Катионные виросомы в качестве системы для переноса генетического материала
Номер патента: 3130
Опубликовано: 27.02.2003
Авторы: Вельти Эрнст Рудольф, Глюк Райнхард, Кляйн Петер

















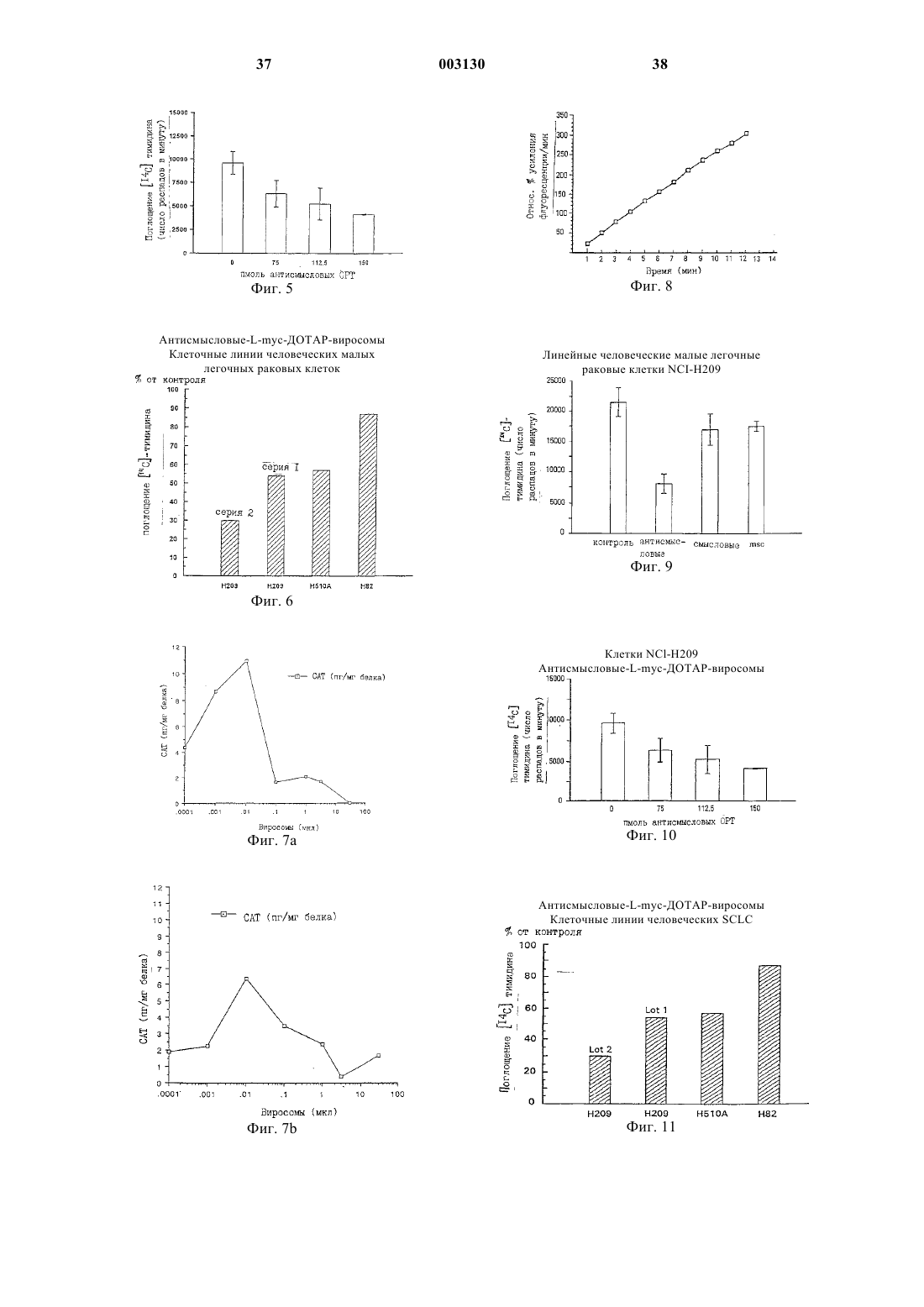
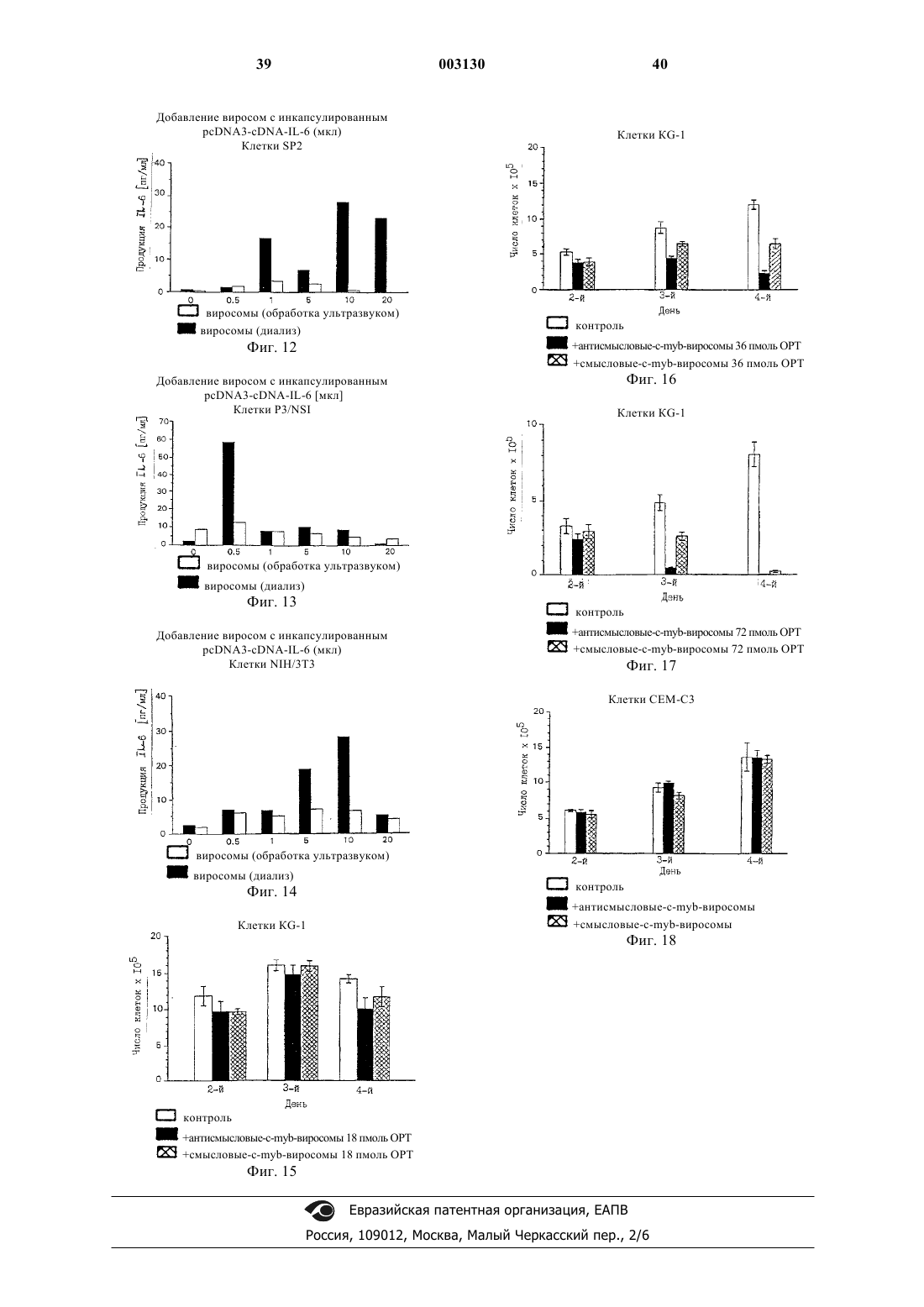
Формула / Реферат
1. Способ получения везикулы, имеющей положительно заряженную липидную бислойную мембрану, которая содержит катионные и/или поликатионные липиды вместе с фосфатидилэтаноламином или фосфатидилхолином и, по меньшей мере, одним функционально активным гемагглютинином вируса гриппа, контролирующим слияние, включенным в указанную мембрану или ковалентно связанным с мембраной, который способен индуцировать и/или способствовать реакции слияния между мембраной везикулы и липидной мембраной клетки-мишени, специфичным клеточным маркером, закрепленным на мембране, и необходимым генетическим материалом, характеризующийся тем, что он включает стадии
а) растворения очищенного вируса гриппа в растворе неионного детергента и выделения фракции, содержащей гемагглютининовый тример;
б) приготовления конъюгированных молекулярных комплексов, состоящих из фосфатидилэтаноламина (PE), бифункционально перекрестно-сшивающего агента и специфичного к клеткам маркера, таким образом, что бифункциональный перекрестно-сшивающий агент одним активным участком связывается с аминогруппой PE и другим - с тиоловой группой специфичного к клеткам маркера, и удаления неконъюгированного материала;
в) приготовления раствора, содержащего забуференный, неионный детергент, гемагглютинин вируса гриппа, полученный на стадии а), катионные и/или поликатионные липиды, фосфатидилэтаноламин (PE), необязательно фосфатидилхолин, перекрестно-сшивающий агент и, по меньшей мере, один специфичный к клеткам маркер в составе получаемых на стадии б) молекулярных комплексов PE/перекрестно-сшивающего агента/маркера, и доведения содержания липидов (к общему количеству липидов) до 90-95% по массе катионных и/или поликатионных липидов и 5-10% по массе фосфатидилэтаноламина; или 45-90% по массе катионных и/или поликатионных липидов, 5-50% по массе фосфатидилхолина и 5-10% по массе фосфатидилэтаноламина;
г) удаления детергента обработкой раствора гранулами-микроносителями, предпочтительно полистирольными гранулами Biobeads SM-2, что приводит к образованию положительно заряженных липидных бислойных везикул; и
д) добавления к везикулам необходимого количества нужного генетического материала для доставки в клетки-мишени, обработки смеси ультразвуком для включения материала в везикулы и удаления невключившегося материала, предпочтительно гель-фильтрацией.
2. Способ по п.1, согласно которому обеспечивают весовое соотношение гемагглютинина и общего содержания липидов в мембране, равное примерно 1 (мг/мг) или меньше.
3. Способ по п.1 или 2, согласно которому специфичный к клеткам маркер способен к избирательному обнаружению и связыванию с клетками-мишенями и выбран из группы, включающей антитело, предпочтительно моноклональное антитело, фрагмент антитела, предпочтительно фрагмент F(ab')2 или Fab', цитокин и фактор роста.
4. Способ по любому из пп.1-3, согласно которому специфичный к клеткам маркер представляет собой фрагмент Fab', а весовое соотношение гемагглютинина и указанного маркера составляет примерно 2:1.
5. Способ по любому из пп.1-4, согласно которому катионные липиды включают, по меньшей мере, одно соединение, выбранное из группы, состоящей из
N-[1,2,3-диолеоилокси)пропил]-N,N,N-триметиламмоний хлорида (DOTMA);
N-[1,2,3-диолеоилоксипропил]-N,N,N-триметиламмонийметилсульфата (DOTAP);
N-t-бутил-N'-тетрадецил-3-тетрадециламинопропионамидина; а
поликатионные липиды включают, по меньшей мере, одно соединение, выбранное из группы, состоящей из
1,3-дипальмитоил-2-фосфатидилэтаноламидоспермина (DPPES);
диоктадециламидоглицилспермина (DOGS);
2,3-диолеоилокси-N-[2-(сперминкарбоксамидо)этил]-N,N-диметил-1-пропанаминотрифторацетата (DOSPA);
1,3-диолеоилокси-2-(6-карбоксиспермил) пропиламида (DOSPER) и
N,N,N'N'-тетраметил-N,N'-бис-(2-гидроксиэтил)-2,3-диолеоилокси-1,4-бутандиаммоний йодида (THDOB-).
6. Способ по любому из пп.1-5, согласно которому указанный раствор неионного детергента включает C12E8 (монододециловый эфир октаэтиленгликоля) или н-октилолигооксиэтилен в концентрации 10-250 мкмоль на 1 мл HEPES буфера.
7. Способ по любому из пп.1-6, согласно которому указанные гранулы-микроносители имеют смачиваемый размер ячеек 20-50 меш. (0,84-0,30 мм) и полученный на стадии в) раствор обрабатывают гранулами-микроносителями до четырех раз.
8. Способ по любому из пп.1-7, согласно которому перекрестно-сшивающий агент является гетеробифункциональной органической молекулой, имеющей, по меньшей мере, одну малеимидную и, по меньшей мере, одну карбоксильнуюгруппу, и предпочтительно выбран из группы, включающей бис-N-сукцинимидильные производные и фотоактивируемые сукцинимидильные производные.
9. Способ по любому из пп.1-8, согласно которому неконъюгированный материал, особенно не прореагировавшие комплексы РЕ/ перекрестно-сшивающего агента такие, как N-[4-(п-малеимидофенил)бутирил]фосфатидилэтаноламин (MPB.PE) или 4-(N-малеимидометил) циклогексан-1-карбоксилатфосфатидилэтаноламин (MCC.PE), отделяют от продуктов реакции с помощью гель-хроматографии, предпочтительно аффинной хроматографией на агарозном носителе, предпочтительно на восстановленной тиопропилсефарозе 6B.
