Растворимый рецептор т- клетки





























Формула / Реферат
1. Растворимый T-лимфоцитный рецептор (soluble T cell receptor, sTCR), который содержит (I) всю a-цепь TCR или ее часть, за исключением ее трансмембранного домена, и (II) всю b-цепь TCR или ее часть, за исключением ее трансмембранного домена, при этом каждый из компонентов (I) и (II) содержит функциональный вариабельный домен и по меньшей мере часть константного домена цепи TCR, и при этом компоненты (I) и (II) соединены дисульфидной связью, расположенной между остатками цистеина, на которые заменены

2. sTCR по п.1, отличающийся тем, что один или оба компонента (I) и (II) содержат весь внеклеточный константный домен Ig цепи TCR.
3. sTCR по п.1 или 2, отличающийся тем, что один или оба компонента (I) и (II) содержат весь внеклеточный домен цепи TCR.
4. Растворимый T-лимфоцитный рецептор ab-формы (sTCR), отличающийся тем, что ковалентная дисульфидная связь соединяет остатки цистеина, на которые заменены

5. Растворимый T-лимфоцитный рецептор (soluble T cell receptor, sTCR), который содержит (I) всю a-цепь TCR или ее часть, за исключением ее трансмембранного домена, и (II) всю b-цепь TCR или ее часть, за исключением ее трансмембранного домена, причем каждый из компонентов (I) и (II) содержит функциональный вариабельный домен и по меньшей мере часть константного домена цепи TCR, и при этом они соединены дисульфидной связью, расположенной между остатками аминокислот константных доменов, которая отсутствует в нативном TCR, причем нативный TCR не содержит межцепочечную дисульфидную связь.
6. sTCR по п.5, отличающийся тем, что один или оба компонента (I) и (II) содержат весь внеклеточный константный домен Ig цепи TCR.
7. sTCR по п.5 или 6, отличающийся тем, что один или оба компонента (I) и (II) содержат весь внеклеточный домен цепи TCR.
8. Растворимый T-лимфоцитный рецептор ab-формы (sTCR), отличающийся тем, что ковалентная дисульфидная связь соединяет остаток аминокислоты в иммуноглобулиновой области константного домена a-цепи с остатком иммуноглобулиновой области константного домена b-цепи, причем нативный TCR не содержит межцепочечную дисульфидную связь.
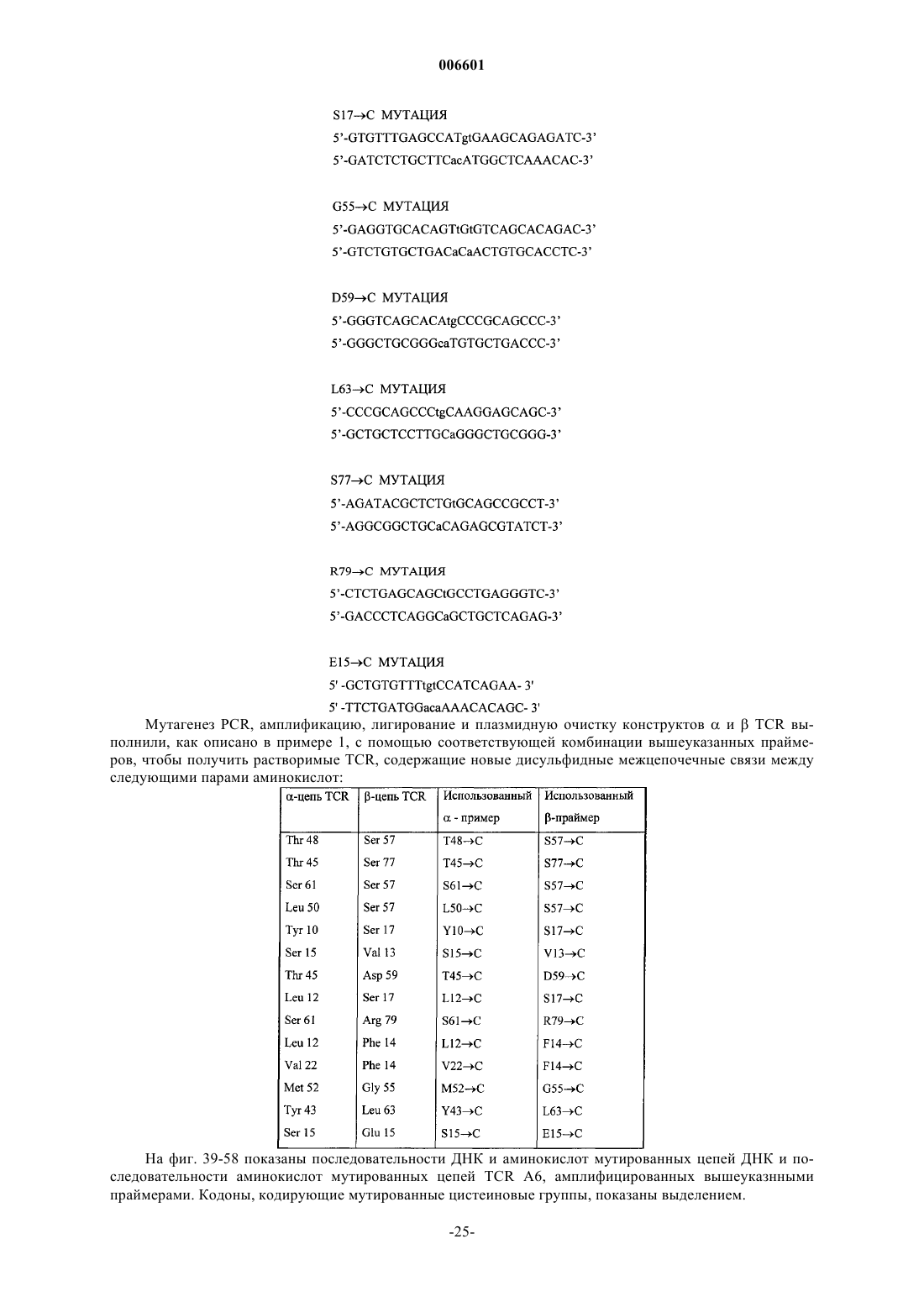
9. sTCR по пп.5-8, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, на которые заменены остатки аминокислот, расстояние между b-атомами углерода которых составляет менее 0,6 нм в нативной структуре TCR.
10. sTCR по пп.5-9, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Thr 48 экзона 1 TRAC*01 и Ser 57 экзона 1 TRBC1*01 или TRBC2*01.
11. sTCR по пп.5-9, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Thr 45 экзона 1 TRAC*01 и Ser 77 экзона 1 TRBC1*01 или TRBC2*01.
12. sTCR по пп.5-9, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Tyr 10 экзона 1 TRAC*01 и Ser 17 экзона 1 TRBC1*01 или TRBC2-01.
13. sTCR по пп.5-9, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Thr 45 экзона 1 TRAC*01 и Asp 59 экзона 1 TRBC1*01 или TRBC2-01.
14. sTCR по пп.5-9, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Ser 15 экзона 1 TRAC*01 и Glu 15 экзона 1 TRBC1*01 или TRBC2*01.
15. sTCR по пп.1-4, отличающийся тем, что нативный TCR не содержит межцепочечную дисульфидную связь.
16. sTCR по пп.5-15, отличающийся тем, что нативные a- и b- цепи TCR усечены в C-концевой области таким образом, что остатки цистеина, которые образуют нативную межцепочечную дисульфидную связь, исключаются.
17. sTCR по пп.5-15, отличающийся тем, что остатки цистеина, которые образуют нативную межцепочечную дисульфидную связь, замещены на остаток другой аминокислоты.
18. sTCR по п.17, отличающийся тем, что остатки цистеина, которые образуют нативную межцепочечную дисульфидную связь, замещены на серин или аланин.
19. sTCR по пп.1-18, отличающийся тем, что неспаренный остаток цистеина, присутствующий в нативной b-цепи TCR, отсутствует.
20. sTCR по пп.1, 2, 5, 6 и 9-19, отличающийся тем, что каждый из компонентов (I) и (II) содержит функциональный вариабельный домен первого TCR, слитый с полным константным доменом или его частью второго TCR, при этом первый и второй TCR происходят от одного и того же вида (животных).
21. sTCR по п.20, отличающийся тем, что константные домены второго TCR усечены в направлении N-конца от остатков, которые образуют ненативную межцепочечную дисульфидную связь.
22. sTCR по пп.1-21, отличающийся тем, что одна или обе цепи дериватизированы какой-либо группой в их C- или N-концевой области или слиты с указанной группой.
23. sTCR по пп.1-22, отличающийся тем, что одна или обе цепи содержат остаток цистеина в их C- или N-концевой области, с которым может быть слита какая-либо группа.
24. sTCR по пп.1-23, отличающийся тем, что содержит также обнаруживаемую метку.
25. sTCR по пп.1-24, отличающийся тем, что связан с терапевтическим агентом.
26. Мультивалентный комплекс T-лимфоцитного рецептора (T cell receptor, TCR), содержащий множество sTCR по пп.1-25.
27. Комплекс по п.26, отличающийся тем, что содержит мультимер sTCR.
28. Комплекс по п.27, отличающийся тем, что содержит две, три, четыре или более молекул T-лимфоцитного рецептора, связанные одна с другой предпочтительно посредством молекулы линкера.
29. Комплекс по пп.26, 27 или 28, отличающийся тем, что растворимые T-лимофцитные рецеторы или их мультимеры находятся в липидном двойном слое (бислое) или присоединены к какой-либо частице.
30. Способ обнаружения комплексов МНС-пептид, включающий
(1) получение растворимого TCR по пп.1-25 или мультивалентного комплекса T-лимфоцитного рецептора по пп.26-29,
(2) контактирование растворимого TCR или мультивалентного комплекса TCR с комплексами МНС-пептид и
(3) детектирование связывания растворимого TCR или мультивалентного комплекса TCR с комплексами МНС-пептид.
31. Фармацевтический состав, содержащий sTCR по пп.1-25 и/или мультивалентный комплекс TCR по пп.26-29 совместно с фармацевтически допустимым носителем.
32. Молекула нуклеиновой кислоты, содержащая последовательность, кодирующую компоненты (I) или (II) sTCR по пп.1-25, или последовательность, комплементарную к ней.
33. Вектор, содержащий молекулу нуклеиновой кислоты по п.32.
34. Клетка-хозяин, содержащая вектор по п.33.
35. Способ получения компонентов (I) или (II) по пп.1-25, включающий инкубацию клетки-хозяина по п.34 в условиях, вызывающих экспрессию пептида, и последующую очистку полипептида.
36. Способ по п.35, отличающийся тем, что включает также смешивание компонентов (I) и (II) в условиях, пригодных для повторной укладки.
37. Способ получения растворимого T-лимфоцитного рецептора (sTCR), включающий
инкубацию клетки-хозяина, которая содержит вектор, содержащий молекулу нуклеиновой кислоты, кодирующей (I) всю a-цепь TCR или ее часть за исключением ее трансмембранного домена, и клетки-хозяина, которая содержит вектор, содержащий молекулу нуклеиновой кислоты, кодирующей (II) всю b-цепь TCR или ее часть за исключением ее трансмембранного домена, при условиях, вызывающих экспресёшю компонентов (I) и (II), при этом каждый из компонентов (I) и (II) содержит функциональный вариабельный домен и по меньшей мере часть константного домена цепи TCR,
очистку компонентов (I) и (II) и
смешивание компонентов (I) и (II) при условиях для повторной укладки, таких что они оказываются соединенными дисульфидной связью, которая расположена между остатками аминокислот константного домена и которая отсутствует в нативном TCR.
38. Способ по п.37, отличающийся тем, что один или оба компонента (I) и (II) содержат весь внеклеточный константный домен Ig цепи TCR.
39. Способ по п.37 или 38, отличающийся тем, что один или оба компонента (I) и (II) содержат весь внеклеточный домен цепи TCR.
40. Способ получения ab-формы растворимого T-лимфоцитного рецептора (sTCR), включающий
инкубацию клетки-хозяина, которая содержит вектор, содержащий молекулу нуклеиновой кислоты, кодирующей a-цепь TCR, и клетки-хозяина, которая содержит вектор, содержащий молекулу нуклеиновой кислоты, кодирующей b-цепь TCR, при условиях, вызывающих экспрессию соответствующих цепей TCR,
очистку соответствующих цепей TCR и
смешивание соответствующих цепей TCR при условиях повторной укладки таким образом, чтобы ковалентная дисульфидная связь соединяла остаток аминокислоты в иммуноглобулиновой области константного домена a-цепи с остатком в иммуноглобулиновой области константного домена b-цепи.
41. Способ по пп.37-40, отличающийся тем, что нативный TCR не содержит межцепочечную дисульфидную связь.
42. Способ по п.41, отличающийся тем, что нативные a- и b- цепи TCR усечены в C-концевой области таким образом, что остатки цистеина, которые образуют нативную межцепочечную дисульфидную связь, исключаются.
43. Способ по п.41, отличающийся тем, что остатки цистеина, которые образуют нативную межцепочечную дисульфидную связь, замещены на остаток другой аминокислоты.
44. Способ по п.43, отличающийся тем, что остатки цистеина, которые образуют нативную межцепочечную дисульфидную связь, замещены на серин или аланин.
45. Способ по пп.37-44, отличающийся тем, что неспаренный остаток цистеина, присутствующий в нативной b-цепи TCR, отсутствует.
46. Способ по пп.37-45, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, на которые заменены остатки аминокислот, расстояние между b-атомами углерода которых составляет менее 0,6 нм в нативной структуре TCR.
47. Способ по пп.37-46, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Thr 48 экзона 1 TRAC*01 и Ser 57 экзона 1 TRBC1*01 или TRBC2*01.
48. Способ по пп.37-46, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Thr 45 экзона 1 TRAC*01 и Ser 77 экзона 1 TRBC1*01 или TRBC2*01.
49. Способ по пп.37-46, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Tyr 10 экзона 1 TRAC*01 и Ser 17 экзона 1 TRBC1*01 или TRBC2-01.
50. Способ по пп.37-46, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Thr 45 экзона 1 TRAC*01 и Asp 59 экзона 1 TRBC1*01 или TRBC2*01.
51. Способ по пп.37-46, отличающийся тем, что дисульфидная связь, отсутствующая в нативном TCR, расположена между остатками цистеина, которые заменяют остатки Ser 15 экзона 1 TRAC*01 и Glu 15 экзона 1 TRBC1*01 или TRBC2*01.
52. Способ по пп.37, 38 и 41-51, отличающийся тем, что каждый из компонентов (I) и (II) содержит функциональный вариабельный домен первого TCR, слитый с полным константным доменом или его частью второго TCR, при этом первый и второй TCR происходят от одного и того же вида (животных).
53. Способ по п.52, отличающийся тем, что константные домены второго TCR усечены в направлении N-конца от остатков, которые образуют ненативную межцепочечную дисульфидную связь.
54. Способ по пп.37-53, отличающийся тем, что одна или обе цепи дериватизированы какой-либо группой в их C- или N-концевой области или слиты с указанной группой.
55. Способ по пп.37-54, отличающийся тем, что одна или обе цепи содержат остаток цистеина в их C- или N-концевой области, с которым может быть слита какая-либо группа.
56. Способ по пп.37-55, отличающийся тем, что sTCR содержит также обнаруживаемую метку.
57. Способ по пп.37-56, отличающийся тем, что sTCR связан с терапевтическим агентом.
58. Способ по любому из пп.30, 35-57, отличающийся тем, что включает также комбинирование множества sTCR для образования мультивалентного комплекса T-лимфоцитного рецептора (TCR).
59. Способ по п.58, отличающийся тем, что sTCR комбинируют с образованием мультимера sTCR.
60. Способ по п.59, отличающийся тем, что две, три, четыре или более молекул T-лимфоцитного рецептора связываются одна с другой, предпочтительно посредством молекулы линкера.
61. Комплекс по пп.58, 59 или 60, отличающийся тем, что растворимые T-лимофцитные рецепторы или их мультимеры комбинируют в липидном двойном слое (бислое) или присоединяют к какой-либо частице.
62. Способ обнаружения комплексов МНС-пептид, включающий
(1) получение растворимого TCR способом по пп.37-57 или мультивалентного комплекса T-лимфоцитного рецептора способом по пп.58-61,
(2) контактирование растворимого TCR или мультивалентного комплекса TCR с комплексами МНС-пептид и
(3) детектирование связывания растворимого TCR или мультивалентного комплекса TCR с комплексами МНС-пептид.
63. Фармацевтический состав, содержащий sTCR, полученный способом по пп.37-57, и/или мультивалентный комплекс TCR, полученный способом по пп.58-61, совместно с фармацевтически допустимым носителем.
