Способ диагностики меланомы человека на основании определения уровня экспрессии гена phf10, диагностический набор
Номер патента: 19599
Опубликовано: 30.04.2014
Авторы: Георгиева София Георгиевна, Набирочкина Елена Николаевна, Сошникова Наталия Валерьевна, Бречалов Александр Викторович







Формула / Реферат
1. Способ диагностики меланомы человека на основании определения уровня экспрессии гена PHF10 в клетках пациента, предусматривающий следующие стадии:
а) получение у пациента с подозрением на меланому образцов морфологически опухолевой и нормальной тканей;
б) выделение мРНК из клеток указанных образцов;
в) синтез кДНК на матрице указанной мРНК;
г) проведение количественной ПЦР на матрице полученной кДНК с использованием по меньшей мере двух пар праймеров, специфичных к гену PHF10, и по меньшей мере одной пары контрольных праймеров;
д) интерпретация полученных данных об уровне экспрессии гена PHF10 в клетках пациента таким образом, что сниженный уровень экспрессии гена PHF10 в клетках образца морфологически опухолевой ткани по сравнению с таковым в клетках образца морфологически нормальной ткани свидетельствует о наличии у пациента меланомы.
2. Способ по п.1, в соответствии с которым две пары праймеров, специфичных к гену PHF10, представляют собой PHF-1_for, PHF-1_rev, PHF-2_for и PHF-2_rev, которым соответствуют последовательности SEQ ID NO: 1-4 соответственно.
3. Набор для диагностики меланомы человека по сниженному уровню экспрессии гена PHF10, выполняемой согласно способу по п.1, включающий по меньшей мере две пары праймеров, специфичных к гену PHF10, и по меньшей мере одну пару контрольных праймеров.
4. Набор по п.3, в котором две пары праймеров, специфичных к гену PHF10, представляют собой PHF-1_for, PHF-1_rev, PHF-2_for и PHF-2_rev, которым соответствуют последовательности SEQ ID NO: 1-4 соответственно.
5. Набор по п.3, который дополнительно содержит другие компоненты для проведения диагностики меланомы человека, такие как отличные от праймеров реагенты, необходимые для проведения количественной ПЦР, включающие буфер для ПЦР, дезоксинуклеотидтрифосфаты, флуоресцентный краситель (например, SYBR-GREEN ´100) и/или полимеразу Taq с "горячим стартом".
Текст
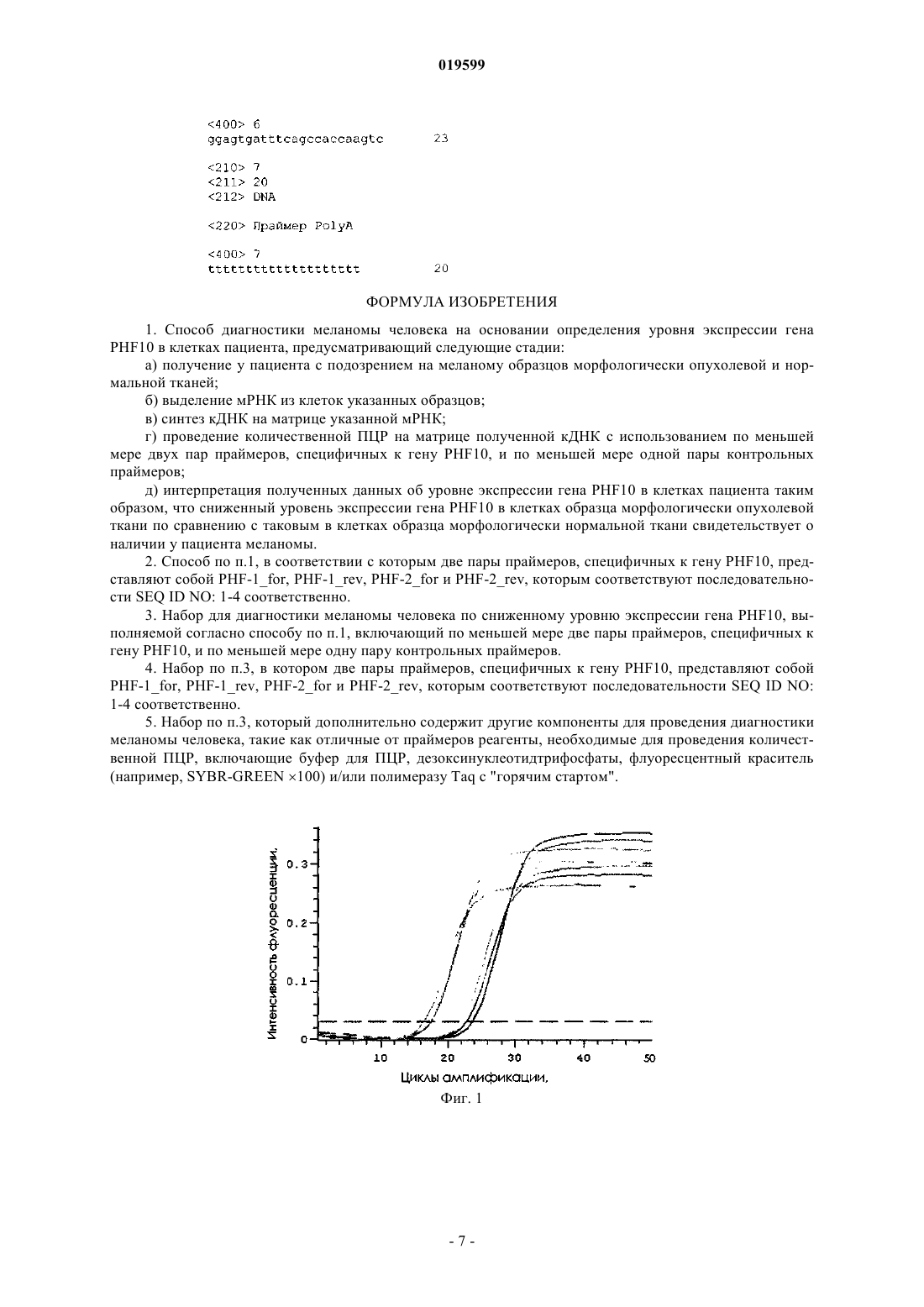
СПОСОБ ДИАГНОСТИКИ МЕЛАНОМЫ ЧЕЛОВЕКА НА ОСНОВАНИИ ОПРЕДЕЛЕНИЯ УРОВНЯ ЭКСПРЕССИИ ГЕНА PHF10, ДИАГНОСТИЧЕСКИЙ НАБОР Изобретение относится к способу диагностики меланомы человека на основании определения уровня экспрессии гена PHF10 в клетках пациента, причем сниженный уровень экспрессии генаPHF10 в клетках образца морфологически опухолевой ткани по сравнению с таковым в клетках образца морфологически нормальной ткани свидетельствует о наличии у пациента меланомы. Также раскрыт набор для проведения диагностики меланомы человека, содержащий по меньшей мере две пары праймеров, специфичных к гену PHF10, и по меньшей мере одну пару контрольных праймеров.(71)(73) Заявитель и патентовладелец: ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ УЧРЕЖДЕНИЕ НАУКИ ИНСТИТУТ БИОЛОГИИ ГЕНА РОССИЙСКОЙ АКАДЕМИИ НАУК; РОССИЙСКАЯ ФЕДЕРАЦИЯ В ЛИЦЕ МИНИСТЕРСТВА ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ (RU) Область техники, к которой относится настоящее изобретение Настоящее изобретение относится к области генной диагностики, в частности к диагностике меланомы с использованием олигонуклеотидов. Предшествующий уровень техники настоящего изобретения Меланома является трудно диагностируемым заболеванием с плохим прогнозом и широким распространением в мире. В настоящее время ведутся активные поиски биологических маркеров меланомы. В одном из исследований сыворотку больных меланомой и здоровых людей исследовали с целью идентификации белковых маркеров меланомы с помощью серологического протеомного подхода. Небольшое количество белков клеток G361, разделенных с помощью двумерного гель-электрофореза и перенесенных на мембрану, инкубировали с сывороткой пациентов. Позитивные сигналы, которые взаимодействовали более чем с 5 образцами сыворотки, идентифицировали с помощью световой массспектрофотометрии. Было обнаружено 12 положительных сигналов и определено 5 белков: эукариотический фактор-2 элонгации (EEF2), энолаза-1 (ENO1), альдолаза (ALDOA), глицеральдегид-3 фосфатдегидрогеназа (GAPDH) и гетерогенный ядерный рибонуклеотид (HNRNP) A2B1. мРНК четырех белков (EEF2, ENO1, ALDOA и HNRNP A2B1) интенсивно экспрессировались в клетках G361 по сравнению с меланоцитами. мРНК EEF2, ENO1 и ALDOA также часто экспрессировались в других культурах клеток меланомы (Suzuki A., Ijzuka A., Komivama M., Kume A., Tai S., Ohshita С., Kurusu A., Nakamura A.,Yamomoto A., Yamazaki N., Ykshikava S., Kivohara Y., Akiyama Y. Identification of Melanoma Antigens Using a Serological Proteome Approach. Cancer Genomics Proteomics. 