Пентаспецифические антитела, способы их получения и применения
Номер патента: 24621
Опубликовано: 31.10.2016
Авторы: Уайт Джон Р., Клегг Стефани Джейн, Гермашевски Фолькер, Добжински Эрик, Эллис Джонатан Х., Йонак Зденка Людмила, Годиллот Алексис Пол, Льюис Алан П.


















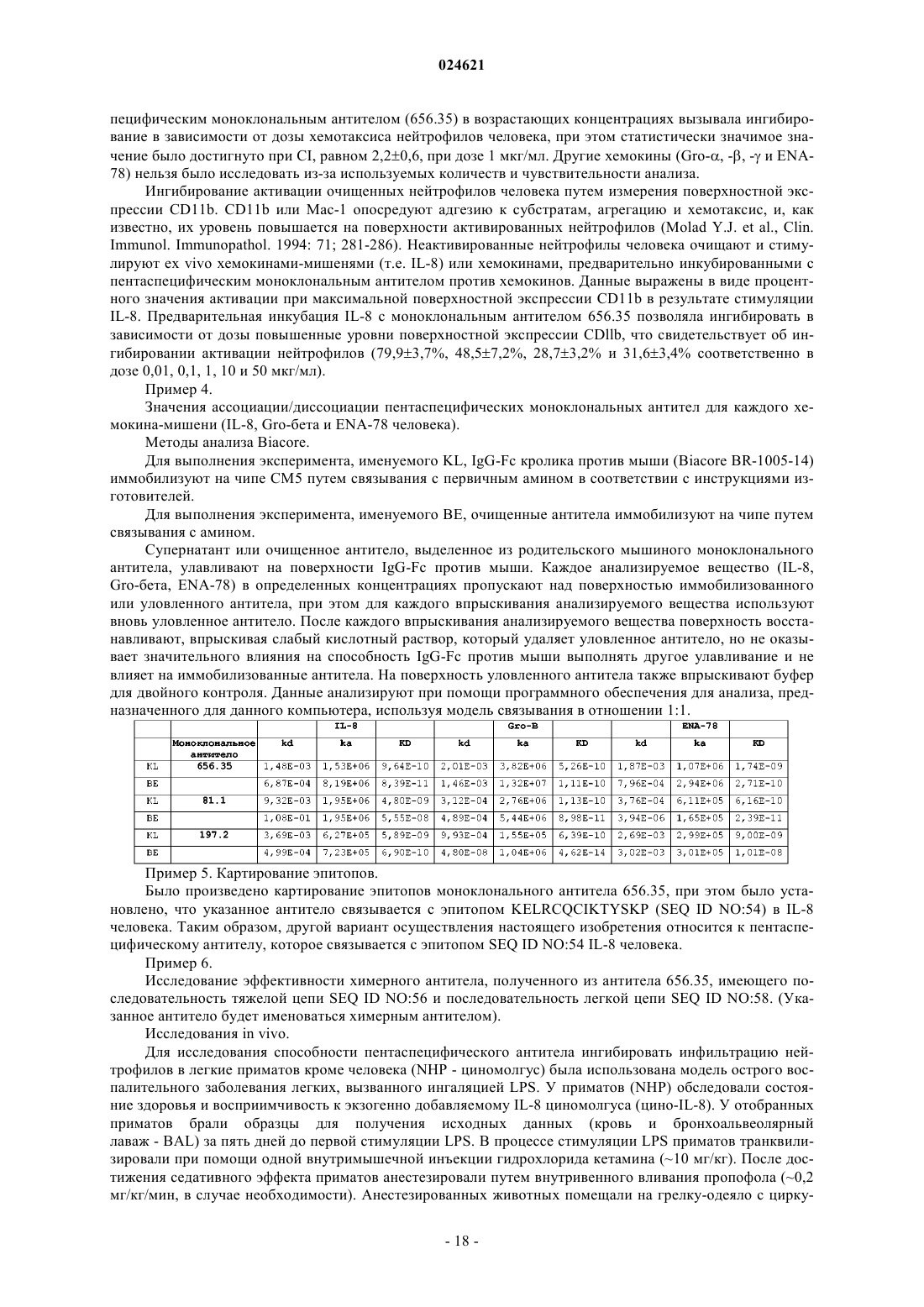








Формула / Реферат
1. Пентаспецифическое антитело, которое перекрестно взаимодействует с IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека, включающее вариабельные области тяжелой и легкой цепей, которые содержат аминокислотные последовательности CDR SEQ ID NO:13, 14 и 15 и SEQ ID NO:16, 17 и 18 соответственно.
2. Антитело по п.1, включающее тяжелую и легкую цепи, содержащие аминокислотные последовательности SEQ ID NO:46 и SEQ ID NO:48, 60, 62 или 64 соответственно.
3. Антитело по п.1, включающее тяжелую и легкую цепи, содержащие аминокислотные последовательности SEQ ID NO:46 и SEQ ID NO:48 соответственно.
4. Антитело по п.1, включающее тяжелую и легкую цепи, содержащие аминокислотные последовательности SEQ ID NO:46 и SEQ ID NO:62 соответственно.
5. Антитело по п.1, включающее тяжелую и легкую цепи, содержащие аминокислотные последовательности SEQ ID NO:46 и SEQ ID NO:60 соответственно.
6. Антитело по п.1, включающее тяжелую и легкую цепи, содержащие аминокислотные последовательности SEQ ID NO:46 и SEQ ID NO:64 соответственно.
7. Антитело по п.1, которое является гуманизированным.
8. Антитело по п.7, включающее константную область IgG1.
9. Антитело по п.7, включающее константную область IgG4.
10. Экспрессирующий вектор, включающий нуклеотидные последовательности, кодирующие вариабельную область тяжелой или легкой цепи антитела, содержащую последовательности CDR SEQ ID NO:13, 14 и 15 или SEQ ID NO:16, 17 и 18 соответственно.
11. Рекомбинантная эукариотическая или прокариотическая клетка, продуцирующая антитело по пп.1-9, содержащая экспрессирующий вектор по п.10.
12. Способ продуцирования антитела по одному из пп.1-9 в единичной клетке-хозяине, включающий следующие стадии:
(i) трансформация указанной единичной клетки-хозяина первой последовательностью ДНК, кодирующей, по меньшей мере, вариабельную область тяжелой цепи антитела, включающую области CDR SEQ ID NO:13, 14 и 15; и второй последовательностью ДНК, кодирующей, по меньшей мере, вариабельную область легкой цепи антитела, включающую области CDR SEQ ID NO:16, 17 и 18; и
(ii) экспрессия указанной первой последовательности ДНК и указанной второй последовательности ДНК, в результате которой указанные тяжелую и легкую цепи антитела получают в виде отдельных молекул в указанной трансформированной единичной клетке-хозяине.
13. Способ по п.12, в котором указанные первая и вторая последовательности ДНК присутствуют в разных векторах или указанные первая и вторая последовательности ДНК присутствуют в одном векторе.
14. Фармацевтическая композиция для лечения или профилактики ХОБЛ, остеоартрита, ревматоидного артрита, эрозивного артрита, астмы, атеросклероза, воспалительного заболевания кишечника, псориаза, отторжения трансплантата, подагры, рака, острого поражения легких, острого заболевания легких, сепсиса, ARDS, заболевания периферических артерий, системного склероза, респираторного дистресс-синдрома у новорожденных, обострения астмы и COPD, муковисцидоза, диффузного панбронхиолита, поражения, вызванного реперфузией, и/или эндометриоза, включающая эффективное количество антитела по одному из пп.1-9 и фармацевтически приемлемый носитель.
15. Способ уменьшения хемотаксиса нейтрофилов у пациента путем введения антитела по одному из пп.1-9.
Текст
ПЕНТАСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ Изобретение относится к пентаспецифическому антителу, обладающему широкой специфичностью. В частности, антитело по настоящему изобретению связывается (перекрестно взаимодействует) с IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека. Клегг Стефани Джейн (GB),Добжински Эрик (US), Эллис Джонатан Х., Гермашевски Фолькер(GB), Годиллот Алексис Пол, Йонак Зденка Людмила (US), Льюис Алан П.(71)(73) Заявитель и патентовладелец: СМИТКЛАЙН БИЧАМ КОРПОРЕЙШН (US) Перекрестная ссылка на родственную заявку По заявке на данное изобретение испрашивается приоритет временной заявки на патент США 60/912229, поданной 17 апреля 2007 г., и 61/044132, поданной 11 апреля 2008 г., которые полностью включены в настоящее описание изобретения путем ссылки. Область техники, к которой относится изобретение Настоящее изобретение относится к антителу, обладающему широкой специфичностью. В частности, антитело по настоящему изобретению связывается (перекрестно взаимодействует) с IL-8, Gro-альфа,Gro-бета, Gro-гамма и ENA-78 человека. Уровень техники Опубликованные данные и отчеты свидетельствуют о том, что в целом ряде заболеваний имеют место повышенные уровни членов надсемейства ELRCXC хемокинов CXCL. Семейство CXCL насчитывает в общей сложности 16 членов. Как сообщалось в научной литературе, уровни хемокинов повышены в целом ряде воспалительных заболеваний, включая COPD, в котором CXCL 1-3, 5 и 8 известны также какOppenheim, J.J. and Feldman, M. ed., Academic Press, London, 1061-1067, Matsushima, K. et el., J. Exp. Med. 1988: 167, 1883-1893) (Am. J. Respir. Crit. Care Med., 163: 349-355, 2001, Am. J. Respir. Crit. Care Med.,168:968-975, 2003, Thorax, 57: 590-595, 2002). Теоретически обосновано, что продолжительная и повышенная экспрессия указанных хемокинов может вызывать развитие таких заболеваний, как COPD, остеоартрит, ревматоидный артрит, эрозивный артрит, астма, атеросклероз, воспалительное заболевание кишечника, псориаз, отторжение трансплантата, подагра, рак, острое поражение легких, острое заболевание легких, сепсис, ARDS, заболевание периферических артерий, системный склероз, респираторный дистресс-синдром у новорожденных, обострение астмы и COPD, муковисцидоз, диффузный панбронхиолит, поражение, вызванное реперфузией, или эндометриоз. Указанные хемокины СХС, как известно,стимулируют хемотаксис нейтрофилов в результате привлечения и активации рецепторов CXCR1 и/илиCXCR2. Таким образом, ингибирование указанных хемокинов может препятствовать инфильтрации клеток воспалительного инфильтрата в ткань легких и, следовательно, предотвращать поражение ткани. Целью настоящего изобретения является ингибирование активации рецепторов CXCR1 и CXCR2 при помощи антитела, способного связываться с IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека, т.е. пентаспецифического антитела. Сущность изобретения Настоящее изобретение относится к антителу (иммуноглобулину), которое обладает широкой специфичностью, заключающейся в одном иммуноглобулине. В частности, антитело по настоящему изобретению связывается (перекрестно взаимодействует) с IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека. Одним объектом настоящего изобретения является выделенное антитело, обладающее широкой специфичностью (или перекрестно взаимодействующее) к IL-8, Gro-альфа, Gro-бета, Gro-гамма иENA-78 человека, поэтому антитело по настоящему изобретению было определено как пентаспецифическое (или пан-ELR) антитело. Указанное определение антитела включает антигенсвязывающую часть(или фрагмент) антитела, которая связывается с IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека. Пентаспецифическое антитело по настоящему изобретению предпочтительно является мышиным моноклональным, химерным, человеческим или гуманизированным антителом. Во избежание неясности следует отметить, что пентаспецифическое антитело по настоящему изобретению не обязательно связывается только с антигенами IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека, оно может также связываться с другими родственными белками (такими как GCP-2 человека, или может даже связываться с отличными от человеческих ортологами IL-8, Gro-альфа, Gro-бета, Gro-гамма, ENA-78 и GCP-2); другими словами, пентаспецифическое антитело по настоящему изобретению, как минимум, связывается с IL8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека. Пентаспецифическое антитело по настоящему изобретению предпочтительно связывается с каждым из IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека при значениях константы равновесияKD менее 107 M, более предпочтительно менее 10-8 М и еще более предпочтительно менее 10-9 М, определяемых путем измерения поверхностного плазмонного резонанса. Измерение поверхностного плазмонного резонанса обычно выполняют в соответствии с приведенным ниже описанием. Один вариант осуществления настоящего изобретения относится к способу уменьшения хемотаксиса нейтрофилов путем ингибирования активации рецепторов CXCR1 и CXCR2 в результате нейтрализации IL-8, Gro-альфа, Gro-бета, Gro-гамма, GCP-2 и ENA-78 человека пентаспецифическим антителом по настоящему изобретению. Один вариант осуществления настоящего изобретения относится к способу уменьшения хемотаксиса нейтрофилов у пациента путем введения пентаспецифического антитела по настоящему изобретению. В одном варианте осуществления изобретения пентаспецифическое антитело связывается с эпитопом KELRCQCIKTYSKP (SEQ ID NO:54) в IL-8 человека. В одном варианте осуществления изобретения пентаспецифическое антитело по настоящему изобретению получают способом по протоколу RIMM (Kilpatrick, K.E., et al. Hybridoma. 1997: 16, 381), используя смесь (коктейль) IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека вместе с набором из пяти множественных антигенных пептидов (MAP), в которых каждое звено содержит одну отдельную последовательность из полипептидов с ID NO:49-53. Не ограничиваясь какой-либо теорией, следует отметить, что MAP выполняют две функции в протоколе иммунизации. Во-первых, MAP обеспечивают селективное множественное представление известной аминокислотной последовательности-мишени иммунной системе хозяина. Во-вторых, вызывают увеличение массы благодаря многочисленным копиям данной последовательности, связанным ядром,таким как лизин, но не ограничиваясь им, что повышает иммуногенность последовательности по сравнению с отдельными пептидами (Francis, J.P., et al., Immunology, 1991: 73; 249, Schott, M.E., et al., Cell.MAP, используемые в настоящем изобретении, состоят из многочисленных копий консервативных последовательностей-мишеней (например, SEQ ID NO:49-53), обнаруженных в областях ELRCXC иGPHCA и рядом с указанными областями хемокинов-мишеней. Типичный набор MAP показан на фиг. 1. В одном варианте осуществления изобретения пентаспецифическое антитело включает вариабельные области тяжелой и легкой цепей, содержащие соответственно аминокислотные последовательностиCDR SEQ ID NO:13, 14 и 15 и SEQ ID NO:16, 17 и 18. Один вариант осуществления настоящего изобретения относится к экспрессирующему вектору,включающему нуклеотидные последовательности, кодирующие вариабельную область тяжелой или легкой цепи пентаспецифического антитела, содержащую соответственно последовательности CDRSEQ ID NO:13, 14 и 15 или SEQ ID NO:16, 17 и 18. В одном варианте осуществления изобретения пентаспецифическое антитело имеет тяжелую и легкую цепи, включающие соответственно аминокислотные последовательности SEQ ID NO:46 иSEQ ID NO:48, 60, 62 или 64. В одном варианте осуществления изобретения пентаспецифическое антитело имеет тяжелую и легкую цепи, включающие соответственно аминокислотные последовательности SEQ ID NO:46 иSEQ ID NO:48. В одном варианте осуществления изобретения пентаспецифическое антитело имеет тяжелую и легкую цепи, включающие соответственно аминокислотные последовательности SEQ ID NO:46 иSEQ ID NO:60. В одном варианте осуществления изобретения пентаспецифическое антитело имеет тяжелую и легкую цепи, включающие соответственно аминокислотные последовательности SEQ ID NO:46 иSEQ ID NO:62. В одном варианте осуществления изобретения пентаспецифическое антитело имеет тяжелую и легкую цепи, включающие соответственно аминокислотные последовательности SEQ ID NO:46 иSEQ ID NO:64. Один вариант осуществления настоящего изобретения относится к рекомбинантной эукариотической или прокариотической клетке-хозяину, продуцирующей пентаспецифическое антитело, имеющее тяжелую или легкую цепь, включающую соответственно аминокислотную последовательностьSEQ ID NO:46 или SEQ ID NO:48, 60, 62 или 64. Один вариант осуществления настоящего изобретения относится к рекомбинантной эукариотической или прокариотической клетке-хозяину, продуцирующей пентаспецифическое антитело, имеющее тяжелую и легкую цепи, включающие соответственно аминокислотные последовательностиSEQ ID NO:46 и SEQ ID NO:48, 60, 62 или 64. Один вариант осуществления настоящего изобретения относится к экспрессирующему вектору,включающему полинуклеотидные последовательности, кодирующие пентаспецифическое антитело,включающее тяжелую и легкую цепи, содержащие соответственно аминокислотные последовательностиSEQ ID NO:46 и SEQ ID NO:48, 60, 62 или 64. Один вариант осуществления настоящего изобретения относится к экспрессирующему вектору,включающему полинуклеотидную последовательность, кодирующую полипептид, содержащий аминокислотную последовательность SEQ ID NO:46, 48, 60, 62 или 64. Один вариант осуществления настоящего изобретения относится к способу продуцирования пентаспецифического антитела (иммуноглобулина) в единичной клетке-хозяине, который включает следующие стадии:(i) трансформацию указанной единичной клетки-хозяина первой последовательностью ДНК, кодирующей, по меньшей мере, вариабельную область тяжелой цепи иммуноглобулина, включающую области CDR SEQ ID NO:13, 14 и 15; и второй последовательностью ДНК, кодирующей, по меньшей мере,вариабельную область легкой цепи иммуноглобулина, включающую области CDR SEQ ID NO:16, 17 и 18; и(ii) экспрессию указанной первой последовательности ДНК и указанной второй последовательности ДНК, в результате которой указанные тяжелую и легкую цепи иммуноглобулина получают в виде отдельных молекул в указанной трансформированной единичной клетке-хозяине,причем этот способ может быть выполнен таким образом, что указанные первая и вторая последовательности ДНК присутствуют в разных векторах или указанные первая и вторая последовательности ДНК присутствуют в одном векторе. Один