Текст
006601 Настоящее изобретение относится к растворимым Т-лимфоцитным рецепторам (Т cell receptor,TCR). Как указано в патентной заявке WO 99/60120, TCR опосредуют распознавание специфических комплексов Главный Комплекс Гистосовместимости (Major Histocompatibility Complex, МНС)-пептид Тлимфоцитами и в этом качестве являются важными для функционирования клеточной ветви иммунной системы. Только два типа молекул - антитела и TCR - могут распознавать антигены специфичным образом,поэтому TCR является единственным рецептором для распознавания конкретных пептидных антигенов,представленных в МНС, при этом чужеродный пептид часто является единственным признаком анормальности клетки. Т-лимфоцитное распознавание происходит, когда Т-лимфоцит и антигенпредставляющая клетка (antigen presenting cell, APC) находятся в прямом физическом контакте, и инициируется лигированием антиген-специфичных TCR с комплексами рМНС (пептид-МНС).TCR представляет собой гетеродимерный белок клеточной поверхности суперсемейства иммуноглобулинов, который ассоциирован с инвариантными белками комплекса CD3, участвующего в опосредовании трансдукции сигналов. TCR существуют в формахи , которые являются сходными по структуре, однако, Т-лимфоциты, экспрессирующие их, имеют совершенно различные анатомические расположения и, вероятно, функции. Внеклеточная часть рецептора состоит из двух проксимальных относительно мембраны констатных доменов и двух дистальных относительно мембраны вариабельных доменов, содержащих полиморфные петли, аналогичные гипервариабельным участкам (complementarydetermining regions, CDR) антител. Именно эти петли образуют связывающий сайт молекулы TCR и определяют пептидную специфичность. Лиганды МНС класса I и класса II также относятся к белкам суперсемейства иммуноглобулинов, однако, они специализируются на представлении антигенов и обладают полиморфной частью, связывающей пептиды и позволяющей им представлять различные наборы коротких пептидных фрагментов на поверхности клеток АРС. Растворимые TCR полезны не только для исследования специфических взаимодействий TCRpMHC, но также в качестве потенциального диагностического инструмента, позволяющего обнаруживать инфекцию или маркеры аутоиммунного заболевания. Растворимые TCR применяют также для окрашивания, например, для окрашивания клеток с целью определения присутствия определенного пептидного антигена, представленного в контексте МНС. Аналогичным образом растворимые TCR можно использовать для доставки терапевтического агента, например цитотоксического соединения или иммуностимулирующего соединения, к клеткам, представляющим определенный антиген. Растворимые TCR можно также использовать для подавления Т-лимфоцитов, например, реагирующих с аутоиммунным пептидньм антигеном. При получении растворимой формы белков, образованных несколькими полипетидными субъединицами и содержащих трансмембранный домен, могут возникнуть сложности, поскольку во многих случаях белок стабилизируется своей трансмембранной областью. Это относится также к TCR, и находит отражение в научной литературе, где описываются усеченные формы TCR, содержащие только внеклеточные домены или внеклеточные и цитоплазматические домены, которые можно распознать антителами, специфическими для данного TCR (с указанием на то, что часть рекомбинантного TCR, распознанного антителом, была уложена соответствующим образом), но которые не удалось получить с хорошим выходом, устойчивыми при низких концентрациях и/или способными распознавать комплексы пептидМНС. Обзор такой литературы приведен в WO 99/60120. В ряде работ описано получение гетеродимеров TCR, которые содержат нативный дисульфидный мостик, связывающий соответствующие субъединицы (Garboczi, et al., (1996), Nature 384(6605): 134-41;US Patent No. 6080840). Однако, хотя такие TCR можно распознать TCR-специфическими антителами,для них не была продемонстрирована способность распознавания ими нативного лиганда, кроме как при относительно высоких концентрациях и/или же они были нестабильными. В WO 99/60120 описан растворимый TCR, который уложен правильным образом и поэтому может распознавать свой нативный лиганд, является стабильным в течение определенного периода времени и может быть получен в достаточных количествах. Этот TCR содержит внеклеточный домен - или -цепиTCR, димеризованный с внеклеточным доменом - или -цепи TCR, соответственно, с помощью пары димеризационных пептидов С-концевой области, в частности пептидов с лейциновыми зипперами. Такая стратегия получения TCR в общем случае применима ко всем TCR.Reiter et al., Immunity, 1995, 2:281-287, подробно описывают конструкцию растворимой молекулы,содержащей дисульфидно-стабилизированные вариабельные - и -домены TCR, один из которых связан с усеченной формой Pseudomonas exotoxin (PE38). Одна из указанных причин получения такой молекулы заключалась в преодолении нестабильности, присущей одноцепочечным TCR. Позицию новой дисульфидной связи в вариабельных доменах TCR идентифицировали путем гомологии с вариабельными доменами антител, в которые их предварительно вводили (см., например, Brinkmann, et al. (1993), Proc.Natl. Acad. Sci. USA 90: 7538-7542, и Reiter, et al. (1994) Biochemistry 33: 5451-5459). Однако такая гомология между антителом и константными доменами отсутствует. Поэтому применение указанного способа с целью идентификации соответствующих сайтов для новых межцепочечных дисульфидных связей,расположенных между константными доменами TCR, было невозможным. Учитывая важность растворимых TCR, было бы желательным обеспечить альтернативный способ получения таких молекул. Согласно первой задаче настоящее изобретение обеспечивает растворимый Т-лимфоцитный рецептор (soluble Т cell receptor, sTCR), который содержит (I) всю -цепь TCR или ее часть, за исключением ее трансмембранного домена и (II) всю -цепь TCR или ее часть, за исключением ее трансмембранного домена, при этом каждый из компонентов (I) и (II) содержит функциональный вариабельный домен и по меньшей мере часть константного домена цепи TCR, и они соединены дисульфидной связью между(аминокислотными) остатками константного домена, которая отсутствует в нативном TCR. Согласно другой задаче изобретение обеспечивает растворимый Т-лимфоцитный рецептор (sTCR) в-форме, в котором ковалентная дисульфидная связь соединяет (аминокислотный) остаток из области иммуноглобулина константного домена -цепи с остатком из области иммуноглобулина константного домена -цепи. Достоинство sTCR согласно изобретению заключается в том, что они не содержат гетерологичных полипептидов, которые могут быть иммуногенньми или которые могут приводить к быстрому выведению sTCR из организма. Кроме того, TCRs согласно настоящему изобретению имеют трехмерную структуру, в высшей степени сходную с нативными TCR, из которых они были получены. Благодаря этому структурному сходству, они маловероятно будут иммуногенными. sTCR в соответствии с изобретением могут быть использованы для распознавания комплексов МНС класса I - пептид или комплексами МНС класса II - пептид.TCR согласно настоящему изобретению являются растворимыми. В контексте данной заявки растворимость определяется как способность TCR к очищению в форме монодисперсного гетеродимера в забуференном фосфатом физиологическом растворе (phosphate buffered saline, PBS) (KCL 2,7 мМ,КН 2 РO4 1,5 мМ, NaCl 137 мМ и Na2PO4 8 мМ, рН 7,1-7,5. Life Technologies, Gibco BRL) при концентрации 1 мг/мл, при этом 90% указанного TCR остается в форме монодисперсного гетеродимера после инкубации при 25 С в течение 1 ч. Чтобы определить растворимость TCR, его сначала очищают, как описано в примере 2. После такой очистки 100 мкг TCR анализируют способом гель-хроматографии, используя, например, колонку Pharmacia Superdex 75 HR, уравновешенную PBS. Другие 100 мкг TCR инкубируют при 25 С в течение 1 ч, а затем анализируют способом гель-хроматографии, как указано выше. Затем с помощью интегрирования анализируют хроматограмму и сравнивают площади под пиками, соответствующими монодисперсному гетеродимеру. Соответствующие пики можно идентифицировать путем сравнения с позицией элюции стандартных белков с известной молекулярной массой. Монодисперсный гетеродимерный растворимый TCR имеет молекулярную массу примерно 50 кДа. Как указано выше, TCR согласно настоящему изобретению являются растворимыми. Однако, как более подробно поясняется далее, TCR могут быть соединены с группой таким образом, что образующийся комплекс является нерастворимым, или они могут быть представлены на поверхности нерастворимой твердой подложки. Нумерация аминокислот TCR, используемая в данном описании, соответствует системе иммуногенетической базы данных (Immunogenetics database, IMGT) описанной в работе The Т Cell ReceptorFactsbook, 2001, LeFrancLeFranc, Academic Press. В этой системе константный домен -цепи имеет следующее обозначение: TRAC01, где TR означает ген Т-лимфоцитного рецептора, А означает ген цепи, С означает константную область и 01 означает аллель 1. Константный домен -цепи имеет следующее обозначение: TRBC101. В данном случае имеется два возможных гена константной области С 1 и С 2. Транслируемый домен, кодируемый каждой аллелью, может быть образован генетическим кодом нескольких экзонов, поэтому они также являются специфическими. Аминокислоты нумеруют согласно экзону определенного домена, в котором они присутствуют. Внеклеточная часть нативного TCR состоит из двух полипептидов ( или ), каждый из которых имеет константный домен, проксимальный по отношению к мембране, и вариабельный домен, дистальный по отношению к мембране (см. фиг. 1). Каждый из константных и вариабельных доменов содержит внутрицепочечную дисульфидную связь. Вариабельные домены содержат высокополиморфные петли,аналогичные гипервариабельным участкам (complementarity determining regions, CDR) антител. CDR3TCR взаимодействует с пептидом, представленным МНС, а CDR 1 и 2 взаимодействуют с пептидом и МНС. Многообразие последовательностей TCR образуется посредством соматической перегруппировки соединенных вариабельных (Variable, V), разнообразных (Diversity, D), соединительных (Joining, J) и константных (Constant, С) генов. Функциональные полипептиды -цепей состоят из перестроенных участков V-J-C, в то время как -цепи состоят из участков V-D-J-C. Внеклеточный константный домен содержит проксимальную по отношению к мембране область и область иммуноглобулина. Проксимальная по отношению к мембране область содержит аминокислоты между трансмембранным доменом и про-2 006601 ксимальной по отношению к мембране остатком цистеина. Константный домен иммуноглобулина состоит из остальной части остатков аминокислот константного домена, занимающих участок от проксимального по отношению к мембране цистеина до начала соединительной области, и отличается наличием укладки иммуноглобулинового типа. Существует один константный домен -цепи, известный как С 1 илиTRAC01, и два различных константных -домена, известных как С 1 или TRBC101 и С 2 илиTRBC201. Различие между этими константными -доменами заключается в остатках аминокислот 4, 5 и 37 экзона 1. Так, TRBC101 содержит в своем экзоне 1 4N, 5 К и 37F, a TRBC201 содержит в своем экзоне 14 К, 5N и 37Y. Размеры всех внеклеточных доменов TCR варьируют в некоторых пределах. Согласно настоящему изобретению дисульфидную связь вводят между остатками, расположенными в константных доменах (или их частях) соответствующих цепей. Соответствующие цепи TCR содержат достаточное количество вариабельных доменов, чтобы взаимодействовать с его комплексом рМНС. Такое взаимодействие можно измерить с помощью прибора BIAcore 3000 или BIAcore 2000, как описано далее в примере 3 или в патентной заявке WO 99/6120 соответственно. В одном из вариантов реализации изобретения соответствующие цепи sTCR согласно изобретению содержат также внутрицепочечные дисульфидные связи. TCR согласно настоящему изобретению могут содержать всю внеклеточную константную область Ig соответствующих цепей TCR и предпочтительно весь внеклеточный домен соответствующих цепей, т.е. включая область, проксимальную относительно мембраны. В нативном TCR имеется дисульфидная связь, которая соединяет консервативные проксимальные относительно мембраны области соответствующих цепей. В одном варианте реализации настоящего изобретения эта дисульфидная связь отсутствует. Это можно обеспечить путем мутации соответствующих цистеиновых остатков (аминоксилота 4, экзон 2 гена TRAC01 и аминокислота 2 обоих генов TRBC101 и TRBC201 соответственно) в другую аминокислоту или усечения соответствующих цепей таким образом, чтобы исключить цистеиновые остатки. Предпочтительный растворимый TCR согласно изобретению содержит нативные - и -цепи TCR, усеченные в С-концевой области таким образом, чтобы исключить цистеиновые остатки, которые образуют нативную межцепочечную дисульфидную связь, т.е. усеченные в остатках 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 N-концевой области с цистеиновыми остатками. Однако следует отметить, что нативная межцепочечная дисульфидная связь может присутствовать в TCR согласно настоящему изобретению и что в некоторых вариантах реализации только одна из цепей TCR имеет нативный остаток цистеина, который образует нативную межцепочечную дисульфидную связь. Этот цистеин можно использовать для присоединения различных групп к TCR. Однако соответствующие цепи TCR могут быть более короткими. Поскольку константные домены не имеют непосредственных контактов с лигандами пептид-МНС, положение точки усечения в Сконцевой области можно существенно изменять без потери функциональности. В альтернативном исполнении может быть представлен более крупный фрагмент константных доменов, чем в предпочтительном случае, т.е. константные домены можно не процессировать непосредственно перед остатками цистеина, которые образуют межцепочечную дисульфидную связь. Так, например, можно включить полный константный домен за исключением трансмембранного домена (т.е. внеклеточные и цитоплазматические домены). В данном случае может оказаться предпочтительным мутировать один или несколько остатков цистеина, образующих межцепочечную дисульфидную связь в клеточном TCR в другой остаток аминокислоты, который не участвует в образовании дисульфидной связи,или удалить один или несколько таких остатков. Сигнальный пептид можно исключить, если растворимый TCR должен быть экспрессирован в прокариотических клетках, например, E.coli, поскольку он не выполняет никакой задачи в зрелом TCR при обеспечении его способности связываться с лигандом и при определенных обстоятельствах может препятствовать образованию функционального растворимого TCR. В большинстве случаев сайт расщепления, по которому сигнальный пептид удаляется из цепей зрелого TCR, можно прогнозировать, но нельзя определить экспериментально. При этом модификация экспрессированных цепей TCR, таким образом,чтобы они были на несколько, например до 10 аминокислот, более длинными или короткими в Nконцевой области, может не иметь существенного значения для функциональности (т.е. способности распознавать рМНС) растворимого TCR. Можно вводить определенные добавки, отсутствующие в последовательности оригинальных белков. Так, например, можно добавить короткую последовательность метки, которая может способствовать очистке цепей TCR при условии, что она не является препятствием для корректной структуры и укладки антигенсвязывающего сайта TCR. Для экспрессии в E.coli в начальную точку N-концевой области прогнозируемой последовательности зрелого белка можно ввести остаток метионина, чтобы обеспечить возможномть инициации трансляции. Далеко не все остатки в вариабельных доменах цепей TCR являются существенными для антигенной специфичности и функциональности. Поэтому значительное количество мутаций можно ввести в этот домен, не оказывая влияния на антигенную специфичность и функциональность. Также далеко не все остатки в константных доменах цепей TCR являются существенными для антигенной специфичности-3 006601 и функциональности. Поэтому значительное количество мутаций можно ввести в эту область, не оказывая влияния на антигенную специфичность.