10. Способ по любому из пп.1-9, согласно которому необходимый генетический материал выбран из группы, включающей короткие цепи ДНК или РНК, дезоксирибонуклеотиды, олигодезоксирибонуклеотиды, олигодезоксирибонуклеотиды селеноатов, олигодезоксирибонуклеотиды фосфоротиоатов, олигодезоксирибонуклеотиды фосфорамидатов, олигодезоксирибонуклеотиды метилфосфонатов, комплексы пептида с нуклеиновой кислотой, рибонуклеотиды, олигорибонуклеотиды, олигорибонуклеотиды фосфоротиоатов, 2'-ОМе-олигорибонуклеотиды фосфатов, 2'-Оме-олигорибонуклеотиды фосфоротиоатов, рибозимы, гены, плазмиды и векторы.
11. Липидная везикула, имеющая положительно заряженную липидную бислойную мембрану, которая содержит катионные и/или поликатионные липиды вместе, по меньшей мере, с одним функционально активным гемагглютинином вируса гриппа, включенным в указанную мембрану или ковалентно связанным с мембраной, который способен индуцировать и/или способствовать реакции слияния между мембраной везикулы и липидной мембраной клетки-мишени, характеризующаяся тем, что
а) мембрана везикулы содержит (к общему количеству липидов) 90-95% по массе катионных и/или поликатионных липидов и 5-10% по массе фосфатидилэтаноламина; или 45-90% по массе катионных и/или поликатионнных липидов, 5-50% по массе фосфатидилхолина и 5-10% по массе фосфатидилэтаноламина;
б) мембрана включает, по меньшей мере, один бифункциональный перекрестно-сшивающий агент, прикрепленный к мембране посредством ковалентного связывания с фосфатидилэтаноламином;
в) везикула включает, по меньшей мере, один специфичный к клеткам маркер, способный к избирательному обнаружению и связыванию с клетками-мишенями, выбранный из группы, включающей антитело, фрагмент антитела, цитокин и фактор роста, причем указанный маркер связан с мембраной с помощью упомянутого перекрестно-сшивающего агента таким образом, что перекрестно-сшивающий агент реагирует со свободной тиоловой группой специфичного к клеткам маркера; и
г) везикула содержит необходимый генетический материал для доставки в клетки-мишени.
12. Везикула по п.11, отличающаяся тем, что
катионные липиды включают, по меньшей мере, одно соединение, выбранное из группы, состоящей из
N-[1,2,3-диолеоилокси)пропил]-N,N,N-триметиламмония хлорида (DOTMA);
N-[1,2,3-диолеоилокси)пропил]-N,N,N-триметиламмонийметилсульфата (DOTAP);
N-t-бутил-N'-тетрадецил-3-тетрадециламинопропионамидина; a
поликатионные липиды включают, по меньшей мере, одно соединение, выбранное из группы, состоящей из
1,3-дипальмитоил-2-фосфатидилэтаноламидоспермина (DPPES);
диоктадециламидоглицилспермина (DOGS);
2,3-диолеоилокси-N-[2-(сперминкарбоксамидо)этил]-N,N-диметил-1-пропанаминотрифторацетата (DOSPA);
1,3-диолеоилокси-2-(6-карбоксиспермил) пропиламида (DOSPER) и
N,N,N',N'-тетраметил-N,N'-бис-(2-гидроксиэтил)-2,3-диолеоилокси-1,4-бутандиаммоний йодида (THDOB).
13. Везикула по любому из пп.11 или 12, характеризующаяся тем, что мембрана включает 1,3-диолеоилокси-2-(6-карбоксиспермил) пропиламид (DOSPER) в качестве поликатионного липида.
14. Везикула по одному из пп.11-13, характеризующаяся тем, что необходимый генетический материал, включенный в указанную везикулу, выбран из группы, включающей цепи ДНК или РНК, дезоксирибонуклеотиды, олигодезоксирибонуклеотиды, олигодезоксирибонуклеотиды селеноатов, олигодезоксирибонуклеотиды фосфоротиоатов, олигодезоксирибонуклеотиды фосфороамидатов, олигодезоксирибонуклеотиды метилфосфонатов, комплексы пептида с нуклеиновой кислотой, рибонуклеотиды, олигорибонуклеотиды, олигорибонуклеотиды фосфоротиоатов, 2'-ОМе-олигорибонуклеотиды фосфатов, 2'-ОМе-олигорибонуклеотиды фосфоротиоатов, рибозимы, гены, плазмиды и векторы.
15. Везикула по любому из пп.11-14, характеризующаяся тем, что ее диаметр находится в пределах 120-180 нм.
16. Везикула по любому из пп.11-15, получаемая способом по любому из пп.1-10.
17. Применение липидной везикулы по любому из пп.11-16 в качестве системы-переносчика для специфической и неинфекционной доставки необходимого генетического материала в клетки-мишени или ткани-мишени, особенно в покоящиеся или пролиферирующие клетки-мишени млекопитающих.
18. Применение по п.17, согласно которому доставку генетического материала осуществляют в целях профилактики и/или терапевтического лечения человека и животных.
19. Применение по любому из пп.17 или 18, согласно которому генетический материал предпочтительно включает, по меньшей мере, один антисмысловой олигонуклеотид, пригодный для антисмысловой терапии заболевания, выбираемого из группы, включающей рак, лейкемию и вирусную инфекцию.
20. Применение по п.19, согласно которому антисмысловой олигонуклеотид направлен на мРНК протоонкогена или онкогена.
21. Способ получения молекулярного комплекса, состоящего из фосфатидилэтаноламина (PE), гетеробифункционального перекрестно-сшивающего агента и специфичного к клеткам маркера, выбранного из группы, включающей антитело, фрагмент антитела, цитокин и фактор роста, включающий следующие стадии:
а) растворение специфического к клеткам маркера, предпочтительно Fab'-фрагмента моноклонального антитела, в буферном растворе, предпочтительно содержащем 100 мМ NaCl, 40 мМ лимонной кислоты, 35 мМ Na2HPO4Ч2H2O, 2 мМ ЭДТА и имеющем рН 5,5;
б) смешивание полученного раствора с раствором детергента, который включает 0,5% неионного детергента, предпочтительно н-октилолигооксиэтилена в цитратном буфере, и с заранее приготовленными структурами, состоящими из перекрестно-сшивающего агента и PE, предпочтительно N-[4-(п-малеимидофенил) бутирил]фосфатидилэтаноламина (MPB.PE), 4-(N-малеимидометил)циклогексан-1-карбоксилатфосфатидилэтаноламина (MCC.PE) или N-Y-малеимидобутирилоксифосфатидилэтаноламина (GMB.PE);
в) инкубирование смеси, предпочтительно при 4шC в течение 16 ч в атмосфере азота при мягком перемешивании для конъюгации специфичного к клеткам маркера с перекрестно-сшивающим агентом, входящим в состав заранее приготовленных структур, с образованием молекулярных комплексов и
г) удаление после инкубации свободных структур, содержащих перекрестно-сшивающий агент и РЕ, неконъюгированных специфичным к клеткам маркером с помощью аффинной хроматографии, используя агарозный матрикс, предпочтительно инкубируя смесь в течение 4 ч при комнатной температуре со свежевосстановленной увлажненной тиопропилсефарозой 6B.
22. Способ по п.21, согласно которому после окончания аффинной хроматографии агарозный матрикс удаляют предпочтительно центрифугированием и оставшийся раствор, содержащий молекулярные комплексы PE/перекрестно-сшивающий агент/маркер, нейтрализуют до pH 7,0 и при необходимости добавляют монододециловый эфир октаэтиленгликоля (C12E8).
23. Способ по любому из пп.21 или 22, согласно которому указанный гетеробифункциональный перекрестно-сшивающий агент является сукцинимидильным производным, имеющим, по меньшей мере, одну малеимидную и, по меньшей мере, одну карбоксильную группу, и предпочтительно выбран из группы, включающей бис-N-сукцинимидильные производные и фотоактивируемые сукцинимидильные производные.
24. Молекулярный комплекс, получаемый в соответствии со способом по любому из пп.21-23, для фиксации специфичного к клеткам маркера с везикулой, имеющей положительно заряженную липидную бислойную мембрану, состоящий из гетеробифункционального перекрестно-сшивающего агента в комбинации с фосфатидилэтаноламином, соединенного посредством его аминогруппы с активным участком перекрестно-сшивающего агента, и специфичного к клеткам маркера, соединенного посредством его тиоловой группы с другим активным участком перекрестно-сшивающего агента, причем гетеробифункциональный перекрестно-сшивающий агент является сукцинимидильным производным, имеющим, по меньшей мере, одну малеимидную и, по меньшей мере, одну карбоксильную группу, и предпочтительно выбран из группы, включающей бис-N-сукцинимидильные производные и фотоактивируемые сукцинимидильные производные, а специфичный к клеткам маркер способен к специфическому обнаружению и связыванию с клетками-мишенями и выбран из группы, включающей антитело, предпочтительно моноклональное антитело, фрагмент антитела, предпочтительно F(ab')2 или Fab'-фрагмент, цитокин и фактор роста.
Текст
1 Область техники, к которой относится изобретение Настоящее изобретение относится к области генной биотехнологии и генной терапии и относится к новым виросомам, т.е. положительно заряженным везикулам, содержащим в мембране вирусные гликопротеины для эффективного переноса генетического материала в местамишени, методам их получения и областям полезного применения. Настоящие катионные виросомы особенно пригодны для специфической и неспецифической, неинфекционной доставки генов в клетки-мишени в условиях in vitro и invivo. Уровень техники Липосомы широко используют для доставки лекарственных препаратов и в качестве защитных оболочек для существующих непродолжительное время фармацевтических соединений или против (био)химической атаки жидкостей организма. Липосомы, содержащие реконструированные мембранные белки или их части из вирусных оболочек, обычно называют виросомами (Sizer et al., Biochemistry, 26: 5106-5113, 1987). Их используют для неспецифической доставки различных лекарственных препаратов и молекул ДНК (Vainstein et al.,Methods Euzymol., 101: 492-512, 1983). Оказалось большим недостатком то, что при слиянии этих виросом с клеточными мембранами клеток-мишеней происходит неконтролируемое высвобождение переносимого вирусной оболочкой, например, белками шиповидных образований, вставленными в мембрану. Обычные методы для включения генетического материала в PC-, РЕ-виросомы страдают недостатком относительно низкой эффективности включения нуклеиновой кислоты. Сущность изобретения Данное изобретение раскрывает способ получения везикулы, имеющей положительно зараженную липидную бислойную мембрану,которая содержит катионные и/или поликатионные липиды вместе с фосфатидилэтаноламином или фосфатидилхолином, по меньшей мере, одним функционально активным гемагглютинином вируса, контролирующим слияние, включенным в указанную мембрану или ковалентно связанным с мембраной, который способен индуцировать и/или способствовать реакции слияния между мембраной везикулы и липидной мембраной клетки-мишени, специфичным клеточным маркером, закрепленным на мембране и необходимым генетическим материалом. Способ включает стадии: а) растворения очищенного вируса гриппа в растворе неионного детергента и выделения фракции, содержащей гемагглютининовый тример; б) приготовления конъюгированных молекулярных комплексов, состоящих из фосфатидилэтаноламина (РЕ), бифункционально пере 003130 2 крестно-сшивающего агента и специфичного к клеткам маркера, таким образом, что бифункциональный перекрестно-сшивающий агент одним активным участком связывается с аминогруппой РЕ и другим - с тиоловой группой специфичного к клеткам маркера, и удаления неконъюгированного материала; в) приготовления раствора, содержащего: забуференный, неионный детергент, гемагглютинин вируса гриппа, полученный на стадии а),катионные и/или поликатионные липиды, фосфатидилэтаноламин (РЕ), необязательно фосфатидилхолин, перекрестно-сшивающий агент и,по меньшей мере, один специфичный к клеткам маркер в составе получаемых на стадии б) молекулярных комплексов РЕ/перекрестносшивающего агента/маркера, и доведение содержания липидов (к общему количеству липидов) до - 90-95% по массе катионных и/или поликатионных - липидов и 5-10% по массе фосфатидилэтаноламина; или 45-90% по массе катионных и/или поликатионных липидов, 5-50% по массе фосфатидилхолина и 5-10% по массе фосфатидилэтаноламина; г) удаления детергента обработкой раствора гранулами-микроносителями, предпочтительно полистирольными гранулами BiobeadsSM-2, что приводит к образованию положительно заряженных липидных бислойных везикул; и д) добавления к везикулам необходимого количества желаемого генетического материала для доставки в клетки-мишени, обработки смеси ультразвуком для включения материала в везикулы и удаления невключившегося материала,предпочтительно гель-фильтрацией. Согласно заявленному способу обеспечивают весовое соотношение гемагглютинина и общего содержания липидов в мембране, равное, примерно, 1 (мг/мг) или меньше. В способе специфичный к клеткам маркер способен к избирательному обнаружению и связыванию с клетками-мишенями и выбран из группы, включающей антитело, предпочтительно моноклональное антитело, фрагмент антитела, предпочтительно фрагмент F(ab')2 или Fab',цитокин и фактор роста. Предпочтительно, что специфичный к клеткам маркер представляет собой фрагментFab', а весовое соотношение гемагглютинина и указанного маркера составляет примерно 2:1. Предпочтительно катионные липиды включают, по меньшей мере, одно соединение,выбранное из группы, состоящей из(DPPES); диоктадециламидоглицилспермина (DOGS); 2,3 диолеоилокси-N-[2-(сперминкарбоксамидо этил]-N,N-диметил-1-пропанаминотрифторацетата (DOSPA); 1,3-диолеоилокси-2-(6-карбоксиспермил)пропиламида (DOSPER); и N,N,N'N'тетраметил-N,N'-бис(2-гидроксиэтил)-2,3-диолеоилокси-1,4-бутандиаммоний йодида (THDOB-). Указанный выше раствор неионного детергента включает C12E8 (монододециловый эфир октаэтиленгликоля) или н-октилолигооксиэтилен в концентрации 10-250 мкмоль на 1 мл HEPES буфера. Указанные выше гранулы-микроносители имеют смачиваемый размер ячеек 20-50 меш.