вариант осуществления настоящего изобретения относится к антителу, которое полностью или частично блокирует связывание любого из вышеуказанных пентаспецифических антител с IL-8,Gro-альфа, Gro-бета, Gro-гамма, ENA-78 и GCP-2 человека при выполнении иммуноанализа, такого как анализ ELISA. В одном варианте осуществления изобретения частичное блокирование происходит тогда,когда антитело блокирует связывание пентаспецифического антитела более чем на 10, 20, 40 или 50%. Один вариант осуществления настоящего изобретения относится к антителу, которое конкурирует за связывание с любым из вышеуказанных пентаспецифических антител с IL-8, Gro-альфа, Gro-бета,Gro-гамма, ENA-78 и GCP-2 человека. Один вариант осуществления настоящего изобретения относится к антителу, которое полностью или частично блокирует связывание любого из вышеуказанных пентаспецифических антител с эпитопомKELRCQCIKTYSKP (SEQ ID NO:54) в IL-8 человека при выполнении иммуноанализа, такого как анализELISA. В одном варианте осуществления изобретения частичное блокирование происходит тогда, когда антитело блокирует связывание пентаспецифического антитела более чем на 10, 20, 40 или 50%. Один вариант осуществления настоящего изобретения относится к антителу, которое конкурирует с любым из вышеуказанных пентаспецифических антител за связывание с эпитопом KELRCQCIKTYSKP(SEQ ID NO:54) в IL-8 человека. Один вариант осуществления настоящего изобретения относится к композиции, включающей вышеуказанное пентаспецифическое антитело и фармацевтически приемлемый носитель. В одном варианте осуществления изобретения вышеуказанным млекопитающим является человек. Описание чертежей На фиг. 1 показан типичный набор MAP, используемый для создания пентаспецифического антитела. Во избежание неясности изображены пять пептидных звеньев MAP. Каждое звено содержит одну идентичную аминокислотную последовательность, выбранную из линейных пептидов SEQ ID NO:49-53. На фиг. 2 приведены данные проточной цитометрии, позволяющие сравнивать нейтрофилы, присутствующие в образцах бронхоальвеолярного лаважа (BAL). Представлены исследования 1, 2 и 3. Столбцы черного цвета показывают исходные данные, полученные за 5 дней до стимуляции LPS. Столбцы серого цвета показывают данные BAL, полученные через 6 ч после стимуляции LPS. Введение химерного антитела (HcLc) значительно и в зависимости от дозы ингибировало инфильтрацию нейтрофилов. Заштрихованные столбцы серого цвета показывают данные, полученные через 24 ч (n=6, р 0,05,р 0,01, планки погрешностей представляют стандартное отклонение). На фиг. 3 показан гематологический анализ всех циркулирующих в крови нейтрофилов (подсчет клеток). Внутривенная инъекция 1 мг/кг (незакрашенные квадраты, ) и 10 мг/кг (закрашенные кружки, ) химерного антитела (HcLc) предотвращала стимуляцию LPS циркулирующих в крови нейтрофилов путем ингаляции. Введение как 1 мг/кг, так и 10 мг/кг вызывает значительное уменьшение симптомов по сравнению с наполнителем NHP и введение 10 мг/кг также вызывает значительное уменьшение по сравнению с дозой, равной 1 мг/кг . Данные представлены в виде общего подсчета клеток в 1 мкл крови (n=6, р 0,05 по сравнению с 1 мг/кг, р 0,01 по сравнению с наполнителем, планки погрешностей представляют стандартное отклонение). На фиг. 4 показано ингибирование активации нейтрофилов, стимулированных IL-8 человека (повышенная поверхностная экспрессия CD11b). Гуманизированные пентаспецифические антитела предварительно инкубировали в разных концентрациях с 10 нМ hIL-8 до добавления к очищенным нейтрофилам человека. Данные выражены в виде средних значений для 4 разных доноров (n=4). Планки представляют стандартную ошибку. Все образцы сравнивали с очищенными нейтрофилами, стимулированными только hIL-8 (отсутствие моноклонального антитела до добавления к очищенным нейтрофилам). Гуманизированные конструкции H0L10 и HOMO более эффективно ингибируют поверхностную экспрессиюCD11b, стимулированную hIL-8. На фиг. 5 А показано связывание моноклонального антитела 656.35 с хемокинами-мишенями человека. На фиг. 5 В показано связывание химерного моноклонального антитела (HcLc) с хемокинамимишенями человека. На фиг. 5 С показано связывание гуманизированных моноклональных антител с хемокинамимишенями человека. Подробное описание изобретения Термин "выделенное пентаспецифическое антитело" или просто "пентаспецифическое антитело" в значении, использованном в настоящем описании изобретения, означает антитело, которое связывается и, следовательно, перекрестно взаимодействует с IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека, по существу, не содержит других антител, обладающих другой антигенной специфичностью, и имеет моносостав. Во избежание неясности следует отметить, что пентаспецифическое антитело по настоящему изобретению не обязательно связывается только с антигенами IL-8, Gro-альфа, Gro-бета, Gro-гамма иENA-78 человека, оно может также связываться с другими родственными белками, такими как GCP-2 человека, или даже может связываться с отличными от человеческих ортологами IL-8, Gro-альфа,Gro-бета, Gro-гамма, ENA-78 и GCP-2; другими словами, пентаспецифическое антитело по настоящему изобретению, как минимум, связывается с IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека. Кроме того, выделенное пентаспецифическое антитело, по существу, не содержит других клеточных и/или химических веществ. Пентаспецифическое антитело в значении, использованном в данном контексте, включает также антигенсвязывающие фрагменты и/или производные, обладающие характеристиками связывания "родительского" пентаспецифического антитела. Пентаспецифическое антитело по настоящему изобретению может включать две идентичные тяжелые цепи и две идентичные легкие цепи,образующие типичную билатерально симметричную молекулу иммуноглобулина, состоящую из двух гетеродимеров, каждый из которых включает тяжелую цепь и легкую цепь. Таким образом, пентаспецифическое антитело по настоящему изобретению может включать две копии одного антигенсвязывающего домена, образованного в результате ассоциации одной тяжелой цепи с одной легкой цепью. Пентаспецифическое антитело по настоящему изобретению, несмотря на наличие связывающего домена только одного типа, может связываться и, следовательно, взаимодействовать с IL-8, Gro-альфа, Gro-бета, Groгамма и ENA-78 человека. В используемом в описании значении термин "антитело" означает также "иммуноглобулин". В используемом в описании значении термин "антитело, перекрестно взаимодействующее с" означает, что указанное антитело связывается не только с одним антигеном, но и также с другими антигенами. Термин "нейтрализация" в используемом в описании значении означает частичное или полное ингибирование по меньшей мере одной биологической активности IL-8, Gro-альфа, Gro-бета, Gro-гамма,GCP-2 или ENA-78 человека. Например, одной из биологических активностей IL-8, Gro-альфа, Gro-бета,Gro-гамма, GCP-2 или ENA-78 человека является их способность индуцировать хемотаксис нейтрофилов. Одним способом измерения кинетики связывания антитела является измерение поверхностного плазмонного резонанса. Термин "поверхностный плазмонный резонанс" в используемом в описании значении означает оптическое явление, которое позволяет анализировать биоспецифические взаимодействия в реальном времени путем обнаружения изменений в концентрации белков в биосенсорной матрице,например, при помощи системы BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, N.J.). Для ознакомления с более подробным описанием см. публикации Jonsson, U., et al. (1993), Ann. Biol. Clin. 51:19-26; Jonsson, U., et al. (1991), Biotechniques 11:620-627; Johnsson, В., et al. (1995), J. Mol.Recognit. 8:125-131 и Johnnson, В., et al. (1991), Anal. Biochem. 198:268-277. Термин "эпитоп" означает детерминанту белка, способную специфически связываться с антителом. Эпитопы обычно состоят из химически активных поверхностных групп молекул, таких как боковые цепи аминокислот или сахаров, и имеют специфическую трехмерную структуру, а также характерные заряды. Термин "моноклональное антитело" или mAb (в отличие от поликлонального антитела) в используемом в описании значении означает молекулы антител, имеющие одинаковый молекулярный состав. Например, моноклональное антитело, выделенное у мыши (мышиное моноклональное антитело), может быть получено методом образования гибридом, таким как стандартная методика образования гибридом Колера и Милстейна (Kohler and Milstein). Антитело-продуцирующие клетки могут быть получены у субъекта и использованы для получения моноклональных антител стандартными методами, такими как метод гибридом, первоначально описанный Колером и Милстейном (1975, Nature, 256:495-497) (см. также публикации Brown et al. (1981), J.Yeh et al. (1982), Int. J. Cancer 29:269-75). Методика получения гибридом моноклонального антитела хорошо известна (см. публикации R.H. Kenneth, in Monoclonal Antibodies: A New Dimension In BiologicalAnalyses, Plenum Publishing Corp., New York, N.Y. (1980); E.A. Lerner (1981), Yale J. Biol. Med., 54:387402; M.L. Gefter et al. (1977), Somatic Cell Genet., 3:231-36). Термин "трансфектома" в используемом в описании значении означает рекомбинантную эукариотическую клетку-хозяина, экспрессирующую антитело, такую как клетки СНО, клетки NS/0, клеткиHEK-293, растительные клетки или грибы, включая дрожжевые клетки. В используемом в описании значении термин "специфическое" связывание означает связывание антитела с заранее определенным антигеном. Антитело обычно связывается при значениях константы равновесия KD, равных примерно 110-7 М или меньше, и связывается с заранее определенным антигеном со сродством, соответствующим значению KD, которое по меньшей мере на два порядка величины меньше сродства связывания с неспецифическим антигеном (например, BSA, казеин), отличным от заранее определенного антигена или близко родственного антигена. Фразы "антитело, узнающее антиген" и"антитело, специфичное к антигену" имеют взаимозаменяемые значения с термином "антитело, которое специфически связывается с антигеном". Термин "kd" (с-1) в используемом в описании значении означает константу скорости диссоциации определенного взаимодействия антитело-антиген. Термин "ka" (Мс-1) в используемом в описании значении означает константу скорости ассоциации определенного взаимодействия антитело-антиген. Термин "KD" (М) в используемом в описании значении означает константу равновесия определенного взаимодействия антитело-антиген, которую получают в результате деления kd на ka. Термин "консервативные модификации последовательностей" для модификаций нуклеотидных и аминокислотных последовательностей означает изменения, которые не оказывают существенного влияния или не изменяют характеристики связывания антитела, кодируемого данной нуклеотидной последовательностью или содержащего такую аминокислотную последовательность. Такие консервативные модификации последовательностей включают замены, добавления и делеции нуклеотидов и аминокислот. Модификации могут быть введены в последовательности стандартными методами, известными в данной области, такими как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез. Консервативные замены аминокислот включают такие замены, в которых аминокислотный остаток заменяется другим аминокислотным остатком, имеющим подобную боковую цепь. В данной области известны семейства аминокислотных остатков, имеющих подобные боковые цепи. Указанные семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, прогнозируемый заменимый аминокислотный остаток в антителе, имеющем рассмотренную последовательность, предпочтительно заменяют другим аминокислотным остатком из семейства с такой же боковой цепью. Таким образом, одним объектом настоящего изобретения является пентаспецифическое антитело по настоящему изобретению, включающее все консервативные модификации рассмотренных аминокислотных последовательностей. В объем настоящего изобретения входят также "производные" описанных аминокислотных последовательностей, в которых один или несколько аминокислотных остатков являются производными остатками, полученными, например, путем ацилирования или гликозилирования, которые не оказывают существенного влияния и не изменяют характеристики связывания антитела, содержащего указанные аминокислотные последовательности. Термин "по существу, идентичные" применительно к нуклеиновым кислотам означает, что две нуклеиновые кислоты или их последовательности являются идентичными при оптимальном совмещении и сравнении с соответствующими инсерциями или делециями нуклеотидов по меньшей мере примерно у 80% нуклеотидов, обычно по меньшей мере примерно у 90-95% и более предпочтительно по меньшей мере примерно у 98-99,5% нуклеотидов. Альтернативно, по существу, идентичными нуклеиновые кислоты являются тогда, когда сегменты гибридизируют в условиях селективной гибридизации с комплементом цепи. Термин "идентичный" применительно к нуклеотидным и аминокислотным последовательностям означает степень идентичности двух нуклеотидных или аминокислотных последовательностей при оптимальном совмещении и сравнении с соответствующими инсерциями или делециями. Альтернативно,по существу, идентичными последовательности являются тогда, когда сегменты ДНК гибридизируют в условиях селективной гибридизации с комплементом цепи. Процентное значение идентичности двух последовательностей является функцией числа идентичных положений в указанных последовательностях (т.