-цепь TCR содержит остаток цистеина, который является неспаренным в клеточном или нативномTCR. Предпочтительно удалять эту цистеиновую группу или мутировать ее в другую группу, чтобы избежать неправильного межцепочечного или внутрицепочечного спаривания. Заместители этой цистеиновой группы, например серин или аланин, могут оказывать существенное положительное влияние на эффективность укладки in vitro. Дисульфидную связь можно получить, мутируя нецистеиновые остатки в соответствующих цепях в цистеин и инициируя образование связи между мутированными группами. Предпочтительными являются группы, в которых расстояние между соответствующими -атомами углерода в нативном TCR составляет примерно 6(0,6 нм) или менее и предпочтительно в пределах от 3,5(0,35 нм) до 5,9(0,59 нм),при этом дисульфидную связь можно получать между остатками цистеина, введенными вместо нативных групп. Предпочтительно, чтобы дисульфидная связь находилась между остатками в константной области иммуноглобулина, однако, она может находиться и между остатками в проксимальной относительно мембраны области. Предпочтительными сайтами, в которые можно вводить остатки цистеина для получения дисульфидной связи, являются следующие остатки в экзоне 1 TRAC01 для -цепи TCR и Один из sTCR согласно настоящему изобретению получают из TCR A6 Tax (Garboczi et al., Nature,1996, 384(6605): 134-141). В одном из вариантов реализации sTCR содержит всю область -цепи TCR в направлении N-конца от экзона 2, остаток 4 TRAC01 (остатки 1-182 аминокислот -цепи согласно нумерации, используемой в работе Garboczi et al.) и всю область -цепи TCR в направлении N-конца от экзона 2, остаток 2 TRAC01 и TRCB201 (остатки 1-210 аминокислот -цепи согласно нумерации, используемой в работе Garboczi et al.). Чтоб получить дисульфидную связь, треонин 48 экзона 1 вTRAC01 (треонин 158 -цепи согласно нумерации, используемой в работе Garboczi et al.) и серии 57 экзона 1 в TRBC101 и TRBC201 (серии 172 -цепи согласно нумерации, используемой в работе Garboczi et al.) могут быть мутированы в цистеин. Эти аминокислоты располагаются в -цепи D константного домена - и -цепей TCR соответственно. Следует отметить, что на фиг. 3 а и 3b группа 1 (согласно нумерации, используемой в работе Garboczi et аl.) представляет собой K и N соответственно. Остаток метионина в N-концевой области не присутствует в нативном TCR A6 Tax и, как указано выше, иногда присутствует, если соответствующие цепи продуцируют в бактериальных системах экспресии. Теперь, когда идентифицированы остатки в TCR человека, которые могут быть мутированы в цистеиновые группы, чтобы получить новую межцепочечную дисульфидную связь, специалисты в данной области могут мутировать любой TCR таким образом, чтобы получить растворимую форму указанногоTCR, содержащего новую межцепочечную дисульфидную связь. В случае организма человека специалистам достаточно только найти следующие мотивы в соответствующих цепях TCR, чтобы определить группу, подлежащую мутации (затененная группа представляет собой группу для мутации в цистеин). У других видов цепи TCR могут не иметь области, которая на 100% идентична вышеуказанным конфигурациям. Однако специалисты смогут использовать вышеуказанные конфигурации для идентификации эквивалентной части - или -цепи TCR и, следовательно, группы, подлежащей мутации в цистеин. Для этой цели можно использовать способ сравнительного анализа первичной структуры. Так, например, можно воспользоваться системой ClustalW, представленной на веб-сайте Европейского института биоинформатики (http://www.ebi.ac.uk/index.html), чтобы сравнить вышеприведенные конфигурации с последовательностью цепей определенного TCR, чтобы определить положение соответствующей части последовательности TCR для мутации. Настоящее изобретение включаетTCR человека с дисульфидной связью, а такжеTCR других млекопитающих с дисульфидной связью, в частности, но без ограничения изобретения, мыши, крысы,свиньи, козы и овцы. Как указано выше, специалисты в данной области смогут определить сайты, эквивалентные вышеописанным сайтам человека, в которые можно ввести цистеиновые группы, чтобы получить межцепочечную дисульфидную связь. Так, например, ниже представлены последовательности аминокислот растворимых доменов С и C мыши совместно с мотивами, показывающими остатки аминокислот мыши, эквивалентные вышеуказанным остаткам аминокислот человека, которые можно мутировать в цистеины с образованием межцепочечной дисульфидной связи TCR (соответствующие группы затенены) В предпочтительном варианте реализации настоящего изобретения оба TCR (I) и (II) содержат функциональный вариабельный домен первого TCR, слитый с полным константным доменом или его частью второго TCR, при этом первый и второй TCR относятся к одному и тому же виду, а межцепочечная дисульфидная связь, между остатками в указанном соответствующем полном константном домене или его части, отсутствует в нативном TCR. В одном из вариантов реализации первый и второй TCR представляют собой TCR человека. Иными словами, константные домены с дисульфидной связью играют роль каркаса, к которому могут присоединяться вариабельные домены. Образующийся в результатеTCR будет практически идентичен нативному TCR, из которого получен первый TCR. Такая система позволяет легко экспрессировать любой функциональный вариабельный домен на стабильном каркасе из константного домена. Константные домены sTCR A6 Tax, описанные выше, или константные домены любых -мутантовsTCR, содержащие новую межцепочечную дисульфидную связь, описанную выше, можно использовать в качестве каркаса, к которым можно присоединять (сливать) гетерологические вариабельные домены. Предпочтительно, чтобы белки слияния сохраняли как можно в большей степени конформацию гетерологических вариабельных доменов. Поэтому предпочтительно, чтобы гетерологические вариабельные домены были связаны с константными доменами в любой точке между введенными цистеиновьми группами и N-концевой областью константного домена. Для TCR A6 Tax цистеиновые группы в - и -цепи предпочтительно располагают в положении треонина 48 экзона 1 в TRAC01 (треонин 158 -цепи согласно нумерации, используемой в работе Garboczi et al.) и серина 57 экзона 1 в TRBC101 и TRBC201(серин 172 -цепи согласно нумерации, используемой в работе Garboczi et al.) соответственно. Поэтому предпочтительно, чтобы точки присоединения гетерологических вариабельных доменов - и -цепи находились между группами 48 (159 согласно нумерации, используемой в работе Garboczi et al.) или 58(173 согласно нумерации, используемой в работе Garboczi et al.) и N-концевой областью константных доменовилисоответственно. Группы в константных доменах гетерологических - и -цепей, соответствующие точкам присоединения в TCR A6 Tax, можно идентифицировать по гомологии последовательностей. Белок слияния предпочтительно конструируют таким образом, чтобы он включал всю гетерологическую последовательность в направлении N-конца к точке присоединения. Как более подробно описано ниже, можно получить производные sTCR согласно настоящему изобретению с некоторой группой в его С- или N-концевой области или сливать с этой группой. С-концевая область предпочтительна, поскольку она является дистальной относительно связывающего домена. В одном из вариантов реализации одна или обе цепи TCR содержат остаток цистеина в своей С- и/или Nконцевой области, с которым может быть слита соответствующая группа. Растворимый TCR (который предпочтительно представляет собой TCR человека) согласно настоящему изобретению можно получить в практически чистой форме или в форме очищенного или выделенного препарата. Так, например, его можно получить в форме, практически не содержащей других белков. Множество растворимых TCR согласно настоящему изобретению можно получить в поливалентном комплексе. Таким образом, одной из задач настоящего изобретения является поливалентный комплекс Т-лимфоцитных рецепторов (Т cell receptor, TCR), который содержит множество растворимых Тлимфоцитных рецепторов согласно данному описанию. Все растворимые TCR в этом множестве предпочтительно являются идентичными. Другой задачей изобретения является способ обнаружения комплексов пептид-МНС, который включает 1) получение растворимого Т-лимфоцитного рецептора или поливалентного комплекса Тлимфоцитных рецепторов, согласно настоящему описанию,2) контактирование растворимого Т-лимфоцитного рецептора или поливалентного комплекса TCR с комплексами пептид-МНС и 3) определение (детектирование) связывания растворимого Т-лимфоцитного рецептора или поливалентного комплекса TCR с комплексами МНС-пептид. В поливалентном комплексе согласно настоящему изобретению TCR могут иметь форму мультимеров и/или могут быть представлены в липидном двойном слое или связаны с ним, например, в форме липосомы. В своей простейшей форме поливалентный комплекс TCR согласно изобретению содержит мультимер из двух, трех, четырех или большего количества молекул Т-лимфоцитного рецептора, связанных(например, ковалентной или иной связью) с другой молекулой, предпочтительно посредством молекулы линкера. Пригодные молекулы линкеров включают, в частности, но без ограничения изобретения, поливалентные присоединительные молекулы, например авидин, стрептавидин, нейтравидин и экстравидин,каждый из которых имеет четыре связывающих сайта для биотина. Таким образом, биотинилированные молекулы TCR можно получать в мультимерах Т-лимфоцитных рецепторов, имеющих множество связывающих сайтов TCR. Количество молекул TCR в мультимере зависит от отношения количества TCR к количеству молекул линкера, используемых для получения мультимеров, а также от наличия или отсутствия других биотинилированных молекул. Предпочтительными мультимерами являются димерные,тримерные или тетрамерные комплексы TCR. Структуры гораздо более крупные, чем тетрамеры TCR, можно использовать для отслеживания или определения клеток, экспрессирующих специфический комплекс МНС-пептид. Предпочтительные структуры имеют диаметр в пределах от 10 нм до 10 мкм. Каждая структура может содержать множество молекул TCR с достаточным расстоянием между ними, чтобы две или несколько молекул TCR в структуре могли одновременно соединяться с двумя или несколькими комплексами пептид-МНС в клетке и тем самым увеличивать сродство многомерной связующей группы для указанной клетки. Пригодные структуры для применения согласно изобретению включают мембранные структуры, в частности липосомы, и твердые структуры, которые представляют собой предпочтительно частицы, например шарики, в частности латексные шарики. Другие структуры, которые могут иметь наружное покрытие молекулами Т-лимфоцитного рецептора, также являются пригодными. Структуры покрывают предпочтительно мультимерами Т-лимфоцитных рецепторов, а не отдельными молекулами Тлимфоцитных рецепторов. В случае липосом молекулы Т-лимфоцитных рецепторов или их изомеры можно присоединить к мембране или иным образом связать с ней. Соответствующие способы хорошо известны специалистам. В поливалентный комплекс TCR согласно настоящему изобретению можно включить маркерную или иную группу, в частности токсическую или терапевтическую группу. Так, например, маркерную или иную группу можно включать в смешанный молекулярный мультимер. Примером такой мультимерной молекулы является тетрамер, содержащий три молекулы TCR и одну молекулу пероксидазы. Ее можно получить путем смешивания TCR и фермента в молярном отношении 3:1, чтобы сформировать тетрамерные комплексы, и отделить желаемый комплекс от комплексов, не содержащих нужного отношения-7 006601 молекул. Эти смеси молекул могут содержать любую комбинацию молекул при условии, что стерическое препятствие не ухудшает или не существенно ухудшает желательную функцию молекул. Позиционирование связующих сайтов на молекуле стрептавидина является пригодным для смешанных тетрамеров, поскольку возникновение стерического препятствия не представляется вероятным. Возможны также альтернативные средства биотинилирования TCR. Так, например, можно использовать химическое биотинилирование. Можно использовать альтернативные биотиновые метки, однако,определенные аминокислоты в последовательности биотиновой метки являются существенными (Schatz,(1993). Biotechnology N Y 11(10): 1138-43). Смесь, применяемую для биотинилирования, также можно изменять. Фермент требует Mg-ATP и низкой ионной силы, однако, оба этих условия можно изменять,например, может оказаться возможным использовать более высокую ионную силу и более длительное время реакции. Возможно применение молекулы, отличной от авидина или стрептавидина, для получения мультимеров TCR. Пригодной является любая молекула, которая связывает биотин в поливалентное соединение. В альтернативном случае может быть использована совершенно иная связь, в частности полигистидиновая метка с хелатным ионом никеля (Quiagen Product Guide 1999, Chapter 3 "Protein Expression, Purification, Detection и Assay" p. 35-37). Метку располагают предпочтительно в направлении Сконцевой области белка, чтобы минимизировать величину стерического препятствия при взаимодействии с комплексами пептид-МНС. Одну или обе цепи TCR можно пометить обнаруживаемой (детектируемой) меткой, например меткой, которая пригодна для диагностических целей. Таким образом, изобретение обеспечивает способ обнаружения комплексов пептид-МНС, при этом указанный способ включает контактирование комплексов пептид-МНС с TCR или мультимерным комплексом TCR согласно изобретению, который специфичен для комплекса пептид-МНС, и определение связывания TCR или мультимерного комплекса TCR с комплексом пептид-МНС. В тетрамерном TCR, образованном с помощью биотинилированных гетеродимеров, можно использовать флуоресцентный стрептавидин (серийно выпускается), чтобы получить обнаруживаемую метку. Тетрамер с флуоресцентной меткой можно использовать для анализа способом клеточного сортера с возбуждением флуоресценции (fluorescent-activated cell sorter, FACS), например,для обнаружения антигенпредставляющих клеток, содержащих пептид, для которого TCR является специфичным. Другой способ, которым можно обнаружить растворимые TCR согласно настоящему изобретению,заключается в применении TCR-специфических антител, в частности моноклональных антител. Существует множество серийно выпускаемых антител против TCR, в частности F1 и F1, которые распознают константные области - и -цепи соответственно.TCR (или его поливалентный комплекс) согласно настоящему изобретению можно альтернативно или дополнительно соединить (например, ковалентной или иной связью) с терапевтическим агентом,который может представлять собой, например, токсическую группу, применяемую для убийства клеток,или иммуностимулирующий агент, в частности интерлейкин или цитокин. Многовалентный комплексTCR согласно настоящему изобретению может иметь увеличенную связывающую способность для рМНС по сравнению с немультимерным гетеродимером Т-лимфоцитного рецептора. Таким образом,поливалентные комплексы TCR согласно изобретению являются особенно полезными для отслеживания или определения (targeting) клеток, представляющих определенные антигены in vitro или in vivo, а также являются полезными в качестве промежуточных соединений для получения других поливалентных комплексов TCR, имеющих такое же применение. При этом TCR или поливалентный комплекс TCR можно получить в фармацевтически допустимом составе для применения in vivo. Изобретение обеспечивает также способ доставки терапевтического агента в клетку-мишень, который включает контактирование потенциальных клеток-мишеней с TCR или поливалентным комплексомTCR в соответствии с изобретением при условиях, которые позволяют присоединять TCR или поливалентный комплекс TCR к клетке-мишени, при этом указанный TCR или поливалентный комплекс TCR являются специфичньми для комплексов пептид-МНС и содержат связанный с ними терапевтический агент. Растворимый TCR или поливалентный комплекс TCR можно использовать, в частности, для доставки терапевтических агентов в клетки, представляющие определенный антиген. Это является полезным во многих случаях и, в особенности, при лечении опухолей. Терапевтический агент можно доставить таким образом, что он будет оказывать свое влияние в определенной области, а не только в тех клетках, с которыми он связан. Таким образом, особая стратегия предусматривает применение противоопухолевых молекул, связанных с Т-лимфоцитными рецепторами или поливалентными комплексамиTCR, специфичными для опухолевых антигенов. Для этой цели можно использовать, например, множество терапевтических агентов, в частности радиоактивные соединения, ферменты (например, перфорин) или химиотерапевтические агенты (например,цис-платин). Чтобы обеспечить проявление токсических эффектов в желаемой области, токсин внутри липосомы может быть связан со стрептавидином с целью медленного выделения соединения. Это пре-8 006601 дотвратит повреждения в процессе транспортировки в организме и обеспечит максимальный эффект токсина после соединения TCR с соответствующими антигенпредставляющими клетками. Другие пригодные терапевтические агенты включают малые цитотоксические агенты, т.е. соединения, способные убивать клетки млекопитающих,имеющие молекулярную массу менее 700 Да. Такие соединения могут также содержать токсические металлы, способные оказывать цитотоксический эффект. Кроме того, следует понимать, что эти цитотоксические агенты для мелких молекул включают также пролекарства, т.е. соединения, которые разлагаются или трансформируются при физиологических условиях, выделяя цитотоксические агенты. Примеры таких агентов включают цис-платин, производные майтансина, рашельмицин, калишамицин, доцетаксель,этопозид, гемцитабин, ифосфамид, иринотекан, мелфалан, митоксантрон, сорфимер-натрий-фотофрин II,темозолмид, топотекан, триметриатглюкоронат, ауристатин Е винкристин и доксорубицин,пептидные цитотоксины, т.е. белки или их фрагменты, способные убивать клетки млекопитающих. Примеры включают рицин, дифтерийный токсин, синегнойный бактериальный экзотоксин А, ДНКазу и РНКазу,радионуклиды, т.е. неустойчивые изотопы элементов, которые разлагаются с параллельным излучением одной или нескольких - или -частиц или -лучей. Примеры включают йод 131, рений 186, индий 111, иттрий 90, висмут 210 и 213, актиний 225 и астат 213,пролекарства (предшественники лекарств), в частности ферментные пролекарства, направленные на антитела,иммуностимуляторы, т.е. группы, которые стимулируют иммунную реакцию. Примеры включают цитокины, в частности IL-2, хемокины, в частности IL-8, тромбоцитный фактор 4, белок-стимулятор роста мелоаномы и т.д., антитела или их фрагменты, комплементные активаторы, ксеногенные белковые домены, аллогенные белковые домены, вирусные/бактериальные белковые домены и вирусные/бактериальные пептиды. Растворимые TCR или поливалентные комплексы TCR согласно изобретению могут быть связаны с ферментом, способным трансформировать пролекарство в лекарство. Это позволяет трансформировать пролекарство в лекарство только в той области, где оно требуется (т.