT CGT. Триплеты представляют первые 13 кодонов ВАР: подчеркнуты три нуклеотида, которые были изменены так, чтобы кодировать последовательность грызуна. Олигонуклеотидную затравку 23 объединяли с затравкой 24 (5'TGACCTACAG GAAAGAGTTA), которая отжигается на участок векторов Y2H, расположенный с 3'-стороны от сайта клонирования, в ПЦР с использованием рЕК.162 в качестве матрицы. Полученный продукт расщепляли рестриктазами NcoI+SalI и лигировали в составpAS2 с получением плазмиды рЕК 240. Была подтверждена точность нуклеотидной последовательности в сегменте, кодирующем ВАР грызуна. Геномное клонирование: метод RACE (быстрая амплификация концов кДНК). Геномная клонотека на -фагах (Stratagene), соответствующая a2,0x.106 pfus, была подвергнута скринингу с зондом из случайного праймированного ЕсоRI/-СlаI-фрагмента, соответствующим нуклеотидам 187-600 (фиг. 2). Этот зонд был помечен 32P-ЦТФ с использованием набора реактивов T7QuickPrimer kit в соответствии с рекомендацией производителя (Pharmacia). Фильтры гибридизовались в условиях высокой жесткости: 40 С в 50%-ном формамиде, 0,12 МNaHPO4, 0,25 М NaCl, 7% SDS и 25 мг/мл облученной ультразвуком ДНК спермы лосося и промывали при 65 С в буфере 0,1xSSC, содержащем 0,1% додецилсульфата натрия и сканировали с использованием пленки Kodak BioMaxMS. Клоны -фагов, гибридизовавшие с зондом,очищали путем успешного высевания и повторного скрининга. Десять позитивных клонов были очищены и подвергнуты дальнейшему анализу путем гибридизации с 45-нуклеотидным олигонуклеотидным зондом, соответствующим 5'-последовательности, известной в составе исходного клона кДНК. Этот олигонуклеотид характеризовался обратным составом нуклеотидов 157-201 (фиг. 2) и характеризовался последовательностью-фага была выделена с помощью стандартных молекулярно-биологических методик и подвергнута прямому секвенированию по методу Сэйнджера с использованием автоматического секвенсора ABI-373. Метод RACE. Синтез первой цепи ДНК был осуществлен с использованием системы термостойкой полимеразы rTth (Perkin Elmer). Следующие реагенты были соединены в 1,5-мл пробирке с достижением объема 10 мкл: 1 х обратнотранскриптазный буфер, 1 мМ MnCl2, 1,6 мМ дНТФ (смесь), 2,5 ед. rTth-полимеразы, 100 25 нг поли-А РНК гиппокампа человека (Clontech),10 мМ олигонуклеотида (нуклеотиды 429-452 фиг. 2: 5'-GTTATGTTGG GTGCTG GAAAACAG). Реакционную смесь инкубировали при 70 С в течение 15 мин и немедленно помещали на лед. Набор для синтеза кДНК Marathon(Conthech) был использован для формирования второй цепи кДНК. Полный объем (10 мкл) реакционной смеси первой цепи был дополнен следующими реагентами: 1 х буфера второй цепи, 0,8 мМ смеси дНТФ, 4 х "коктейля" второй цепи (ДНК-полимераза I E.coli, ДНК-лигазаE.coli, РНКаза-Н E.coli) и дистиллированная вода до объема 80 мкл. Пробирку инкубировали при 16 С в течение полутора часов, после чего была добавлена ДНК-полимераза фага Т 4 (10 ед.), и инкубировали в течение еще 45 мин при 16 С. Для остановки реакции 4 мкл 20 х смесиEDTA/гликогена (0,2 М EDTA + 2 мг/мл гликогена) добавляли к реакционным смесям с последующей экстракцией смесью фенола/хлороформа/изо-амилового спирта для удаления ферментов и других примесей. ДНК преципитировали добавлением 0,1 х 3 М ацетата натрия (рН=5,2) и 2,5 х химически чистого этанола с помещением в температуру -70 С. ДНК промывали один раз 70%-ным этанолом, высушивали и ресуспендировали в 10 мкл дистиллированной воды. Половину ДНК использовали для лигирования с адаптером Marathon с последующим использованием в реакциях RACEПЦР в соответствии с протоколом фирмы Clontech следующим образом: 5 мкл кДНК добавляли к 2 мкл (10 мМ) Marathon (5'-CTAATACGACAGGT), 1x буфера ДНК-лигирования и 1 мкл (1 ед.) ДНК-лигазы Т 4. Реакционную смесь инкубировали на протяжении ночи при 16 С. Смесь разводили 1:50 для исходной реакции RACE объединяли в 0,2-мл пробирке для ПЦР со следующими реактивами: 40 мкл дистиллированной воды, 1 мкл 10 х ДНК-полимеразы Klentaq(Clontech), 1 мкл (10 мМ) AP1 затравки (5'ССАТССТААТ ACGACTCACT ATAGGGC), 1 мкл (10 мМ) BBP1-специфичной затравки (соответствующей нуклеотидам 187-209- фиг. 2: 5'CCFGACGGCCA GGCGGCCGCC AT), 5 мкл 10 х буфера полимеразы Klentaq, 1 мкл 10 мМ смеси дНТФ, 1 мкл разведенной кДНК, полученной в предыдущей реакции. Следующие циклические условия были осуществлены с использованием термоциклера GeneAmp 2400(Perkin Elmer): цикл денатурации при 94 С в течение 1 мин с последующими 5 циклами при 94 С в течение 30 с, при 72 С в течение 3 мин, 5 циклами при 94 С 30 