е. % идентичности = число идентичных положений/общее число положений, умноженное на 100) с учетом числа разрывов и длины каждого разрыва,которые должны быть введены для оптимального сравнения двух последовательностей. Сравнение последовательностей и определение процентного значения идентичности двух последовательностей может быть выполнено при помощи математического алгоритма, описанного в приведенных ниже неограничивающих примерах. Процентное значение идентичности двух нуклеотидных или полипептидных последовательностей может быть определено при помощи программы GAP в пакете программного обеспечения GCG (представлен на сайте http://www.gcg.com) с использованием матрицы NWSgapdna. CMP, весового коэффициента разрыва, равного 40, 50, 60, 70 или 80, и весового коэффициента длины разрывов, равного 1, 2, 3, 4,5 или 6. Процентное значение идентичности двух нуклеотидных или аминокислотных последовательностей можно определить, используя алгоритм Е. Мейерса и В. Миллера (Е. Meyers and W. Miller, Comput.Appl. Biosci., 4:11-17 (1988 который включен в программу ALIGN (версия 2.0), таблицу весовых коэффициентов остатков РАМ 120, штраф за длину разрыва, равный 12, и штраф за разрыв, равный 4. Кроме того, процентное значение идентичности двух аминокислотных последовательностей можно определить,используя алгоритм Нидлмана и Ванша (Needleman and Wunsch, J. Mol. Biol. 48:444-453 (1970, который включен в программу GAP пакета программного обеспечения GCG (представлен на сайтеhttp://www.gcg.com), матрицу Blossum 62 или матрицу РАМ 250, весовой коэффициент разрыва, равный 16, 14, 12, 10, 8, 6 или 4, и весовой коэффициент длины разрыва, равный 1, 2, 3, 4, 5 или 6. Нуклеиновая кислота является "функционально связанной" при нахождении в функциональном взаимодействии с последовательностью другой нуклеиновой кислоты. Например, промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию данной последовательности. Что касается транскрипции регуляторных последовательностей, то термин "функционально связанный" означает, что связанные последовательности ДНК являются смежными и, при необходимости, соединяют кодирующие области двух белков, которые являются смежными и находятся в рамке считывания. Термин "функционально связанные" применительно к переключающим последовательностям означает, что указанные последовательности могут осуществлять рекомбинацию на этапе переключения. Термин "вектор" в используемом в описании значении означает молекулу нуклеиновой кислоты,способную транспортировать связанную с ним другую нуклеиновую кислоту. Одним типом вектора является "плазмида", которая представляет собой кольцевую петлю двухцепочечной ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы могут автономно реплицировать в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную природу репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть встроены в геном клеткихозяина при введении в клетку-хозяина, после чего указанные векторы реплицируют вместе с геномом хозяина. Кроме того, некоторые векторы могут направлять экспрессию генов, с которыми они функционально связаны. Такие векторы определяются в настоящем описании изобретения как "рекомбинантные экспрессирующие векторы" (или просто "экспрессирующие векторы"). Как правило, экспрессирующие векторы, используемые в методах рекомбинантных ДНК, часто имеют форму плазмид. В настоящем описании изобретения термины "плазмида" и "вектор" могут иметь взаимозаменяемые значения, так как плазмида является наиболее часто используемой формой вектора. Однако в объем настоящего изобретения входят также другие формы экспрессирующих векторов, такие как вирусные векторы (например,ретровирусы с дефектом репликации, аденовирусы и аденоассоциированные вирусы), которые выполняют такие же функции. Термин "рекомбинантная клетка-хозяин" (или просто "клетка-хозяин") в используемом в описании значении означает клетку, в которую введен рекомбинантный экспрессирующий вектор. Очевидно, что такие термины относятся не только к определенной клетке, но и к потомству такой клетки. Так как в последующих поколениях могут появиться определенные модификации вследствие мутации или влияния окружающих условий, такое потомство фактически не может быть идентично материнской клетке, но все же входит в определение термина "клетка-хозяин" в значении, использованном в настоящем описании изобретения. Рекомбинантные клетки-хозяева включают, например, трансфектомы, такие как клетки СНО, клетки NS/0 и лимфоциты. В используемом в описании значении термин "субъект" означает человека или животное, отличное от человека. Термин "животное, отличное от человека" означает всех позвоночных, например млекопитающих и немлекопитающих, таких как приматы кроме человека, овцы, собаки, коровы, куры, амфибии, рептилии и т.д. 1. Структуры антител. Интактные антитела. Интактные антитела обычно являются гетеромногомерными гликопротеинами, имеющими по меньшей мере две тяжелые и две легкие цепи. Помимо IgM интактные антитела являются гетеротетрамерными гликопротеинами с длиной цепи около 150 кДа, состоящими из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь обычно связана с тяжелой цепью одной ковалентной дисульфидной связью, причем число дисульфидных связей между тяжелыми цепями разных изотипов иммуноглобулина может быть различным. Все тяжелые и легкие цепи имеют также дисульфидные мостики внутри цепи. У одного конца каждой тяжелой цепи находится вариабельная область (VH), за которой следует ряд константных областей. Каждая легкая цепь имеет вариабельную об-6 024621 ласть (VL) и константную область у другого конца; константная область легкой цепи совмещается с первой константной областью тяжелой цепи, и вариабельная область легкой цепи совмещается с вариабельной областью тяжелой цепи. Легкие цепи антител у большинства позвоночных могут быть отнесены к одному из двух типов, именуемых каппа и лямбда на основании аминокислотной последовательности константной области. В зависимости от аминокислотной последовательности константной области тяжелых цепей человеческие антитела могут быть отнесены к пяти разным классам: IgA, IgD, IgE, IgG и IgM.IgG и IgA могут быть далее подразделены на подклассы IgG1, IgG2, IgG3 и IgG4 и IgA1 и IgA2. Варианты существуют у мышей и крыс, имеющих по меньшей мере IgG2a, IgG2b. Вариабельная область сообщает антителу специфичность связывания с определенными областями, характеризующимися конкретной вариабельностью, которые именуются областями, определяющими комплементарность (CDR). Более консервативные части вариабельной области именуются каркасными областями (FR). Вариабельные области тяжелых и легких цепей интактных антител включают четыре каркасные области, соединенные тремя CDR. CDR в каждой цепи удерживаются в непосредственной близости друг от друга каркасными областями и вместе с CDR из другой цепи способствуют образованию антигенсвязывающего центра антител. Константные области непосредственно не участвуют в связывании антитела с антигеном, но выполняют разные эффекторные функции, например опосредуют антителозависимую клеточноопосредованную цитотоксичность (ADCC), участвуют в фагоцитозе благодаря связыванию с рецепторомFc, определяют период полувыведения/клиренс при помощи неонатального рецептора Fc (FcRn) и комплементзависимую цитотоксичность при помощи компонента C1q пути активации комплемента. Константная область IgG2 человека не может активировать комплемент по классическому пути активации комплемента или опосредовать антителозависимую клеточно-опосредованную цитотоксичность. Константная область IgG4 не может активировать комплемент по классическому пути активации комплемента и может лишь слабо опосредовать антителозависимую клеточно-опосредованную цитотоксичность. Антитела, у которых, по существу, отсутствуют указанные эффекторные функции, могут именоваться"нелитическими" антителами. Человеческие антитела. Человеческие антитела могут быть получены рядом методов, известных специалистам в данной области. Человеческие антитела могут быть получены методом гибридом с использованием линий миеломных клеток человека или гетеромиеломных клеток мыши-человека; см. публикации Kozbor, J. Immunol. 133, 3001 (1984) и Brodeur, Monoclonal Antibody Production Techniques and Applications, p.51-63 (MarcelDekker Inc, 1987). Альтернативные методы включают использование фаговых библиотек или трансгенных мышей на основе наборов V-областей человека (см. публикации Winter G. (1994), Annu. Rev. Immunol. 12, 433-455,Green L.L. (1999), J. Immunol. methods 231, 11-23). В настоящее время имеется несколько линий трансгенных мышей, у которых мышиные локусы иммуноглобулина заменены сегментами генов иммуноглобулина человека (см. публикации Tomizuka K.Genetics, 15, 146-156). В результате антигенной стимуляции такие мыши могут продуцировать набор человеческих антител, из которых могут быть выбраны антитела, представляющие интерес. Особого внимания заслуживает система Trimera (см. публикацию Eren R. et al. (1998),Immunology 93:154-161), в соответствии с которой лимфоциты человека трансплантируют облученным мышам, а именно система отобранных лимфоцитных антител (SLAM, см. публикацию Babcook et al.,PNAS (1996), 93:7843-7848), в которой лимфоциты человека (или другие разновидности) эффективно подвергают процедуре массового образования антител in vitro с последующим выполнением процедуры неразрушающего, ограниченного разведения и отбора, и система Xenomouse II (Abgenix Inc). Альтернативным подходом является метод Morphodoma компании Morphotek Inc. Для получения человеческих антител (и их фрагментов) может быть использован метод отображения на фаге, см. публикации McCafferty; Nature, 348, 552-553 (1990) и Griffiths A.D. et al. (1994), EMBO,13:3245-3260. В соответствии с указанным методом гены V-области антитела клонируют в рамке считывания в главном или минорном гене оболочки белка нитевидного бактериофага, такого как М 13 или fd, и отображают (обычно при помощи фага-хелпера) в виде функциональных фрагментов антитела на поверхности фаговой частицы. На основании функциональных свойств антитела отбирают ген, кодирующий антитело, обладающее указанными свойствами. Метод отображения на фаге может быть использован для отбора антиген-специфических антител из библиотек, полученных из В-клеток человека, взятых у субъектов, страдающих вышеуказанным заболеванием или нарушением, или, альтернативно, у неиммунизированных людей-доноров (см. публикацию Marks; J. Mol. Bio. 222, 581-597, 1991). Если желательно получить интактное человеческое антитело, включающее Fc-область, необходимо реклонировать фрагмент, выделенный из антитела, отображенного на фаге, в экспрессирующих векторах млекопитающего, включающих требуемые константные области, и получить линии клеток, устойчиво экспрессирующих данное антитело. Метод созревания аффинности (Marks; Bio/technol. 10, 779-783 (1992 может быть использован для улучшения сродства связывания, при этом сродство первичного человеческого антитела улучшается в результате последовательной замены V-областей Н- и L-цепей естественными вариантами и отбора, выполняемого на основании улучшенного сродства связывания. В настоящее время известны также варианты рассмотренного метода, такие как "импринтинг эпитопа"; см. WO 93/06213. См. также публикациюWaterhouse; Nucl. Acids Res. 21, 2265-2266 (1993). Химерные и гуманизированные антитела. Использование интактных антител, отличных от человеческих, при лечении заболеваний или нарушений у человека создает хорошо известные проблемы возможной иммуногенности, особенно при повторном введении антитела, которые заключаются в том, что иммунная система пациента может узнать чужеродное интактное антитело, отличное от человеческого, как несобственное и вызвать нейтрализующую реакцию. Помимо создания полностью человеческих антител (см. выше), в течение последних лет были разработаны разные методы, позволяющие преодолеть вышеуказанные проблемы, которые обычно предполагают уменьшение содержания аминокислотных последовательностей, отличных от человеческих, в интактном терапевтическом антителе при сохранении относительной простоты получения антител, отличных от человеческих, у иммунизированных животных, например мыши, крысы или кролика. Для достижения указанной цели широко используются два метода. Первый метод включает создание химерных антител, которые обычно содержат вариабельную область, отличную от человеческой (например, грызуна, такого как мышь), слитую с константной областью человека. Так как антигенсвязывающий центр антитела локализован в вариабельных областях, химерное антитело сохраняет сродство связывания с антигеном, но приобретает эффекторные функции константной области человека и поэтому может выполнять вышеописанные эффекторные функции. Химерные антитела обычно получают методами рекомбинантных ДНК. ДНК, кодирующую антитела (например, кДНК), выделяют и секвенируют стандартными методами (например, используя олигонуклеотидные зонды, способные специфически связываться с генами, кодирующими вариабельные области Н- и L-цепей антитела по настоящему изобретению. Клетки гибридомы являются типичным источником такой ДНК. Выделенную ДНК вводят в экспрессирующие векторы, которые трансфицируют в клетки-хозяева, такие как E.coli, клетки COS, клетки СНО или миеломные клетки, которые в противном случае не продуцируют белок иммуноглобулина, вызывая синтез указанного антитела. ДНК может быть модифицирована путем замены L- и Н-цепей человека кодирующей последовательностью для соответствующих константных областей Н- и L-цепей, отличных от человеческих (например, мыши); см., например, публикацию Morrison; PNAS 81, 6851 (1984). Таким образом, другой вариант осуществления изобретения относится к химерному антителу, включающему VH-область, имеющую последовательность SEQ ID NO:2, 6 или 10, и VL-область, имеющую последовательность SEQ ID NO:4, 8 или 12, слитые с константной областью человека (которая может быть изотипом IgG, например IgG1). Второй метод включает создание гуманизированных антител, в которых содержание части, отличной от человеческой, уменьшено путем гуманизации вариабельных областей. Широко применяются два метода гуманизации. Первый метод включает гуманизацию путем трансплантации CDR. CDR формируют петли рядом с N-концом антитела, где они образуют поверхность в каркасных областях. Антигенсвязывающая специфичность антитела определяется, главным образом, топографией и химическими характеристиками поверхности CDR. Указанные свойства, в свою очередь, определяются конформацией отдельных CDR, относительным расположением CDR, характером и расположением боковых цепей остатков, образующих CDR. Значительное уменьшение иммуногенности может быть достигнуто в результате трансплантации только CDR антител, отличных от человеческих (например, мыши)("донорские" антитела) в подходящую каркасную область человека ("каркасная область акцепторного антитела") и константные области (см. публикации Jones et al. (1986), Nature, 321, 522-525 иVerhoeyen M. et al. (1988), Science, 239, 1534-1536). Однако сама по себе трансплантация CDR не может гарантировать полного сохранения антигенсвязывающих свойств, поэтому часто оказывается, что необходимо сохранить некоторые остатки каркасной области донорского антитела (иногда определяемые как"обратные мутации") в гуманизированной молекуле для достижения значительного антигенсвязывающего сродства (см. публикации Queen С. et al. (1989), PNAS 86, 10029-10033, Со, М. et al. (1991), Nature, 351,501-502). В данном случае V-области человека, характеризующиеся наибольшей гомологией последовательностей (обычно 60% или больше) с донорским антителом, отличным от человеческого, могут быть выбраны из базы данных для создания каркасной области человека (FR). FR человека могут быть выбраны из консенсусных или индивидуальных антител человека. При необходимости основные остатки из донорского антитела вводят в каркасную область акцепторного антитела человека для сохранения конформаций CDR. Для идентификации таких структурно важных остатков может быть использовано компьютерное моделирование антитела; см. WO 99/48523. Альтернативно, гуманизация может быть выполнена методом "маскировки". Статистический анализ уникальных вариабельных областей тяжелых и легких цепей иммуноглобулина человека и мыши показал, что точные паттерны остатков различны в антителах человека и мыши и в большинстве случаев предпочитают положения с небольшим числом разных остатков (см. публикации Padlan E.A. et al. (1991),-8 024621Mol. Immunol. 