е. целенаправленное действиеsTCR). Примеры пригодных мишеней пептид-МНС для TCR согласно изобретению включают, в частности,но без ограничения изобретения, вирусные эпитопы, например эпитопы HTLV (вирус человеческого Тклеточного лейкоза) - 1 (в частности, пептид Tax, ограниченный по HLA-A2; HTLV-1 - связан с лейкемией), ВИЧ-эпитопы, эпитопы EBV, эпитопы CMV, эпитопы меланомы и другие эпитопы, специфичные для раковых заболеваний, (например, связанный антиген G250, ограниченный по HLA-A2), а также эпитопы, связанные с аутоиммунными нарушениями, в частности с ревматическим артритом. Кроме того,мишени рМНС, связанные с заболеваниями и пригодные для применения согласно настоящему изобретению, перечислены в работе HLA Factbook (Barclay (Ed) Academic Press), при этом также идентифицированы многие другие мишени. Во многих случаях лечение заболевания можно сделать гораздо более эффективным за счет локализации лекарственного препарата, благодаря специфичности растворимых TCR. Для лечения вирусных болезней, для которых существуют лекарства, например, ВИЧ, SIV (SimianImmunodefiency Virus, вирус иммунодефицита обезьяны), EBV (Epstein-Barr Virus, вирус ЭпштейнаБарра), CMV (Cytomegalovirus, цитомегаловирус) полезно выделение или активирование лекарства в непосредственной близости от инфицированных клеток. В случае рака локализация в непосредственной близости от опухолей или метастаз усиливает эффективность токсинов или иммуностимуляторов. При аутоиммунных заболеваниях возможность медленного выделения иммунодепрессивных препаратов обеспечивает повышенный локальный эффект в течение более длительного периода времени при минимальном воздействии на общую иммунную активность пациента. Для предотвращения отторжения трансплантанта таким же образом можно оптимизировать влияние иммунодепрессивных препаратов. Для доставки вакцины антиген, используемый для приготовления вакцины, можно локализовать вблизи от антигенпредставляющих клеток, повышая таким образом эффективность антигена. Способ можно применять также для визуализации изображений (тканей). Растворимые TCR согласно настоящему изобретению можно использовать для модулирования Тлимфоцитной активации путем связывания со специфическим рМНС и тем самым для ингибирования активации Т-лимфоцитов. Этот подход применим к аутоиммунным болезням, включая воспаление, опосредованное Т-лимфоцитами, и/или повреждение ткани, например диабет типа I. Для такого применения необходимо знание специфического пептидного эпитопа, представленного соответствующим рМНС. Медикаменты в соответствии с изобретением обычно поставляют как часть стерильного фармацевтического состава, который обычно содержит фармацевтически допустимый носитель. Этот фармацевтический состав, как правило, может иметь любую пригодную форму (в зависимости от желаемого способа введения пациенту). Он может поставляться в стандартной дозированной форме, обычно в герметичном контейнере и может представлять собой часть набора. Такой набор обычно (но не обязательно) содержит инструкцию по применению. Он может включать множество указанных стандартных доз.-9 006601 Фармацевтический состав можно адаптировать для ведения любым подходящим способом, например перорально (в том числе трансбуккально или сублингвально), ректально, назально, локально (в том числе трансбуккально, сублингвально или трансдермально), вагинально или парентерально (в том числе подкожно, внутримышечно, внутривенно или внутрикожно). Такие составы можно приготовлять любым способом, известным в фармации, например путем смешивания активного ингредиента с одним или несколькими носителями или наполнителями в стерильных условиях. Фармацевтические составы, предназначенные для перорального введения, могут быть представлены в форме дискретных доз, например капсул или таблеток, а также порошков, гранул, растворов, сиропов или суспензий (в водных или неводных жидкостях, или в форме съедобной пены, крема или эмульсий). Пригодные наполнители для таблеток или твердых желатиновых капсул включают лактозу, маисовый крахмал или его производные, а также стеариновую кислоту или ее соли. Пригодные наполнители для применения с мягкими желатиновыми капсулами включают, например, растительные масла, воски,жиры, полутвердые или жидкие многоатомные спирты. Примеры наполнителей, которые можно использовать для приготовления растворов и сиропов,включают воду, многоатомные спирты и сахара. Для получения масляных суспензий (например, растительных масел) можно использовать суспензии типа "масло в воде" или "вода в масле". Фармацевтические составы для трансдермального введения могут быть представлены в форме дискретных накладок,предназначенных для обеспечения плотного контакта с эпидермисом реципиента в течение длительного периода времени. Активный ингредиент можно доставлять, в частности, из накладки путем ионофореза,как описано в общем случае в Pharmaceutical Research, 3(6): 318 (1986). Фармацевтические составы для местного введения могут быть приготовлены в форме мазей, кремов, суспензий, лосьонов, порошков,растворов, паст, гелей, спреев, аэрозолей или масел. В случае инфекции глазной или других наружных тканей, например полости рта и кожи, составы наносят предпочтительно в форме локальной мази или крема. Для приготовления мази активный ингредиент можно использовать с парафиновой или со смешивающейся с водой масляной основой. Альтернативно активный ингредиент можно вводить в крем с эмульсионной основой типа "масло в воде" или "вода в масле". Фармацевтические составы для местного введения в глаз включают глазные капли, в которых активный ингредиент растворен или взвешен в соответствующем носителе, в частности в водном растворителе. Фармацевтические составы для местного введения в полость рта включают таблетки, пастилки и растворы для полоскания полости рта. Фармацевтические составы для ректального введения могут быть представлены в форме свечей или клизм. Фармацевтические составы для назального введения, в которых носитель является твердым веществом, включают крупные порошки с размером частиц, например, от 20 до 500 мкм, которые вводят также, как нюхательный табак, т.е. быстрым вдыханием через носовой ход из контейнера с порошком, который держат вблизи носа. Пригодные составы, в которых носителем является жидкость и которые предназначены для введения в качестве назальных спреев или капель, включают водные или масляные растворы активного ингредиента. Фармацевтические составы для введения путем ингаляции включают тонкодисперсную пыль или туман, которые можно получить с помощью различных типов дозированных, поступающих под давлением аэрозолей, распылителей или инсуффляторов. Фармацевтические составы для вагинального введения могут быть представлены в форме пессариев, тампонов, кремов, гелей, паст, пен или спреев. Фармацевтические растворы для парентерального введения включают водные и неводные стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостаты и растворенные вещества, которые делают раствор практически изотоническим с кровью соответствующего пациента, а также водные и неводные суспензии, которые могут включать взвешенные агенты и загустители. Наполнители, которые могут быть использованы в растворах для инъекций, включают, например, воду, спирты, многоатомные спирты, глицерин и растительные масла. Составы могут находиться в контейнере с одной или несколькими дозами, например, в герметичных ампулах и флаконах, могут храниться в сублимированном (лиофилизированном) состоянии и непосредственно перед применением требуют только добавления стерильной жидкости, например, воды для инъекций. Растворы для инъекций и суспензии для немедленного приема можно приготовить из стерильных порошков, гранул и таблеток. Фармацевтические составы могут содержать консерванты, солюбилизаторы, стабилизаторы, смачивающие вещества, эмульгаторы, подсластители, красители, отдушки, соли (вещества согласно настоящему изобретению сами могут быть представлены в форме соли), буферы, глазировочные агенты или антиоксиданты. Они могут содержать также другие терапевтические активные агенты кроме веществ согласно настоящему изобретению. Дозирование веществ согласно настоящему изобретению может изменяться в широких пределах в зависимости от заболевания или нарушения, подлежащего лечению, возраста и состояния пациента, которому требуется такое лечение, и т.д., поэтому терапевт должен окончательно определить дозы для приема. Дозировку можно повторять по мере необходимости. В случае возникновения побочных эффектов величину дозы и частоту приема можно уменьшить в соответствии с обычной клинической практикой. Для получения sTCR согласно изобретению, предпочтительно в практически чистой форме, можно использовать способы генного клонирования. Эти способы описаны в литературе, например, J. Sambrooket al. Molecular Cloning 2nd Edition, Cold Spring Harbor Laboratory Press (1989). Таким образом, согласно следующей задаче настоящее изобретение обеспечивает молекулу нуклеиновой кислоты, которая содержит последовательность, кодирующую цепь растворимого TCR согласно настоящему изобретению, или комплементарную последовательность. Такие последовательности нуклеиновых кислот можно получить путем выделения нуклеиновой кислоты, кодирующей TCR, из Т-лимфоцитных клонов и создания соответствующих мутаций (с помощью вставки, делеции или замещения). Молекула кислоты может быть представлена в изолированной или в рекомбинантной форме. Она может быть введена в вектор, который, в свою очередь, может быть введен в клетку-хозяина. Такие векторы и соответствующие хозяева представляют собой другие задачи настоящего изобретения. Изобретение обеспечивает также способ получения цепи TCR, включающий инкубацию указанной клетки-хозяина при условиях, вызывающих экспрессию цепи TCR, и последующую очистку полипептида. Растворимые TCR согласно настоящему изобретению можно получить путем экспрессии в бактерии, в частности, в Е. coli, в форме телец включения с последующей повторной укладкой in vitro. Повторная укладка цепей TCR может происходить in vitro в режимах, пригодных для повторной укладки. В одном из вариантов реализации TCR с корректной конформацией получают путем повторной укладки растворимых цепей TCR в буфере повторной укладки, содержащем солюбилизатор, например мочевину. Мочевина может быть представлена в концентрации предпочтительно по меньшей мере 0,1 М,или по меньшей мере 1 М, или по меньшей мере 2,5 М, или около 5 М. Альтернативно в качестве солюбилизатора может быть использован гуанидин с концентрацией от 1 до 8 М, предпочтительно по меньшей мере 1 М или по меньшей мере 2,5 М. Перед повторной укладкой предпочтительно используют восстановитель, чтобы обеспечить полное восстановление цистеиновых групп. Кроме того, в случае необходимости можно использовать денатурирующие агенты, в частности DTT и гуанидин. Различные денатуранты и восстановители можно использовать перед операцией повторной укладки (например, мочевину,-меркаптоэтанол). Альтернативно при повторной укладке можно использовать окислительновосстановительные пары, в частности окислительно-восстановительные пары цистамин/цистеамин, DTT или -меркаптоэтанол/атмосферный кислород и цистеин в восстановленной и окисленной формах. Эффективность укладки можно также увеличить путем добавления в повторно укладываемую смесь некоторых других белковых компонентов, например белков шаперонов. Улучшения повторной укладки удалось достичь путем пропускания белка через колонки с иммобилизованными мини-шаперонамиUSA 94(8): 3576-8). Растворимые TCR согласно настоящему изобретению можно получить альтернативно путем экспрессии в системе эукариотических клеток, в частности клеток насекомых. Очистку TCR можно произвести множеством различных способов. Так, например, можно использовать различные варианты ионного обмена или другие способы очистки белков, в частности гельхроматографию или аффинную хроматографию. Растворимые TCR и поливалентные комплексы TCR согласно настоящему изобретению также находят применение в скрининге агентов, в частности, малых химических соединений, способных подавлять связывание TCR с его комплексом рМНС. Таким образом, согласно следующей задаче настоящее изобретение обеспечивает способ скрининга агента, который подавляет связывание Т-лимфоцитного рецептора с комплексом пептид-МНС, при этом указанный способ включает мониторинг связывания растворимого Т-лимфоцитного рецептора согласно настоящему изобретению с комплексом пептид-МНС в присутствии соответствующего агента и селекцию агентов, которые подавляют указанное связывание. Пригодные способы для такого скрининга включают способ, основанный на поверхностном плазменном резонансе и описанный в WO 01/22084. Другими хорошо известными методиками, которые могут представлять собой основу для указанного способа скрининга, являются сцинтилляционный экспресс-анализ (Scintillation Proximity Analysis, SPA) и экспресс-анализ с усиленной люминесценцией (Amplified Luminiscent Proximity Assay). Агенты, селектированные способами скрининга согласно изобретению, могут быть использованы в качестве лекарственных препаратов или в качестве основы для программы разработки лекарственных препаратов, будучи модифицированными или усовершенствованными иным образом с целью получения характеристик, которые сделают их более пригодными для введения в качестве медикамента. Такие медикаменты можно использовать для лечения состояний, которые включают нежелательный компонент Тлимфоцитной реакции. Эти состояния включают рак (например, почки, яичника, кишечника, области головы и шеи, яичка, легкого, желудка, шейки матки, мочевого пузыря, простаты или меланомный рак кожи), аутоиммунные болезни, отторжение трансплантанта и трасплантант против болезни хозяина. Предпочтительные характеристики каждой указанной задачи изобретения, как и других задач, являются mutatis mutandis (можно получить, сделав соответствующие изменения). Известные документы,упомянутые в данном описании, включены во всей полноте, разрешенной законом.-11 006601 Примеры Ниже приведены примеры реализации изобретения, ни в какой степени не ограничивающие области распространения изобретения. Приводятся ссылки на прилагаемые фигуры, где показано следующее. Фиг. 1 - схематическое изображение структуры растворимого TCR с введенной межцепочечной дисульфидной связью согласно изобретению; фиг. 2 а и 2b - последовательности нуклеиновых кислот - и -цепей, соответственно, растворимогоTCR A6, мутированного таким образом, чтобы ввести кодон цистеина. Затенение указывает введенный цистеиновый кодон; фиг. 3 а - последовательность аминокислот внеклеточной части -цепи TCR A6, включающая мутацию Т 48 С (подчеркнута), которую используют для получения новой дисульфидной межцепочечной связи; фиг. 3b - последовательность аминокислот внеклеточной части -цепи TCR A6, включающая мутацию S57 С (подчеркнута), которую используют для получения новой дисульфидной межцепочечной связи; фиг. 4 - кривая, полученная в результате анионообменной хроматографии растворимого TCR A6,показывающая элюцию белка из колонки POROS 50HQ с использованием градиента NaCl 0-500 мМ, как указано пунктирной линией; фиг. 5 - А. Восстанавливающий электрофорез в полиакриламидном геле с додецисульфатом натрия(Sodium Dodecyi Sulfate - Polyacrylamide Gel Electrophoresis, SDS-PAGE) (с окрашиванием кумасси) фракций элюата из колонки, показанного на фиг. 4. В. Невосстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций элюата из колонки, показанного на фиг. 4. Пик 1 определенно содержит главным образом -цепь без дисульфидной связи, пик 2 содержит гетородимер TCR с межцепочечной дисульфидной связью, а плечо обусловлено загрязнениямиE.coli, смешанными с sTCR, содержащим межцепочечную дисульфидную связь, и плохо видимыми на этой репродукции; фиг. 6 - кривая, полученная в результате гель-хроматографии объединенных фракций пика 1 на фиг. 5. Белок элюирует как одинарный основной пик, соответствующий гетеродимеру; фиг. 7 - кривая реакции BIAcore специфического связывания растворимого TCR А 6 с дисульфидной связью и комплекса HLA-A2-tax. На вставке показана реакция связывания по сравнению с контрольным образцом для одной инъекции растворимого TCR A6 с дисульфидной связью; фиг. 8 а - последовательность -цепи TCR, содержащая новый остаток цистеина, мутированный для включения в сайт BamH1 рестрикции. Затенение показывает мутации, введенные для получения сайтаBamH1 рестрикции; фиг. 8b и 8 с - последовательность ДНК - и -цепи TCR JM22, мутированная для включения дополнительных остатков цистеина с целью получения ненативной дисульфидной связи; фиг. 9 а и 9b - последовательности аминокислот внеклеточной части - и -цепей, соответственно,TCR JM22, полученных из последовательностей ДНК с фиг. 8 а и 8b; фиг. 10 - кривая, полученная в результате анионообменной хроматографии растворимого TCR JM22 с дисульфидной связью, показывающая элюцию белка из колонки POROS 50HQ с использованием градиента NaCl 0-500 мМ, как указано пунктирной линией; фиг. 11 а - восстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций элюата из колонки,показанного на фиг. 10; фиг. 11b - невосстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций элюата из колонки, показанного на фиг. 10. Пик 1 определенно содержит гетородимер TCR, который присоединен межцепочечной дисульфидной связью; фиг. 12 - кривая, полученная в результате гель-хроматографии объединенных фракций пика 1 на фиг. 10. Белок элюирует как одинарный основной пик, соответствующий гетеродимеру. Выход составляет 80%; фиг. 13 - А. Кривая реакции BIAcore специфического связывания растворимого TCR JM22 с дисульфидной связью и комплекса HLA-Flu. В. Реакция связывания по сравнению с контрольным образцом для одной инъекции растворимогоTCR JM22 с дисульфидной связью; фиг. 14 а и 14b - последовательность ДНК - и -цепи NY-ESO, мутированную для включения дополнительных остатков цистеина с целью получения ненативной дисульфидной связи; фиг. 15 а и 15b - последовательности аминокислот внеклеточной части - и -цепей, соответственно,TCR NY-ESO, полученных из последовательностей ДНК с фиг. 14 а и 14b; фиг. 16 - кривая, полученная в результате анионообменной хроматографии растворимого TCR NYESO с дисульфидной связью, показывающая элюцию белка из колонки POROS 50HQ с использованием градиента NaCl 0-500 мМ, как указано пунктирной линией;-12 006601 фиг. 17 - А. Восстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций элюата из колонки, показанного на фиг. 16. В. Невосстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций элюата из колонки, показанного на фиг. 10. Пик 1 и 2 определенно содержат гетородимер TCR, который присоединен межцепочечной дисульфидной связью; фиг. 18 - кривая, полученная в результате гель-хроматографии объединенных фракций пика 1 (А) и пика 2 (В) на фиг. 17. Белок элюирует как одинарный основной пик, соответствующий гетеродимеру; фиг. 19 - кривая