Стрептококковые антигены
Номер патента: 6232
Опубликовано: 27.10.2005
Авторы: Бродеур Бернард, Хамел Джози, Оуиллет Катерина, Чарлэнд Натали, Мартин Денис







Формула / Реферат
1. Изолированный полинуклеотид, включающий полинуклеотид, выбранный из
(а) полинуклеотида, кодирующего полипептид, обладающий по крайней мере 95% идентичностью со вторым полипептидом, выбранным из таблицы B, E или H;
(б) полинуклеотида, кодирующего полипептид, обладающий последовательностью аминокислот, выбранной из таблицы B, E или H;
(в) полинуклеотида, кодирующего полипептид, способный вырабатывать антитела, обладающие специфическим связыванием с полипептидом, имеющим последовательность, выбранную из таблицы B, E или H;
(г) полинуклеотида, кодирующего несущий эпитоп участок полипептида, выбранного из таблицы B, E или H; а также
(д) полинуклеотида, комплементарного полинуклеотиду из пп.(а), (б), (в) или (г).
2. Изолированный полинуклеотид по п.1, отличающийся тем, что указанный полинуклеотид представляет собой (а).
3. Изолированный полинуклеотид по п.1, отличающийся тем, что указанный полинуклеотид представляет собой (б).
4. Изолированный полинуклеотид по п.1, отличающийся тем, что указанный полинуклеотид представляет собой (в).
5. Изолированный полинуклеотид по п.1, отличающийся тем, что указанный полинуклеотид представляет собой (г).
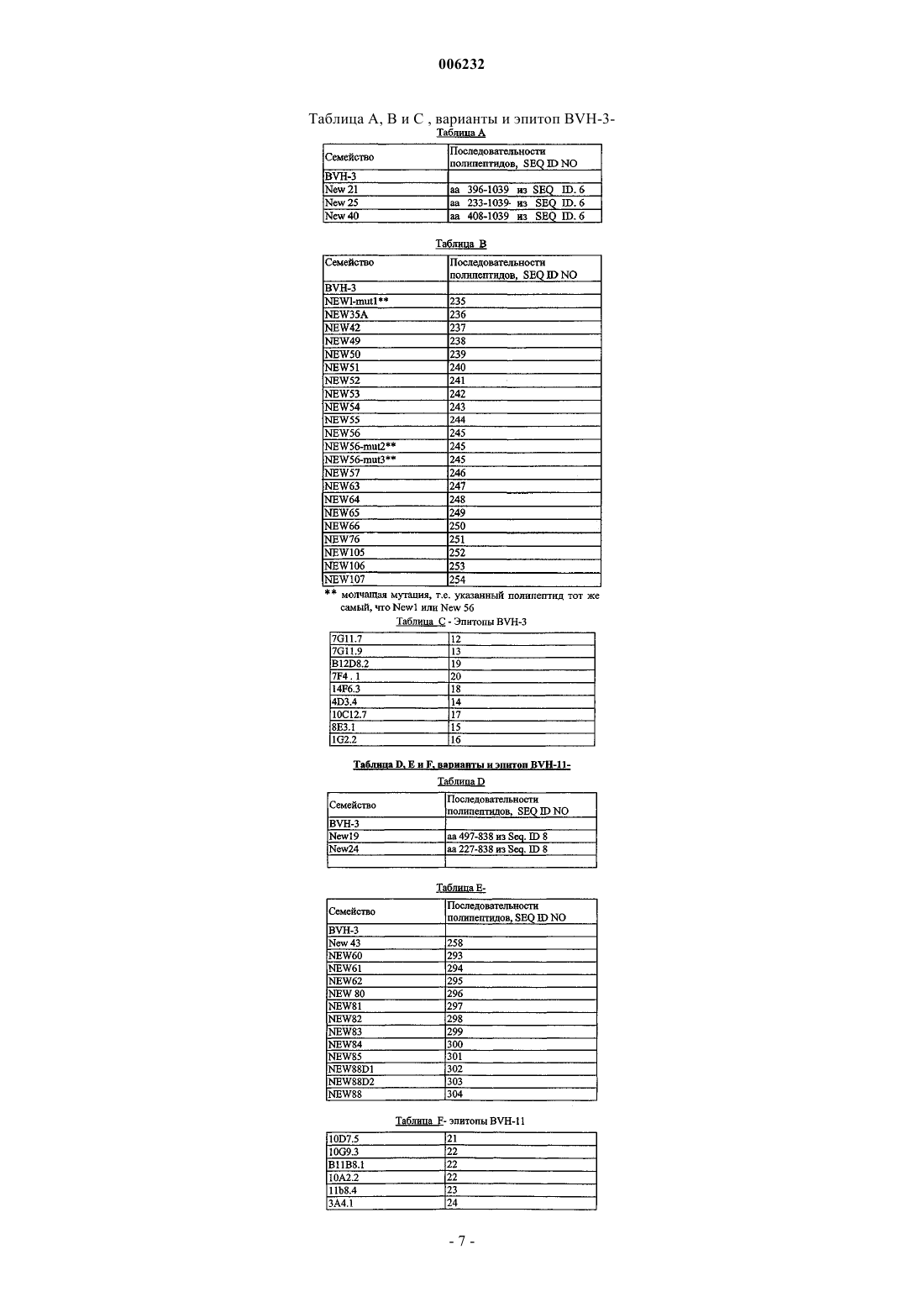
6. Изолированный полинуклеотид по п.5, отличающийся тем, что указанный полинуклеотид выбирают из таблицы B.
7. Изолированный полинуклеотид по п.6, отличающийся тем, что указанный участок, несущий эпитоп, выбирают из таблицы C.
8. Изолированный полинуклеотид по п.5, отличающийся тем, что указанный полинуклеотид выбирают из таблицы E.
9. Изолированный полинуклеотид по п.8, отличающийся тем, что указанный участок, несущий эпитоп, выбирают из таблицы F.
10. Полинуклеотид по любому из пп.1-9, отличающийся тем, что указанный полинуклеотид представляет собой ДНК.
11. Полинуклеотид по любому из пп.1-9, отличающийся тем, что указанный полинуклеотид представляет собой РНК.
12. Вектор, содержащий полинуклеотид по п.10, отличающийся тем, что указанная ДНК реально связана с областью контролируемой экспрессии.
13. Клетка-хозяин, подверженная трансфекции вектором по п.12.
14. Процесс продуцирования полипептида, включающий культивирование клетки-хозяина по п.13 в условиях, пригодных для экспрессии указанного полипептида.
15. Химерный полипептид, включающий элемент, выбранный из
(а) полипептида, обладающего по крайней мере 70% идентичностью со вторым полипептидом, имеющим последовательность аминокислот, выбранную из таблицы B, E или H;
(б) полипептида, обладающего по крайней мере 95% идентичностью со вторым полипептидом, имеющим последовательность аминокислот, выбранную из таблицы B, E или H;
(в) полипептида, обладающего последовательностью аминокислот, выбранной из таблицы B, E или H;
(г) полипептида, обладающего последовательностью аминокислот, выбранной из таблицы B, E или H;
(д) полипептида, способного вырабатывать антитела, обладающие специфическим связыванием со вторым полипептидом, имеющим последовательность, выбранную из таблицы B, E или H;
(е) несущего эпитоп участка полипептида, обладающего последовательностью аминокислот, выбранной из таблицы B, E или H;
(ж) полипептида из (а), (б), (в), (г), (д) или (е), в котором N-концевой метиониновый остаток претерпел делецию; или
(з) полипептида из (а), (б), (в), (г), (д) или (е), в котором секреторная последовательность аминокислот претерпела делецию.
16. Полипептид по п.15, отличающийся тем, что указанный полипептид представляет собой (е).
17. Полипептид по п.16, отличающийся тем, что указанный полипептид выбирают из таблицы B.
18. Полипептид по п.17, отличающийся тем, что указанный участок, несущий эпитоп, выбирают из таблицы C.
19. Полипептид по п.16, отличающийся тем, что указанный полипептид выбирают из таблицы E.
20. Полипептид по п.19, отличающийся тем, что указанный участок, несущий эпитоп, выбирают из таблицы F.
21. Химерный полипептид, содержащий два или более полипептидов, выбранных из таблицы B, E или H при условии, что эти полипептиды соединены для образования химерного полипептида.
22. Состав вакцины, содержащей полипептид по любому из пп.15-21, а также фармацевтически приемлемый носитель, разбавитель или адъювант.
23. Способ терапевтического или профилактического лечения менингита, среднего отита, бактериемической или пневмонийной инфекции у субъекта, восприимчивого к менингиту, среднему отиту, бактериемической или пневмонийной инфекции, включающий введение указанному субъекту терапевтического или профилактического количества состава по п.22.
24. Способ терапевтического или профилактического лечения стрептококковой бактериальной инфекции у субъекта, восприимчивого к стрептококковой бактериальной инфекции, включающий введение указанному субъекту терапевтического или профилактического количества состава по п.22.
25. Способ по п.23, отличающийся тем, что указанным субъектом является млекопитающее.
26. Способ по п.24, отличающийся тем, что указанным субъектом является млекопитающее.
27. Способ по п.23, отличающийся тем, что указанным субъектом является человек.
28. Способ по п.24, отличающийся тем, что указанным субъектом является человек.
29. Способ по п.24, отличающийся тем, что указанная бактериальная инфекция представляет собой S.pneumoniae группу A стрептококк (pyogenes), группу B стрептококк (GBS или agalactiae), dysgalactiae, uberis, nocardia, или золотистый стафилококк.