28, 489-498 и Pedersen J.T. et al. (1994), J. Mol. Biol. 235; 959-973). Поэтому иммуногенность Fv-фрагмента, отличного от человеческого, можно уменьшить, заменяя в каркасных областях остатки, отличающиеся от остатков, обычно обнаруживаемых в антителах человека. Так как антигенность белка можно коррелировать с доступностью поверхности, замена поверхностных остатков может быть достаточной для того, чтобы сделать вариабельную область мыши "невидимой" для иммунной системы человека (см. также публикацию Mark G.E. et al. (1994), in Handbook of Experimental Pharmacology vol. 113: The pharmacology of monoclonal Anthibodies, Springer-Verlag, p. 105-134). Данный процесс гуманизации определяется как "маскировка", поскольку изменяется только поверхность антитела, при этом несущие остатки остаются без изменения. Другой альтернативный метод описан в WO 04/006955. Другие альтернативные подходы включают методы, рассмотренные в WO04/006955, и метод Humaneering(Kalobios), который благодаря использованию бактериальных экспрессирующих систем позволяет получить антитела, в которых последовательность близка к зародышевой линии человека (Alfenito-MAdvancing Protein Therapeutics, January 2007, San Diego, California). Другой метод гуманизации включает отбор каркасных областей акцепторного антитела человека на основании структурного сходства областей CDR человека с областями CDR донорского антитела мыши, а не на гомологии других областей антитела, таких как каркасные области. Указанный метод также известен как супергуманизация (Evogenix Inc.; Hwang et al. (2005), Methods 36:35-42). Специалистам в данной области должно быть понятно, что термин "выделенный" означает не только источник физического происхождения материала, но также служит для определения материала, который идентичен в структурном отношении данному материалу, но получен из другого источника. Таким образом, "остатки, обнаруженные в донорском антителе", необязательно должны быть очищены от донорского антитела. Фрагменты антитела. Некоторые варианты осуществления изобретения относятся к терапевтическому антителу, которое является антигенсвязывающим фрагментом. Такие фрагменты могут быть функциональными антигенсвязывающими фрагментами интактных, гуманизированных и/или химерных антител, такими как Fab,Fd, Fab', F(ab')2, Fv, ScFv фрагменты вышеописанных антител. Фрагменты, у которых отсутствует константная область, утрачивают способность активировать комплемент по классическому пути или опосредовать антителозависимую клеточно-опосредованную цитотоксичность. Такие фрагменты традиционно получают в результате протеолитического расщепления интактных антител, например, папаином(см., например, WO 94/29348), но фрагменты могут быть получены непосредственно из рекомбинантно трансформированных клеток-хозяев. Для получения ScFv см. публикацию Bird et al. (1988), Science, 242,423-426. Кроме того, фрагменты антитела могут быть получены разными методами генной инженерии,описанными ниже.Fv-фрагменты, по-видимому, характеризуются более низкой энергией взаимодействия двух цепей,чем Fab-фрагменты. Чтобы стабилизировать ассоциацию VH- и VL-областей, указанные области связывают пептидами (Bird et al., (1988), Science 242, 423-426, Huston et al., PNAS, 85, 5879-5883), дисульфидными мостиками (Glockshuber et al. (1990), Biochemistry, 29, 1362-1367) и мутациями "выступ в отверстии" (Zhu et al. (1997), Protein Sci., 6, 781-788). ScFv-фрагменты могут быть получены методами, хорошо известными специалистам в данной области; см. публикации Whitlow et al. (1991), Methods companionMethods Enzymol., 2, 97-105 и Huston et al. (1993), Int.Rev.Immunol. 10, 195-217. ScFv-фрагменты могут быть получены в бактериальных клетках, таких как E.Coli, но обычно указанные фрагменты получают в эукариотических клетках. Одним недостатком ScFv-фрагментов является одновалентность продукта, что делает невозможной повышенную авидность из-за многовалентного связывания и определяет короткий период полувыведения. Попытки преодолеть указанные проблемы привели к использованию двухвалентного (ScFv')2-фрагмента, полученного из ScFv-фрагмента, содержащего дополнительный С-концевой цистеин, путем химического связывания (Adams et al. (1993), Can. Res. 53, 4026-4034 и McCartney et al.Biophys, 26, 187-204). Альтернативно, из ScFv-фрагмента могут быть получены многомеры в результате укорачивания пептидного линкера между остатками 3-12 с образованием "диател"; см. публикациюHolliger et al., PNAS (1993), 90, 6444-6448. Уменьшение линкера может далее привести к получению тримеров ScFv ("триатела"; см. публикацию Kortt et al. (1997), Protein Eng., 10, 423-433) и тетрамеровScFv могут быть также созданы путем генетического слияния с димеризующими белок фрагментами с образованием "миниантител" (см. публикацию Pack et al. (1992), Biochemistry 31, 1579-1584) и "минител"(см. публикацию Hu et al. (1996), Cancer Res. 56, 3055-3061). Могут быть также получены тандемы ScFvSc-Fv ScFv)2) путем связывания двух звеньев ScFv третьим пептидным линкером; см. публикациюKurucz et al. (1995), J. Immol. 154, 4576-4582. Биспецифические диатела могут быть получены путем нековалентного связывания двух одноцепочечных продуктов слияния, состоящих из VH-области одного антитела, соединенного коротким линкером с VL-областью другого антитела; см. публикацию Kipriyanovet al. (1998), Int. J. Can. 77, 763-772. Устойчивость таких биспецифических диател может быть увеличена путем введения дисульфидных мостиков или мутаций "выступ в отверстии", описанных выше, или образования одноцепочечных диател (ScDb), в которых два гибридных ScFv-фрагмента соединены пептидным линкером; см. публикацию Kontermann et al. (1999), J. Immunol. Methods 226, 179-188. Четырехвалентные биспецифические молекулы могут быть получены, например, путем слияния ScFv-фрагмента с СН 3-областью молекулы IgG или с Fab-фрагментом при помощи шарнирной области; см. публикациюColoma et al. (1997), Nature Biotechnol. 15, 159-163. Альтернативно, четырехвалентные биспецифические молекулы были созданы путем слияния биспецифических одноцепочечных диател (см. публикацию Altet al., (1999), FEBS Lett, 454, 90-94). Более мелкие четырехвалентные биспецифические молекулы могут быть также образованы путем димеризации тандемов ScFv-ScFv при помощи линкера, содержащего фрагмент спираль-петля-спираль (DiBi-миниантитела; см. публикацию Muller et al. (1998), FEBS Lett 432,45-49), или одноцепочечной молекулы, содержащей четыре вариабельные области антитела (VH и VL) в ориентации, препятствующей внутримолекулярному спариванию (тандемное антитело; см. публикациюKipriyanov et al., (1999), J. Mol. Biol. 293, 41-56). Биспецифические F (ab')2-фрагменты могут быть созданы путем химического связывания Fab'-фрагментов или гетеродимеризации при помощи лейциновых зипперов (см. публикации Shalaby et al. (1992), J. Exp. Med. 175, 217-225 и Kostelny et al. (1992), J.Immunol. 148, 1547-1553). Кроме того, могут быть получены так называемые антитела-домены, созданные на основе выделенных VH- или VL-областей (Domantis Ltd.); см. патенты США 6248516, 6291158,6172197. Гетероконъюгатные антитела. Гетероконъюгатные антитела являются производными антителами, которые также относятся к одному варианту осуществления настоящего изобретения. Гетероконъюгатные антитела состоят из двух ковалентно связанных антител, образованных при помощи любых известных методов перекрестного связывания. См. патент США 4676980. Другие модификации. Антитела по настоящему изобретению могут также включать любые другие модификации в константных областях. Например, известно, что гликозилирование антител в консервативных положениях константных областей оказывает сильное воздействие на функцию антител, в частности, на вышеописанные эффекторные функции; см., например, публикацию Boyd et al. (1996), Mol. Immunol. 32, 13111318. Варианты гликозилирования терапевтических антител по настоящему изобретению включают добавление, замену, делецию или модификацию одной или нескольких углеводных частей. Введение фрагмента аспарагин-Х-серин или аспарагин-Х-треонин создает потенциальный сайт для ферментативного присоединения углеводных частей и поэтому может быть использовано для управления гликозилированием антитела. В публикации Raju et al. (2001), Biochemistry, 40, 8868-8876 указано, что сиалилирование конца иммуноадгезина TNFR-IgG было увеличено в результате регалактозилирования и/или ресиалилирования с использованием бета-1,4-галактозилтрансферазы и/или альфа-2,3-сиалилтрансферазы. Считается, что повышенное сиалилирование конца увеличивает период полувыведения иммуноглобулина. Антитела, наряду с большинством гликопротеинов, обычно образуются в природе в виде смеси гликоформ. Указанная смесь является особенно очевидной при получении антител в эукариотических клетках, в частности в клетках млекопитающих. Были разработаны разные методы получения определенных гликоформ; см. публикации Zhang et al., Science (2004), 303, 371, Sears et al., Science (2001), 291,2344, Wacker et al. (2002), Science, 298, 1790, Davis et al. (2002), Chem. Rev. 102, 579, Hang et al. (2001),Acc. Chem. Res. 34, 727. Таким образом, настоящее изобретение относится к множеству терапевтических антител (которые могут быть изотипом IgG, например, IgG1), включающему определенное число (например, 7 или меньше, в частности 5 или меньше, а именно две или одну) гликоформ указанных антител. Производные по настоящему изобретению включают также терапевтические антитела по настоящему изобретению, связанные с небелковым полимером, таким как полиэтиленгликоль (PEG), полипропиленгликоль или полиоксиалкилен. Конъюгация белков с PEG является известным методом увеличения периода полувыведения белков, а также уменьшения антигенности и иммуногенности белков. Метод обработки полиэтиленгликолем с разными молекулярными массами и разных типов (линейные или разветвленные) был исследован с использованием интактных антител, а также Fab'-фрагментов; см. публикацию Koumenis I.L. et al. (2000), Int. J. Pharmaceut. 198:83-95. Конкретный вариант осуществления изобретения относится к антигенсвязывающему фрагменту по настоящему изобретению без эффекторных функций а) активации комплемента по классическому пути и b) опосредования антителозависимой клеточно-опосредованной цитотоксичности (такому как Fab-фрагмент или scFv), связанному с PEG. Считается, что взаимодействие Fc-области антитела с разными Fc-рецепторами (FcR) опосредует эффекторные функции антитела, которые включают антителозависимую клеточно-опосредованную цитотоксичность (ADCC), фиксацию комплемента, фагоцитоз и период полувыведения/клиренс антитела.Fc-область антител по настоящему изобретению может быть подвергнута разным модификациям в зависимости от требуемой эффекторной функции. В частности, константные области человека, у которых отсутствуют функции а) активации комплемента по классическому пути и b) опосредования антителозависимой клеточно-опосредованной цитотоксичности, включают константную область IgG4, константную область IgG2 и константную область IgG1, содержащие специфические мутации, например мутации в положениях 234, 235, 236, 237, 297, 318, 320 и/или 322, описанные в EP 0307434 (WO 88/07089),ЕР 0629240 (WO 93/17105) и WO 2004/014953. Кроме того, было отмечено, что мутации в положении остатков 235 или 237 СН 2-домена константной области тяжелой цепи (нумерация Кабата; европейская система индексации) уменьшают связывание с FcRI, FcRII и FcRIII и, таким образом, уменьшают антителозависимую клеточно-опосредованную цитотоксичность (ADCC) (Duncan et al. Nature 1988, 332; 563-564; Lund et al. J. Immunol. 1991, 147; 2657-2662; Chappel et al. PNAS, 1991, 88; 9036-9040; Burton andWoof, Adv. Immunol. 1992, 51; 1-84; Morgan et al., Immunology 1995, 86; 319-324; Hezareh et al., J. Virol. 2001, 75 (24); 12161-12168). В некоторых научных отчетах было описано участие некоторых из указанных остатков в рекрутинге или опосредовании комплементзависимой цитотоксичности (CDC) (Morgan etal., 1995; Xu et al., Cell. Immunol. 2000; 200:16-26; Hezareh et al., J. Virol. 