(0,84-0,30 мм), и полученный на стадии в) раствор обрабатывают гранулами-микроносителями до четырех раз. Указанный выше перекрестно-сшивающий агент является гетеробифункциональной органической молекулой, имеющей, по меньшей мере, одну малеимидную и, по меньшей мере,одну карбоксильную группу и, предпочтительно, выбран из группы, включающей бис-Nсукцинимидильные производные и фотоактивируемые сукцинимидильные производные. В способе неконьюгированный материал,особенно не прореагировавшие комплексы РЕ/ перекрестно-сшивающего агента, такие как N[4-(п-малеимидофенил)бутирил]фосфатидилэтаноламин (МРВ.РЕ) или 4-(N-малеимидометил)циклогексан-1-карбоксилатфосфатидилэтаноламин (МСС.РЕ) отделяют от продуктов реакции с помощью гель-хроматографии, предпочтительно аффинной хроматографией на агарозном носителе, предпочтительно на восстановленной тиопропилсефарозе 6 В. Необходимый генетический материал, выбран из группы, включающей короткие цепи ДНК или РНК, дезоксирибонуклеотиды, олигодезоксирибонуклеотиды, олигодезоксирибонуклеотиды селеноатов, олигодезоксирибонуклеотиды фосфоротиоанатов, олигодезоксирибонуклеотиды фосфорамидатов, олигодезоксирибонуклеотиды метилфосфонатов, комплексы пептида с нуклеиновой кислотой, рибонуклеотиды,олигорибонуклеотиды, олигорибонуклеотиды фосфоротиоатов, 2'-ОМе-олигорибонуклеотиды фосфатов, 2'-Оме-олигорибонуклеотиды фосфоротиоатов, рибозимы, гены, плазмиды и векторы. Изобретение раскрывает липидную везикулу, имеющую положительно заряженную липидную бислойную мембрану, которая содержит катионные и/или поликатионные липиды вместе, по меньшей мере, с одним функционально активным гемагглютинином вируса гриппа, включенным в указанную мембрану или ковалентно связанным с мембраной, который способен индуцировать и/или способствовать реакции слияния между мембраной везикулы и 4 липидной мембраной клетки-мишени, характеризующуюся тем, что а) мембрана везикулы содержит (к общему количеству липидов) - 90-95% - по массе катионных и/или поликатионных липидов и 5-10% по массе фосфатидилэтаноламина; или 45-90% по массе катионных и/или поликатионнных липидов, 5-50% по массе фосфатидилхолина и 510% по массе фосфатидилэтаноламина; б) мембрана включает, по меньшей мере,один бифункциональный перекрестно-сшивающий агент, прикрепленный к мембране посредством ковалентного связывания с фосфатидилэтаноламином; в) везикула включает, по меньшей мере,один специфичный к клеткам маркер, способный к избирательному обнаружению и связыванию с клетками-мишенями, выбранный из группы, включающей антитело, фрагмент антитела,цитокин и фактор роста, причем указанный маркер, связан с мембраной с помощью упомянутого перекрестно-сшивающего агента таким образом, что перекрестно-сшивающий агент реагирует со свободной тиоловой группой специфичного к клеткам маркера; и г) везикула содержит необходимый генетический материал для доставки в клеткимишени. Предпочтительно в везикулe катионные липиды включают, по меньшей мере, одно соединение, выбранное из группы, состоящей изN,N,N',N'-тетраметил-N,N'-бис-(2-гидроксиэтил)-2,3-диолеоилокси-1,4-бутандиаммоний йодида (THDOB). Предпочтительно мембрана везикулы включает 1,3-диолеоилокси-2-(6-карбоксиспермил)пропиламид (DOSPER) в качестве поликатионного липида. Необходимый генетический материал,включенный в указанную везикулу, выбран из группы, включающей цепи ДНК или РНК, дезоксирибонуклеотиды, олигодезоксирибонуклеотиды, олигодезоксирибонуклеотиды селеноатов, олигодезоксирибонуклеотиды фосфоротиоатов, олигодезоксирибонуклеотиды фосфороа 5 мидатов, олигодезоксирибонуклеотиды метилфосфонатов, комплексы пептида с нуклеиновой кислотой, рибонуклеотиды, олигорибонуклеотиды, олигорибонуклеотиды фосфоротиоатов,2'-ОМе-олигорибонуклеотиды фосфатов, 2'ОМе-олигорибонуклеотиды фосфоротиоатов,рибозимы, гены, плазмиды и векторы. Предпочтительно диаметр везикулы находится в пределах 120-180 нм. Изобретение также касается применения новой липидной везикулы в качестве системыпереносчика для специфической и неинфекционной доставки необходимого генетического материала в клетки-мишени или ткани-мишени,особенно в покоящиеся или пролиферирующие клетки-мишени млекопитающих, в том числе в целях профилактики и/или терапевтического лечения человека и животных. Применение согласно изобретению предполагает, что генетический материал предпочтительно включает, по меньшей мере, один антисмысловой олигонуклеотид, пригодный для антисмысловой терапии заболевания, выбираемого из группы, включающей рак, лейкемию и вирусную инфекцию при этом: антисмысловой олигонуклеотид направлен на мРНК протоонкогена или онкогена. Далее изобретение раскрывает способ получения молекулярного комплекса, состоящего из фосфатидилэтаноламина (РЕ), гетеробифункционального перекрестно-сшивающего агента и специфичного к клеткам маркера, выбранного из группы, включающей антитело, фрагмент антитела, цитокин и фактор роста, включающий следующие стадии: а) растворение специфического к клеткам маркера, предпочтительно Fab'-фрагмента моноклонального антитела, в буферном растворе,предпочтительно содержащем 100 мМ NaCl, 40 мМ лимонной кислоты, 35 мМ Nа 2 НРO42 Н 2 О, 2 мМ ЭДТА и имеющем рН 5,5; б) смешивание полученного раствора с раствором детергента, который включает 0,5% неионного детергента, предпочтительно ноктилолигооксиэтилена в цитратном буфере с заранее приготовленными структурами, состоящими из перекрестно-сшивающего агента и РЕ, предпочтительноN-Y-малеимидобутирилоксифосфатидилэтаноламина (GMB.PE); в) инкубирование смеси, предпочтительно при 4 С в течение 16 ч в атмосфере азота при мягком перемешивании для конъюгации специфичного к клеткам маркера с перекрестносшивающим агентом, входящим в состав заранее приготовленных структур с образованием молекулярных комплексов и 6 г) удаление после инкубации свободных структур, содержащих перекрестно-сшивающий агент и РЕ, неконьюгированных специфичным к клеткам маркерам с помощью аффинной хроматографии, используя агарозный матрикс, предпочтительно инкубируя смесь в течение 4 ч при комнатной температуре со свежевосстановленной увлажненной тиопропилсефарозой 6 В. Указанный способ предусматривает, что после окончания аффинной хроматографии агарозный матрикс удаляют, предпочтительно,центрифугированием и оставшийся раствор,содержащий молекулярные комплексы РЕ/ перекрестно-сшивающий агент/маркер, нейтрализуют до рН 7,0 и при необходимости добавляют монододециловый эфир октаэтиленгликоля(C12/E8). Далее указанный способ предусматривает,что указанный гетеробифункциональный перекрестно-сшивающий агент является сукцинимидильным производным, имеющим, по меньшей мере, одну малеимидную и, по меньшей мере,одну карбоксильную группу и, предпочтительно, выбран из группы, включающей бис-Nсукцинимидильные производные и фотоактивируемые сукцинимидильные производные. Далее изобретение предусматривает молекулярный комплекс, получаемый в соответствии со способом по любому из пп.21-23, для фиксации специфичного к клеткам маркера с везикулой, имеющей положительно заряженную липидную бислойную мембрану, состоящий из гетеробифункционального перекрестно-сшивающего агента в комбинации с фосфатидилэтаноламином, соединенного посредством его аминогруппы с активным участком перекрестносшивающего агента, и специфичного к клеткам маркера, соединенного посредством его тиоловой группы с другим акивным участком перекрестно-сшивающего агента, причем гетеробифункциональный перекрестно-сшивающий агент является сукцинимидильным производным, имеющим, по меньшей мере, одну малеимидную и, по меньшей мере, одну карбоксильную группу и, предпочтительно, выбран из группы, включающей бис-N-сукцинимидильные производные и фотоактивируемые сукцинимидильные производные, а специфичный к клеткам маркер способен к специфическому обнаружению и связыванию с клеткамимишенями и выбран из группы, включающей антитело, предпочтительно, моноклональное антитело, фрагмент антитела, предпочтительно,F(ab')2 или Fab'-фрагмент, цитокин и фактор роста. Таким образом, настоящее изобретение относится к новой катионной виросоме, которая вследствие специфического состава мембраны,может очень эффективно нагружаться любым желаемым генетическим материалом, включающим длинную и короткую цепь ДНК или РНК, олигодезоксинуклеотиды, рибонуклеоти 7 ды, комплексы пептида с нуклеиновой кислотой(PNA), рибозимы (молекулы РНК с ферментной активностью), гены, плазмиды и векторы, и таким образом, оно убедительно преодолевает недостатки предшествующего уровня. Кроме того, изобретение относится к методу эффективной реконструкции гемагглютинина вируса гриппа А, особенно штамма А/Сингапур, в основном в одноламеллярные катионные липидные везикулы, что приводит к образованию катионных виросом со средним диаметром примерно 120-180 нм и непрерывного липидного бислоя, который в основном свободен от недостатков вытекания, что наблюдается со многими обычными виросомными препаратами. Структура - предпочтительно одноламеллярной катионной бислойной мембраны является таковой, что гидрофильные, положительно заряженные головки липидов ориентированы в водной фазе(ам), а гидрофобные хвосты жирных кислот ориентированы к центру бислоя. С помощью электронной микроскопии можно было показать, что реконструированные вирусные белки шиловидных образований (гемагглютинин) включаются в липидный бислой и выступают за поверхность катионных везикул (фиг. 1). В липидный состав мембран везикул входят катионные и/или поликатионные липиды и необязательно фосфолипиды, такие как фосфатидилэтаноламин и фосфатидилхолин. В большинстве случаев оказалось выигрышным выбрать состав липидов мембраны, включающий по общему содержанию липидов - любой изi) 100% по массе катионных и/или поликатионных липидов; илиii) 90-95% по массе катионных и/или поликатионных липидов и 5-10% по массе фосфатидилэтаноламина; илиiii) 45-90% по массе катионных и/или поликатионных липидов, 5-10% по массе фосфатидилэтаноламина и 5-50% по массе фосфатидилхолина. В предпочтительном варианте осуществления настоящее изобретение также относится к необратимому ковалентному связыванию специфичных к клеткам маркеров с катионными виросомами, включая, но не ограничиваясь таковыми, моноклональные антитела, фрагменты антител, такие как фрагменты F(ab')2 или Fab' цитокины и/или факторы роста, пригодные для избирательного обнаружения и связывания с клетками-мишенями. Они связываются с мембраной везикулы таким образом, что выступают наружу и проявляют в основном полную функциональную активность в отношении рецепторов обнаружения и связывания. Связывание специфичных к клеткам маркеров таких, как фрагменты антител, с предварительно образованными везикулами, как описано, например, у Martin et al. (J.Biol. Chem.,257: 286-288, 1982) приводит к низкому и часто 8 невоспроизводимому связыванию - осложнение,которое налагает значительное ограничение для мишеневой стратегии. Следовательно, согласно предпочтительному варианту осуществления настоящего изобретения, маркеры связываются с предварительно образованными молекулами фосфатидилэтаноламина - перекрестно-сшивающего агента такими, например, как N-[4-(пмалеимидофенилбутирил)]фосфатидилэтаноламин (МРВ.РЕ) в присутствии детергента. Для достижения наилучших возможных результатов оказалось преимущественным осторожно выделять и очищать вирусные гликопротеины перед реконструкцией в целях избежания инактивации под действием протеолитического расщепления или восстановления внутримолекулярных S-S связей. Следовательно,является предпочтительным, чтобы конъюгированные маркеры, например, маркер МРВ.РЕ,отделяли от неконъюгированных молекул фосфатидилэтаноламина - перекрестно-сшивающего агента (например, МРВ.