Способ лечения ревматоидного артрита
Номер патента: 5734
Опубликовано: 30.06.2005
Авторы: Браунинг Джеффри Л., Хохман Паула С., Бенджамин Кристофер Д.




















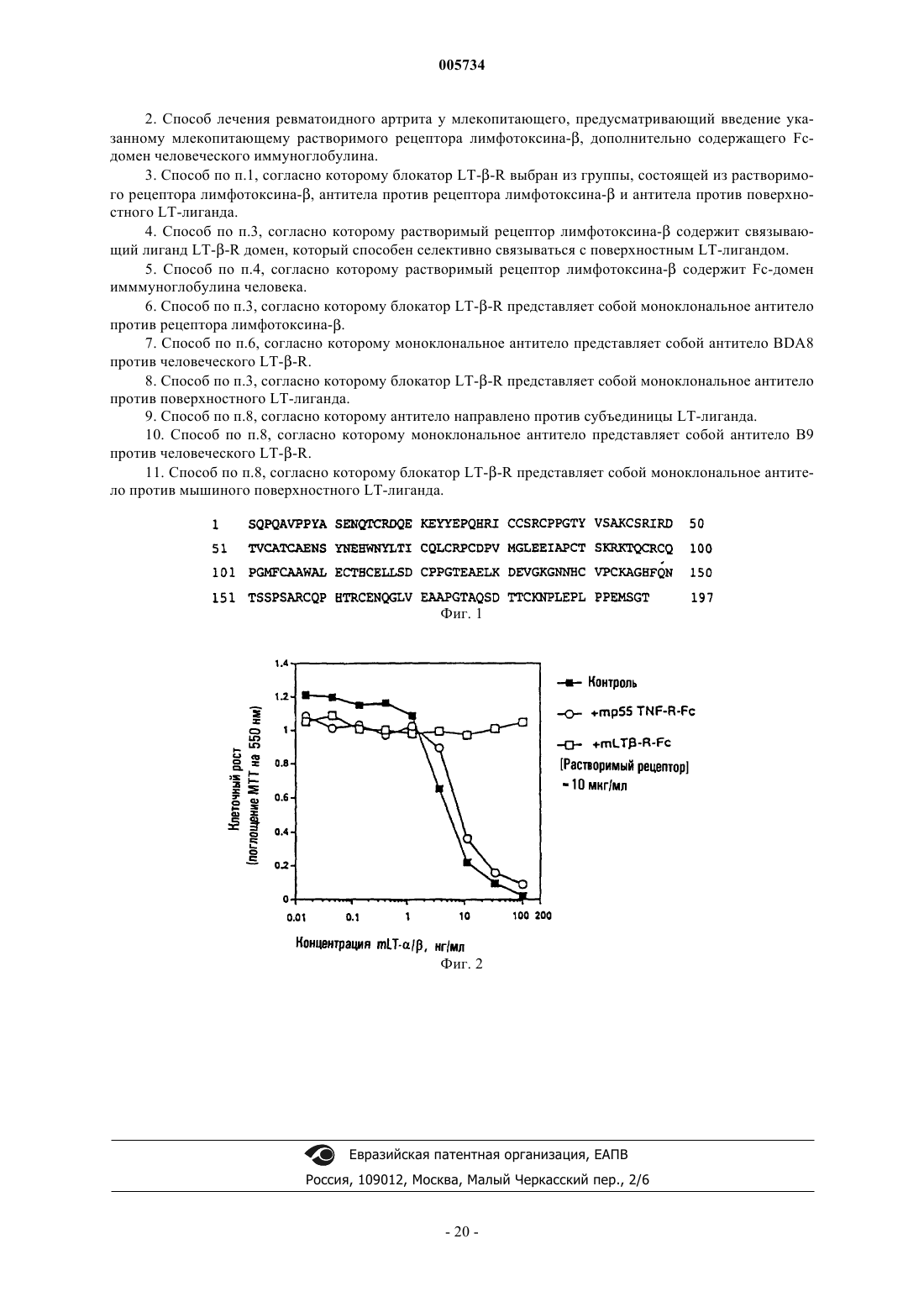
Формула / Реферат
1. Способ лечения ревматоидного артрита у млекопитающего, предусматривающий введение указанному млекопитающему фармацевтической композиции, содержащей терапевтически эффективное количество блокатора рецептора лимфотоксина-b (LT-b-R) и фармацевтически приемлемый носитель.
2. Способ лечения ревматоидного артрита у млекопитающего, предусматривающий введение указанному млекопитающему растворимого рецептора лимфотоксина-b, дополнительно содержащего Fc-домен человеческого иммуноглобулина.
3. Способ по п.1, согласно которому блокатор LT-b-R выбран из группы, состоящей из растворимого рецептора лимфотоксина-b, антитела против рецептора лимфотоксина-b и антитела против поверхностного LT-лиганда.
4. Способ по п.3, согласно которому растворимый рецептор лимфотоксина-b содержит связывающий лиганд LT-b-R домен, который способен селективно связываться с поверхностным LT-лигандом.
5. Способ по п.4, согласно которому растворимый рецептор лимфотоксина-b содержит Fc-домен имммуноглобулина человека.
6. Способ по п.3, согласно которому блокатор LT-b-R представляет собой моноклональное антитело против рецептора лимфотоксина-b.
7. Способ по п.6, согласно которому моноклональное антитело представляет собой антитело BDA8 против человеческого LT-b-R.
8. Способ по п.3, согласно которому блокатор LT-b-R представляет собой моноклональное антитело против поверхностного LT-лиганда.
9. Способ по п.8, согласно которому антитело направлено против субъединицы LT-лиганда.
10. Способ по п.8, согласно которому моноклональное антитело представляет собой антитело В9 против человеческого LT-b-R.
11. Способ по п.8, согласно которому блокатор LT-b-R представляет собой моноклональное антитело против мышиного поверхностного LT-лиганда.
Текст
005734 Область техники Данное изобретение относится к способу лечения ревматоидного артрита у млекопитающего путем введения эффективного количества блокатора рецептора лимфотоксина- (LTR). Блокаторы рецепторов к лимфотоксину- применимы для лечения лимфоцит-опосредованных иммунологических заболеваний и, более конкретно, для ингибирования иммунных ответов, опосредованных Th1-клетками. Данное изобретение также относится к способу лечения ревматоидного артрита у млекопитающего путем введения растворимого рецептора лимфотоксина-, содержащего Fc-домен Ig человека. Предпосылки изобретения Профиль цитокинов, высвобождаемых при иммунной стимуляции, может повлиять на последующий выбор активируемого иммунного эффекторного пути. Выбор между иммунными эффекторными механизмами опосредован CD4-положительными хелперными Т-лимфоцитами (Т-хелперными клетками,или Th-клетками). Th-клетки взаимодействуют с антиген-презентирующими клетками (АПК), которые презентируют на своей поверхности пептидные фрагменты процессированного чужеродного антигена в комплексе с молекулами ГКГ класса II. Th-клетки активируются, когда распознают определенные эпитопы чужеродного антигена, презентированные на поверхности подходящей АПК, на которые Th-клетки экспрессируют специфичный рецептор. Активированные Th-клетки, в свою очередь, секретируют цитокины (лимфокины), которые запускают подходящие иммунные эффекторные механизмы.Th-клетки способны запускать различные эффекторные механизмы, включая активацию киллерных Т-клеток, активацию продукции В-клетками антител и активацию макрофагов. Выбор между иммунными эффекторными механизмами зависит в значительной степени от того, какие цитокины продуцируют активированные Th-клетки.Th-клетки могут быть подразделены на три подгруппы на основании профилей секретируемых ими цитокинов (Fitch et al., Ann. Rev. Immunol., 11, стр. 29-48 (1993. Данные подгруппы называют ThO, Th1 и Th2. У мышей нестимулированные "наивные" Т-хелперные клетки продуцируют IL-2. Кратковременная стимуляция приводит к преобразованию в клетки-предшественники ThO, которые продуцируют широкий спектр цитокинов, включая IFN-, IL-2, IL-4, IL-5 и IL-10. Длительно стимулируемые ThO-клетки способны дифференцироваться либо в клеточный тип Th1, либо в клеточный тип Th2, после чего изменяется профиль экспрессируемых цитокинов. Некоторые цитокины секретируются как Th1-, так и Th2-клетками (например, IL-3, GM-CSF и TNF). Другие цитокины продуцируются исключительно одной либо другой подгруппой Th-клеток. Специализированные эффекты подгрупп Т-хелперных клеток впервые были выявлены у мышей. Сходное разделение применимо и к Т-хелперным клеткам человека (Romagriani et al., Ann. Rev. Immunol., 12, стр. 227-57 (1994.Th-клетки продуцируют LT-, IL-2 и IFN-. У человека профиль секретируемых цитокинов Th1 обычно связывали с клеточным иммунитетом и резистентностью к инфекции. Цитокины Th1 имеют тенденцию к активации макрофагов и запуску определенных видов воспалительного ответа, таких как гиперчувствителъность IV типа (замедленного типа) (смотри ниже). Цитокины Th1 играют важную роль в клеточном отторжении трансплантатов тканей и органов. Тh2-клетки продуцируют цитокины IL-4, IL-5, IL-6 и IL-10. Цитокины Th2 увеличивают продукцию эозинофилов и тучных клеток и стимулируют пролиферацию и созревание В-клеток (Howard et al., "Тcell-derived cytokines and their receptors". Fundamental Immunology, 3d ed., Raven Press, New York (1993. Цитокины Th2 также увеличивают продукцию антител, включая антитела IgE, связанные с аллергическими реакциями, и антитела к трансплантату. Th2-клетки могут также участвовать в супрессии иммунитета и толерантности к персистирующим антигенам.Th1- и Th2-ассоциированные цитокины играют роль в развитии определенных видов реакций по типу гиперчувствительности - неадекватного или диспропорционального иммунного ответа, развивающегося при контакте с ранее встречавшимся антигеном. Выделяют четыре типа гиперчувствительности(Roitt et al., Immunology, стр. 19.1-22.12 (Mosby-Year Book Europe Ltd., 3d ed. 1993. Гиперчувствительностъ I типа (немедленного типа) включает в себя аллерген-индуцированные активацию Th2-клеток и секрецию Тh2-клетками цитокинов. Цитокин IL-4, продуцируемый Th2, стимулирует претерпевание В-клетками переключения изотипа на продукцию IgE, который активирует тучные клетки с развитием острых воспалительных реакций, как, например, ведущие к развитию экземы, бронхиальной астмы и ринита. Гиперчувствительность II и III типов вызывается антителами IgG и IgM к клеточной поверхности или к специфическим тканевым антигенам (II тип) или к растворимым сывороточным антигенам (III тип). Считается, что данные типы реакций гиперчувствительности не опосредованы Th-клетками. Гиперчувствительность IV типа (замедленного типа) (ГЗТ) является опосредованной Th1-клетками. Развитие реакций ГЗТ занимает более 12 ч, и о них говорят как о "клеточно-опосредованных", поскольку они могут быть перенесены от одной мыши к другой лишь путем переноса Th1-клеток, а не только сыворотки. Реакции ГЗТ IV типа обычно подразделяют на три типа: контактную, туберкулинового типа и гранулематозную гиперчувствительность.-1 005734 Многие клеточно-опосредованные реакции, которые способны вызывать заболевание, можно индуцировать у здоровых мышей путем переноса лимфоцитов от больной мыши (например, инсулинзависимый сахарный диабет и экспериментальный аутоиммунный энцефалит). Данное свойство отличает ГЗТ IV типа от остальных трех типов гиперчувствительности, которые представляют собой гуморальные иммунные реакции, вызываемые преимущественно антителами, которые могут быть перенесены в бесклеточной сыворотке. Т-хелперные клетки также участвуют в регуляции переключения изотипа иммуноглобулина denovo. Различные подгруппы Th могут влиять на соотношение иммуноглобулинов определенного изотипа, продуцируемых в ответ на иммунную стимуляцию. Например, цитокин IL-4, продуцируемый Th2,может переключать активированные В-клетки на изотип IgG1 и подавлять другие изотипы. Как обсуждалось выше, IL-4 также активирует избыточную продукцию IgE в реакциях гиперчувствительности I типа. Цитокин IL-5, продуцируемый Th2, индуцирует изотип Ig-A. Данное влияние цитокинов, продуцируемых Th2, на переключение изотипа уравновешено влиянием IFN-, продуцируемого Th1-клетками. Создается впечатление, что отличные профили цитокинов, секретируемых Th1- и Тh2-клетками,направляют реакцию по различным иммунным эффекторным механизмам. Переключение, которое активирует либо клеточно-опосредованный, либо гуморальный эффекторный механизм, является чувствительным к процессам перекрестной супрессии между Th1- и Тh2-клетками:IFN-, продуцируемый Th1-клетками, подавляет пролиферацию Th2-клеток, a IL-10, секретируемый Тh2-клетками, снижает секрецию цитокинов Тh1-клетками. В зависимости от относительных аффинностей цитокинов к их молекулярным мишеням круги отрицательной регуляции Тh1 и Th2 могут умножать эффекты малых различий в концентрациях цитокинов, продуцируемых Тh1- и Тh2-клетками. Возможность контролировать данное переключение путем изменения относительных концентраций цитокинов, продуцируемых Тh1- и Th2-клетками, была бы полезна для лечения дисбаланса при многих иммунных реакциях, зависимых от Тh1- и Тh2-клеток, которые способны приводить к развитию иммунных нарушений и заболеваний. Патологические реакции, опосредованные Тh1, ассоциированы с рядом органоспецифических и системных аутоиммунных состояний, хронических воспалительных заболеваний и реакций гиперчувствительности замедленного типа. Как обсуждалось выше, реакции, опосредованные Тh1, также способствуют развитию клеточных реакций, приводящих к отторжению трансплантированных тканей и органов. До сих пор лечение данных различных иммунных состояний, опосредованных Тh1-клетками, заключается, главным образом, в применении иммуномодуляторов и иммуносупрессоров, равно как и ряда лекарственных средств с плохо изученными механизмами действия (например, золото или пеницилламин). Тремя основными применяемыми в настоящее время иммуносупрессорами являются стероиды,циклоспорин и азатиоприн. Стероиды представляют собой плейотропные противовоспалительные средства, которые подавляют активированные макрофаги и ингибируют активность антиген-презентирующих клеток так, что обращают многие эффекты цитокина IFN-, продуцируемого Th1. Циклоспорин - мощный иммуносупрессор подавляет продукцию цитокинов и снижает экспрессию рецепторов к IL-2 на лимфоцитах при их активации. Азатиоприн представляет собой противовоспалительное средство, которое ингибирует синтез ДНК. Обычно требуется введение высоких доз данных неспецифических иммуносупрессоров, что повышает их токсичность (например, нефро- и гепатотоксичность) и приводит к развитию нежелательных побочных эффектов. Таким образом, они являются неприменимыми в проведении длительных курсов лечения. В порядке решения трудностей, вызываемых традиционными методами лечения с помощью неспецифических иммуносупрессоров, многие современные стратегии лечения направлены на подавление или активацию отдельных функций иммунной системы. Особенно привлекательной задачей является манипулирование балансом между цитокинами, продуцируемыми Тh1 и Th2, в целях сдвигания баланса между клеточно-опосредованными и гуморальными эффекторными механизмами. Для достижения сдвига между клеточно-опосредованными и гуморальными эффекторными механизмами полезной была бы возможность модулирования активности молекулы, которая способна изменять относительные активности подклассов Тh1- и Th2-клеток. Кандидаты на роль таких молекул включают цитокины и рецепторы к ним. Последние данные позволяют предположить, что LT-, IL-12, IFNи IFN- способствуют развитию Th1-опосредованных реакций, тогда как IL-1 и IL-4 направляют ответ по Тh2-опосредованному эффекторному механизму (Romagnani et al., Ann. Rev. Immunol., 12, стр. 227-57 (1994. Многие из цитокинов Th-клеток представляют собой плейотропные регуляторы развития и функционирования иммунной системы, и ингибирование их продукции окажет вредное действие на неопосредованные Т-клетками реакции. Желательная и эффективная мишень для избирательного модулирования выбора между Тh1- и Th2-опосредованными эффекторными механизмами не определена. Сущность изобретения Настоящее изобретение относится к способу лечения ревматоидного артрита у млекопитающего,предусматривающему введение указанному млекопитающему фармацевтической композиции, содержащей эффективное количество блокатора рецептора лимфотоксина- (LTR) и фармацевтически приемлемый носитель.