2010. 7(1):17-23). Возникновение злокачественных опухолей обычно является следствием утраты контроля за интегрированностью генома, что приводит к его злокачественным изменениям. Сравнительная геномная гибридизация (CGH) является методом, который может быть осуществлен на ДНК из фиксированных тканей для определения полного генома на предмет изменения количества копий ДНК. Анализ по методуCGH позволил определить, что меланома отличается от невусов наличием часто повторяющихся хромосомных аберраций (Bauer J, Bastain ВС. Distinguishing melanocytic nevi from melanoma by DNA copy number changes: comparative genomic hybridization as a research and diagnostic tool. Dermatology Therapy. 2006. 19:40-49). С развитием наследственной меланомы ассоциирован ген ингибитора клеточного цикла CDKN2A(p16INK4alpha). Было показано, что в спорадических меланомах происходят изменения уровня экспрессии р 16. Проведены иммуногистохимические исследования с целью изучения ассоциации экспрессии р 16 с клеточной пролиферацией, опухолевой пролиферацией и выживаемостью пациентов. Обнаружено,что для первичных опухолей характерно отсутствие окрашивания белка р 16. Также отсутствие экспрессии р 16 обнаружено и в метастазах. Отсутствие/минимальное окрашивание ядра по р 16 позволяет предсказывать низкую выживаемость пациентов (Straume О., Sviland L., Aksen L.A. Loss of nuclear p16 proteingrowth phase melanoma.Clinical Cancer Research. 2000. 6:1845-1853). Эпигенетический контроль динамики хроматина в процессе синтеза ДНК, репликации и репарации является фундаментальным для правильной последовательности клеточной пролиферации. Фактор-1 сборки хроматина (CAF-1) действует как главный регулятор этого процесса. Как было показано, его субъединица р 60 является новым пролиферативным и прогностическим маркером для нескольких типов опухолей. С помощью иммуногистохимического метода, выполненного на парафиновых срезах первичной меланомы, было показано, что CAF-1/р 60 усиленно экспрессируется во всех изучаемых образцах(130 образцов), что позволяет рассматривать его как новый прогностический маркер для меланомы (Mascolo M., Vecchione M.L., Ilardi G., Scalvenzi M., Molea G., Di Benedetto M., Nugnes L., Siano M., De RosaG., Staibano S. Overexpression of Chromatin Assembly Factor- l/p60 helps to predict the prognosis of melanoma patients. BMC Cancer. 2010. 24:63-69). Полноразмерный MITF-M (ассоциированный с микрофтальмией транскрипционный фактор М) и кДНК его сплайс-варианта были клонированы из клеток меланомы 624mel с помощью ОТ-ПЦР. Его экспрессию исследовали с помощью обычной ПЦР и количественной ОТ-ПЦР в нормальных меланоцитах(NHEM), в клеточных линиях меланомы, в образцах тканей меланомы, в крови здоровых доноров и в клеточных линиях опухолей, отличных от меланомы. Был идентифицирован новый сплайс-вариант, названный MITF-Mdel, который широко экспрессировался в меланоцитах, клеточных линиях и тканях меланомы, но полностью отсутствовал в немеланомных клеточных линиях и в крови здоровых доноров. Обе изоформы значительнее экспрессируются в тканях меланомы по сравнению с другими клеточными линиями (Wang Y., Radfar S., Liu S., Riker A.L., Khong H.T. Mitf-Mdel, a novel melanocyte/melanomaspecific isoform of microphthalmia-associated transcription factor-M, as a candidate biomarker for melanoma.BMC Medicine. 2010. 