Пептид, индуцирующий противоопухолевый иммунный ответ, и его применение
Номер патента: 23013
Опубликовано: 29.04.2016
Авторы: Вайншенк Тони, Шоор Оливер, Сингх Харприт, Хильф Норберт, Вальтер Штеффен, Траутвайн Клаудия





























Формула / Реферат
1. Пептид, индуцирующий противоопухолевый иммунный ответ, где указанный пептид имеет общую длину между 8 и 16 аминокислот и включает последовательность, которая выбирается из группы SEQ ID No. 1-9, 11-13, 15-24 и 26-30, где указанный пептид связывается с молекулой главного комплекса гистосовместимости человека (МНС) класса I или II и где указанный пептид способен стимулировать Т-клетки CD4 и/или CD8.
2. Пептид по п.1, где указанный пептид имеет аминокислотную последовательность согласно SEQ ID No. 1-9, 11-13, 15-24 и 26-30.
3. Пептид по п.1 или 2, где указанный пептид включает непептидные связи, выбранные из группы -CH2-NH, -CH2S-, -СН2СН2-, -СН=СН-, -СОСН2-, -СН(ОН)СН2- и -CH2SO-.
4. Пептид по любому из пп.1-3, где указанный пептид является белком слияния, в частности включающего N-терминальные аминокислоты антиген-ассоциированной инвариантной цепи (Ii) HLA-DR.
5. Нуклеиновая кислота, кодирующая пептид по любому из пп.1-4.
6. Вектор, способный экспрессировать нуклеиновую кислоту по п.5.
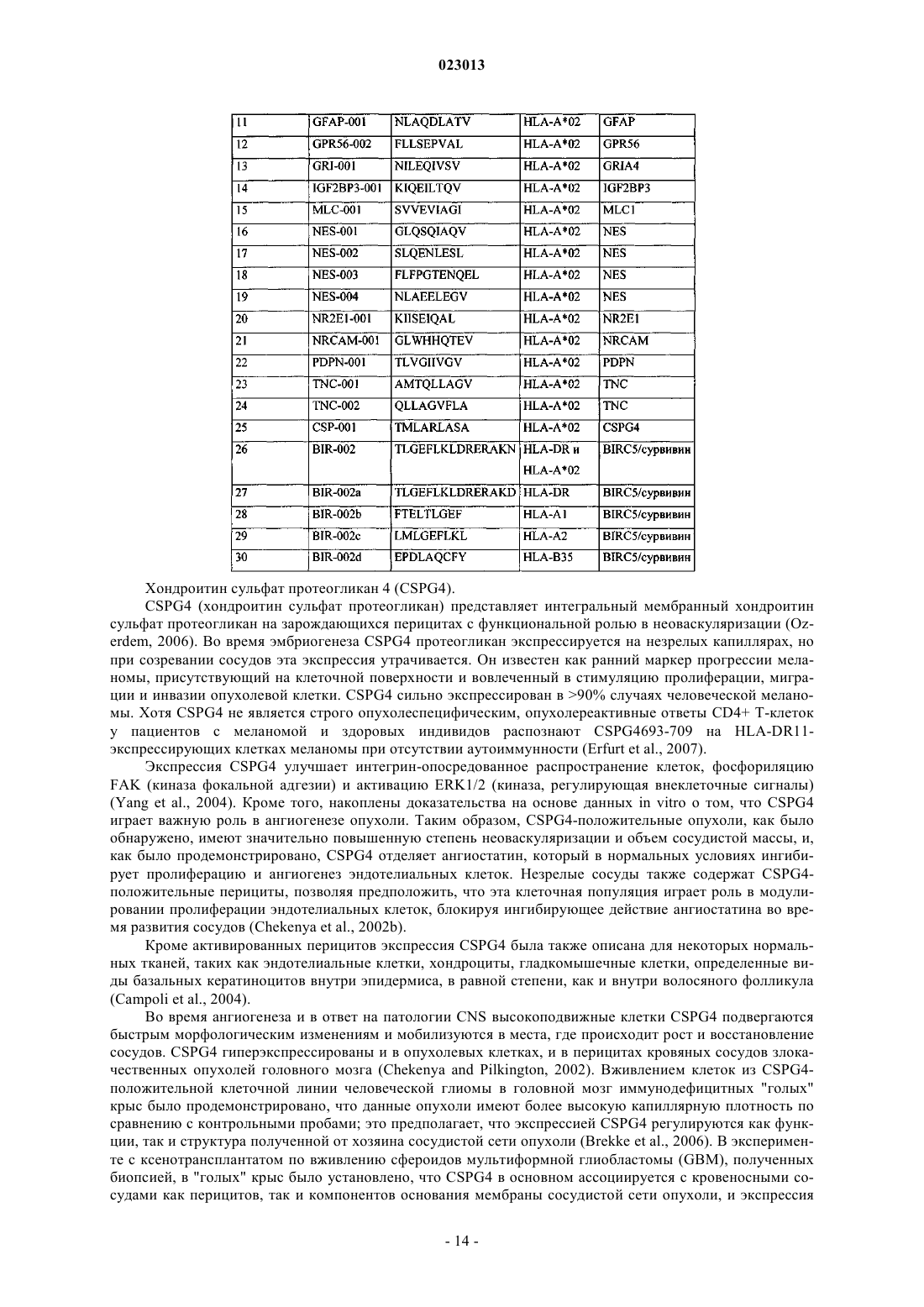
7. Клетка-хозяин, включающая нуклеиновую кислоту по п.5 или вектор по п.6, где указанная клетка-хозяин является антигенпрезентирующей клеткой или дендритной клеткой.
8. Способ получения пептида по любому из пп.1-4, включающий культивирование клетки-хозяина по п.7, которая экспрессирует нуклеиновую кислоту по п.5 или вектор по п.6, и выделение пептида из клетки-хозяина или его культуральной среды.
9. Способ получения активированных цитотоксических Т-лимфоцитов (ЦТЛ) in vitro, включающий контактирование ЦТЛ in vitro с нагруженными антигеном молекулами человека МНС I или II класса, экспрессированными на поверхности подходящей антигенпрезентирующей клетки или искусственной конструкции, имитирующей антигенпрезентирующую клетку, на период времени, достаточный для активации указанных ЦТЛ антиген-специфическим образом, где указанный антиген является пептидом по любому из пп.1-3.
10. Активированный цитотоксический Т-лимфоцит (ЦТЛ), полученный с помощью способа по п.9, который селективно распознает клетку, которая аберрантно экспрессирует полипептид, включающий аминокислотную последовательность, представленную в любом из пп.1-3.
11. Антитело, которое специфически связывается с пептидом по пп.1, 2 в комплексе с главным комплексом гистосовместимости человека (МНС) класса I или II, где указанным антителом предпочтительно является поликлональное антитело, моноклональное антитело и/или химерное антитело.
12. Применение пептида по любому из пп.1-3 для изготовления вакцины для лечения рака.
13. Применение нуклеиновой кислоты по п.5 для изготовления вакцины для лечения рака.
14. Применение вектора по п.6 для изготовления вакцины для лечения рака.
15. Применение клетки-хозяина по п.7 для изготовления вакцины для лечения рака.
16. Применение активированного цитотоксического Т-лимфоцита по п.10 для изготовления вакцины для лечения рака.
17. Применение антитела по п.11 для изготовления вакцины для лечения рака.
18. Применение по любому из пп.12-17, где указанный рак выбирается из астроцитомы, пилоидной астроцитомы, дисэмбриопластической нейроэпителиальной опухоли, олигодендроглиомы, эпендимомы, мультиформной глиобластомы, смешанных глиом, олигоастроцитомы, медуллобластомы, ретинобластомы, нейробластомы, герминомы, тератомы, ганглиоглиомы, ганглиоцитомы, центральной ганглиоцитомы, примитивных нейроэктодермальных опухолей (PNET, к примеру, медуллобластомы, медуллоэпителиомы, нейробластомы, ретинобластомы, эпендимобластомы), опухолей паренхимы шишковидной железы (к примеру, пинеоцитомы, пинеобластомы), опухолей, развивающихся из эпендимных клеток, опухолей хориоидного сплетения, нейроэпителиальных опухолей неопределенного происхождения (к примеру, глиоматоза головного мозга, астробластомы), глиобластомы, опухоли предстательной железы, рака груди, рака пищевода, колоректального рака, светлоклеточной почечно-клеточной карциномы, рака легких, ЦНС, яичника, меланомы, рака поджелудочной железы, плоскоклеточной карциномы, лейкемии или медуллобластомы, опухолей толстого кишечника, прямого кишечника, желудка, почек, легких, поджелудочной железы, предстательной железы, кожи.
19. Набор для индукции противоопухолевого иммунного ответа, включающий:
(a) контейнер, который содержит фармацевтическую композицию, содержащую пептид по любому из пп.1-4, нуклеиновую кислоту по п.5, вектор по п.6, клетку по п.7, активированный цитотоксический Т-лимфоцит по п.10 или антитело по п.11 в буферном растворе или в лиофилизованной форме;
(b) второй контейнер, содержащий буферный разбавитель или восстанавливающий раствор для лиофилизованного состава.
20. Набор по п.19, дополнительно включающий инструкции по использованию раствора или восстановлению и использованию лиофилизованного состава.
21. Набор по п.19 или 20, дополнительно содержащий фильтр, иглу или шприц.