-2 005734 Настоящее изобретение также относится к способу лечения ревматоидного артрита у млекопитающего, предусматривающему введение указанному млекопитающему эффективного количества растворимого LTR, слитого с Fc-доменом человеческого Ig. Краткое описание фигур Фиг. 1. Последовательность внеклеточной части рецептора к LT- человека, которая кодирует лигандсвязывающий домен. Фиг. 2. Растворимый мышиный рецептор к LT-, присоединенный к Fc-домену IgG1 человека(mLTR-Fc), блокирует передачу сигнала через LTR в клетках мышей WEHI 164, индуцированного растворимым мышиным LT-/-лигандом. Клетки WEHI 164 гибнут как функция повышения концентрации LT-лиганда (mLT-/). Растворимый mLTR-Fc (10 мкг/мл) блокирует такую индуцированнуюLT-лигандом клеточную гибель. Гибридный белок растворимого мышиного рецептора к TNF (p55TNFR-Fc) обладает небольшим эффектом на блокирование LT-/-стимулированной клеточной гибели. Рост количественно оценивали по прошествии 3 дней путем измерения оптической плотности (OD 550) прореагировавшей МТТ, которая пропорциональна количеству клеток. Подробное описание изобретения В целях полнейшего понимания описываемого здесь изобретения приведено следующее подробное описание. Термин "цитокин" относится к молекуле, которая опосредует взаимодействия между клетками."Лимфокин" представляет собой цитокин, секретируемый лимфоцитами. Термин "Т-хелперные (Th) клетки" относится к функциональному подклассу Т-клеток, которые способствуют наработке цитотоксических Т-клеток и которые взаимодействуют с В-клетками, стимулируя продукцию антител. Хелперные Т-клетки распознают антиген, ассоциированный с молекулами ГКГII класса. Термин "Th1" относится к подклассу Т-хелперных клеток, которые продуцируют LT-, интерферон- и IL-2 (и другие цитокины) и которые вовлечены в воспалительные реакции, связанные с клеточным, т.е. не с иммуноглобулиновым ответом на иммунную стимуляцию. Термин "Th2" относится к подклассу Т-хелперных клеток, которые продуцируют цитокины, включая IL-4, IL-5, IL-6 и IL-10, которые связаны с иммуноглобулин-опосредованным (гуморальным) ответом на иммунную стимуляцию. Термин "клеточно-опосредованный" относится к тем реакциям в иммунной системе, которые развиваются в результате прямых эффектов Т-клеток и их продуктов с развитием ответа. Данный тип ответа, главным образом (но не исключительно), связан с классом Тh1 Т-клеток. В данную категорию не входят хелперные эффекты Т-клеток на дифференцировку В-клеток и пролиферацию В-клеток, которые,главным образом, связаны с классом Th2 Т-клеток. Термин "гиперчувствительность замедленного типа (ГЗТ)" относится к иммунному ответу, который характеризуется медленным развитием реакции на антиген с полным проявлением эффекта в течение 13-суточного периода. Данное медленное развитие реакции отличается от относительно быстрого развития реакции, наблюдаемого при иммуноглобулин-опосредованной (гуморальной) аллергической реакции. Существует три типа реакций ГЗТ: реакции по типу контактной гиперчувствителъности, гиперчувствительности туберкулинового типа и гранулематозные реакции. Термины "иммуноглобулин-опосредованный ответ" или "гуморальный ответ" относятся к иммунному ответу организма животного на чужеродный антиген, в ходе которого в организме животного продуцируются антитела на чужеродный антиген. Т-клетки класса Th2 принципиально важны для эффективной продукции высокоаффинных антител. Термин "Fc-домен антитела" относится к части молекулы, содержащей шарнир, СН 2- и СН 3 домены, но не содержащей антигенсвязывающие участки. Подразумевается, что термин также включает эквивалентные участки IgM или антитела другого изотипа. Термин "антитело к рецептору к LT-" относится к любому антителу, которое специфично связывается, по крайней мере, с одним эпитопом рецептора к LT-. Термин "антитело к LT" относится к любому антителу, которое специфично связывается, по крайней мере, с одним эпитопом LT-, LT- или комплекса LT-/. Термин "передача сигналов через LTR" относится к молекулярным реакциям, связанным с LT-R-путем, и последующим реакциям, которые развиваются вследствие первых. Термин "блокатор LTR" относится к агенту, который способен ослабить связывание лиганда сLTR, объединение LTR клеточной поверхности в кластеры или передачу сигналов через LTR либо который способен повлиять на интерпретацию сигнала внутри клетки. Блокатор LTR, который действует на стадии лиганд-рецепторного связывания, способен ингибировать связывание LT-лиганда с LTR, по крайней мере, на 20%. Блокатор LTR, который действует после стадии лиганд-рецепторного связывания, способен ингибировать цитотоксические эффекты стимуляции LTR на опухолевую клетку, по крайней мере, на 20%. Примеры блокаторов LTR включа-3 005734 ют растворимые молекулы LTR-Fc и Ab к LT-, LT-, LT-/ или LTR. Предпочтительно, антитела не вступают в перекрестные реакции с секретируемой формой LT-. Термин "биологическая активность LTR" относится к 1) способности молекулы LTR или производного конкурировать за связывание с растворимым или поверхностным LT-лигандом с растворимыми или поверхностными молекулами LTR; или 2) способности стимулировать развитие иммунорегуляторного ответа или цитотоксической активности наподобие нативной молекулы LTR. Термины "гетеромерный комплекс LT-/" и "гетеромерный комплекс LT" относятся к стойкому комплексу, по крайней мере, одной субъединицы LT- и одной или более субъединиц LT-, включая растворимые, мутантные, измененные и химерные формы одной или более субъединиц. Субъединицы могут объединяться посредством электростатических, вандерваальсовых или ковалентных взаимодействий. Предпочтительно, гетеромерный комплекс LT-/ содержит, по крайней мере, две соседние субъединицы LT- и не содержит соседних субъединиц LT-. Когда гетеромерный комплекс LT-/ служит в качестве стимулятора LTR в анализе клеточного роста, комплекс является, предпочтительно, растворимым и описывается стехиометрическим соотношением LT-1/2. Растворимые гетеромерные комплексы LT-/ не содержат трансмембранного домена и могут быть секретированы подходящей клеткойхозяином, которая экспрессирует субъединицы LT- и/или LT- (Crowe et аl., J. Immunol. Methods, 168,стр. 78-89 (1994. Термин "LT-лиганд" относится к гетеромерному комплексу LT или его производному, который специфично связывается с рецептором к LT-. Термин "лигандсвязывающий домен LTR" относится к части или частям LTR, которые вовлечены в процесс специфичного распознавания и взаимодействия с LT-лигандом. Термины "поверхностный комплекс LT-/" и "поверхностный комплекс LT" относятся к комплексу, содержащему субъединицы LT- и мембранно-связанные субъединицы LT-, включая мутантные,измененные и химерные формы одной или более субъединиц, которые экспрессируются на клеточной поверхности. Термин "поверхностный LT-лиганд" относится к поверхностному комплексу LT или его производному, который способен специфично связываться с рецептором к LT-. Термин "субъект" относится к животному или к одной или более клеткам животного происхождения. Предпочтительно, животное представляет собой млекопитающее. Клетки могут находиться в любом виде, включая, но не ограничиваясь, клетки, оставленные в ткани, кластеры клеток, иммортализованные,трансфицированные или трансформированные клетки и клетки, полученные из животного, которое было физически или фенотипически изменено. Лимфотоксин-: Член семейства TNF. Выяснилось, что цитокины, родственные фактору некроза опухоли (TNF), представляют собой большое семейство плейотропных медиаторов иммунной защиты и регуляции иммунной системы. Члены данного семейства существуют в мембранно-связанной форме, которая действует местно посредством межклеточных контактов, или в виде секретируемых белков, которые способны действовать на удаленные мишени. Параллельно семейство TNF-родственных рецепторов взаимодействует с данными цитокинами и запускает разнообразные пути, включая клеточную гибель, клеточную пролиферацию, дифференцировку ткани и провоспалительные реакции.TNF, лимфотоксин- (LT-, также называемый TNF-) и лимфотоксин- (LT-) являются членами семейства TNF лигандов, которые также включают лиганды рецепторов Fas, CD27, CD30, CD40, ОХ-40 и 4-1 ВВ (Smith et al., Cell, 76, стр. 959-62 (1994. Передача через некоторые члены семейства TNF, включая TNF, LT-, LT- и Fas, может индуцировать гибель опухолевой клетки по механизму некроза или апоптоза (запрограммированной клеточной гибели). В неопухолевых клетках TNF и многие лигандрецепторные взаимодействия в пределах семейства TNF влияют на развитие иммунной системы и ответы на различные антигенные стимулы. Большинство мембранно-связанных комплексов LT-/ ("поверхностных LT") отвечают стехиометрическому соотношению LT-1/2 (Browning et al., Cell, 72, стр. 847-56 (1993); Browning et al., J. Immunol., 154, стр. 33-46 (1995. Поверхностные LT-лиганды не связываются с TNF-R с высокой аффинностью и не стимулируют передачу сигналов через TNF-R. Другой TNF-родственный рецептор, называемый рецептором к LT- (LTR), связывает данные поверхностные лимфотоксиновые комплексы с высокой аффинностью (Crowe et al., Science, 264, стр. 707-10 (1994. Передача сигналов через LTR, как и передача сигналов через TNF-R, обладает антипролиферативным эффектом и может являться цитотоксической по отношению к опухолевым клеткам. В совместно поданной заявке на патент Соединенных Штатов, серийный номер 0/378968, авторов настоящей заявки описаны композиции и способы для избирательной стимуляции LTR с применением стимуляторовLTR. Стимуляторы LTR применимы для ингибирования роста опухолевых клеток без коактивации-4 005734 Что касается воздействия на неопухолевые клетки, TNF и TNF-родственные цитокины стимулируют развитие широкого круга иммунных реакций. Как TNF, так и LTлиганды связываются и стимулируют TNF-рецепторы (р 55 или р 60 и р 75 или р 80; называемые здесь TNF-R). TNF и LT- продуцируются макрофагами в ранней и быстрой реакции на бактериальную инфекцию, которая усиливает бактерицидную активность макрофагов и нейтрофилов. TNF и LT-, продуцируемые макрофагами или цитотоксическими Т-лимфоцитами (ЦТЛ или "киллерными Т-клетками"), связываются с рецепторами к TNF на поверхности клеток-мишеней и запускают гибель восприимчивых клеток.TNF и TNF-родственные цитокины способны также запускать воспалительные каскады в ответ на инфекцию или стресс. Выброс TNF, LT- и IFN- изменяет адгезивные свойства клеток сосудистого эндотелия и лимфоцитов определенных типов. Повышенная адгезивность облегчает миграцию фагоцитов и лейкоцитов из кровотока в ткани, окружающие очаг воспаления. Сходные воспалительные реакции играют основную роль в клеточном отторжении трансплантатов тканей и органов и при определенных иммунных нарушениях. Лимфотоксиновые (LT) комплексы клеточной поверхности были охарактеризованы на клеткахCD4+ Т-клеточной гибридомы (11-23.D7), которые экспрессируют высокие уровни LT (Browning et аl., J.Immunol., 147, стр. 1230-37 (1991); Androlewicz et аl., J. Biol. Chem., 267, стр. 2542-47 (1992. Данные по экспрессии и биологической роли LTR, субъединиц LT и поверхностных LT комплексов приведены в обзоре C.F. Ware et al., "The ligands and receptors of the lymphotoxin system", в Pathways for Cytolysis, CurrentTopics Microbiol. Immunol., Springer-Verlag, стр. 175-218 (1995). Экспрессия LT- индуцируется, а LT- секретируется преимущественно активированными Т- и Влимфоцитами и естественными киллерами (NK). Оказывается, что среди подклассов Т-хелперных клетокLT- продуцируется Th1-, но не продуцируется Тh2-клетками. LT- также был определен в меланоцитах. Анти-LT- антитела могут быть также сорбированы на клетки микроглии и Т-клетки в очагах рассеянного склероза. Лимфотоксин- (также называемый р 33) был выявлен на поверхности Т-лимфоцитов, Т-клеточных линий, В-клеточных линий и лимфокин-активированных киллерных (LAK) клеток. LT-3 является субъектом совместно поданных международных заявок авторов настоящей заявки PCT/US91/04588, опубликованной 9 января 1992 года, как WO92/00329; и PCT/US93/11669, опубликованной 23 июня 1994 года,как WO94/13808, которые включены здесь в качестве ссылки. Поверхностные LT комплексы преимущественно экспрессируются активированными Т- и В-лимфоцитами и естественными киллерами (NK), что может быть определено с помощью FACS-анализа или в соответствии с иммуногистологическими методиками с применением анти-LT- антител или растворимых гибридных белков LTR-Fc. Поверхностный LT был также описан в клонах цитотоксических Тлимфоцитов (CTL) человека, на активированных периферических мононуклеарных лимфоцитах (PML),на IL-2-активированных периферических лимфоцитах крови (LAK-клетках), на периферических Влимфоцитах (PBL), активированных митогеном фитолакки или активированных анти-CD40, и в различных лимфоидных опухолях из Т- и В-клеточных рядов. Контакт с аллоантиген-несущими клетками мишенями специфично индуцирует экспрессию поверхностного LT клонами CD8+ и CD4+ CTL. Рецептор к LT-, член TNF-семейства рецепторов, специфически связывает поверхностные LTлиганды. LTR связывает гетеромерные комплексы LT (преимущественно LT-1/2 и LT-2/1), но не связывает TNF или LT- (Crowe et аl., Science, 264, стр. 707-10 (1994. Передача сигнала через LTR может играть роль в развитии периферических лимфоидных органов и в развитии гуморальных иммунных реакций. Исследования экспрессии LTR находятся на ранних стадиях. мРНК, кодирующая LTR, обнаружена в селезенке, тимусе и других важнейших органах человека. Характер экспрессии LTR сходен с таковым, показанным для p55-TNF-R, за исключением того, что LTR отсутствует на Т-клетках периферической крови и на Т-клеточных линиях. Продукция растворимых LT-комплексов. Растворимые гетеромерные комплексы LT-/ содержат субъединицы LT-, которые преобразованы из мембранно-связанной в растворимую форму. Данные комплексы подробно описаны в совместно поданной международной заявке авторов настоящей заявки (PCT/US93/11669, опубликованной 23 июня 1994 года, как WО 94/13808). Растворимые LTпептиды определяются аминокислотной последовательностью лимфотоксина-, где последовательность расщепляется в любой точке между концом трансмембранного фрагмента (т.