8:14-20). В одном из исследований проводили флуоресцентную in situ гибридизацию на невусах слизистой оболочки глаза и меланомах. Были использованы зонды к 6 р 25 (RREB1), 6q23 (MYB), 11q13 (CCND1) и центромере 6 (СЕР 6). Хромосомные перестройки были выявлены только в меланомах (Busam K., Fang Ранее белок SOX-9 рассматривали только как регуляторный белок, который играет роль в дифференцировке мезенхимальных клеток в хондроциты. В настоящее время к его функциям относят и регуляцию транскрипции в ходе меланогенеза. Была проанализирована экспрессия SOX-9 на срезах метастазирующей меланомы и проведено сравнение с 3 используемыми маркерами меланоцитов (MART-1, антигеном НМВ-45 и белком S-100). Опухоли, негативные по двум или трем маркерам, были позитивны поSOX-9. Было показано, что SOX-9 экспрессируется практически во всех меланомах (Rao P., Fuller G.N.,Prieto V.G. Expression of SOX-9 in Metastatic Melanoma-A Potential Diagnostic Pitfall. American JournalDermatopathology. 2010. 1: 16-21). Также было показано, что в развитие рака вовлечена аберрантная экспрессия хроматинремодулирующего комплекса SWI/SNF. Обнаружено, что SNF5, основная субъединица комплексаSWI/SNF, участвует в регуляции клеточной дифференцировки, контроле клеточного цикла и апоптоза. С помощью иммуногистохимических методов изучали экспрессию SNF5 в диспластических невусах, первичных меланомах и метастатических меланомах. Оказалось, что экспрессия SNF5 в меланомах значительно редуцирована по сравнению с таковой в невусах. Таким образом, SNF5 является важным маркером развития меланомы (Lin H., Wong R.P., Martinka M., Li G. Loss of SNF5 expression correlates with poorpatient survival in melanoma. Clin. Cancer Research. 2009. 15:6404-6411). Регуляция клеточного цикла имеет большое значение в развитии опухоли. Это послужило основанием для проведения исследования взаимосвязи изменения уровня экспрессии циклина D1, p14, CDK4 иRB с пролиферацией опухолевых клеток и прогрессированием опухоли. Иммуногистохимическим методом было выявлено, что в опухолевых клетках происходит усиление экспрессии этих белков (Bahmannnodular melanomas. NT Journal Oncology. 2004. 25(6): 1559-1565). В одном из исследований с помощью иммуногистохимических методов был изучен профиль экспрессии белков, участвующих в апоптозе, регуляции клеточного цикла, транскрипции и репарации ДНК. Было обнаружено, что белки S-100, HMB45, melan А, специфические циклины (циклин А, циклин D1,циклин D3), белки клеточного цикла RB, pRB, HDM-2, Ki-67, Cyc D3, белок апоптоза сурвивин, транскрипционный фактор STAT-1, белок репарации ДНК MLH-1 характеризуются более высоким уровнем экспрессии в меланомах по сравнению с таковым в невусах, тогда как уровень экспрессии белков клеточного цикла Cyc D1, CDK2, CDK6, р 16, р 27, мембранного рецептора C-KIT, транскрипционных факторов РКСВ, MUM-1 в меланомах ниже (Alonso S., Ortiz P., Pollan M., Perez-Gomez В., Sanchez L., AcunaM., Pajares R., Martinez-Tello F., Hortelano C., Piris M., Rodriguez-Perrato J. Progression in cutaneous malignant melanoma is associated with distinct expression profile. American Journal of Pathology. 2004. 164: 193203). Первичные меланомы исследовали методом микромассивов кДНК. Было показано, что в меланомах изменен уровень экспрессии 174 генов. Среди них гены белков, вовлеченных в регуляцию клеточного цикла (CKS2, CDC2, CCNB1, CENPF, DHFR) и митоз (HCAP-G, STK6), ингибирующих (BIRC5) и стимулирующих (GPR105) апоптоз, регулирующих репликацию ДНК (ТОР 2 А, RRM2, TYMS, PCNA,MCM4, МСМ 6), изменяющих уровень экспрессии в ответ на стресс (GLRX2, DNAJA1, HSPA4, HSPA5,HSPD1, TXNIP), передающих сигнал внутри клетки (STMN2), ингибирующих сигнальный путь Wnt(CTNNBIP1), ингибирующих экспрессию MITF (EMX2), регулирующих протеолиз (TNA), подавляющих метастазирование (NME1) (Winnepenninckx V., Lazar V., Michiels S. et al. Gene expression profiling of primary cutaneous melanoma and clinical outcome. Journal Natl Cancer Inst. 2006. 98:472-482). Методом микромассивов кДНК, Вестерн-блот анализа, количественной ПЦР в режиме реального времени было показано, что в линиях клеток меланомы происходит снижение уровня транскрипции генов, участвующих в развитии, таких как MITF, EDNRD, DCT и TYR, и увеличение уровня транскрипции генов, участвующих во взаимодействии с клеточным окружением, таких как PLAUR, VCAN, HIFa (Jeffsmelanoma cells. PLosOne. 2009. 