с, при 70 С 3 мин, с последующими 25 циклами при 94 С 30 с, при 68 С 3 мин, с конечной достройкой цепи при 72 С в течение 7 мин. После этого этапа осуществляли RACE-ПЦР со "вложенными" затравками по такой схеме: 40 мкл дистиллированной воды, 1 мкл (1 ед.) 10 х ДНК-полимеразы Am 004256RACE-продукта циклические условия проведения ПЦР были такими: исходный цикл денатурации - 9 мин при 94 С, 25 циклов по 30 с при 94 С, 30 с при 68 С, 2 мин при 72 С, с последующей достройкой цепи при 72 С в течение 7 мин. ПЦР-продукт был перенесен в 1%-ный агарозный гель в буфере 1xTBE. Полученный в результате продукт, состоящий из 350 пар нуклеотидов, был очищен в геле и напрямую клонирован с использованием набора для клонирования ТА Cloning kit (Invitrogen). Лигационными смесями трансформировали клетки OneShot(Invitrogen), которые высевали на агар с LBампициллином (100 мкг/мл) на планшеты, содержащие X-gal. Минипрепараты ДНК были получены и подвергнуты секвенированию по методу Сэйнджера на автоматическом секвенсоре ABI-373. Анализ методом Нозерн-блоттинга. Нозерн-блоты мРНК из различных тканей человека и из тканей различных участков головного мозга были получены от Clontech (Palo Alto, CA). Последовательности ВВР 1, продолжающиеся от исходного слитого соединения до полиаденилового сигнала, были выделены в EcoRI-фрагмент из ТА-клона, производного от плазмиды рЕК 196. ДНК -актина была предоставлена производителем. Радиоактивно помеченные зонды были получены на материале таких ДНК с использованием метода случайного прайминга с целью введения 32 Р-дЦТФ (Pharmacia Biotech,Piscataway, NJ). Гибридизация была осуществлена согласно инструкциям производителя(Clontech) в растворе Express Hyb при 68 С. Блоты промывали в буфере 2xSSC (1xSSC - 0,15 М хлорида натрия, 0,015 М цитрата натрия),0,05% SDS при комнатной температуре с последующим промыванием в 0,1xSSC, 0,1% SDS при 50 С. Гибридизационные сигналы были визуализованы с использованием фотопленки KodakBioMax. Гибридизация in situ. Матрицы ДНК для синтеза рибонуклеиновых зондов были получены с помощью ПЦР с использованием плазмидного клона, включающего полноразмерную кДНК ВВР человека. Был использован единственный рибозонд, метящий 3'-UTR (3'нетранслируемый сегмент) данной кДНК. Последовательности зондов были проверены сопоставлением с базой данных GenBank для того,чтобы быть уверенным в том, что они только распознают подходящие метки среди всех депонированных последовательностей. Для формирования рибозондов для ВВР 1 была сконструирована пара олигонуклеотидных затравок, кото 27 рые позволяют амплифицировать участок, состоящий из 275 нуклеотидов от 3'-UTR кДНКBBP1, и добавлять промоторные последовательности к Т 7 (смысловой) и Т 3 (антисмысловой) полимеразе. Эти затравки имели следующие последовательности: 5'-TAATACGACTCAAGTGGCAA CTTGCCTTTG (обратная). ПЦР-продукты были очищены в 1,5%-ном легкоплавком агарозном геле, содержащие продукт бэнды вырезали, экстрагировали фенолом и смесью фенола/хлороформа и преципитировали этиловых спиртом. Осадок высушивали и ресуспендировали в 1 х буфере ТЕ (10 мМ ТрисHCl, 1 мМ EDTA - рН=7,4). АРР-рибозондовая матрица включала DdeI-XhoI-фрагмент из состава кодирующей последовательности в соответствии с описанным у Якобсена с соавт. (Jacobsen et aL, 1991). Пятьдесят нг ДНК-матрицы использовали для реакции транскрипции с использованием меченного 35S-ЦТФ (New EnglandGemini (Promega, Madison, WI). Гистохимическую гибридизацию in situ с использованием срезов ткани гиппокампа человека, полученных post mortem, осуществляли в соответствии с ранее описанным (Rhodes, 1996). Срезы получали толщиной в 10 мкм с использованием криостат-ультратома Hacker-Brights и по растаиванию были распластаны на охлажденные до -20 С предметные сткла, покрытые реагентом Vectabond (Vector Labs, Burlingame, CA). Все растворы были получены в дистиллированной воде, обработаны 0,1%-ным (объемные проценты) диэтил-пирокарбонатом и автоклавированы. Срезы были зафиксированы погружением в 4%-ный параформальдегид в фосфатном буфере (рН=7,4), затем их последовательно погружали в 2xSSC, дистиллированную воду и 0,1 М триэтиламин (рН=8,0). Затем срезы подвергали ацетилированию погружением в 0,1 М триэтаноламин, содержащий 0,25% (объемные проценты) уксусного ангидрида, промывали в 0,2xSSC, обезвоживали в 50%-ном, 70%-ном и 90%-ном этаноле и быстро высушивали. Один мл прегибридизационного раствора, содержащего 0,9 М NaCl, 1 мМ EDTA, 5x раствора Данхардта, 0,25 мг/мл одноцепочечной ДНК спермы сельди (Gibco/BRL, Gaithersberg, MD), 