е. приблизительно у аминокислотного остатка No 44) и первым TNF-гомологичным фрагментом (т.е. у аминокислотного остатка No 88) в соответствии с системой нумерации Browninget al., Cell, 72, стр. 847-56 (1993). Растворимые LTполипептиды могут быть получены путем усечения N-конца LT- с удалением цитоплазматического фрагмента и трансмембранного фрагмента (Crowe et al., Science, 264, стр. 707-10(1994. Альтернативно, трансмембранный домен может быть инактивирован путем делеции или замещения гидрофобных аминокислотных остатков, которые содержат трансмембранный домен в норме,-5 005734 гидрофильными остатками. В другом случае создается существенно гидрофильный профиль гидрофобности, который снижает сродство к липидам и повышает водорастворимость. Делеция трансмембранного домена является предпочтительной по сравнению с замещением гидрофильными аминокислотными остатками, поскольку это позволяет избежать внесения потенциально иммуногенных эпитопов. Исключенный или инактивированный трансмембранный домен может быть замещен или присоединен к лидерной последовательности типа I (например, к лидерной последовательности VCAM-1) таким образом, что последовательность секретируемого белка начинается с остатка, находящегося междуval40 и рrо 88. Растворимые LTполипептиды могут включать в себя любое количество хорошо известных лидерных последовательностей на N-конце. Такая последовательность позволит пептидам экспрессироваться и служить мишенью для каскада секреции в эукариотической системе. Смотри, например,Ernst et al., патент Соединенных Штатов 5082783 (1992). Растворимые гетеромерные комплексы LT-/ могут быть получены путем совместного трансфицирования подходящей клетки-хозяина ДНК, кодирующей LT- и растворимый LT- (Crowe et al., J. Immunol.Methods, 168, стр. 78-89 (1994. Растворимый LT-, ceкретируемый в отсутствие LT-, сильно олигомеризован. Однако, когда он экспрессируется совместно с LT-, образуется гримерная структура массой 70 кДа,которая содержит оба белка. Также можно получить растворимые гетеромерные комплексы LT-1/2 путем трансфицирования клеточной линии, которая в норме экспрессирует только LT- (такая, как линия RPMI 1788, обсуждавшаяся выше), геном, кодирующим растворимый полипептид LT-. Полипептиды LT- и LT- могут быть синтезированы по отдельности, денатурированы с использованием нестойких детергентов, смешаны вместе и ренатурированы путем удаления детергента с образованием смешанных гетеромерных комплексов LT, которые могут быть выделены (смотри ниже). Очистка комплексов LT-1/2. Растворимые гетеромерные комплексы LT-1/2 выделяют из совместно экспрессируемых комплексов с различными стехиометрическими соотношениями субъединиц по методу хроматографии, применяя рецепторы к TNF и LT- в качестве реагентов для аффинной очистки. TNF-рецепторы связываются лишь с /-щелями комплексов LT. Рецептор к LT- с высокой аффинностью связывается с /-щелями и с меньшей аффинностью с /-щелями гетеромерных комплексов LT-/. Соответственно, LT-3 иLT-2/1 будут связываться с TNF-R. LTR также способен связывать тримеры LT-2/1 (с /-щелями), но не способен связывать LT-3. В дополнение к этому, LTR (но не TNF-R) связывает LT-1/2 иLT-n (точный состав такого соединения неизвестен, однако, оно представляет собой крупные агрегаты). Аффинные реагенты на основе рецепторов могут быть получены либо в виде растворимого внеклеточного домена (смотри, например, Loetscher et al., J. Biol. Chem., 266, стр. 18324-29 (1991, либо в виде химерных белков, состоящих из внеклеточного лигандсвязывающего домена, присоединенного к Fcдомену иммуноглобулина (Loetscher et al., J. Biol. Chem., 266, стр. 18324-29 (1991); Crowe et al., Science,264, стр. 707-10 (1994. Рецепторы связывают с аффинными матрицами посредством химического поперечного сшивания, применяя устоявшиеся методики. Существуют две схемы, по которым LT-1/2-лиганд может быть очищен с применением рецепторов и иммуноаффинной хроматографии. В соответствии с первой схемой супернатант из системы с подходящей экспрессией, экспрессирующей как LT-, так и усеченную форму LT-, пропускают через колонку с TNF-R. TNF-R будет связывать LT-3 и тримеры LT-2/1. Элюат, прошедший через колонку сTNF-R, будет содержать LT-(n) и LT-/2. В соответствии со второй схемой все LTсодержащие формы (LT-(n), LT-1/2 и LT-2/1) сажают на колонку с LTR и элюируют с нее, применяя классические методики изменения chaotrophe или рН (LT-3 элюируется сквозь колонку). Элюат нейтрализуют или удаляют chaotrophe, а элюат затем пропускают через колонку с TNF-R, который связывает лишь тримеры LT-2/1. Элюат, прошедший через данную колонку, будет содержать LT-(n) и LT-1/2. В обоих случаях чистые тримеры LT-1/2 могут быть отделены от LT- с помощью последовательного выполнения методик гель-фильтрации и/или ионообменной хроматографии, известных в данной области. Альтернативно, различные формы гетеромерных комплексов LT-/ могут быть разделены и очищены в соответствии с множеством традиционных хроматографических методик. Также предпочтительным может являться объединение серии традиционных схем очистки с одной из стадий иммуноаффинной очистки, описанных выше. Скрининг для поиска блокаторов LTR. В одном осуществлении данного изобретения блокатор LTR содержит антитело (Аb) к LTR,которое ингибирует передачу сигнала через LTR. Предпочтительно, анти-LTR Аb представляет собой моноклональное антитело (mAb). Одним из таких анти-LTR mAb является mAb BDA8. Ингибирующие анти-LTR Аb и другие блокаторы LTR могут быть идентифицированы с применением методов скрининга, которые определяют способность одного или более агентов либо связываться с LTR или с LT-лигандом, либо ингибироватъ эффекты передачи сигнала через LTR на клетки.-6 005734 В одном скрининговом методе применяются цитотоксические эффекты передачи сигнала через LTR на опухолевые клетки, экспрессирующие LTR. Опухолевые клетки подвергают воздействию одного или более стимуляторов LTR с тем, чтобы вызвать передачу сигнала через LTR. Стимуляторы LTR включают гетеромерные комплексы LT-/ (предпочтительно, растворимые LT-1/2) в присутствииIFN- или стимулирующее анти-LTR Аb (смотри ниже; также описанное в совместно поданной заявке на патент Соединенных Штатов серийный номер 08/378968 авторов настоящей заявки). Антитела и другие агенты, которые способны блокировать передачу сигнала через LTR, выбирают, основываясь на их способности ингибировать цитотоксический эффект передачи сигнала через LTR на опухолевые клетки в следующем анализе: 1) oпухолевые клетки, такие как клетки НТ 29, культивируют в течение от 3 до 4 суток в серии лунок для тканевой культуры, содержащих среду и, по крайней мере, один стимулятор LTR в присутствии или в отсутствие серийных разведении тестируемого агента; 2) к опухолевым клеткам добавляют витальный краситель, по которому определяют функцию митохондрий, такой как МТТ, и реакции дают протекать в течение нескольких часов; 3) количественно определяют оптическую плотность смеси в каждой лунке при длине волны 550 нм(ОП 550). Значение ОП 550 пропорционально количеству опухолевых клеток, выживших в присутствии стимулятора LTR и тестируемого блокатора LTR в каждой лунке. Агент или комбинация агентов,которые в данном анализе способны уменьшить LTR-стимулированную цитотоксичность по отношению к опухолевым клеткам, по крайней мере, на 20%, являются блокаторами LTR, входящими в сферу данного изобретения. Любой агент или комбинация агентов, которые стимулируют передачу сигнала через LTR, могут быть применены в описанном выше анализе для идентификации блокаторов LTR. Стимуляторы LTR,которые индуцируют передачу сигнала через LTR (такие, как стимулирующие анти-LTR mAb),могут быть выбраны на основании их способности - одного или в сочетании с другими агентами - потенциироватъ цитотоксичность по отношению к опухолевым клеткам с применением анализа выживаемости опухолевых клеток, описанного выше. Другой способ выбора блокатора LTR представляет собой исследование способности предполагаемого агента непосредственно вмешиваться в процесс LT-лиганд-рецепторного связывания. Агент или комбинация агентов, которые способны блокировать лиганд-рецепторное связывание, по крайней мере,на 20%, являются блокаторами LTR, входящими в сферу данного изобретения. Для проведения анализа конкуренции с предполагаемыми блокаторами LTR может быть применена любая из ряда методик проведения анализа, с помощью которого измеряют прочность лигандрецепторного связывания. Прочность связывания между рецептором и лигандом может быть измерена с помощью твердофазного иммуноферментного анализа (ELISA) или радиоиммуноанализа (RIA). Параметры специфичного связывания также могут быть измерены с помощью комплексов антиген-антитело,меченных флюоресцентной меткой, или с помощью выполнения проведения анализа на клеточном сортере с возбуждением флюоресценции (FACS), или путем постановки других подобных методик иммунологического анализа, каждая из которых представляет собой методику, хорошо известную в данной области. Параметры лиганд-рецепторного связывания также могут быть измерены с помощью прибора BIAcore(Pharmacia Biosensor), в котором используется явление plasmon резонансного определения (Zhou et al.,Biochemistry, 32, стр. 8193-98 (1993); Faegerstram and O'Shannessy "Surfase plasmon resonance detection inaffinity technologies", в Handbook of Affinity Chromatography, стр. 229-52, Marcel Dekker, Inc., New York (1993. Технология BIAcore позволяет посадить рецептор на золотую поверхность и пропускать над ним лиганд. Plasmon резонансное определение позволяет непосредственно количественно определить массу,связавшуюся с поверхностью за определенное время. Данная методика позволяет определить константы скорости как прямой, так и обратной реакции, и, таким образом, константа диссоциации и константа сродства комплекса лиганд-рецептор могут быть непосредственно определены в присутствии и в отсутствие предполагаемого блокатора LTR. С помощью данных или иных методик для измерения параметров связи в комплексе рецепторлиганд можно оценить способность блокатора LTR, одного или в сочетании с другими агентами, ингибировать связывание поверхностных или растворимых LT-лигандов с поверхностными или растворимыми молекулами LTR. Такие анализы также могут быть применены для тестирования блокаторовLTR или производных таких агентов (например, конденсированных, химерных, мутантных или химически модифицированных форм), одних или в сочетании с другими агентами, для оптимизации способности данного модифицированного агента блокировать стимуляцию LTR. Продукция растворимых молекул LTR. В одном осуществлении данного изобретения блокаторы LTR содержат растворимые молекулы рецептора к LT-. Фиг. 1 показывает последовательность внеклеточного фрагмента человеческого LTR, которая кодирует лигандсвязываюший домен. Применяя информацию последовательности, приведенную на фиг. 1,-7 005734 и методики получения рекомбинантной ДНК, хорошо известные в данной области, функциональные фрагменты, кодирующие лигандсвязывающий домен LTR, могут быть клонированы в векторе и экспрессированы подходящим хозяином с получением растворимой молекулы LTR. Растворимые молекулы LTR, которые по данным описанных здесь анализов могут конкурировать с нативными рецепторами к LT- за связывание LT-лиганда, отбирают как блокаторы LTR. Растворимый рецептор к LT-, содержащий аминокислотные последовательности, выбранные из таковых, приведенных на фиг. 1, может быть присоединен к одному или более гетерологичных белковых доменов ("гибридных доменов") в целях увеличения устойчивости рецепторного гибридного белка invivo или в целях модулирования его биологической активности или локализации. Предпочтительно, для создания рецепторных гибридных белков применяют стойкие белки плазмы,которые, как правило, имеют период полужизни в циркуляторном русле