4(12):e8461). С использованием микромассивов кДНК из образцов рака прямой кишки было показано, что у больных изменена экспрессия 8 генов, кодирующих белки ITGB2, MRPSU, NPR1, TXNL2, PHF10,PRSS8, KCNK3, JAK3. Уровень экспрессии РНК измеряли методом ОТ-ПЦР в режиме реального времени. (Cavalieri D., Dolara P., Mini E., Luceri С., Castagnini С., Toti S., Maciaq K., De Filippo С., Nobili S.,Morganti M., Napoli C., Tonini G., Baccini M., Biggeri A., Tonelli F., Valazano R., Orlando C., Gelmini S.,Cianchi F., Messerini L., Luzzatto L. Analysis of gene expression profiles reveals novel correlations with theclinical course of colorectal cancer. Oncology Research. 2007. 16:535-548). Было показано, что ингибирование пути JAK-STAT приводит к ингибированию роста раковых клеток и к их апоптозу (Bromberg, J., and Darnell, J. E., Jr. The role of STATs in transcriptional control and theirtreatment of leukemias. Oncogene, 2000. 19: 2496-2504). При этом ингибирование пути JAK-STAT в нормальных фибробластах хотя и вызывает прекращение роста клеток, но не вызывает их апоптоз. Одно из объяснений этого эффекта заключается в том, что раковые клетки становятся необратимо связанными с работой пути JAK-STAT для поддержания своего существования, в то время как в нормальной клетке ингибирование пути JAK-STAT просто приводит к ее выходу из митотического цикла. Таким образом,ингибиторы пути JAK-STAT приводят к гибели только раковых клеток, что делает их важным потенциальным средством терапии опухолей. Также были обнаружены селективные ингибиторы тирозинкиназ, что дает возможность воздействовать на первый этап пути JAK-STAT. Было показано, что подавление функции JAK-киназы приводит к гибели клеток миеломы, рака предстательной и молочной желез и т.д. Тем не менее, недостатком ингибирования тирозинкиназ является неспецифичность этого воздействия. Кроме JAK-STAT, оно влияет и на многие другие пути и, таким образом, может негативно влиять и на нормальные клетки. Более специфическим является репрессирование пути JAK-STAT с помощью ингибирования экспрессии белков STAT. Так, было показано, что воздействие на экспрессию STAT с помощью антисмысловых олигонуклеотидов приводит к блокированию роста и жизнеспособности клеток ряда опухолей(Epling-Burnette, P.K., Liu J. H., Catlett-Falcone, et al. Inhibition of STAT3 signaling leads to apoptosis of leukemic large granular lymphocytes and decreased Mcl-1 expression. Journal Clinic Investigations. 2001. 107: 351-362). Ингибирование STAT с помощью белкового ингибитора (доминантно-негативный вариантSTAT) приводит к ингибированию роста опухоли и ее регрессии. Хотя только 15% опухолевых клеток были транформированы доминантно-негативным вариантом STAT, результатом воздействия была массовая гибель клеток меланомы. Новый транскрипционный фактор эукариот PHF10, открытый и охарактеризованный в ИБГ РАНGeorgieva, and Y.V. Shidlovskii. The novel regulator of metazoan development SAYP organizes a nuclear coactivator supercomplex. Cell Cycle. 2009. 8: 2152-2156), является регулятором клеточного цикла и взаимодействует с путем JAK-STAT. Учитывая, что нарушение работы генов пути JAK-STAT в клетках человека связано с развитием различных злокачественных новообразований, в частности меланом, было сделано предположение о том, что вероятность образования меланомы значительно увеличивается при возникновении мутации гена PHF10. Белок PHF10 входит в состав комплекса SWI/SNF, ремоделирующего хроматин. В пролиферирующих нервных стволовых клетках, а также их предшественниках PHF10 в составе этого комплекса регулирует промоторы генов компонентов сигнальных путей, таких как Notch, SHH (J. Lessard, J.I. Wu, J.A.switch in subunit composition of a chromatin remodeling complex during neural development. Neuron. 2007. 55: 201-215). Эти сигнальные пути играют большую роль в развитии организма, отвечают за правильную дифференцировку клеток и тканей. Как только нервная стволовая клетка начинает дифференцироваться,PHF10 в комплексе SWI/SNF заменяется другой субъединицей. Экспрессия PHF10 полностью не прекращается, но в дифференцированных клетках значительно снижена. В большом количестве PHF10 содержится и в других пролиферирующих клетках, например в иммортализованных фибробластах человека. Выключение PHF10 с помощью РНК-интерференции приводило к уменьшению пролиферативной способности этих клеток, что доказывает важную функцию PHF10 в правильном росте клеток (S.S.SV40-immortalized human fibroblast cells. Cytogenetic and genome research. 