50% деионизированного формамида (ЕМ Sciences,Gibbstown, NJ) в 10 мМ Трис (рН=7,6) были пипеткой раскапаны на каждое предметное стекло,которые затем инкубировали в течение 3 ч при 50 С во влажном инкубаторе. Затем срезы были обезвожены погружением в 50%-ный, 70%-ный и 90%-ный этанол и высушены на воздухе. Помеченные рибозонды добавляли в конечной концентрации 50000 имп/мин на 1 мкл к гибридизационному раствору, содержащему 0,9 М 28 чечной ДНК спермы лосося, декстрансульфат(BSA), 10 мМ дитиотреитола (Boehringer Mannheim, Indianapolis, IN) и 50% деионизированного формамида в 10 мМ Трис (рН=7,6). Затем зонды были денатурированы при 95 С (1 мин), помещены на лед (5 мин) и раскапаны на срезы: они были оставлены для гибридизации на протяжении ночи при 55 С во влажном инкубаторе. Срезы последовательно промывали один раз в течение 45 мин при 37 С в 2xSSC, включающем 10 мМ дитиотреитола, затем один раз в течение 30 мин при 37 С в 1xSSC, содержащем 50% формамида, и один раз в течение 30 мин при 37 С в 2xSSC. Одноцепочечные и неспецифически гибридизованные рибозонды расщепляли путем погружения в 10 мМ Трис (рН=8,0), содержащий РНКазу-А поджелудочной железы быка (Boehringer Mannheim; 40 мг/мл), 0,5 МNaCl и 1 мМ EDTA. Срезы промывали в 2xSSC в течение 1 ч при 60 С, затем - в 0,1xSSC, содержащий 0,5% (весовые проценты) тиосульфата натрия, в течение 2 ч при 60 С. Затем срезы обезвоживали 50-70-90%-ным этанолом, содержащим 0,3 М ацетата аммония, и высушивали. Предметные стекла были заложены в рентгенографические кассеты и подвергнуты контакту сHyperfilm b-Max (Amersham) в течение 14-30 дней. Когда был получен удовлетворительный результат, предметные стекла были покрыты эмульсией для выявления ядерных треков (NTB2; Kodak) и тестированы в течение 7-21 дней при 4 С. Эмульсионные авторадиограммы были получены и зафиксированы в соответствии с инструкциями производителя, а расположенные под эмульсией срезы тканей окрашивали гематоксилином. Для оценки неспецифического мечения контрольный зонд был сформирован на основе матрицы, взятой из набора RiboprobeGemini System kit (Promega). Этот вектор был линеаризован с использованием рестриктазыScaI и транскрибирован с использованием полимеразы Т 3. В полученных транскрипционных реакциях получены два продукта - рибозонды,состоящие из 250 и 1525 нуклеотидов, содержащие только векторную последовательность. Смесь с этим контрольным зондом была помечена так, как это описано выше, добавлена к гибридизационному раствору при конечной концентрации 50000 имп/мин в 1 мкл. На контрольных срезах специфическая гибридизация не была выявлена, т.е. эти срезы дают очень слабый однообразный гибридизационный сигнал, который не соответствует каким-либо нейро-анатомическим маркерам (данные не включены). Пример 1. Клонирование и выделение ВАР-связывающего белка (ВВР 1). Метод дигибридного дрожжевого скрининга (Y2H) был разработан для целей идентификации белков, которые взаимодействуют с ВАР 42 человека - состоящим из 42 аминокислот 29 протеолитический фрагментом АРР, который потенциально существенно более токсичен в агрегационной форме ВАР. ВАР 42 был экспрессирован в виде химеры в соединении с ДНКсвязывающимся доменом Gal 4, а также был экспрессирован в виде свободного пептида (фиг. 1). Данный штамм был трансформирован Y2 Нбиблиотекой кДНК плодного головного мозга человека. Единственный клон, обозначенный, в соответствии с отмеченным выше, как клон 14,из приблизительно миллиона независимых трансформантов, проявлял существенную активацию репортерного гена и по сути включал открытую кодирующую рамку, непрерывно переходящую в последовательность, кодирующую домен GAL4. Вставка кДНК включала 984 пары нуклеотидов и заканчивалась полиадениловым трактом. Последовательность кодировала 201 аминокислоту (аминокислотные остатки 68-269 в SEQ ID NO 2) с наличием двух участков, характеризующихся длиной и уровнем гидрофобности, достаточными для проникновения через клеточную мембрану. Производная от библиотеки плазмида была выделена из клона 14 и использована для реконструкции штаммов, включенных в тест Y2H. Тестирование этих штаммов показало, что слитый белок ВАР специфическим образом взаимодействует с белком, кодируемым клоном 14,хотя ответ оказался достаточно слабым. С учетом того, что обладающие высоким уровнем гидрофобности белковые домены, такие как трансмембранные участки, подавляют ответы в тесте Y2H (Ozenberger, неопубликованные данные), то клон 14 был укорочен (обозначен какBBP1tm) с целью удаления участка жесткой гидрофобности с