Текст
ПЕПТИД, ИНДУЦИРУЮЩИЙ ПРОТИВООПУХОЛЕВЫЙ ИММУННЫЙ ОТВЕТ, И ЕГО ПРИМЕНЕНИЕ Изобретение относится к пептидам, индуцирующим противоопухолевый иммунный ответ, где указанные пептиды связываются с молекулой главного комплекса гистосовместимости человека(МНС) класса I или II и где указанные пептиды способны стимулировать Т-клетки CD4 и/ или CD8, и их применению в вакцинах для лечения рака. Изобретение относится также к нуклеиновым кислотам, кодирующим указанные пептиды, векторам, экспрессирующим указанные пептиды, клеткам-хозяевам, включающим указанные нуклеиновые кислоты, и их применению в вакцинах для лечения рака, а также к способам получения указанных пептидов и активированных цитотоксических Т-лимфоцитов и наборам для индукции противоопухолевого иммунного ответа.(71)(73) Заявитель и патентовладелец: ИММАТИКС БАЙОТЕКНОЛОДЖИЗ ГМБХ (DE) 023013 Изобретение относится к пептидам, индуцирующим противоопухолевый иммунный ответ, где указанные пептиды связываются с молекулой главного комплекса гистосовместимости человека (МНС) класса I или II и где указанные пептиды способны стимулировать Т-клетки CD4 и/или CD8, и их применению в вакцинах для лечения рака. Изобретение относится также к нуклеиновым кислотам, кодирующим указанные пептиды, векторам, экспрессирующим указанные пептиды, клеткам-хозяевам, включающим указанные нуклеиновые кислоты, и их применению в вакцинах для лечения рака, а также к способам получения указанных пептидов и активированных цитотоксических Т-лимфоцитов и наборам для индукции противоопухолевого иммунного ответа. Уровень техники Глиомы являются опухолями головного мозга, развивающимися из глиальных клеток нервной системы. Глиальные клетки, называемые обычно нейроглией или просто глией, являются не-нейрональными клетками, которые обеспечивают поддержку и питание, поддерживают гомеостаз, формируют миелин и участвуют в передаче сигналов в нервной системе. Две наиболее важные подгруппы глиом - это астроцитомы и олигодендроглиомы, называемые в соответствии с видами нормальных глиальных клеток, из которых они образуются (астроциты или олигодендроциты соответственно). Относящаяся к подгруппе астроцитом мультиформная глиобластома (называемая далее глиобластомой) является наиболее распространенной злокачественной опухолью головного мозга взрослых людей и на ее счет приходится приблизительно 40% всех злокачественных опухолей головного мозга и приблизительно 50% глиом. Она агрессивно поражает центральную нервную систему и имеет наивысший уровень злокачественности(степень IV) среди всех глиом. Хотя наблюдается постоянный прогресс в их лечении благодаря усовершенствованию нейровизуализации, микрохирургии, различным терапевтическим возможностям, таким как Темозоломид или облучение, глиобластомы остаются неизлечимыми. Процент летальных исходов при этой опухоли головного мозга очень высок: средняя ожидаемая продолжительность жизни после постановки первого диагноза составляет 9-12 месяцев. 5-летний срок выживаемости в течение периода наблюдения с 1986 по 1990 гг. составил 8,0%. На данный момент пятилетний срок выживаемости вследствие агрессивной терапии, включая макроскопическое удаление опухоли, все еще ниже 10%. Соответственно, существует высокая потребность медицины в альтернативном и эффективном методе лечения. Опухолевые клетки глиобластомы являются наиболее недифференцированными среди опухолей головного мозга, так что опухолевые клетки имеют высокий потенциал для миграции и пролиферации и являются высокоинвазивными, приводя к очень неутешительному прогнозу. Глиобластомы приводят к смерти из-за быстрого, агрессивного и инфильтрирующего роста в головном мозге. Паттерны инфильтрирующего роста являются причиной нерезектабельности этих опухолей. Глиобластомы также относительно резистентны к облучению и химиотерапии, поэтому высок процент рецидивов после лечения. Кроме того, иммунный ответ на неопластические клетки довольно неэффективен для полного устранения всех неопластических клеток после резекции и лучевой терапии. Глиобластома подразделяется на первичную глиобластому (de novo) и вторичную глиобластому в зависимости от различий в генном механизме во время злокачественной трансформации недифференцированных астроцитов или глиальных клеток-предшественников. Вторичная глиобластома возникает в более молодом возрасте, до 45 лет. В течение 4-5 лет, в среднем, вторичная глиобластома развивается из астроцитомы низкой степени, проходя стадию недифференцированной астроцитомы. Первичная глиобластома, напротив, возникает в более пожилом возрасте, в среднем это 55 лет. В целом, первичная глиобластома возникает как молниеносная глиобластома, характеризуясь прогрессированием опухоли в течение 3 месяцев, начиная с состояния без клинических или патологических отклонений (Pathology andGenetics of the Nervous Systems. 29-39 (IARC Press, Lyon, France, 2000. Глиобластома мигрирует вдоль миелинизированных нервов и широко распространяется по центральной нервной системе. В большинстве случаев хирургическое вмешательство приводит лишь к ограниченно устойчивому терапевтическому эффекту. Злокачественные клетки глиомы ускользают от обнаружения иммунной системой хозяина за счет выработки иммуноподавляющих веществ, которые нарушают пролиферацию Т-клеток и выработку иммуностимулирующего цитокина ИЛ-2. Внутричерепные новообразования могут возникнуть из любых структур или видов клеток, присутствующих в ЦНС, включая головной мозг, мягкие мозговые оболочки, мозговой придаток, череп и даже остаточную эмбриональную ткань. Общая ежегодная частота заболеваемости первичными опухолями головного мозга в Соединенных Штатах составляет 14 случаев на 100000. Наиболее распространенные первичные опухоли головного мозга - менингиомы, составляющие 27% всех первичных опухолей головного мозга, и глиобластомы, составляющие 23% всех первичных опухолей головного мозга (причем на счет глиобластомы приходятся 40% всех случаев злокачественных опухолей головного мозга взрослого населения). Многие из этих опухолей агрессивны и имеют высокую степень злокачественности. Первичные опухоли головного мозга являются наиболее распространенными солидными опухолями у детей и второй по частоте причиной смерти от рака после лейкемии у детей. Поиск методов эффективного лечения пациентов от глиобластом продолжается и сейчас. Была изучена иммунотерапия, или лечение посредством стимуляции иммунной системы, в целях борьбы с данными неопластическими клетками. Первые обнадеживающие результаты от использования иммунотера-1 023013 певтического подхода для пациентов, страдающих от глиобластомы, были получены "Northwest Therapeutics" с использованием "DCVax Brain", который представляет собой основанную на клетках вакцинацию с использованием полученных от пациента дендритных клеток, нагруженных лизатами аутологичных опухолевых клеток, и "Celldex", в котором использовался пептид из EGFRvIII для индуцирования антигенспецифических ответов ЦТЛ, которые, в свою очередь, соотносились с увеличенным средним сроком выживаемости по сравнению со сроками при использовании стандартного лечения (Heimberger etal., 2006). Колоректальная карцинома. По данным Американского общества рака, колоректальный рак (КРР) занимает третье место по частоте распространения в США, это заболевание поражает ежегодно более 175000 новых пациентов. В США, Японии, Франции, Германии, Италии, Испании и Великобритании оно поражает более 480000 пациентов. Это одна из наиболее частых причин смертности от рака в развитых странах. 1-годичный и 5 летний срок выживаемости для людей с колоректальным раком составляет 84 и 64% соответственно. Срок выживаемости продолжает снижается после 5 лет до 57% по истечении 10 лет после постановки диагноза. Если колоректальный рак обнаруживают на ранней, локализованной стадии, то 5-летняя выживаемость составляет 90%; однако на этой стадии диагностируются лишь 39% случаев заболевания колоректальным раком, по причине слишком низкого процента обследований. После того как рак распространился регионально, поразив соседние органы или лимфатические узлы, 5-летняя выживаемость сокращается до 68%. Для лиц с отдаленными метастазами 5-летняя выживаемость составляет 10%. Исследователи предполагают, что возникновение колоректального рака является результатом взаимодействия между наследственными факторами и факторами окружающей среды. В большинстве случаев появляются аденоматозные полипы, становясь предшественниками колоректальных опухолей, хотя процесс их перехода в опухоль может занять много лет. Первостепенный фактор риска для заболевания колоректальным раком - это возраст, в 90% случаев диагноз был поставлен в возрасте более 50 лет. Другие факторы риска заболевания колоректальным раком по данным Американского общества рака включают употребление алкоголя, питание с высоким содержанием жиров и/или сырого мяса и неадекватное потребление фруктов и овощей. Число новых случаев продолжает расти, в особенности на территории таких стран, как Япония, где виновником может быть внедрение западных образцов питания с избыточным потреблением жира и мяса и снижение потребления волокнистой пищи. Несмотря на это частота заболеваемости растет не так быстро, как раньше, что может быть связано с увеличением количества скринингов и удалением полипов, предотвращающими прогрессирование заболевания и переход полипов в рак. Как и при большинстве солидных опухолей, терапия первой линии - это операция, однако только пациенты на ранней стадии могут получить от этого пользу, значительной же доле пациентов диагноз ставится лишь на поздних стадиях заболевания. В химиотерапии колоректального рака на поздних стадиях стандартом лечения являются лечебные схемы, основанные на фторурациле. Большинство подобных схем лечения - это т.н. протоколы FOLFOX (инфузия S-FU/лейковорин плюс оксалиплатин) иFOLFIRI (иринотекан, лейковорин, болюсное и длительная инфузия 5-FU). Внедрение цитотоксических препаратов третьего поколения, таких как иринотекан и оксалиплатин,увеличило надежду на значительное улучшение эффективности, однако прогнозы до сих пор относительно плохие. Срок выживаемости при метастатической болезни, как правило, составляет приблизительно 20 месяцев, в результате чего беспомощность медицины при данном заболевании остается высокой. Недавно стало доступным новое поколение медикаментов, действующих на молекулярные мишени,таких как Avastin (бевацизумаб) и Erbitux (цетуксимаб), а около 40 соединений находятся на поздней стадии клинических исследований для лечения различных стадий колоректального рака. Комбинации из нескольких таких соединений повышают число потенциальных возможностей лечения, ожидаемых в будущем. Подавляющее большинство веществ находятся на 2-й фазе исследований, при этом данные соединения чаще, чем другие разрабатываемые медикаменты против колоректального рака, адресованы против EGFR (рецептор эпидермального фактора роста), что связано с тем фактом, что у 80% пациентов с колоректальным раком наблюдается повышенная экспрессия EGFR. На данный момент проводятся клинические исследования с участием пациентов II стадии с комбинацией химиотерапии с одобренными недавно моноклональными антителами (mAbs) (цетуксимаб + иринотекан или FOLFOX4; бевацизумаб в качестве монотерапии или в комбинации с FOLFOX4). Для получения статистически достоверных результатов по данным исследованиям необходимы трех-четырехгодичные периоды наблюдения. Моноклональные антитела (mAb), используемые в настоящее время в онкологии, в целом имеют превосходные шансы на то, что они не будут создавать помех активной иммунотерапии. Действительно, имеются доклинические (GABRILOVICH 1999) и клинические данные, предполагающие, что элиминация фактора роста эндотелия сосудов VEGF (в случае бевацизумаба) вносит положительный вклад в опосредованную ДК (дендритные клетки) активацию Т-клеток (Osada T., Chong G.,Tansik R., Hong T., Spector N., Kumar R., Hurwitz H.I., Dev I., Nixon A.B., Lyerly H.K., Clay T., Morse M.A.The effect of anti-VEGF therapy on immature myeloid cell and dendritic cells in cancer patients. Cancer Immunol Immunother. 