2001, 75(24); 12161-12168). Поэтому остатки 235 и 237 были мутированы с введением остатков аланина (Brett et al., Immunology 1997,91; 346-353; Bartholomew et al., Immunology 1995, 85; 41-48 и WO 99/58679 для уменьшения комплементопосредованных и FcR-опосредованных воздействий. Антитела, включающие указанные константные области, могут именоваться "нелитическими" антителами. В антитело можно ввести эпитоп связывания с рецептором реутилизации для увеличения периода полувыведения из кровяного русла; см. патент США 5739277. В настоящее время известны пять узнаваемых рецепторов Fc, а именно, FcR(I), FcRIIa, FcRIIb,FcRIIIa и неонатальный рецептор FcRn. В публикации Shields et al. (2001), J. Biol. Chem. 276, 6591-6604 было продемонстрировано, что общий набор остатков IgG1 участвует в связывании всех рецепторовFcR, в то время как FcRII и FcRIII используют другие сайты, не входящие в указанный общий набор. Одна группа остатков IgG1 уменьшала связывание со всеми рецепторами FcR при замене на аланин:Pro-238, Asp-265, Asp-270, Asn-297 и Pro-239. Все указанные остатки находятся в СН 2-домене IgG и образуют кластер рядом с шарнирным соединением СН 1 и СН 2. В то время как FcRI использует только общий набор остатков IgG1 для связывания, FcRII и FcRIII взаимодействуют также с другими остатками помимо общего набора. Замена некоторых остатков уменьшала связывание только с FcRII (например, Arg-292) или FcRIII (например, Glu-293). Некоторые варианты характеризовались лучшим связыванием с FcRII или FcyRIII, но не влияли на связывание с другим рецептором (например, замена Ser-2 67 аланином (Ala) улучшала связывание с FcRII, не влияя на связывание с FcRIII). Другие варианты характеризовались лучшим связыванием с FcRII или FcRIII при уменьшении связывания с другим рецептором (например, замена Ser-298 аланином (Ala) улучшала связывания с FcRIII и уменьшала связывание с FcRII). Варианты IgG1, обеспечивающие лучшее связывание с FcRIIIa, характеризовались общим введением аланина в положениях Ser-298, Glu-333 и Lys-334. Считается, что неонатальный рецептор FcRn защищает молекулы IgG от разрушения и, таким образом, увеличивает период полувыведения из кровяного русла и трансцитоз в тканях (см. публикации Junghans R.P. (1997), Immunol. Res. 16, 29-57 иGhetie et al. (2000), Annu. Rev. Immunol. 18, 739-766). Установлено, что остатки IgG1 человека, непосредственно взаимодействующие с FcRn человека, включают Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435. Терапевтическое антитело по настоящему изобретению может включать любые вышеуказанные модификации константных областей. 2. Способы получения. Антитела по настоящему изобретению могут быть продуцированы в трансгенных организмах, таких как козы (см. публикацию Pollock et al. (1999), J. Immunol. Methods 231:147-157), цыплята (см. публикацию Morrow KJJ (2000), Genet. Eng. News 20:1-55), мыши (см. вышеуказанную публикацию Pollock) или растения (см. публикации Doran P.M. (2000), Curr. Opinion Biotechnol. 11, 199-204, Ma JK-C (1998),Nat. Med. 4; 601-606, Baez J. et al., BioPharm (2000), 13:50-54, Stoger E. et al. (2000), Plant Mol. Biol. 42:583-590). Антитела могут быть также получены методами химического синтеза. Однако антитела по настоящему изобретению обычно получают методом культивирования рекомбинантных клеток, хорошо знакомым специалистам в данной области. Полинуклеотид, кодирующий антитело, выделяют и вводят в реплицирующийся вектор, такой как плазмида, для последующего клонирования (амплификации) или экспрессии в клетке-хозяине. Одной приемлемой экспрессирующей системой является система на основе глутаматсинтетазы (компании Lonza Biologics), особенно при использовании в качестве клетки-хозяина СНО или NS0 (см. ниже). Полинуклеотид, кодирующий антитело, легко выделяют и секвенируют стандартными методами (например, при помощи олигонуклеотидных зондов). Применяемые векторы включают плазмиду, вирус, фаг, транспозоны, минихромосомы, из которых наиболее типичными являются плазмиды. Такие векторы обычно дополнительно включают сигнальную последовательность, ориджин репликации, один или несколько генов-маркеров, энхансер, промотор и последовательности терминации транскрипции, функционально связанные с полинуклеотидом легкой и/или тяжелой цепи для облегчения экспрессии. Полинуклеотид, кодирующий легкую и тяжелую цепи, может быть вставлен в отдельные векторы и одновременно или последовательно введен (например, путем трансформации, трансфекции,электропорации или трансдукции) в ту же клетку-хозяина, либо, при желании, тяжелая и легкая цепи могут быть вставлены в один и тот же вектор до введения в клетку-хозяина. Специалистам в данной области должно быть очевидно, что вследствие избыточности генетического кода, помимо вышеуказанных полинуклеотидов, также существуют другие полинуклеотиды, которые кодируют полипептиды по настоящему изобретению. Сигнальные последовательности. Антитела по настоящему изобретению могут быть получены в виде слитого белка с гетерологичной сигнальной последовательностью, имеющей специфический сайт расщепления у N-конца зрелого белка. Сигнальная последовательность должна быть узнана и процессирована клеткой-хозяином. Для прокариотических клеток-хозяев сигнальной последовательностью может быть щелочная фосфатаза, пенициллиназа или лидерные последовательности теплостойкого энтеротоксина II. Для секреции в дрожжах сигнальными последовательностями могут быть лидерная последовательность инвертазы дрожжей, лидерная последовательность -фактора или лидерные последовательности кислой фосфатазы; см., например,WO 90/13646. В системах клеток млекопитающих могут быть использованы секретирующие лидерные последовательности вирусов, такие как сигнальная последовательность вируса простого герпеса gD и сигнальная последовательность природного иммуноглобулина (например, тяжелая цепь Ig человека). Сигнальная последовательность обычно лигирована в рамке считывания с полинуклеотидом, кодирующим антитело по настоящему изобретению. Ориджин репликации. Ориджины репликации хорошо известны в данной области, при этом pBR322 пригоден для большинства грамотрицательных бактерий, 2 плазмида пригодна для большинства дрожжей, и разные вирусы, такие как SV40, вирус полиомы, аденовирус, VSV или BPV, пригодны для большинства клеток млекопитающих. Ориджин репликации обычно не нужен для интегрированных экспрессирующих векторов млекопитающих, за исключением введения вектора в E.Coli. Однако ориджин репликации SV40 может быть использован, так как данный вирус содержит ранний промотор. Селектирующий маркер. Типичные селектирующие гены кодируют белки, которые (а) сообщают устойчивость к антибиотикам или другим токсинам, таким как ампициллин, неомицин, метотрексат или тетрациклин, или (b) восполняют недостаток ауксотрофов или предоставляют питательные вещества, отсутствующие в комплексной среде, или (с) выполняют обе функции. Схема селекции может включать замедление роста клеток-хозяев, которые не содержат вектора или векторов. Клетки, которые были успешно трансформированы генами, кодирующими терапевтическое антитело по настоящему изобретению, выживают, например, благодаря лекарственной устойчивости, сообщенной доставленным селектирующим маркером. Одним примером является DHFR-селектирующая система, в которой трансформанты образуются в DHFRотрицательных линиях хозяев (см., например, публикацию Page and Sydenham, 1991, Biotechnology, 9:6468). В указанной системе ген DHFR доставляют вместе с полинуклеотидными последовательностями антитела по настоящему изобретению и затем отбирают DHFR-положительные клетки путем удаления нуклеозида. При необходимости, для отбора трансформантов с амплификацией гена DHFR также используют ингибитор DHFR метотрексат. В результате функционального связывания гена DHFR с кодирующими последовательностями антитела по настоящему изобретению или функциональными производными указанных последовательностей амплификация гена DHFR вызывает сопутствующую амплификацию представляющих интерес последовательностей требуемого антитела. Клетки СНО представляют собой особенно пригодную линию клеток для отбора с помощью DHFR/метотрексата, и в данной области хорошо известны методы амплификации и отбора клеток-хозяев с использованием системы DHFR; см. публикацию Kaufman R.J. et al., J. Mol. Biol. (1982), 159, 601-621; для обзора см. публикациюbiopharmaceuticals", Arzneimittel-Forschung, 48(8): 870-80, 1988 Aug. Другим примером является экспрессирующая система на основе глутаматсинтетазы (Lonza Biologies). Подходящим селектирующим геном,предназначенным для использования в дрожжах, является ген trp1; см. публикацию Stinchcomb et al.,Nature, 282, 38, 1979. Промоторы. Подходящие промоторы для экспрессии антител по настоящему изобретению функционально связаны с ДНК/полинуклеотидом, кодирующим данное антитело. Промоторы для прокариотических хозяев включают промотор phoA, промоторные системы на основе бета-лактамазы и лактозы, щелочную фосфатазу, триптофан и гибридные промоторы, такие как Тас. Промоторы, пригодные для экспрессии в дрожжевых клетках, включают 3-фосфоглицераткиназу или другие гликолитические ферменты, например, энолазу, глицеральдегид-3-фосфатдегидрогеназу, гексокиназу, пируватдекаробоксилазу, фосфофруктокиназу, глюкоза-6-фосфатизомеразу, 3-фосфоглицератмутазу, глюкокиназу и др. Индуцибельные дрожжевые промоторы включают алкогольдегидрогеназу-2, изоцитохром С, кислую фосфатазу, металлотионеин, ферменты, отвечающие за азотистый обмен или утилизацию мальтозы/галактозы, и др. Промоторы, предназначенные для экспрессии в клеточных системах млекопитающих, включают РНК-полимеразу II, в том числе вирусные промоторы, такие как вирус полиомы, птичий поксвирус и аденовирусы (например, аденовирус 2), вирус папилломы крупного рогатого скота, птичий вирус саркомы, цитомегаловирус (в частности, промотор предраннего гена), ретровирус, вирус гепатита В, актин,вирус саркомы Рауса (RSV) и ранний или поздний вакуолизирующий обезьяний вирус 40, и невирусные промоторы, такие как EF-1-альфа (Mizushima and Nagata, Nucleic Acids Res., 1990, 18(17): 5322), и др. Выбор промотора может быть основан на подходящей совместимости с клеткой-хозяином, используемой для экспрессии. Энхансер. В случае необходимости, например, для экспрессии в высших эукариотических клетках могут быть использованы дополнительные энхансерные элементы вместо промоторов или энхансерные элементы,обнаруженные в описанных выше промоторах. Подходящие энхансерные последовательности млекопитающих включают энхансерные элементы из глобина, эластазы, альбумина, фетопротеина, металлотионина и инсулина. Альтернативно можно использовать энхансерный элемент из вируса эукариотической клетки, такой как энхансер SV40, энхансер раннего промотора цитомегаловируса, энхансер вируса полиомы, энхансер бакуловируса или локус IgG2a мыши (см. WO 04/009823). Несмотря на то что такие энхансеры обычно находятся в векторе в сайте вверху от промотора, они могут быть расположены в любом месте, например, в нетранслированной области или внизу от сигнала полиаденилирования. Энхансер и его местоположение могут быть выбраны с учетом подходящей совместимости с клеткой-хозяином,используемой для экспрессии. Полиаденилирование/терминация. В эукариотических системах сигналы полиаденилирования функционально связаны с полинуклеотидом, кодирующим антитело по настоящему изобретению. Такие сигналы обычно находятся у 3'-конца открытой рамки считывания. В системах млекопитающих неограничивающие примеры сигналов включают последовательности, выделенные из гормонов роста, альфа-фактора элонгации 1 и вирусных (например, SV40) генов или длинных концевых повторов ретровирусов. В дрожжевых системах неограничивающие примеры сигналов полиаденилирования/терминации включают последовательности, выделенные из генов фосфоглицераткиназы (PGK) и алкогольдегидрогеназы 1 (ADH). В прокариотической системе сигналы полиаденилирования обычно не требуются, и вместо них обычно используют более короткие и более определенные терминаторные последовательности. Выбор последовательностей полиаденилирования/терминации может быть основан на подходящей совместимости с клеткой-хозяином, используемой для экспрессии. Другие методы/элементы для повышения выхода. Помимо вышеизложенного, другие признаки, которые позволяют увеличить выход, включают элементы ремоделирования хроматина, интроны и модификацию конкретного кодона клетки-хозяина. Кодон, используемый в антителе по настоящему изобретению, может быть модифицирован для соответствия смещению кодона клетки-хозяина, например, для увеличения выхода транскрипта и/или продукта(см., например, публикацию Hoekema A. et al., Mol. Cell Biol., 1987, 7(8):2914-24). Выбор кодонов может быть основан на подходящей совместимости с клеткой-хозяином, используемой для экспрессии. Клетки-хозяева. Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитела по настоящему изобретению, являются, например, прокариотическими, дрожжевыми или высшими эукариотическими клетками. Подходящие прокариотические клетки включают эубактерии, например, энтеробактерии, такие как Escherichia, например, E.Coli (например, АТСС 31446, 31537, 27325), Enterobacter,Erwinia, Klebsiella Proteus, Salmonella, например Salmonella typhimurium, Serratia, например, Serratiamarcescans, и Shigella, а также Bacilli, такие как В.subtilis и В.licheniformis (см. DD 266710), Pseudomonas,такие как P.aeruginosa и Streptomyces. Из дрожжевых клеток-хозяев могут быть также использованыAspergillus, такие как A.nidulans и A.niger. Несмотря на то что в объем настоящего изобретения входят прокариотические и дрожжевые клетки-хозяева, обычно клетками-хозяевами по настоящему изобретению являются клетки позвоночных. Подходящие клетки-хозяева позвоночных включают клетки млекопитающих, такие как COS-1 (АТССCRL 1650), COS-7 (АТСС CRL 1651), линия клеток почек эмбриона человека 293, PerC6 (Crucell),клетки почек детеныша хомячка (BHK) (АТСС CRL 1632), BHK570 (АТССCRL 10314), 293 (АТССCRL 1573), клетки яичника китайского хомячка СНО (например, CHO-K1, АТССCCL 61), линия клеток DHFR-CHO, такая как DG44 (см. приведенную выше публикацию Urlaub et al. (1986, в частности линии клеток СНО, адаптированные для суспензионной культуры, клетки Сертоли мыши, клетки почки обезьяны, клетки почки африканской зеленой мартышки (АТСС CRL 1587), клетки HELA, клетки почки собаки (АТСС CCL 34), клетки легких человека (АТСС CCL 75), клетки Hep G2 и миеломные или лимфомные клетки, например, NS0 (см. патент США 5807715), Sp2/0, Y0. Таким образом, один вариант осуществления изобретения относится к устойчиво трансформированной клетке-хозяину, содержащей вектор, кодирующий тяжелую цепь и/или легкую цепь терапевтического антитела по настоящему изобретению. Такие клетки-хозяева обычно содержат первый вектор, кодирующий легкую цепь, и второй вектор, кодирующий тяжелую цепь. Такие клетки-хозяева могут быть также созданы или адаптированы для изменения качества, функции и/или выхода антитела по настоящему изобретению. Неограничивающие примеры включают экспрессию специфических модифицирующих (например, гликозилирующих) ферментов и чаперонов, производящих укладку белка. Методы культивирования клеток. Клетки-хозяева, трансформированные векторами, кодирующими терапевтические антитела по настоящему изобретению, можно культивировать любым методом, известным специалистам в данной области. Клетки-хозяева можно культивировать во вращающихся колбах, встряхиваемых колбах, роллерфлаконах или системах полых волокон, но для широкомасштабного производства предпочтительны реакторы с мешалками или реакторы с корзинами (см., например, Wave Biotech. Somerset, New Jersey,USA), особенно для суспензионных культур. Баки с мешалкой обычно приспосабливают для аэрации при помощи барботеров, дефлекторов или импеллеров со слабым сдвигающим усилием. Для барботажных колонн и реакторов с перемешиванием струей воздуха может быть использована прямая аэрация пузырьками воздуха или кислорода. При культивировании клеток-хозяев в бессывороточной культуральной среде желательно ввести в среду защищающий клетки агент, такой как плюроник F-68, для предотвращения разрушения клеток в результате процесса аэрации. В зависимости от характеристик клеткихозяина в качестве субстратов роста для якорной подложки могут быть использованы микроносители,или клетки могут быть адаптированы для суспензионной культуры (которая является типичной). При культивировании клеток-хозяев, в частности, клеток-хозяев позвоночных, могут быть использованы разные методы, такие как периодическое культивирование, культивирование с подпиткой, повторяющееся периодическое культивирование (см. публикацию Drapeau et al. (1994), Cytotechnology, 15:103-109), продолжительное периодическое культивирование или перфузионное культивирование. Хотя рекомбинантно трансформированные клетки-хозяева млекопитающих можно культивировать в сывороточной среде,которая включает фетальную телячью сыворотку (FCS), клетки-хозяева предпочтительно культивируют в синтетической бессывороточной среде, аналогичной описанной в публикации Keen et al. (1995),Cytotechnology, 17:153-163, или в коммерчески доступной среде, такой как ProCHO-CDM илиUltraCHO (Cambrex NJ, USA), содержащей при необходимости источник энергии, такой как глюкоза, и синтетические факторы роста, такие как рекомбинантный инсулин. Культивирование клеток-хозяев в бессывороточной среде может потребовать адаптации указанных клеток для роста в бессывороточных условиях. Одним методом адаптации является культивирование таких клеток-хозяев в сывороточной среде и последовательная замена 80% культуральной среды бессывороточной средой, в результате чего клетки-хозяева будут адаптироваться к бессывороточным условиям (см., например, Scharfenberg K. et al.(1995), in Animal Cell technology: Developments towards the 21st century (Beuvery E.C. et al., eds), p. 619623, Kluwer Academic publishers). Антитела по настоящему изобретению, секретируемые в среду, могут быть выделены и очищены от среды разными методами с достижением степени чистоты, подходящей для предполагаемого применения. Например, применение терапевтических антител по настоящему изобретению для лечения людей требует по меньшей мере 95% чистоты, определяемой методом SDS-PAGE, обычно 98 или 99% чистоты по сравнению с культуральной средой, содержащей терапевтические антитела. В первом случае клеточный дебрис из культуральной среды обычно удаляют центрифугированием с последующим выполнением стадии очистки супернатанта при помощи микрофильтрации, ультрафильтрации и/или глубинной фильтрации. Альтернативно антитело может быть получено микрофильтрацией, ультрафильтрацией или глубинной фильтрацией без предшествующего центрифугирования. Существует целый ряд других методов,таких как диализ и гель-электрофорез, и хроматографических методов, таких как хроматография на колонке с гидроксиапатитом (НА), аффинная хроматография (необязательно включающая аффинную систему мечения, такую как полигистидин) и/или хроматография гидрофобного взаимодействия (HIC, см. патент США 5429746). В одном варианте осуществления антитела по настоящему изобретению получают после выполнения разных стадий очистки при помощи аффинной хроматографии белков А или G с последующим выполнением ионообменной хроматографии и/или хроматографии на колонке с гидроксиапатитом, анионообменной или катионообменной хроматографии, гель-хроматографии и осаждения сульфатом аммония. Кроме того, могут быть использованы разные стадии удаления вируса (например,нанофильтрация с использованием, например, фильтра DV-20). В результате выполнения вышеуказанных стадий получают очищенный (обычно моноклональный) препарат, содержащий по меньшей мере 10 мг/мл или больше, например, 100 мг/мл или больше антитела по настоящему изобретению, который представляет собой еще один вариант осуществления изобретения. Концентрация до 100 мг/мл или больше может быть достигнута ультрацентрифугированием. Такие препараты, по существу, не содержат агрегированные формы антител по настоящему изобретению. Бактериальные системы особенно пригодны для экспрессии фрагментов антитела. Такие фрагменты локализованы внутри клетки или в периплазме. Нерастворимые периплазматические белки могут быть экстрагированы и подвергнуты повторной укладке с образованием активных белков согласно методам,известным специалистам в данной области; см. публикации Sanchez et al. (1999), J. Biotechnol. 72, 13-20 иCupit P.M. et al. (1999), Lett. Appl. Microbiol., 29, 273-277. 3. Фармацевтические композиции и способ введения. Очищенные препараты антител по настоящему изобретению, описанные выше, могут быть введены в фармацевтические композиции для применения при лечении вышеуказанных заболеваний и нарушений у человека. Такие композиции обычно дополнительно включают фармацевтически приемлемый (т.е. инертный) носитель, используемый в общепринятой фармацевтической практике; см., например,Remingtons Pharmaceutical Sciences, 16th ed. (1980), Mack Publishing Co. Примеры таких носителей включают стерилизованные носители, такие как физиологический раствор, раствор Рингера или раствор декстрозы, содержащие подходящие буферы для достижения значения рН, равного 5-8. Фармацевтические композиции для инъекций (например, внутривенных, внутрибрюшинных, внутрикожных, подкожных,внутримышечных или внутрипортальных) или вливаний не содержат видимых частиц и могут включать от 0,1 мг до 10 г терапевтического антитела, обычно от 5 до 25 мг антитела. Методы получения таких фармацевтических композиций хорошо известны специалистам в данной области. В одном варианте осуществления изобретения фармацевтические композиции содержат от 0,1 мг до 10 г терапевтических антител по настоящему изобретению в стандартной лекарственной форме с прилагаемой инструкцией по применению. Фармацевтические композиции по настоящему изобретению могут быть лиофилизованы(высушены вымораживанием) для восстановления перед введением способами, хорошо известными специалистам в данной области. Когда варианты осуществления изобретения относятся к антителам по настоящему изобретению с изотипом IgG1, в фармацевтическую композицию может быть добавлен хелатор меди, такой как цитрат (например, цитрат натрия), EDTA или гистидин, для уменьшения степени опосредуемого медью разрушения антител данного изотипа; см. ЕР 0612251. Эффективные дозы и схемы введения антитела по настоящему изобретению обычно определяются эмпирически и зависят от таких факторов, как возраст, масса тела и состояние здоровья пациента, подлежащее лечению заболевание или нарушение. Такие факторы должны учитываться лечащим врачом. Руководство по выбору соответствующих доз приведено, например, в публикации Smith et al. (1977),Antibodies in human diagnosis and therapy, Raven Press, New York, но приемлемая доза обычно находится в пределах от 0,1 мг до 1 г. В одном варианте осуществления изобретения схема введения при лечении человека составляет от 0,1 мг до 10 г терапевтического антитела по настоящему изобретению, вводимого подкожно один раз в неделю или через две недели, либо в виде внутривенного вливания через 1 или 2 месяца. Композиции по настоящему изобретению могут быть также использованы в профилактических целях. 4. Клинические применения. Настоящее изобретение относится к антителу, которое связывается с IL-8, Gro-альфа, Gro-бета,Gro-гамма и ENA-78 человека. Настоящее изобретение относится также к способам лечения указанным антителом заболеваний или нарушений, характеризующихся повышенным или несбалансированным уровнем одного или нескольких хемокинов, включающих IL-8, Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека, а именно хронического обструктивного заболевания легких (COPD), остеоартрита, ревматоидного артрита, эрозивного артрита, астмы, атеросклероза, воспалительного заболевания кишечника, псориаза, отторжения трансплантата, подагры, рака, острого поражения легких, острого заболевания легких,сепсиса, респираторного дистресс-синдрома у взрослых (ARDS), заболевания периферических артерий,системного склероза, респираторного дистресс-синдрома у новорожденных, обострения астмы и COPD,муковисцидоза, диффузного панбронхиолита, поражения, вызванного реперфузией, и/или эндометриоза,к фармацевтическим композициям, содержащим указанное антитело, и способам приготовления таких композиций. Настоящее изобретение относится также к применению антитела для приготовления лекарственного средства для лечения заболеваний или нарушений, характеризующихся повышенным или несбалансированным уровнем одного или нескольких хемокинов, включающих IL-8, Gro-альфа, Gro-бета, Groгамма, GCP-2 и ENA-78 человека, а именно, хронического обструктивного заболевания легких (COPD),остеоартрита, ревматоидного артрита, эрозивного артрита, астмы, атеросклероза, воспалительного заболевания кишечника, псориаза, отторжения трансплантата, подагры, рака, острого поражения легких, острого заболевания легких, сепсиса, респираторного дистресс-синдрома у взрослых (ARDS), заболевания периферических артерий, системного склероза, респираторного дистресс-синдрома у новорожденных,обострения астмы и COPD, муковисцидоза, диффузного панбронхиолита, поражения, вызванного реперфузией, и/или эндометриоза. Несмотря на то что настоящее изобретения описано, главным образом,применительно к лечению заболеваний или нарушений у человека, настоящее изобретение может быть также использовано для лечения аналогичных заболеваний или нарушений у млекопитающих, отличных от человека. Конкретные варианты осуществления изобретения Пример 1. Получение мышиного моноклонального антитела 656.35, 197.2 и 81.1. Получение пентаспецифического моноклонального антитела против хемокинов-мишеней (IL-8,Gro-альфа, Gro-бета, Gro-гамма и ENA-78 человека). Были использованы разные методы и схемы иммунизации мышей с целью создания панспецифических моноклональных антител (mAb). Панспецифические моноклональные антитела были получены с использованием разных смесей множественных антигенных пептидов (MAP) и/или интактных хемокинов-мишеней (IL-8, Gro-, -, - иENA-78), смешанных в полном или неполном адъюванте Фрейнда (cFA или iFA), в соответствии с измененным протоколом повторной иммунизации нескольких сайтов (RIMMS) в линии мышей SJL/JOrlCrl.MAP или множественные антигенные пептиды выполняют две функции в протоколе иммунизации. Во-первых, MAP обеспечивают селективное множественное представление известной аминокислотной последовательности-мишени. Во-вторых, вызывают увеличение массы благодаря многочисленным копиям последовательности, связанным, например, при помощи лизинового ядра, что также повышает иммуногенность последовательности по сравнению с отдельными пептидами (Francis, J.P. et al., Immunology,1991: 73; 249, Schott, M.E. et al., Cell. Immunol. 1996: 174, 99-209, Tam, J.P. Proc. Natl. Acad. Sci. 1988: 85; 5409-5413). На фиг. 1 представлено схематическое изображение набора MAP, имеющих аминокислотные последовательности SEQ ID NO:49-53. Линкер в MAP может быть любым линкером, кроме лизинов. Общая временная последовательность иммунизации. Панспецифические моноклональные антитела были получены в соответствии с двумя разными протоколами иммунизации в указанной временной последовательности. 1. Первичная иммунизация (0-й день) состояла из нескольких подкожных инъекций (конечности,спина и шея) всех хемокинов-мишеней, смешанных с cFA (по 10 мкг каждого). Нижеследующие 4 бустер-дозы состояли из смеси всех хемокинов-мишеней, смешанных в iFA (по 10 мкг каждого). Пятая и все последующие бустер-дозы состояли из коктейля всех хемокинов-мишеней и всех 5 линейных MAP (по 10 мкг каждого) в iFA. Конечная бустер-доза, вводимая при помощи внутрибрюшинной (IP) инъекции за 3 дня до умерщвления и слияния, состояла из всех хемокинов-мишеней и линейных MAP в PBS. 2. Первичная иммунизация (0-й день) состояла из нескольких подкожных инъекций (конечности,спина и шея) всех пяти линейных MAP, смешанных в cFA (по 10 мкг каждого). Следующие 4 бустердозы состояли из смеси всех пяти линейных MAP в iFA (по 10 мкг каждого). Пятая и все последующие бустер-дозы состояли из коктейля всех 5 линейных MAP и всех хемокинов-мишеней (по 10 мкг каждого) в iFA. Последняя бустер-доза, вводимая при помощи внутрибрюшинной (IP) инъекции за 3 дня до умерщвления и слияния, состояла из всех 5 линейных MAP и всех хемокинов-мишеней (по 10 мкг каждого) вPBS. Пример 2. Пансвязывание моноклонального антитела с хемокинами-мишенями.