последующим повторным тестированием по взаимодействию с ВАР. Существенно более мощный ответ в тесте Y2H был отмечен с вариантом BBP1tm (фиг. 2): это подтверждает предположение о том, что делегированные последовательности кодировали вероятный трансмембранный "якорь" ("tm") . Нуклеотидная последовательность клона 14 была скринирована по данным GenBank: в результате ВАР-связывающий белок (ВВР 1) был определен как новый. Пример 2. Выделение и подтверждение 5'конца ВР 1. Последовательности кДНК ВВР 1, включавшие клон 14, описанные в примере 1, были лишены 5'-кодирующего участка, на что указывает отсутствие потенциального инициирующего кодона метионина. Многочисленные попытки применения стандартного метода 5'-RACE ("быстрая амплификация концов кДНК") с применением обычной обратной транскриптазы позволили добавить всего 27 нуклеотидов. Эти последовательности включали триплет ATG, но не включали расположенный выше стоп-кодон в той же транслируемой кодирующей рамке, что 30 подтверждает точность определения этого триплета как инициирующего кодона. Методика геномного клонирования была применена для выделения 5'-концевой части гена ВР 1. Гибридизация геномной -библиотеки человека с зондом, полученным методом случайного прайминга, соответствующим 400 парам нуклеотидов (п.н.) 5'-последовательности клона 14 привела к выявлению 10 позитивных клонов. Дальнейшее охарактеризование этих клонов с использованием 45-мерного олигонуклеотидного зонда, направленного на самую верхнююBPl-последовательность клона 14 (и соответствующей 5'-"верхней" последовательности 400 п.н.), этот зонд показал, что 6 клонов из 10 выделенных включают концевые 5'-последовательности, находящиеся в пределах тех, которые были идентифицированы ранее. Было установлено, что другие четыре -клона представляют другие экзоны, которые входят в состав исходного производного из кДНК зонда, полученного с помощью случайного прайминга 400 п.н. (данные здесь не показаны). Прямое секвенирование ДНК -фага из клонов, соответствующих 5'-концу ВВР 1, представляет примерно 500 нуклеотидов, находящихся "сверху" и перекрывающихся с известным клоном 14. Эта дополнительная последовательность предположительно кодирует 62 аминокислоты, расположенные в N-сторону по отношению к ранее охарактеризованному метионину, выявленному до обнаружения MET, предваряющего внутрирамочный стоп-кодон. Хотя по отношению в "верхнему" MET имеются два других "нижних" остатка метионина, с помощью стандартных подходов заявители экспериментально определили последовательности Nконцевой части гена ВВР 1 человека, как включающую первый из 5'-метиониновых кодонов,следующий за внутрирамочным стоп-кодоном. Полный кодирующий сегмент и расшифрованная аминокислотная последовательность показаны в SEQ ID NO 1 и 2. Плазмида (обозначенная как BBP1-fl), включающая данную аминокислотную последовательность, была внесена в Американскую коллекцию типовых культур под депозитарным номером АТСС 98617. Поскольку 5'-кодирующие последовательности происходили от геномной клонотеки, то существует вероятность того, что этот сегмент включает интроны. Эта вероятность была оценена с применением двух методов. Во-первых,прямая затравка, соответствующая сегменту 5'MET, и обратная затравка, соответствующая участку, находящемуся в пределах клона 14,были использованы для амплификации последовательностей из состава кДНК головного мозга, равно как и геномной ДНК. Из обоих образцов были получены продукты идентичного размера: это указывает на отсутствие интронов в пределах данного участка и подтверждает связь"верхней" последовательности с исходной последовательностью. Во-вторых, последовательности кДНК были выделены в экспериментах на основе модифицированного метода 5'-RACE(см. раздел "Материалы и методы"), которые оказались идентичными тем, которые были выделены из геномного клона. Эти данные подтвердили точность установления "верхних" последовательностей (из источников как библиотеки кДНК, так и геномной клонотеки) и отсутствие интронов в этом сегменте. Пример 3. Характеристика ВВР 1. Последовательности ВВР 1 сравнивали с данными GenBank с применением программы локального сопоставления (BLAST: Altschul etal., 1990). Были идентифицированы две геномные последовательности свободноживущей нематоды Caenorhabditis elegans и одна геномная последовательность дрозофилыmelanogaster, а также большое количество экспрессирующихся маркеров геномов человека,мыши и других видов млекопитающих. Однако не было найдено полных последовательностей кДНК, равно как и каких-либо функциональных