2009. 126: 227-242). Также было показано, что изменение экспрессии PHF10 коррелирует с ростом опухолевых клеток рака желудка. В частности, уменьшение количества PHF10 приводило к апоптозу этих клеток in vitro и in vivo. После выключения же PHF10 происходило повышение индукции каспазы-3, важнейшего модулятора апоптоза.PHF10 локализуется на промоторе каспазы-3 и ингибирует его, являясь репрессором экспрессии каспазы 3 (М. Wei, В. Liu, J. Li et al. A novel plant homeodomain finger 10 -mediated antiapoptotic mechanism involving repression of caspase-3 in gastric cancer cells. Molecular Cancer Therapeutics. 2010. 9:1764-1774).PHF10 содержит два PHD-домена, которые также называются ассоциированными с лейкозом доменами. Таким образом, изменение экспрессии PHF10 негативно влияет на нормальный рост клеток, а снижение количества PHF10 приводит к разбалансировке и нарушению нормального функционирования клеток. Это позволило авторам настоящего изобретения предположить, что изменение экспрессии PHF10 может служить диагностическим критерием состояния, при котором у пациента имеет место нарушение клеточного деления, в частности злокачественной опухоли, например меланомы. Раскрытие настоящего изобретения Таким образом, настоящее изобретение относится к способу диагностики меланомы человека на основании определения уровня экспрессии гена PHF10 в клетках пациента, предусматривающему следующие стадии: а) получение у пациента с подозрением на меланому образцов морфологически опухолевой и нормальной тканей; б) выделение мРНК из клеток указанных образцов; в) синтез кДНК на матрице указанной мРНК; г) проведение количественной ПЦР на матрице полученной кДНК с использованием по меньшей мере двух пар праймеров, специфичных к гену PHF10, и по меньшей мере одной пары контрольных праймеров; д) интерпретация полученных данных об уровне экспрессии гена PHF10 в клетках пациента таким образом, что сниженный уровень экспрессии гена PHF10 в клетках образца морфологически опухолевой ткани по сравнению с таковым в клетках образца морфологически нормальной ткани свидетельствует о наличии у пациента меланомы. Кроме того, настоящее изобретение относится к набору для диагностики меланомы человека, содержащему по меньшей мере две пары праймеров, специфичных к гену PHF10, и по меньшей мере одну пару контрольных праймеров и необязательно содержащему другие компоненты для проведения диагностики. Другие компоненты для проведения диагностики меланомы человека, которые могут входить в состав набора, например, могут включать в себя отличные от праймеров реагенты, необходимые для проведения количественной ПЦР, такие как буфер для ПЦР, дезоксинуклеотидтрифосфаты, флуоресцентный краситель (например, SYBR-GREEN 100) и полимераза Taq с "горячим стартом". В соответствии с предпочтительным вариантом осуществления настоящего изобретения две пары праймеров, специфичных к гену PHF10, могут представлять собой PHF-1for, PHF-1rev, PHF-2for иPHF-2rev. В соответствии с одним из вариантов осуществления настоящего изобретения пара контрольных праймеров может представлять собой HopMfor и HopMrev. Краткое описание фигур На фиг. 1 приведены кривые амплификации. При обработке результатов использовались данные достижения кривыми порогового значения, установленного на уровне 0,025. Проекция точки пересечения кривых с пунктиром на ось абсцисс в каждом случае соответствует значению c(t). На фиг. 2 проиллюстрированы уровни экспрессии PHF10, измеренные методом количественной ПЦР на праймерах PHF-1. В качестве контроля использовали материал невусов и мышечной ткани. По оси ординат отложены значения уровней экспрессии PHF10 относительно уровня экспрессии нормировочного (контрольного) гена актина в процентах. Из фигуры наглядно видна разница между уровнями экспрессии PHF10 в клетках различных меланом, уровнями экспрессии в клетках доброкачественных невусов и уровнем экспрессии в клетках мышечной ткани. На фиг. 3 проиллюстрированы уровни экспрессии PHF10, измеренные методом количественной ПЦР на праймерах PHF-2. В качестве контроля использовали материал невусов и мышечной