более чем 20 ч. Такие белки плазмы включают, но не ограничиваются ими, иммуноглобулины, сывороточный альбумин, липопротеины, аполипопротеины и трансферрин. Последовательности, которые способны нацеливать растворимую молекулуLTR на определенную клетку или тип ткани, также могут быть присоединены к лигандсвязывающему домену LTR в целях создания специфически локализованного растворимого гибридного белка LTR. Весь внеклеточный фрагмент LTR или его функциональная часть (фиг. 1), содержащая лигандсвязывающий домен LTR, могут быть гибридизованы с постоянным доменом тяжелой цепи иммуноглобулина, таким как Fc-домен тяжелой цепи IgG1 человека (Browning et al., J. Immunol., 154, стр. 33-46 (1995. Растворимые гибридные белки рецептор-Ig-G являются предпочтительными и представляют собой распространенные иммунологические реагенты, а способы их создания известны специалистам в данной области (смотри, например, патент Соединенных Штатов 5225538, включенный здесь в качестве ссылки). Функциональный лигандсвязывающий домен LTR может быть гибридизован с Fc-доменом иммуноглобулина (Ig), относящегося к классу или подклассу иммуноглобулинов, отличному от IgG1. Fcдомены или антитела, относящиеся к различным классам или подклассам Ig, способны активировать различные вторичные эффекторные функции. Активация происходит, когда Fc-домен связывается сходнымFc-рецептором. Вторичные эффекторные функции включают способность активировать систему комплемента, проникать через плаценту и связывать различные белки микробного происхождения. Свойства различных классов или подклассов иммуноглобулинов описаны Roitt et al., Immunology, стр. 4.8 (MosbyYear Book Europe Ltd., 3d ed. 1993). Активация системы комплемента запускает каскады ферментативных реакций, которые опосредуют воспаление. Продукты системы комплемента обладают множеством функций, включая связывание бактерий, эндоцитоз, фагоцитоз, цитотоксичность, продукцию свободных радикалов и солюбилизацию иммунных комплексов. Ферментативный каскад системы комплемента может быть активирован Fc-доменами антигенсвязанных антител классов IgG1, IgG3 и IgM. Выяснилось, что Fc-домен IgG2 является менее эффективным,а Fc-домены IgG4, IgA, IgD и IgE являются неэффективными для активации системы комплемента. Таким образом, Fc-домен выбирают на основании того, являются ли связанные с его применением вторичные эффекторные функции желаемыми при конкретной иммунной реакции или заболевании, которые лечат с помощью гибридного белка LTR-Fc. Если предпочтительно будет повредить или убить LT-лиганд-несущие клетки-мишени, для создания гибридного белка LTR-Fc можно выбрать особенно активный Fc-домен (IgG1). Альтернативно,если предпочтительно будет нацелить гибридный белок LTR-Fc на клетку без активации системы комплемента, можно выбрать неактивный Fc-домен IgG4. Были описаны мутации в Fc-домене, которые снижают или прекращают связывание с Fcрецепторами и активацию системы комплемента (S. Morrison, Annu. Rev. Immunol., 10, стр. 239-65(1992. Данные или другие мутации могут быть применены одни или в сочетании для оптимизации активности Fc-домена, применяемого для создания гибридного белка LTR-Fc. Получение растворимого человеческого гибридного белка LTR, содержащего лигандсвязывающие последовательности, конденсированные с Fс-доменом человеческого иммуноглобулина (hLTR-Fc),описано в примере 1. Одну линию СНО, созданную в соответствии с примером 1, которая секретируетhLTR-Fc, называют "hLT; R-hGl CHC14". Образец данной линии был принят на хранение 21 июля 1995 года в American Type Culture Collection (ATCC) (Rockville, MD) в соответствии с положениями Будапештского договора (the Budapest Treaty), и ему был присвоен номер по каталогу ATCC CRL11965. Получение молекулы растворимого мышиного гибридного белка LTR (mLTR-Fc) описано в примере 2. Линию СНО, созданную в соответствии с примером 1, которая секретирует mLTR-Fc, называют "mLT; R-hGl СНО 1.3.ВВ". Образец данной линии был принят на хранение 21 июля 1995 года вAmerican Type Culture Collection (ATCC) (Rockville, MD) в соответствии с положениями Будапештского договора (the Budapest Treaty), и ему был присвоен номер по каталогу ATCC CRL11964. Все ограничения доступности широкому кругу исследователей вышеуказанных хранилищ ATCC будут окончательно сняты по выдаче патента на данную заявку.-8 005734 Различные аминокислотные остатки, образующие точку соединения в гибридном белке рецептор-Ig могут изменять структуру, устойчивость и предельную биологическую активность растворимого LT-рецепторного гибридного белка. На С-конец выбранного фрагмента LTR может быть добавлена одна или более аминокислот в целях модификации точки соединения выбранным доменом слияния.N-конец гибридного белка LTR также может быть модифицирован путем изменения положения,по которому выбранный фрагмент ДНК, кодирующей LTR, расщепляется на 5'-конце для вставки в рекомбинантный экспрессирующий вектор. Устойчивость и активность каждого гибридного белка LTR могут быть протестированы и оптимизированы с помощью устоявшихся методик постановки эксперимента и анализов для выбора описываемых здесь блокаторов LTR. Применяя последовательности лигандсвязывающего домена LTR во внеклеточном домене, показанном на фиг. 1, можно также создать варианты аминокислотных последовательностей с тем, чтобы модифицировать сродство растворимого рецептора к LT- или гибридного белка к LT-лиганду. Растворимые молекулы LTR по данному изобретению способны конкурировать за связывание поверхностного LT-лиганда с эндогенными рецепторами к LT- на клеточной поверхности. Представляется, что любая растворимая молекула, содержащая лигандсвязывающий домен LTR, которая способна конкурировать с рецепторами к LT- на клеточной поверхности за связывание поверхностного LT-лиганда,является блокатором LTR, который входит в сферу настоящего изобретения. Растворимые молекулы LTR как блокаторы LTR. Растворимый гибридный белок, состоящий из рецептора к LT- и иммуноглобулина человека(hLTR-Fc), получали в соответствии с методиками, приведенными в примере 1, и тестировали на предмет его способности блокировать LTR-индуцированную цитотоксичность по отношению к человеческим опухолевым клеткам НТ 29. В таблице (пример 3) приводится сравнение способности растворимых гибридных белков рецептора к LT- (hLTR-Fc) и рецептора к TNF (p55-TNF-R-Fc) блокировать ингибирующие эффекты различных TNF- и растворимых LT-лигандов на рост опухолевых клеток НТ 29. Среди данных, изложенных в таблице, указаны концентрации, при которых растворимый рецептор к LT- (hLTR-Fc) способен предотвращать гибель опухолевых клеток, вызванную взаимодействием между LТ-1/2-лигандом и рецепторами к LT- клеточной поверхности, на 50%. Способность блокировать рост опухолевых клеток, по крайней мере, на 20% свидетельствует о данном растворимом рецепторе к LT- как о блокаторе LTR по данному изобретению. Как и ожидалось, растворимый гибридный белок TNF-R (p55-TNF-R-Fc) полностью блокирует TNF-индуцированное ингибирование роста путем связывания с TNF и предотвращения его взаимодействия с поверхностным рецептором. Растворимый гибридный белок TNF-R не влияет на опосредованные LT-лигандом (LT-1/2) антипролиферативные эффекты. В противоположность этому, гибридный белок LTR блокировал эффектыLT-лиганда, но не блокировал эффекты TNF или LT-. Таким образом, растворимые гибридные белкиLTR человека не вмешиваются в процесс активации TNF-R TNF- и LT-лигандами. Для того чтобы определить, является ли передача сигнала через LTR также цитотоксической для опухолевых клеток у мышей и способны ли растворимые гибридные белки LTR блокировать LTRиндуцированную цитотоксичность, проводили сходный эксперимент, применяя мышиные опухолевые клетки. Растворимый мышиный гибридный белок LTR-Fc (mLTR-Fc; смотри пример 2) тестировали на предмет его способности предотвращать гибель мышиных клеток WEHI 164, обработанных LTлигандом (пример 4). На фиг. 2 показаны эффекты растворимого мышиного LTR (mLTR-Fc) на LT-лиганд-индуцированную передачу сигнала через LTR на мышиных клетках WEHI 164. Как показано данным анализом, клетки WEHI 164 гибнут при обработке растворимым LT-1/2-лигaндoм. Добавление mLTR-Fc блокирует LT-лиганд-стимулированную клеточную гибель. Контрольный гибридный белок TNFрецептора обладает слабым эффектом на блокирование клеточной гибели. Эти данные показывают, что растворимый гибридный белок LTR способен эффективно конкурировать с поверхностными молекулами LTR за связывание LT-лиганда. Таким образом, растворимый гибридный белок LTR-Fc действует у мышей как блокатор LTR. Источник антител к человеческому LTR. В другом осуществлении данного изобретения антитела к человеческому рецептору к LT- (антиLTR Аb) функционируют как блокатор LTR. Анти-LTR Ab по данному изобретению могут являться поликлональными или моноклональными (mAb) и могут быть модифицированы в целях оптимизации их способности блокировать передачу сигнала через LTR, их биологической доступности invivo, устойчивости и других желаемых свойств. Поликлональные антисыворотки к человеческому рецептору к LT- получают, применяя традиционные методики подкожного введения животным, таким как козы, кролики, крысы, хомяки, мыши, гибридного белка человеческого рецептора к LT- и Fc-домена иммуноглобулина (пример 1) в полном адъюванте Фройнда с последующим реиммунизирующим внутрибрюшинным или подкожным введением в-9 005734 неполном адъюванте Фройнда. Поликлональные антисыворотки, содержащие желаемые антитела к рецептору к LT-, тестируют с помощью традиционных иммунологических методик. Гибридомная клеточная линия (BD.A8.AB9), которая продуцирует мышиные mAb BDA8 к человеческому LTR, была принята на хранение 12 января 1995 года в American Type Culture Collection(ATCC) (Rockville, MD) в соответствии с положениями Будапештского договора (the Budapest Treaty), и ей был присвоен номер по каталогу ATCC НВ 11798. Все ограничения доступности широкому кругу исследователей вышеуказанных хранилищ ATCC будут окончательно сняты по выдаче патента на данную заявку. Различные виды анти-LTR антител также могут быть созданы с применением стандартных методик рекомбинантной ДНК (Winter and Milstein, Nature, 349, стр. 293-99 (1991. Например, могут быть сконструированы "химерные" антитела, в которых антигенсвязывающий домен антитела животного сшит с постоянным доменом иммуноглобулина человека (например, Cabilly et al., US 4816567; Morrisonet al., Proc. Natl. Acad. Sci. U.S.A., 81, стр. 6851-55 (1984. Химерные антитела снижают выраженность наблюдаемых иммуногенных реакций, вызываемых антителами животных, если последние применяют в клинических лечебных процедурах на человеке. В дополнение к этому, могут быть синтезированы рекомбинантные "гуманизированные" антитела,которые распознают LTR. Гуманизированные антитела представляют собой химерные молекулы, содержащие, в основном, последовательности человеческого IgG, в которые введены фрагменты, ответственные за специфичное связывание антигена (например, WO 94/04679). Животных иммунизируют желаемым антигеном, соответствующие антитела выделяют и удаляют часть последовательностей вариабельных доменов, ответственную за специфичное связывание антигена. Антигенсвязывающие фрагменты животного происхождения затем клонируют в подходящее положение генов человеческих антител, из которых были удалены антигенсвязывающие фрагменты. Наличие гуманизированных антител сводит к минимуму применение гетерологичных (межвидовых) последовательностей в человеческих антителах, а индукция ими иммунного ответа у обрабатываемого субъекта менее вероятна. Конструирование различных классов рекомбинантных анти-LTR антител может быть также усовершенствовано созданием химерных или гуманизированных антител, содержащих вариабельные домены анти-LTR антител и постоянные домены человеческих антител (СН 1, СН 2, СН 3), выделенных из иммуноглобулинов различных классов. Например, анти-LTR IgM антитела с повышенной валентностью антигенсвязывающего сайта могут быть рекомбинантно получены путем клонирования антигенсвязывающего сайта в векторах, несущих ДНК, кодирующую постоянные домены -цепи человеческого(1993); Traunecker et al., Nature, 339, стр. 68-70 (1989. В дополнение к этому, стандартные методики рекомбинантной ДНК могут применяться для изменения значений сродства связывания рекомбинантных антител с антигенами путем замены аминокислотных остатков поблизости от антигенсвязывающих сайтов. Сродство связывания гуманизированного антитела с антигеном может быть увеличено с помощью мутагенеза, основанного на молекулярном моделировании (Queen et al., Proc. Natl. Acad. Sci. U.S.A., 86, стр. 10029-33 (1989); WO 94/04679). Может оказаться желательным повысить или понизить сродство анти-LTR Аb к LTR в зависимости от типа ткани-мишени или конкретной предполагаемой схемы лечения. Например, может являться