2008 Jan 10.). Рак предстательной железы и другие опухоли. Число смертей от рака предстательной железы в 2007 году составит 27050, что делает его наиболее частой причиной смерти от рака у мужчин. Несмотря на то что процент смертности среди белого и афроамериканского населения снижается с начала 1990-х годов, до сих пор число мужчинафроамериканцев более чем в два раза превышает число белых мужчин. Рак предстательной железы является наиболее часто диагностируемым видом рака у мужчин. По неизвестным причинам частота заболеваемости значительно выше среди афроамериканцев, чем среди белых мужчин. Частота заболеваемости раком предстательной железы постоянно менялась на протяжении последних 20 лет: быстрый рост с 1988 по 1992 г., резкое снижение с 1992 по 1995 г. и умеренный рост с 1995 года. Данные тенденции отражают большей частью увеличение числа обследований на рак предстательной железы с помощью анализа крови на простатоспецифический антиген (PSA). Умеренный рост частоты заболеваемости за последнее десятилетие связан, скорее всего, с распространенным скринингом PSA среди мужчин моложе 65 лет. Частота заболеваемости раком предстательной железы стабилизируется у мужчин 65 лет и старше. Число случаев достигло своего пика для белых мужчин в 1992 г. (237,6 случаев на 100000 человек) и для афроамериканцев - в 1993 г. (342,8 случаев на 100000 человек). Лечение рака предстательной железы может включать внимательное наблюдение, операцию, лучевую терапию, высокоинтенсивный фокусированный ультразвук (HIFU), химиотерапию, криохирургию,гормональное лечение или какую-либо их комбинацию. Какая из возможностей является наилучшей зависит от стадии заболевания, оценки по шкале Глисона и уровня PSA. Другие важные факторы - это возраст мужчины, общее состояние его здоровья и его отношение к возможным способам лечения и побочным эффектам от них. Так как все способы лечения могут иметь побочные эффекты, такие как эректильная дисфункция и недержание мочи, то в центре внимания при дискуссиях о способе лечения часто стоит возможность нахождения баланса между целью лечения и рисками, связанными с изменением стиля жизни. Если рак распространился за пределы простаты, то возможности для лечения значительно изменяются, так что большинство врачей, лечащих рак предстательной железы, используют различные номограммы для предсказания вероятности распространения. Лечение с использованием бдительного наблюдения, высокоинтенсивного фокусированного ультразвука (HIFU), лучевой терапии, криохирургии и операции предлагаются обычно мужчинам, у которых рак остается в пределах предстательной железы. Гормональная терапия и химиотерапия часто предусматриваются для лечения заболевания, которое распространилось за пределы простаты. Однако возможны исключения: лучевая терапия может использоваться для некоторых опухолей на поздних стадиях, а гормональная терапия - для опухолей на ранней стадии. Криотерапия, гормональная терапия и химиотерапия также могут быть предложены, если первоначальное лечение было неудачным и рак прогрессирует. У значительного числа пациентов с раком предстательной железы, которые подвергаются простатэктомии из-за клинического подозрения на рост новообразования в пределах органа, окончательное гистологическое обследование операционного материала показывает локально интенсивное распространение опухоли за пределы органа. Для этих пациентов существует высокий риск быстрого локального рецидива, обнаруживаемого обычно как повышение уровня PSA с точки зрения биохимического рецидивирования заболевания. Терапевтические возможности в данной ситуации включают наружную лучевую терапию и гормональную абляцию; однако значимость данных терапевтических подходов, в особенности в отношении продления жизни пациента в долгосрочной перспективе, не подтверждена. Кроме того, необходимо учитывать возможность связанных с лечением осложнений, таких как появление стриктуры уретры (лучевая терапия), потеря полового влечения и импотенция, риск снижения содержания солей кальция в костях в связи с остеопорозом и существенно возросший риск патологической хрупкости костей (гормональная абляция). Более 90% всех случаев заболевания раком предстательной железы обнаруживаются на локальной и региональной стадиях; 5-летняя относительная выживаемость для пациентов, опухоли которых были диагностированы на этих стадиях, достигает 100%. В течение последних 25 лет 5-летняя выживаемость для всех стадий в целом увеличилась с 69 до примерно 90%. В соответствии с последними данными относительная 10-летняя выживаемость составляет 93%, а 15-летняя - 77%. Разительное увеличение выживаемости, в особенности 5-летней, отчасти связано с ранней постановкой диагноза и улучшениями в лечении. Несмотря на это сроки выживаемости значительно сокращаются после распространения на другие ткани и органы. Рак легких. В 2007 году ожидаются приблизительно 210000 новых случаев в США, что составляет около 15% всех диагнозов рака. Частота заболеваемости значительно снижается среди мужчин, со 102 случаев на 100000 человек в 1984 г. до 78,5 в 2003 г. Среди женщин частота заболеваемости стабилизировалась после долгого периода роста. В целях лечения рак легких классифицируется клинически на мелкоклеточный (13%) или немелкоклеточный (87%).-3 023013 Рак легких является наиболее частой причиной смертности от рака как среди мужчин, так и среди женщин. Около 160390 смертей, составляющих приблизительно 29% всех летальных исходов от рака,ожидаются в 2007 году. Начиная с 1987 г., от рака легких ежегодно умирало больше женщин, чем от рака молочной железы. Число смертей среди мужчин значительно снижалось в период с 1991 по 2003 гг.,примерно на 1,9% ежегодно. Смертность от рака легких среди женщин сохраняется на неизменном уровне после продолжительного роста на протяжении нескольких десятилетий. Данные тенденции по смертности от рака легких отражают снижение числа курящих на протяжении последних 30 лет. Вид лечения определяется типом (мелкоклеточный или немелкоклеточный) и стадией ракового заболевания и включают хирургическое вмешательство, лучевую терапию, химиотерапию, а также целенаправленные биологические терапии, такие как бевацизумаб (Avastin) и эрлотиниб (Tarceva). Для локализованного рака в качестве терапии обычно выбирается операция. Последние исследования указывают на то, что выживаемость с немелкоклеточным раком легких ранней стадии улучшается, если за операцией следует химиотерапия. Так как на момент своего обнаружения заболевание обычно распространилось, часто используются лучевая терапия и химиотерапия, иногда в сочетании с операцией. Химиотерапия сама по себе или в сочетании с лучевой терапией является стандартным лечением, выбираемым для мелкоклеточного рака легких; при такой схеме лечения большой процент пациентов испытывает ремиссию, которая в некоторых случаях бывает продолжительной. Одногодичная относительная выживаемость для рака легких слегка возросла с 37% в 1975-1979 гг. до 42% в 2002 г., во многом благодаря усовершенствованиям в хирургической технике и комбинированным способам лечения. Однако 5-летний срок выживаемости для всех стадий в целом составил лишь 16%. Процент выживаемости для всех случаев, обнаруживаемых, когда заболевание все еще локализовано, составляет 49%; однако только 16% всех случаев рака легких диагностируются на этой ранней стадии. Таким образом, существует потребность в новом, эффективном и безопасном способе лечения глиобластомы, опухоли предстательной железы, рака молочной железы, рака пищевода, рака толстого кишечника, светлоклеточной почечно-клеточной карциномы, рака легких, ЦНС, яичника, меланомы, рака поджелудочной железы, плоскоклеточной карциномы, лейкемии и медуллобластомы, а также других видов опухолей, демонстрирующих гиперэкспрессию сурвивина и/или других белков по настоящему изобретению, который улучшал бы самочувствие пациентов без применения химиотерапевтических средств или других веществ, которые могут вызывать серьезные побочные эффекты. Краткое изложение сущности изобретения В первом своем аспекте настоящее изобретение относится к пептиду, индуцирующему противоопухолевый иммунный ответ, где указанный пептид имеет общую длину между 8 и 16 аминокислот и включает последовательность, которая выбирается из группы SEQ ID No. 1-9, 11-13, 15-24 и 26-30, где указанный пептид связывается с молекулой главного комплекса гистосовместимости человека (МНС) классаI или II, и где указанный пептид способен стимулировать Т-клетки CD4 и/или CD8. Во втором своем аспекте настоящее изобретение относится к нуклеиновой кислоте, кодирующей пептид в соответствии с настоящим изобретением, или вектору экспрессии, способному экспрессировать указанную нуклеиновую кислоту. В третьем своем аспекте настоящее изобретение относится к клетке-хозяину, включающей нуклеиновую кислоту или вектор экспрессии в соответствии с настоящим изобретением, где указанная клеткахозяин предпочтительно является антигенпрезентирующей клеткой, в частности, дендритной клеткой или антигенпрезентирующей клеткой. В четвертом своем аспекте настоящее изобретение относится к способу получения активированных цитотоксических Т-лимфоцитов (ЦТЛ) in vitro, включающему контактирование ЦТЛ in vitro с нагруженными антигеном молекулами человека МНС I класса, экспрессированными на поверхности подходящей антигенпрезентирующей клетки или искусственной конструкции, имитирующей антигенпрезентирующую клетку, на период времени, достаточного для активации указанных ЦТЛ антиген-специфическим образом, где указанный антиген является пептидом в соответствии с настоящим изобретением. В пятом своем аспекте настоящее изобретение относится к применению пептида в соответствии с настоящим изобретением, нуклеиновой кислоты или вектора экспрессии в соответствии с настоящим изобретением, клетки в соответствии с настоящим изобретением или активированного цитотоксического Т-лимфоцита, полученного в соответствии с настоящим