ткани. По оси ординат отложены значения уровней экспрессии PHF10 относительно уровня экспрессии нормировочного (контрольного) гена актина в процентах. Из фигуры наглядно видна разница между уровнями экспрессии PHF10 в клетках различных меланом, уровнями экспрессии в клетках доброкачественных невусов и уровнем экспрессии в клетках мышечной ткани. Описание конкретных вариантов осуществления настоящего изобретения Пример 1. Конструирование праймеров для амплификации методом количественной ПЦР. Для определения уровня экспрессии гена человека PHF10 в нормальных и злокачественных клетках были сконструированы олигонуклеотидные последовательности для их последующего использования в амплификации методом количественной ПЦР. Конструирование проводили с помощью программы Vector NTI (Invitrogen, США). В результате для дальнейшего анализа были выбраны пары праймеров, указанные в табл. 1. Таблица 1 Праймеры PHF-1 и PHF-2 отжигаются на разные фрагменты транскрипта PHF10: PHF-1 в районе 720, и длина продукта составляет 228 п.н.; PHF-2 в районе 1100, и длина продукта составляет 233 п.н. Для достоверной оценки количества PHF10 в тканях необходимо проводить ПЦР в режиме реального времени с обеих пар праймеров и результаты обсчитывать относительно нормировочных (контрольных) праймеров (HopMfor, HopMrev) отдельно. В качестве нормировочных (контрольных) были использованы праймеры к транскрипту гена ENY2,экспрессия которого постоянна в разных типах клеток и тканей на разных стадиях развития и клеточного цикла. Пример 2. Проведение амплификации методом количественной ПЦР. Выделение мРНК из культур клеток меланомы, полученных от пациентов III-IV стадий меланомы(примерно из 5 млн. клеток каждой линии: меланома IL, меланома ME, меланома Р, меланома G, меланома KOR) и тканей (объемом 100 мкл мышечной ткани и невусов) производили по следующему протоколу. Образец ткани в керамической ступке заливали жидким азотом и растирали в порошок. Жидкий азот доливали и добавляли 10 объемов буфера GTB. Полученную смесь заливали жидким азотом и растирали в порошок. Лизат переносили в пробирку и расплавляли. Добавляли 0,1 объема раствора ацетата натрия, рН 5,0 (3 М). Компоненты смешивали (и заодно раздробляли ДНК), несколько (3-4) раз пропустив лизат через 19G иглу. К смеси добавляли 1 объем водонасыщенного фенола и перемешивали. Полученную смесь оставляли при комнатной температуре на 5 мин. Затем добавляли 0,2 объема хлороформа и интенсивно перемешивали на вортексе (в течение 3-4 мин). Смесь центрифугировали при 1000 об/мин при комнатной температуре в течение 5-10 мин, а затем при 4000 об/мин при комнатной температуре в течение 10 мин. Водную фазу переносили в пробирку, содержащую 1 объем водонасыщенного фенола, и перемешивали. Смесь центрифугировали при 4000 об/мин при комнатной температуре в течение 10 мин. Затем смесь экстрагировали добавлением хлороформа. Добавляли 1 объем изопропанола, перемешивали и выдерживали при -20 С в течение 1 ч. Смесь центрифугировали при 4000 об/мин при 4 С в течение 10-20 мин. Осадок промывали 75%-ным этанолом, растворяли в не содержащей РНКаз Н 2 О и хранили при -70 С. Затем реакцию синтеза кДНК проводили с использованием выделенной мРНК каждой линии(1 мкг/мкл). Смесь мРНК (1 мкг), праймера polyA 10 мМ (1 мкл) и воды (9 мкл) прогревали в течение 5 мин при 65 С, затем быстро охлаждали до 4 С и добавляли реакционную смесь (общий объем 20 мкл),содержащую 5 буфер M-MuLV RT (4 мкл, Fermentas), ингибитор РНКаз RiboLockRNase Inhibitor (0,5 мкл, Fermentas), обратную транскриптазу RevertAid M-MuLV RT (1 мкл, Fermentas), dNTP 10 мМ (2 мкл),воду, не содержащую РНКаз (0,5 мкл). Синтез кДНК проводили на программируемом амплификаторе фирмы Eppendorf в режиме: 42 С, 90 мин; затем 70 С, 10 мин. Полученную кДНК (разведенную в 5 раз) использовали в качестве матрицы для определения уровня экспрессии PHF10 в линиях клеток методом количественной ПЦР. Реакционная смесь объемом 30 мкл содержала кДНК (3 мкл), 10 буфер для ПЦР (TrisCl, рН 8,6, КС 1 0,5 М, MgCl2 15 мМ, Tween 20 1%) (3 мкл), dNTP 10 мМ (0,6 мкл), праймерfor 10 мкМ (0,9 мкл), праймерrev 10 мкМ (0,9 мкл), флуоресцентный краситель SYBR-GREEN 100 (0,06 мкл), полимераза Taq HotStart (5 