реакции BIAcore специфического связывания растворимого TCR NY-ESO с дисульфидной связью и комплекса HLA- NY-ESO. А - пик 1. В - пик 2; фиг. 20 а и 20b - последовательности ДНК - и -цепей растворимого TCR NY-ESO, мутированного таким образом, чтобы ввести новый цистеиновый кодон (показан затенением). Последовательности включают цистеин, содержащийся в нативной дисульфидной межцепочечной связи (кодон показан полужирным шрифтом); фиг. 21 а и 21b - последовательности аминокислот внеклеточной части - и -цепей, соответственно,TCR NY-ESO, полученных из последовательностей ДНК с фиг. 20 а и 20b; фиг. 22 - кривая, полученная в результате анионообменной хроматографии растворимогоTCRcyscys NY-ESO, показывающая элюцию белка из колонки POROS 50HQ с использованием градиента NaCl 0-500 мМ, как указано пунктирной линией; фиг. 23 - кривая, полученная в результате анионообменной хроматографии растворимого TCRcysNY-ESO, показывающая элюцию белка из колонки POROS 50HQ с использованием градиента NaCl 0500 мМ, как указано пунктирной линией; фиг. 24 - кривая, полученная в результате анионообменной хроматографии растворимого TCRcysNY-ESO, показывающая элюцию белка из колонки POROS 50HQ с использованием градиента NaCl 0500 мМ, как указано пунктирной линией; фиг. 25 - восстанавливающий SDS-PAGE (с окрашиванием кумасси) TCRcyscys, TCRcys и TCRcysNY-ESO фракций из элюатов анионообменной колонки, показанных на фиг. 22-24 соответственно. Дорожки 1 и 7: маркеры MW, дорожка 2: пик NYESOdsTCR1g4 -cys(ЕВ/084/033); дорожка 3: малый пик(EB/084/035); фиг. 26 - восстанавливающий SDS-PAGE (с окрашиванием кумасси) TCR NY-ESOcys cys, TCRcys и TCRcys элюатов фракций из анионообменной колонки, показанных на фиг. 22-24 соответственно. Дорожки 1 и 7: маркеры MW, дорожка 2: пик NYESOdsTCR1g4 -cys 13 (ЕВ/084/033); дорожка 3: малый пик NYESOdsTCR1g4 -cys(ЕВ/084/033), дорожка 4: NYESOdsTCR1g4-cys (ЕВ/084/034), дорожка 5: малый пик NYESOdsTCR1g4 -cys -cys (EB/084/035) и дорожка 6: пик NYESOdsTCR1g4 -cys -cys(EB/084/035); фиг. 27 - кривая, полученная результате обменной гель-хроматографии растворимого TCR NYESOcys cys, показывающая элюцию объединенных фракций с фиг. 22. Белок элюирует как одинарный основой пик, соответствующий гетеродимеру; фиг. 28 - кривая, полученная в результате обменной гель-хроматографии растворимого TCR NYESOcys, показывающая элюцию объединенных фракций с фиг. 22. Белок элюирует как одинарный основой пик, соответствующий гетеродимеру; фиг. 29 - кривая, полученная результате обменной гель-хроматографии растворимого TCR NYESOcys, показывающая элюцию объединенных фракций с фиг. 22. Белок элюирует как одинарный основой пик, соответствующий гетеродимеру; фиг. 30 - реакция BIAcore специфического связывания TCR NY-ESOcys cys с комплексом HLANY-ESO; фиг. 31 - реакция BIAcore специфического связывания TCR NY-ESOcys с комплексом HLA-NYESO; фиг. 32 - реакция BIAcore специфического связывания TCR NY-ESOcys с комплексом HLA-NYESO; фиг. 33 а и 33b - последовательности ДНК - и -цепи, соответственно, растворимого TCR АН-1.23,мутированного таким образом, чтобы ввести новый цистеиновый кодон (показан затенением). Последовательности включают цистеин, содержащийся в нативной дисульфидной межцепочечной связи (кодон показан полужирным шрифтом); фиг. 34 а и 34b - внеклеточные последовательности аминокислот - и -цепи TCR АН-1.23, полученные из последовательностей ДНК с фиг. 33 а и 33b; фиг. 35 - кривая, полученная в результате анионообменной хроматографии растворимого TCR АН 1.23, показывающая элюцию из колонки POROS 50HQ с использованием градиента 0-500 мМ NaCl, как указано пунктирной линией;TCR АН-1.23 из элюата анионообменной колонки с фиг. 35. Исследуемые белки представляют собой анионообменные фракции TCR 1.23 S-S из повторной укладки 3. Дорожка 1: маркеры MW, дорожка 2: В 4, дорожка 3: С 2, дорожка 4: С 3, дорожка 5: С 4, дорожка 6: С 5, дорожка 7: С 6, дорожка 8: С 7, дорожка 9: С 8 и дорожка 10: С 9; фиг. 37 - невосстанавливающий SDS-PAGE (10% бис-трис гель, с окрашиванием кумасси) фракцийTCR АН-1.23 из элюата анионообменной колонки с фиг. 35. Исследуемые белки представляют собой анионообменные фракции TCR 1.23 S-S из повторной укладки 3. Дорожка 1: маркеры MW, дорожка 2: В 4, дорожка 3: С 2, дорожка 4: С 3, дорожка 5: С 4, дорожка 6: С 5, дорожка 7: С 6, дорожка 8: С 7, дорожка 9: С 8 и дорожка 10: С 9; фиг. 38 - кривая, полученная результате обменной гель-хроматографии растворимого TCR АН-1.23,показывающая элюцию объединенных фракций с фиг. 35. Белок элюирует как одинарный основой пик,соответствующий гетеродимеру; фиг. 39 а и 39b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR А 6, мутированного таким образом, чтобы ввести новый цистеин в положение 48 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 40 а и 40b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR А 6, мутированного таким образом, чтобы ввести новый цистеин в положение 45 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 41 а и 41b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR А 6, мутированного таким образом, чтобы ввести новый цистеин в положение 61 в экзоне 1,TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 42 а и 42b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR A6, мутированного таким образом, чтобы ввести новый цистеин в положение 50 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 43 а и 43b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR A6, мутированного таким образом, чтобы ввести новый цистеин в положение 10 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 44 а и 44b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR A6, мутированного таким образом, чтобы ввести новый цистеин в положение 15 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 45 а и 45b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR, мутированного таким образом, чтобы ввести новый цистеин в положение 12 в экзоне 1 TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 46 а и 46b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR A6, мутированного таким образом, чтобы ввести новый цистеин в положение 22 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а затененная аминокислота показывает введенный цистеин; фиг. 47 а и 47b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR A6, мутированного таким образом, чтобы ввести новый цистеин в положение 52 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 48 а и 48b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR A6, мутированного таким образом, чтобы ввести новый цистеин в положение 43 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 49 а и 49b - соответственно, последовательности ДНК и аминокислот -цепи растворимогоTCR A6, мутированного таким образом, чтобы ввести новый цистеин в положение 57 в экзоне 1TRAC01. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 50 а и 50b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 77 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин;A6, мутированного таким образом, чтобы ввести новый цистеин в положение 17 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 52 а и 52b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 13 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 53 а и 53b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 59 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 54 а и 54b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 79 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 55 а и 55b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 14 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 56 а и 56b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 55 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 57 а и 57b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 63 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 58 а и 58b - соответственно, последовательности ДНК и аминокислот -цепи растворимого TCRA6, мутированного таким образом, чтобы ввести новый цистеин в положение 15 в экзоне 1 TRBC201. Затененные нуклеотиды показывают введенный новый цистеиновый кодон, а подчеркнутая аминокислота показывает введенный цистеин; фиг. 59-64 - кривые, полученные в результате анионообменной хроматографии растворимого TCRA6, содержащего новую дисульфидную межцепочечную связь между остатками 48 экзона 1 TRAC01 и 57 экзона 1 TRBC201; остатками 45 экзона 1 TRAC01 и 77 экзона 1 TRBC201; остатками 10 экзона 1TRAC01 и 17 экзона 1 TRBC201; остатками 45 экзона 1 TRAC01 и 59 экзона 1 TRBC201; остатками 52 экзона 1 TRAC01 и 55 экзона 1 TRBC201; остатками 15 экзона 1 TRAC01 и 15 экзона 1 TRBC201,соответственно, при элюции белка из колонки POROS 50 с помощью градиента 0-500 мМ NaCl, как указано пунктирной линией; фиг. 65 а и 65b - соответственно, результаты восстановительного и невосстановительного SDSPAGE (с окрашиванием кумасси) растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 48 экзона 1 TRAC01 и 57 экзона 1 TRBC201, элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 59; фиг. 66 а и 66b - соответственно, результаты восстановительного и невосстановительного SDSPAGE (с окрашиванием кумасси) растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 45 экзона 1 TRAC01 и 77 экзона 1 TRBC201, элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 60; фиг. 67 а и 67b - соответственно, результаты восстановительного и невосстановительного SDSPAGE (с окрашиванием кумасси) растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 10 экзона 1 TRAC01 и 17 экзона 1 TRBC201, элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 61; фиг. 68 а и 68b - соответственно, результаты восстановительного и невосстановительного SDSPAGE (с окрашиванием кумасси) растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 45 экзона 1 TRAC01 и 59 экзона 1 TRBC201, элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 62; фиг. 69 а и 69b - соответственно, результаты восстановительного и невосстановительного SDSPAGE (с окрашиванием кумасси) растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 52 экзона 1 TRAC01 и 55 экзона 1 TRBC201, элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 63; фиг. 70 а и 70b - соответственно, результаты восстановительного и невосстановительного SDSPAGE (с окрашиванием кумасси) растворимого TCR A6, содержащего новую дисульфидную межцепо-15 006601 чечную связь между остатками 15 экзона 1 TRAC01 и 15 экзона 1 TRBC201, элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 64; фиг. 71 - кривая, полученная в результате гель-хроматографии растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 48 экзона 1 TRAC01 и 57 экзона 1TRBC201, показывающая элюцию белка из колонки Superdex 200 HL гель-фильтрации. Элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 59; фиг. 72 - кривая, полученная в результате гель-хроматографии растворимого TCR А 6, содержащего новую дисульфидную межцепочечную связь между остатками 45 экзона 1 TRAC01 и 77 экзона 1TRBC201, показывающая элюцию белка из колонки Superdex 200 HL гель-фильтрации. Элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 60; фиг. 73 - кривая, полученная в результате гель-хроматографии растворимого TCR А 6, содержащего новую дисульфидную межцепочечную связь между остатками 10 экзона 1 TRAC01 и 17 экзона 1TRBC201, показывающая элюцию белка из колонки Superdex 200 HL гель-фильтрации. Элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 61; фиг. 74 - кривая, полученная в результате гель-хроматографии растворимого TCR А 6, содержащего новую дисульфидную межцепочечную связь между остатками 45 экзона 1 TRAC01 и 59 экзона 1TRBC201, показывающая элюцию белка из колонки Superdex 200 HL гель-фильтрации. Элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 62; фиг. 75 - кривая, полученная в результате гель-хроматографии растворимого TCR А 6, содержащего новую дисульфидную межцепочечную связь между остатками 52 экзона 1 TRAC01 и 55 экзона 1TRBC201, показывающая элюцию белка из колонки Superdex 200 HL гель-фильтрации. Элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 63; фиг. 76 - кривая, полученная в результате гель-хроматографии растворимого TCR А 6, содержащего новую дисульфидную межцепочечную связь между остатками 15 экзона 1 TRAC01 и 15 экзона 1TRBC201, показывающая элюцию белка из колонки Superdex 200 HL гель-фильтрации. Элюат фракций отобрали из элюата анионообменной колонки, показанного на фиг. 64; фиг. 77-80 - кривые реакции BIAcore, показывающие, соответственно, соединение растворимогоTCR А 6, содержащего новую дисульфидную межцепочечную связь между остатками 48 экзона 1TRBC201 с рМНС HLA-A2-tax; фиг. 81 - кривая BIAcore, показывающая неспецифическое связывание растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 52 экзона 1 TRAC01 и 55 экзона 1 TRBC201 с HLA-A2-tax и с рМНС HLA-A2-NY-ESO; фиг. 82 - кривая реакции BIAcore, показывающая связывание растворимого TCR A6, содержащего новую дисульфидную межцепочечную связь между остатками 15 экзона 1 TRAC01 и 15 экзона 1TRBC201 с рМНС HLA-A2-tax; фиг. 83 а - карта электронной плотности модели с последовательностью 1BD2 (цепь A Thr164, цепь В Ser 174). На карте нанесены контуры при значениях 1.0, 2.0 и 3.0; фиг. 83b - карта электронной плотности после усовершенствования модели за счет введения Cys в две позиции А 164 и В 174. На карте нанесены контуры при таких же значениях , как и на фиг. 83 а; фиг. 84 - сравнение структур TCR 1BD2 и TCR NY-ESO согласно настоящему изобретения путем наложения указанных структур при изображении в виде ленты и спирали; фиг. 85 а и 85b - последовательности ДНК и аминокислот, соответственно, -цепи TCR NY-ESO,включающие сайт узнавания биотина. Сайт узнавания биотина выделен; фиг. 86 а и 86b - последовательности ДНК и аминокислот, соответственно, -цепи TCR NY-ESO,включающие гексагистидиновую метку. Гексагистидиновая метка вьщелена; фиг. 87 - элюция растворимого TCR NY-ESO, содержащего новую дисульфидную связь и последовательность узнавания биотина, из анионообменной колонки POROS 50HQ с помощью градиента 0-500 мМ NaCl, как указано пунктирной линией; фиг. 88 - элюция растворимого TCR NY-ESO, содержащего новую дисульфидную связь и гексагистидиновую метку, из анионообменной колонки POROS 50HQ с помощью градиента 0-500 мМ NaCl, как указано пунктирной линией; фиг. 89 - профиль элюции белка, полученный в результате гель-хроматографии объединенных фракций меченого биотином NY-ESO из элюата анионообменной колонки, показанного на фиг. 87; фиг. 90 - профиль элюции белка, полученный в результате гель-хроматографии объединенных фракций меченого гексагистидином NY-ESO из элюата анионообменной колонки, показанного на фиг. 88; фиг. 91a-h - гистограммы FACS, иллюстрирующие интенсивность окрашивания, полученную в 25000 случаев для линии (РР LCL) В-клеток, трансформированных положительным EBV HLA-A2 и культивированных при следующих концентрациях пептида NY-ESO и флуоресцентных тетрамеров TCRNY-ESO, соответственно, NYESO 0 - TCR 5 мкг, NYESO 10-4 М - TCR 5 мкг, NYESO 10-5 М - TCR 5 мкг,NYESO 10-6 М - TCR 5 мкг, NYESO 0 - TCR 10 мкг, NYESO 10-4 М - TCR 10 мкг, NYESO 10-5 М - TCR 10 мкг, NYESO 10-6 М - TCR 10 мкг; фиг. 92 - последовательность ДНК -цепи TCR A6, содержащего константную область TRBC101; фиг. 93 - кривая анионообменной хроматографии растворимого TCR A6, содержащего константную область TRBC101, показывающая элюцию из колонки POROS 50HQ с использованием градиента 0-500 мМ NaCl, как указано пунктирной линией; фиг. 94 - А. Восстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций из элюата из колонки, как показано на фиг. 93. В. Невосстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций из элюата из колонки, как показано на фи. 93; фиг. 95 - гель-хроматограмма объединенных фракций пика 2 с фиг. 93. Пик 1 содержит гетеродимерTCR, который присоединен межцепочечной дисульфидной связью; фиг. 96 - А. Анализ BIAcore специфического соединения растворимого TCR A6, содержащего дисульфидную связь, с комплексом HLA-Flu. В. Сравнение реакции связывания с контрольным образцом для одной инъекции растворимого TCRA6 с дисульфидной связью; фиг. 97 - последовательность нуклеиновых кислот мутированной -цепи TCR А 6, содержащей"свободный" цистеин; фиг. 98 - анионообменная хроматограмма растворимого TCR А 6, содержащего "свободный" цистеин, показывающая элюцию из колонки POROS 50HQ с использованием градиента 0-500 мМ NaCl, как указано пунктирной линией; фиг. 99 - А. Восстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций из элюата из колонки, показанного на фиг. 98. В. Невосстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций из элюата из колонки,показанного на фиг. 98; фиг. 100 - гель-хроматограмма объединенных фракций, представленных пиком 2 на фиг. 98. Пик 1 содержит гетеродимер TCR, который присоединен межцепочечной дисульфидной связью; фиг. 101 - А. Анализ BIAcore специфического соединения растворимого TCR A6, содержащего дисульфидную связь, с комплексом HLA-Flu. В. Сравнение реакции связывания с контрольным образцом для одной инъекции растворимого TCRA6 с дисульфидной связью; фиг. 102 - последовательность нуклеиновых кислот мутированной -цепи TCR A6, содержащей остаток серина, мутированный в "свободный" цистеин; фиг. 103 - анионообменнная хроматограмма растворимого TCR A6, содержащего остаток серина,мутированный в "свободный" цистеин, показывающая элюцию из колонки POROS 50HQ с использованием градиента 0-500 мМ NaCl, как указано пунктирной линией; фиг. 104 - А. Восстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций из элюата из колонки, показанного на фиг. 103. В. Невосстанавливающий SDS-PAGE (с окрашиванием кумасси) фракций из элюата из колонки,показанного на фиг. 103. Пик 2 определенно содержит гетородимер TCR, который присоединен межцепочечной дисульфидной связью; фиг. 105 - гель-хроматограмма объединенных фракций, представленных пиком 2 на фиг. 103. Пик 1 содержит гетеродимер TCR, который присоединен межцепочечной дисульфидной связью; фиг. 106 - А. Анализ BIAcore специфического соединения растворимого TCR A6, содержащего остаток серина. мутированный в "свободный" цистеин, с комплексом HLA-Flu. В. Сравнение реакции связывания с контрольным образцом для одной инъекции растворимого TCRA6 с дисульфидной связью; фиг. 107 - нуклеотидная последовательность pYX112; фиг. 108 - нуклеотидная последовательность pYX122; фиг. 109 - последовательности ДНК и белка pre-pro mating фактора альфа, слитого с -цепью TCR; фиг. 110 - последовательности ДНК и белка pre-pro mating фактора альфа, слитого с -цепью TCR; фиг. 111 - вестерн блоттинг растворимого TCR, экспрессированного в штамме SEY6210 S.cerevisiae. Дорожка С содержит 60 нг очищенного растворимого TCR NY-ESO, используемого в качестве контрольного образца. Дорожки 1 и 2 содержат белки, собранные из двух отдельных трансформированных дрожжевых культур TCR; фиг. 112 - последовательность нуклеиновых кислот KpnI к вставке EcoRI плазмиды рЕХ 172. Остаток плазмиды представляет собой pBlueScript II KS-; фиг. 113 - схематическое изображение цепей TCR для клонирования в бакуловирус; фиг. 114 - последовательность нуклеиновых кислот дисульфидного конструкта TCRA6 в форме вставки BamHI для инсерции в плазмиду экспрессии рАсАВ 3;-17 006601 фиг. 115 - дисульфидный конструкт TCRA6 в форме BamHI для инсерции в плазмиду экспрессии рАсАВ 3; фиг. 116 - гель, окрашенный кумасси, и вестерн блоттинг для полученного бактериальным путем дисульфидного TCR A6 и дисульфидного TCR A6 насекомого. Во всех следующих примерах, если не указано иного, полученные цепи растворимого TCR подвергают усечению непосредственно перед С-концевой областью цистеиновых групп, образующих нативную межцепочечную дисульфидную связь. Пример 1. Конструкция праймеров и мутагенез - и -цепей TCR А 6 Tax. Для мутации треонина 48 А 6 Tax экзона 1 в TRAC01 в цистеин сконструировали следующие праймеры (мутация показана строчными буквами): Для мутации серина 57 А 6 Tax экзона 1 в TRBC101 и TRBC201 в цистеин сконструировали следующие праймеры (мутация показана строчными буквами): Мутагенез PCR (polimerase chain reaction, полимеразная цепная реакция). Экспрессионные плазмиды, содержащие гены для - или -цепей TCR A6, мутировали с помощью праймеров -цепи или праймеров -цепи, соответственно, следующим образом. 