изобретением для лечения рака или для изготовления медикамента против рака, где указанный медикамент предпочтительно является вакциной. Предпочтительно, если указанный рак выбирается из астроцитомы, пилоидной астроцитомы, дисэмбриопластической нейроэпителиальной опухоли, олигодендроглиомы, эпендимомы, мультиформной глиобластомы, смешанных глиом, олигоастроцитом, медуллобластом, ретинобластом, нейробластом, термином,тератом, ганглиоглиом, ганглиоцитом, центральной ганглиоцитомы, примитивных нейроэктодермальных опухолей (PNET, к примеру, медуллобластом, медуллоэпителиом, нейробластом, ретинобластом, эпендимобластом), опухолей паренхимы шишковидной железы (к примеру, пинеоцитом, пинеобластом),опухолей, развивающихся из эпендимных клеток, опухолей хороидного сплетения, нейроэпителиальных опухолей неопределенного происхождения (к примеру, глиоматоз головного мозга, астробластом), гли-4 023013 областом, рака предстательной железы, рака молочной железы, рака пищевода, рака толстого кишечника,почечно-клеточной карциномы, светлоклеточной почечно-клеточной карциномы, рака легких, ЦНС,яичника, меланомы, рака поджелудочной железы, плоскоклеточной карциномы, лейкемии и медуллобластомы, а также других видов опухолей или рака, демонстрирующих гиперэкспрессию сурвивина и/или других белков настоящего изобретения. В шестом своем аспекте настоящее изобретение относится к набору для индукции противоопухолевого иммунного ответа, включающему: (а) контейнер, который содержит фармацевтическую композицию, содержащую заявленный пептид, заявленную нуклеиновую кислоту, заявленный вектор, заявленную клетку, заявленный активированный цитотоксический Т-лимфоцит или заявленное антитело в буферном растворе или в лиофилизованной форме; и (b) второй контейнер, содержащий буферный разбавитель или восстанавливающий раствор для лиофилизованного состава. При необходимости, набор дополнительно включает инструкции по использованию раствора или восстановлению и использованию лиофилизованного состава, фильтр, иглу или шприц. В седьмом своем аспекте настоящее изобретение относится к антителу, которое специфически связывается с главным комплексом гистосовместимости человека (МНС) класса I или II в комплексе с рестриктированным по HLA антигеном, где антитело предпочтительно является поликлональным антителом,моноклональным антителом и/или химерным антителом. Краткое описание чертежей Фиг. 1 демонстрирует масс-спектры ESI-жидкостной хроматографии, идентифицирующие опухолеассоциированные пептиды (TUMAPs) IGF2BP3-001 из пробы глиобластомы GB6010, которые были презентированы рестриктированным по МНС класса I способом. Вкратце, А) цикл сканирования начинался с полного сканирования с высокой точностью масс на спектрометре Orbitrap (R = 30000), за чем следовало В) сканирование MS/MS на Orbitrap (R = 7500) на 5 особенно многочисленных ионахпредшественниках с динамическим исключением отобранных ранее ионов. Идентифицированную последовательность пептида подтверждали сравнением генерированного фрагментационного образца естественного пептида С) с фрагментационным образцом синтетического контрольного пептида с идентичной последовательностью D); на фиг. 2 показан профиль экспрессии мРНК генов-мишеней по изобретению, которые в высокой степени гиперэкспрессированы в пробах глиобластомы. Экспрессия этих генов отсутствует или находится на очень низком уровне в нормальных тканях, в то время как она сильно повышена в пробах глиобластомы. Относительная экспрессия мРНК, измеренная с помощью генных чипов, показана для нескольких нормальных тканей и отдельных проб мультиформной глиобластомы (GBM). Значения являются относительными по отношению к уровням экспрессии на нормальной почке (значение произвольно приравнивается к 1,0). Значения для нормальных тканей были установлены с помощью имеющихся в продаже пулов мРНК. Буквы в скобках обозначают "наличие сигнала", как указано в программе анализа. "Наличие сигнала" обозначает, был ли вообще в пробе специфически детектирован транскрипт, или же не могло наблюдаться значимой детекции. Оно может принимать значения "Р" (присутствует), "А" (отсутствует) или "М" (детектируемо в незначительной степени); фиг. 2 а(а) демонстрирует профиль экспрессии FABP7, фиг. 2 а(b) демонстрирует профиль экспрессии NR2E1, фиг. 2b(а) демонстрирует профиль экспрессии SLCO1C1, и фиг. 2b(b) демонстрирует профиль экспрессии IGF2BP3. Перевод английских терминов, употребляющихся на фиг. 2 а - 2b, представлен после фиг. 2b(b); на фиг. 3 показан тетрамерный анализ стимулированной микросферой пролиферации CSP-001 иNLGN4X-001-специфических CD8+ лимфоцитов из периферической крови здорового донора. ПК, обогащенные на лунку 1106 CD8+, стимулировали еженедельно микросферами, связанными с анти-CD28 плюс опухолевый антиген высокой плотности A0201/CSP-001 (левая секция) или анти-CD28 плюс опухолевый антиген высокой плотности A0201/NLGN4X-001 (правая секция). После трех стимуляций invitro клетки были окрашены антителом CD8 FITC и тетрамерами с флуоресцентными меткамиA0201/CSP-001 и А 0201/NLGN4X-001. Клетки высаживают на лимфоциты CD8+; цифрами представлена процентная доля клеток в обозначенном квадранте среди лимфоцитов CD8+; на фиг. 4 показана аффинность пептидов HLA I класса по изобретению к молекуле МНС, закодированной аллелем HLA-A0201. Константы диссоциации (KD) пептидов TUMAP HLA класса I из изобретения и контрольного пептида HBV-001 (сильная связывающая сила А 02) были измерены основанным на методике ELISA анализом по рефолдингу МНС (см. примеры). Детальное описание изобретения Все термины, используемые здесь, если не указано иное, имеют значения, данные ниже. Термин "пептид" используется здесь для обозначения серий аминокислотных остатков, связанных друг с другом типично пептидными связями между альфа-аминными и карбонильными группами смежных аминокислот. Пептиды типично имеют длину 9 аминокислот, но могут быть короче - длиной 8 аминокислот и длиннее - 16 или 10, 11, 12, 13, 14 или 15 аминокислот. Термин "олигопептид" используется здесь для обозначения серий аминокислотных остатков, связанных один с другим типично пептидными связями между альфа-аминными и карбонильными группа-5 023013 ми смежных аминокислот. Длина олигопептида не особенно важна для изобретения до тех пор, пока в нм сохраняются надлежащие эпитоп или эпитопы. Олигопептиды типично бывают менее чем около 30 аминокислотных остатков в длину и более чем около 14 аминокислот в длину. Термин "полипептид" обозначает серии аминокислотных остатков, связанных один с другим типично пептидными связями между альфа-аминными и карбонильными группами смежных аминокислот. Длина полипептида не особенно важна для изобретения до тех пор, пока сохраняются надлежащие эпитопы. В отличие от терминов "пептид" или "олигопептид", термин "полипептид" введн для обозначения молекул, содержащих более приблизительно 30 аминокислотных остатков. Пептид, олигопептид, белок или полинуклеотид, кодирующий для такой молекулы, является "иммуногенным" (и, таким образом, "иммуногеном" в рамках настоящего изобретения), если он способен индуцировать иммунный ответ. В случае настоящего изобретения иммуногенность получает более специфическое определение как способность индуцировать Т-клеточный ответ. Таким образом, "иммуноген" будет представлять собой молекулу, которая способна индуцировать иммунный ответ, и, в случае настоящего изобретения, молекулу, способную индуцировать ответ Т-клетки. Для Т-клеточного "эпитопа" необходим короткий пептид, который связан с рецептором МНС I илиII класса, образующий трехчленный комплекс (альфа-цепь МНС класса I, бета-2-микроглобулин и пептид), который может быть распознан Т-клеткой, несущей подходящий Т-клеточный рецептор, связывающийся с комплексом МНС/пептид с подходящей аффинностью. Пептиды, связывающиеся с молекулами МНС I класса, типично имеют длину в 8-14 аминокислот и, особенно типично, длину в 9 аминокислот. Т-клеточные эпитопы, которые связываются с молекулами МНС II класса, типично имеют длину в 12-30 аминокислот. В случае пептидов, которые связываются с молекулами МНС II класса, один и тот же пептид и соответствующий Т-клеточный эпитоп могут иметь общий центральный сегмент, но различаться по общей длине из-за примыкающих последовательностей с различными длинами, расположенными перед аминным концом центральной последовательности и после ее карбоксильного конца соответственно. Рецепторы МНС II класса имеют более открытую структуру, пептиды, связанные с рецепторами МНС II класса, соответствующим образом, не полностью углублены в структуру пептидсвязывающей бороздки молекулы МНС II класса, как то имеет место быть с пептидсвязывающей бороздкой молекулы МНС I класса. Как ни удивительно, это не применимо к пептиду в соответствии с SEQ ID1, так как небольшие изменения в длине пептида ведут к экстремальному снижению активности (см. ниже). У человека имеется три различных генетических локуса, которые кодируют для молекул МНС I класса (молекулы МНС человека называются также человеческими лейкоцитарными антигенами (HLA:HLA-A, HLA-B и HLA-C. HLA-A01, HLA-A02 и HLA-A11 являются примерами различных аллелей МНС I класса, которые могут экспрессироваться этими локусами. Человеческий геном имеет 3 различных локуса для генов МНС II класса: HLA-DR, HLA-DQ и HLADP. Рецепторы МНС II класса являются гетеродимерами, состоящими из альфа- и бета-цепи, которые обе закрепляются в клеточной мембране с помощью трансмембранного участка. HLA-DRB104 и HLADRB107 - это два примера различных бета-аллелей МНС II класса, о которых известно, что они кодируются в данных локусах. Аллели II класса сильно полиморфны, к примеру, было описано несколько сотен различных аллелей HLA-DRB1. Поэтому для терапевтических и диагностических целей крайне желателен пептид, который связывается с подходящей аффинностью с несколькими различными рецепторами HLA II класса. Пептид, связывающийся с несколькими различными молекулами HLA II класса,называется беспорядочно связывающимся пептидом. Используемая здесь ссылка на последовательность ДНК включает как однонитевую, так и двунитевую ДНК. Таким образом, специфическая последовательность, если в контексте не указано иное, относится к однонитевой ДНК такой последовательности, дуплексу такой последовательности с его комплементом (двунитевая ДНК) и комплементу такой последовательности. Термин "кодирующая область" относится к тому участку гена, который естественно или обычно кодирует для продукта экспрессии того гена в его естественном геномном окружении, т.