30. Способ по п.24, отличающийся тем, что указанная бактериальная инфекция представляет собой S.pneumoniae
31. Применение состава вакцины по п.22 для профилактического или терапевтического лечения стрептококковой бактериальной инфекции у животного, восприимчивого к стрептококковой бактериальной инфекции или инфицированного ей, включающее введение указанному животному профилактического или терапевтического количества этого состава.
Текст
006232 Область применения Настоящее изобретение касается антигенов, эпитопов и антител, специфических для этих эпитопов,в частности, полипептидных антигенов стрептококковой патогенной пневмонии, которые могут быть полезными для профилактики, диагностики или лечения стрептококковой инфекции. Предпосылки изобретенияS. pneumoniae представляет собой важный возбудитель заболевания людей, особенно детей, пожилых людей и лиц с нарушениями иммунитета. Эти бактерии часто выделяют у пациентов с инвазивными заболеваниями, типа бактериемической/септической пневмонии и менингита, которые во всем мире характеризуются высокой заболеваемостью и смертностью. Пневмококковые инфекции даже при соответствующей антибиотикотерапии по-прежнему являются причиной многих смертей. Несмотря на то, что появление противомикробных лекарственных средств снизило общую смертность от пневмококковых заболеваний, главной проблемой в мире в настоящее время стало существование резистентных пневмококковых организмов. Эффективные пневмококковые вакцины могут оказать значительное влияние на заболеваемость и смертность, вызванные S. pneumoniae. Такие вакцины должны также быть пригодны,чтобы предотвратить развитие среднего отита у детей раннего и младшего возраста. Обычно усилия по созданию пневмококковой вакцины концентрировались на выработке иммунной реакции на капсульный пневмококковый полисахарид. Более 80 пневмококковых капсульных серотипов было проидентифицировано по принципу антигенных различий. Доступная в настоящее время пневмококковая вакцина, которая содержит 23 капсульных полисахарида, наиболее часто являющихся причиной заболевания, обладает существенными недостатками. Они касаются, в первую очередь, низкой иммуногенности некоторых капсульных полисахаридов, разнообразия серотипов и отличиями в распределении серотипов по времени, географическим зонам и возрастным группам. В частности, неспособность вакцин, существующих в настоящее время, а также разрабатываемых капсульных сопряженных вакцин защитить детей младшего возраста от всех серотипов, стимулировала разработку других компонентов S.pneumoniae. Хотя иммуногенность капсульных полисахаридов можно повысить, но главным ограничением для вакцин на основе полисахаридов будет по-прежнему представлять собой специфичность серотипа. Применение антигенно-консервативного иммуногенного антигена пневмококкового белка или в чистом виде, или в сочетании с другими дополнительными компонентами, предполагает возможность создания пневмококковой вакцины на основе белка. Патент РСТ WO 98/18930, опубликованный 7 мая 1998 г. и озаглавленный Антигены и вакцины стрептококковой пневмонии, описывает определенные полипептиды, и заявляется, что они обладают антигенностью. Однако никаких сведений относительно биологической активности этих полипептидов не сообщается. Аналогично не называется никакая консервативная последовательность, являющаяся необходимым биотипом для стандартных вариантов вакцин. Патент WO 00/39299 описывает полипептиды, а также полинуклеотиды, кодирующие эти полипептиды. В этом патенте показано, что полипептиды, обозначенные как BVH-3 и BVH-11, в эксперименте обеспечивают защиту от смертельной инфекции пневмококков. Следовательно, сохраняется неудовлетворенная потребность стрептококковых антигенов, которые могут быть полезны в качестве компонентов для профилактики, диагностики и/или терапии стрептококковой инфекции. Краткое изложение настоящего изобретения Изолированный полинуклеотид, включающий полинуклеотид, выбранный из(а) полинуклеотида, кодирующего полипептид, обладающий по крайней мере 70% идентичностью со вторым полипептидом, выбранным из таблицы А, В, D, Е или Н;(б) полинуклеотида, кодирующего полипептид, обладающий по крайней мере 95% идентичностью со вторым полипептидом, выбранным из таблицы А, В, D, Е или Н;(г) полинуклеотида, кодирующего полипептид, выбранный из таблицы А, В, D, Е или Н;(д) полинуклеотида, кодирующего полипептид, способный вырабатывать антитела, обладающие специфическим связыванием с полипептидом, имеющим последовательность, выбранную из таблицы А,В, D, Е или Н;(е) полинуклеотида, кодирующего несущий эпитоп участок полипептида, выбранного из таблицы А, В, D, Е или Н; а также(ж) полинуклеотида, комплементарного полинуклеотиду из пп.(а), (б), (в), (г), (д) или (е). Другими аспектами обеспечиваются новые полипептиды, кодируемые полинуклеотидами по настоящему изобретению; фармацевтические составы или вакцины; векторы, содержащие полинуклеотиды по настоящему изобретению и реально связанные с областью контролируемой экспрессии, а также клетки-хозяина, в которые проведена трансфекция указанными векторами; и способы получения полипептидов, включая культивирование указанных клеток-хозяина в условиях, пригодных для экспрессии.-1 006232 Краткое описание рисунков На фиг. 1 приведена ДНК-последовательность гена SP64 BVH-3: SEQ ID NO:1. На фиг. 2 приведена ДНК-последовательность, содержащая полностью последовательность генаSP64 ВVН-3, начиная с 1777 нуклеотида до 4896 нуклеотида; SEQ ID NO:2. На фиг. 3 приведена ДНК-последовательность гена SP64 BVH-11; SEQ ID NO:3. На фиг. 4 приведена ДНК-последовательность, содержащая полностью последовательность генаSP64 BVH-11, начиная с 45 нуклеотида до 2567 нуклеотида; SEQ ID NO:4. На фиг. 5 приведена ДНК-последовательность, содержащая полностью последовательность генаSP64 BVH-11-2, начиная с 114 нуклеотида до 2630 нуклеотида; SEQ ID NO:5. На фиг. 6 приведена последовательность аминокислот полипептида SP64 BVH-3; SEQ ID NO:6. На фиг. 7 приведена последовательность аминокислот полипептида SP64 BVH-11; SEQ ID NO:7. На фиг. 8 приведена последовательность аминокислот полипептида SP64 BVH-11-2; SEQ ID NO:8. На фиг. 9 приведена ДНК-последовательность гена SP63 BVH-3; SEQ ID NO:9. На фиг. 10 приведена последовательность аминокислот полипептида SP63 BVH-3 ; SEQ ID NO:10. На фиг. 11 приведена последовательность аминокислот полипептида 4D4.9;SEQ ID NO:11. На фиг. 12 приведена последовательность аминокислот полипептида 7G11.7;SEQ ID NO:12. На фиг. 13 приведена последовательность аминокислот полипептида 7G11.9;SEQ ID NO:13. На фиг. 14 приведена последовательность аминокислот полипептида 4D3.4;SEQ ID NO:14. На фиг. 15 приведена последовательность аминокислот полипептида 8E3.1;SEQ ID NO:15. На фиг. 16 приведена последовательность аминокислот полипептида 1G2.2;SEQ ID NO:16. На фиг. 17 приведена последовательность аминокислот полипептида 10C12.7;SEQ ID NO:17. На фиг. 18 приведена последовательность аминокислот полипептида 14F6.3;SEQ ID NO:18. На фиг. 19 приведена последовательность аминокислот полипептида B12D8.2;SEQ ID NO:19. На фиг. 20 приведена последовательность аминокислот полипептида 7F4.1;SEQ ID NO:20. На фиг. 21 приведена последовательность аминокислот полипептида 10D7.5;SEQ ID NO:21. На фиг. 22 приведена последовательность аминокислот полипептида 10G9.3, полипептида 10 А 2.2 и полипептида B11B8.1; SEQ ID NO:22. На фиг. 23 приведена последовательность аминокислот полипептида 11B8.4;SEQ ID NO:23. На фиг. 24 приведена последовательность аминокислот целевого эпитопа моноклонального антитела H11B-11B8; SEQ ID 163. На фиг. 25 схематически изображен ген BVH-3, а также приведено расположение последовательностей гена, кодирующих полипептиды по всей их длине и укороченные полипептиды. Соотношение фрагментов ДНК дано относительно друг друга. На фиг. 26 схематическое изображен ген BVH-11, а также приведено расположение последовательностей гена, кодирующих полипептиды по всей их длине и укороченные полипептиды. Соотношение фрагментов ДНК дано относительно друг друга. На фиг. 27 схематически изображен ген BVH-11-2, а также приведено расположение последовательностей гена, кодирующих полипептиды по всей их длине и укороченные полипептиды. Соотношение фрагментов ДНК дано относительно друг друга. На фиг. 28 схематически изображен белок BVH-3, а также приведено расположение внутреннего и поверхностных эпитопов, распознаваемых определенными моноклональными антителами. На фиг. 29 схематически изображен белок BVH-11-2, а также приведено расположение защитных поверхностных эпитопов, распознаваемых определенными моноклональными антителами. Фиг. 30 представляет карту плазмиды PURV22.HIS. KanR -резистентная канамицину кодирующая область; с 1857 термочувствительный репрессивный ген бактериофагас 1857;pL -промотор транскрипции бактериофага ; His-tag обозначает кодирующую область 6-гистидин; терминатор - терминатор транскрипции Т 1; ори -оригинал репликации colEl. На фиг. 31 представлено сравнение последовательностей аминокислот белков BVH-3M (Sp64)иBVH-3 (Sp63), проведенное с использованием программы Clustal W, программное обеспечение MacVector по секвенированию (версия 6.5.3). Внизу находится обобщающая линия, на которой значкамии . соответственно обозначены идентичные и похожие остатки аминокислот. На фиг. 32 представлено сравнение последовательностей аминокислот белков BVH-3, BVH-11 иBVH-11-2, проведенное с использованием программы Clustal W, программное обеспечение MacVector по секвенированию (версия 6.5.3). Внизу находится обобщающая