100 нг плазмиды смешали с 5 мкл 10 мМ dNTP, 25 мкл буфера 10Pfu (Stratagene), 10 единицами полимеразы Pfu (Stratagene) и довели конечный объем водой до 240 мкл. В 48 мкл указанной смеси добавили праймеры и разбавили,получив конечную концентрацию 0,2 мкмоль в 50 мкл конечного объема реактива. После начального этапа денатурации в течение 30 с при 95 С провели 15 циклов денатурации смеси (95 С, 30 с), отжиг(55 С, 60 c) и элонгацию (73 С, 8 мин) в экспресс-установке PCR - Hybaid PCR. Затем продукт подвергли ферментации в течение 5 ч при 37 С с 10 единицами рестрикционного фермента DpnI (New England Biolabs). 10 мкл продукта реакции ферментации трансформировали в компетентные бактерии XL I-Blue и выращивали в течение 18 ч при 37 С. Отобрали единичную колонию и выращивали в течение ночи в 5 мл TYP + ампициллин (бактотриптон 16 г/л, дрожжевой экстракт 16 г/л, NaCl 5 г/л, K2 НРO4 2,5 г/л, ампициллин 100 мг/л). Плазмидную ДНК очистили в минипрепараторной колонке Qiagen согласно инструкциям изготовителя и проконтролировали последовательность с помощью автоматического секвенирования на оборудовании для секвенирования факультета биохимии Оксфордского университета. Соответствующие мутированные последовательности нуклеиновых кислот и аминокислот показаны на фиг. 2 а и 3 а для -цепи и фиг. 2b и 3b для -цепи. Пример 2. Экспрессия, повторная укладка и очистка растворимого TCR. Экспрессионные плазмиды, содержащие мутированную -цепь и -цепь, соответственно, трансформировали раздельно в штамм E.coli BL21pLysS, и вырастили отдельные ампициллин-резистентные колонии при 37 С в среде TYP (ампициллин 100 мкг/мл) до ОD600 0,4 перед индуцированном экспрессии белка при 0,5 мМ IPTG. Клетки отобрали через 3 ч после индукции путем центрифугирования в течение 30 мин со скоростью 4000 об./мин в центрифуге Beckman J-6B. Осадок, содержащий клетки, повторно суспендировали в буфере, содержащем 50 мМ Tris-HCl, 25% (мас./об.) сахарозы, 1 мМ NaEDTA (этилендиаминтетрауксуснокислого натрия), 0,1% (мас./об.) азида натрия, 10 мМ DTT (дитиотрентола), рН 8,0. После циклического замораживания - оттаивания в течение ночи повторно суспендированные клетки обработали ультразвуковыми импульсами длительностью 1 мин в течение примерно 10 мин в ультразвуковой установке Milsonix XL2020 с помощью стандартного зонда диаметром 12 мм. Осажденные тельца включения извлекли путем центрифугирования в течение 30 мин при 13000 об./мин в центрифуге Beckman J2-21. Затем для удаления дебриса и компонентов мембраны осадок промыли тремя порциями детергента. Каждый раз осадок телец включений гомогенизировали в буфере Triton (500 мМ Tris-HCl, 0,5%Triton-X100, 200 мМ NaCl, 10 мМ NaEDTA, 0,1% (мас./об.) азида натрия, 2 мМ DDT, рН 8,0) перед осаждением путем центрифугирования в течение 15 мин при 13000 об./мин в центрифуге Beckman J2-21. После этого детергент и соль удалили с помощью аналогичной промывки следующим буфером: 50 мМ TrisHCl, 1 мМ NaEDTA, 0,1% (мас./об.) азида натрия, 2 мМ DDT, рН 8,0. И, наконец, тельца включений разделили на аликвоты по 30 мг и заморозили при -70 С. Количественное определение выхода белка телец включения провели путем растворения в 6 М растворе гуанидин-НСl и анализа связывания с красителемBradford (PerBio). Произвели оттаивание примерно 30 мг (т.е. 1 мкмоль) растворенных телец включения каждой цепи из замороженных аликвот, затем перемешали образцы и разбавили смесь 15 мл раствора гуанидина (6 М гуанидинхлорида, 10 мМ ацетата натрия 10 мМ EDTA), чтобы обеспечить полную денатурацию цепи. Затем ввели раствор гуанидина, содержащий полностью восстановленные и денатурированные цепи TCR в 1 л следующего буфера для повторной укладки: 100 мМ Tris pH 8.5, 400 мМ L-аргинина, 2 мМ EDTA, 5 мМ восстановленного глутатиона, 0,5 мМ окисленного глутатиона, 5 М мочевины, 0,2 мМ PMSF (фе-18 006601 нилметансулфонилфторида). Раствор выдержали в течение 24 ч. После этого смесь для повторной укладки дважды подвергли диализу, вначале против 10 л 100 мМ мочевины, а затем против 10 л 100 мМ мочевины и 10 мМ Tris pH 8,0. Операции повторной укладки и диализа проводили при 6-8 С.sTCR отделили от продуктов разложения и загрязнений путем загрузки диализованного повторно уложенного продукта в анионообменную колонку POROS 50HQ и элюирования связанного белка с градиентом 0-400 мМ NaCl в 50 объемах колонки, используя очистительную систему Akta (Pharmacia), как показано на фиг. 4. Пиковые фракции до объединения и концентрирования хранили при 4 С и анализировали с помощью SDS-PAGE с окрашиванием кумасси (фиг. 5). И, наконец, sTCR очистили и проанализировали с помощью колонки Superdex 200HR гель-фильтрации (фиг. 6), предварительно сбалансировав в буфере HBS-EP (10 мМ HEPES pH 7,4, 150 мМ NaCl, 3,5 мМ EDTA, 0,05% nonidet p40). Фракции пика элюции с относительной молекулярной массой около 50 кДа объединили и сконцентрировали перед проведением анализа с помощью поверхностного плазменного резонанса на установке BIAcore. Пример 3. Исследование связывания sTCR со специфичным рМНС с помощью поверхностного плазменного резонанса на установке BIAcore. Поверхностный плазменный резонансный биосенсор (BIAcore 3000) использовали для анализа связывания sTCR с его лигандом пептид-МНС. Это обеспечили путем создания одиночных комплексов рМНС (описаны ниже), которые иммобилизовали на связывающей поверхности, покрытой стрептавидином, в полуориентированной форме, что позволило проводить эффективный контроль связывания растворимого Т-лимфоцитного рецептора одновременно с четырьмя различными рМНС (иммобилизованными в отдельных проточных кюветах). Ручное введение комплекса HLA (human leukocyte antigens,главный комплекс гистосовместимости человека) позволяет легко регулировать точный уровень иммобилизованных молекул класса I. Такие иммобилизованные комплексы способны связывать как Т-лимфоцитные рецепторы, так и корецептор CD8, при этом все они могут быть введены в растворимую фазу. Специфическое связываниеTCR получали даже при низких концентрациях (по меньшей мере 40 мкг/мл), что указывало на то, чтоTCR является относительно устойчивым. Наблюдения показали, что свойства sTCR связывать рМРС являются аналогичными с количественной и качественной точек зрения, если TCR используют в растворимой или иммобилизованной фазе. Это является важным средством контроля парциальной активности растворимых форм, и также указывает на то, что биотинилированные комплексы рМНС так же активны биологически, как и небиотинилированные комплексы. Биотинилированные пептидные комплексы HLA-A2 класса I повторно уложили in vitro из бактериально экспрессированных телец включения, содержащих конститутивные субъединицы белков и синтетический пептид, с последующей очисткой и ферментным биотинилированием in vitro (O'Callaghan et al.(1999) Anal. Biochem. 266: 9-15). Тяжелую цепь HLA, которая замещает трансмембранные и цитоплазматические домены белка в соответствующем конструкте, экспрессировали с биотинилированной меткой в С-концевой области. Получили уровни экспрессии телец включения около 75 мг/л бактериальной культуры. Легкую цепь HLA или 2-микроглобулин также экспрессировали в форме телец включений в E.coli из соответствующего конструкта на уровне примерно 500 мг/л бактериальной культуры. Клетки Е. coli лизировали, и очистили тельца включения до чистоты, равной примерно 80%. Белок из телец включений денатурировали в 6 М ганидин-HCl, 50 мМ Tris рН 8,1, 100 мМ NaCl, 10 мМ DTT, 10 мМ EDTA и произвели повторную укладку при концентрации 30 мг/л тяжелой цепи, 30 мг/л 2m в 0,4 МL-аргинин-HCl, 100 мМ Tris рН 8,1, 3,7 мМ циастамина, 6,6 мМ -цистеамина, 4 мг/мл пептида (например,tax 11-19), путем добавления одной порции денатурированного белка в буфер повторной укладки при температуре ниже 5 С. Повторную укладку удалось завершить при 4 С в течение по меньшей мере 1 ч. Буфер заменили путем диализа в 10 объемах 10 мМ Tris рН 8.1. Для существенного уменьшения ионной силы раствора потребовалось дважды заменить буфер. Затем раствор белка профильтровали через фильтр 1,5 мкм из ацетата целлюлозы и загрузили в анионообменную колонку POROS 50HQ (объем слоя 8 мл). Белок элюировал с линейным градиентом 0-500 мМ NaCl. Комплекс НLА-А 2-пептид элюировал примерно при 250 мМ NaCl. Отобрали пиковые фракции, добавили смесь ингибиторов протеазы(Calbiochem) и охладили фракции на льду. В комплексах HLA, меченых биотинилированием, замену буфера производили в 10 мМ Tris рН 8,1,5 мМ NaCl с помощью колонки быстрого обессоливания (Pharmacia), сбалансированной в том же самом буфере. Непосредственно после элюции фракции, содержащие белок, охладили на льду и добавили смесь ингибиторов протеазы (Calbiochem). Затем добавили агенты для биотинилирования: 1 мМ биотина, 5 мМ АТР (забуференного до рН 8), 7,5 мМ MgCl2 и 5 мкг/мл фермента BirA (очищенного согласно O'Callaghan et al. (1999) Anal. Biochem. 266: 9-15). После этого смесь выдержали при комнатной температуре в течение ночи. Биотинилированные комплексы HLA очистили с помощью гель-хроматографии. Колонку Superdex 75 HR 10/30 (Pharmacia) предварительно сбалансировали фильтрованным забуференным фосфатом физиологическим раствором (phosphate-buffered saline, PBS) и 1 мл реакционной смеси для биотинилирования и обработали колонну PBS при 0,5 мл/мин. Биотинилированные комплексы HLA элюировали, обра-19 006601 зуя один пик примерно при 15 мл. Фракции, содержащие белок объединили, охладили на льду и добавили смесь ингибиторов протеазы. Концентрацию белка определили способом связывания кумасси(PerBio). Аликвоты биотинилированных комплексов HLA хранили замороженными при -20 С. Стрептавидин иммобилизовали стандартными способами путем образования аминовой связи. Взаимодействия между sTCR Tax A6, содержащим новую межцепочечную связь, и его комплексом лиганд/МНС или посторонней комбинацией HLA- пептид, получение которых описано выше, анализировали с помощью поверхностного плазменного резонанса (SPR) на биосенсоре Biacore 3000. Измерения SPR основаны на изменении коэффициента преломления, выраженного в реактивных единицах (RU),вблизи поверхности сенсора в миниатюрной проточной кювете - принцип, который можно использовать для определения взаимодействий лиганда рецептора и анализа их сродства и кинетических параметров. Пробные проточные кюветы подготовили путем иммобилизации отдельных комплексов HLA-пептид в раздельных проточных кюветах посредством связывания между биотином, сшитым с 2m, и стрептавидином, который был химическим образом связан с активированной поверхностью проточных кювет. Затем провели анализ путем пропускания sTCR над поверхностью различных проточных кювет при постоянной скорости потока, измеряя при этом реакцию SPR. Вначале проверили специфичность взаимодействия путем пропускания sTCR при постоянной скорости потока, равной 5 мкл мин-1, над двумя различными поверхностями: одна с покрытием около 5000 RU специфичного комплекса пептид-HLA, вторая - с покрытием около 5000 RU неспецифичного комплекса пептид-HLA (фиг. 7, вставка) Введение растворимого sTCR над комплексом пептид-HLA при постоянной скорости потока и различных концентрациях использовали для определения фонового резонанса. Результаты этих контрольных измерений вычли из значений, полученных со специфичным комплексом пептид-HLA и использовали для расчета сродства к соединению, выраженного константой диссоциации, Кд (PriceDwek, Principles и Problems in PhysicalChemistry for Biochemists (2nd Edition) 1979, Clarendon Press, Oxford), как показано на фиг. 7. Полученное значение Кд (1,8 мкмоль) является близким к опубликованному в литературе для взаимодействия между TCR Tax A6 без новой дисульфидной связи и рМНС (0,91 мкмоль - Ding et al., 1999,Inmmunity 11:45-56). Пример 4. Получение растворимого TCR JM22, содержащего новую дисульфидную связь.-цепь растворимого TCR A6, полученного в примере 1, содержит в нативной последовательности рестрикционный сайт BglII (AAGCTT), пригодный для применения в качестве сайта лигирования. Мутагенез PCR выполнили, как подробно описано ниже, для введения рестрикционного сайтаBamH1 (GGATCC) в -цепь растворимого TCR A6, 5' нового цистеинового кодона. Последовательность,показанную на фиг. 2 а, применили в качестве матрицы для этого мутагенеза. Использовали следующие праймеры: 100 нг плазмиды смешали с 5 мкл 10 мМ dNTP, 25 мкл буфера 10Pfu (Stratagene), 10 единицамиPfu полимеразы (Stratagene) и довели конечный объем водой до 240 мкл. В 48 мкл указанной смеси добавили праймеры и разбавили, получив конечную концентрацию 0,2 мкмоль в 50 мкл конечного объема реактива. После начального этапа денатурации в течение 30 с при 95 С провели 15 циклов денатурации смеси (95 С, 30 с), отжиг (55 С, 60 c) и элонгацию (73 С, 8 мин) в экспресс-установке PCR - Hybaid PCR. Затем продукт подвергли ферментации в течение 5 ч при 37 С с 10 единицами рестрикционного фермента DpnI (New England Biolabs). 10 мкл продукта реакции ферментации трансформировали в компетентные бактерии XL1-Blue и выращивали в течение 18 ч при 37 С. Выбрали одну колонию и выращивали в течение ночи в 5 мл TYP + ампициллин (бактотриптон 16 г/л, дрожжевой экстракт 16 г/л, NaCl 5 г/л,K2 НРO4 2,5 г/л, ампициллин 100 мг/л). Плазмиду ДНК очистили в минипрепараторной колонке Qiagen согласно инструкциям изготовителя и проверили последовательность с помощью автоматического секвенирования на оборудовании для секвенирования факультета биохимии Оксфордского университета. Мутации, введенные в -цепь, были "молчащими", поэтому последовательность аминокислот этой цепи осталась такой же, как показано на фиг. 3 а. Последовательность ДНК для мутированной -цепи показана на фиг. 8 а. Для получения растворимого TCR JM22, содержащего новую дисульфидную связь, в качестве матрицы использовали плазмиды TCR A6, содержащие рестрикционный сайты -цепи BamHI и -цепиPCR провели с помощью вышеуказанных праймеров и матриц, содержащих цепи TCR JM22. ПродуктыPCR рестрикционно ферментировали и клонировали в pGMT7, чтобы получить экспрессионные плазмиды. Последовательность плазмидных вставок подтвердили автоматическим секвенированием ДНК. Фиг. 8b и 8 с показывают последовательность ДНК мутированных - и -цепей TCR JM22, соответственно, а фиг. 9 а и 9b - результирующие последовательности аминокислот. Соответствующие цепи TCR экспрессировали, повторно уложили совместно и очистили, как описано в примерах 1 и 2. На фиг. 10 показана элюция белка растворимого TCR JM22 с дисульфидной связью из колонки POROS 50HQ с использованием градиента 0-500 мМ NaCl, как показано пунктирной линией. На фиг. 11 показаны результаты восстанавливающего SDS-PAGE (с окрашиванием кумасси) и невосстанавливающего SDS-PAGE (с окрашиванием кумасси) гелей фракций из элюата из колонки, показанного на фиг. 10. Пик 1 определенно содержит гетродимер TCR, который присоединен межцепочечной дисульфидной связью. На фиг. 12 показана элюция белка из объединенных фракций пика 1 с фиг. 10. Анализ BIAcore связывания TCR JM22 с рМНС провели, как описано в примере 3. Фиг. 13 а показывает результат анализа способом BIAcore специфического связывания растворимого TCR JM22 с дисульфидной связью с комплексом HLA-Flu. Фиг. 13b показывает реакцию связывания по сравнению с контрольным образцом для одной инъекции растворимого TCR JM22 с дисульфидной связью. Значение Кд этого TCR с дисульфидной связью, для комплекса HLA-flu получили равным 7,90,51 мкмоль. Пример 5. Получение растворимого TCR NY-ESO содержащего новую дисульфидную связь. кДНК, кодирующую TCR NY-ESO, выделили из Т-лимфоцитов, полученных от Enzo Cerundolo(Институт молекулярной медицины Оксфордского университета), в соответствии с известной методикой. кДНК, кодирующую TCR NY-ESO, получили путем обработки мРНК обратной транскриптазой. Для получения растворимого