ед/мкл, Сибэнзим) (0,15 мкл),воду (21,4 мкл). Каждый образец амплифицировали в двух повторностях с двух пар праймеров для определения количества транскриптов PHF10 (PHF-1forPHF-1rev, PHF-2forPHF-2rev), используя нормировочные (контрольные) праймеры к (HopMforHopMrev). Реакцию проводили на программируемом амплификаторе фирмы Bio-Rad для количественной ПЦР в следующем режиме: плавление 94 С, 2 мин; амплификация 50 циклов с шагами -94 С, 20 с; 63 С, 20 с; 70 С, 20 с; считывание сигнала флуоресценции; заключительный синтез 70 С, 10 мин. Данные повторностей усредняли и количество транскриптов вычисляли по следующей формуле: где c(t)HopM - среднее значение достижения кривыми амлификации порогового цикла продукта с нормировочных (контрольных) праймеров;c(t)PHF - среднее значение достижения кривыми амлификации порогового цикла PHF-1 или PHF-2. Пороговый цикл был установлен на уровне 0,025. Полученные кривые амплификации приведены на фиг. 1. Результаты анализа экспрессии PHF10 представлены на фиг. 2 и 3 и в табл. 2. В нормальной мышечной ткани экспрессия PHF10 находится на достаточно высоком уровне: для праймеров PHF-1 и PHF2 соответственно 93% и 55% относительно экспрессии нормировочного (контрольного) гена. ЭкспрессияPHF10 в невусах снижена примерно в два раза по сравнению с мышечной тканью. Экспрессия же PHF10 в клетках линий меланом снижена по сравнению с экспрессией в невусах в среднем в два-три раза, а по сравнению с мышечной тканью в 5-10 раз. Таким образом, анализ уровня экспрессии PHF10 возможно использовать в качестве одного из способов диагностики меланомы, a PHF10 - в качестве маркера. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ диагностики меланомы человека на основании определения уровня экспрессии генаPHF10 в клетках пациента, предусматривающий следующие стадии: а) получение у пациента с подозрением на меланому образцов морфологически опухолевой и нормальной тканей; б) выделение мРНК из клеток указанных образцов; в) синтез кДНК на матрице указанной мРНК; г) проведение количественной ПЦР на матрице полученной кДНК с использованием по меньшей мере двух пар праймеров, специфичных к гену PHF10, и по меньшей мере одной пары контрольных праймеров; д) интерпретация полученных данных об уровне экспрессии гена PHF10 в клетках пациента таким образом, что сниженный уровень экспрессии гена PHF10 в клетках образца морфологически опухолевой ткани по сравнению с таковым в клетках образца морфологически нормальной ткани свидетельствует о наличии у пациента меланомы. 2. Способ по п.1, в соответствии с которым две пары праймеров, специфичных к гену PHF10, представляют собой PHF-1for, PHF-1rev, PHF-2for и PHF-2rev, которым соответствуют последовательности SEQ ID NO: 1-4 соответственно. 3. Набор для диагностики меланомы человека по сниженному уровню экспрессии гена PHF10, выполняемой согласно способу по п.1, включающий по меньшей мере две пары праймеров, специфичных к гену PHF10, и по меньшей мере одну пару контрольных праймеров. 4. Набор по п.3, в котором две пары праймеров, специфичных к гену PHF10, представляют собойPHF-1for, PHF-1rev, PHF-2for и PHF-2rev, которым соответствуют последовательности SEQ ID NO: 1-4 соответственно. 5. Набор по п.3, который дополнительно содержит другие компоненты для проведения диагностики меланомы человека, такие как отличные от праймеров реагенты, необходимые для проведения количественной ПЦР, включающие буфер для ПЦР, дезоксинуклеотидтрифосфаты, флуоресцентный краситель
МПК / Метки
МПК: G01N 33/50, C12Q 1/68
Метки: основании, человека, диагностический, phf10, экспрессии, определения, гена, способ, диагностики, уровня, меланомы, набор
Код ссылки
<a href="https://eas.patents.su/9-19599-sposob-diagnostiki-melanomy-cheloveka-na-osnovanii-opredeleniya-urovnya-ekspressii-gena-phf10-diagnosticheskijj-nabor.html" rel="bookmark" title="База патентов Евразийского Союза">Способ диагностики меланомы человека на основании определения уровня экспрессии гена phf10, диагностический набор</a>
Способ получения сыворотки для диагностики сибирской язвы и диагностический набор

Номер патента: 17617
Опубликовано: 30.01.2013
Авторы: Захарченко Олег Сергеевич, Соболев Виктор Васильевич
МПК: G01N 33/531, A61K 35/16, A61B 10/00...