линия, на которой значкамии . соответственно обозначены идентичные и похожие остатки аминокислот. На фиг. 33 приведена ДНК последовательность гена NEW43 (SEQ ID No 257). На фиг. 34 приведена логически установленная последовательность аминокислот полипептидаNEW43 (SEQ ID No 258). Подробное описание настоящего изобретения Было установлено, что участки полипептидов BVH-3 и BVH-11 являются внутренними. В штаммах,типа инкапсулированного s.pneumonia, вызывающих заболевания, другие участки, не содержались. Было-2 006232 бы желательно располагать таким полипептидом, который содержал бы участок, не являющийся внутренним. Если значительные участки полипептида являются внутренними, то эти участки не подвержены действию бактерий. Однако в рекомбинантном полипептиде эти участки могут быть в высокой степени иммуногенными и не будут создавать защиту от инфекций. Было бы также желательно иметь в распоряжении полипептид, включающий такой участок, который содержится в большинстве штаммов. Настоящее изобретение касается полипептидов, в которых, в целях получения специфической иммунной реакции, ненужные участки уничтожены и/или модифицированы. В соответствии с настоящим изобретением обеспечиваются также полипептиды или полинуклеотиды, кодируемые такими полипептидами, содержащими защищенные домены. Удивительно, что когда ненужные участки полипептидов уничтожаются или модифицируются, то эти полипептиды обладают требуемыми биологическими свойствами. Это удивительно с точки зрения того факта, что в описании патента РСТ WO 98/18930 некоторые из этих участков описаны как участки,несущие эпитоп. В других публикациях (типа РСТ WO 00/37105) утверждалась важность участков, идентифицированных как триада гистидина и области витка, свернутые в кольцо. Настоящим изобретением установлено, что варианты полипептидов BVH-3 и BVH-11, в которых были уничтожены и/или модифицированы определенные участки, а также химеры этих полипептидов, обладают биологическими свойствами, и вызывают специфическую иммунную реакцию. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полипептид, обладающий по крайней мере 70% идентичностью со вторым полипептидом, содержащим последовательность, раскрываемую в настоящем описании, его таблицах и рисунках. В соответствии с одним аспектом настоящего изобретения обеспечивается изолированный полинуклеотид, включающий полинуклеотид, выбранный из(а) полинуклеотида, кодирующего полипептид, обладающий по крайней мере 70% идентичностью со вторым полипептидом, выбранным из таблицы В, Е или Н;(б) полинуклеотида, кодирующего полипептид, обладающий по крайней мере 95% идентичностью со вторым полипептидом, выбранным из таблицы В, Е или Н;(д) полинуклеотида, кодирующего полипептид, способный производить антитела, обладающие специфическим связыванием с полипептидом, имеющим последовательность, выбранную из таблицы В, Е или Н;(е) полинуклеотида, кодирующего несущий эпитоп участок полипептида, выбранного из таблицы В, Е или Н; а также(ж)полинуклеотида, комплементарного полинуклеотиду из пп.(а), (б), (в), (г), (д) или (е). Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, кодирующий полипептид, обладающий по крайней мере 70% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. А, В, D, E, G или Н, или их аналогов или производных. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, кодирующий полипептид, обладающий по крайней мере 95% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. А, В, D, E, G или Н, их аналогов или производных. Согласно одному аспекту настоящее изобретение касается полипептидов, характеризующихся последовательностью аминокислот, выбранной из табл. А, В, D, E, G или Н, или их аналогов или производных. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, кодирующий полипептид, обладающий по крайней мере 70% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. А, В, D, E, G или Н. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, кодирующий полипептид, обладающий по крайней мере 95% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. А, В, D, E, G или Н. Согласно одному аспекту настоящее изобретение касается полипептидов, характеризующихся последовательностью аминокислот, выбранной из табл. А, В, D, E, G или Н. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, кодирующий полипептид, обладающий по крайней мере 70% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. В, Е или Н, или их аналогов или производных. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, кодирующий полипептид, обладающий по крайней мере 95% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. В, Е или Н, или их аналогов или производных. Согласно одному аспекту настоящее изобретение касается полипептидов, характеризующихся последовательностью аминокислот, выбранной из табл. В, Е или Н, или их аналогов или производных. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, ко-3 006232 дирующий полипептид, обладающий по крайней мере 70% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. В, Е или Н. Согласно одному аспекту настоящее изобретение обеспечивает изолированный полинуклеотид, кодирующий полипептид, обладающий по крайней мере 95% идентичностью со вторым полипептидом,содержащим последовательность, выбранную из табл. В, Е или Н. Согласно одному аспекту настоящее изобретение касается полипептидов, характеризующихся последовательностью аминокислот, выбранной из табл. В, Е или Н. В соответствии с настоящим изобретением все нуклеотиды, кодирующие полипептиды и химерные полипептиды, входят в границы настоящего изобретения. По другому варианту настоящего изобретения полипептиды или химерные полипептиды в соответствии с настоящим изобретением являются антигенными. По другому варианту настоящего изобретения полипептиды или химерные полипептиды в соответствии с настоящим изобретением являются иммуногенными. По еще одному варианту настоящего изобретения полипептиды или химерные полипептиды в соответствии с настоящим изобретением способны вызывать у субъекта иммунную реакцию. По другому варианту настоящее изобретение касается также полипептидов, которые способны мобилизовать антитела, обладающие специфическим связыванием с определенными выше полипептидами или химерными полипептидами по настоящему изобретению. В еще одном варианте фрагменты полипептидов по настоящему изобретению должны содержать один или более несущих эпитоп участков, которые проидентифицированы в табл. С и F. Этот участок должен включать по крайней мере 15 заменимых аминокислот полипептида из табл. С и F. Этот участок должен включать по крайней мере 20 заменимых аминокислот полипептида из табл. С и F. В другом варианте настоящего изобретения создающий несущий эпитоп участок полипептида из таблицы A(BVH-3) содержит по крайней мере один полипептид, приведенный в табл. С. В другом варианте настоящего изобретения создающий несущий эпитоп участок полипептида из таблицы B(BVN-11) содержит по крайней мере один полипептид, приведенный в табл. F. Антитело, обладающее специфическим связыванием, представляет собой антитело, которое в некоторой пробе (типа биологической пробы) распознает и связывает выбранные полипептиды, но в значительной степени не распознает и не связывает другие молекулы. Специфическое связывание можно измерить с помощью твердофазного иммуноферментного анализа (ТФИФА) в котором выбранный полипептид используется в качестве антигена. Если не приведено другого определения, то все используемые здесь технические и научные термины имеют то же самое значение, которое обычно понимается людьми, квалифицированными в области, к которой относится настоящее изобретение. Все публикации, описания патентов, патенты и другие упомянутые здесь ссылки даны по всей полноте. В случае конфликта следует сделать проверку настоящего описания, включая содержащиеся в нем определения. Кроме того, материалы, способы и примеры являются иллюстративными и не предназначены для ограничения настоящего изобретения. В соответствии с настоящим изобретением термин защита в проведенных биологических исследованиях определяется значительным ростом кривой выживаемости, коэффициента или периода выживаемости. Для того, чтобы вычислить значение вероятности Р и определить, является ли разница между двумя группами существенной со статистической точки зрения, может быть полезным применение статистического анализа с использованием логарифмического рангового критерия (для сравнения кривых выживаемости) и точного критерия Фишера (для сравнения коэффициентов выживаемости и количества дней вплоть до смерти). Величины Р, составляющие 0,05 считаются несущественными. Используемые здесь термины фрагменты, производные, или аналоги полипептидов по настоящему изобретению включает такие полипептиды, в которых один аминокислотный остаток или большее их количество заменены консервативным или неконсервативным аминокислотным остатком(предпочтительно консервативным),и такие полипептиды, которые могут быть природными или неприродными. По одному варианту производные и аналоги полипептидов по настоящему изобретению должны обладать приблизительно 70% идентичностью с последовательностями, приведенными на рисунках или их фрагментами. Это означает, что 70% этих остатков одинаковы. По другому варианту полипептиды должны обладать более чем 75% гомологией. По еще одному варианту полипептиды должны обладать более чем 80% гомологией. По другому варианту полипептиды должны обладать более чем 85% гомологией. По другому варианту полипептиды должны обладать более чем 90% гомологией. По еще одному варианту полипептиды должны обладать более чем 95% гомологией. По другому варианту полипептиды должны обладать более чем 99% гомологией. В еще одном варианте производные и аналоги полипептидов по настоящему изобретению должны содержать меньше, чем приблизительно 20 замененных, модифицированных аминокислотных остатков или аминокислотных остатков, претерпевших делецию, а более предпочтительно, чтобы их количество было меньше 10. Предпочтительными заменами являются такие замены, которые в данной технологии известны как консервативные, а именно, замененные остатки наделены физическими или химическими свойствами (типа гидрофобности, размера, заряда) функциональных групп.