е. участку, кодирующему in vivo для нативного продукта экспрессии гена. Кодирующая область может быть из нормального, мутировавшего или измененного гена или может даже быть из последовательности ДНК, или же гена, целиком синтезированного в лаборатории с использованием методов, хорошо известных специалистам из области синтеза ДНК. Термин "нуклеотидная последовательность" относится к гетерополимеру дезоксирибонуклеотидов. Нуклеотидная последовательность, кодирующая для конкретного пептида, олигопептида или полипептида, может быть встречающейся в природе или может быть получена синтетически. В целом, фрагменты ДНК, кодирующие пептиды, полипептиды и белки данного изобретения, собраны из фрагментов кДНК и коротких олигонуклеотидных линкеров или же из серий олигонуклеотидов для обеспечения синтетического гена, который способен экспрессироваться в рекомбинантной транскрипционной единице,включающей регуляторные элементы, образованные из микробиального или вирусного оперона. Термин "продукт экспрессии" означает полипептид или белок, являющийся естественным продуктом трансляции гена и кодирующих эквивалентов любой последовательности нуклеиновой кислоты, образующихся в результате вырожденности генетического кода и, таким образом, кодирующих для той/тех-6 023013 же самой(-ых) нуклеиновой(-ых) кислот(-ы). Термин "фрагмент", если относится к кодирующей последовательности, означает участок ДНК,включающий меньше, чем полную кодирующую область, продукт экспрессии которого обязательно сохраняет ту же самую биологическую функцию или активность, что и продукт экспрессии целой кодирующей области. Термин "сегмент ДНК" относится к полимеру ДНК в виде отдельного сегмента или в качестве компонента более крупной конструкции ДНК, которая была образована из ДНК, изолированной по крайней мере один раз в существенно чистой форме, т.е. без контаминирующих эндогенных материалов и в количестве или с концентрацией, позволяющей идентификацию, манипуляцию и получение сегмента и его составных нуклеотидных последовательностей стандартными биохимическими методами, например с использованием вектора для клонирования. Такие сегменты обеспечиваются в форме открытой рамки считывания, не прерываемой внутренними не-транслированными последовательностями или нитронами,которые типично присутствуют в эукариотических генах. Последовательности нетранслированной ДНК могут быть представлены по ходу транскрипции из открытой рамки считывания, где она не интерферирует с манипуляцией или экспрессией кодирующих областей. Термин "праймер" означает короткую последовательность нуклеиновой кислоты, которая может быть спарена с одной нитью ДНК и обеспечивает свободный конец 3'ОН, на котором ДНК-полимераза начинает синтезировать дезоксирибонуклеотидную цепь. Термин "промотор" означает участок ДНК, задействованный в связывании РНК-полимеразы для инициации транскрипции. Термин "открытая рамка считывания (ОРС)" означает серии триплетов, кодирующих для аминокислот без каких-либо терминирующих кодонов, и является последовательностью (потенциально), способной транслироваться в белок. Понятие "изолированный" означает, что материал удален из его исходного окружения (к примеру,естественное окружение, если он встречается в природе). Например, встречающийся в природе полинуклеотид или полипептид, представленный в живых организмах, не является изолированным, но тот же самый полинуклеотид или полипептид, выделенный из некоторых или всех сосуществующих материалов природной системы, является изолированным. Такие полинуклеотиды могли быть частью вектора и/или такие полинуклеотиды или полипептиды могли быть частью композиции и все-таки могли быть изолированы, так что такой вектор или композиция не является частью своего естественного окружения. Полинуклеотиды и рекомбинантные или иммуногенные полипептиды, раскрытые в соответствии с настоящим изобретением, могут также быть в "очищенной" форме. Термин "очищенный" не требует абсолютной чистоты; скорее он предназначен для дачи относительного определения и может включать препараты с высокой очисткой или препараты только с частичной очисткой, в соответствии с тем, как эти термины понимаются специалистами в соответствующей области. Например, отдельные клоны, изолированные из библиотеки кДНК, обычно очищались до электрофоретической чистоты. Очистка исходного материала или природного материала до по крайней мере одного порядка величины, предпочтительно двух или трех порядков и более предпочтительно четырех или пяти порядков величины определенно рассматривается. Более того, определенно рассматривается заявленный полипептид, чистота которого составляет предпочтительным образом 99,999 или по крайней мере 99,99 или 99,9 и еще более желательно 99 мас.% или более. Нуклеиновые кислоты и продукты экспрессии полипептида, раскрываемые в соответствии с настоящим изобретением, в равной степени как и векторы экспрессии, содержащие такие нуклеиновые кислоты и/или такие полипептиды, могут быть в "обогащенной форме". Используемый здесь термин "обогащенный" означает, что концентрация материала по крайней мере в приблизительно 2, 5, 10, 100 или 1000 раз выше его естественной концентрации (например), преимущественно 0,01, предпочтительно по крайней мере около 0,1 мас.%. Рассматриваются также препараты с около 0,5, 1, 5, 10 и 20 мас.%. Последовательности, конструкции, векторы, клоны и другие материалы, включенные в настоящее изобретение,могут предпочтительно быть в обогащенной или изолированной форме. Термин "активный фрагмент" означает фрагмент, который дает иммунный ответ (т.е. обладает иммуногенной активностью), если он введен отдельно или опционально с подходящим адъювантом животному, такому как млекопитающее, например кролику или мыши, также включая человека; таковой иммунный ответ, принимающий форму стимуляции ответа Т-клетки, у животного-реципиента, такого как человек. Альтернативно "активный фрагмент" может также быть использован для инициации ответа Тклетки in vitro. Используемые здесь термины "участок", "сегмент" и "фрагмент", если они использованы по отношению к полипептидам, относятся к непрерывной последовательности остатков, таких как аминокислотные остатки, последовательность которых формирует подкласс более крупной последовательности. Например, если полипептид был подвергнут обработке любой из известных эндопептидаз, таких как трипсин или химотрипсин, то полученные в результате такой обработки олигопептиды будут представлять участки, сегменты или фрагменты исходного полипептида. Это означает, что любой таковой фрагмент будет обязательно содержать как часть его аминокислотной последовательности сегмент, фрагмент илиSEQ ID1 по 30, которая соответствует встречающимся в природе или "материнским" белкам последовательностей с SEQ ID1 по 30. При использовании по отношению к полинуклеотидам такие термины относятся к продуктам, полученным при обработке указанных полинуклеотидов любой из известных эндонуклеаз. В соответствии с настоящим изобретением термин "процентная доля идентичности" или "идентичный с процентной долей", если он относится к последовательности, означает, что последовательность сравнивается с заявленной или описанной последовательностью после противопоставления сравниваемой последовательности ("Сравниваемая последовательность") описанной или заявленной последовательностью ("Контрольная последовательность"). Процентная доля идентичности определяется затем по следующей формуле: Процентная доля идентичности = 100 [I -(C/R)],где С является числом различий между Контрольной последовательностью и Сравниваемой последовательностью по длине противопоставления между Контрольной последовательностью и Сравниваемой последовательностью, где(i) каждое основание или аминокислота в Контрольной последовательности, которые не имеют соответствующего противопоставленного основания или аминокислоты в Сравниваемой последовательности, и(iii) каждое противопоставленное основание или аминокислота в Контрольной последовательности,которые отличаются от противопоставленного основания или аминокислоты в Сравниваемой последовательности, представляют собой различие; иR - это число оснований или аминокислот в Контрольной последовательности по длине противопоставления Сравниваемой последовательности с любой брешью, образующейся в Контрольной последовательности, считающейся также за основание или аминокислоту. Если существует противопоставление между Сравниваемой последовательностью и Контрольной последовательностью, для которых процентная доля идентичности по расчетам приблизительно равна или выше установленной минимальной Процентной доли идентичности, тогда Сравниваемая последовательность имеет установленную минимальную процентную долю идентичности с Контрольной последовательностью, если даже могут существовать противопоставления, в которых вышеупомянутая расчетная Процентная доля идентичности меньше, чем установленная Процентная доля идентичности. Исходные пептиды, раскрываемые здесь, могут быть модифицированы путм замещения одного или нескольких остатков в различных, по возможности отобранных, местах пептидной цепи, если не заявлено иное. Такие замещения могут носить консервативный характер, например, где одна аминокислота заменяется аминокислотой с похожей структурой и характеристиками, так же как при замене гидрофобной аминокислоты на другую гидрофобную аминокислоту. Ещ более консервативным будет замещение аминокислот одинакового или похожего размера и химического характера, такое как при замене лейцина на изолейцин. В исследованиях вариаций последовательностей внутри семейств встречающихся в природе гомологичных белков определнные замещения аминокислот переносятся гораздо чаще, чем другие, и они часто проявляют взаимосвязь со сходствами по размеру, заряду, полярности и гидрофобности между исходной аминокислотой и е заменой; и таковой является основа определения "консервативных замещений". Консервативные замещения определены здесь как обмены внутри одной из последующих пяти групп: группа 1 - малые, алифатические, неполярные или слабо полярные остатки (Ala, Ser, Thr, Pro,Gly); группа 2 - полярные, отрицательно заряженные остатки и их амиды (Asp, Asn, Glu, Gln); группа 3 полярные, положительно заряженные остатки (His, Arg, Lys); группа 4 - крупные, алифатические, неполярные остатки (Met, Leu, Ile, Val, Cys); и группа 5 - крупные, ароматические остатки (Phe, Tyr, Trp). Менее консервативные замещения могут охватывать замещение одной аминокислоты другой, имеющей похожие характеристики, но отличающейся в какой-то степени по размеру, как в случае замещения аланина остатком изолейцина. Высоко неконсервативные замещения могут охватывать замещение кислой аминокислоты другой, которая имеет полярность, или даже такой, которая имеет щелочной характер. Такие "радикальные" замещения не могут, однако, быть отвергнуты как потенциально неэффективные из-за того, что химические эффекты не полностью предсказуемы, и радикальные замещения могут привести к увеличению случайных эффектов, в противном случае не предсказуемых, исходя из обычных химических принципов. Разумеется, в таких замещениях могут участвовать другие структуры, отличающиеся от обычныхL-аминокислот. Таким образом, D-аминокислоты могут быть замещены L-аминокислотами, обычно встречающимися в антигенных пептидах по изобретению и также охватываемые настоящим раскрытием сущности изобретения. Кроме того, аминокислоты, содержащие нестандартные R-группы (т.