РЕ) аффинной хроматографией на активированном агарозовом матриксе, предпочтительно на восстановленной тиопропилсефарозе 6 В. Аликвоты очищенных конъюгированных маркеров молекулярных комплексов фосфатидилэтаноламина - перекрестно-сшивающего агента - маркера) затем добавляют к раствору детергента, содержащего смесь растворенных мембранных липидов, белков слияния и других желаемых ингридиентов,перед образованием из них катионных виросом. Оказалось выигрышным проводить связывание бифункционального перекрестно-сшивающего агента с фосфолипидом и специфичным к клеткам маркером, в отдельном процессе,предшествующем приготовлению виросом. Этот метод позволяет лучше контролировать и оптимизировать поверхностную плотность виросомных мембран, особенно в отношении числа специфичных к клеткам маркеров, связанных с ней. Более совершенный контроль концентрации белковых молекул, включенных в или связанных с мембраной, является важным, поскольку несбалансированное соотношение содержания пептидов слияния (например, гемагглютинина) и специфичных к клеткам маркеров(например, фрагментов антител Fab') на виросомной мембране может снизить или даже нарушить их селективные свойства и - в экстремальных случаях - может привести к коагуляции и осаждению везикул. Использование фрагментов антител F(ab')2 и Fab' вместо цельных молекул антител в качестве специфичных к клеткам маркеров имеет особое преимущество, поскольку они значительно менее иммуногенны по сравнению с цельным антителом. Кроме того, отсутствие Fc домена устраняет ряд нежелательных биологических и иммунологических активностей, осуществляемых через Fc, таких, например, как активация комплемента классическим путем и 9 острые гуморальные ответные реакции, в конечном итоге приводит - среди прочего - к клиренсу прикрепляемых виросом с поверхности клеток-мишеней посредством взаимодействия между антителом и его Fc рецептором на клетке-мишени. В отличие от известных липосомальных композиций для доставки нуклеиновых кислот,настоящие катионные виросомы обычно не нуждаются в слиянии или дестабилизации клеточной мембраны для вхождения в цитоплазму. Они способны поступать в клетки хозяина с помощью двух механизмов: 1. прикрепления и 2. проникновения. На первой стадии они связываются через пептиды слияния (например, гемагглютинин) и/или специфичные к клеткам маркеры, с клеточными рецепторами, особенно с мембранными гликопротеинами или гликолипидами с концевой сиаловой кислотой и затем очень эффективно включаются посредством эндоцитоза, осуществляемого через рецепторы. В случае виросом, несущих специфичные к клеткам маркеры, например, фрагменты антител, эти маркеры будут дополнительно распознавать антигенные структуры на поверхности клеток-мишеней, что приводит к прикреплению с помощью двух различных механизмов связывания. Таким образом, настоящие специфичные к клеткам виросомы проявляют избирательность в отношении различных типов клеток за счет имеющихся в их мембране специфичных к клеткам маркеров и одновременно высокую способность к проникновению в клетку посредством эндоцитоза за счет вирусного пептида слияния, например, гемагглютинина. Виросомы с Fab' фрагментами, которые распознают связанные с опухолями антигены такие, как TAG 72, СЕА, 17-1 А, СА 19-9, или связанные с лейкемией антигены, такие как CD10 (CALLA= антиген обычной острой лимфоцитарной лейкемии) и CD20, будут избирательно связываться с опухолевыми или лейкемическими клетками,несущими вышеупомянутые антигены на их клеточной поверхности. На второй стадии, поступая в клетки хозяина посредством эндоцитоза, осуществляемого через рецепторы, виросомы включаются в эндосомы. Затем рН внутри эндосом снижается примерно до 5-6, что активирует гемагглютининовый пептид слияния и запускает слияние виросомальной мембраны с эндосомальной мембраной. Реакция слияния мембраны открывает липидную оболочку виросом и высвобождает включенный генетический материал в цитозоль. Этот механизм в значительной мере улучшает возможности для генетического материала достичь ядра, не будучи расщепленным в результате пищеварительного разложения и/или экзоцитоза. Описание изобретения Основной целью настоящего изобретения является получение положительно заряженных 10 липидных везикул, включающих катионные или поликатионные липиды и внутреннее - обычно водное - пространство, и дополнительно включающих, по меньшей мере, один вирусный пептид слияния, включенный или интегрированный в или ковалентно связанный с мембраной везикул. Предпочтительно везикула, кроме того,содержит в мембране, по меньшей мере, один специфичный к клеткам маркер. Дополнительной целью настоящего изобретения является получение везикул, имеющих полную биологическую активность слияния, т.е. имеющих в основном аналогичную активность слияния, как интактный вирус гриппа. Пептид слияния является вирусным гликопротеином, таким как гемагглютинин, или его производным или синтетическим пептидом слияния, способный индуцировать быстрое слияние упомянутых везикул с эндосомальными мембранами клетокмишеней после эндоцитоза. Новые везикулы или виросомы являются особенно пригодными для переноса любого желаемого генетического материала к местам мишеням, особенно к клеткам и тканям животных и человека в условиях in vitro и in vivo. Особое значение придается тому, что новые виросомы не только способны проникать в пролиферирующие, т.е. делящиеся клетки, но также в непролиферирующие, т.е. покоящиеся клетки,это свойство делает возможным широко использовать их в биологических науках, в фармакологии и медицине, как в качестве исследовательского и/или диагностического средства, так лекарственного препарата. При использовании в качестве лекарственного препарата, настоящие виросомы могут быть частью фармацевтической композиции, которая дополнительно включает обычные добавки и фармацевтически приемлемые носители. Предпочтительно, чтобы фармацевтическую композицию готовили в форме инъекционного раствора, но для накожного и системного назначения преимущественными могут быть другие формы приготовления, например, эмульсии, кремы, гели, мази. Следовательно, целью настоящего изобретения также является использование настоящих виросом для получения фармацевтической композиции, пригодной для профилактического и/или терапевтического лечения животных и людей, для которых подобное лечение может принести пользу. Другой целью настоящего изобретения является использование настоящих виросом для получения диагностического набора для применения в условиях in vitro и in vivo. В одном варианте осуществления настоящие везикулы получают способом, включающим эффективную реконструкцию гемагглютинина (ГА) вируса гриппа А. Соответственно,целью настоящего изобретения также является обучение методу приготовления катионных виросом. В предпочтительном варианте осуществления метод дополнительно включает стадию 11 включения генетического материала в катионные липидные везикулы. В основном, метод приготовления включает следующие стадии: 1. Растворение катионных липидов в неионном детергенте, предпочтительно октаэтиленгликоле-моно-н-додецилэфире (OEG, C12E8) ,вместе с предпочтительно очищенными вирусными гликопротеинами из шиповидных образований, генетическим материалом, желаемым для доставки, и необязательно заранее полученными молекулярными комплексами, приготовленными из фосфатидилэтаноламина, перекрестносшивающего агента и специфичного к клеткам маркера; и 2. Образование везикул - предпочтительно повторное - путем удаления детергента с помощью адсорбирующих гранул - микроносителей,предпочтительно полистирольных гранул типаSM-2 Biobeads с предпочтительным размером ячеек (смачиваемым) 20-50 (0,84-0,30 мм). В предпочтительном варианте изобретения приемлемый бифункциональный перекрестносшивающий агент используют для необязательного связывания специфичного к клеткам маркера с мембраной везикул. Специфичный к клеткам маркер, который направлен к клеточному рецептору, ответственному за избирательное связывание виросом с клеткой, соединяется с перекрестно-сшивающим агентом таким образом, что он по-прежнему является полностью биологически активным. Предпочтительно, чтобы перекрестно-сшивающий агент использовался в форме заранее полученного молекулярного комплекса, где перекрестно-сшивающий агент ковалентно соединяется с фосфатидилэтаноламином или как с фосфатидилэтаноламином,так и специфичным к клеткам маркером. Благодаря функционально активным пептидам слияния в настоящих виросомах, инкапсулированный материал высвобождается в цитозоль клеток-мишеней в основном в результате понижения эндосомального рН (как указано выше). Подобное контролируемое высвобождение, с одной стороны, пролонгирует время сохранения доставленного материала внутри клеток-мишеней и, с другой стороны, позволяет избежать нежелательного длительного нахождения виросом внутри эндосом, и таким образом, при этом снижается опасность неспецифической деградации ценных веществ, транспортируемых виросомами. Еще в одном варианте осуществления настоящее изобретение относится к везикулам, где мембранные липиды дополнительно включают фосфатидилхолин и фосфатидилэтаноламин,что дополнительно улучшает возможности специфической конструкции виросом и/или способствует связыванию пептидов слияния и/или специфичных к клеткам маркеров с мембраной. Термин "пептид слияния" относится к пептидам или белкам, способным индуцировать и/или способствовать реакции слияния между 12 мембраной виросом и липидной мембраной клеток-мишеней. Наиболее предпочтительно, если он относится к вирусным гликопротеинам, содержащим пептид слияния, особенно к полному гемагглютининовому тримеру вирусных поверхностных шиповидных образований, его мономеру или одной или обеим гидролизованным субъединицам, гликопептидам НА 1 и НА 2, содержащим функциональный пептид слияния. В другом варианте осуществления настоящего изобретения термин относится к самому чистому пептиду слияния как выделенному из естественных источников, так и синтезированному. В особо предпочтительном варианте осуществления настоящего изобретения эти полипептиды,содержащие пептид слияния, относятся к гемагглютининам вируса гриппа, особенно к одному из субтипов A-H1N1. Синтетические пептиды слияния предпочтительно выбираются из аминокислотных последовательностей, перечисленных ниже в табл. 1, где аминокислоты обозначены соответствующими кодами из одной буквы (смотри также пример 6 и фиг. 2 вWO 92/13525). Термин "перекрестно-сшивающий агент" относится к органической гетеробифункциональной молекуле, способной присоединяться к поверхности везикул, приготовленных по настоящему изобретению, и способной связывать полипептиды. В предпочтительном варианте осуществления настоящего изобретения, эта молекула содержит N-гидроксисукцинимидную группу для связывания с аминогруппой фосфатидилэтаноламина и малеимидную группу для конъюгации с фрагментами моноклональных антител, такие как сукцинимидил 4-(N-малеимидометил)циклогексан-1-карбоксилат, м-малеимидобензоил-N-гидроксисукцинимидэфир,м-малеимидобензоил-N-гидроксисульфосукцинимидэфир, сукцинимидил 4-(п-малеимидофенил)бутират, сульфосукцинимидил 4(п-малеимидофенил)бутират; или он содержит Nгидроксисукцинимидную группу и фотореактивную азидогруппу для связывания с цитокинами, такие как N-гидроксисукцинимидилсуберат (NHS-SA), N-гидроксисукцинимидил-4 азидобензоат (HASAB), N-сукцинимидил-6-(4'азидо-2'-нитрофениламино)гексаноат Предпочтительно,если перекрестносшивающий агент используется в форме заранее приготовленного молекулярного комплекса перекрестно-сшивающего агента и липида, особенно перекрестно-сшивающего агента и фосфатидилэтаноламина или липида плюс перекрестно-сшивающий агент плюс специфичный к клеткам маркер. Термин "специфичный к клеткам" белок или маркер относится к белку, способному связываться с перекрестно-сшивающим агентом или комплексом перекрестно-сшивающего агента - липида соответственно, и мишеней. В предпочтительном варианте осуществления настоящего изобретения эта молекула относится к соединению, специфичному в отношении клеточного рецептора такому, как моноклональное антитело, фрагмент антитела, цитокин или фактор роста. Специфичный к клеткам маркер обеспечивает избирательное обнаружение и связывание с клетками-мишенями и, таким образом, улучшает действие пептида слияния,присутствующего в качестве сопутствующего компонента в виросомальной мембране. Предпочтительные фрагменты антител включают фрагменты F(ab')2 и Fab', в то время как в число специфичных к клеткам маркеров дополнительно входят интерлейкины и другие цитокины,особенно перечисленные ниже в табл. 2. Таблица 2 Цитокины (международные сокращения) 14 Использованный здесь термин "катионный липид" относится к органической молекуле,которая содержит катионный компонент и неполярный "хвост", так называемое от "головки" до "хвоста" амфифильное соединение такое, как(Commun., 203:1622-1628, 1994). Если точно не указано, термин также включает указанные ниже поликатионные липиды. Термин "поликатионный липид" относится к органической молекуле, которая содержит поликатионный компонент и неполярный(DOGS) (Behr et al., Proc. Natl. Acad. USA,86:6982-6986, 1989); 2,3-диолеоилокси-N-[2(сперминкарбоксиамидо)этил]-N,N-диметил-1 пропанаммоний трифторацетат (DOSPA); 1,3 диолеоилокси-2-(6-карбоксиспермил)пропиламид (DOSPER); N,N,N',N'-тетраметил-N,N'бис(2-гидроксиэтил)-2,3-диолеоилокси-1,4-бутандиаммоний йодид (THDOB). Термин "нуклеиновая кислота" или "генетический материал", использованные здесь,включает короткую цепь ДНК или РНК, дезоксирибонуклеотиды, олигодезоксирибонуклеотиды, олигодезоксирибонуклеотиды