-4 006232 Квалифицированные люди должны понимать, что аналоги или производные белков или полипептидов по настоящему изобретению также должны найти применение в контексте настоящего изобретения,а именно в качестве антигенного иммуногенного материала. Таким образом, белки или полипептиды,содержащие одно или более дополнений, делеций, замен и т.п.входят в настоящее изобретение. Помимо этого может быть возможной замена одной аминокислоты другой аминокислотой того же типа. Например, замена одной гидрофобной аминокислоты другой гидрофобной аминокислотой. Для сравнения последовательности аминокислот можно использовать программу, типа программыClustal. Эта программа сравнивает последовательности аминокислот и находит оптимальное выравнивание, внедряя, в качестве уместных, пробелы в любую последовательность. Для оптимального выравнивания можно подсчитать идентичность аминокислот или их схожесть (идентичность плюс консервативность типа аминокислоты). Программа типа BLASTx должна выровнять самый длинный фрагмент секвенирования сходных последовательностей и установить величину нужного соответствия. Таким образом можно провести сравнение, при котором обнаруживаются несколько схожих областей, каждая из которых имеет разное положение. Настоящее изобретение рассматривает оба типа исследования идентичности. При альтернативном подходе аналогами или производными могут являться слитые белки, которые включают компоненты, более легко подвергающиеся очистке, например эффективным нанесением метки на целевой белок или полипептид. Может быть необходимым удаление этой метки, или может быть так, что сам слитый белок сохраняет антигенность, достаточную для его полезности. В дополнительном аспекте настоящего изобретения обеспечиваются антигенные/иммуногенные фрагменты белков или полипептидов по настоящему изобретению, или их аналогов или фрагментов. Для того, чтобы сохранить свои антигенные/иммуногенные свойства, фрагменты по настоящему изобретению должны включать один такой эпитопный участок (или большее их количество), или они должны быть в достаточной степени похожи на такие участки. Таким образом, согласно настоящему изобретению для фрагментов возможна нерелевантная степень идентичности, поскольку эти фрагменты могут иметь 100% идентичность с определенной частью белка или полипептида, его аналога или производного. Вновь существенным результатом является то, чтобы конкретный фрагмент сохранял антигенные/иммуногенные свойства. Таким образом для аналогов, производных и фрагментов существенно то, чтобы они по крайней мере обладали степенью антигенности/иммуногенности того белка или полипептида, производным которого являются. В соответствии с настоящим изобретением в полипептиды по настоящему изобретению входят и полипептиды, и химерные полипептиды. Входят также и полипептиды, слитые с другими соединениями, меняющими биологические или фармакологические свойства полипептидов, а именно с полиэтиленгликолем - для увеличения периода полупревращения, с лидерной или секреторной последовательностью аминокислот - для облегчения процесса очистки, препро- и пропоследовательностями, а также с (поли)сахаридами. Более того, в тех ситуациях, когда обнаруживается, что области аминокислот полиморфны, то для более эффективного копирования различных эпитопов разных стрептококковых штаммов желательным может быть изменение одной конкретной аминокислоты или большего их количества. Помимо этого, для того, чтобы обеспечить стабильность и, в целях сцепления или связывания с подложкой или другой молекулой, увеличить гидрофобность, полипептиды по настоящему изобретению можно модифицировать NH2 -концевым ацилированием (например, ацилированием, амидирование тиогликолевой кислотой, концевым карбоксиамидированием, к примеру аммиаком или метиламином). Рассматриваются также гетеро- и гомополипептидные мультимеры полипептидных фрагментов,аналогов и производных. Эти полимерные формы включают, например, один или более полипептидов,сшитых сшивающим линкером (таким, как авидин/биотин, глютеральдегид или диметисуперимидат). В такие полимерные формы включены также полипептиды, которые содержат две или более последовательно соединенные последовательности или обращенные и следующие друг за другом последовательности, продуцированные мультицистронными мРНК, полученными генной инженерией. Предпочтительно, чтобы фрагмент, аналог или производное полипептида по настоящему изобретению содержал по крайней мере одну антигенную область, а именно по крайней мере один эпитоп. Для того, чтобы осуществить получение антигенных полимеров (а именно, синтетических мультимеров) можно использовать полипептиды с бис-галоацетильными группами, нитроарилгалогенидами или т.п.группами, в которых указанные реагенты являются специфическими для тиогрупп. Поэтому связь двух меркаптогрупп различных полипептидов может представлять собой одинарную химическую связь,или она может быть образована связывающей группой, содержащей по крайней мере 2 атома углерода,обычно по крайней мере 4, но не более 16 атомов углерода (обычно не более чем приблизительно 14 атомов углерода). В частном варианте полипептидные фрагменты, аналоги и производные по настоящему изобретению не содержат метиониновый (Мет) остаток как стартовый. Предпочтительно, чтобы полипептиды не содержали лидерной или секреторной последовательности (сигнальной последовательности). Сигналь-5 006232 ный участок полипептида по настоящему изобретению можно определить, используя утвержденные методику молекулярной биологии. В общих чертах для того, чтобы установить первичный остаток зрелого белка, а следовательно и последовательность зрелого полипептида, из стрептококковой культуры можно выделить представляющий интерес полипептид, а затем его секвенировать. Согласно другому аспекту настоящего изобретения обеспечиваются составы вакцин, которые содержат один или более стрептококковых полипептидов по настоящему изобретению в смеси с фармацевтически приемлемым носителем, разбавителем или адъювантом. Подходящие адъюванты включают масла, а именно полный или неполный адъювант Фрейнда; соли, а именно AlK(SO4)2, AlNa(SO4)2,AlNH4(SO4)2; кремнезем, каолин, углеродные полинуклеотиды, а именно поли-IC и поли-AU. Предпочтительные адъюванты включают QuilA и Alhydrogel. Вакцины по настоящему изобретению можно вводить парентеральной инъекцией, быстрой инфузией, назофарингеальным всасыванием, дермоабсорбцией, трансбуккально или перорально. В фармацевтически приемлемые носители входит также столбнячный анатоксин. Термин вакцина подразумевает также включение антител. В соответствии с настоящим изобретением в целях лечения или профилактики стрептококковой инфекции и/или заболеваний и симптомов,опосредованных стрептококковой инфекцией, обеспечивается применением одного или более антител,обладающих специфическим связыванием с полипептидами по настоящему изобретению. Составы вакцин по настоящему изобретению используются для лечения или профилактики стрептококковой инфекции и/или заболеваний и симптомов, опосредованных стрептококковой инфекцией таким образом, как это описано в руководстве по клинической микробиологии (P. R. Murray (Ed, in chief),E. J. Baron, M. A. Pfaller, F. C.Tenover and R. H. Yolken. Manual of Clinical Microbiology, ASM Press,Washington, D. С sixth edition, 1995, 1482 p), ссылка на которое дана здесь полностью. В одном варианте составы вакцины по настоящему изобретению используют для лечения или профилактики менингита,среднего отита, бактериемии или пневмонии. В одном варианте составы вакцины по настоящему изобретению используют для лечения и профилактики стрептококковой инфекции и/или заболеваний и симптомов, опосредованных стрептококковой инфекцией, в частности S. pneumoniae, группой А стрептококков (pyogenes), группой В стрептококков (GBS или agalactiae), disgalactiae, uberis, nocardia, а также золотистым стафилококком. В другом варианте указанная стрептококковая инфекция представляет собой S.pneumoniae. В частном варианте вакцины вводят субъектам, имеющим фактор риска стрептококковой инфекции, таким, как дети, люди старшего возраста и лица с нарушением иммунитета. Согласно использованию в настоящем описании термин субъекты включает млекопитающих. В другом варианте таким млекопитающим является человек. Составы вакцины предпочтительно представляют собой стандартную дозировку, составляющую приблизительно от 0,001 до 100 мкг/кг (антиген/масса тела), более предпочтительно от 0,01 до 10 мкг/кг,а наиболее предпочтительно от 0,1 до 1 мкг/кг. Эту дозировку вводят от 1 до 3 раз с интервалом между иммунизациями, равным приблизительно от 1 до 6 недель. Составы вакцины предпочтительно представляют собой стандартную дозировку, составляющую приблизительно от 0,01 мкг до 10 мг, более предпочтительно от 1 мкг до 1 мг, а наиболее предпочтительно от 10 до фрагмент антитела обозначается антитело или фрагмент антитела, продуцированные