данных, связанных с геном. Белок ВВР 1 и продукты трансляции представленных экспрессирующихся маркеров были сопоставлены, тестированы на наличие консервативных сегментов и оценены с применением программы MoST (Tatusov et al., 1994), нацеленной на поиск специфичных белковых мотивов. Эти анализы показали наличие вероятного эволюционной связи с семейством G-связанных белковых рецепторов. Конкретно эти анализы показали, что ВВР 1 включает два потенциальных трансмембранных домена, эквивалентных трансмембранным доменам 3 и 4, имеющимся в составе G-связанных белковых рецепторов. Расположенная между ними гидрофильная петля включает хорошо известный мотив, включающий три аминокислоты - аспарагиновая (D) или глутаминовая кислоты, далее аргинин (R) и затем ароматический остаток (Y или F) (обычно обозначаемый как последовательность "DRY"), который консервативен у почти всех членов данного рецепторного семейства, и для него ранее было показано участие в молекулярном механизме опосредования активации G-белков (AcharyaKarnik,1996). Эти данные указывают на то, что белок ВВР 1 представляет новый белок, потенциально включающий функциональный модуль, общий с членами суперсемейства G-связанных белковых рецепторов. Конкретно, считается, что белок ВВР 1 сохраняет критическую последовательность DRF между двумя его предполагаемыми доменами так, что он может быть потенциальным лигандом G-связанных белковых, участвующих в регуляции сигнального пути. Структурный анализ белка ВВР 1 показал,что он включает структурный мотив, известный как активаторная последовательность G-белка 32 в составе родственных G-связанных белковых рецепторов. Дигибридные дрожжевые тесты,показывающие наличие взаимодействия ВВР 1 с различными членами семейства G-связанных белковых рецепторов, проиллюстрированы на фиг. 3. Основываясь на структурных предсказаниях, белок ВВР 1 определен как белок, дважды проникающий сквозь мембрану обоими своими концами в ком-партмент просвета. Но и другие ориентации этого белка полностью не могут быть отвергнуты. Описанные выше потенциальные межбелковые взаимодействия были исследованы в тестах Y2H. Два перекрывающихся сегмента последовательностей ВВР 1, входящие в состав клона BBP1tm, были амплифицированы и клонированы в состав Y2H-вектора рАСТ 2-табл. 2; и соответствующие белки BBP1N и ВВР 1 С - фиг. 4). Конструкция С утрачивает оба трансмембранных домена, а конструкцияN кодирует первый трансмембранный домен плюс предшествующие 52 аминокислоты. Эти слитые белки были тестированы на взаимодействие со слитыми белками ВАР, а полученные ответы сравнивали с такими ответами штаммов,экспрессирующих больший белок BBP1tm. Полученные результаты подтверждают, что основной детерминант связывания с белком ВАР находится в пределах того сегмента белка ВВР 1,который предположительно топологически сходен с сегментом ВАР в пределах предшественника АРР дикого типа. Пример 4. Тканевое распределение экспрессии ВВР 1 человека. Экспрессия мРНК ВВР 1 была исследована в качестве исходного этапа выявления активности данного гена и кодируемого им продукта. Основной транскрипт длиной 1250 нуклеотидов был найден во всех тканях (фиг. 5 А). Отмечен высокий уровень экспрессии в сердце. Цельный головной мозг характеризовался средним уровнем экспрессии. Все образцы отдельных отделов головного мозга проявляли экспрессию ВВР 1 (фиг. 5 В). Интересно, что все лимбические участки характеризовались относительно большим количеством мРНК ВВР 1. Имелись участки головного мозга, в которых исходно имеются агрегации ВАР и связанное с ними проявление нейротоксичности. Анализ авторентгенограмм гибридизации in situ, полученные с использованием специфичного рибозонда ВВР 1, показал, что в гиппокампе человека энторинальном отделе полушарий головного мозга мРНК ВВР 1 экспрессирована в клетках от среднего до крупного размера по типу, согласующемуся с экспрессией в нейронах в противоположность глиальным клеткам (фиг. 6). Более того,мРНК ВВР 1 виртуально экспрессирована во всех гиппокампных и энторинальных нейронах,поскольку не удалось выявить каких-либо реальных или ламинарных различий в интенсив 33 ности гибридизационного сигнала. Интересно,что параметры экспрессии ВВР 1 оказались в значительной степени сходными с параметрами,выявленными с использованием рибозонда, направленного на мРНК, кодирующую предшественник амилоидного белка - АРР (фиг. 6). В целом, мРНК ВВР 1 была выявлена во всех тканях и всех тестированных сегментах головного мозга. Анализ in situ экспрессии мРНК ВВР 1 также показал наличие