TCR NY-ESO, содержащего новую дисульфидную связь, в качестве матрицы использовали плазмиды TCR A6, содержащие рестрикционные сайты -цепи BamHI и -цепи Конструкты - и -цепей TCR NY-ESO получили клонированием PCR следующим образом. Реакции PCR провели с помощью вышеуказанных праймеров и матриц, содержащих цепи TCR NY-ESO. Продукты PCR рестрикционно ферментировали и клонировали в pGMT7, чтобы получить экспрессирующие плазмиды. Последовательность плазмидных вставок подтвердили автоматическим секвенированием ДНК. Фиг. 14 а и 14b показывают последовательность ДНК мутированных - и -цепей TCR NYESO, соответственно, а фиг. 15 а и 15b - результирующие последовательности аминокислот. Соответствующие цепи TCR экспрессировали, совместно уложили повторно и очистили, как описано в примерах 1 и 2 с внесением следующих изменений в протокол. Денатурация растворимых TCR: произвели оттаивание 30 мг растворенного тельца включения цепи TCR и 60 мг растворенного тельца включения -цепи TCR из замороженных аликвот. Тельца включения разбавили до конечной концентрации 5 мг/мл в 6 М растворе гуанидина и добавили DTT (2 М маточный раствор) до конечной концентрации 10 мМ. Смесь выдержали при 37 С в течение 30 мин. Повторная укладка растворимых TCR: 1 л буфера для повторной укладки интенсивно перемешали при 5 С 3 С. Примерно за 5 мин до введения денатурированных цепей TCR добавили окислительновосстановительную пару 2-меркаптоэтиламина и цистамина (до конечных концентраций 6,6 мМ и 3,7-21 006601 мМ соответственно). Затем выдержали состав при перемешивании в течение примерно 15 ч 15 мин при 53 С, чтобы обеспечить повторную укладку белка. Диализ повторно уложенных растворимых TCR. Повторно уложенный TCR подвергли диализу в мембране Spectrapor 1 (Spectrum; продукт 132670) против 10 л 10 мМ Tris pH 8,1 при 53 С в течение 18-20 ч. По истечении указанного времени диализный буфер заменили на свежий 10 мМ Tris pH 8,1 (10 л) и продолжили диализ при 53 С в течение дополнительных 20-22 ч. Фиг. 16 иллюстрирует элюцию белка растворимого TCR NY-ESO, присоединенного дисульфидной связью, из колонки POROS 50HQ с использованием градиента 0-500 мМ NaCl, как показано пунктирной линией. На фиг. 17 показаны результаты восстанавливающего SDS-PAGE (с окрашиванием кумасси) и невосстанавливающего SDS-PAGE (с окрашиванием кумасси) гелей фракций из элюата из колонки, показанного на фигуре 16. Пики 1 и 2 определенно содержат гетеродимер TCR, который присоединен межцепочечной дисульфидной связью. Фиг. 18 показывает результаты гель-хроматографии объединенных фракций, представленных пиком 1 (А) и пиком 2 (В) на фиг. 17. Белок элюирует, образуя одинарный главный пик, соответствующий гетеродимеру. Анализ BIAcore для определения связывания TCR NY-ESO, присоединенного дисульфидной связью, с рМНС выполнили, как описано в примере 3. Фиг. 19 показывает результат анализа BIAcore специфичного связывания растворимого TCR NY-ESO, присоединенного дисульфидной связью, с комплексом HLA-NYESO. А. пик 1, В. пик 2. Значение Кд этого TCR, присоединенного дисульфидной связью, для комплекса HLA-NY-ESO получили равным 9,40,84 мкмоль. Пример 6. Получение растворимого TCR NY-ESO, содержащего новую дисульфидную связь и по меньшей мере одну из двух цистеиновых групп, необходимых для образования нативной дисульфидной межцепочечной связи. Для получения растворимого TCR NY-ESO, содержащего новую дисульфидную связь и по меньшей мере одну из двух цистеиновых групп, участвующих в образовании нативной дисульфидной межцепочечной связи, плазмиды, содержащие рестрикционные сайты BamHI -цепи и BglII -цепи, использовали в качестве основы, как описано в примере 4. Использовали следующие праймеры: Конструкты - и -цепей TCR NY-ESO получили клонировавшем PCR следующим образом. Реакции PCR провели с помощью вышеуказанных праймеров и матриц, содержащих цепи TCR NY-ESO. Продукты PCR рестрикционно ферментировали и клонировали в pGMT7, чтобы получить экспрессионные плазмиды. Последовательность плазмидных вставок подтвердили автоматическим секвенированием ДНК. Фиг. 20 а и 20b показывают последовательность ДНК мутированных - и -цепей TCR NY-ESO соответственно, а фиг. 21 а и 21b - результирующие последовательности аминокислот. Для получения растворимого TCR NY-ESO, содержащего ненативную дисульфидную межцепочечную связь и нативную дисульфидную межцепочечную связь, использовали ДНК, выделенную с помощью обоих вышеуказанных праймеров. Для получения растворимых TCR NY-ESO с ненативной дисульфидной межцепочечной связью и только одной из цистеиновых групп, участвующих в образовании нативной дисульфидной межцепочечной связи, использовали ДНК, выделенную с помощью одного из вышеуказанных праймеров совместно с соответствующим праймером из примера 5. Соответствующие цепи TCR экспрессировали, повторно уложили совместно и очистили, как описано в примере 5. Фиг. 22-24 показывают элюцию белка растворимого TCR NY-ESOcys cys (т.е. ненативно и нативной цистеиновой группы в обеих цепях), TCRcys (с ненативными цистеиновыми группами в обеих цепях и нативной цистеиновой группой только в -цепи), и TCRcys (с ненативными цистеиновыми группами в обеих цепях и нативной цистеиновой группой только в -цепи) из анионообменных колонок POROS 50HQ с использованием градиента 0-500 мМ NaCl, как показано пунктирной линией. Фиг. 25 и 26 показывают, соответственно, результаты восстанавливающего SDS-PAGE (с окрашиванием кумасси) и невосстанавливающего SDS-PAGE (с окрашиванием кумасси) гелей фракций TCR NY-ESOcyscys, TCRcys и TCRcys из элюатов из колонки, показанных на фиг. 22-24. На них четко видно образование гетеродимеров TCR, которые присоединены межцепочечной дисульфидной связью. Фиг. 27-29 показывают профили элюции белка, полученные в результате гель-хроматографии объединенных фракций TCR NY-22 006601ESOcys cys, TCRcys и TCRcys из элюатов ионообменных колонок, показанных на фиг. 22-24 соответственно. Белок элюирует как одинарный главный пик, соответствующий гетеродимеру TCR. Анализ BIAcore связывания sTCR с рМНС провели, как описано в примере 3. Фиг. 30-32 показывают результаты анализа BIAcore специфического связывания TCRcys cys, TCRcys и TCRcys NY-ESO,соответственно, с комплексом HLA-NYESO.TCRcys cys имел Кд 18,082,075 мкмоль, TCRcys имел Кд 19,242,01 мкмоль и TCRcys имел Кд 22,54,0692 мкмоль. Пример 7. Получение растворимого TCR АН-1.23, содержащего новую дисульфидную межцепочечную связь. кДНК, кодирующую TCR АН-1.23, выделили из Т-лимфоцитов, полученных от Hill Gaston (медицинское училище, больница Addenbrooke, Кембридж), в соответствии с известной методикой. кДНК, кодирующую TCR NY-ESO, получили путем обработки мРНК обратной транскриптазой. Для получения растворимого TCR АН-1.23, содержащего новую дисульфидную связь, плазмидыTCR, содержащие рестрикционные сайты BamHI -цепи и BglII -цепи, использовали в качестве каркаса, как описано в примере 4. Использовали следующие праймеры: Конструкты - и -цепей TCR АН-1.23 получили клонированием PCR следующим образом. Реакции PCR провели с помощью вышеуказанных праймеров и матриц, содержащих цепи TCR АН-1.23. Продукты PCR рестрикционно ферментировали и клонировали в pGMT7, чтобы получить экспрессионные плазмиды. Последовательность плазмидных вставок подтвердили автоматическим секвенированием ДНК. Фиг. 33 а и 33b показывают последовательность ДНК мутированных - и -цепей TCR АН-1.23 соответственно, а фиг. 34 а и 34b - результирующие последовательности аминокислот. Соответствующие цепи TCR экспрессировали, повторно уложили совместно и очистили, как описано в примере 5. Фиг. 35 иллюстрирует элюцию белка растворимого TCR АН-1.23, присоединенного дисульфидной связью, из колонки POROS 50HQ использованием градиента 0-500 мМ NaCl, как показано пунктирной линией. На фиг. 36 и 37 показаны результаты восстанавливающего SDS-PAGE (с окрашиванием кумасси) и невосстанавливающего SDS-PAGE (с окрашиванием кумасси) гелей соответственно, фракций из элюата из колонки, показанного на фиг. 35. Указанные гели определенно указывают на присутствие гетеродимера TCR, который присоединен межцепочечной дисульфидной связью. Фиг. 38 показывает профиль элюции из колонки Superdex 75 HR гель-хроматографии объединенных фракций из элюата анионообменной колонки, показанного на фиг. 35. Белок элюирует, образуя одинарный главный пик, соответствующий гетеродимеру. Пример 8. Получение растворимых TCR A6, содержащих новую дисульфидную межцепочечную связь в альтернативных позициях в иммуноглобулиновой области константного домена. Следующие эксперименты провели для выяснения возможности получения функциональных растворимых TCR, содержащих новую дисульфидную связь в области ммуноглобулина TCR в позиции, отличной от промежуточной позиции между треонином 48 экзона 1 в TRAC01 и серином 57 экзона 1 вTRBC101/TRBC201. Для мутации -цепи TCR A6 сконструировали следующие праймеры (числа в названиях праймеров относятся к позиции остатка аминокислоты, подлежащего мутации в экзоне 1 TRAC01, мутированные остатки указаны строчными буквами) Для мутации -цепи TCR А 6 сконструировали следующие праймеры (числа в названиях праймеров относятся к позиции остатка аминокислоты, подлежащего мутации в экзоне 1 TRBC201. Мутированные остатки указаны строчными буквами): Мутагенез PCR, амплификацию, лигирование и плазмидную очистку конструктовиTCR выполнили, как описано в примере 1, с помощью соответствующей комбинации вышеуказанных праймеров, чтобы получить растворимые TCR, содержащие новые дисульфидные межцепочечные связи между следующими парами аминокислот: На фиг. 39-58 показаны последовательности ДНК и аминокислот мутированных цепей ДНК и последовательности аминокислот мутированных цепей TCR A6, амплифицированных вышеуказнными праймерами. Кодоны, кодирующие мутированные цистеиновые группы, показаны выделением.-25 006601 Соответствующие цепи TCR экспрессировали, повторно уложили совместно и очистили, как описано в примере 5. После очистки в анионообменной колонке POROS 50HQ полученные белки пропустили на SDS-Page гелях, чтобы проверить образование растворимого белка с правильной повторной укладкой. Указанные гели проанализировали также, чтобы определить наличие или отсутствие в очищенном материале белка, содержащего дисульфидную связь, и обладающего требуемой молекулярной массой. При использовании данной системы бактериальной экспрессии исследуемые TCR, содержащие следующие новые дисульфидные межцепочечные связи, не образовали белка, содержащего дисульфидную связь, с соответствующей молекулярной массой, поэтому их исключили из дальнейших экспериментов. Однако существуют альтернативные прокариотические или эукариотические системы экспресии. Фиг. 59-64 показывают элюцию растворимых TCR, содержащих новые дисульфидные межцепочечные связи между следующими группами: Thr 48-Ser 57, Thr 45-Ser 77, Туr 10-Ser 17, Thr 45-Asp 59, Met 52-Gly 55 и Ser 15-Glu 15, соответственно, из анионообменной колонки POROS 200HQ с помощью градиента 0-500 мМ NaCl, как показано пунктирной линией. Фиг. 65-70 показывают результаты восстанавливающего SDS-PAGE (с окрашиванием кумасси) и невосстанавливающего SDS-PAGE (с окрашиванием кумасси) гелей соответственно, фракций из элюатов из колонки, показанных на фиг. 59-64. Гели определенно указывают на присутствие гетеродимеров TCR, которые присоединены межцепочечной дисульфидной связью. Фиг. 71-76 показывают профили элюции из колонки Superdex 200 HR гель-фильтрации объединенных фракций из элюатов анионообменной колонки, показанных на фиг. 59-64. Анализ BIAcore связывания TCR с рМНС выполнили, как описано в примере 3. На фиг. 77-82 представлены кривые BIAcore, демонстрирующие способность очищенных растворимых TCR связываться с комплексами рМНС HLA-A2 tax.Thr 48-Ser 57 имел Кд, равную 7,8 мкмоль, Thr 45-Ser 77 имел Кд, равную 12,7 мкмоль, Туr 10-Ser 17 имел Кд, равную 34 мкмоль, Thr 45-Asp 59 имел Кд, равную 14,9 мкмоль и Ser 15-Glu 15 имел Кд,равную 6,3 мкмоль. Met 52-Gly 55 мог связываться со своей естественной "мишенью" - комплексомHLA-A2 tax, однако, он связывался также аналогичным образом с "посторонней" мишенью - комплексомHLA-A2-NY-ESO (см. фиг. 81). Пример 9. Рентгеноструктурный анализ Т-лимфоцитного рецептора NY-ESO, присоединенного дисулъфдиной связью и специфичного для комплекса NY-ESO-HLA-А 2.dsTCR NY-ESO клонировали, как описано в примере 5, и экспрессировали следующим образом. Экспрессионные плазмиды, содержащие мутированные -цепь и -цепь соответственно, трансформировали раздельно в штамм E.coli BL21pLysS и вырастили отдельные ампициллин-резистентные колонии при 37 С в среде TYP (ампициллин 100 мкг/мл) до ОD600 0,7 перед индуцированном экспрессии белка при 0,5 мМ IPTG. Клетки отобрали через 18 ч после индукции путем центрифугирования в течение 30 мин со скоростью 4000 об./мин в центрифуге Beckman J-6B. Клеточный осадок повторно суспендировали в лизисном буфере, содержащем 10 мМ Tris-HCl рН 8,1, 10 мМ MgCl2, 150 мМ NaCl, 2 мМ DTT, 10% глицерина. На каждый литр бактериальной фигуры добавили 100 мкл лизоцима (20 мг/мл) и 100 мкл дезоксирибонуклеазы I (20 мкг/мл). После выдержки на льду в течение 30 мин бактериальную суспензию обработали ультразвуковыми импульсами длительностью 1 мин в течение 10 мин в ультразвуковой установке Milsonix XL2020 с помощью стандартного зонда диаметром 12 мм. Осажденные тельца включения извлекли путем центрифугирования в течение 30 мин при 13000 об./мин в центрифуге Beckman J2-21(4 С). Затем для удаления дебриса и компонентов мембраны осадок промыли тремя порциями промывочного буфера Triton (50 мМ Tris-HCl pH 8,1, 0,5% Triton-Х 100, 100 мМ NaCl, 10 мМ NaEDTA, 0,1%(мас./об.), 2 мМ DDT). Каждый раз осадок телец включения гомогенизировали в промывочном буфереTriton перед осаждением путем центрифугирования в течение 15 мин при 13000 об./мин в центрифугеBeckman J2-21. После этого детергент и соль удалили с помощью аналогичной промывки в буфере повторного суспендирования (50 мМ Tris-HCl pH 8,1, 100 мМ NaCl, 100 мМ NaCl, 10 мМ NaEDTA, 0,1%(мас./об.) азида натрия, 2 мМ DDT). И, наконец, тельца включения растворили в 6 М гуанидиновом буфере (6 М гуанидин-гидрохлорид, 50 мМ Tris pH 8,1,100 мМ NaCl, 10 мМ EDTA, 10 мМ DDT), разделили-26 006601 на аликвоты по 120 мг и заморозили при -70 С. Количественное определение телец включения провели путем растворения в 6 М растворе гуанидин-НСl и анализа соединения с красителем Bradford (PerBio). Примерно 60 мг (т.е. 2,4 мкмоль) замороженной растворенной -цепи смешали с 30 мг (т.е. 1,2 мкмоль) замороженной растворенной -цепи. Смесь TCR разбавили до конечного объема 18 мл 6 М гуанидиновым буфером и нагрели до 37 С в течение 30 мин, чтобы обеспечить полную денатурацию цепей. Затем ввели раствор гуанидина, содержащий полностью восстановленные и денатурированные цепиTCR, в 1 л холодного буфера повторной укладки (100 мМ Tris pH 8,1, 400 мМ L-аргинин-НСl, 2 мМEDTA, 6,6 мМ 2-меркаптоэтиламина, 3,7 мМ цистамина, 5 М мочевины) при перемешивании. Раствор выдержали в течение 5 ч в холодном помещении (53 С), чтобы обеспечить проведение повторной укладки. После этого повторно уложенный продукт подвергли диализу против 12 л воды в течение 18-20 ч,а затем - против 12 л 10 мМ Tris pH 8,1 в течение 18-20 ч (53 С). Для указанного процесса диализа использовали диализную мембрану Spectrapor 1 (изделие 132670, Spectrum Laboratories), которая обеспечивает отсечку молекулярной массы 6-8000 кДа. Диализованный белок профильтровали через фильтры с размером пор 0,45 мкм (Schleicher and Schuell,10 404012), установленные на фильтрационной установке Nalgene. Повторно уложенный TCR NY-ESO отделили от продуктов разложения и загрязнений путем загрузки диализованного повторно уложенного продукта в анионообменную колонку POROS 50HQ (Applied Biotech). Перед загрузкой белка колонку POROS 50HQ сбалансировали 10 объемами колонки буфера А (10 мМ Tris рН 8,1). Связанный белок элюировал с градиентом 0-400 мМ NaClI в 7 объемах колонки. Пиковые фракции (1 мл) анализировали с помощью денатурирующего SDS-PAGE, используя восстанавливающий и невосстанавливающий буфер для образца. Затем очистили пиковые фракции, содержащие гетеродимерный альфа-бета комплекс, с помощью гель-фильтрационной колонки Superdex 75HR,предварительно сбалансированной в 25 мМ MES, рН 6,5. Пиковый белок, элюирующий с относительной молекулярной массой около 50 кДа, объединили, сконцентрировали до 42 мг/мл в центробежных концентраторах Ultrafree (Millipore, изделиеUFV2BGC40) и хранили при -80 С. Кристаллизацию TCR NY-ESO выполнили способом висячей капли при 18 С, используя 1 мкл раствора белка (8,4 мг/мл) в 5 мМ Mes при рН 6,5, смешанного с эквивалентным объемом буфера для кристаллизации. Кристаллы появлялись в нескольких различных режимах при использовании буферовCrystal Screen (Hampton Research). Одинарные кубические кристаллы (100 мкм) выращивали в буфере,содержащем 30% PEG 4000, 0,1 М цитрата натрия с рН 5,6 и 0,2 М ацетата аммония, и использовали для определения структуры. Кристаллы TCR NY-ESO подвергли сверхбыстрому замораживанию и провели их дифракционный анализ в рентгеновских лучах на синхротроне Daresbury. Кристаллы дифрагировали до разрешения 0,25 нм (2,5 ). Полученный набор данных обработали и получили 98,6% полного набора амплитуд, приемлемых примерно до 0,27 нм (2,7 ), но пригодных для применения (расчета) до 0,25 нм (2,5 ). Объединенный R-фактор, т.