с помощью молекулярной биологии. Указанное антитело или фрагменты антитела могут быть поликлональными, или, предпочтительно, моноклональными. Антитело может быть специфическим для нескольких эпитопов, связанных с полипептидами стрептококковой пневмонии, но предпочтительно, чтобы оно было специфическим для одного эпитопа. Далее приводятся таблицы, суммирующие последовательности, раскрытые в настоящем описании. которые сшивают в целях получения полного полипептида (блочное сшивание). Стандартные способы получения и расшифровки полинуклеотидов и полипептидов описаны в приведенных ниже работах: Sambrook et al., (1989) Molecular cloning: A Laboratory Manual, 2nd ed, Cold SpringColigan J. E. et al., John WileySons Inc., New York, ссылки на которые здесь приводятся. Для продуцирования рекомбинантов осуществляют трансфекцию клетки-хозяина векторами, кодирующими указанный полипептид, а затем проводят культивирование в питательной среде, модифицированной так, как это необходимо для активации промоторов, отбора трансформантов или для амплификации генов. Подходящими векторами являются вектора, жизнеспособные в выбранной клетке-хозяине и способные к репликации в ней; они включают последовательности хромосомных, нехромосомных и синтетических ДНК, например бактериальные плазмиды, ДНК фагов, бакуловирусов, плазмиды дрожжей,векторы, полученные из объединений плазмид и ДНК фагов. Используя рестрикционные ферменты, последовательность полипептидов можно внедрить в вектор на определенное место так, чтобы она была реально сцеплена с содержащей промотор областью контролируемой экспрессии, сайтом связывания рибосомы (согласованная область или последовательность Шайн-Далгарно), а также (необязательно) с оператором (контролирующий элемент). В соответствии с установленными принципами молекулярной биологии (Sambrook et al., (1989) Molecular cloning: A Laboratory Manual, 2nd ed, Cold Spring Harbor, N.Y.;Current Protocols in Molecular Biology, (1999) Edited by Ausubel F. M. et al., John WileySons, Inc., N.Y.) можно подобрать отдельные компоненты области контролируемой экспрессии, пригодные для данной клетки-хозяина и вектора. Подходящие промоторы включают (однако ими не ограничены): промоторLTR или SV40, lac, tac или trp промоторы E.coli, а также PL промотор -фага. Векторы должны предпочтительно содержать исходные селективные маркеры (а именно ген, резистентный ампициллину) или их репликацию. Пригодные бактериальные вектора включают: рЕТ, pQE70, pQE60, pQE-9, pbs, pD10phagescript, psiX174, pblueacript SK, pbsks, pNH8A, pNH16a, pNH18A, pNH46A, ptrc99a, pKK223-3,pKK233-3, pDR540, pRIT5, a также вектора эукариот : pBlueBacIII, pWLNEO, pSV2CAT, pOG44, pXTl,pSG, pSVK3, pBPV, pMSG and pSVL. Клетки-хозяина могут представлять собой бактерии, а именно:E.Coli, Bacillus subtilis, Streptomyces; грибы: Aspergillus niger, Aspergillus nidulins; дрожжи, а именно Saccharomyces или эукариоты, а именно СНО, COS. При экспрессии полипептида в культуре полученные клетки обычно собирают центрифугированием, а затем разрушают физическим или химическим способом (если прошедший экспрессию полипептид не выделяют в среду). Получившийся в результате этого неочищенный экстракт оставляют для выделения нужного полипептида. Очистку полипептида от культуральной среды или лизата можно провести по общепринятой технологии в зависимости от свойств самого полипептида. Для этого используют осаждение сульфатом аммония или этанолом, экстракцию кислотой, анионную или катионную обменную хроматографию, фосфоцеллюлозную хроматографию, гидрофобную хроматографию, гидроксиапатитовую хроматографию и лектиновую хроматографию. Окончательную очистку можно провести с помощью жидкостной хроматографии высокого разрешения (ВЭЖХ). Полипептид можно подвергнуть экспрессии с лидерной или секретирующей последовательностью или без нее. В первом случае указанную лидерную последовательность можно удалить посттрансляционной обработкой (см. патенты США 4,431,739; 4,425,437 и 4,338,397, на которые здесь сделана ссылка),или (после очистки прошедшего экспрессию полипептида), используя химические способы удаления. Согласно другому аспекту стрептококковые полипептиды по настоящему изобретению можно использовать как диагностический тест на стрептококковую инфекцию, в частности, на инфекцию S. pneumoniae. Для того, чтобы выявить, например, стрептококковый организм в биологическом образце, возможно применение нескольких диагностических способов. Возможно выполнение следующей методики:(а) получение от пациента биологического образца;(б) инкубирование антитела или его фрагмента, реагирующего со стрептококковым полипептидом по настоящему изобретению, с биологическим образцом в целях образования смеси; а также(в) выявление в указанной смеси специфически связанного антитела или связанного фрагмента, что указывает на присутствие стрептококков. Или же, способ выявления антитела, специфичного для стрептококкового антигена в биологическом образце, содержащем указанное антитело, или предположительно содержащем его, можно провести следующим образом:(а) получение от пациента биологического образца;(б) инкубирование одного или более стрептококковых полипептидов по настоящему изобретению или их фрагментов с биологическим образцом в целях образования смеси; а также(в) выявление в указанной смеси специфически связанного антитела или связанного фрагмента, что указывает на присутствие стрептококков. Квалифицированный в этой технологии человек поймет, что этот диагностический тест может иметь разнообразные формы, они включают иммунологический тест типа твердофазного иммуноферментного анализа (ТФИФА), радиоиммунологический тест, или латексный анализ, и, в основном, предназначены для определения являются ли присутствующие в организме антитела специфическими для указанного полипептида. Последовательности ДНК, кодирующие полипептиды по настоящему изобретению, можно также использовать чтобы сконструировать ДНК-зонды, применяемые для обнаружения стрептококков в биологическом образце, предположительно содержащем такие бактерии. Способ выявления согласно настоящему изобретению включает(а) получение от пациента биологического образца;(б) инкубирование одного или более ДНК-зондов, обладающих последовательностью ДНК, кодирующей полипептиды по настоящему изобретению или их фрагменты, с биологическим образцом в целях образования смеси; а также(в) выявление в указанной смеси специфически связанного ДНК-зонда, что указывает на присутствие стрептококков. ДНК-зонды по настоящему изобретению можно также использовать для выявления в образце циркулирующих стрептококков, а именно нуклеиновых кислот S.pneumoniae. В качестве способа диагностики стрептококковых инфекций можно использовать, например цепную реакции полимеразы (ПЦР). Такие зонды можно синтезировать, используя стандартную технологию, и можно провести их иммобилизацию на твердой фазе, или на них можно нанести выявляемую метку. Предпочтительным ДНКзондом для такого применения может служить олигомер, последовательность которого комплементарна по крайней мере 6 соседним нуклеотидам стрептококковых полипептидов по настоящему изобретению. Другой диагностический способ выявления стрептококков у пациента включает(а) нанесение выявляемой метки на антитело, взаимодействующее с полипептидом по настоящему(б) введение пациенту указанного меченного антитела или меченного фрагмента; а также(в) выявление у указанного пациента специфически связанного антитела или связанного фрагмента,что указывает на присутствие стрептококков. Следующим аспектом настоящего изобретения является применение стрептококковых полипептидов по настоящему изобретению в качестве иммуногенов при продуцировании специфических антител в целях диагностики, и, в частности, лечения стрептококковых инфекций. Подходящие антитела можно определить стандартными методами скриннинга, например, измеряя на опытной модели способность конкретного антитела к пассивной защите от стрептококковой инфекции. Одним примером модели на животных является описанная ниже модель на мыши. Указанным антителом может быть антитело целиком или его антигенсвязывающий фрагмент, и оно может относиться к любому иммунологическому классу. Такое антитело или его фрагмент может антителом животного, в частности, млекопитающего, а более конкретно - мыши, крысы или человека. Это может быть природное антитело или его фрагмент,или, если это необходимо, рекомбинантное антитело или фрагмент антитела. Термином рекомбинатное антитело или фрагмент антитела обозначается антитело или фрагмент антитела, продуцированные с помощью молекулярной биологии. Указанное антитело или фрагменты антитела могут быть поликлональными, или, предпочтительно, моноклональными. Антитело может быть специфическим для нескольких эпитопов, связанных с полипептидами стрептококковой пневмонии, но предпочтительно, чтобы оно было специфическим для одного эпитопа. Другим аспектом настоящего изобретения является использование для пассивной иммунизации антител, специфических к стрептококковым полипептидам по настоящему изобретению. Можно использовать антитела, описанные в настоящем изобретении. Подходящие антитела можно определить стандартными методами скриннинга, например, измеряя на опытной модели способность конкретного антитела к пассивной защите от стрептококковой инфекции. Одним примером модели на животных является описанная ниже модель на мыши. Указанным антителом может быть антитело целиком или его антигенсвязывающий фрагмент, и оно может принадлежать любому иммунологическому классу. Такое антитело или его фрагмент может антителом животного, в частности, млекопитающего, а более конкретно - мыши,крысы или человека. Это может быть природное антитело или его фрагмент, или, если это необходимо,рекомбинантное антитело или фрагмент антитела. Термином рекомбинатное антитело или фрагмент антитела обозначается антитело или фрагмент антитела, продуцированные с помощью молекулярной биологии. Указанное антитело или фрагменты антитела могут быть поликлональными, или, предпочтительно, моноклональными. Антитело может быть специфическим для нескольких эпитопов, связанных с полипептидами стрептококковой пневмонии, но предпочтительно, чтобы оно было специфическим для одного эпитопа. Далее приводятся таблицы, суммирующие последовательности, раскрытые в настоящем описании.