высокого уровня экспрессии в сегменте гиппокампа. Пример 5. Распределение экспрессии ВВР 1 в линиях клеток. Экспрессия ВВР 1 также была исследована в ряде клеточных линий, а также были взяты данные из dbEST - коллекции экспрессирующихся маркеров, имеющейся в Национальном Центре биотехнологической информации. Методы полимеразной цепной реакции с ревертированием (ОТ-ПЦР) были применены для количественного анализа экспрессии мРНК ВВР 1 в клеточных линиях, обычно используемых для экспрессии рекомбинантных белков, а также в ряде линий опухолевых клеток. Белок ВВР 1 был обнаружен в клетках яичника китайского хомяка и клетках НЕК 293 человека. Сигналы были обнаружена в линии эмбриональных стволовых клеток Ntera-2 и нейробластомных линияхIMRS2 и SK-N-SH. Экспрессия ВВР 1 была выявлена в линиях опухолевых клеток, происходящих из следующих тканей: толстая кишка(Сх-1, Colo205, MIP101, SW948, СаСо, SW620,LS174T), яичники (A2780S, A2780DDP), молочная железа (MCF-7, SKBr-3, T47-D, В 7474), легкие (Lx-1, A5439), меланома (Lox, Skmel30),лейкоз (HL60, СЕМ), простата (LNCAP, Du145,PC-3). Во всех образцах была обнаружена экспрессия ВВР 1 при использовании Нозернблоттинга с мРНК, выделенными из следующих линий опухолевых клеток: промиелоцитозный лейкоз (HL-60), карцинома (HeLa S3), хронический миелогенный лейкоз (К-562), лимфобластоидный лейкоз (MOLT-4), лимфома Беркитта(G361). Пример 6. Избирательное взаимодействие ВВР 1 с ВАР человека в противопоставлении ВАР грызуна. В составе ВАР грызуна имеются три аминокислотные замены (G5R, F10Y и R13H) по сравнению с последовательностью человека. Пептид грызуна проявляет сниженную нейротоксичность и отсутствие связывания с гомогенатами головного мозга человека (Maggio et al,1992). Таким образом представлялось интересным оценить связывание ВАР грызуна с белком ВВР 1 в тест-системе Y2H. Последовательность ВАР человека в составе рЕК 162 была изменена таким образом, чтобы кодировать пептид грызуна, с применением олигонуклеотиднаправленного мутагенеза с помощью ПЦР, описанном 34 выше. Полученная в результате плазмида рЕК 240 была идентична плазмиде, экспрессирующей слитый белок ВАР человека, использованной в настоящем изобретении, за исключением трех кодонов, продуцирующих три аминокислотные замены, соответствующие аминокислотной последовательности грызуна. Взаимодействия между слитым белком BBFI и слитыми белками ВАР человека и грызуна сопоставляли с помощью биотеста Y2H. Штаммы, экспрессирующие ВВР 1 и ВАР грызуна, не проявляли способности ответу по интенсивности роста (фиг. 7). Эти данные подтверждают предположение о том, что ВВР 1 может являться специфическим медиатором нейротоксического действия ВАР, а также указывает на механизм,объясняющий сниженную нейротоксичность ВАР грызуна. Важно также, что полученные данные демонстрируют высокую степень специфичности взаимодействия ВВР 1/ВАР в тестахY2H при том, что замены трех аминокислот были достаточны для полного нарушения такой связи (фиг. 7). Ясно, что настоящее изобретение может быть применено на практике другими путями в сравнении с тем конкретными вариантами, которые имеются в представленных выше описаниях и примерах. Различные модификации и изменения настоящего изобретения являются возможными в свете представленных описаний и, соответственно, они попадают в масштаб прилагаемой формулы изобретения. БиблиографияSEQ ID NO:1, приведенной в Списке последовательностей, кодирующий фрагмент белка, связывающегося с -амилоидным пептидом (ВВР),включающим аминокислоты 68-269 последовательности SEQ ID NO:2, приведенной в данном Списке, а также вариант указанного фрагмента кДНК, полученный на основании вырожденности генетического кода. 2. Фрагмент молекулы кДНК по п.1, представляющий собой вставку клона рЕК 196 АТСС 98399. 3. Молекула кДНК, имеющая последовательность нуклеотидов SEQ ID NO:1, приведенную в Списке последовательностей, кодирующая белок, связывающийся с -амилоидным пептидом (ВВР), имеющим аминокислотную последовательность SEQ ID NO: 2, приведенную в данном Списке, а также вариант указанной молекулы кДНК, полученный на основании вырожденности генетического кода. 4. Молекула кДНК по п.3, представляющая собой вставку клона BBP1-fl АТСС 98617. 5. Полинуклеотид, который гибридизуется в жестких условиях с молекулой кДНК по любому из пп.1-4 или с комплементарной ей молекулой, при том, что указанный полинуклеотид имеет длину, составляющую по крайней мере 75% длины указанной молекулы кДНК, и кодирует -амилоидный пептид. 