е. R-группы,отличающиеся от обнаруженных в распространнных 20 аминокислотах природных белков) могут быть также использованы в целях замещения для получения иммуногена и иммуногенных полипептидов в соответствии с настоящим изобретением.-8 023013 Если были произведены замещения в более чем одной позиции для получения пептида с практически эквивалентной или большей антигенной активностью, чем та, что определена ниже, то комбинации таких замещений будут проанализированы для определения того, приведут ли эти комбинации замещений к дополнительным или синергетическим эффектам по отношению к антигенности пептида. По большей части не более 4 позиций внутри пептида должны замещаться одновременно. Термин "Т-клеточный ответ" означает специфическую пролиферацию и активацию эффекторных функций, индуцированных пептидом in vitro или in vivo. Для ЦТЛ, рестриктированных по МНС класса I,эффекторными функциями может быть лизис клеток-мишеней с введенным импульсным способом пептидом, с введенным импульсным способом предшественником пептида или клеток-мишеней, естественно презентирующих пептид, секреция цитокинов, предпочтительно интерферона-гамма, TNF-альфа или ИЛ-2, индуцированная пептидом, секреция эффекторных молекул, предпочтительно гранзимов или перфоринов, индуцированная пептидом, или дегрануляция. Для Т-хелперных клеток, рестриктированных по МНС класса II, эффекторными функциями может быть индуцированная пептидом секреция цитокинов,предпочтительно, IFN-гамма, TNF-альфа, ИЛ-4, ИЛ-5, ИЛ-10 или ИЛ-2, или индуцированная пептидом дегрануляция. Возможные эффекторные функции ЦТЛ и Т-хелперных клеток не ограничиваются данным списком. Предпочтительно, чтобы ЦТЛ, специфичные для пептида с SEQ ID1 по 30, были тестированы относительно замещнных пептидов; концентрация пептида, при которой замещнные пептиды достигают половины максимального роста лизиса относительно фона, составляет не более чем около 1 мМ,предпочтительно не более чем около 1 мкМ, более предпочтительно не более чем около 1 нМ и ещ более предпочтительно не более чем около 100 пМ и наиболее предпочтительно не более чем около 10 пМ. Также предпочтительно, чтобы замещнный пептид распознавался ЦТЛ более чем одного индивида, по крайней мере двух и более предпочтительно трх индивидов. Таким образом, эпитопы настоящего изобретения могут быть идентичны встречающимся в природе опухолеассоциированным или опухолеспецифическим эпитопам или могут включать эпитопы, отличающиеся не более чем 4 остатками от контрольного пептида, при условии, что они имеют, по существу,идентичную антигенную активность. Стимуляция иммунных ответов находится под воздействием присутствия антигенов, распознаваемых иммунной системой хозяина как чужеродные. Открытие существования опухолеассоциированных антигенов улучшило возможности для использования иммунной системы хозяина для способствования иммунному ответу, специфическому для антигенов-мишеней, экспрессированных на поверхности опухолевых клеток, который благодаря данному механизму действия способен вызывать регрессию, задержку или замедление роста опухоли. Различные механизмы объединения обеих ветвей иммунной системы, как гуморальной, так и клеточной, исследуются в настоящее время для иммунотерапии рака. Специфические элементы клеточных иммунных ответов способны к специфическому распознаванию и уничтожению опухолевых клеток. Изоляция цитотоксических Т-клеток (ЦТЛ) из популяций опухоль-инфильтрирующих клеточных популяций или из периферической крови предполагает, что такие клетки играют важную роль в естественной иммунной защите против рака (Cheever et al., 1993; Zeh, III etal., 1999). Основываясь на 415 тканевых препаратах пациентов, страдающих от колоректального рака,Galon и соавторы были в состоянии продемонстрировать, что вид, плотность и местонахождение иммунных клеток в опухолевой ткани являются действительно лучшим прогностическим фактором выживаемости пациентов, чем широко используемая классификация опухолей по TNM (Galon et al., 2006). МНС класса I презентируют пептиды, образующиеся из протеолитического расщепления преимущественно эндогенных белков, DRIPs и более крупных пептидов. Молекулы МНС II класса могут встречаться преимущественно на профессиональных антигенпрезентирующих клетках (АПК) и, в первую очередь, презентировать пептиды экзогенных или трансмембранных белков, которые поглощаются АПК в период эндоцитоза и впоследствии процессируются (Cresswell, 1994). Комплексы из пептида и молекул МНС класса I распознаются CD8-положительными цитотоксическими Т-лимфоцитами, несущими подходящий ТКР (Т-клеточный рецептор), а комплексы из пептида и молекул МНС класса II распознаютсяCD4-положительными хелперными Т-клетками, несущими подходящий ТКР. Хорошо известно, что ТКР,пептид и МНС часто встречаются в стехиометрическом соотношении 1:1:1.CD4-положительные хелперные Т-клетки играют важную роль в индуцировании и поддержании эффективных ответов CD8-положительных цитотоксических Т-клеток (Wang and Livingstone, 2003; Sunand Bevan, 2003; Shedlock and Shen, 2003). Первоначально прайминг и экспансия ЦТЛ в лимфатических узлах поддерживается CD4+ Т-клетками (Schoenberger et al., 1998). Один механизм поэтому может служить руководством для наивных CD8+ клеток к месту функционального взаимодействия CD4+ Т-клеток- АПК (Castellino et al., 2006). В итоге, образование функциональных клеток памяти CD8+ в большинстве случаев зависит от поддержки CD4+ Т-клеток (Sun and Bevan, 2003; Janssen et al., 2003). По этим причинам идентификация CD4-положительных Т-клеточных эпитопов, получаемых из опухолеассоциированных антигенов (ТАА), может быть чрезвычайно важна для разработки фармацевтических препаратов для инициации противоопухолевых иммунных ответов (Kobayashi et al., 2002; Qin et al., 2003; Gnjatic et al.,2003). В месте опухоли Т-хелперные клетки поддерживают благоприятное для ЦТЛ цитокиновое окру-9 023013 жение (Qin and Blankenstein, 2000; Mortara et al., 2006) и привлекают эффекторные клетки, к примеру,ЦТЛ, естественные киллерные клетки (NK), макрофаги, гранулоциты (Marzo et al., 2000; Hwang et al.,2007). При отсутствии воспаления экспрессия молекул МНС II класса преимущественно рестриктирована по клеткам иммунной системы, в особенности профессиональным антигенпрезентирующим клеткам(АПК), например моноцитам, образованным из моноцитов клеткам, макрофагам, дендритным клеткам. Неожиданно было обнаружено, что клетки опухоли раковых пациентов экспрессируют молекулы МНС класса II (Dengjel et al., 2006). На моделях млекопитающих животных, например мышах, было показано, что даже при отсутствии эффекторных клеток (т.е. CD8-положительных Т-лимфоцитов), CD4-положительных Т-клеток достаточно для ингибирующего проявления опухолей посредством ингибирования ангиогенеза при секреции интерферон-гамма (IFN) (Qin and Blankenstein, 2000). Также предлагалось прямое уничтожение опухолевых клеток цитотоксическими CD4+ Т-клетками посредством лимфотоксинов и гранзимов В (Penna et al.,1992; Littaua et al., 1992). К тому же было показано, что CD4-положительные Т-клетки, распознающие пептиды из опухолеассоциированных антигенов, презентированные молекулами HLA класса II, могут препятствовать опухолевой прогрессии посредством индукции ответов антител (Ab) (Kennedy et al., 2003). В отличие от опухолеассоциированных пептидов, связывающихся с молекулами HLA класса I, до сих пор было описано лишь небольшое число лигандов класса II опухолеассоциированных антигенов(ТАА). Так как конститутивная экспрессия молекул HLA класса II обычно ограничена клетками иммунной системы (Mach et al., 1996), то возможность изоляции пептидов класса II напрямую из первичных опухолей считалась невозможной. Тем не менее Dengjel с соавторами недавно удалось идентифицировать ряд эпитопов МНС класса II напрямую из опухолей (WO 2007/028574, ЕР 1760088 B1; (Dengjel et al., 2006). Антигены, которые распознаются опухолевыми специфическими цитотоксическими Тлимфоцитами, т.е. их эпитопами, могут быть молекулами, образованными из любого класса белков, таких как ферменты, рецепторы, факторы транскрипции и т.д., которые экспрессируются и, по сравнению с не измененными клетками того же происхождения, находятся в повышенном количестве в клетках соответствующей опухоли. Актуальная классификация опухолеассоциированных антигенов (ТАА) включает следующие основные группы (Novellino et al., 2005). 1. Раковые антигены семенника: первые в истории идентифицированные ТАА, которые могут распознаваться Т-клетками (van der Bruggen et al., 1991), принадлежат к этому классу, называвшемуся первоначально "антиген ракового семенника" (СТ), так как его члены экспрессируются в отличных по гистологической структуре опухолях человека, а среди нормальных тканей - только в сперматоцитах/сперматогониях семенника и изредка в плаценте. Так как клетки семенника не экспрессируют молекулы HLA I и II класса, то эти антигены не могут быть распознаны Т-клетками в нормальных тканях и поэтому могут рассматриваться как иммунологически опухолеспецифические. Хорошо известными примерами антигенов СТ являются члены семейства MAGE или NY-ESO-1. 2. Дифференциационные антигены: данные ТАА встречаются в опухолевых и нормальных тканях,из которых образуется опухоль; большинство из них имеется в меланомах и нормальных меланоцитах. Многие из этих линиеспецифических белков меланоцитов участвуют в биосинтезе меланина и поэтому не являются опухолеспецифическими, однако, несмотря на это, они широко применяются в противораковой терапии. Примеры включают, но не ограничиваются, тирозиназой и Melan-A/MART-1 для меланомы или PSA для рака простаты. 3. Гиперэкспрессированные ТАА: гены, кодирующие широко экспрессированные ТАА, были обнаружены в отличных по гистологической структуре опухолях, а также во многих нормальных тканях, в основном, с более низким уровнем экспрессии. Возможно, что многие эпитопы, процессируемые и потенциально презентируемые нормальными тканями, находятся ниже порогового уровня для распознавания Т-клетками, в то время как их гиперэкспрессия в опухолевых клетках может инициировать противораковый ответ, нарушая установившуюся ранее толерантность. Известными примерами ТАА этого класса являются Her-2/neu, сурвивин, теломераза или WT1. 4. Опухолеспецифические антигены: данные уникальные ТАА образуются из мутаций нормальных генов (таких как -катенин, CDK4 и т.д.). Некоторые из этих молекулярных изменений ассоциированы с неопластической трансформацией и/или прогрессией. Опухолеспецифические антигены, в основном,способны индуцировать сильные иммунные ответы, не заключая в себе риска аутоиммунных реакций по отношению к нормальным тканям. С другой стороны, данные ТАА в большинстве случаев релевантны только для определенной опухоли, на которой они были идентифицированы, и обычно не являются общими для многих отдельных опухолей. 