предпочтительной обработка пациента постоянными уровнями анти-LTR Аb со сниженной способностью к передаче сигнала через LTпуть в порядке полупрофилактического лечения. Кроме того, ингибирующие анти-LTR Аb с повышенным сродством к LTR могут являться предпочтительными для кратковременных курсов лечения. Анти-LTR антитела как блокаторы LTR. Анти-LTR антитела могут быть отобраны путем тестирования их способности ингибироватъ LT-R-индуцированную цитотоксичностъ, направленную на опухолевые клетки. В предпочтительном осуществлении данного изобретения композиции и способы содержат мышиные mAb к человеческому LTR BDA8. Показано, что mAb BDA8 действует как блокатор LTR, что определено данным изобретением. Опухолевые клетки WiDr прекращают рост в присутствии IFN- и растворимого LT-1/2-лигaндa. Контрольные антитела (IgG1) не влияют на данное ингибирование роста. В противоположность этому, анти-LTR mAb BDA8 блокирует способность растворимого LT1/2-лигaндa ингибировать рост клеток WiDr. Таким образом, антитело к человеческому рецептору кLT- способно действовать как блокатор LTR, что определено настоящим изобретением. Тестируя другие антитела к человеческому рецептору к LT-, можно ожидать, что дополнительные анти-LTR антитела, которые действуют как блокаторы LTR у людей, могут быть идентифицированы с помощью устоявшихся методик постановки эксперимента и анализов, описанных здесь. Источник антител к поверхностному LT-лиганду. Другое предпочтительное осуществление данного изобретения содержит композиции и способы,которые включают в себя антитела к LT-лиганду, которые действуют как блокаторы LTR. Как описано выше для случая анти-LTR Аb, антитела к LT-лиганду, которые действуют как блокаторы LTR,- 10005734 могут являться поликлональными или моноклональными и могут быть модифицированы в соответствии с устоявшимися методиками в целях модулирования их антигенсвязывающих свойств и их иммуногенности. Антитела к LT по данному изобретению могут быть выработаны по отдельности к одной из двух субъединиц LT, включая растворимые, мутантные, измененные и химерные формы субъединицы LT. Если в качестве антигена применяют субъединицы LT, они предпочтительно представляют собой субъединицы LT-. Если применяют субъединицы LT-, предпочтительно, чтобы полученные анти-LT- антитела связывались с поверхностным LT-лигандом, и не вступали в перекрестную реакцию с секретируемымLT-, и не модулировали активность TNF-R (в соответствии с анализами, описанными в примере 3). Альтернативно, антитела к гомомерному (LT-) или гетеромерному (LT-/) комплексу, содержащему одну или более субъединиц LT, могут быть выработаны и протестированы на предмет LTRблокирующей активности. Предпочтительно, комплексы LT-1/2 применяют в качестве антигена. Как обсуждалось выше, предпочтительно, чтобы полученные анти-LT-1/2 антитела связывались с поверхностным LT-лигандом, и не связывались с секретируемым LT-, и не влияли на активность TNF-R. Продукция поликлональных антител к человеческому LT- описана в совместно поданной заявке авторов настоящей заявки (WO 94/13808). Моноклональные анти-LT- и анти-LT- антитела также были описаны (Browning et аl., J. Immunol., 154, стр. 33-46 (1995. Гибридомная клеточная линия (В 9.С 9.1), которая продуцирует мышиные mAb B9 к человеческомуLTR, была принята на хранение 21 июля 1995 года в American Type Culture Collection (ATCC) (Rockville,MD) в соответствии с положениями Будапештского договора (the Budapest Treaty), и ей был присвоен номер по каталогу ATCC HB11962. Гибридомная клеточная линия (BB.F6.1), которая продуцирует mAb хомяка к мышиному LT-/BB.F6, была принята на хранение 21 июля 1995 года в American Type Culture Collection (ATCC) (Rockville,MD) в соответствии с положениями Будапештского договора (the Budapest Treaty), и ей был присвоен номер по каталогу ATCC НВ 11963. Все ограничения доступности широкому кругу исследователей вышеуказанных хранилищ ATCC будут окончательно сняты по выдаче патента на данную заявку. Антитела к LT-лиганду как блокаторы LTR. Проводили анализ на клеточном сортере с возбуждением флюоресценции (FACS) в целях скрининга на предмет антител к субъединицам LT или комплексам LT, которые действуют как блокаторы LTR. В данном анализе растворимый гибридный белок LTR-Fc добавляют к активированным митогеном фитолакки клеткам 11-23, которые экспрессируют поверхностные LT-комплексы (Browning et al., J. Immunol.,154, стр. 33-46 (1995, в присутствии возрастающих концентраций тестируемого антитела. Антитело,которое способно ингибировать взаимодействие рецептора к LT- и его лиганда, по крайней мере, на 20%, отбирают как блокатор LTR. Результаты данного анализа, проведенного в целях тестирования мышиных mAb B9 к человеческому LT-, показывают, что анти-LT- mAb B9 способно избирательно ингибировать связывание растворимых гибридных белков LTR-Fc с поверхностными LT-лигандами, индуцированное на активированных клетках. Данные результаты подтверждают представление о том, что антитела к субъединице LTлиганда будут действовать как блокаторы LTR. Описанный выше FACS-анализ также применяли для тестирования LT, выработанных в организме хомяка к растворимому комплексу мышиных LT-/. Результаты данного анализа, проведенного в целях тестирования mAb хомяка к мышиному LT-/ BB.F6, показывают, что анти-LT-/ mAb BB.F6 способны эффективно ингибировать связывание растворимых гибридных белков LTR-Fc (пример 2) с поверхностными LT-лигандами, экспрессируемыми на клетках мышиной Т-клеточной гибридомы, и, таким образом, являются блокаторами LTR по данному изобретению. Предпочтительное применение комплекса LT-/ по сравнению с субъединицей LT в качестве антигена для иммунизации животного может привести к более эффективной иммунизации или может привести к продукции антител, обладающих большим сродством к поверхностному LT-лиганду. Представляется возможным, что в случае иммунизации комплексом LT-/ могут быть выделены антитела, которые распознают аминокислотные остатки на обеих субъединицах, LT- и LT- (например, остатки, которые образуют щель в комплексе LT-/). Тестируя антитела к человеческому гетеромерному комплексуLT-/, можно ожидать, что дополнительные анти-LT антитела, которые действуют как блокаторы LTR у людей, могут быть идентифицированы с помощью устоявшихся методик постановки эксперимента и анализов, описанных здесь. Блокаторы LTR ингибируют опосредованную Тh1-клетками контактную гиперчувствительность у мышей. Блокаторы LTR по данному изобретению способны ингибировать опосредованные Th1-клетками иммунные реакции. Одной такой опосредованной Th1-клетками реакцией является гиперчувствительность замедленного типа (ГЗТ; Cher and Mosmann, J. Immunol., 138, стр. 3688-94-(1987); смотри также I.Roitt et al., Immunology, стр. 22.1-22.12, Mosby-Year Book Europe Ltd., 3d ed. (1993) для общего обсужде- 11005734 ния). ГЗТ индуцируется, когда сенсибилизированные антигеном Th1-клетки секретируют цитокины после повторного контакта с тем же антигеном. Цитокины, продуцируемые Тh1, являются аттрактантами и стимуляторами макрофагов, которые секретируют дополнительные эффекторные молекулы, которые запускают воспалительные реакции. Реакции, протекающие по типу ГЗТ, подразделяют на три различных типа: контактную гиперчувствительность, гиперчувствительность туберкулинового типа и гранулематозные реакции. Три типа гиперчувствительности (ГЧ) можно различить по скорости и природе ответа на чужеродный антиген, когда его наносят непосредственно на кожу или вводят под кожу сенсибилизированного субъекта. За ходом реакции ГЗТ следят путем определения скорости и степени отека кожи. Реакции ГЧ туберкулинового типа представляют собой кожные реакции, которые развиваются в месте введения чужеродного антигена из микроорганизма, с которым субъект ранее контактировал (например, Mycobacterium tuberculosis или М. leprae). Данную кожную реакцию, которая достигает своего максимума в интервале от 48 до 72 ч после введения антигена, зачастую применяют как базовую для диагностических тестов чувствительности к ранее встречавшимся микроорганизмам (например, туберкулиновый кожный тест). Если развивается повреждение туберкулинового типа, оно может перейти в гранулематозную реакцию при условии, что антиген будет персистировать в ткани. Гранулематозные реакции являются клинически наиболее серьезными реакциями, протекающими по типу ГЗТ, поскольку они способны привести к развитию множества патологических эффектов, связанных с заболеваниями, опосредованными Th1-клетками. Гранулематозные реакции развиваются в случае, если макрофаги оказываются неспособными элиминировать антигены или иммунные комплексы, и последние продолжают стимулировать секрецию цитокинов Th1. Хроническое воспаление и скопление активированных макрофагов в месте нахождения стимулирующего агента характеризуют гранулематозные реакции. Ядро из эпителиальных клеток и макрофагов, которое может быть окружено лимфоцитами и фиброзными отложениями, формирует твердое образование, называемое гранулемой. Иногда в центре гранулемы происходит интенсивная клеточная гибель (например, в пораженной туберкулезом ткани легкого). Формирование твердого образования в ткани-мишени гранулематозной реакции протекает в течение приблизительно 4 недель. Агенты, которые влияют на частоту образования гранулем, могут быть идентифицированы с использованием инфицированных шистосомами мышей (Amiri et al., Nature, 356, стр. 604-607 (1992. Гельминты шистосомы способны вызывать паразитарное заболевание, которое ведет к образованию гранулем вокруг яиц шистосом в воротных венулах инфицированной печени. Агенты, которые способны ингибировать данную опосредованную Th1-клетками реакцию ГЗТ, способны снижать размер гранулем или частоту или скорость образования гранулем в инфицированной шистосомами печени мышей. Клеточная реакция на яйца шистосом может быть оценена с помощью определения изменения количества и размера гранулем, образующихся у мышей, обрабатываемых возрастающими концентрациями предполагаемого блокатора LTR с течением времени. Контактная гиперчувствительность (КГЧ) представляет собой класс ГЗТ, в котором органоммишенью является кожа. При КГЧ воспалительная реакция вызывается местным нанесением реактивного гаптена на кожу. Аллергены, как правило, состоят из, по крайней мере, одной молекулы гаптена, который обычно очень мал, чтобы являться антигеном самим по себе. Гаптен проникает через эпидермис и взаимодействует под кожей с нормальным белком с образованием нового антигенного комплекса. Повторный контакт сенсибилизированного субъекта с гаптеном запускает реакцию ГЗТ. Конъюгат гаптена и белка-носителя в сочетании с антиген-презентирующими клетками активирует эффекторные механизмы, которые запускают секрецию цитокинов (включая IL-2, IL-3, IFN- и GM-CSF). Каскад секретируемых цитокинов стимулирует пролиферацию CD4+ Т-клеток, изменение профилей экспрессии различных молекул адгезии клеточной поверхности и миграцию Т-клеток и макрофагов в очаг воспаления в коже. Активация цитокинового каскада и, как следствие, вазодилятация, клеточная инфильтрация и отек дермы и эпидермиса приводят к образованию припухлости и воспалению ткани-мишени, благодаря которым происходит измеримое утолщение кожи в процессе развития реакций ГЗТ. Степень, до которой конкретный гаптен способен сенсибилизировать индивидуума, зависит от множества факторов. Данные факторы включают параметры прохождения гаптена через кожу и его взаимодействия с белком-носителем хозяина с образованием конъюгата. Один гаптен, который сенсибилизирует почти всех индивидуумов, представляет собой 2,4-динитрофторбензол (ДНФБ). Кожная реакция на гаптен, такой как ДНФБ, по типу КГЧ представляет собой классическую животную модель клеточно-опосредованного иммунитета. Локализация данной реакции КГЧ на ухе сенсибилизированной мыши позволяет просто, точно и воспроизводимо количественно оценивать данный клеточно-опосредованный иммунный ответ in vivo путем измерения толщины уха. Данные об особенностях реакции КГЧ у мышей и гистопатологии ДНФБ-индуцированной воспалительной реакции были опубликованы (Chisholm et al., Eur. J. Immunol., 23, стр. 682-688 (1993.