- 13006232 Пример 1. В этом примере описаны использованные штаммы бактерий, плазмиды, праймеры цепной реакции полимеразы (ПЦР), рекомбинантные белки, а также гибридома, продуцирующая антитела. Клинические изоляты S. pneumoniae SP64 (серогруппа 6) и SP63 (серогруппа 9) были предоставлены by the Laboratoire de la Sant Publique du Qubec, Sainte-Anne-de-Bellevue; штамм Rxl - неинкапсулированное производное типа 2 штамма D39, а также типа 3 штамма WU2 были получены от David E. BrilesRecherche en Infectiologie du Centre Hospitalier de l'Universite Laval, Sainte-Foy. В настоящем исследовании использовались штаммы Е. coli: DHS (Gibco BRL, Gaithesburg, MD), AD494 (DE3) (Novagen, Madison,WI) и BL21 (JDE3) (Novagen); а также вектор pSL301 суперлинкера плазмиды (Invitrogen, San Diego,CA); вектор pCMV-GH (предоставлен Dr. Stephen A. Johnston, Department for Biochemistry, University ofTexas, Dallas, Texas); pET32 и pET21 (Novagen), а также вектора экспрессии pURV22.HIS (рис. 30). Этот вектор pURV22.HIS содержит кассету бактериофагасI857, термочувствительного репрессивного гена,из которого была произведена делеция функционального промотора PR. Дезактивация репрессора сI857 путем увеличения температуры от 30-37 С до 37-42 С вызывает индукцию гена под контролем промотора PL. Праймеры ПЦР, используемые для получения рекомбинантных плазмид, у 5'-конца содержат сайт рестрикции эндонуклеазы, что позволяет направленное клонирование амплифицированного продукта в гидролизованный вектор плазмиды. Использованные олигонуклеотидные праймеры ПЦР приведены ниже в табл. 1. Расположение последовательностей гена, кодирующих генные продукты BVH-3, BVH-11 и BVH-11-2, показаны на рис. 25, 26 и 27 соответственно. Таблица 1. Перечень олигонуклеотидных праймеров ЦПР Приведенные здесь и далее в тексте описания, а также на рисунках символы а, с, g и t обозначают аденозин, цитозин, гуанозин и тимидин соответственно (примеч. переводчика). Методики молекулярной биологии были осуществлены стандартным образом. См., например, руководство Sambrook, J., Fritsch, E. F., Maniatis, Т., "Molecular cloning. A laboratory manual" Vol. 1-2-3 (secondedition) Cold Spring Harbour laboratory Press, 1989, New York, которое включено здесь в виде ссылки. Амплифицированные продукты ПЦР гидролизовали рестрикционной эндонуклеазой и провели их сшивание или с линеаризованной плазмидой pSL30l, pCMV-GH, рЕТ, или с вектором экспрессии pURV22.HIS,гидролизованным аналогично, или гидролизованным ферментом, продуцирующим совместимые липкие концы. Рекомбинант pSLSOl и рекомбинантные плазмиды pCMV-GH гидролизовали рестрикционными ферментами для того, чтобы осуществить рамочное клонирование в вектор экспрессии рЕТ. При использовании векторов рЕТ клоны вначале стабилизировали в E.coli DH5, это проводилось до введения в E.coli ВL21(DE3) или AD494 (DE3) в целях экспрессии молекул BVH-3, BVH-11 или BVH-11-2 по всей их длине или укороченных молекул. Каждый из полученных в результате конструктов плазмид был подтвержден секвенированием нуклеотидов. Рекомбинантые белки обозначены как N-концевые слитые с тиоредоксином и His-tag (pET32 система экспрессии) белки; как С-концевые слитые с His-tag (сис- 17006232 тема экспрессии рЕТ 21) белки; или как N-концевые слитые с His-tag (система экспрессии рURV22.НIS)белки. Полученные экспрессией рекомбинантные белки очистили от супернатантных фракций, образовавшихся после центрифугирования обработанных ультразвуком IPIG- (системы рЕТ) или термообработанных (pURV22.HIS) индуцированных E. coli; для очистки использовались хелатная металлическая His-Bind смола (QIflgen, Chatsworth, СА). Генные продукты, полученные из S. pneumonae SP64,приведены ниже в табл. 2. Фрагменты гена, кодирующего белок BVH-3-Sp63 (аминокислотные остатки с 21 по 840 из последовательности SQ ID NO: 10), были получены из S. pneumonae SP63, используяOCRR479-OCRR480 совокупности праймеров ЦПР, а также вектор клонирования pSL301. Рекомбинант pSL301-BVH-3Sp63 гидролизовали для рамочного клонирования в вектор рЕТ 32 в целях экспрессии молекулы BVH-3-Sp63. Таблица 2. Перечень укороченных BVH-3, BVH-11, BVH-11-2. а также химерных генных продуктов, полученных из S. pneumoniae SP64- 19006232 Секретирующие моноклональные антитела гибридомы получали слиянием клеток селезенки иммунизированных мышей и несекретирующих HGPRT-неполных клеток миеломы мыши SP2/0, используя методы Fazekas De St-Groth and Scheidegger (J. Immunol Methods 35 : 1-21, 1980) с модификациями (J.pURV22His), или NEW1 генными продуктами штамма S. pneumonae SP64 (слитый белок NEW1-HisTag-/ система рЕТ 21). Это проводится для выработки моноклональных антител серий Н 3-, Н 11-, Н 112-, H11B-,H3V- и HN1- соответственно. Сначала супернатанты культур подвергли скриннингу с помощью твердофазного иммуноферментного метода по описанной ранее методике (Hamel et al.); использовались пластины, покрытые препаратами из очищенных рекомбинантных белков BVH-3, BVH-11 и/или BVH-11-2,или из суспензий клеток термообработанных клеток S. pneumoniae. Отобранные для дальнейшего изучения гибридомы, секретирующие моноклональные антитела, представлены в табл. 3 и 4 из приведенного ниже примера 2. Класс и подкласс моноклональных иммуноглобулинов определяли твердофазным иммуноферментным анализом, используя для этого коммерчески доступные реагенты (Southern Biotechnology Associates, Birmingham, AL). Далее описывается клонирование и экспрессия химерного гена (генов), кодирующих химерные полипептиды, а также защита, выявленная после вакцинации указанными химерными полипептидами. Фрагменты генов BVH-3 и BVH-11, соответствующие 3'-концу этих генов, амплифицировали с помощью ПЦР, используя для этого пары олигонуклеотидов, сконструированных таким образом, чтобы амплифицировать те фрагменты генов, которые должны содержаться в указанных химерных генах. Используемые праймеры содержали сайт рестрикции эндонуклеазы у 5'-конца, что допускает направленное рамочное клонирование амплифицированного продукта в прошедшие гидролиз вектора плазмид (таблицы 1 и 2). Амплифицированные с помощью ПЦР продукты гидролизовали рестрикционной эндонуклеазой и осуществили сшивание с линеаризованной плазмидой рЕТ 21 или вектором pSL301. Полученные в результате конструкты плазмид были подтверждены секвенированием нуклеотидов. Рекомбинантные плазмиды рЕТ 21, содержащие продукт ПЦР, линеаризовали гидролизом рестрикционными ферментами с целью рамочного клонирования второго фрагмента ДНК и получения химерного гена, кодирующего химерную молекулу пневмококкового белка. Рекомбинантные плазмиды pSL301, содержащие продукт ПЦР, подвергли гидролизу рестрикционными ферментами для получения включений в ДНК. Полученные в результате включенные фрагменты ДНК очистили, а включения, соответствующие данному химерному гену, сшили с вектором рЕТ 21 в целях получения химерного гена. Рекомбинантные химерные полипептиды, приведенные в табл. 2, представляли собой С-концевые слияния с His-tag. Полученные экспрессией рекомбинантные белки очистили от супернатантных фракций, образовавшихся после центрифугирования обработанных ультразвуком IPIG-(системы рЕТ) или термообработанных (pURV22.HIS) индуцированных Е. coli; для очистки использовалась хелатная металлическая His-Bind смола (QIAgen,Chatsworth, CA). Группу из 8 женских особей BALB/c мышей (Charles River) подкожно иммунизировали, вводя 25 мкг очищенного слитого Histag белка любого сродства (проидентифицированного в присутствии 15-20 мкг адъюванта QuilA), введение проводили дважды с 3-недельным интервалом. Спустя 10-14 дней после последней иммунизации было осуществлено пробное заражение мышей, его проводили, вводя внутривенно 10 Е 5-10 Е 6 колониеобразующих единиц (КОЕ) S.pneumoniae 3 типа, штамм WU2. Полипептиды и фрагменты оказались способными вызывать защитную иммунную реакцию. Таблица 2 А Пример 2. В этом примере описана идентификация пептидного домена, содержащего целевые эпитопы. Идентификацию проводили, используя моноклональные антитела и описанные в примере 1 рекомбинантные укороченные белки. Гибридомы исследовали твердофазным иммуноферментным методом на наличие в них укороченных генных продуктов BVH-3, BVH-11, или ВVН-11-2, это делалось для того, чтобы описать эпитопы,распознаваемые указанными моноклональными антителами. Укороченные генные продукты получали из штамма S. pneumoniae SP64, за исключением BVH-3-Sp63, полученного из штамма S. pneumoniae SP63. Реакционноспособность каждого антитела испытывалась на полных (неукороченных) рекомбинантных белках BVH-3, BVH-11 или BVH-11-2, которые использовали в качестве позитивного контроля. В некоторых случаях реакционную способность моноклональных антител оценивали вестерн-иммуноблоттингом, который проводили после отделения генного продукта с