5. ТАА, образующиеся в результате аномальных пост-трансляционных модификаций: такие ТАА могут образоваться из белков, которые не являются ни специфическими, ни гиперэкспрессированными в- 10023013 опухолях, однако, несмотря на это, становятся опухолеассоциированными в ходе пост-трансляционных процессов, происходящих первоначально в опухолях. Примеры для этого класса берут свое начало в измененных паттернах гликозилирования, приводящих к новым эпитопам в опухолях, как в случае MUC1,или таких процессах как белковый сплайсинг во время деградации, который может или не может быть опухолеспецифическим (Hanada et al., 2004; Vigneron et al., 2004). 6. Онковирусные белки: данные ТАА являются вирусными белками и могут играть ведущую роль в онкогенном процессе, и, так как они являются чужеродными (не человеческого происхождения), они могут провоцировать Т-клеточный ответ. Примерами таких белков являются вирусные белки человеческой папилломы типа 16, Е 6 и Е 7, которые экспрессированы в карциноме шейки матки. Для того чтобы белки были узнаны цитотоксическими Т-лимфоцитами в качестве опухолеспецифического или -ассоциированного антигена, и в целях применения в терапии должны выполняться особые предварительные требования. Антиген должен экспрессироваться преимущественно опухолевыми клетками, а не здоровыми тканями или в сравнительно малом объеме. В дальнейшем желательно, чтобы соответствующий антиген не только присутствовал в каком-либо виде опухоли, но и также имел высокую концентрацию (т.е. число копий соответствующего пептида на клетку). Опухолеспецифические и опухолеассоциированные антигены часто образованы из белков, напрямую задействованных в трансформации нормальной клетки в опухолевую, в связи с функцией, например, при контроле клеточного цикла или подавлении апоптоза. Кроме того, мишени нисходящего пути белков, напрямую являющихся причиной трансформации, могут быть также представлены в повышенном количестве и, таким образом, косвенно опухолеассоциированными. Такие косвенно опухолеассоциированные антигены могут также быть мишенями вакцинационного подхода (Singh-Jasuja et al., 2004). В обоих случаях необходимо, чтобы эпитопы присутствовали в аминокислотной последовательности антигена, поскольку такой пептид ("иммуногенный пептид"), который образован из опухолеассоциированного антигена, должен вести in vitro или invivo к Т-клеточному ответу. В основном, любой пептид, способный связываться с молекулой МНС, может выполнять функцию Т-клеточного эпитопа. Предварительным условием для индукции Т-клеточного ответа in vitro или in vivo является присутствие Т-клетки с соответствующим ТКР и отсутствие иммунологической толерантности к данному конкретному эпитопу. Поэтому опухолеассоциированные антигены (ТАА) являются отправным пунктом для разработки противораковой вакцины. Методы идентификации и характеристики ТАА основаны на использовании ЦТЛ, которые могут быть изолированы у пациентов или здоровых субъектов, или же они основаны на генерировании дифференциальных транскрипционных профилей или дифференциальных паттернов пептидной экспрессии между опухолевыми и нормальными тканями (Lemmel et al., 2004; Weinschenk et al.,2002). Однако идентификация генов, гиперэкспрессированных в опухолевых тканях или человеческих опухолевых клеточных линиях или же селективно экспрессированных в таких тканях или клеточных линиях, не дает точной информации об использовании антигенов, транскрибированных с данных генов, в иммунотерапии. Это обусловлено тем, что только отдельная субпопуляция эпитопов этих антигенов подходит для такого применения, так как Т-клетка с соответствующим ТКР должна быть в наличии, и необходимо, чтобы отсутствовала или была минимальной иммунологическая толерантность к этому конкретному эпитопу. Поэтому важно выбрать лишь те пептиды из гиперэкспрессированных или селективно экспрессированных белков, которые презентируются в соединении с молекулами МНС, против которых может быть обнаружена функциональная Т-клетка. Такая функциональная Т-клетка определяется как Тклетка, которая при стимуляции специфическим антигеном может быть распространена клонально и способна к выполнению эффекторных функций ("эффекторная Т-клетка"). Т-хелперные клетки играют важную роль в управлении эффекторной функцией ЦТЛ в противоопухолевом иммунитете. Эпитопы Т-хелперных клеток, инициирующие ответы Т-хелперных клеток типа ТН 1, поддерживают эффекторные функции CD8-положительных киллерных Т-клеток, которые включают цитотоксические функции, направленные против опухолевых клеток, проявляющих опухолеассоциированные пептиды/МНС-комплексы на их клеточной поверхности. Таким образом, опухолеассоциированные пептидные эпитопы Т-хелперных клеток, одни или в комбинации с другими опухолеассоциированными пептидами, могут служить в качестве активных фармацевтических ингредиентов вакцинных композиций, которые стимулируют противоопухолевые иммунные ответы. Так как оба вида ответов, зависящие от CD8 и от CD4, вносят свой вклад в противоопухолевый эффект сообща и усиливая друг друга, то идентификация и характеристика опухолеассоциированных антигенов, распознаваемых как CD8+ ЦТЛ (лиганд: молекула МНС I класса + пептидный эпитоп), так и CD4 положительными хелперными Т-клетками (лиганд: молекула МНС II класса + пептидный эпитоп) являются важными при разработке противоопухолевых вакцин. Ввиду серьезных побочных эффектов и расходов, связанных с лечением рака, крайне необходимы улучшенные методы прогнозирования и диагностики. Поэтому существует необходимость в идентификации других факторов, репрезентирующих биомаркеры рака вообще и глиобластомы в частности. Кроме того, существует необходимость в идентификации факторов, которые могут быть использованы при- 11023013 лечении рака вообще и глиобластомы в частности. Кроме того, не существует признанного подхода к лечению пациентов с раком предстательной железы с биохимическим рецидивом после радикальной простатэктомии, обычно вызываемым резидуальным остатком опухоли in situ при наличии локально прогрессирующего роста опухоли. Желательны были бы новые терапевтические подходы, которые бы сопровождались низкой смертностью с адекватной терапевтической эффективностью по сравнению с имеющимися на данный момент терапевтическими подходами. Данным изобретением обеспечиваются пептиды, которые полезны для лечения глиобластомы, рака предстательной железы и других опухолей, которые гиперэкспрессируют сурвивин и/или CSP и/или другие пептиды по изобретению. Как непосредственно показал масс-спектрометрический анализ, эти пептиды естественно презентировались молекулами HLA на пробах первичной глиобластомы человека (см. пример 1 и фиг. 1) или в случае SEQ ID : 26, предсказанной по алгоритму SYFPEITHI (Rammensee etal., 1995), они должны беспорядочно связываться с аллелями HLA-DR, HLA-DRB101, DRB103,DRB104, DRB111 и DRB115. Основываясь на этих данных и частоте этих часто встречающихся аллелей DRB1, можно предположить, что 92% А 02-положительных европеоидов экспрессируют по крайней мере один аллель DRB1, который связывается с пептидом в соответствии с SEQ ID26. Исходный ген, из которого образованы SEQ ID26 по 30 - сурвивин - был представлен в гиперэкспрессированном состоянии при глиобластоме, опухоли предстательной железы, раке молочной железы, раке пищевода, колоректальном раке, светлоклеточной почечной карциноме, раке легких, ЦНС, яичника, меланоме (Tamm et al. 1998), раке поджелудочной железы, плоскоклеточной карциноме, лейкемии и медуллобластоме в сравнении с нормальными тканями (см. пример 2 и фиг. 2), демонстрируя высокую степень ассоциации данных пептидов с опухолью, т.е. эти пептиды в большом количестве презентируются на опухолевой ткани, но не на нормальных тканях. В WO 2004/067023 описываются рестриктированные по МНС класса I пептиды, образованные из опухолеассоциированного антигена сурвивина, пептиды которого способны связываться с высокой аффинностью с молекулами I класса HLA. Связанные с HLA пептиды могут распознаваться иммунной системой, специально Тлимфоцитами/Т-клетками. Т-клетки могут разрушать клетки, презентирующие распознанный комплексHLA/пептид; к примеру, опухолевые клетки глиобластомы, презентирующие полученные пептиды. Тхелперные клетки, активированные образованными из сурвивина пептидами, могут ингибировать васкуляризацию опухоли, могут привлекать эффекторные клетки иммунной системы и способствовать праймингу ЦТЛ, пролиферации и устойчивому CD8+ Т-клеточному ответу. Показано, что все пептиды настоящего изобретения в состоянии стимулировать Т-клеточные ответы (см. пример 3 и фиг. 3). Таким образом, пептиды полезны для генерирования иммунного ответа у пациента, который может уничтожать опухолевые клетки. Иммунный ответ у пациента может быть индуцирован при непосредственном введении описанных пептидов или подходящих веществпредшественников (к примеру, удлиненных пептидов, белков или нуклеиновых кислот, кодирующих эти пептиды) пациенту, в идеальном случае в комбинации с веществом, усиливающим иммуногенность (т.е. адъювантом). От иммунного ответа, вызванного такой терапевтической вакцинацией, может ожидаться,что он будет высоко специфическим против опухолевых клеток, так как целевые пептиды настоящего изобретения не презентируются на нормальных тканях со сравнимыми количествами копий, предотвращая тем самым риск нежелательных аутоиммунных реакций против нормальных клеток у пациента. Фармацевтические композиции включают пептиды как в свободной форме, так и в форме фармацевтически приемлемой соли. Используемое здесь понятие "фармацевтически приемлемая соль" относится к производным раскрытых пептидов, причм пептид модифицирован путм получения кислотных или щелочных солей агента. Например, кислотные соли получают из свободного основания (типично, где нейтральная форма лекарственного средства имеет нейтральную группу -NH2) с применением реакции с подходящей кислотой. Подходящие кислоты для получения кислотных солей включают как органические кислоты, например уксусную кислоту, пропионовую кислоту, гликолевую кислоту, пировиноградную кислоту, щавелевую кислоту, оксиянтарную кислоту, малоновую кислоту, янтарную кислоту, малеиновую кислоту, фумаровую кислоту, винную кислоту, лимонную кислоту, бензойную кислоту, коричную кислоту, миндальную кислоту, метансульфоновую кислоту, этансульфоновую кислоту, р-толуолсульфокислоту, салициловую кислоту и подобные, так и неорганические кислоты, например хлористоводородную кислоту,бромистоводородную кислоту, серную кислоту, азотную кислоту, фосфорную кислоту и подобные. И наоборот, приготовление щелочных солей кислотных составляющих, которые могут присутствовать на пептиде, приготовляются при использовании фармацевтически приемлемого основания, такого как гидроксид натрия, гидроксид калия, гидроксид аммония, гидроксид кальция, триметиламин или подобных. В одном особенно предпочтительном воплощении фармацевтические композиции включают пептиды в виде солей уксусной кислоты (ацетаты) или хлористоводородной кислоты (хлориды). Кроме того, пептиды настоящего изобретения полезны не только для лечения рака, но и также в качестве диагностических средств. Так как пептиды были получены из глиобластомы, и так как было определено, что данные пептиды не присутствуют в нормальных тканях, то эти пептиды