е. согласованность между множеством изменений кристаллографически эквивалентных отражений, составлял 10,8% для всех данных. Это является минимальным для самой высокой области разрешения. Установили, что пространственная группа соответствует P21 с размерами ячейки а=4,25 нм (42,5 ), b=5,95 нм (59,5 ), с=8,17 нм (81,7 ), =91,5. Размеры ячейки и симметрия указывают на наличие в ячейке двух копий. Асимметричный элемент, аu или минимальный объем, который подлежит исследованию, содержит только одну молекулу, а другая молекула в ячейке образуется в результате симметрии 21. Позиционирование молекулы в направлении оси у в аu является произвольным. До тех пор пока она находится в правильной позиции в плоскости x-z, ее можно произвольно транслировать в направлении оси у. Это называется свободным параметром в данной "полярной" пространственной группе. База данных PDB имеет только одну позицию, содержащую гетеродимерный TCR А/В, а именно 1BD2. Эта позиция имеет также координаты HLA-родственного пептида в комплексе с TCR. Цепь В TCR была такой же, как в NY-ESO, в то время как цепь А имела небольшие отличия в домене С и существенные отличия в домене N. При использовании модели 1BD2 А/В для молекулярного замещения (molecularreplacement, MR) получили некорректное разрешение, что показало обширное перекрытие молекул с эквивалентной симметрией. Использование только цепи В дало лучшее разрешение, при котором отсутствовали существенные коллизии с соседями. Коэффициент корреляции был равен 49%, кристаллографический R-фактор - 50%, и минимальное приближение (центра тяжести к центру тяжести) составляло 0,49 нм (49 ). К цепи А применили операции вращения и трансляции, необходимые для трансформации начальной модели цепи В в эквивалентное MR. Полученное при этом гибридное разрешение MR хорошо укладывалось в ячейку при минимальных коллизиях. Карты электронной плотности в целом согласовывались с моделью и позволяли производить ее корректировку для получения соответствия с последовательностью TCR NY-ESO. Однако исходная модель имела множество разрывов, в частности, в ней отсутствовали боковые цепи, что характеризует плохое упорядочение частей модели. Многие петли в форме шпильки (для волос) в промежутках между нитями имели низкую плотность и были сложны для моделирования. Кристаллографический R-фактор мо-27 006601 дели был равен 30%. Этот R-фактор является остаточным, т.е. представляет собой разность между расчетной и наблюдаемой амплитудами. Как показывают фиг. 83 а и 83b, входная последовательность из 1BD2 не очень хорошо соответствует плотности. Изменение модели на Cys в позициях 164 в цепи А и 174 в цепи В с последующим дополнительным усовершенствованием однозначно показало, что такое задание последовательности гораздо лучше согласуется с плотностью. При этом отличия размеров боковой цепи являются минимальными,что привело к небольшим пертурбациям в модели. Электронная плотность в этой области имела также небольшие изменения. Наиболее важный аспект настоящей работы состоит в том, что новый TCR очень похож по структуре на опубликованную модель (1BD2). Сравнение включало весь TCR, константные домены или мелкие фрагменты вблизи точки мутации. Отклонения значений параметров r.m.s представлены в приведенной ниже таблице. Сравнение структур показано на фиг. 84. Короткий участок означает одинарную нить цепи А (от А 157 до А 169) и одинарную нить цепи В (от В 170 до В 183), которые соединены дисульфидным мостиком. Отклонения рассчитаны только для главных атомов цепей. Эти результаты подтверждают, что введение дисульфидной связи оказывает минимальное влияние на локальную структуру в зоне связи. Несколько более существенное влияние наблюдается при сравнении TCR с опубликованной структурой (1BD2) TCR A6, однако, увеличение смещения RMS в большой степени вызвано отличиями петлевых конформаций (см. фиг. 84). Эти петли не образуют части структуры ядра TCR, которая формируется последовательностью -складок, создающих характерную укладку иммуноглобулина. Отклонение RMS для целой -цепи особенно велико, что вызвано различием последовательности вариабельных доменов в A6 (1BD2) и TCR NY-ESO. Однако A6 и TCR NY-ESO имеют одинаковый вариабельный -домен, и отклонения RMS для целой -цепи показывают, что структура этого вариабельного домена сохраняется также в TCR с новой дисульфидной связью. Таким образом, полученные данные показывают, что структура ядра TCR сохраняется в кристаллической структуре TCR с новой дисульфидной связью. Пример 10. Получение растворимых TCR NY-ESO, содержащих новую дисульфидную межцепочечную связь и меченые сайты в С-концевой области -цепи. Для получения растворимого TCR NY-ESO, содержащего новую дисульфидную связь, в качестве основы использовали плазмиды TCR A6, содержащие рестрикционные сайты BamHI -цепи и BglII цепи, как описано в примере 4. Конструкции -цепи TCR NY-ESO получили клонированием PCR следующим образом. РеакцииPCR провели с помощью указанных ниже праймеров и матриц, содержащих цепи TCR NY-ESO. Продукты PCR рестрикционно ферментировали и клонировали в pGMT7, содержащий биотиновую последовательность узнавания, чтобы получить экспрессионные плазмиды. Последовательность плазмидных вставок подтвердили автоматическим секвенированием ДНК. Фиг. 85 а показывает последовательность ДНК -цепи TCR NY-ESO, содержащую биотниновый сайт узнавания, а фиг. 85b - результирующую последовательность аминокислот. Конструкт -цепи получили, как описано в примере 5. Соответствующие цепи TCR экспрессировали, повторно уложили совместно и очистили, как описано в примере 5. Для получения растворимого TCR NY-ESO, содержащего ненативную дисульфидную межцепочечную связь и гексагистидиновую метку в С-концевой области -цепи, использовали такие же праймеры и матрицы NY-ESO, как указано выше. Продукты PCR рестрикционно ферментировали и клонировали вpGMT7, содержащий гексагистидиновую последовательность, чтобы получить экспрессирующие плазмиды. Фиг. 86 а показывает последовательность ДНК -цепи TCR NY-ESO, содержащую гексагистидиновую метку, а фиг. 86b - результирующую последовательность аминокислот.-28 006601 Фиг. 87 показывает элюцию растворимого TCR NY-ESO, содержащего новую дисульфидную связь и последовательность узнавания биотина из анионообменной колонки POROS 50HQ, с помощью градиента 0-500 мМ NaCl, как указано пунктирной линией. Фиг. 88 иллюстрирует элюцию растворимого TCRNY-ESO, содержащего новую дисульфидную связь и гексагистидиновую метку, из анионообменных колонок POROS 50HQ с помощью градиента 0-500 мМ NaCl, как указано пунктирной линией. Фиг. 89 и 90 представляют собой профили гель-хроматографии объединенных фракций элюатовNY-ESO, меченых биотином и гексагистидином из анионообменной колонки и показанных на фиг. 87 и 88 соответственно. Белок элюирует, образуя одинарный основной пик, соответствующий гетеродимеруTCR. Анализ BIAcore sTCR, связанного с рМНС, выполнили, как описано в примере 3. TCR NY-ESO, меченый биотином, имел Кд, равную 7,5 мкМ, TCR NY-ESO, меченый гексагистидином, имел Кд, равную 9,6 мкМ. Пример 11. Окрашивание клеток с использованием флуоресцентных меченых тетрамеров растворимого TCR NY-ESO, содержащего новую дисульфидную межцепочечную связь. Получение тетрамера TCR Растворимые TCR NY-ESO, содержащие новую дисульфидную связь и биотиновую последовательность узнавания, полученные, как описано в примере 10, использовали для получения тетрамеров растворимых TCR, которые требуются для окрашивания клеток. Произвели буферную замену 2,5 мл раствора очищенного растворимого TCR (0,2 мг/мл) в буфере реакции биотинилирования (50 мМ Tris pH 8,0,10 мМ MgCl2) с помощью колонки PD-10 (Pharmacia). Элюат (3,5 мл) сконцентрировали до 1 мл с помощью концентратора Centricon (Amicon) с отсечкой молекулярной массы 10 кДа. Эту операцию выполняли до получения 10 мМ при добавлении АТР из маточного раствора (0,1 г/мл, откорректированное значение рН 7,0). Затем добавили смесь ингибиторов протеазы (набор 1 смеси ингибиторов протеазы, Calbiochem Biochemicals) в объеме, достаточном для получения конечной концентрации смеси ингибиторов протеазы, равной 1/100 маточного раствора в состоянии поставки, а после этого - 1 мМ биотина (из 0,2 М маточного раствора) и 20 мкг/мл фермента (из 0,5 мг/мл маточного раствора). Смесь выдержали в течение ночи при комнатной температуре. Удалили из раствора избыток биотина с помощью гельхроматографии в колонке S75 HR. Уровень биотинилирования TCR NY-ESO определили способом жидкостной хроматографии высокого разрешения (HPLC), как описано ниже. Аликвоту 50 мкл биотинилированного TCR NY-ESO (2 мг/мл) выдержали с 50 мкл гранул агарозы, покрытых стрептовидином(Sigma), в течение 1 ч. Затем гранулы осадили центрифугированием и 50 мкл несвязанного образца пропустили через колонку TSK 2000 SW (Tosoohaas) при скорости потока 0,5 мл/мин (200 мМ фосфатный буфер с рН 7,0) в течение 30 мин. Присутствие биотинилированного TCR NY-ESO определяли с помощью ультрафиолоетового спектрофотометра при длине волны 214 и 280 нм. Биотинилированный NYESO пропускали против контрольного образца небиотинилированного TCR NY-ESO. Процент биотинилирования рассчитывали как разность площадей пиков биотинилированного и небиотинилированного белка. Тетрамеризацию биотинилированного растворимого TCR проводили с помощью нейтравидинфикоэритринового (ФЭ) конъюгата (Cambridge Biosciences, UK). Концентрацию биотинилированного растворимого TCR определяли способом окрашивания белка кумасси (Pierce), и рассчитали содержание нейтравидин-фикоэритринового конъюгата 0,8 мг/мг растворимого TCR, чтобы обеспечить насыщение нейтравидин-ФЭ биотинилированным TCR в отношении 1:4. 19,5 мкл раствора с концентрацией 6,15 мг/мл биотинилированного растворимого TCR NY-ESO в забуференном фосфатом физиологическом растворе (phosphate buffered saline, PBS) медленно добавили к 150 мкл растворимого нейтравидин-ФЭ конъюгата на льду при легком перемешивании. Затем к этому раствору добавили 100,5 мкл PBS, чтобы получить конечную концентрацию тетрамера TCR NY-ESO 1 мг/мл. Протокол окрашивания Четыре аликвоты 0,3106 HLA-A2 положительных В-клеток линии (РР LCL), трансформированных(SLLMWITQC) HLA-A2 NYESO в течение 2 ч при 37 С. Затем дважды промыли эти клетки РР LCL в сбалансированном солевом растворе Хенкса (Hank's buffered Saline solution, HBSS) (Gibco, UK). Каждую из четырех аликвот разделили на равные части и окрасили биотинилированным, свежететрамеризованным нейтравидин-фикоэритрином TCR NY-ESO, присоединенным дисульфидной связью. Клетки инкубировали с 5 или 10 мкг фикоэритрина, меченого тетрамерными комплексами dsTCR, на льду в течение 30 мин и промыли HBSS. Затем клетки промыли еще раз, повторно суспендировали вHBSS и проанализировали с помощью системы FACS Vantage. Отобрали 25,000 единиц и проанализировали данные с помощью программного обеспечения WinMIDI. Результаты На фиг. 91a-h показаны гистограммы данных, полученных при анализе FACSVantage для каждого образца, приготовленного, как описано выше. В следующей таблице указано процентное содержание положительно окрашенных клеток, наблюдавшихся в каждом из образцов Эти данные определенно показывают, что увеличение доли клеток, меченых тетрамерами TCR NYESO, коррелирует с концентрацией пептида (SLLMWITQC), в котором они были культивированы. Таким образом, указанные тетрамеры TCR NY-ESO представляют собой компоненты, пригодные для специфического мечения клеток на основании экспрессии комплекса HLA-A2 NY-ESO. В данном примере использовали флуоресцентный конъюгированный тетрамер TCR NY-ESO. Однако аналогичных уровней связывания клеток можно ожидать, если эту метку заменить пригодным терапевтическим компонентом. Пример 12. Получение растворимого TCR A6 с новой дисулъфидной связью, содержащего константную область С 1. Все предшествующие примеры описывают получение растворимых TCR с новой дисульфидной связью, содержащих константную область С 2. Настоящий пример показывает, что растворимые TCR,содержащие константную область С 1, также могут быть успешно получены. Конструкции праймеров для сращивания с помощью PCR вариабельного домена -цепи TCR А 6 с С 1 Для конструкта PCR вариабельного домена -цепи TCR A6 создали следующие праймеры: Для конструкта PCR C1 создали следующие праймеры: Конструкт -VTCR и конструкт С 1 получили отдельно путем амплификации вариабельного и постоянного домена каждой цепи с использованием стандартной технологии PCR. Их взаимно соединили с помощью сращивания PCR (stitching-PCR). Плазмиду ДНК очистили в минипрепараторной колонкеQiagen согласно инструкциям изготовителя и проконтролировали последовательность с помощью автоматического секвенирования на оборудовании для секвенирования факультета биохимии Оксфордского университета. Последовательность А 6+С 1 показана на фиг. 92. Затем цепь А 6+С 1 спарили с -цепью TCR A6 с помощью межцепочечной дисульфидной связи после введения цистеина в С-область обеих цепей. Растворимый TCR экспрессировали и повторно уложили, как описано в примере 2. Очистка повторно уложенного растворимого TCRsTCR отделили от продуктов разложения и загрязнений путем загрузки диализованного повторно уложенного продукта в анионообменную колонку POROS 50HQ и элюирования связанного белка с градиентом 0-400 мМ NaCl в 50 объемах колонки, используя очистительную систему Akta (Pharmacia), как показано на фиг. 4. Пиковые фракции до объединения и концентрирования хранили при 4 С и анализировали с помощью SDS-PAGE с окрашиванием кумасси (фиг. 94). И, наконец, sTCR очистили и проанализировали с помощью колонки Superdex 200HR гель-фильтрации (фиг. 95), предварительно сбалансировав ее в буфере HBS-EP (10 мМ HEPES рН 7,4, 150 мМ NaCl, 3,5 мМ EDTA, 0,05% nonidet p40). Фракции пика элюции с относительной молекулярной массой около 50 кДа объединили и сконцентрировали перед проведением анализа с помощью поверхностного плазменного резонанса на установке BIAcore. Анализ BIAcore соединения TCR A6 с дисульфидной связью с рМНС выполнили, как описано в примере 3. На фиг. 96 показан результат анализа BIAcore специфического связывания растворимого TCR
МПК / Метки
МПК: C07K 14/725, A61K 38/17, G01N 33/68
Метки: рецептор, растворимый, клетки
Код ссылки
<a href="https://eas.patents.su/30-6601-rastvorimyjj-receptor-t-kletki.html" rel="bookmark" title="База патентов Евразийского Союза">Растворимый рецептор т- клетки</a>
Растворимый гетеродимерный рецептор цитокина

Номер патента: 5496
Опубликовано: 24.02.2005
Авторы: Новак Джулия Э., Чандрасехер Ясмин А., Зу Венфенг, Джасперс Стивен Р., Фостер Доналд К.
МПК: C07K 14/00, C12N 15/00
Метки: растворимый, цитокина, рецептор, гетеродимерный
Формула / Реферат:
1. Выделенный растворимый рецептор, состоящий из субъединицы IL-22R и субъединицы IL-20RB, где субъединица IL-22R состоит из полипептида, имеющего аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 12, 13, 25, 26, 31 и 32, а субъединица IL-20RB состоит из полипептида, имеющего аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO 15-21, 23, 28, 29, 34 и 35. 2. Растворимый рецептор по п.1, где...
Применение лиганда мнс класса ii, представляющего собой lag-3 или производное, мутантную форму или растворимый фрагмент lag-3, для приготовления иммунотерапевтического лекарственного средства

Номер патента: 3740
Опубликовано: 28.08.2003
Автор: Триебель Фредерик
МПК: C12N 15/63, A61K 39/00, A61P 37/02...
Метки: средства, мнс, лиганда, фрагмент, растворимый, лекарственного, мутантную, форму, приготовления, иммунотерапевтического, lag-3, собой, класса, применение, производное, представляющего
Формула / Реферат:
1. Применение лиганда МНС класса II, способного усиливать антигенспецифичный иммунный ответ, в сочетании с антигеном, способным индуцировать указанный антигенспецифичный иммунный ответ, для производства лекарственного средства, предназначенного для профилактики или лечения патологических состояний, связанных с индукцией и усилением антигенспецифичного иммунного ответа, где лиганд МНС класса II представляет собой LAG-3 или производное, мутантную...
Рецептор baff (bcma), иммунорегуляторный агент

Номер патента: 4635
Опубликовано: 24.06.2004
Авторы: Амброуз Кристин, Маккэй Фабьенн, Чопп Юрг, Шнайдер Паскаль, Браунинг Джеффри, Томпсон Джеффри
МПК: A61K 38/17, C12N 15/12, C07K 14/715...
Метки: иммунорегуляторный, bcma, рецептор, агент
Формула / Реферат:
1. Способ ингибирования роста B-клеток у животного, включающий в себя стадию введения терапевтически эффективного количества композиции, содержащей соединение, выбранное из группы, состоящей из (a) полипептида BCMA или его фрагмента; (b) химерной молекулы, содержащей полипептид BCMA или его фрагмент, слитый с гетерологичной аминокислотной последовательностью; и (c) гомолога антитела против BCMA. 2. Способ ингибирования продукции иммуноглобулина...
Димерные тромбопоэтиновые пептидные миметики, связывающие мpl рецептор и имеющие тромбопоэтичеcкую активность

Номер патента: 3998
Опубликовано: 25.12.2003
Авторы: Лиу Чуан-Фа, Фейдже Улрих, Читхэм Дженет
МПК: A61K 38/19, A61P 7/06, C07K 14/52...
Метки: пептидные, связывающие, активность, имеющие, миметики, тромбопоэтиновые, димерные, тромбопоэтичеcкую, рецептор
Формула / Реферат:
1. Соединение, которое связывается с рецептором mpl, имеющее структуру TMP1(L1)n-TMP2, где TMP1 и TMP2, каждый независимо друг от друга, выбран из группы коровых соединений, имеющих структуру X2 - X3 - X4 - X5 - X6 - X7 - X8 - X9 - X10, причем X2 выбран из группы, включающей Glu, Asp, Lys и Val; X3 выбран из группы, включающей Gly и Ala; X4 представляет собой Pro; X5 выбран из группы, включающей Thr и Ser; X6 выбран из группы, включающей...
Рецептор нейротрофного фактора, полученного из глиальной клеточной линии

Номер патента: 2924
Опубликовано: 31.10.2002
Авторы: Уен Дуанжи, Джинг Шукьян, Фокс Гэри М.
МПК: A61K 38/17, C07K 14/71, C12N 15/12...
Метки: нейротрофного, рецептор, клеточной, глиальной, фактора, линии, полученного
Формула / Реферат:
1. Выделенный и очищенный белок рецептора нейротрофного фактора человека, полученного из глиальной клеточной линии (GDNFR), имеющий аминокислотную последовательность SEQ ID No: 2, приведенную на фиг. 2, или его аналог, полученный исключением, дополнением и/или заменой аминокислот в указанной последовательности при сохранении конечной молекулой активности GDNFR, причем указанный белок способен к образованию комплекса с нейротрофным фактором,...
Предыдущий патент: Фармацевтические композиции фибринолитического агента
Следующий патент: Применение стафилококковых и энтерококковых вакцин для защиты иммунокомпромиссного субъекта от бактериальных инфекций
Случайный патент: Гербицидно активные бициклические 1,3-дионы