Метки: диагностический, сыворотки, диагностики, набор, язвы, получения, сибирской, способ
Формула / Реферат:
1. Способ получения сыворотки для диагностики сибирской язвы путем гипериммунизации продуцентов антигеном из штамма Bacillus anthracis в возрастающих дозах с последующим взятием крови у продуцента, отделением сыворотки и выделением целевого продукта, отличающийся тем, что для гипериммунизации используют волов и антиген из штамма Bacillus anthracis M-71, причем гипериммунизацию проводят следующим образом: первоначально антиген вводят подкожно в...
Выделенное антитело и его фрагмент, способы и набор для оценки уровня перекисного окисления липидов в биологическом образце и способ диагностики заболеваний, обусловленных перекисным окислением липидов

Номер патента: 2520
Опубликовано: 27.06.2002
Авторы: Партхасаратхи Сампатх, Александер Уэйн Р., Медфорд Рассел М.
МПК: G01N 33/543, C07K 16/00, A61P 43/00...
Метки: окислением, способы, липидов, заболеваний, окисления, диагностики, способ, обусловленных, выделенное, фрагмент, уровня, биологическом, набор, перекисным, образце, перекисного, оценки, антитело
Формула / Реферат:
1. Выделенное антитело, которое взаимодействует с антигеном, образовавшимся в результате реакции гидроперекиси липида с первичным амином, и при этом в какой-либо значительной степени не связывается с альдегидмодифицированным белком. 2. Антитело по п.1, отличающееся тем, что конъюгировано с дополнительным агентом. 3. Антитело по п.2, отличающееся тем, что дополнительный агент представляет собой детектируемую метку. 4. Антитело по п.3,...
Днк, кодирующая белок-7 морфогенеза кости человека, содержащая промоторную область гена, кодирующего данный белок, и способ ее получения

Номер патента: 4509
Опубликовано: 29.04.2004
Авторы: Каваи Синдзи, Сугиура Такеюки
МПК: A61P 19/00, C12N 15/85, C12Q 1/68...
Метки: данный, кодирующего, промоторную, белок, способ, белок-7, человека, морфогенеза, содержащая, кодирующая, получения, гена, кости, область, днк
Формула / Реферат:
1. ДНК, содержащая нуклеотиды 1-10877 SEQ ID NO:1, приведенной в списке последовательностей, кодирующая белок-7 морфогенеза кости (ВМР-7) человека и содержащая промоторную область гена, кодирующего указанный белок. 2. Способ получения ДНК, охарактеризованной в п.1, предусматривающий следующие стадии: (1) расщепление геномной ДНК клеток плаценты человека рестриктазой HindIII, (2) выделение фрагмента ДНК размером приблизительно 11 т.п.н. с помощью...
Способ диагностики немелкоклеточного рака легких и набор для его осуществления

Номер патента: 10571
Опубликовано: 30.10.2008
Авторы: Зиновьева Марина Валерьевна, Зборовская Ирина Борисовна, Машкова Тамара Дмитриевна, Зиновьева Ольга Леонидовна, Виноградова Татьяна Викторовна, Копанцев Евгений Павлович, Опарина Нина Юрьевна
МПК: C12N 15/11, C12Q 1/68
Метки: легких, способ, осуществления, немелкоклеточного, рака, диагностики, набор
Формула / Реферат:
1. Способ диагностики немелкоклеточного рака легких, включающий следующие стадии: а) получение исходной пары образцов ткани от пациента, где один из образцов получен из предположительно пораженной раком ткани, а второй получен из прилегающей гистологически нормальной (условно нормальной) ткани; б) выделение и очистка препаратов РНК из исходной пары образцов; в) синтез одноцепочечной или двуцепочечной кДНК на матрице РНК с использованием...
Способ одновременного выявления и идентификации антибиотиков различных классов in vitro и соответствующий диагностический набор

Номер патента: 11656
Опубликовано: 28.04.2009
Автор: Гранье Бенуа
МПК: G01N 33/558, G01N 33/94
Метки: соответствующий, одновременного, способ, диагностический, набор, различных, vitro, антибиотиков, классов, идентификации, выявления
Формула / Реферат:
1. Диагностический набор для одновременного определения антибиотиков различных классов, по меньшей мере, b-лактамов и тетрациклинов, отличающийся тем, что он включает во-первых, единственную реакционную смесь, включающую, по меньшей мере, первый рецептор, специфически распознающий b-лактамы, второй рецептор, специфическим и конкурентным образом распознающий тетрациклин и биотинилированный фрагмент нуклеиновой кислоты, причем оба рецептора...
Предыдущий патент: Способ изготовления роторных и статорных пластин
Следующий патент: Воздухораспределитель пневматического тормоза рельсового транспортного средства
Случайный патент: Устройство для тренировки мышц