помощью SDS-PAGE и переноса на нитроцеллюлозную бумагу. Выявленная в результате этого реакционная способность приведена далее в табл. 3 и 4. Логически установленные расположения эпитопов показаны на рис. 28 и 29. Как видно из данных табл. 3, моноклональные антитела, реагирующие с BVH-3, можно разделить на две группы: моноклональные антитела, реагирующие с BVH-3A и моноклональные антитела, реагирующие с BVH-3B, за исключением моноклональных антител H11-7G-11 и H3V-15A10, которые реагируют как с молекуламиBVH-3A, так и с молекулами BVH-3B. Моноклональные антитела, реагирующие с BVH-3A, можно разделить на 2 подгруппы в зависимости от их способности реагировать с рекомбинантным белком BVH3C, или отсутствию такой способности. Моноклональные антитела, реагирующие с белком BVH-3C,распознают эпитопы, которыми совместно владеют как белок BVH-3, так и белок ВVH-11. Из данных таблицы 4 видно, что моноклональные антитела, обладающие перекрестной реакционной способностью с BVH-3, и с BVH-11, способны взаимодействовать также с рекомбинантными белками BVH-11 А иBVH-11-2M. Моноклональные антитела, реагирующие с BVH-3B, можно разделить на 3 подгруппы с учетом их способности взаимодействовать с рекомбинантыми белками NEW1, NEW2 и NEW3. Некоторые моноклональные антитела способны реагировать только с белком NEW1, в то время как другие антитела обладают реакционной способностью в отношении любого из рекомбинантных белков NEW1 иNEW2, или NEW1 и NEW3. Моноклональные антитела H11-7G11 взаимодействуют с эпитопами, занимающими более одной позиции в BVH-3. Реакционная способность H11-7G11 с молекулами BVH-3AD, BVH-3B, BVH-3C,BVH-11A и BVH-11-2 М предполагает, что эпитоп H11-7G11 может содержать последовательность НХХНХН. Эта последовательность повторяется соответственно 6 и 5 раз в последовательностях белковBVH-3 и BVH-11/BVH-11-2. Отсутствие реакционной способности моноклональных антител H11-7G11 с молекулой NEW10 предполагает, что эпитоп содержит последовательность HGDHXH. Многопозиционное картирование эпитопа H3V-15A10 на BVH-3 предполагает, что моноклональное антитело взаимодействует с двумя неперекрывающимися фрагментами BVH-3. Интересно, что моноклональные антитела H3-7G2, H3V-9C6 и H3V-16 А 7 не взаимодействовали сBVH-3 Sp63; это допускает возможность расположения их соответствующих эпитопов на фрагменте 177 аминокислоты, находящемся между 244 и 420 аминокислотами в молекуле BVH-3 штамма S.pneumoniaeSP64 (рис. 31). Как видно из данных табл. 4, моноклональные антитела, которые обладают реакционной способностью относительно BVH-11- и/или BVH-11-2, и не распознают молекулы BVH-3, можно разделить на 3 подгруппы с учетом их способности взаимодействовать с рекомбинантыми белками BVH-11A и NEW10. Некоторые моноклональные антитела способны реагировать только с белком BVH-11 А или NEW10, в то время как другие антитела обладают реакционной способностью в отношении и рекомбинантного белкаBVH-11A и NEW10. Пример 3. В этом примере описано создание библиотек генов BVH-3 и BVH-11-2 в целях картирования эпитопов. Библиотеки генов BVH-3 и BVH-11-2 создавали с использованием рекомбинантных плазмид ДНК:pCMV-GH и PSL301, которые содержат соответственно последовательность гена BVH-3, охватывающую нуклеотиды с 1837 по 4909 (SEQ ID NO:2) или последовательность гена BVH-11-2, охватывающую нук- 21006232 леотиды с 172 по 2630 (SEQ ID NO:5); а также систему создания библиотеки Novatope и систему отбора (Novagen). Таблица 3. Определенная методом ТФИФА реакционная способность BVH-3-реактивных моноклональных антител (МКА) с 11 генными продуктами BVH-3, а также с молекулой BVH-11M- 22006232 Таблица 4. Определенная методом ТФИФ реакционная способность BVH-11- и/или BVH-11-2 реактивных моноклональных антител (МКА) с 14 генными продуктами BVH-11 и BVH-11-2, а также с молекулой BVH-3M Рекомбинантные плазмиды, содержащие фрагменты генов BVH-3 или BVH-11-2 очистили, используя для этого QIAgen набор (Chartsworth, CA), а затем подвергли гидролизу рестрикционными ферментами BglII и XbaI соответственно. Полученные в результате BglII-XbaI фрагменты ДНК очистили, проведя экстракцию QIAquick гелем из QIAgen набора, а затем для получения случайным образом расщепленной ДНК осуществили гидролиз ДНКазы I. Фрагменты ДНК из 50-200 базовых точек очистили, и обработали Т 4 ДНК-полимеразой, чтобы затупить концы целевой ДНК. Затем добавили один 3'dA остаток и осуществили сшивание в вектор pSCREEN-T (Novagen); при этом действовали по методикам,предложенным производителем(Novatope System, Novagen). Библиотеки клонов E.coli, каждая из которых отображает небольшой пептид, полученный из генов BVH-3 или BVH-11-2, отсортировали стандартным способом, используя для этого моноклональные антитела как иммунологические зонды. Сортировка колоний моноклональными антителами, создающими очень высокий фон на возвышенностях колоний, не была успешной. Более того, в нескольких случаях не удалось выявить колоний, экспресси- 24006232 рующих эпитоп. Потерю реакционной способности можно вероятно объяснить небольшим количеством продуцированных рекомбинантных белков, или распознаванием конформационно-зависимых эпитопов,содержащих различные белковые домены. Секвенирование включений ДНК из положительных клонов определяет расположение сегмента, кодирующего целевой эпитоп. Эти данные приведены в таблице 5. Пептиды, кодируемые включениями ДНК в рекомбинантный вектор pSCREEN-T, можно очистить и использовать как иммуногены, это описано ниже в примере 6. Последовательности пептидов, полученные в результате скриннинга библиотек генов BVH-3 иBVH-11-2 с антителами, согласуются с данными ТФИФА, касающимися реакционной способности в отношении укороченных генных продуктов. Как и ожидалось, последовательности аминокислот, полученные из H11-7G11, содержали последовательность HGDHXH. Эти данные обеспечивают дополнительное доказательство расположению эпитопов, распознаваемых антителами. Интересно, что хотя антителаH112-10G9, H112-10 А 2 и Н 11 В-11 В 8 и обладали реакционной способностью в отношении одной и той же последовательности пептидов (аминокислотные остатки с 594 до 679 на последовательности белкаBVH-11-2), клоны, соответствующие последовательности, охватывающей аминокислотные остатки с 658 до 679 были собраны только антителами Н 11 В-11 В 8, что выявляет размещение эпитопа Н 11 В-11 В 8 между аминокислотными остатками 658-679 (SEQ ID NO: 163). Антитела H112-10G9, Н 112-10 А 2 и Н 11 В 11 В 8 являются специфическими в отношении трех отдельных неперекрывающихся эпитопов, расположенных непосредственно на последовательности пептидов, соответствующей аминокислотным остаткам с 594 до 679 (SEQ ID NO:22). Таблица 5. Последовательности пептидов, полученные из скриннинга библиотек генов BVH-3 Пример 4. В этом примере описана иммунизация животных рекомбинантными белками в целях выработки антител, реакционноспособных в отношении BVH-3, BVH-11 и/или BVH-11-2. Была проведена подкожная иммунизация NZW кроликов (Charles River Laboratories, St-Constant,Quebec, Canada). Ее проводили несколькими сериями 50 мкг или 10 мкг очищенного рекомбинантного белка BVH-3M, L-BVH-3AD, NEW1, NEW13 или L-BVH-11 в присутствии 80 мкг адъюванта QulA (Cedarlane Laboratories Ltd, Hornby, Canada). Дважды с трехнедельным интервалом кроликов подпитывали тем же антителом, образцы крови собирали перед каждой иммунизацией, а также с 6 по 28 день после последней иммунизации. Были разработаны образцы предъиммунной сыворотки, и сывороток после первой, второй и третьей инъекции. Иммунную реакцию кроликов оценивали методом твердофазного иммуноферментного анализа, используя рекомбинанты белка BVH-3M (BVH-3M-HisTag) слитый белок/система рЕТ 21) или белка BVH-11M (BVH-11M-HisTag)слитый белок/система рЕТ 21), или суспензии клеток S. pneumoniae Rx-1, ослабленных тепловым действием, в качестве антигенов с покрытием. Титр метода определяли как значение, соответствующее самому высокому разведению, при котором величина поглощения А 410 была на 0,1 выше фона. Антитела, взаимодействующие с эпитопом BVH-3 и/или с эпитопом BVH-11, выявляли после такой иммунизации у всех животных; это показано в приведенной далее таблице 6. В предъиммунной сыворотке не содержались антитела, взаимодействующие с рекомбинантами или антигенами пневмококков. Иммунную реакцию на иммунизацию можно было выявить в сыворотке каждого кролика после однократной инъекции рекомбинантного антигена. Иммунная реакция после вторичной инъекции любым из тестируемых антигенов характеризовалась значительным ростом титра антитела. Интересно, что были получены хорошие титры антител, способных к реакции с клеткамиS.pneumoniae. причем после третьей иммунизации среднее значение титра составляло 52000. Это показывает, что нативные пневмококковые эпитопы прошли экспрессию на рекомбинантых генных продуктах E.coli. Эти данные подтверждают возможность использования генных продуктов BVH-3, BVH-11 и/или BVH-11-2, и антител, полученных на генных продуктах BVH-3, BVH-11 и/или BVH-11-2, в качестве вакцин для предотвращения и лечения пневмококковых инфекций соответственно.