(IL-8, Gro-альфа, Gro-бета, Gro-гамма, GCP-2 и ENA-78 человека) было подтверждено при помощи иммунофлуоресцентного анализа с флуоресцентным усилением и разрешением по времени. Каждый антиген (IL-8, Gro-альфа, Gro-бета, Gro-гамма, GCP-2, ENA-78 человека и hIL-18, используемый в качестве отрицательного контрольного образца) отдельно наносили на 96-луночный иммунофлуоресцентный планшет. Затем планшеты промывали и блокировали коммерчески доступным блокирующим раствором. Блокирующий раствор удаляли, в каждую лунку добавляли образцы, содержащие моноклональное антитело, и инкубировали в течение 40-60 мин (в течение указанного времени моноклональное антитело связывалось с хемокинами-мишенями). Планшеты снова промывали и в каждую лунку добавляли детектирующее антитело. В качестве детектирующих реагентов было использовано соответственно 2-кратное количество антитела 656.35, химерные (HcLc) и гуманизированные моноклональные антитела:биотинилированный IgG против мыши, биотинилированный Fab2 IgG против человека и IgG против человека, обработанный европием. Детектирующее антитело было конъюгировано с европием или биотином. Планшет снова промывали, чтобы удалить несвязанное детектирующее антитело, и в анализируемые образцы с обнаруживаемым биотином добавляли раствор стрептавидина-европия. Смесь стрептавидина-европия связывается с биотином, позволяя обнаружить указанный реагент. После выполнения последней промывки для удаления несвязанного стрептавидина-европия или в случае анализа гуманизированного моноклонального антитела после последующей промывки конъюгата с европием все лунки заполняли раствором хелатирующего детергента для активации флуоресценции, продуцируемой европием. Более сильная флуоресценция пропорциональна количеству детектирующего антитела, присутствующего в лунке, которое пропорционально количеству хемокин-связывающего моноклонального антитела. На фиг. 5 А показано связывание мышиного моноклонального антитела 656.35 с хемокинамимишенями человека. На фиг. 5 В показано связывание химерного антитела (HcLc) с хемокинами-мишенями человека. На фиг. 5 С показано связывание гуманизированных моноклональных антител с хемокинамимишенями человека. Пример 3. Функциональное панингибирование мышиным моноклональным антителом было подтверждено разными методами: CXCR2-опосредованная мобилизация Са 2+, хемотаксис нейтрофилов человека и активация нейтрофилов человека (поверхностная экспрессия CD11b). Анализ мобилизации кальция на титрационном микропланшете FLIPR (спектрофотометр для прочтения планшетов с флуорометрической визуализацией, Molecular Devices, Sunnyvale, СА [Schroeder,1996] выполняли для функционального исследования нейтрализующего воздействия антител на [Ca2+]iмобилизацию, индуцированную ELR + хемокином, в клетках CHO-K1, трансфицированных hCXCR2 иG16 и устойчиво экспрессирующих указанные вещества. За один день до анализа клетки высевали на 96-луночные планшеты с черными стенками и светлым основанием (Packard View) в количестве 40000 клеток/лунку. Через 18-24 ч из лунок отсасывали среду и заменяли 100 мкл рабочей среды, содержащей минимальную эссенциальную среду Игла (ЕМЕМ) с солями Эрла и L-глутамином, 0,1% BSA (Serologicals Corporation), 4 мкМ флуоресцентного индикаторного красителя на основе флуо-4-ацетоксиметилового эфира (флуо-4 AM) и 2,5 мМ пробенецида. Клетки инкубировали в указанной среде, содержащей краситель, в течение 1 ч при 37 С. Среду, содержащую краситель, отсасывали и заменяли такой же средой без флуо-4 AM, содержащей 0,1% желатина (BSA удаляли) и 2,5 мМ пробенецида. Клетки инкубировали в течение 10 мин при 37 С и затем трижды промывали буфером для анализа KRH [буфер Кребса-Рингера-Хенселейта (120 мМ NaCl, 4,6 мМ KCl, 1,03 мМKH2PO4, 25 мМ NaHCO3, 1 мМ CaCl2, 1,1 мМ MgCl2, 11 мМ глюкозы, 20 мМ HEPES (рН 7,4, содержащий 0,1% желатина и 2,5 мМ пробенецида]. После последней промывки буфером к клеткам добавляли 100 мкл буфера для анализа KRH, содержащего 0,1% желатина и 2,5 мМ пробенецида, нагревали до 37 С в течение 10 мин и вводили в FLIPR, где клетки, окрашенные красителем, подвергали воздействию возбуждающего излучения (488 нм), создаваемого аргонным лазером мощностью 6 Вт.[Са 2+]i-мобилизацию измеряли в виде увеличения интенсивности излучения флуо-4, связанного с Са 2+,при 516 нм. Изменение интенсивности излучения непосредственно связано с уровнями цитозольного кальция [Ca2+]i. После измерения исходного значения в течение 10 секунд в планшет добавляли 50 мкл 3-кратного количества смеси ERL + хемокин, которую предварительно инкубировали с антителом в разных концентрациях, и регистрировали данные каждую секунду в течение 1 мин, затем еще в течение 0,5 мин с интервалами в 3 с. Максимальную клеточную реакцию, которая была выше исходного значения, использовали для построения графика в программе GraphPad Prism (версия 4.03). Значение IC50 определяли в виде концентрации антитела, необходимой во время предварительной обработки 3-кратным количеством хемокина в концентрации EC80 для нейтрализации CXCR2 опосредованного стимулирующего действия смеси ELR + хемокин в концентрации EC80 на 50%. Вторичную клеточную реакцию на 25 мкМ АТР измеряли для исследования жизнеспособности клеток[Sarau, 1999]. Ингибирование индуцированного смесью ERL + хемокин кальциевого потока у человека (геометрически усредненное значение IC50, мкг/мл) тремя мышиными моноклональными антителами. (n=3-6)cloning, expression, and characterization of a cysteinyl leukotrien receptor. Mol. Pharmacol. 1999; 56:657-663. Ингибирование стимулированного IL-8 хемотаксиса нейтрофилов человека было также продемонстрировано при использовании микрокамеры Бойдена. Микрокамера Бойдена состоит из двух небольших камер, расположенных выше и ниже пористой мембраны толщиной 3 мкм. В нижнюю камеру помещают агент, вызывающий хемотаксис (т.е. IL-8), или смесь хемокина с пентаспецифическим моноклональным антителом. В верхней камере находятся очищенные неактивированные нейтрофилы человека. После сборки камеры возникает градиент концентраций из нижней части в верхнюю, стимулирующий движение нейтрофилов через мембрану к хемотаксину. Результаты анализа выражены в виде CI (хемотаксического индекса), который представляет собой отношение числа клеток, подверженных хемотаксису под воздействием стимулятора, к числу нестимулированных клеток. В процессе предварительной инкубации хемокина (т.е. IL-8) с пентаспецифическим моноклональным антителом в нижней камере указанное антитело ингибировало в зависимости от дозы стимулируемый IL-8 хемотаксис нейтрофилов. IL-8 в количестве 10 нМ обеспечивал достижение CI в пределах 3,60,8. Предварительная инкубация IL-8 с пентас- 17024621 пецифическим моноклональным антителом (656.35) в возрастающих концентрациях вызывала ингибирование в зависимости от дозы хемотаксиса нейтрофилов человека, при этом статистически значимое значение было достигнуто при CI, равном 2,20,6, при дозе 1 мкг/мл. Другие хемокины (Gro-, -, - и ENA78) нельзя было исследовать из-за используемых количеств и чувствительности анализа. Ингибирование активации очищенных нейтрофилов человека путем измерения поверхностной экспрессии CD11b. CD11b или Мас-1 опосредуют адгезию к субстратам, агрегацию и хемотаксис, и, как известно, их уровень повышается на поверхности активированных нейтрофилов (Molad Y.J. et al., Clin.Immunol. Immunopathol. 1994: 71; 281-286). Неактивированные нейтрофилы человека очищают и стимулируют ex vivo хемокинами-мишенями (т.е. IL-8) или хемокинами, предварительно инкубированными с пентаспецифическим моноклональным антителом против хемокинов. Данные выражены в виде процентного значения активации при максимальной поверхностной экспрессии CD11b в результате стимуляцииIL-8. Предварительная инкубация IL-8 с моноклональным антителом 656.35 позволяла ингибировать в зависимости от дозы повышенные уровни поверхностной экспрессии CDllb, что свидетельствует об ингибировании активации нейтрофилов (79,93,7%, 48,57,2%, 28,73,2% и 31,63,4% соответственно в дозе 0,01, 0,1, 1, 10 и 50 мкг/мл). Пример 4. Значения ассоциации/диссоциации пентаспецифических моноклональных антител для каждого хемокина-мишени (IL-8, Gro-бета и ENA-78 человека). Методы анализа Biacore. Для выполнения эксперимента, именуемого KL, IgG-Fc кролика против мыши (Biacore BR-1005-14) иммобилизуют на чипе СМ 5 путем связывания с первичным амином в соответствии с инструкциями изготовителей. Для выполнения эксперимента, именуемого BE, очищенные антитела иммобилизуют на чипе путем связывания с амином. Супернатант или очищенное антитело, выделенное из родительского мышиного моноклонального антитела, улавливают на поверхности IgG-Fc против мыши. Каждое анализируемое вещество (IL-8,Gro-бета, ENA-78) в определенных концентрациях пропускают над поверхностью иммобилизованного или уловленного антитела, при этом для каждого впрыскивания анализируемого вещества используют вновь уловленное антитело. После каждого впрыскивания анализируемого вещества поверхность восстанавливают, впрыскивая слабый кислотный раствор, который удаляет уловленное антитело, но не оказывает значительного влияния на способность IgG-Fc против мыши выполнять другое улавливание и не влияет на иммобилизованные антитела. На поверхность уловленного антитела также впрыскивают буфер для двойного контроля. Данные анализируют при помощи программного обеспечения для анализа, предназначенного для данного компьютера, используя модель связывания в отношении 1:1. Пример 5. Картирование эпитопов. Было произведено картирование эпитопов моноклонального антитела 656.35, при этом было установлено, что указанное антитело связывается с эпитопом KELRCQCIKTYSKP (SEQ ID NO:54) в IL-8 человека. Таким образом, другой вариант осуществления настоящего изобретения относится к пентаспецифическому антителу, которое связывается с эпитопом SEQ ID NO:54 IL-8 человека. Пример 6. Исследование эффективности химерного антитела, полученного из антитела 656.35, имеющего последовательность тяжелой цепи SEQ ID NO:56 и последовательность легкой цепи SEQ ID NO:58. (Указанное антитело будет именоваться химерным антителом). Исследования in vivo. Для исследования способности пентаспецифического антитела ингибировать инфильтрацию нейтрофилов в легкие приматов кроме человека (NHP - циномолгус) была использована модель острого воспалительного заболевания легких, вызванного ингаляцией LPS. У приматов (NHP) обследовали состояние здоровья и восприимчивость к экзогенно добавляемому IL-8 циномолгуса (цино-IL-8). У отобранных приматов брали образцы для получения исходных данных (кровь и бронхоальвеолярный лаваж - BAL) за пять дней до первой стимуляции LPS. В процессе стимуляции LPS приматов транквилизировали при помощи одной внутримышечной инъекции гидрохлорида кетамина (10 мг/кг). После достижения седативного эффекта приматов анестезировали путем внутривенного вливания пропофола (0,2 мг/кг/мин, в случае необходимости). Анестезированных животных помещали на грелку-одеяло с цирку- 18024621 лирующей теплой водой и/или обертывали одеялом с циркулирующим теплым воздухом и в каждый глаз вводили глазную мазь. Затем животных интубировали и искусственно вентилировали легкие для достижения стимуляции. Для стимуляции использовали дыхательный аппарат с положительным давлением и регулируемым объемом (дыхательный аппарат для кошек/кроликов компании Stoelting,www.stoeltingco.comпо каталогу 5019510). Стимуляцию липополисахаридом (LPS) выполняли путем ингаляции при помощи ультразвукового распылителя Ultraneb-99 компании DeVilbiss. Аэрозоль LPS вводили в течение 5 мин в количестве 100 мкг/мл. Во время стимуляции контролировали и регистрировали частоту сердечных сокращений,температуру тела и частоту дыхания. Первичная стимуляция подтвердила, что приматы реагируют наLPS (повышенная инфильтрация нейтрофилов в легкие и повышенные уровни цино-IL-8). Индивидуальные реакции приматов были подтверждены образцами, полученными через 6 и 24 ч после стимуляции. Приматов (NHP), подвергнутых первичной стимуляции LPS, произвольно распределяли в две группы. После восстановления в течение 4 недель у всех приматов брали образцы крови и BAL. Через четыре дня после измерения исходных показателей подопытным животным внутривенно инъецировали наполнитель или 1 мг/кг химерного антитела. На следующий день приматов снова стимулировали ингалируемым LPS и через 6 и 24 ч после стимуляции брали образцы для анализа. После дополнительного периода восстановления у приматов, которым вводили наполнитель, брали исходные образцы и через 4 дня указанным приматам делали одну внутривенную инъекцию ударной дозы химерного антитела в количестве 10 мг/кг. На следующий день животных стимулировали LPS и через 6 и 24 ч брали образцы для анализа. Ингаляция LPS представляет собой модель острого воспалительного заболевания, стимулирующую повышение хемотаксических хемокинов, таких как IL-8, которые вызывают повышенную инфильтрацию нейтрофилов в легкие. Первичным параметром эффективности химерного антитела является ингибирование инфильтрации нейтрофилов в легкие приматов, стимулированных LPS. Инфильтрация нейтрофилов в указанной модели острого воспалительного заболевания происходит в течение первых 24 ч после стимуляции LPS, после чего более выраженными становятся другие воспалительные процессы. Предварительное введение химерного антитела значительно и в зависимости от дозы ингибирует инфильтрацию нейтрофилов в легкие приматов, стимулированных LPS. Введение химерного антитела также предотвращает повышение уровней циркулирующих нейтрофилов, не влияя на действительную функцию нейтрофилов (т.