селеноаты,олигодезоксирибонуклеотиды фосфортиоаты(ORT), олигодезоксирибонуклеотиды фосфорамидаты, олигодезоксирибонуклеотиды метилфосфонаты, комплексы пептида с нуклеиновой кислотой (PNA), рибонуклеотиды, олигорибонуклеотиды, олигорибонуклеотиды фосфортиоаты, 2'-ОМе-олигорибонуклеотиды фосфаты, 2'ОМе-олигорибонуклеотиды фосфортиоаты, рибозимы (молекулы РНК с ферментной активностью), гены, плазмиды и векторы (носители для клонирования). Использованный здесь термин "виросомы" относится - в его самой упрощенной форме - к липосомальным везикулам с бислойной мембраной, включающей катионные липиды и внутреннее, предпочтительно водное, пространство, где мембрана дополнительно включает вирусные белки, особенно вирусные гликопротеины. В предпочтительном олицетворении,вирусные белки включают, по меньшей мере,один контролирующий слияние пептид или белок, имеющий полную биологическую активность слияния, особенно гликопротеины из шиповидных образований и/или нейраминидаза вируса гриппа А (например, А/Сингапур). Следует понимать, что вирусные белки также включают синтетические аминокислотные последовательности, соответствующие или анало 15 гичные пептиду слияния вируса гриппа, как здесь описано. Мембранные липиды включают катионные липиды, указанные выше, но они могут необязательно дополнительно включать другие природные и/или синтетические липиды,предпочтительно фосфолипиды, такие как фосфатидилхолин (PC) и фосфатидилэтаноламин(РЕ). Несмотря на то, что катионные виросомы по настоящему изобретению во многих случаях- особенно для опытов с клеточной культурой invitro - могут успешно использоваться без специфичных к клеткам маркеров в мембране, особенно предпочтительно для применения in vivo,чтобы они дополнительно включали, по меньшей мере, один специфичный к клеткам маркер в мембране, как определено здесь выше. Средний диаметр везикул находится в пределах 120180 нм, что установлено с помощью электронной микроскопии и динамического светового рассеивания. Использованный здесь термин "полная(биологическая) активность слияния" означает то, что виросомы по настоящему изобретению,включающие реконструированные вирусные белки в мембране везикулы, имеют в основном,ту же активность слияния относительно клетокмишеней, как интактный вирус, из которого они обычно реконструированы. Предпочтительно,если контролирующие слияние свойства катионных виросом приближались к интактному вирусу гриппа А. Активность слияния определяется известными методами, особенно о которых сообщают Hoekstra et al. (Biochemistry,23:5675-5681, 1984) или Luscher et al. (Arch. Virol., 130:317-326, 1993). Прежде чем пациенту в лечебных или профилактических целях применяют антисмысловую технологию, необходимо заранее решить ряд технических проблем, особенно относящихся к разработке приемлемой системыносителя. Например, генетический материал такой, как антисмысловые олигонуклеотиды,может быть нестабильным и разрушаться или в других случаях в большей или меньшей степени инактивироваться прежде, чем он достигнет клеток-мишеней, и таким образом может быть необходимым использовать большие количества подобного материала, включенного в обычные катионные липосомы. В результате этих больших количеств возникает вопрос о потенциальной токсичности для человека и животного. При использовании катионных виросом по настоящему изобретению в качестве носителей для генетического материала, эти проблемы можно легко преодолеть и нежелательные побочные эффекты в результате токсичности можно предотвратить или, по меньшей мере, в значительной мере снизить. Этот выигрышный эффект достигается, потому, что настоящие катионные виросомы имеют - по сравнению с липосомами или виросомами, известными ранее 003130 16 значительно более высокую, в 1000-20000 раз,активность и эффективность переноса включенного генетического материала такого, как антисмысловые олигонуклеотиды в клетки-мишени. Как следствие, практически невозможно сравнивать продуктивность обычных виросом или катионных липосом с продуктивностью катионных виросом по настоящему изобретению. Краткое описание чертежей Фиг. 1 показывает микрофотографиюDOTAR-виросом с вирусными белками из шиповидных образований. Фиг. 2 показывает рН-индуцированную активность слияния DOTAP-виросом, меченых октадецил родамином В, с модельными липосомами. Фиг. 3 показывает DOTAP-виросомы с инкапсулированными FITC-OPT, включенными в линейные человечьи малые легочные раковые клетки. Фиг. 4 и 5 показывают необычную эффективность поглощения и трансфекции антисмысловых-L-myc-DOTAP-виросом для линейных человечьих малых легочных раковых клеток. Фиг. 6 показывает инкубацию различных линейных человечьих малых легочных раковых клеток с антисмысловыми- L-mус-виросомами. Фиг. 7 а, 7b показывают эффективность трансфекции pRSVcat - DOTAP-виросом с клетками Jurkat. Фиг. 8 - слияние DOTAP-виросом с фосфолипидными липосомами. Слияние определяли R18 методом при 37 С. Фиг. 9 - включение тимидина в клеткиNCI-H209, обработанные виросомами. 75 мкл виросом, содержащих 200 пикомоль любых из антисмысловых или msc FITC-OPT и 625 мкл свежей среды, содержащей 0,5 мкCi 14 Стимидина, добавили к 5 х 104 клеткам/лунку. Фиг. 10 - дозазависимое ингибирование включения тимидина в клетки NCI-H209, при добавлении виросом, содержащих антисмысловые-L-myc-OPT. Клетки NCI-H209 инкубировали при исходной клеточной концентрации 1 х 105 на лунку и на мл. Значения являются средними величинамисредние отклонения трех опытов. Фиг. 11 - инкубация различных клеточных линий человечьих малых легочных раковых клеток с 75 мкл антисмысловых-L-mус-виросом. Экспрессия онкогена L-myc снизилась в клеточных линиях в указанном порядке Н 82 Н 510 АН 209. Виросомы серии 1 содержали меньшее количество антисмыслового-L-myc, чем в серии 2. Фиг. 12 - трансфекция клеток Sp2 двумя препаратами виросом, полученными разными способами. Фиг. 13 - трансфекция клеток P3/NS1 двумя препаратами виросом, полученными разными способами. 17 Фиг. 14 - трансфекция клеток NIH/3T3 двумя препаратами виросом, полученными разными способами. Фиг. 15, 16, 17 - рост клеток KG1 при обработке смысловыми и антисмысловыми c-mybDOTAP-виросомами. Добавление 25, 50 или 100 мкл раствора виросом, содержащего 18, 36 или 72 пикомоль OPT, соответственно. Значения являются средними величинамистандартными отклонениями трех опытов. Фиг. 18 - рост клеток СЕМ-С 3 при обработке DOTAP-виросомами при добавлении 50 мкл раствора смысловых и антисмысловых cmyb-виросом. Значения являются средними величинамисредние отклонения трех опытов. Для того чтобы лучше понять описанное здесь изобретение ниже приводятся следующие примеры. Примеры представлены только для целей иллюстрации, и ни в каком отношении их не следует рассматривать в качестве ограничения этого изобретения. Пример 1. Приготовление катионных липидных везикул с вирусными гемагглютининовыми тримерами с полной активностью слияния из вируса гриппа, содержащих инкапсулированные антисмысловые L-myc-FITC-(=меченные флуоресцином)-олигодезоксинуклеотиды. Приготовление DOTAP-виросом и DOTAPфосфатидилхолин(PC)-виросом Гемагглютинин (НА) из вируса гриппа штамма А/Сингапур/6/86 выделяли, как описаноUSA, 79:968-972. Кратко, вирус рос в аллантоисной полости куриных яиц, и его очищали дважды ультрацентрифугированием в градиенте сахарозы. Очищенный вирус стабилизировали в буфере, содержащем 7,9 мг/мл NaCl, 4,4 мг/мл цитрата тринатрия. 2 Н 2 О, 2,1 мг/мл морфолинэтансульфокислоты и 1,2 мг/мл N-гидроскиэтилпиперазин-N'-2-этансульфокислоты, рН 7,3. 53 мл суспензии вируса, содержащей 345 мкг НА на мл, осаждали ультрацентрифугированием при 100000 g в течение 10 мин. 7,7 мл забуференного раствора детергента, содержащего 145 мМ NaCl, 2,5 мМ HEPES и 54 мг/мл неионного детергента октаэтиленгликольмонододецилэфира (OEG=C12E8), рН 7,4, добавили к осадку вируса гриппа. Осадок полностью растворяли с помощью обработки ультразвуком в течение 2 мин при комнатной температуре. Раствор подвергли ультрацентрифугированию при 100000 хg в течение 1 ч. Полученный супернатант содержал солюбилизированный НА тример (1,635 мг НА/мл) и следовые количества нейраминидазы. 6 мг DOTAP добавили к 3,7 мл супернатанта (6 мг НА) и растворили. Раствор стерилизовали прохождением через 0,2 мкм фильтр и затем переносили в стеклянный резервуар, содержащий 1,15 г стерильных гранул-микроносителей,предпочтительно Biobeads SM-2. Резервуар встряхивали в течение 1 ч, используя встряхиватель REAX2 производство Heidolph (Kelheim, 003130 18 Германия). При необходимости эту процедуру повторяли до 3 раз с 0,58 мг Biobeads. После этих процедур получали слегка прозрачный раствор DOTAP-виросом. Для получения DOTAP-РС-виросом 3 мгDOTAP и 3 мг PC добавили к супернатанту,содержащему 6 мг НА и растворили. Последующие стадии аналогичны описанным дляDOTAP-виросом. Приготовление виросом с синтетическим пептидом слияния Одна из возможностей приготовления катионных виросом, несущих синтетические пептиды слияния в мембране, включает следующие стадии: 1. Активацию фосфатидилэтаноламинаN-[-малеимидобутирилокси]сукцинимидэфира (GMBS) в реакции РЕ+GMBSMBS-РЕ+N-гидроксисукцинимид,как описано Martin et al., необратимое связывание фрагментов иммуноглобулина с заранее образованными везикулами; J. Biol. Сhеm.,257: 286-288 (1982). 2. Приготовление липидных везикул с активированным РЕ (GMB - РЕ), где 20% фосфатидилхолина, 70% DOTAP и 10% GMB -РЕ растворяют в забуференном растворе детергента,как описано выше, для DOTAP-виросом, включающих НА. Последующие стадии приготовления липидных везикул являются аналогичными,описанным выше для DOTAP-виросом. 3. Соединение синтетического пептида слияния с липидными везикулами, где используется пептид, включающий 20 аминокислот, и имеющий аминокислотную последовательностьG-L-F-E-A-I-A-G-F-I-E-N-G-W-E-G-M-I-D-C и который содержит свободную аминогруппу наN-конце и амидную группу на С-конце. Поскольку аминокислотой на С-конце является цистеин, имеется свободная тиоловая группа,доступная для соединения пептида к GMP-PE в мембране липидных везикул. Для осуществления реакций связывания РЕ-СМВмембрана+HS-Суs-пептидРЕGМВмембрана-S-Суs-пептид Раствор свежеприготовленных липидных везикул в буфере (40 мМ лимонной кислоты, 35 мМ однозамещенного фосфата натрия, 100 мМNaCl и 2 мМ ЭДТА, рН 5,5) смешивают с раствором пептида в том же буфере. Смесь осторожно перемешивают в атмосфере азота в течение ночи при 4 С. Липидные везикулы отделяют от неконъюгированных пептидов гельфильтрацией на колонке с High Load Superdex 200. В качестве альтернативы пептиды слияния можно также соединить с липидными везикулами с помощью заранее приготовленных комплексов РЕ-перекрестно-сшивающий агент 19 пептид, как описано ниже в этом примере для фрагментов Fab'. Включение олигодезоксирибонуклеотидов фосфоротиоатов в DOTAP-виросомы Антисмысловые и смысловые олигодезоксирибонуклеотиды фосфортиоаты (ОРТ) гена Lmyc использовали в качестве примера для демонстрации высокой эффективности трансфекции катионных виросом. 5'-FITC-OPTs синтезировали при помощи фосфорамидитной химии(Mycrosynth GmbH, Balgach, Швейцария). В качестве антисмыслового ОРТ и смыслового ОРТ соответственно использовали пентадекамер (5'FITC-GTAGTCCATGTCCGC-3') и пентадекамер(5'-FITC-GCGGACATGGACTAC-3'). Синтезировали смешанные контрольные (msc) OPT той же нуклеотидной длины, что и антисмысловые и смысловые OPT. 1 мл DOTAP-виросом или DOTAP-PSвиросом добавили к каждому изFITC-OPT растворяли и растворы затем обрабатывали ультразвуком в течение 2 мин при 26 С. Неинкапсулированные FITC-OPT отделяли от виросом гель-фильтрацией на колонке сHigh Load Superdex 200 (Pharmacia, Швеция). Колонку уравновешивали стерильным PBS. Свободные объемные фракции содержащиеFITC-OPT, элюировали PBS и собирали. Концентрацию включенных FITC-OPT в виросомы определяли флуорометрически после того, как виросомы полностью растворяли в 0,1 М NaOH,содержащем 0,1% (объем/объему) Тритона Х 100. Для калибровки шкалы флуоресценции устанавливали на 0 флуоресценцию пустыхDOTAP-виросом, которые растворили в упомянутом растворе детергента. Связывание фрагментов Fab' с виросомами с помощью заранее образованных молекулярных комплексов с фосфатидилэтаноламина бифункционального перекрестно-сшивающего агента 3 мг свежевосстановленных Fab' из мышиных моноклональных анти-СД 10 (анти-CALLA) антител, растворенных в 2,8 мл раствора цитратного буфера (100 мМ NaCl, 40 мМ лимонной кислоты, 35 мМ Na2HPO42 Н 2O, 2 мМ ЭДТА, рН 5,5), добавили к раствору 0,524 мг N-[4-(пмалеимидофенил)бутирил]фосфатидилэтаноламина (МРВ.РЕ) в 215 мкл лимонной кислоты буфера, содержащего 0,5% н-октил-олигооксиэтилена. Смесь затем инкубировали в атмосфере азота в течение 16 ч при 4 С при мягком перемешивании. После инкубации несвязанный МРВ.