- 27006232 Пример 5. В этом примере описана защита животных в эксперименте от смертельной инфекции стрептококков путем введения им антител, созданных из генных продукте BVH-3, BVH-11 или BVH-11-2. Препараты антител с высоким титром были получены из асцитной жидкости мышей, инокулированных внутрибрюшинно клетками гибридомы, выделяющими антитела. Процедуру проводили по методу, описанному Brodeur et al. (J. Immunol Methods 71:265-272,1984). У кроликов, иммунизированныхBVH-3M, собрали образцы сыворотки, это проводилось так, как описано в примере 4. Сыворотку кроликов, собранную после третьей иммунизации, а также асцитную жидкость, применяли для очистки антител осаждением, был использован насыщенный 45-50% сульфат аммония. Препараты антител растворили и подвергли диализу в отношении фосфатно-солевого буфера (ФСБ).CBA/N (xid) мышам (National Cancer Institute, Frederick, MA) внутрибрюшинно вводили или 0,1 мл очищенных антител кролика, или 0,2 мл асцитной жидкости; это осуществляли до проведения внутривенного контрольного заражения, осуществляемого приблизительно 200 КОЕ.S. Pneumoniae. тип 3 штамма WU2. Контрольные мыши получали стерильный ФСБ или антитела,очищенные от предъиммунной сыворотки кроликов или от сыворотки кроликов, иммунизированых антигеном, неродственным рекомбинантному белку N. meningitis. Образцы инокулята S. pneumoniae поместили на пластины с шоколадным агаром для того, чтобы определить количество колониеобразующих единиц и проверить дозу контрольного заражения. Мышей CBA/N выбрали из-за их высокой предрасположенности к инфекции S. pneumoniae. Было установлено, что доза, приводящая к смерти 50% мышейCBA/N, инфицированных внутривенно WU2, составляет 10 КОЕ. Смерти регистрировались с 24 ч интервалом в течение по крайней мере 7 дней. Данные по защите мышей, которым делали инъекции антителом кролика в отношении BVH-3, приведены далее в табл. 7. Из 10 мышей, получавших анти-ВVН-3 антитело, 9 выжили при дозе контрольного заражения; напротив, ни одна из 10 мышей, которым делали инъекцию контрольным антителом или фосфатно-солевым буфером, не выжила. Результаты, показывающие, что антитело, относящееся к молекуле BVH-3-M, осуществляет пассивную защиту даже в том случае, когда введение было сделано после контрольного заражения, продемонстрировали способность анти-ВVН-3 антитела предотвращать смерть даже от уже установленной инфекции. Таблица 7. Защитное действие антител кролика к гену BVH-3-M у CBA/N мышей после проведения контрольного внутривенного заражения пневмококком WU2 В других экспериментах мышам CBA/N вводили внутрибрюшинно 0,1 мл антитела, приготовленного из предъиммунной и иммунной сыворотки; введение проводили за 4 ч до интраназального контрольного заражения 280 КОЕ штамма S. pneumoniae P424, тип 3. Как видно из приведенной далее таблицы 8,все иммунизированные мыши выжили после контрольного заражения, в то время как ни одна из 9 мышей, получавших только антитело предъиммунной сыворотки или буфер, не осталась в живых к 6 дню после инфицирования. Гемокультуры S. pneumoniae на 11 день после контрольного заражения были негативными у всех выживших мышей. Более того, у мышей, получавших моноклональные антитела H11210G9 или смесь H112-10G9 и Н 11 В-7 Е 11, специфические в отношении BVH-11/BVH-11-2, наблюдалась 100 % защита. Таблица 8. Защитное действие пассивной передачи антитела кролика генному продукту BVH-3-M или специфическим антителам анти-BVH-11/BVH-11-2 на мышей CBA/N, испытавших контрольное интраназальное заражение пневмококком Р 4241 Результаты табл. 7 и 8 отчетливо показывают, что иммунизация животных генным продуктом BVH3 (типа BVH-3M) создает защитные антитела, способные в эксперименте предотвратить бактериемическую и пневмонийную инфекции. Данные по защите, полученные для мышей, которым делали инъекцию асцитной жидкостью, приведены далее в табл. 9. Введение 0,2 мл асцитной жидкости несколькими сериями предотвращает смерть от инфекции в эксперименте. Например, серии H112-3 А 4 + H112-10G9 и H112-10G2 + H112-10D7 из 2 моноклональных антител создают полную защиту от инфекции в эксперименте. Эти данные показывают,что антитела, целевым объектом которых являются эпитопы BVH-11 и/или BVH-11-2, создают эффективную защиту. Моноклональные антитела Н 112-3 А 4, H112-10G9, H112-10D7, Н 112-10 А 2, Н 112-3 Е 8,Н 112-10 С 5, Н 11 В-11 В 8, H11B-15G2, Н 11 В-1 С 9, Н 11 В-7 Е 11, H11B-13D5 и Н 11-10 В 8 находились по крайней мере в одной паре моноклональных антител, и оказалось, что они обладают защитными свойствами и реакционной способностью в отношении защитных эпитопов. В табл. 10 и на фиг. 29 приведены итоговые сведения о расположении на молекулах BVH-11-2 эпитопов, обеспечивающих защиту. Все защитные моноклональные антитела Н 112-3 А 4, H112-10G9, H112-10D7, Н 112-10 А 2, Н 112-3 Е 8, Н 11210 С 5, Н 11 В-11 В 8, H11B-15G2, Н 11 В-1 С 9, Н 11 В-7 Е 11, H11B-13D5, а также Н 11-10 В 8 обладали реакционной способностью в отношении белка NEW10, которому в молекуле BVH-11-2 соответствуют аминокислотные остатки с 271 до 838. Из 12 приведенных моноклональных антител 6 антител обладали специфичностью в отношении эпитопа, присутствующего в белке NEW10, а 3 защитных моноклональных антитела распознавали NEW14. Интересно, что моноклональные антитела Н 112-3 А 4 и Н 112-10 С 5 взаимодействовали с отдельными эпитопами, принадлежащими BVH-11-2 и расположенными у карбоксильного конца между аминокислотными остатками с 769 до 837. Моноклональные антитела H11-7G1, Н 116 Е 7 и H3-4F9, реагирующие с эпитопами, охватывающими пневмококковые молекулы BVH-3, BVH-11 иBVH-11-2, не могут осуществлять защиту, даже при их комбинации с защитными моноклональными антителами H112-10G9 или Н 112-11 В 8. Эти моноклональные антитела распознают эпитопы, расположенные у аминного конца молекул BVH-3, BVH-11 и BVH-11-2, содержащих первые 225, 228 и 226 аминокислотных остатка соответственно. Сравнение белковых последовательностей BVH-3, BVH-11 и BVH11-2 выявило, что на аминном участке конца, содержащем эти 225-228 остатки, большое количество аминокислот сохраняется неизменными, их общая идентичность составляет 72,8% (рис. 32). Данные табл. 9 и 10, приведенных далее, предполагают, что защита, выявляемая эпитопами BVH-11 и BVH-11-2, заключается в карбокси-концевом продукте, на белках которого BVH-11 и BVH-11-2 находятся аминокислоты с 229 до 840 и с 227 до 838 соответственно.- 29006232 Таблица 9. Пассивная иммунизация специфическими моноклональными антителами.BVH-11- и/или BVH-11-2- способна защитить мышей от смертельной пневмококковой инфекции в эксперименте Таблица 10. Логически установленное расположение создающих защиту эпитопов на молекулахBVH-11-2
МПК / Метки
МПК: A61K 39/09, C07K 14/315, C12N 15/31
Метки: стрептококковые, антигены
Код ссылки
<a href="https://eas.patents.su/30-6232-streptokokkovye-antigeny.html" rel="bookmark" title="База патентов Евразийского Союза">Стрептококковые антигены</a>
Модифицированные вич-пептиды на основе белка gag p24, антигены, вакцинные композиции, набор для иммуноанализа и способ определения вич-индуцированных антител

Номер патента: 4802
Опубликовано: 26.08.2004
Автор: Сёренсен Биргер
МПК: C07K 14/16, A61K 39/21, A61P 31/18...
Метки: антигены, основе, вич-пептиды, антител, вакцинные, набор, белка, иммуноанализа, способ, композиции, модифицированные, вич-индуцированных, определения
Формула / Реферат:
1. Пептид на основе белка ВИЧ gag p24, включающий по крайней мере девять расположенных подряд аминокислот модифицированной по сравнению с природной аминокислотной последовательности SEQ ID No 1: Xaa1 Xaa2 Xaa3 Xaa4 Xaa5 Xaa6 Ala Xaa8 Xaa9 Gln Thr Pro Trp Xaa14 Xaa15 Xaa16 Xaa17 Xaa18 Val Xaa20 (SEQ ID NO 1), где Xaa в положении 1 пептидного производного означает Lys или Arg; Xaa в положении 2 означает Ala, Gly, Ser или Arg; Xaa в положении 3...
Регуляторные и вспомогательные пептиды вич, антигены, вакцинные композиции, набор для иммуноанализа и способ обнаружения антител, индуцируемых вич

Номер патента: 6210
Опубликовано: 27.10.2005
Автор: Сёренсен Биргер
МПК: G01N 33/569, A61K 39/21, C07K 14/16...
Метки: индуцируемых, композиции, регуляторные, иммуноанализа, вакцинные, антигены, обнаружения, вспомогательные, способ, вич, набор, пептиды, антител
Формула / Реферат:
1. Пептид, происходящий из регуляторного и вспомогательного белка ВИЧ-1, отличающийся тем, что он включает по меньшей мере одну аминокислотную последовательность, которая содержит модификации по сравнению с нативной последовательностью и выбрана из следующих групп последовательностей: Xaa1 Xaa2 Xaa3 Xaa4 Xaa5 Xaa6 Xaa7 Leu Glu Pro Trp Xaa12 His Pro Xaa15 Xaa16 Xaa17 Xaa18 Xaa19 Xaa20 Xaa21 Xaa22 Xaa23 Xaa24 (SEQ ID No. 1), где аминокислоты цепи...
Выделенные, по существу очищенные, поверхностные антигены белковой природы neisseria meningitidis и neisseria gonorrhoeae, их фрагменты и последовательности днк

Номер патента: 1789
Опубликовано: 27.08.2001
Авторы: Риу Клеман, Бродор Бернар Р., Хамел Хосе, Мартэн Денис
МПК: C07K 14/22, A61K 38/16, C12N 1/21...
Метки: природы, neisseria, белковой, днк, последовательности, meningitidis, gonorrhoeae, антигены, очищенные, существу, поверхностные, фрагменты, выделенные
Формула / Реферат:
1. Выделенный, по существу очищенный, высокоустойчивый к протеиназе K поверхностный антиген белковой природы Neisseria meningitidis мол. массы 22 кДа, определенной методом гель-электрофореза в полиамидном геле, иммунологически доступный в клетках более чем 50%, в частности примерно 99%, известных штаммов N. meningitidis, или выделенный, по существу очищенный, высокоустойчивый к протеиназе K поверхностный антиген белковой природы Neisseria...
Предыдущий патент: Способ получения 16,17-[(циклогексилметилен)бис(окси)] -11,21-дигидроксипрегна-1,4-диен-3,20-диона или его 21-изобутирата транскетализацией.
Следующий патент: Комбинированные адъювантные композиции для усиления иммунного ответа у позвоночного животного
Случайный патент: Таблетка меченной изотопом мочевины