- 12005734 Способность ДНФБ вызывать реакцию контактной гиперчувствительности у большинства индивидуумов может быть применена для определения агентов, которые снижают или устраняют воспалительные реакции, связанные с опосредованными Th1-клетками реакциями ГЗТ. Растворимый мышиный гибридный белок LTR-Fc эффективно ингибирует ДНФБ-индуцированную реакцию контактной гиперчувствительности у мышей. Первоначально мышей сенсибилизировали путем нанесения ДНФБ на нижнюю часть каждой задней лапы в течение 2 дней подряд. Через 5 дней после первичной сенсибилизации на поверхность левого уха наносили субирритантную дозу ДНФБ в растворе-носителе. Раствор-носитель в отдельности наносили на правое ухо в качестве контроля. Затем возрастающие концентрации блокатора LTR mLTR-Fc (пример 2) внутривенно вводили мышам. Инъекции фосфатно-солевого буфера в отдельности или гибридного белка IgG человека (LFA3Fc) служили в качестве отрицательных контролей, а инъекция mАb, специфичных к VLA4, (mAb PS/2),которые ингибируют КГЧ, служила в качестве положительного контроля. Через 24 ч после иммунной стимуляции измеряли толщину каждого уха (ДНФБ-стимулированного и нестимулированного). Об ингибировании реакции отека уха блокатором LTR судили по сравнению обрабатываемых групп с группой отрицательного контроля. Показано, что mLTR-Fc вызывает значительное уменьшение реакции отека уха у мышей, обработанных ДНФБ, по сравнению с ДНФБ-обработанными контрольными животными, не получившими инъекции ингибитора, (PBS и LFA3-Fc). Растворимый LTR способен ингибировать данную реакцию КГЧ так же эффективно, как и ингибирующее mАb, специфичное к VLA4 (mАb PS/2), которое действует путем ингибирования миграции Т-клеток в место действия стимула (Chisholm et al., Eur. J. Immunol., 23,стр. 682-688 (1993. Эти данные показывают, что растворимый гибридный белок LTR, который действует как блокатор LTR in vitro, также способен эффективно ингибировать опосредованную Тh1-клетками иммунную реакцию, будучи введен в организм животного. Блокаторы LTR по данному изобретению, идентифицированные in vitro, могут быть протестированы с помощью данного анализа отека уха или других анализов на ГЗТ, как описанные выше, для выбора дополнительных блокаторов LTR, которые были бы применимы для уменьшения тяжести связанных с Th1-клетками иммунных реакций in vivo. Блокаторы LTR не ингибируют опосредованный Тh2-клетками (гуморальный) иммунный ответ. Как показано выше, блокаторы LTR по данному изобретению способны ингибировать опосредованный Th1-клетками эффекторный механизм, такой как контактная гиперчувствительность замедленного типа. Данная опосредованная Th1-клетками реакция ингибируется без оказания значительного влияния на зависимые от ThS-клеток реакции. Дифференциальный эффект блокаторов LTR на опосредованные Th1-клетками иммунные реакции был продемонстрирован с помощью наблюдения за зависимыми от Тh2-клеток реакциями, такими как первичный гуморальный ответ и переключение изотипов, в присутствии блокатора LTR. Мышам 5 раз в течение десятидневного промежутка времени вводили либо растворимый гибридный белок LTR (mLTR-Fc; пример 2), либо контрольный гибридный белок IgG человека (LFA3FC), либо оставляли необработанными. После второго введения всем мышам вводили в основание хвоста 100 мкл полного адъюванта Фройнда, содержащего 100 мкг овальбумина. Через 11 дней анализировали первичные сывороточные титры антител, специфичных к овальбумину, с помощью ELISA, специфичного к изотипам IgG1, IgG2a и IgM. Показан также эффект блокатора мышиного LTR mLTR-Fc на продукцию сывороточных антител к овальбумину у мышей, иммунизированных овальбумином. Введение блокатора LTR не влияет значительно на первичные титры антител после иммунизации овальбумином. В качестве сравнения,вмешательство в процесс СD40-лиганд-индуцированной передачи сигнала через рецептор CD40 полностью ингибирует антиген-специфичный Ig-G ответ у мышей (Renshaw et al., J. Exp. Med., 180, стр. 18891900 (1994. CD40 представляет собой еще одну пару лиганд/рецептор из семейства TNF. Весь процесс продукции и созревания иммуноглобулинов, несомненно, является зависимым от Тh2 клеток. Однако также очевидно, что продуцируемый Th1 цитокин IFN- принимает участие, но не является абсолютно необходимым для переключения на подкласс IgG2a (Huang et al., Science, 259, стр. 174245 (1993. Блокатор LTR mLTR-Fc не ингибировал переключение на IgG2a в данных экспериментах. Возможно, что блокаторы LTR по данному изобретению не ингибируют данное гуморальное направление опосредованной Th1-клетками реакции. В дополнение к этому, параметры пролиферативных реакций лимфоцитов из мышей, обработанных mLTR-Fc, не снижались. Результаты данных экспериментов указывают на то, что лечение, основанное на введении блокаторов LTR по данному изобретению, не окажет неблагоприятного влияния на функции Тh2-зависимой продукции антител при развитии иммунного ответа. Нормальный профиль гуморального ответа также указывает, что интенсивная обработка растворимым mLTR-Fc была нетоксичной для мышей, что далее указывает на применимую в терапии природу композиций и способов, изложенных далее в данном изобретении.- 13005734 Заболевания, опосредованные Т-хелперными клетками. Оказывается, что во многие органоспецифичные аутоиммунные состояния вовлечен патологический Тh1-ответ. По этим данным был составлен обзор (Modin and Nutman, Current Opinion in Immunol., 5,стр. 511-17 (1993); Romagnani et al., Ann. Rev. Immunol., 12, стр. 227-57 (1994. Данные органоспецифичные аутоиммунные состояния включают в себя рассеянный склероз, инсулинзависимый сахарный диабет, симпатическую офтальмию, увеит и псориаз. Инсулинзависимый сахарный диабет представляет собой аутоиммунное заболевание, при котором инсулин-продуцирующие бета-клетки поджелудочной железы разрушены лейкоцитами, инфильтрующими островки Лангерганса. Диабет может быть с легкостью индуцирован у новорожденных нетучных диабетических (neonatal nonobese diabetic) (NOD) мышей путем переноса активированных преддиабетических спленоцитов. Недавно Тh1- и Тh2-подобные клетки, по другим признакам генетически идентичные, переносили новорожденным мышам NOD. Быстрое развитие диабета индуцировали только Тh1 клетки, причем почти у всех реципиентов (Katz et al., Science, 268, стр. 1185-88 (1995. Данный факт указывает на то, что блокаторы LTR по данному изобретению, которые способны ингибировать эффекты опосредованной Тh1-клетками иммунной реакции in vivo, будут применимы для лечения или предотвращения инсулинзависимого сахарного диабета. Некоторые системные аутоиммунные заболевания, включая различные поражения кожи ревматического происхождения, являются ассоциированными с Th1-клетками. Оказывается, что в развитие как ревматоидного артрита, так и сухого кератоконъюнктивита вовлечены ThO- и Th1-клетки. В противоположность этому, системная красная волчанка (СКВ) протекает по типу аберрантной реакции с преобладанием роли ТhО/Тh2. Некоторые хронические воспалительные заболевания также протекают по типу аберрантной Тh1 опосредованной реакции, включая синдром воспаленной (раздраженной) толстой кишки, саркоидоз легкого и отторжение аллотрансплантата. Синдром воспаленной (раздраженной) толстой кишки (СВТК) у людей включает в себя, по крайней мере, две категории: язвенный колит и болезнь Крона. Считается, что оба заболевания являются результатом иммунопатологических нарушений аутоиммунного типа. В некоторых моделях СВТК на мышах очевидно, что некоторые агенты, которые ингибируют Th1 опосредованные реакции, способны ингибировать развитие или течение заболевания (F. Powrie et al.,Immunity 1:553 1994). Возможно, что ингибирование Th1-зависимого компонента иммунного ответа возымеет благоприятные эффекты при СВТК у человека. Многие модели СВТК были описаны и включены в обзоры (С. Elson et al., Gastroenterology 109:1344 1995). Существует, по крайней мере, три группы моделей: индуцированные химическим агентом, индуцированные полимерным/микробным агентом и модели иммунологического типа с использованием мутантных мышей. В одной часто применяемой индуцированной полимерным/микробным агентом модели раствор декстрансульфата вводят мышам в питьевую воду, и после приема внутрь эпителиальная выстилка кишечника раздражается, что ведет к массивному иммунному ответу на повреждение. У животных развивается колит, который проявляется в виде диарреи, стула с кровью, потери массы тела и уменьшения длины кишечника благодаря утолщению стенки кишечника. В данной модели развиваются признаки язвенного колита: левосторонний колит и дисплазия эпителия, которая может привести к образованию злокачественной опухоли. Вторая модель включает в себя трансплантацию отобранной группы CD4 Т-клеток мышам scid, т.е. мышам, у которых отсутствуют Т- и В-клетки (F. Powrie et al., International Immunology 5:1461-1471 1993;Morrissey et al., J. Exp. Med. 178:237 1993). Так как отобранные клетки, называемые CD45RBhi-клетками,распространяются и заселяют лимфоидные органы мышей scid, нормальные механизмы предотвращения возникновения аутореактивных Т-клеток не способны функционировать нормально и аутореактивные клетки развиваются. У крыс наблюдается появление клеток, реактивных ко многим органам, тогда как у мышей реактивность наблюдается преимущественно к кишечнику. В данной модели эффективными будут являться либо агенты, которые изменяют характер распространения и развития аутореактивных клеток, либо агенты, которые ингибируют способность клеток атаковать клетки кишечника. Более того, поскольку данная модель, по крайней мере, частично имитирует патологическое развитие аутореактивных иммунокомпетентных клеток, обработка, с помощью которой ингибируется данная модель, фактически может иметь модифицирующий течение заболевания эффект у человека. В данной модели развитие заболевания может быть ингибировано с помощью антител к TNF (F. Powrie et al., Immunity 1, 552 1994), и было обнаружено, что данные антитела эффективны в лечении заболеваний человека (Н.М. van Dullemenet al., Gastroenterology 109:109 1995). Таким образом, с помощью данной модели можно предсказать, какие агенты могут найти терапевтическое применение при СВТК. Более того, поскольку модель CD45RB является примером развития Тh1-опосредованного заболевания и фактически у крыс в данной модели развиваются заболевания многих органов, эффективность LTR-Ig в данной системе указывает на то,что LTR-Ig или другие средства, ингибирующие взаимодействие LTR со своим лигандом, могут оказать благоприятный эффект в лечении широкого круга родственных иммунных заболеваний.- 14005734 Вообще, точный размер вклада аутоантител по сравнению со специфичными Т-клетками при данных аутоиммунных заболеваниях не был описан. Клеточные реакции могут вносить основной вклад в патогенез тех системных аутоиммунных заболеваний, которые в настоящее время считаются вызываемыми преимущественно антителами, например различные поражения кожи ревматического происхождения. Нормальный иммунный ответ на некоторые патогенные инфекционные агенты также включает в себя Th1-опосредованный ответ, который может стать избыточным и начать представлять медицинскую проблему сам по себе. Примеры гранулематозных реакций (класса реакции ГЗТ, описанного выше), которые ведут к развитию тяжелых проблем со здоровьем, включают в себя лепру, образование гранулем в легких больных туберкулезом, саркоидоз и шистосомоз (Roitt et al., Immunology, стр. 22.5-6 (Mosby-YearBook Europe Ltd., 3d ed. 1993. Видимо, течение псориаза также опосредовано Th1-клетками. Цитолитические Т-клетки, т.е. CTL (CD8-положительные Т-клетки) также могут быть подразделены на Th1- и Th2-подобные популяции. Таким образом, возможно, что многое из того, что известно о группах Th, также применимо и к СD8+-клеткам, которые преимущественно вовлечены в антивирусный ответ и реакцию отторжения трансплантированной ткани. Способы лечения с применением блокаторов LTR. Композиции по данному изобретению будут вводиться в эффективной дозе для направленного лечения конкретного клинического состояния. Определение предпочтительного фармацевтического состава и терапевтически эффективной схемы приема лекарственного средства для данного применения проводят по методике, известной в данной области, принимая во внимание, например, состояние и массу тела пациента, объем желаемого лечения и переносимость пациентом такого лечения. Ожидается, что дозы растворимого LTR приблизительно в 1 мг/кг будут являться подходящими начальными точками для оптимизации терапевтических доз. Определение терапевтически эффективной дозы также может быть проведено путем постановки экспериментов in vitro, с помощью которых измеряют концентрацию блокатора LTR, необходимую для того, чтобы покрыть клетки-мишени (LTR- или LT-лиганд-положительных клеток в зависимости от блокатора) на срок от 1 до 14 суток. Анализы лиганд-рецепторного связывания, описанные здесь, могут быть применены для контроля за реакцией покрытия клеток. LTR- или LT-лиганд-положительные клетки могут быть отобраны из популяций активированных лимфоцитов с помощью FACS. На основании результатов данных анализов связывания in vitro можно выбрать интервал подходящих концентраций блокатора LTR для испытания на животных в соответствии с анализами, описанными здесь. Введение растворимых молекул LTR Аb к LT-лиганду и к LT-B-R по данному изобретению,одних или в сочетании, включая выделенные и очищенные формы антител или комплексов, их солей и их фармацевтически пригодных производных, может быть завершено с помощью любого общепринятого способа введения агентов, которые проявляют иммуносупрессивную активность. Фармацевтические композиции, применяемые в данных способах лечения, также могут находиться во множестве форм. Они включают в себя, например, твердые, полутвердые и жидкие дозированные формы, такие как таблетки, пилюли, порошки, жидкие растворы или суспензии, суппозитории и растворы для инъекций и инфузий. Предпочтительная форма зависит от предполагаемого способа введения и терапевтического применения. Способы введения могут включать пероральное, парентеральное, подкожное, внутривенное, внутриочаговое и местное введение. Растворимые молекулы LTR Аb к LT-лиганду и к LTR по данному изобретению могут быть помещены, например, в стерильные изотонические составы, содержащие или не содержащие кофакторы,которые стимулируют поглощение или устойчивость. Состав является, предпочтительно, жидким или может представлять собой лиофилизированный порошок. Например, растворимые молекулы LTR Ab к LT-лиганду и к LTR по данному изобретению могут быть разбавлены буферным составом, содержащим 5,0 мг/мл моногидрата лимонной кислоты, 2,7 мг/мл трехзамещенного цитрата натрия, 41 мг/мл маннитола, 1 мг/мл глицина и 1 мг/мл полисорбата 20. Данный раствор может быть лиофилизирован,положен на хранение в замороженном состоянии, а объем может быть доведен до введения стерильнымWater-For-Injection (USP). Также композиции будут, предпочтительно, содержать традиционные фармацевтически пригодные носители, хорошо известные в данной области (смотри, например, Remington's Pharmaceutical Sciences,16th Edition, 1980, Mac Publishing Company). Такие фармацевтически пригодные носители могут включать другие лекарственные средства, носители, генетические носители (genetic carriers), адъюванты, разбавители и т.