РЕ удаляли порцией в 400 мкл свежевосстановленной увлажненной тиопропилсефарозы 6 В (Pharmacia, Швеция). Смесь инкубировали в 20 течение 4 ч при комнатной температуре. Тиопропилсефарозу 6 В удаляли центрифугированием и образовавшийся раствор нейтрализовали до рН 7,0. В нейтрализованный раствор добавляли OEG (54 мг/мл). Растворы, приготовленные, как описано выше, добавляли к растворам для приготовления DOTAP-виросом. Молекулы Fab'-МРВ.РЕ вставляли в липидный бислой во время образования виросом. Электронно-микроскопические исследования Микрофотографии DOTAP-виросом подтверждают предпочтительную одноламелярную структуру везикул со средним диаметром примерно 120-180 нм, как определено лазерным рассеиванием света. Хорошо видны НА белковые шиповидные образования вируса гриппа(фиг. 1). Определение активности слияния у DOTAP-виросом Активность слияния настоящих DOTAPвиросом определяли количественным методом,основанным на усилении флуоресценции, описанным Hoekstra et al. 1984, Biochemistry, 23: 5675-5681 и Luscher et al. (1993), Arch. Virol.,130: 317-326. Флуоресцирующий зонд октадецилродамин В хлорид (18) (полученный из Molecular Probes Inc., Eugene, США) вставляли в высокой плотности мембрану DOTAP-виросом при добавлении забуференного раствора OEG(C12E8), содержащего DОТАР и НА к тонкой сухой пленке флуоресцентного зонда с последующим встряхиванием в течении 5-10 мин для растворения зонда, затем продолжали, как описано в "Приготовлении катионных липидных везикул". Наблюдали усиление флуоресценции родамина при инкубации меченных родамином(соотношение DOTAP: липосомальный фосфолипид=1:20). Флуоресценцию измеряли на спектрофлуориметре Perkin Elmer 1000 при 560 и 590 нм соответственно длины волн возбуждения и эмиссии. На фиг. 2 показана рН-индуцированная реакция слияния DOTAP-виросом,выражено в процентах усиление флуоресценции(% FDQ). Поглощение клетками инкапсулированных антисмысловых -L-myc-FITC-OPT Оказалось очень полезным метить ОРТ флуоресцином для изучения механизма клеточного захвата DOTAP-виросом. Линейные человечьи малые легочные раковые клетки (ATCCNCI-H209), которые экспрессируют высокие уровни гена L-myc (Nau et al., 1985, Nature, 318,69-73) выращивали в 2-луночных камерных планшетах для тканевых культур (Nunc, Naperville, IL 60566, США). 50 мкл FITC-OPTвиросом добавили к клеткам. Их инкубировали в течение 5, 15 и 30 мин при 37 С, дважды промывали PBS и затем просматривали под флуоресцентным микроскопом. DOTAP-виросомы с инкапсулированными антисмысловыми FITC 21OPT быстро включались в клетки, как можно видеть на фиг. 3. Исследование биологического эффекта антисмысловых -L-mуc-FITC-ОРТ-DОТАР-виросом методом включения тимидина Линейные человеческие малые легочные раковые клетки (АТСС-NCI-H209 AmericanType Culture Collection, Rockville, США) культивировали в 24-луночных планшетах Costar при начальной концентрации 1 х 105 на лунку и на мл. После инкубации в течение 24 ч среду удаляли и добавляли 625 мкл свежей среды,содержащей 0,5 мСi 14 С-тимидина (приготовленного из [2-14C] тимидина, 52,0 MкCi/ммоль;Amersham, Англия) и 75 мкл DОТАР-виросом,включающих 0,2 нмоль каждого из антисмысловых, смысловых или msc FITC-OPT. Культуры мягко встряхивали при очень медленном встряхивании в течение 1 ч при 37 С и затем переносили в инкубатор. Через 48 ч клеточные суспензии извлекали, переносили в центрифужные пробирки и центрифугировали. Полученные клеточные осадки два раза промывали. Когда клетки невозможно было в достаточной мере диспергировать в однородную клеточную суспензию, их подвергали быстрому воздействию раствора трипсина/ЭДТА. Клеточные осадки растворили в 1,5 мл 0,1 М раствораNaOH/тритон Х-100 (0,1%). 3 мл жидкой сцинтилляционной смеси (готовый белок + Beckman,Fullerton, CA, США), добавили к 1 мл раствора 14 С-радиоактивность определяли на жидкостном сцинтилляционном счетчике(Beckman,Fullerton, CA, США). Фиг. 4 и 5 ясно показывают необычную эффективность поглощения трансфекции антисмыловых -L-mуc-DОТАР-виросом. Фиг. 6 показывает, что клетки, которые не экспрессируют L-myc, не подверглись влиянию и не ингибировались антисмысловыми -L-mуcвиросомами. Кроме того, пустые виросомы не показали каких-либо эффектов на раковые клетки и нормальную клетку. Следовательно, оказалось, что противораковая терапия антисмысловыми ОРТ, инкапсулированными в настоящие виросомы, может иметь высокий потенциал,особенно в результате отсутствия вышеупомянутых недостатков обычных катионных липосом. Пример 2. Приготовление катионных липидных везикул с полной активностью слияния вирусных гемагглютининовых тримеров из вируса гриппа, включающих инкапсулированный вектор pcDNA3 с человеческим геном IL-6, клонированным в полилинкерный сайт (=pcDNA3IL-6). Приготовление DOTAP-виросом и включение рсDNA3-IL-6pcDNA3 (Invitrogen Corporation, San Diego,США) является вектором размером в 5,4 Т.п.н.,предназначенный для высокостабильной и кратковременной экспрессии у хозяев эукариот. НА 22 выделили и очищали, как описано в примере 1. 4 мг DОТАР растворяли в 0,5 мл забуференного раствора детергента, содержащего 145 мМ NaCl,2,5 мМ Hepes и 54 мг/мл OEG (=C12E8), рН 7,4 и добавили к 2 мл супернатанта, содержащего 4 мг НА. К образовавшейся смеси добавили 100 мкг pcDNA3-IL-6 и растворили. Раствор подвергли обработке ультразвуком в течение 30 с.pcDNA-IL-6, в мышиные миеломные клетки Полученный раствор,включающийAmerican Type Culture Collection Rockville,США). Через 48 ч инкубации супернатанты клеточных культур тестировали на человеческийDOTAP-липосом, нагруженных pcDNA-IL-6 В культурах миеломных клеток, подвергшихся трансфекции обычными DOTAPлипосомами (которые лишены вирусных пептидов слияния), содержащих то же количествоpcDNA-IL-6, что и DOTAP-виросомы, IL-6 не был обнаружен. Для того чтобы получить те же результаты по трансфекции, что и с DOTAPвиросомами, нагруженными pcDNA-IL-6, необходимо было увеличить pcDNA-IL-6, включенное в DOTAP-липосомы, в 1000 раз. Пример 3. Приготовление катионных липидных везикул с полной активностью слияния вирусного гемагглютининового тримера из вируса гриппа, содержащих инкапсулированный вектор pRSVcat. Приготовление DOTAP-виросом и включение pRSVcat Экспрессирующий вектор pRSVcat (из АТСС, Rockville, США) содержит ген CAT, который кодирует хлорамфениколацетилтрансферазу (CAT). Фермент катализирует перенос ацетильной группы с ацетил-СоА в 3'-гидроксиположение хлорамфеникола. Векторы CAT являются полезными для мониторинга эффективности трансфекции в целом. 23 Фиг. 7 а и 7b показывают максимальную трансфекцию, достигаемую при добавлении 0,01 мкл DOTAP-виросом. В противоположность вышеупомянутому, добавление 0,01 мклDOTAP-липосом, нагруженных pRSVcat, в клетки Jurkat, при тех же условиях инкубации,не привело к какой-либо заметной активностиCAT. Пример 4. Поглощение виросом клетками. Вхождение виросом в клетки-мишени можно разделить на две отдельные стадии: 1. Прикрепление. 2. Проникновение. Прикрепление вовлекает связывание виросом посредством НА с клеточными рецепторами, которые являются мембранными гликопротеинами или гликолипидами с концевой сиаловой кислотой. В случае специфических виросом с фрагментами Fab' будут дополнительно распознаваться антигенные структуры на поверхности клеток-мишеней, что приводит к прикреплению к клеткам-мишеням двумя различными механизмами связывания. Таким образом специфические виросомы проявляют избирательность в отношении отдельных клеточных типов. Виросомы с фрагментами Fab', которые распознают антигены, связанные с опухолями такие,как TAG72, СЕА, 17-1 А, СА 19-9 или антигены,связанные с лейкемией такие, как CD10 (Calla) иCD20, будут избирательно связываться с опухолевыми или лейкемическими клетками, несущими упомянутые антигены на их клеточной поверхности. Гемагглютининовые гликопротеины осторожно выделяют и очищают. Не имеется инактивации как в результате протеолитического расщепления, так и восстановления их внутримолекулярных дисульфидных связей (-S-S-). В проникновение входит поступление виросом в клетки путем эндоцитоза, опосредованного через рецепторы. Виросомы попадают в эндосомы. Кислое значение рН (5-6) внутри эндосом запускает слияние виросомальной мембраны с эндосомальной мембраной. Слияние осуществляется при помощи вирусного гликопротеина шиповидных образований гемагглютинина (НА). Реакция слияния мембраны в эндосоме освобождает виросомы от их липидной оболочки и обеспечивает поступление инкапсулированных препаратов в цитозоль. Активность слияния этих виросомальных препаратов тестировали с помощью усиления флуоресценции. Виросомы метили флуоресцентным зондом октадецилродамином В (R18) и активность слияния НА прослеживали по усилению флуоресценции в результате проникновения зонда из виросомальной в липосомальную мембранумишень. Фиг. 8 показывает флуоресценцию,наблюдаемую при добавлении DOTAP-виросом,меченых R18, к фосфолипидным липосомам. Начавшая быстро увеличиваться флуоресценция указывает на интактное опосредованное через НА слияние. 24 Зависимое от времени поглощение клетками Поглощение виросом определяли при инкубации клеток с 14 С-мечеными виросомами. Клетки P3/NSI в концентрации 1 х 105/мл инкубировали с 40 мкл виросом при 37 С в течение 5, 10, 15, 20 и 30 мин. После отмывания клетки лизировали и определяли количество 14 Смеченых виросом. Как видно из табл. 3, клеточный захват очень быстрый: в течение первых 5 мин включалось 10% виросом. Более длительные периоды инкубации не усиливали дальнейшего поглощения. 1 мл виросом содержал примерно 1011-1012 виросом, следовательно 400040000 виросом включалось в клетку в течение 5 мин. Таблица 3 Время инкубации, мин Число распадов в минуту 5 6414 10 6832 15 6096 20 6610 30 6626 Пример 5. Антисмысловая стратегия при лечении опухолей. Так называемые "антисмысловые" олигодезоксинуклеотиды (ODN) являются короткими нуклеотидными последовательностями ДНК,синтезированными в качестве реверсивных комплементарных для желаемой мишеневой нуклеотидной последовательности mРНК. Путем образования дуплекса РНК-ДНК трансляция информации предотвращается и облегчается разрушение молекулы под воздействием РНКазы Н. Перенос ODN, направленных на онкоген,кодируемый mРНК, в опухолевые клетки может привести к ингибированию клеточной пролиферации и, в некоторых случаях, гибели клетки. Антисмысловые ODN имеют большой потенциал в качестве терапевтических средств. Многие доклинические испытания на животных, а также клинические исследования на стадиях I-III развития рака показали, что антисмысловые ODN, направленные против онкогенов и вирусных генов, терапевтически активны. Перенос молекул функциональной ДНК в клетки с помощью нагруженных ДНК липосом или обычных виросом не очень эффективен. Следовательно были разработаны виросомы с положительно зараженным бислоем (катионным) для переноса генетического материала. Положительно заряженные липидный бислой взаимодействует с нуклеиновыми кислотами и побуждает их концентрироваться внутри образованных везикул. Антисмысловые -L-myc-виросомы Ген L-myc, впервые открытый в линейных человеческих малых легочных раковых клетках(SCLC), часто амплифицирован и сверхэкспрессирован в SCLC. 5'-FITC-олигодезоксирибонуклетиды фосфортиоаты (ОРТ) были синтези 25 рованы посредством фосфорамидитной химии(Microsynth GmbH, Balgach, Швейцария). В качестве антисмысловых ОРТ и смысловых ОРТ использовали соответственно пентадекамер (5'FITC-GTAGTCCATGTCCGC-3') и пентадекамер(5'-FITC-GCGGACATGGACTAC-3'). Синтезировали смешанную контрольную последовательность (msc) OPT, состоящую из аналогичной нуклеотидной длины, что и антисмысловые и смысловые ОРТ. Антисмысловые ОРТ,имеющие сайт инициации трансляции, действуют путем ингибирования рибосомальной трансляции мишеневой mРНК. АнтисмысловыеL-myc-олигодезоксирибонуклеотиды фосфортиоаты были инкапсулированы в виросомы. Оценивали антипролиферативный эффект виросом, в которые была инкапсулирована L-myc антисмысловая ДНК, в SCLC клеточных линиях Н 209, Н 510 и Н 82. Антисмысловые -L-mycвиросомы добавили к линейным человечьим легочным раковым клеткам. Смысловые -Lmyc-виросомы и msc (смешанные контрольные последовательности) использовали в качестве контролей (фиг. 9). Антисмысловые -L-myc-виросомы были в 20000 раз более активны по сравнению с неинкапсулированными антисмысловыми ОРТ. Как показано на фиг. 10, для индукции аналогичных эффектов необходимо было добавить в клеточные культуры некапсулированных -L-mycантисмысловых ОРТ в концентрациях на уровне микромолей. Следовательно, катионные виросомы значительно более эффективны в доставкеODN, по сравнению с поглощением некапсулированных ODN и, кроме того, более эффективны, чем катионные липосомы. Эффект ингибирования роста под воздействием антисмысловых -L-myc-виросом коррелировал с уровнями экспрессии L-myc в трехSCLC клеточных линиях Н 209, Н 510 и Н 82. На основе фиг. 11 сделан вывод, что те клетки, которые не экспрессируют ген L-myc, не