е. способность клеток к фагоцитозу). Введение химерного антитела также не оказывало сильного воздействия на клетки других типов, такие как макрофаги/моноциты, в легких или кровотоке. См. фиг. 2. Пример 7. Дальнейшие исследования гуманизированных моноклинальных антител. Было получено несколько конструкций на основе гуманизированного пентаспецифического антитела, четыре из которых, как было показано, прочно связываются со всеми хемокинами-мишенями и ингибируют указанные хемокины. (Для ознакомления с каждой последовательностью см. приведенную ниже информацию о последовательностях). Указанный анализ состоял из измерений сродства (BiaCore), анализов мобилизации кальция (функциональный анализ in vitro, FLIPR) и анализа стимуляции нейтрофиловBiaCore. Кинетику гуманизированных и химерных конструкций определяли методом улавливания белка А,выполняемым следующим образом. Белок А иммобилизуют на чипе СМ 5 путем связывания с первичным амином в соответствии с инструкциями изготовителя. Очищенное гуманизированное или химерное антитело улавливают на поверхности белка А. Каждое анализируемое вещество (IL-8, Gro- и ENA-78) в определенных концентрациях пропускают над поверхностью уловленного антитела, выполняя отдельное улавливание для каждого впрыскивания анализируемого вещества. После каждого впрыскивания анализируемого вещества поверхность регенерируют, впрыскивая слабый кислотный раствор, который удаляет уловленное антитело,но не оказывает значительного воздействия на способность белка А выполнять следующее улавливание. На поверхность уловленного антитела также впрыскивают буфер для двойного контроля. Данные анализируют при помощи программного обеспечения для анализа, предназначенного для данного компьютера,используя модель связывания в отношении 1:1. Сродство (KD) определяют, исследуя скорость ассоциации (ka) и скорость диссоциации (kd) белков.HcLc означает химерное антитело, полученное из антитела 656.35, имеющего последовательность тяжелой цепи SEQ ID NO:56 и последовательность легкой цепи SEQ ID NO:58. Функциональный анализ: мобилизация кальция (FLIPR) с использованием CXCR2 (с G16) в клетках CHO-K1. При выполнении анализа мобилизации кальция на титрационном микропланшете FLIPR (спектрофотометр для прочтения планшетов с флуорометрической визуализацией, Molecular Devices, Sunnyvale,СА, [Schroeder, 1996]) был использован для функционального исследования нейтрализующего воздействия антител на [Ca2+]i-мобилизацию, индуцированную ELR+ хемокином, в клетках CHO-K1, трансфицированных hCXCR2 и G16 и устойчиво экспрессирующих указанные вещества. За один день до анализа клетки высевали на 96-луночные планшеты с черными стенками и светлым основанием (Packard View) в количестве 40000 клеток на лунку. Через 18-24 ч из лунок отсасывали среду и заменяли 100 мкл рабочей среды, содержащей минимальную эссенциальную среду Игла (ЕМЕМ) с солями Эрла и L-глутамином, 0,1% BSA (Serologicals Corporation), 4 мкМ флуоресцентного индикаторного красителя на основе флуо-4-ацетоксиметилового эфира (флуо-4 AM) и 2,5 мМ пробенецида. Клетки инкубировали в указанной среде, содержащей краситель, в течение 1 ч при 37 С. Среду, содержащую краситель, отсасывали и заменяли такой же средой без флуо-4 AM, содержащей 0,1% желатина (BSA удаляли) и 2,5 мМ пробенецида. Клетки инкубировали в течение 10 мин при 37 С и затем трижды промывали буфером для анализа KRH [буфер Кребса-Рингера-Хенселейта (120 мМ NaCl, 4,6 мМ KCl,1,03 мМ KH2PO4, 25 мМ NaHCO3, 1,0 мМ CaCl2, 1,1 мМ MgCl2, 11 мМ глюкозы, 20 мМ HEPES (рН 7,4,содержащий 0,1% желатина и 2,5 мМ пробенецида]. После последней промывки буфером к клеткам добавляли 100 мкл буфера для анализа KRH, содержащего 0,1% желатина и 2,5 мМ пробенецида, нагревали до 37 С в течение 10 мин и вводили в FLIPR, где клетки, окрашенные красителем, подвергали воздействию возбуждающего излучения (488 нм), создаваемого аргонным лазером мощностью 6 Вт.[Са 2+]i-мобилизацию измеряли в виде увеличения интенсивности излучения флуо-4, связанного с Са 2+,при 516 нм. Изменение интенсивности излучения непосредственно связано с уровнями цитозольного кальция [Ca2+]i. После измерения исходного значения в течение 10 секунд в планшет добавляли 50 мкл 3-кратного количества смеси ERL + хемокин, которую предварительно инкубировали с антителом в разных концентрациях, и регистрировали данные каждую секунду в течение 1 мин, затем еще в течение 0,5 мин с интервалами в 3 с. Максимальную клеточную реакцию, которая была выше исходного значения, использовали для построения графика в программе GraphPad Prism (версия 4.03). Значение IC50 определяли в виде концентрации антитела, необходимой во время предварительной обработки 3-кратным количеством хемокина в концентрации ЕС 80 для нейтрализации CXCR2 опосредованного стимулирующего действия смеси ELR + хемокин в концентрации ЕС 80 на 50%. Вторичную клеточную реакцию на 25 мкМ АТР измеряли для исследования жизнеспособности клеток[Sarau, 1999]. Ингибирование индуцированного смесью ERL + хемокин кальциевого потока человека (геометрически усредненное значение IC50, мкг/мл) четырьмя конструкциями гуманизированного моноклонального антитела (n=3)cloning, expression, and characterization of a cysteinyl leukotrien receptor. Mol Pharmacol. 1999;56:657-663. Анализ стимуляции нейтрофилов человека CD11b ex vivo. Способность гуманизированных пентаспецифических антител предотвращать индуцированную хемокинами активацию очищенных нейтрофилов человека исследовали при помощи проточной цитометрии, анализируя физические изменения клеток (размер и форма - грануляция) и маркеры поверхностной активации (повышенная поверхностная экспрессия CD11b). Контрольный стимулятор нейтрофиловfMLP использовали для подтверждения способности очищенных нейтрофилов человека активироваться,см. фиг. 4. Информация о последовательностях. Полную РНК экстрагировали из клеток гибридомы 656.35, 197.2 и 81.1, затем получали последовательность кДНК вариабельной области тяжелой и легкой цепей, выполняя обратную транскрипцию и полимеразную цепную реакцию (RT-PCR). Верхняя затравка для RT-PCR представляла собой смесь вырожденных затравок, специфичных для лидерных последовательностей гена иммуноглобулина мыши, и нижняя затравка была специфична для константных областей антитела, в данном случае изотипаIgG2a/K. Затравки были созданы в соответствии с описанием, приведенным в публикации Джонса и Бендига (Bio/Technology 9:88, 1991). RT-PCR выполняли дважды для обеих последовательностей V-области,чтобы можно было произвести последующую проверку правильных последовательностей V-области. Продукты V-области, полученные при помощи RT-PCR, клонировали (набор для клонирования ТАInvitrogen) и получали данные для последовательностей. Данный процесс оказался безуспешным для вариабельной области легкой цепи 81.1. Таким образом, аминокислотная последовательность, показанная ниже (SEQ ID NO:12), была получена в результате секвенирования белков легкой цепи, выделенной методом электрофореза в полиакриламидном геле с додецилсульфатом натрия в восстановительных условиях. Области, определяющие комплементарность, (CDR) подчеркнуты. Полинуклеотидные последовательности для вариабельных областей тяжелой и легкой цепей (соответственно SEQ ID NO:1 и 3) для антитела 656.35. Полипептидные последовательности для вариабельных областей тяжелой и легкой цепей (соответственно SEQ ID NO:2 и 4) для антитела 656.35. Полипептидные последовательности для CDR тяжелой цепи (соответственно SEQ ID NO:13, 14, 15) для антитела 656.35.SEQ ID NO:15: Полипептидные последовательности для CDR легкой цепи (SEQ ID NO:16, 17, 18) для антитела 656.35. Полипептидные последовательности для CDR тяжелой цепи (SEQ ID NO:31, 32, 33) для антитела 656.35.SEQ ID NO:33: Полинуклеотидные последовательности для CDR легкой цепи (SEQ ID NO:34, 35, 36) для антитела 656.35. SEQ ID NO:34SEQ ID NO:36: Полинуклеотидные последовательности для вариабельных областей тяжелой и легкой цепей (соответственно SEQ ID NO:5 и 7) для антитела 197.2. Полипептидные последовательности для вариабельных областей тяжелой и легкой цепей (соответственно SEQ ID NO:6 и 8) для антитела 197.2. Полипептидные последовательности для CDR тяжелой цепи (соответственно SEQ ID NO:19, 20, 21) для антитела 197.2.SEQ ID NO:21: Полипептидные последовательности для CDR легкой цепи (SEQ ID NO:22, 23, 24) для антитела 197.2.SEQ ID NO:24: Полинуклеотидные последовательности для CDR тяжелой цепи (SEQ ID NO:37, 38, 39) для антитела 197.2.SEQ ID NO:39: Полинуклеотидные последовательности для CDR легкой цепи (SEQ ID NO:40, 41, 42) для антитела 197.2.SEQ ID NO:42: Полинуклеотидная последовательность для вариабельной области тяжелой цепи (SEQ ID NO:9) для антитела 81.1. Полипептидная последовательность для вариабельной области тяжелой цепи (SEQ ID NO:10) для антитела 81.1. Полипептидные последовательности для CDR тяжелой цепи (соответственно SEQ ID NO:25, 26, 27) для антитела 81.1.SEQ ID NO:27: Полинуклеотидные последовательности для CDR тяжелой цепи (SEQ ID NO:43, 44, 45) для антитела 81.1.SEQ ID NO:45 Полипептидная последовательность для вариабельной области легкой цепи (SEQ ID NO:12) для антитела 81.1. Полипептидные последовательности для CDR легкой цепи (соответственно SEQ ID NO:28, 29 и 30) для антитела 81.1.SEQ ID NO:30: Полинуклеотидная последовательность химерного антитела (вариабельная область тяжелой цепи + Полипептидная последовательность химерного антитела (вариабельная область тяжелой цепи + Полинуклеотидная последовательность химерного антитела (вариабельная область легкой цепи + Полипептидная последовательность химерного антитела (вариабельная область легкой цепи + cK человека с оптимизированным кодоном) SEQ ID NO:58: Химерное антитело является химерным антителом, полученным из мышиного антитела 656.35. Последовательность ДНК Н 0 зрелой тяжелой цепи SEQ ID NO:11: Белковая последовательность Н 0 зрелой тяжелой цепи SEQ ID NO:46: Последовательность ДНК L7 зрелой легкой цепи SEQ ID NO:47: Белковая последовательность L7 зрелой легкой цепи SEQ ID NO:48: Последовательность ДНК L8 зрелой легкой цепи SEQ ID NO:59: Белковая последовательность L8 зрелой легкой цепи SEQ ID NO:60:
МПК / Метки
МПК: C12N 15/63, C12N 15/13, C07K 16/24, A61K 39/395, C12P 21/02
Метки: антитела, способы, получения, пентаспецифические, применения
Код ссылки
<a href="https://eas.patents.su/30-24621-pentaspecificheskie-antitela-sposoby-ih-polucheniya-i-primeneniya.html" rel="bookmark" title="База патентов Евразийского Союза">Пентаспецифические антитела, способы их получения и применения</a>
Моноклональные антитела к растворимому рецептору лпнп человека, способы их получения и применения и гибридомы, продуцирующие указанные антитела

Номер патента: 7494
Опубликовано: 27.10.2006
Авторы: Смоларски Моше, Дреано Мишель, Бельзер Илана, Суисса Дани, Антонетти Франческо, Йонах Начум
МПК: C07K 16/28, A61P 31/20, C12N 5/20...
Метки: указанные, моноклональные, получения, продуцирующие, применения, рецептору, гибридомы, способы, лпнп, растворимому, антитела, человека
Формула / Реферат:
1. Моноклональное антитело, химерное антитело, полностью гуманизированное антитело, антитело к антиидиотипическому антителу или его фрагмент, которое специфически распознает и связывает растворимый рецептор ЛПНП человека и его фрагменты, которое получают путем иммунизации животных формой +291 растворимого рецептора ЛПНП человека, включающей аминокислотную последовательность от Asp +4 до Glu +291 последовательности ЛПНП человека, и которое...
Антитела против каспазы-8 и способы их получения и применения

Номер патента: 9770
Опубликовано: 28.04.2008
Авторы: Колумам Ганеш, Гончаров Таня, Уоллах Дэвид
МПК: C07K 14/47, C12N 9/64, C07K 16/40...
Метки: получения, антитела, применения, каспазы-8, против, способы
Формула / Реферат:
1. Антитело против каспазы-8, полученное путем иммунизации животного С-концевым пептидом субъединицы Sub-1 каспазы-8, или его фрагмент, где пептид, используемый для иммунизации, имеет аминокислотную последовательность CQGDNYQKGIPVETD (SEQ ID NO: 4), способное совместно иммунопреципитировать указанную каспазу (как активную каспазу-8, так и прокаспазу-8) вместе со связанным с каспазой-8 белком р72 (SEQ ID NO: 3) и эффективно высвобождать каспазу и...
Молекула антитела, обладающая специфичностью по отношению к kdr человека, и способы применения антитела

Номер патента: 10970
Опубликовано: 30.12.2008
Авторы: Моррисон Роберт Кендалл, Попплвелл Эндрю Джордж, Зинкевич-Пеотти Карен, Тикл Саймон Питер
МПК: A61K 39/395, A61K 47/48, A61P 17/06...
Метки: молекула, специфичностью, человека, обладающая, антитела, применения, способы, отношению
Формула / Реферат:
1. Молекула антитела, обладающая специфичностью по отношению к KDR человека, содержащая тяжелую цепь, в которой вариабельный домен содержит по крайней мере одну CDR, имеющую последовательность, указанную как H1 на фиг. 1 (SEQ ID NO: 1) для CDRH1, H2 на фиг. 1 (SEQ ID NO: 2) для CDRH2 или H3 на фиг. 1 (SEQ ID NO: 3) для CDRH3. 2. Молекула антитела, обладающая специфичностью по отношению к KDR человека, содержащая легкую цепь, в которой...
Способ получения обогащённого поликлонального антитела, высокоспецифичного к антигену поверхностного полисахарида липоарабиноманнана микобактерий, и способы его применения

Номер патента: 13228
Опубликовано: 30.04.2010
Авторы: Кульчин Владимир А., Молокова Елена В., Керрик Джиль Л.
МПК: G01N 33/569, C07K 16/12
Метки: липоарабиноманнана, применения, полисахарида, поверхностного, микобактерий, способ, высокоспецифичного, поликлонального, способы, антитела, антигену, получения, обогащённого
Формула / Реферат:
1. Способ получения обогащенного поликлонального антитела, высокоспецифичного к антигену поверхностного полисахарида липоарабиноманнана (LAM) микобактерии, включающийнанесение сыворотки из млекопитающего, зараженного микобактериями, на первый аффинный матрикс, приготовленный с выделенным антигеном LAM микобактерии, так что поликлональное антитело, специфичное к выделенному антигену LAM, удерживается первым аффинным матриксом;выделение...
Антитела к cd154, их фрагменты и способы применения антител и фрагментов

Номер патента: 14226
Опубликовано: 29.10.2010
Авторы: Ханф Карл Дж.М., Ван Влеймен Герман, Луговской Алексей Александрович
МПК: A61K 39/395, C07K 16/28
Метки: способы, фрагментов, cd154, антитела, применения, фрагменты, антител
Формула / Реферат:
1. Легкая цепь антитела к CD154, содержащая аминокислотную последовательность, которая отличается от последовательности SEQ ID NO: 1 по крайней мере одной аминокислотой в положениях 24, 26, 27, 28, 30, 31, 32, 33, 34, 35, 36, 38, 54, 57, 58, 59, 60, 93, 95, 96, 97, 98, 100 и 101 и которая по меньшей мере на 80% идентична SEQ ID NO: 3, причем указанная легкая цепь способна образовывать комплекс с тяжелой цепью антитела 5с8 (АТСС НВ 10916),...
Предыдущий патент: Сахаридсодержащий продукт и способ его получения
Следующий патент: Регулирующий клапан для регулирования потока текучей среды