6. Зонд или праймер, включающий нуклеотидную последовательность, способную гибридизоваться с нуклеотидами 172-194 молекулы кДНК по п.3. 7. Полинуклеотид, включающий по крайней мере одну последовательность, регулирующую экспрессию, функционально связанную по крайней мере с одним полинуклеотидом, выбранным из группы, состоящей из молекулы кДНК по любому из пп.1-4. 8. Штамм клетки-хозяина, трансформированный полинуклеотидом по п.7, при том, что указанная клетка представляет собой прокариотическую клетку. 9. Штамм клетки-хозяина, трансформированный полинуклеотидом по п.7, при том, что указанная клетка представляет собой эукариотическую клетку. 10. Фрагмент белка, связывающегося с амилоидным пептидом (ВВР), включающийID NO:2, приведенной в Списке последовательностей, кодируемый фрагментом молекулы кДНК, включающим нуклеотиды 202-807 последовательности SEQ ID NO:1, приведенной в данном Списке, или кодируемый вариантом указанного фрагмента кДНК, полученного на основании вырожденности генетического кода. 11. Фрагмент белка, связывающегося с амилоидным пептидом (ВВР), кодируемый фрагментом молекулы кДНК по п.1, представляющим собой вставку клона рЕК 196 АТСС 98399. 12. Белок,связывающийся с амилоидным пептидом (ВВР), имеющий аминокислотную последовательность SEQ ID NO: 2,приведенную в Списке последовательностей,кодируемый молекулой кДНК, имеющей последовательность нуклеотидов SEQ ID NO:1, приведенную в данном Списке, или кодируемый вариантом указанной молекулы кДНК, полученным на основании вырожденности генетического кода. 13. Белок ВВР, включающий аминокислотную последовательность, кодируемую молекулой кДНК по п.3, представляющей собой вставку клона BBP1-fl АТСС 98617. 14. Способ получения белка по любому из пп.12-13 или фрагмента данного белка по любому из пп.10-11, заключающийся в том, что выращивают штамм клетки-хозяина по любому из пп.8-9 в подходящей культуральной среде и очищают продуцируемый указанным штаммом белок или его фрагмент. 15. Белок, полученный в соответствии со способом по п.14. 16. Слитый белок ВВР, включающий белок по любому из пп.12-13, 15 или фрагмент данного белка по любому из пп.10-11, связанный с гетерологичной белковой или пептидной последовательностью. 17. Олигонуклеотид, представляющий собой ингибитор экспрессии гена ВВР 1, включающий нуклеотидную последовательность,состоящую из 15-35 нуклеотидов, которая гибридизуется в жестких условиях с молекулой кДНК по любому из пп.1-4. 18. Способ определения количества полинуклеотида, кодирующего белок, связывающегося с -амилоидным пептидом (ВВР), в образце, включающий следующие стадии:(a) гибридизацию зонда по п.6 с образцом в жестких условиях и(b) определение количества указанного зонда, связавшегося с полинуклеотидами в образце. 19. Антитело, которое специфически связывается с внеклеточной областью белка по любому из пп.12-13, 15-16 или фрагмента данного белка по любому из пп.10-11. 20. Способ определения в образце количества белка по любому из пп.12-13, 15-16 или фрагмента данного белка по любому из пп.1011, включающий следующие стадии:a) инкубацию антитела по п.19 с образцом и контролем, при том, что указанный контроль включает различные количества известного белка ВВР или фрагмента белка ВВР и где указанное антитело связано с репортерной молекулой;b) определение количества указанного антитела, связанного с указанным белком ВВР или фрагментом белка в образце иc) количественное определение указанного белка ВВР или фрагмента белка ВВР путем сравнения количества антитела, определенного в b), с указанными контролями. 21. Способ лечения пациента, нуждающегося в ингибировании накопления -амилоидного пептида в головном мозге, включающий введение пациенту терапевтически эффективного количества белка по любому из пп.12-13, 1516 или фрагмента данного белка по любому из пп.10-11.
МПК / Метки
МПК: A61K 48/00, A61P 25/28, G01N 33/53, C12Q 1/68, C07K 14/47, C12N 15/12
Метки: использования, белок, beta-амилоидным, полинуклеотид, связывающийся, пептидом, кодирующий, способы
Код ссылки
<a href="https://eas.patents.su/22-4256-belok-svyazyvayushhijjsya-s-beta-amiloidnym-peptidom-kodiruyushhijj-ego-polinukleotid-i-sposoby-ih-ispolzovaniya.html" rel="bookmark" title="База патентов Евразийского Союза">Белок, связывающийся с &beta-амилоидным пептидом, кодирующий его полинуклеотид и способы их использования</a>
Усеченный по амино-концу моноцитарный хемотаксический белок (mcp-2) и способы его использования

Номер патента: 3942
Опубликовано: 30.10.2003
Авторы: Прост Поль, Ван Дамме Йо, Стрюйф Софи
МПК: A61P 43/00, A61K 38/19, C07K 14/52...