подвергаются воздействию антисмысловых -L-mycвиросом. Пустые катионные виросомы не показали какие-либо или показали только незначительные эффекты на нормальные клетки и раковые клетки. Поскольку ген L-mуc часто амплифицирован и сверхэкспрессирован в SCLC и очень ограничен и экспрессируется на низком уровне в тканях взрослого человека, L-mуc может быть хорошей мишенью для антисмысловой виросомной терапии. Пример 6. Неинфекционный перенос плазмидных векторов для генной терапии: трансфекция векторов для экспрессии в тканях млекопитающих с помощью катионных виросом. В современных подходах к генной терапии рака используются плазмидные векторы для экспрессии пригодных генов-мишеней в человеческих раковых клетках как в условиях ех 26 для мишеневой терапии: чувствительность генов таких, как гены тимидинкиназы вируса простого герпеса (HSV-TK) (Moolten F.L.; CancerRes., 46: 5276-5281, 1986); гены, направленные на иммунную систему для уничтожения раковых клеток, такие как гены цитокина (TepperR.I. et al.; Cell, 57: 503-512, 1989), гены, кодирующие костимуляторные молекулы (TownsendS.E. et al.; Science, 259: 368-370, 1993); гены чужеродной гистосовместимости (Plautz G.E. et al.;Proc. Natl. Acad. Sci. USA, 90: 4645-4649, 1993) и, замена супрессорных генов опухолей дикого типа таких, как р 53 (Chen P.L. et al.; Science,250: 1576-1580, 1990). Вследствие некоторых ограничений в отношении используемых в настоящее время вирусных векторов для генной терапии таких как,например, отсутствие специфичности при переносе генов для опухолевых клеток мишеней, и вследствие безопасности, касающейся возможной индукции вторичных злокачественных новообразований, возможности рекомбинации с образованием вирусов, способных к репликациии, технология неинфекционной доставки генов в условиях in vivo с помощью плазмидных экспрессирующих векторов, нуждается в совершенствовании. Использование раскрытых здесь катионных виросом является обещающей альтернативой для технологии неинфекционного осуществляемого через рецепторы переноса генов. Типичная эффективность трансфекции при использовании имеющихся в продаже липидов находится в пределах 5-50%. Виросомы обеспечивают не только более высокую эффективность трансфекции по сравнению с имеющимися в продаже липосомами, но также включение ДНК в виросомы приводит к более стабильной трансформации клеток. Ген человеческого интерлейкина 6 (IL-6) клонировали в полилинкерный сайт pcDNA3,вектор в 5,4 Т.п.н., предназначенный для высокостабильной и кратковременной экспрессии у эукариотных хозяев. Вектор содержит маркер резистентности к неомицину, экспрессированный из начального промотора SV40 для отбора стабильных трансформантов в присутствииG418. Инкапсулирование вектора проводили 3 различными методами: 1. Диализ: плазмиды инкапсулировали во время образования виросом. Детергент актилРОЕ (от Alexis Corp., Laeufelfingen, Швейцария) удаляли диализом. 2. Biobeads: плазмиды инкапсулировали во время образования виросом. Детергент OEG удаляли с помощью Biobeads. 3. Обработка ультразвуком: плазмиды инкапсулировали с помощью DOTAP и проводили слияние полученных DOTAP-липосом с 27 Для определения количества инкапсулиро 14 ванных плазмид получили С-меченыйpcDNA3-cIL-6-DNA. Метод Количество инкапсуинкапсулирования лированных плазмид 0,02 мкг ДНК на мкл Диализ (1) виросом 0,009 мкг ДНК на мклBiobeads (2) виросом 0,04 мкг ДНК на мкл Обработка ультразвуком виросом Клетки Sp2/0-Ag14 (гибридные несекретирующие, мышь; ID-No:ATCC CRL - 1581; здесь обозначены Sp2), клетки P3/NSI/1-Аg4-1 (несекретирующая миелома, мышь; ID-No:ATCC TIB003130(эмбриональные,контактно-ингибированные,мышь NIH Swiss; ID - No:ATCC CRL -1658) при концентрации клеток 1 х 105 в 1 мл среды трансфектировали препаратами виросом (1)-(3). Через 10 дней после трансфекции определяли количество экспрессированного IL-6 с помощьюELISA (фиг. 12, 13, 14). Все клеточные линии получили от АТСС, 12301 Parklawn Drive,Rockville, Maryland, США. Подвергшиеся трансфекации клетки Sp2 иP3/NS1 дважды отбирали с помощью G418. Через 2 месяца культивирования вновь определяли продукцию IL-6. Значения представлены в табл. 4. Таблица 4 Продукция IL-6 клетками, подвергшимися трансфекции, после повторного отбора с помощью G418 Клеточная Метод Кол-во Общее кол-во IL-6 Общее кол-во IL-6 [пг/106 линия приготовления клеток на мл клетокBiobeads 1,6 х 106 4 мл буфера Обработка 8,0 х 106 1,54 х 106 289 1156 144 ультразвуком 4 мл буфера 5 млBiobeads 1,8 х 106 4,5 мл буфера Обработка 1,37 х 107 2,8 х 106 680 4760 347 ультразвуком 7 мл буфера 5 мл 268 1340 119Sp2 Супернатант Диализ супернатанта 5,2 мл 8 42 4,4Biobeads супернатанта Обработка 4,9 мл 651 3190 233 ультразвуком супернатанта Объем буфера для лизиса, добавленного к клеточным осадкам, подобрали таким образом,что полученное количество клеток составило примерно 2 х 106 на мл. Пример 7. Антисмысловые стратегии для лечения лейкемий. Наиболее распространенной генетической аномалией при лейкемиях человека является транслокация филадельфийской хромосомы(Ph1). Транслокация протоонкогена аbl из хромосомы 9 в кластерный участок точки разрыва(bсr) в хромосоме 22 приводит к образованию гибридных генов bсr-аbl. Протоонкоген аbl в норме кодирует белок с тирозинкиназной активностью, которая усиливается в клетках, не сущих гибридные гены bcr-аbl. Транскриптыbcr-аbl обнаружены у подавляющего большинства пациентов с хронической миелоидной лейкемией (CML) и у пациентов с острой филадельфийской лимфоцитарной лейкемией. Наиболее рациональной терапевтической процедурой при CML четко является мишеневое воздействие на гены bcr-abl. Синтетические ODN,комплементарные соединениюbcr-аblтранскриптов, полученные в результате сплайсинга либо второго, либо третьего экзона генаbcr во второй экзон с-abl, подавляют пролиферацию филадельфийских 1 лейкемических клеток в условиях in vitro и сберегают рост нормальных предшественников костного мозга(Szczylic С. et al.; Science, 253: 562-565, 1991). Однако антисмысловая терапия bcr-abl ограничена пациентами с CML. Другой молекулярной мишенью для антисмысловой терапии является ген myb. Myb, кодирующий продукт протоонкогена с-myb,функционирует как ДНК-связывающий специфический фактор транскрипции. Предпочтительно он экспессируется в гематопоэтических клетках и необходим для пролиферации гематопоэтических клеток. Антисмысловые 18-merODN, направленные на 2-7 кодоны c-myb, сильно ингибируют или полностью прекращают клоногенный рост линейных лейкемических Тклеток (CCRF-CEM), а также в 78% обследованных случаях первичной острой миелоидной лейкемии и в 4 из 5 случаев хронической миелоидной лейкемии (CML) в период злокачественного кризиса (Calabretta В. et al.; Proc. Natl.Acad. Sci. USA, 88: 2351-2355, 1991). Очистку костного мозга используют в качестве компонента при лечении некоторых опухолевых заболеваний, включая острые и хронические лейкемии. В настоящее время костный мозг очищают от лейкемических клеток с помощью различных средств таких, как иммунологические реагенты и хемотерапевтические лекарственные препараты. Виросомы, включающие ODN, направленные против онкогена,который дает преимущественную возможность для злокачественного роста лейкемических клеток, окажутся терапевтически пригодными и что наиболее важно, более избирательными, чем обычные хемотерапевтические средства в уничтожении лейкемических клеток при сохранении нормальных клеточных предшественников. Антисмысловые -с-myb-виросомы Были получены смысловые и антисмысловые ОРТ, соответствующие 2-9 кодонам с-myb. Смысловыми и антисмысловыми последовательностями c-myb соответственно были 5'GCCCGAAGACCCCGGCAC-3' и 5'TGTGCCGGGGTCTTCGGGC-3'. Было проведено инкапсулирование ОРТ в DOTAP-виросомы тем же методом, использованным для L-mycDOTAP-виросом. Воздействие смысловых и антисмысловых c-myb-виросом подвергли клетки линии KG-1 от человека с миелоидной лейкемией и клетки линии СЕМ-С 3 от человека с острой лимфобластной лейкемией. Пролиферация клеток KG-1 зависит от гена протоонкогенаmyb, в то время как клетки СЕМ-С 3 не зависят от продукта гена c-myb. Клетки KG-1 инкубировали с 25, 50 и 100 мкл смысловых и антисмысловых c-myb-виросом, содержащих 18, 36 и 72 пмоль смысловых и антисмысловых ОРТ,соответственно. Количество клеток определяли на 2-й, 3-й и 4-й дни. Добавление 25 мкл смысловых и антисмысловых с-myb-виросом имело лишь незначительный эффект на рост клеток (фиг. 15). Однако добавление 50 мкл (фиг. 16) и 100 мкл (фиг. 30 17) значительно ингибировало рост клеток. Более высокие дозы смысловых с-mуb-виросом также показывали ингибирующие эффекты. Полагают, что эти эффекты не являлись проявлением действия виросомальной мембраны, поскольку клетки СЕМ-С 3 не подверглись воздействию аналогичных препаратов виросом (фиг. 18). Сокращения, использованные в описании 2'-Omе 2'-о Метил Антиген обычной острой лимфобластной лейкемии ХлорамфениколацетилтрансферазаN-[(1,2,3-диолеоилокси)-пропил]-N,N,Nтриметиламмонийхлорид Меченый флуоресцином олигодезоксирибонуклеотид фосфортиоат Генетицин дисульфат (антибиотик G418) Гемагглютинин Интерлейкин-6N-[4-(4-малеимидо)фенилбутирил]фосфатидилэтаноламин (=комплекс перекрестно-сшивающего агента - фосфолипида) Смешанная контрольная последовательность Нейраминидаза н-Октилолигооксиэтилен Олигодезоксинуклеотиды Октаэтиленгликольмонододецилэфир (С 12 Е 8) Олигодезоксирибонуклеотид фосфортиоат(ы) Фосфатидилхолин Фосфатидилэтаноламин Комплекс пептида с нуклеиновой кислотой Малые легочные раковые клетки Обезьяний вирус 40 ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ получения везикулы, имеющей положительно заряженную липидную бислойную мембрану, которая содержит катионные и/или поликатионные липиды вместе с фосфатидилэтаноламином или фосфатидилхолином и,по меньшей мере, одним функционально активным гемагглютинином вируса гриппа, контролирующим слияние, включенным в указанную мембрану или ковалентно связанным с мембраной, который способен индуцировать и/или способствовать реакции слияния между мембраной везикулы и липидной мембраной клеткимишени, специфичным клеточным маркером,закрепленным на мембране, и необходимым генетическим материалом, характеризующийся тем, что он включает стадии а) растворения очищенного вируса гриппа в растворе неионного детергента и выделения фракции, содержащей гемагглютининовый тример; б) приготовления конъюгированных молекулярных комплексов, состоящих из фосфатидилэтаноламина (РЕ), бифункционально перекрестно-сшивающего агента и специфичного к клеткам маркера, таким образом, что бифункциональный перекрестно-сшивающий агент 31 одним активным участком связывается с аминогруппой РЕ и другим - с тиоловой группой специфичного к клеткам маркера, и удаления неконъюгированного материала; в) приготовления раствора, содержащего забуференный, неионный детергент, гемагглютинин вируса гриппа, полученный на стадии а),катионные и/или поликатионные липиды, фосфатидилэтаноламин (РЕ), необязательно фосфатидилхолин, перекрестно-сшивающий агент и,по меньшей мере, один специфичный к клеткам маркер в составе получаемых на стадии б) молекулярных комплексов РЕ/перекрестносшивающего агента/маркера, и доведения содержания липидов (к общему количеству липидов) до 90-95% по массе катионных и/или поликатионных липидов и 5-10% по массе фосфатидилэтаноламина; или 45-90% по массе катионных и/или поликатионных липидов, 5-50% по массе фосфатидилхолина и 5-10% по массе фосфатидилэтаноламина; г) удаления детергента обработкой раствора гранулами-микроносителями, предпочтительно полистирольными гранулами BiobeadsSM-2, что приводит к образованию положительно заряженных липидных бислойных везикул; и д) добавления к везикулам необходимого количества нужного генетического материала для доставки в клетки-мишени, обработки смеси ультразвуком для включения материала в везикулы и удаления невключившегося материала,предпочтительно гель-фильтрацией. 2. Способ по п.1, согласно которому обеспечивают весовое соотношение гемагглютинина и общего содержания липидов в мембране, равное примерно 1 (мг/мг) или меньше. 3. Способ по п.1 или 2, согласно которому специфичный к клеткам маркер способен к избирательному обнаружению и связыванию с клетками-мишенями и выбран из группы, включающей антитело, предпочтительно моноклональное антитело, фрагмент антитела, предпочтительно фрагмент F(ab')2 или Fab', цитокин и фактор роста. 4. Способ по любому из пп.1-3, согласно которому специфичный к клеткам маркер представляет собой фрагмент Fab', а весовое соотношение гемагглютинина и указанного маркера составляет примерно 2:1. 5. Способ по любому из пп.1-4, согласно которому катионные липиды включают, по меньшей мере, одно соединение, выбранное из группы, состоящей изN,N,N'N'-тетраметил-N,N'-бис-(2-гидроксиэтил)-2,3-диолеоилокси-1,4-бутандиаммоний йодида (THDOB-). 