д., такие как человеческий сывороточный альбумин или препараты плазмы. Композиции, предпочтительно, находятся в форме разовых доз и обычно будут вводиться 1 или более раз в сутки. Фармацевтические композиции по данному изобретению могут также быть введены с применением микросфер, липосом, других систем доставки на основе микрочастиц или составов продолжительного высвобождения непосредственно в пораженные ткани, в их окружение или иным образом в контакт с пораженными тканями или с кровотоком. Показательные примеры носителей продолжительного высвобождения включают полупроницаемые полимерные матрицы в форме заостренных предметов, таких как- 15005734 суппозитории или микрокапсулы. Имплантируемые или микрокапсулярные матрицы продолжительного высвобождения включают полилактиды (патент США 3773319; ЕР 58481), сополимеры Lглютаминовой кислоты и этил-L-глютамата (Sidman et al., Biopolymers, 22, стр. 547-56 (1985; поли(2 гидроксиэтилметакрилат) или этиленвинилацетат (Langer et al., J. Biomed. Mater. Res., 15, стр. 167-277(1981); Langer, Chem. Tech., 12, стр. 98-105 (1982. Липосомы, содержащие растворимые молекулы LTR Аb к LT-лиганду и к LTR по данному изобретению, одни или в сочетании, могут быть получены в соответствии с хорошо известными способами (смотри, например, DE 3218121; Epstein et al., Proc. Natl. Acad. Sci. U.S.A., 82, стр. 3688-92 (1985);Hwang et al., Proc. Natl. Acad. Sci. U.S.A. 77, стр. 4030-34 (1980); патенты США 4485045 и 4544545). Обычно липосомы относятся к типу небольших (приблизительно 200-800 ) однословных липосом, содержание холестерина составляет более 30 мол.% от содержания липидов. Пропорцию содержания холестерина выбирают так, чтобы контролировать оптимальную скорость высвобождения растворимых молекул LTR Аb к LT-лиганду и к LTR. Растворимые молекулы LTR Аb к LT-лиганду и к LTR по данному изобретению также могут быть присоединены к липосомам, содержащим другие блокаторы LTR, иммуносупрессивные средства или цитокины для модулирования LTR-ингибирующей активности. Присоединение к липосомам молекул LTR, Аb к LT-лиганду и к LTR может быть проведено с помощью любого известного поперечно-сшивающего агента, такого как гетеробифункциональные агенты, которые широко применяли для присоединения токсинов или химиотерапевтических средств к антителам для направленной доставки. Конъюгация с липосомами также может быть проведена с помощью углеводнаправленного поперечносшивающего реагента гидразида 4-(4-малеимидофенил)масляной кислоты (ГМФМ) (Duzgunes et al., J.Cell. Biochem. Abst. Suppl. 16E 77 (1992. Преимущества терапевтических композиций, содержащих блокаторы LTR. Блокаторы LTR по данному изобретению способны избирательно ингибировать зависимые отTh1- и Th2-клеток иммунные эффекторные механизмы. Блокаторы LTR будут применимы в лечении состояний, которые обостряются благодаря активностям цитокинов, продуцируемых Th1 (например, IL-2 и IFN-). Поскольку цитокины, продуцируемые Тh1, способны ингибировать реакции, зависимые от Тh2 клеток, блокаторы LTR могут также косвенно стимулировать определенные реакции, зависимые от Тh2-клеток, которые в норме ингибируются Тh1-индуцированными цитокиновыми каскадами. Способность избирательно подавлять клеточные реакции, опосредованные Тh1 (или косвенно стимулировать таковые, опосредованные Th2), будет применима для лечения аномалий при разнообразных клеточно-опосредованных иммунных реакциях, включая различные аутоиммунные и хронические воспалительные состояния, толерантность к антигенам и клеточную реакцию отторжения трансплантатов тканей и органов. Как обсуждалось выше, в лечении иммунных состояний с преобладающей ролью Тh1-клеток обычно применяют иммуномодуляторы и иммуносупрессоры, которые обладают плейотропными эффектами на широкий круг типов клеток и иммунных реакций. Обычно требуется применение данных неспецифических иммуносупрессоров в высоких и зачастую цитотоксических дозах, что вызывает неблагоприятные побочные эффекты. Наличие способности изменять характер иммунного ответа подтверждено недавними исследованиями сахарного диабета у мышей, что обсуждалось выше (Katz et al., Science, 268, стр. 1185-88 (1995,и модели аллогенной трансплантации (Sayegh et al., J. Exp. Med., 181, стр. 1869-74 (1995. В последнем из них было показано, что гибридный белок, который ингибирует костимуляторный путь CD28-B7 Тклеток, вызывает толерантность к трансплантату почки. Толерантность коррелировала со снижением концентрации цитокинов, продуцируемых Th1, и повышением концентрации цитокинов, продуцируемыхTh2, in vivo. Эти данные указывают на то, что блокаторы LTR по данному изобретению будут применимы для подавления клеточной реакции отторжения трансплантатов тканей и органов путем подавления секреции цитокинов Th1-клетками. Блокаторы LTR из композиций и способов по данному изобретению могут быть модифицированы с получением желаемого уровня передачи сигнала через LTR в зависимости от состояния, подвергающегося лечению нарушения или заболевания. Представляется, что абсолютный уровень передачи сигнала через LTR может быть точно настроен путем манипулирования концентрацией и значениями сродства блокаторов LTR к соответствующим им молекулярным мишеням. Например, в одном осуществлении данного изобретения субъекту вводят композиции, содержащие растворимые молекулы LTR. Растворимый рецептор к LT- способен эффективно конкурировать с рецепторами к LT- клеточной поверхности за связывание поверхностных LT-лигандов. Способность конкурировать за связывание поверхностных LT-лигандов зависит от относительных концентраций растворимых молекул LTR и молекул LTR клеточной поверхности и от их относительных аффинностей к связыванию лиганда.- 16005734 Растворимые молекулы LTR, несущие мутации, которые повышают или снижают сродство связывания данного мутантного растворимого LTR с поверхностным лигандом, могут быть получены с применением стандартных методик рекомбинантной ДНК, хорошо известных специалистам в данной области. Большие количества молекул с сайт-специфичными или случайными мутациями могут быть протестированы на предмет их способности действовать как блокаторы LTR с применением устоявшихся методик постановки эксперимента и анализов, описанных здесь. Сходным образом, в другом осуществлении данного изобретения антитела либо к рецептору к LT, либо к одной или более субъединицам LT-лиганда функционируют как блокаторы LTR. Способность данных антител ингибироватъ передачу сигнала через LTR может быть модифицирована с помощью мутации, химической модификации или других методов, с помощью которых можно изменять эффективную концентрацию или активность антител, введенных субъекту. Способность снижать передачу сигнала через LTR без ее полного ингибирования может оказаться важной для установления или поддержания сниженных уровней передачи сигнала через LTR, которые поддерживают нормальное функционирование иммунной системы, при ингибировании реакций,опосредованных Th1-клетками, когда те являются избыточными или аномальными. Разрушения гена LT- у мышей приводит к отклонению от нормы в развитии периферических лимфоидных органов (De Togni et аl., Science, 264, стр. 703-7 (1994. У таких мышей отсутствовали лимфатические узлы, а в их селезенке отсутствовало обычно четкое разделение между Т- и В-зонами в фолликулах. Авторы считают, что данный фенотип связан с утратой индуцированной поверхностнымLT-лигандом передачи сигнала через LTR, поскольку при модулировании активности TNF-R сходные фенотипы не наблюдались. Таким образом, способность избирательно или частично ингибировать LT-R-путь может быть применена в лечении аномалий развития лимфоидных органов, связанных с аномальной или избыточной экспрессией передачи сигнала через LTR-путъ. Некоторые Th1-ассоциированные реакции являются принципиально важными компонентами ряда клеточно-опосредованных иммунных ответов (Romagriani et al., Ann. Rev. Immunol., 12, стр. 227-57(1994, и полное ингибирование Th1-клеточной активности при определенных обстоятельствах может быть нежелательным. Например, если может развиться значительный Тh1-ответ, мышь может эффективно противостоять паразитарной инфекции. Инфекционные агенты, такие как Listeria и Toxoplasma, также вызывают сильные ответы Тh1-типа. У человека в ответе на инфекцию Mycobacterium tuberculosis преобладающую роль играют Тh1. Патогенность возбудителя лейшманиоза коррелирует с реакциями, сходными с Th1-ответами, охарактеризованными у мышей (Reed and Scott, Current Opinion in Immunol., 5, стр. 524-31 (1993. Возможность оказывать влияние на уровень ингибирования Тh1 путем ингибирования передачи сигнала через LTR может оказаться важной для получения предельно полезных результатов, которые только могут быть получены путем обработки блокаторами LTR по данному изобретению. Далее следуют примеры, которые иллюстрируют растворимые молекулы LTR Ab к LT-лиганду и к LTR по данному изобретению и способы, примененные для их охарактеризования. Данные примеры не следует истолковывать как ограничивающие: примеры включены в целях иллюстрации, а настоящее изобретение ограничено лишь формулой изобретения. Пример 1. Получение растворимых человеческих рецепторов к LT- в виде гибридных белков с Fcдоменом иммуноглобулина. Последовательность из клона человеческой кДНК, выделенную из библиотеки человеческих транскрибируемых последовательностей 12 р, взятой из гибридной соматической клетки (Baens et al., Genomics,16, стр. 214-18 (1993, ввели в GenBank, а затем идентифицировали как последовательность, которая кодирует человеческий LTR. Последовательность данного полного клона кДНК человеческого LTR доступна с 1992 года как ввод в GenBank L04270. Внеклеточный домен LTR до трансмембранного фрагмента (фиг. 1) амплифицировали с помощью PCR с клона кДНК с применением праймеров, которые включали в себя сайты рестрикции NotI иSalI на 5'- и 3'-концах соответственно (Browning et al., J. Immunol., 154, стр. 33-46 (1995. Амплифицированный продукт вырезали с помощью NotI и SalI, очищали и лигировали к NotI-линеаризованному вектору pMDR901 вместе с SalI-NotI-фрагментом, кодирующим Fc-домен человеческого IgG1. Полученный вектор содержал ген дигидрофолатредуктазы и ген гибридного белка LTR-Fc, транскрипция которых запускалась отдельными промоторами. Вектор электропорировали в клети СНО dhfr- и выделяли метотрексат-резистентные клоны согласно стандартным методикам. LTR-Fc секретировался в среду, и для выбора клеточных линий, продуцирующих наивысший уровень рецепторного гибридного белка, применяли ELISA. Клеточные линии с высоким уровнем продукции культивировали в больших количествах и кондиционную среду отбирали. Чистый гибридный белок рецептора к LT- выделяли по методу высокоскоростной аффинной хроматографии на Protein A Sepharose (Pharmacia).