Метки: хемотаксический, усеченный, использования, амино-концу, способы, белок, mcp-2, моноцитарный
Формула / Реферат:
1. Усеченный по амино-концу моноцитарный хемотаксический белок (MCP-2), у которого отсутствуют NH2-концевые аминокислоты, соответствующие аминокислотным остаткам 1-5 природного MCP-2, имеющий аминокислотную последовательность SEQ ID No:3 или SEQ ID No:4, который обладает антагонистической активностью в отношении хемокинов MCP-1, MCP-2, MCP-3 и RANTES. 2. Усеченный по амино-концу MCP-2 по п.1 в гликозилированной форме. 3. Молекула ДНК, кодирующая...
Гибридный белок, образующий гетеродимеры, способ его получения и использования

Номер патента: 2474
Опубликовано: 27.06.2002
Авторы: Джеймсон Бредфорд А., Кэмпбелл Роберт К., Чаппел Скотт К.
МПК: C07K 14/59, A61P 5/00, A61K 38/24...
Метки: гетеродимеры, использования, белок, образующий, способ, гибридный, получения
Формула / Реферат:
1. Гибридный белок, содержащий две коэкспрессируемые аминокислотные последовательности, образующие гетеродимер, каждая из которых представляет собой: а) по крайней мере, одну аминокислотную последовательность, выбранную из группы, состоящей из цепи гомомерного рецептора или его фрагмента, цепи гетеродимерного рецептора или его фрагмента, лиганда, другого, чем гонадотропный гормон или его фрагмент, легкой или тяжелой цепи антитела, легкой цепи...
Способ получения дроспиренона (6&beta, 7β 15&beta, 16&beta – диметилен -3 – оксо -17α-прегн- 4 -ен-21,17-карболактона, drsp) и 6&beta,7β15&beta,16&beta – диметилен -5&beta-гидрокси -3- оксо – 17&alpha – андростан – 21,17 – карболактон (zk 90965) в качестве промежуточного продукта

Номер патента: 1947
Опубликовано: 22.10.2001
Авторы: Никкиш Клаус, Мор Йорг-Торштен
МПК: C07J 53/00
Метки: получения, карболактон, 6&beta, андростан, 17&alpha, диметилен, оксо, 90965, drsp, 21,17, продукта, 15&beta, 5&beta-гидрокси, дроспиренона, способ, 17α-прегн, 16&beta, промежуточного, ен-21,17-карболактона, 7&beta, 6&beta,7β15&beta,16&beta, качестве
Формула / Реферат:
1. Способ получения дроспиренона (6b,7b;15b,16b-диметилен-3-оксо-17a-прегн-4-ен-21,17-карболактон DRSP) осуществляемый каталитическим гидрированием 17a-(3-гидрокси-1-пропинил)-6b,7b;15b,16b-диметилен-5b-андростан-3b,5,17b-триола (ZK 34506) с получением 17a-(3-гидрокси-1-пропил)-6b,7b;15b,16b-диметилен-5b-андростан-3b,5,17b-триола (ZK 92836) окислением в присутствии соли рутения до...
Усеченный по амино-концу хемокин rantes и способы его использования

Номер патента: 3940
Опубликовано: 30.10.2003
Авторы: Стрюйф Софи, Прост Поль, Вам Дамме Йо
МПК: A61P 43/00, C07K 14/52, A61K 38/19...
Метки: способы, усеченный, амино-концу, хемокин, использования, rantes
Формула / Реферат:
1. Усеченный по амино-концу хемокин RANTES, у которого отсутствуют NH2-концевые аминокислоты, соответствующие аминокислотным остаткам 1-2 природного RANTES, имеющий аминокислотную последовательность SEQ ID No:2, который обладает антагонистической активностью в отношении хемокинов MCP-3, MCP-1a , MCP-1b и RANTES. 2. Усеченный по амино-концу RANTES по п.1 в гликозилированной форме. 3. Молекула ДНК, кодирующая усеченный по амино-концу...
Ген, кодирующий новый маркер рака

Номер патента: 3735
Опубликовано: 28.08.2003
Автор: Радосевич Джеймз А.
МПК: A61K 35/12, C07K 5/00, C12N 15/52...
Метки: новый, рака, ген, кодирующий, маркер
Формула / Реферат:
1. Применение молекулы ДНК с нуклеотидной последовательностью, представленной на фиг. 1, либо с последовательностью, имеющей, как минимум, приблизительно 70% гомологию с упомянутой нуклеотидной последовательностью, представленной на фиг. 1, для диагностирования рака. 2. Молекула кДНК с нуклеотидной последовательностью, представленной на фиг. 1. 3. Молекула кДНК с нуклеотидной последовательностью, имеющей приблизительно 70% гомологию с упомянутой...
Случайный патент: Способ лечения остеохондроза позвоночника