6. Способ по любому из пп.1-5, согласно которому указанный раствор неионного детергента включает C12E8 (монододециловый эфир октаэтиленгликоля) или н-октилолигооксиэтилен в концентрации 10-250 мкмоль на 1 млHEPES буфера. 7. Способ по любому из пп.1-6, согласно которому указанные гранулы - микроносители имеют смачиваемый размер ячеек 20-50 меш.(0,84-0,30 мм) и полученный на стадии в) раствор обрабатывают гранулами - микроносителями до четырех раз. 8. Способ по любому из пп.1-7, согласно которому перекрестно-сшивающий агент является гетеробифункциональной органической молекулой, имеющей, по меньшей мере, одну малеимидную и, по меньшей мере, одну карбоксильную группу, и предпочтительно выбран из группы, включающей бис-N-сукцинимидильные производные и фотоактивируемые сукцинимидильные производные. 9. Способ по любому из пп.1-8, согласно которому неконъюгированный материал, особенно не прореагировавшие комплексы РЕ/ перекрестно-сшивающего агента такие, как N-[4(п-малеимидофенил)бутирил]фосфатидилэтаноламин (МРВ.РЕ) или 4-(N-малеимидометил) циклогексан-1-карбоксилатфосфатидилэтаноламин (МСС.РЕ), отделяют от продуктов реакции с помощью гель-хроматографии, предпочтительно аффинной хроматографией на агарозном носителе, предпочтительно на восстановленной тиопропилсефарозе 6 В. 10. Способ по любому из пп.1-9, согласно которому необходимый генетический материал выбран из группы, включающей короткие цепи ДНК или РНК, дезоксирибонуклеотиды, олигодезоксирибонуклеотиды, олигодезоксирибонуклеотиды селеноатов, олигодезоксирибонуклеотиды фосфоротиоатов, олигодезоксирибонуклеотиды фосфорамидатов, олигодезоксирибонуклеотиды метилфосфонатов, комплексы пептида с нуклеиновой кислотой, рибонуклеотиды, олигорибонуклеотиды, олигорибонуклеотиды фосфоротиоатов,2'-ОМе-олигорибонуклеотиды фосфатов, 2'-Оме-олигорибонуклеотиды фосфоротиоатов, рибозимы, гены, плазмиды и векторы. 33 11. Липидная везикула, имеющая положительно заряженную липидную бислойную мембрану, которая содержит катионные и/или поликатионные липиды вместе, по меньшей мере, с одним функционально активным гемагглютинином вируса гриппа, включенным в указанную мембрану или ковалентно связанным с мембраной, который способен индуцировать и/или способствовать реакции слияния между мембраной везикулы и липидной мембраной клеткимишени, характеризующаяся тем, что а) мембрана везикулы содержит (к общему количеству липидов) 90-95% по массе катионных и/или поликатионных липидов и 5-10% по массе фосфатидилэтаноламина; или 45-90% по массе катионных и/или поликатионнных липидов, 5-50% по массе фосфатидилхолина и 5-10% по массе фосфатидилэтаноламина; б) мембрана включает, по меньшей мере,один бифункциональный перекрестно-сшивающий агент, прикрепленный к мембране посредством ковалентного связывания с фосфатидилэтаноламином; в) везикула включает, по меньшей мере,один специфичный к клеткам маркер, способный к избирательному обнаружению и связыванию с клетками-мишенями, выбранный из группы, включающей антитело, фрагмент антитела,цитокин и фактор роста, причем указанный маркер связан с мембраной с помощью упомянутого перекрестно-сшивающего агента таким образом, что перекрестно-сшивающий агент реагирует со свободной тиоловой группой специфичного к клеткам маркера; и г) везикула содержит необходимый генетический материал для доставки в клеткимишени. 12. Везикула по п.11, отличающаяся тем,что катионные липиды включают, по меньшей мере, одно соединение, выбранное из группы,состоящей из 34 13. Везикула по любому из пп.11 или 12,характеризующаяся тем, что мембрана включает 1,3-диолеоилокси-2-(6-карбоксиспермил)пропиламид (DOSPER) в качестве поликатионного липида. 14. Везикула по одному из пп.11-13, характеризующаяся тем, что необходимый генетический материал, включенный в указанную везикулу, выбран из группы, включающей цепи ДНК или РНК, дезоксирибонуклеотиды, олигодезоксирибонуклеотиды, олигодезоксирибонуклеотиды селеноатов, олигодезоксирибонуклеотиды фосфоротиоатов, олигодезоксирибонуклеотиды фосфороамидатов, олигодезоксирибонуклеотиды метилфосфонатов, комплексы пептида с нуклеиновой кислотой, рибонуклеотиды,олигорибонуклеотиды, олигорибонуклеотиды фосфоротиоатов, 2'-ОМе-олигорибонуклеотиды фосфатов, 2'-ОМе-олигорибонуклеотиды фосфоротиоатов, рибозимы, гены, плазмиды и векторы. 15. Везикула по любому из пп.11-14, характеризующаяся тем, что ее диаметр находится в пределах 120-180 нм. 16. Везикула по любому из пп.11-15, получаемая способом по любому из пп.1-10. 17. Применение липидной везикулы по любому из пп.11-16 в качестве системыпереносчика для специфической и неинфекционной доставки необходимого генетического материала в клетки-мишени или ткани-мишени,особенно в покоящиеся или пролиферирующие клетки-мишени млекопитающих. 18. Применение по п.17, согласно которому доставку генетического материала осуществляют в целях профилактики и/или терапевтического лечения человека и животных. 19. Применение по любому из пп.17 или 18, согласно которому генетический материал предпочтительно включает, по меньшей мере,один антисмысловой олигонуклеотид, пригодный для антисмысловой терапии заболевания,выбираемого из группы, включающей рак, лейкемию и вирусную инфекцию. 20. Применение по п.19, согласно которому антисмысловой олигонуклеотид направлен на мРНК протоонкогена или онкогена. 21. Способ получения молекулярного комплекса, состоящего из фосфатидилэтаноламина(РЕ), гетеробифункционального перекрестносшивающего агента и специфичного к клеткам маркера, выбранного из группы, включающей антитело, фрагмент антитела, цитокин и фактор роста, включающий следующие стадии: а) растворение специфического к клеткам маркера, предпочтительно Fab'-фрагмента моноклонального антитела, в буферном растворе,предпочтительно содержащем 100 мМ NaСl, 40 мМ лимонной кислоты, 35 мМ Na2HPO42 Н 2 О, 2 мМ ЭДТА и имеющем рН 5,5; б) смешивание полученного раствора с раствором детергента, который включает 0,5%(GMB.РЕ); в) инкубирование смеси, предпочтительно при 4 С в течение 16 ч в атмосфере азота при мягком перемешивании для конъюгации специфичного к клеткам маркера с перекрестносшивающим агентом, входящим в состав заранее приготовленных структур, с образованием молекулярных комплексов и г) удаление после инкубации свободных структур, содержащих перекрестно-сшивающий агент и РЕ, неконъюгированных специфичным к клеткам маркерoм с помощью аффинной хроматографии, используя агарозный матрикс, предпочтительно инкубируя смесь в течение 4 ч при комнатной температуре со свежевосстановленной увлажненной тиопропилсефарозой 6 В. 22. Способ по п.21, согласно которому после окончания аффинной хроматографии агарозный матрикс удаляют предпочтительно центрифугированием и оставшийся раствор, содержащий молекулярные комплексы РЕ/перекрестно-сшивающий агент/маркер, нейтрализуют до рН 7,0 и при необходимости добавляют монододециловый эфир октаэтиленгликоля (C12E8). 23. Способ по любому из пп.21 или 22, согласно которому указанный гетеробифункцио Фиг. 1 36 нальный перекрестно-сшивающий агент является сукцинимидильным производным, имеющим,по меньшей мере, одну малеимидную и, по меньшей мере, одну карбоксильную группу, и предпочтительно выбран из группы, включающей бис-N-сукцинимидильные производные и фотоактивируемые сукцинимидильные производные. 24. Молекулярный комплекс, получаемый в соответствии со способом по любому из пп.2123, для фиксации специфичного к клеткам маркера с везикулой, имеющей положительно заряженную липидную бислойную мембрану, состоящий из гетеробифункционального перекрестно-сшивающего агента в комбинации с фосфатидилэтаноламином, соединенного посредством его аминогруппы с активным участком перекрестно-сшивающего агента, и специфичного к клеткам маркера, соединенного посредством его тиоловой группы с другим активным участком перекрестно-сшивающего агента, причем гетеробифункциональный перекрестно-сшивающий агент является сукцинимидильным производным, имеющим, по меньшей мере, одну малеимидную и, по меньшей мере, одну карбоксильную группу, и предпочтительно выбран из группы, включающей бис-N-сукцинимидильные производные и фотоактивируемые сукцинимидильные производные, а специфичный к клеткам маркер способен к специфическому обнаружению и связыванию с клетками-мишенями и выбран из группы, включающей антитело,предпочтительно моноклональное антитело,фрагмент антитела, предпочтительно F(ab')2 или Фиг. 3 Линейные человеческие малые легочные paковые клетки Н 209 Антисмысловые-L-myc-ДОТАР-виросомы Клеточные линии человеческих малых легочных раковых клеток Линейные человеческие малые легочные раковые клетки NCI-Н 209Aнтисмысловые-L-myc-ДОТАР-виросомы Клеточные линии человеческих SCLC
МПК / Метки
МПК: A61K 9/127, C12N 15/88
Метки: катионные, качестве, переноса, материала, виросомы, генетического, системы
Код ссылки
<a href="https://eas.patents.su/21-3130-kationnye-virosomy-v-kachestve-sistemy-dlya-perenosa-geneticheskogo-materiala.html" rel="bookmark" title="База патентов Евразийского Союза">Катионные виросомы в качестве системы для переноса генетического материала</a>
Амиды, ингибирующие секрецию аполипопротеина в (аро в ) и/или микросомного протеина переноса триглицеридов (мтр)

Номер патента: 1539
Опубликовано: 23.04.2001
Авторы: Кволлич Джордж Джозеф, Чанг Джордж
МПК: A61P 3/10, C07D 217/04, A61K 31/472...
Метки: микросомного, аполипопротеина, протеина, амиды, триглицеридов, мтр, переноса, ингибирующие, аро, секрецию
Формула / Реферат:
1. Соединение формулы или его стереоизомеры, фармацевтически приемлемые соли или гидраты, где G выбирается из (а) фенила или гетероциклического кольца, где указанное гетероциклическое кольцо содержит в общей сложности от 3 до 14 атомов в кольце, причем указанное гетероциклическое кольцо включает в общей сложности от 1 до 4 гетероатомов в кольце, выбираемых независимо из кислорода, азота и серы, причем отдельные кольца из указанных...
Способ слежения для радиолокационной системы

Номер патента: 517
Опубликовано: 28.10.1999
Авторы: Эванс Робин Джон, Палфорд Грэхэм Уильям, Джерротт Роберт Кейт
МПК: G01S 13/06
Метки: радиолокационной, способ, системы, слежения
Формула / Реферат:
1. Способ слежения для системы эхо-сигналов, заключающийся в том, что формируют множество строб-импульсов для соответствующих мод распространения на основе предсказания состояния цели в течение времени облучения цели и формируют оценки состояния цели в течение указанного времени облучения цели на основе точек измерения цели, попадающих в указанные строб-импульсы. 2. Способ слежения по п.1, отличающийся тем, что получают исходные точки измерения...
Системы обработки контейнеров

Номер патента: 341
Опубликовано: 29.04.1999
Автор: Росс Аллан Симпсон
МПК: B66C 23/36
Метки: системы, контейнеров, обработки
Формула / Реферат:
1. Механизм для погрузки, разгрузки или транспортировки контейнера (7) на и с поддона настила (1), прикрепленного или могущего быть прикрепленным к грузовому автомобилю или трейлеру, причем на каждом конце поддона механизм включает в себя поперечный блок (8), содержащий установленную по центру телескопическую стрелу (12), которая смонтирована на оси поворота (11), отличающийся тем, что нижняя часть (26) стрелы имеет удлинение (28), с которым...
Звукоизлучающие устройства и/или системы

Номер патента: 2176
Опубликовано: 28.02.2002
Авторы: Колломз Мартин, Бэнк Грэхэм, Эзима Генри, Хэррис Нейл
МПК: H04R 1/40
Метки: звукоизлучающие, системы, устройства
Формула / Реферат:
1. Излучатель звука, содержащий зонально распределенную решетку многочисленных акустических преобразователей, имеющих индивидуально снабжаемые энергией средства возбуждения, средство для обеспечения сигналов возбуждения со сдвигами фаз, приводящих к излучениям звука со сдвигами фаз из акустических преобразователей, отличающийся тем, что средство для обеспечения сигналов возбуждения выполняет наложение на указанные сигналы возбуждения...
Способ и устройство для оптимизации имплантации биологического материала

Номер патента: 3119
Опубликовано: 27.02.2003
Автор: Вальдес Гонсалес Рафаэль
МПК: A61F 2/00
Метки: имплантации, материала, биологического, оптимизации, устройство, способ
Формула / Реферат:
1. Устройство для оптимизации имплантации биологического материала, отличающееся тем, что содержит центральное предпочтительно трубчатое тело, закрывающие средства на концах этого пористого тела, поршень и закрывающие элементы корпуса устройства. 2. Устройство для оптимизации имплантации биологического материала по п.1, отличающееся тем, что центральное тело представляет собой пористое тело предпочтительно на основе сетки цилиндрической формы с...
Предыдущий патент: Замещенные трициклические соединения.
Следующий патент: Лиганды, включая антитела, проявляющие реактивность в отношении эндокринных клеток
Случайный патент: Способ определения двигательной активности человека (спортсмена) и устройство для его реализации