- 17005734 Пример 2. Получение растворимых мышиных рецепторов к LT- в виде гибридных белков с Fcдоменом иммуноглобулина. Полный клон кДНК mLTR получали путем лигирования 5'-NotI/ApalI- и 3'-ApalI/MolIфрагментов из двух частичных изолятов кДНК в сайт NotI pCDMA3 (InVitrogen, San Diego, CA). Последовательность данного клона кДНК доступна как ввод в GenBank U29173. При сравнении с другой последовательностью, кодирующей mLTR, находящейся в GenBank под номером L38423, не было отмечено различий в кодирующих последовательностях. Растворимый гибридной белок mLTR (hIgG1) получали путем амплификации с помощью PCR,используя полный клон кДНК mLTR в качестве матрицы и праймеры 5'-AACTGCAGCGGCCGCCATGCGCCTGCCC-3' и 5'-GACTTTGTCGACCATTGCTCCTGGCTCTGGGGG-3'. Амплифицированный продукт очищали и вырезали с помощью NotI/SalI и aктивировали с SalI/NotI Fc-доменом человеческогоIgG1 в NotI-линеаризованный и обработанный фосфатазой вектор SAB132 с получением JLB122. В целях обеспечения устойчивой экспрессии в сайт NotI/pMDR901 переносили кассету NotI, содержащую mLT-R-Fc-фрагмент, с получением PSH001 и данным вектором трансфицировали клетки СНО, как было описано (Browning et al., J. Immunol., 154, стр. 33-46 (1995. Клоны клеток, секретирующих mLTR-Fc,идентифицировали с помощью ELISA. Очищенный рецепторный гибридный белок выделяли из супернатанта клеток СНО по методу высокоскоростной аффинной хроматографии на Protein A Sepharose (Pharmacia). Пример 3. Применение растворимого человеческого LTR-Fc для ингибирования взаимодействия рецептора к LT- и его лиганда. Растворимый hLTR-Fc тестировали на предмет его способности ингибировать связывание LTлиганда с рецептором к LT- с помощью анализа цитотоксичности, направленной на опухолевые клетки,описанного выше. В данном анализе растворимую форму LT-лиганда (LT-1/2), которая стимулирует передачу сигнала через LTR, применяют для цитолиза человеческих опухолевых клеток. Ингибиторы передачи сигнала через LTR способны снижать LTR-индуцированную цитотоксичность, направленную на опухолевые клетки. Растворимые LT-1/2-лиганды состоят из усеченных или модифицированных субъединиц LTбез функционального трансмембранного домена. Растворимые LТ-1/2-лиганды связываются с LTR и стимулируют передачу через него сигнала так же, как и поверхностные формы LT-лиганда (Browning etal., J. Immunol., 154, стр. 33-46 (1995. Серийные разведения hLT-1/2, hTNF или hLT- готовили в 0,05 мл в 96-луночных планшетах и 5000 трипсинизированных клеток НТ 29 (АТСС) вносили в 0,05 мл среды, содержащей 80 Ед./мл (в антивирусных единицах) hu-INF-. Через 4 дня содержание восстановленного митохондриями красителя МТТ измеряли следующим образом: добавляли 10 мкл МТТ и через 3 ч восстановленный краситель растворяли 0,09 мл изопропанола с 10 мМ НСl и ОП измеряли на 550 нм. До внесения клеток добавляли растворимые формы рецептора или чистый человеческий Ig-G в 10 мкл с получением конечной концентрации 5 мкг/мл. В таблице ниже сравнивается способность hLTR-Fc и химерных молекул p55-TNF-R-Fc (с человеческим IgG в качестве контроля) блокировать ингибирующие эффекты различных растворимых TNF- иLT-лигандов на рост опухолевых клеток НТ 29. Способность белков LTR и p55-TNF-R, гибридизованных с иммуноглобулином, блокировать ингибирующие эффекты различных TNF- и LT-лигандов на рост НТ 29. Концентрация цитотоксического агента (нг/мл), приводящая к 50%-ному ингибированию роста. В присутствии а. Контрольный hIgGLT-1/2 а Каждый цитотоксический агент предварительно смешивали с белками, гибридизованными с Ig, за 10 мин до добавления к клеткам. Конечная концентрация гибридного белка составила 5 мкг/мл.b Более высокие концентрации не тестировали. Данные в таблице указывают на то, что растворимый гибридный белок человеческого LTR (hLT-R-Fc) способен эффективно ингибироватъ взаимодействие между LT-лигандом (LT-1/2) и рецепторами к LT- клеточной поверхности и, таким образом, является блокатором LTR по данному изобретению. Как и ожидалось, растворимый гибридный белок TNF-R (р 55-TNF-R-Fc) полностью блокировалTNF-индуцированное ингибирование роста путем связывания с TNF и предотвращения его взаимодействия с поверхностными рецепторами к TNF. Данный растворимый рецептор к TNF не оказывает влияния на LT-лиганд-опосредованные антипролиферативные эффекты. В противоположность этому, LTR-Fc ингибировал LT-лиганд-индуцированные цитотоксические эффекты, но не таковые, индуцированныеTNF или LT-. Таким образом, гибридные белки человеческого LTR не вмешиваются в активациюTNF-R TNF и лигандами LT-. Пример 4. Применение растворимого мышиного LTR-Fc для ингибирования взаимодействия мышиного рецептора к LT- и его лиганда. Растворимый мышиный рецептор к LT-, сшитый с Fc-доменом человеческого IgG1 (mLTR-Fc: смотри пример 2) тестировали на предмет его способности ингибироватъ связывание рецептора к LTсо своим лигандом с помощью анализа цитотоксичности, направленной на мышиные клетки (фиг. 2). Анализ цитотоксичности проводили на клетках WEHI 164, применяя, по существу, ту же методику, которая была применена в анализе на клетках НТ 29, описанном в примере 3 (смотри также Browning andRibolini, J. Immunol., 143, стр. 1859-67 (1989. На фиг. 2 показаны эффекты mLTR-Fc на лиганд-индуцированную передачу сигнала через LTR на мышиных клетках WEHI 164. Как показано данным анализом, клетки WEHI 164 гибнут при обработке растворимым LТ-1/2-лигандом в концентрациях в интервале от 1 до 100 нг/мл. mLTR-Fc ингибирует LT-лиганд-стимулированную клеточную гибель. Добавление растворимого мышиного гибридного белка p55-TNF-R-Fc или контрольных антител IgG (каждого по 10 /мл) обладает незначительным или не обладает эффектом на ингибирование клеточной гибели. Эти данные показывают, что гибридный белок mLTR-Fc способен эффективно конкурировать с поверхностными молекулами LTR за связывание LT-лиганда. Эти данные также показывают, что LТ/-индуцированная цитотоксичность является LTR-опосредованной и может быть ингибирована растворимым mLTR-Fc, который действует как блокатор LTR по настоящему изобретению. Перечень последовательностей ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ лечения ревматоидного артрита у млекопитающего, предусматривающий введение указанному млекопитающему фармацевтической композиции, содержащей терапевтически эффективное количество блокатора рецептора лимфотоксина- (LTR) и фармацевтически приемлемый носитель.- 19005734 2. Способ лечения ревматоидного артрита у млекопитающего, предусматривающий введение указанному млекопитающему растворимого рецептора лимфотоксина-, дополнительно содержащего Fcдомен человеческого иммуноглобулина. 3. Способ по п.1, согласно которому блокатор LTR выбран из группы, состоящей из растворимого рецептора лимфотоксина-, антитела против рецептора лимфотоксина- и антитела против поверхностного LT-лиганда. 4. Способ по п.3, согласно которому растворимый рецептор лимфотоксина- содержит связывающий лиганд LTR домен, который способен селективно связываться с поверхностным LT-лигандом. 5. Способ по п.4, согласно которому растворимый рецептор лимфотоксина- содержит Fc-домен имммуноглобулина человека. 6. Способ по п.3, согласно которому блокатор LTR представляет собой моноклональное антитело против рецептора лимфотоксина-. 7. Способ по п.6, согласно которому моноклональное антитело представляет собой антитело BDA8 против человеческого LTR. 8. Способ по п.3, согласно которому блокатор LTR представляет собой моноклональное антитело против поверхностного LT-лиганда. 9. Способ по п.8, согласно которому антитело направлено против субъединицы LT-лиганда. 10. Способ по п.8, согласно которому моноклональное антитело представляет собой антитело В 9 против человеческого LTR. 11. Способ по п.8, согласно которому блокатор LTR представляет собой моноклональное антитело против мышиного поверхностного LT-лиганда.
МПК / Метки
МПК: A61K 38/17, A61K 39/395, A61P 1/04, G01N 33/577, G01N 33/68
Метки: лечения, способ, ревматоидного, артрита
Код ссылки
<a href="https://eas.patents.su/21-5734-sposob-lecheniya-revmatoidnogo-artrita.html" rel="bookmark" title="База патентов Евразийского Союза">Способ лечения ревматоидного артрита</a>
Способ лечения артрита при использовании блокирующего моноклонального антитела к интегрину vla-1 или его фрагмента

Номер патента: 5242
Опубликовано: 30.12.2004
Авторы: Лобб Рой, Готвэлс Филип, Де Фужеролль Антонин, Котелянский Виктор
МПК: C07K 16/28, A61P 19/02, A61K 39/395...
Метки: использовании, блокирующего, лечения, интегрину, артрита, антитела, фрагмента, способ, vla-1, моноклонального
Формула / Реферат:
1. Способ лечения артрита, включающий введение субъекту, страдающему артритом, композиции, которая включает моноклональное антитело AJH10 или его фрагмент, взаимодействующие с эпитопом домена a1-I антигена "очень поздней активации" (VLA-1) семейства интегринов, включающим аминокислотные остатки 91-96, показанные на фиг. 15, (SEQ ID NO:8), в количестве, эффективном для обеспечения снижения показателя артрита приблизительно на 65% или более по...
Средство для лечения артрита и супрессии продукции ммр-9, содержащее гиалуронатный комплекс цинка и способы на его основе

Номер патента: 5010
Опубликовано: 28.10.2004
Авторы: Серизава Исао, Несмели Эржебет, Маекава Кейсей, Иллеш Янош
МПК: A61P 19/02, A61K 33/30
Метки: гиалуронатный, супрессии, основе, продукции, лечения, комплекс, ммр-9, средство, артрита, цинка, содержащее, способы
Формула / Реферат:
1. Средство для лечения артрита, включающее гиалуронатный комплекс цинка. 2. Средство по п.1, в котором гиалуронатный комплекс цинка имеет мол. массу от 100 до 2000 kD. 3. Средство по п.2, в котором гиалуронатный комплекс цинка имеет мол. массу около 1000 kD. 4. Средство по любому из пп.1-3, в котором весовое соотношение гиалуроновой кислоты и цинка составляет от 5:1 до 20:1. 5. Средство по п.4, в котором весовое соотношение гиалуроновой кислоты...
Стабильный однородный крем на основе нитроглицерина для локального применения , способ его приготовления , способ лечения эректильной дисфункции у мужчин, способ лечения аноргазма у женщин , способ лечения микроваскулярных заболеваний и способ лечения ран или хирургических рассечений

Номер патента: 566
Опубликовано: 29.12.1999
Автор: Аллан Майкл П.
МПК: A61K 31/21
Метки: аноргазма, мужчин, крем, способ, дисфункции, эректильной, приготовления, нитроглицерина, женщин, хирургических, лечения, локального, основе, микроваскулярных, рассечений, однородный, стабильный, заболеваний, применения, ран
Формула / Реферат:
1. Стабильный однородный крем на основе нитроглицерина для локального применения, отличающийся тем, что содержит следующие компоненты, в мас.%: Нитроглицерин 0,1-3 Усилитель пенетрации 5-24 Загуститель 0,5-3 Эмульгатор 0,4-2 Вода Остальное и имеет значение рН от приблизительно 6,0 до приблизительно 9,0. 2. Крем по п.1, отличающийся тем, что он содержит нитроглицерин...
Вакцина для лечения воспаления и доброкачественной гиперплазии предстательной железы на основе бактерий lactobacilli и способ лечения указанных заболеваний

Номер патента: 5198
Опубликовано: 30.12.2004
Авторы: Бартуш Йожеф, Лазар Эрика Агнеш, Варга Дьюла Арпад, Ийхейи Карой
МПК: A61K 39/02, A61P 13/08
Метки: указанных, способ, лечения, бактерий, основе, заболеваний, гиперплазии, доброкачественной, lactobacilli, воспаления, железы, предстательной, вакцина
Формула / Реферат:
1. Вакцина для лечения воспаления предстательной железы и доброкачественной гиперплазии предстательной железы (I и II стадии), содержащая штаммы Lactobacillus, входящие в состав вакцин GynevacR, GynatrenR и SolcoTrichovacR, в инактивированной форме, и носители и/или эксципиенты, обычно используемые в вакцинных препаратах. 2. Применение штаммов Lactobacillus, входящих в состав вакцин GynevacR, GynatrenR и SolcoTrichovacR, в инактивированной форме...
Способ лечения болезни сердца, способ лечения недостаточности периферических сосудов и способ ограничения доставки и экспрессии трансгенной конструкции в определенном органе или структуре

Номер патента: 1616
Опубликовано: 25.06.2001
Авторы: Диллмэн Вольфганг Х., Хэммонд Х.Кэрк, Джордано Фрэнк Дж.
МПК: C12N 15/00, A61K 48/00, A61P 9/10...
Метки: определенном, способ, лечения, конструкции, болезни, структуре, сосудов, ограничения, периферических, экспрессии, недостаточности, трансгенной, доставки, сердца, органе
Формула / Реферат:
1. Способ лечения болезни сердца путем стимуляции развития коронарных коллатеральных сосудов у пациента с ишемией миокарда, включающий в себя доставку аденовирусного вектора, дефектного по репликации, в миокард пациента посредством интракоронарной инъекции, причем указанную интракоронарную инъекцию осуществляют непосредственно в одну или обе коронарные артерии, а указанный вектор содержит трансгенную конструкцию, кодирующую ангиогенный белок или...
Предыдущий патент: Блок воздушного насоса для вентилируемой обуви
Следующий патент: Получение сверхчистого висмута-213 для использования в терапевтической ядерной медицине
Случайный патент: Устройство и способ распыления порошкообразного вещества в газе-носителе