могут быть исполь- 12023013 зованы для постановки диагноза о наличии рака. Присутствие заявленных пептидов на тканевых биоптатах может помочь патологу в постановке диагноза рака. Детекция конкретных пептидов с помощью антител, масс-спектрометрии или других методов, известных из уровня техники, могут дать знать патологу, что ткань является злокачественной или воспаленной или же поражена заболеванием общего порядка. Присутствие групп пептидов может позволить классифицировать или выделить подклассы пораженных тканей. Детекция пептидов на образцах пораженной заболеванием ткани может позволить принять решение о преимуществах от терапии, воздействующей на иммунную систему, в особенности, если Т-лимфоциты,как известно или ожидается, задействованы в механизме действия. Отсутствие экспрессии МНС является хорошо описанным механизмом, при котором инфицированные или злокачественные клетки уклоняются от иммунного надзора. Таким образом, присутствие пептидов показывает, что этот механизм не используется проанализированными клетками. Пептиды могут использоваться в анализе ответов лимфоцитов против таковых пептидов, таких как Т-клеточные ответы или ответы антител против пептида или пептида в комплексе с молекулами МНС. Такие иммунные ответы лимфоцитов могут использоваться в качестве прогностических маркеров для принятия решения о дальнейших этапах терапии. Данные иммунные ответы могут также использоваться в качестве суррогатных маркеров в иммунотерапевтических подходах, направленных на индуцирование ответов лимфоцитов с помощью различных средств, как, например, вакцинации белком, нуклеиновыми кислотами, аутологичными материалами, адоптивного переноса лимфоцитов. При генной терапии ответы лимфоцитов на пептиды могут быть рассмотрены в рамках оценки побочных эффектов. Мониторинг ответов лимфоцитов может также быть ценным инструментом для последующих обследований в случае трансплантации, к примеру, для детекции реакций "хозяин против трансплантата" и "трансплантат против хозяина". Пептиды могут использоваться для генерации и разработки специфических антител к комплексам МНС/пептид. Они могут быть использованы в терапии, нацеливающей токсины или радиоактивные вещества на пораженную ткань. Другим видом использования данных антител может быть "нацеливание" радионуклидов на пораженную ткань в целях визуализации, такой как PET (позитронно-эмиссионная томография). Это может помочь в обнаружении небольших метастазов или в определении размера и точного расположения пораженных тканей. Кроме того, пептиды могут быть использованы для верификации диагноза патолога, поставленного на основании биоптата. В табл. 1 показаны пептиды в соответствии с настоящим изобретением, соответствующие им номера последовательностей SEQ ID ; аллели HLA, с которыми связываются соответствующие пептиды, и исходные белки, из которых могут быть образованы данные пептиды. Особенно интересен факт, что пептид в соответствии с SEQ ID1 связывается с HLA-DR в равной степени, как и с HLA-A02, таким образом, вызывая два различных ответа. Таблица 1. Пептиды изобретения Хондроитин сульфат протеогликан 4 (CSPG4).CSPG4 (хондроитин сульфат протеогликан) представляет интегральный мембранный хондроитин сульфат протеогликан на зарождающихся перицитах с функциональной ролью в неоваскуляризации (Ozerdem, 2006). Во время эмбриогенеза CSPG4 протеогликан экспрессируется на незрелых капиллярах, но при созревании сосудов эта экспрессия утрачивается. Он известен как ранний маркер прогрессии меланомы, присутствующий на клеточной поверхности и вовлеченный в стимуляцию пролиферации, миграции и инвазии опухолевой клетки. CSPG4 сильно экспрессирован в 90% случаях человеческой меланомы. Хотя CSPG4 не является строго опухолеспецифическим, опухолереактивные ответы CD4+ Т-клеток у пациентов с меланомой и здоровых индивидов распознают CSPG4693-709 на HLA-DR11 экспрессирующих клетках меланомы при отсутствии аутоиммунности (Erfurt et al., 2007). Экспрессия CSPG4 улучшает интегрин-опосредованное распространение клеток, фосфориляцию(Yang et al., 2004). Кроме того, накоплены доказательства на основе данных in vitro о том, что CSPG4 играет важную роль в ангиогенезе опухоли. Таким образом, CSPG4-положительные опухоли, как было обнаружено, имеют значительно повышенную степень неоваскуляризации и объем сосудистой массы, и,как было продемонстрировано, CSPG4 отделяет ангиостатин, который в нормальных условиях ингибирует пролиферацию и ангиогенез эндотелиальных клеток. Незрелые сосуды также содержат CSPG4 положительные перициты, позволяя предположить, что эта клеточная популяция играет роль в модулировании пролиферации эндотелиальных клеток, блокируя ингибирующее действие ангиостатина во время развития сосудов (Chekenya et al., 2002b). Кроме активированных перицитов экспрессия CSPG4 была также описана для некоторых нормальных тканей, таких как эндотелиальные клетки, хондроциты, гладкомышечные клетки, определенные виды базальных кератиноцитов внутри эпидермиса, в равной степени, как и внутри волосяного фолликула(Campoli et al., 2004). Во время ангиогенеза и в ответ на патологии CNS высокоподвижные клетки CSPG4 подвергаются быстрым морфологическим изменениям и мобилизуются в места, где происходит рост и восстановление сосудов. CSPG4 гиперэкспрессированы и в опухолевых клетках, и в перицитах кровяных сосудов злокачественных опухолей головного мозга (Chekenya and Pilkington, 2002). Вживлением клеток из CSPG4 положительной клеточной линии человеческой глиомы в головной мозг иммунодефицитных "голых" крыс было продемонстрировано, что данные опухоли имеют более высокую капиллярную плотность по сравнению с контрольными пробами; это предполагает, что экспрессией CSPG4 регулируются как функции, так и структура полученной от хозяина сосудистой сети опухоли (Brekke et al., 2006). В эксперименте с ксенотрансплантатом по вживлению сфероидов мультиформной глиобластомы (GBM), полученных биопсией, в "голых" крыс было установлено, что CSPG4 в основном ассоциируется с кровеносными сосудами как перицитов, так и компонентов основания мембраны сосудистой сети опухоли, и экспрессия- 14023013 ассоциирована с участками высокой клеточной пролиферации (Chekenya et al., 2002 а). Кроме того, экспрессия CSPG4 сравнима с прогрессией опухоли в модели с имплантированной глиомой (Wiranowska etal., 2006). Прогрессия злокачественного характера сохраняется при перекрестных помехах между опухолью и ее стромой, где активированная строма питает пролиферативные и инвазивные клетки новообразования, обеспечивая образование новых сосудов, внеклеточных матричных компонентов и стимулирующих факторов роста. В данном контексте CSPG4 играет ведущую роль в активации опухолевой стромы посредством изменений в клеточной адгезии, миграции, пролиферации и морфогенезе сосудовCSPG4 дифференциально экспрессирован в человеческих глиомах с более высокой экспрессией в глиомах высокой степени злокачественности по сравнению с глиомами низкой степени злокачественности (Chekenya et al., 1999). Высокая экспрессия CSPG4 соотносится с мультилекарственной резистентностью, опосредованной возросшей активацией сигнальных процессов с участием 31 интегрин/PI3K и их мишеней по ходу транскрипции, способствуя выживанию клеток (Chekenya et al., 2008).CSP-001 был обнаружен в следующих органах/тканях и видах рака: головной мозг: глиобластома; вторичная глиобластома (образованная из астроцитомы); толстая кишка: аденокарцинома (за исключением муцинозного типа), первичная; прямая кишка: аденокарцинома с метастазами; желудок: аденокарцинома (за исключением перстневидно-клеточного типа), первичная; почка: почечно-клеточная карцинома, клеточная линия; почечно-клеточная карцинома, светлоклеточный тип с метастазами, все вторичные участки; почечно-клеточная карцинома, светлоклеточный тип,первичная; почечно-клеточная карцинома, первичная; легкие: аденокарцинома, первичная; аденосквамозная карцинома, первичная; первичный рак; мелкоклеточная карцинома, первичная; сквамозная карцинома, первичная; поджелудочная железа: аденокарцинома, первичная; рак из островковых клеток, злокачественный с метастазами; предстательная железа: аденокарцинома, первичная; кожа: метастатическая злокачественная меланома, метастазы лимфатических узлов. Поэтому фармацевтическая композиция, содержащая пептид в соответствии с SEQ ID1, особенно предпочтительна для лечения головного мозга: глиобластома; вторичная глиобластома (образованная из астроцитомы); толстой кишки: аденокарцинома (за исключением муцинозного типа), первичная; прямой кишки: аденокарцинома с метастазами; желудка: аденокарцинома (за исключением перстневидно-клеточного типа), первичная; почек: почечно-клеточная карцинома, клеточная линия; почечно-клеточная карцинома, светлоклеточный тип с метастазами, все вторичные участки; почечно-клеточная карцинома, светлоклеточный тип,первичная; почечно-клеточная карцинома, первичная; легких: аденокарцинома, первичная; I-й стадии, аденосквамозная карцинома, первичная; первичный рак; мелкоклеточная карцинома, первичная; сквамозная карцинома, первичная; поджелудочной железы: аденокарцинома, первичная; рак из островковых клеток, злокачественный с метастазами; предстательной железы: аденокарцинома, первичная; кожи: метастатическая злокачественная меланома, метастазы лимфатических узлов. Ацил-СоА синтетаза, член семейства bubblegum 1 (ACSBG1). Белок, кодируемый этим геном, обладает активностью длинноцепочной ацил-СоА синтетазы. Считается, что он играет центральную роль в головном мозге при активации метаболизма жирных кислот с очень длинными цепями и при миелинизации. Активация жирных кислот при реакции тиоэтерификации до Ацил-СоА является предпосылкой для большинства реакций, в которых участвует этот класс молекул. Специфические для рака функции или гиперэкспрессия до сих пор не были описаны в литературе. Паттерны экспрессии ACSBG1 в головном мозге, надпочечной железе, семеннике и яичнике и его функция предполагают роль данного белка в биохимической патологии Х-сцепленной адренолейкодистрофии(XALD). XALD - тяжелое, зачастую оканчивающееся летальным исходом нейродегенеративное нарушение, характеризующееся повышенным уровнем насыщенных жирных кислот очень длинных цепей в плазме и тканях (Asheuer et al., 2005; Pei et al., 2003). Бревикан (BCAN). Бревикан является внеклеточным матричным белком, сильно экспрессируемым при рождении, который экспрессируется от рождения в течение первых 8 лет жизни, а к 20 годам его активность снижается до низких уровней, которые сохраняются в нормальном взрослом корковом веществе. Изоформа GPI экспрессирована на постоянно низких уровнях во время развития (Gary et al., 2000). Злокачественные глиомы агрессивно поражают окружающий нормальный головной мозг, что может быть опосредованно ткане-или опухолеспецифическими внеклеточными белками. Таким образом, внеклеточный матрикс отчасти может модулировать терпимость ткани по отношению к движению клетки. Соответственно, способность глиом модифицировать внеклеточный матрикс ЦНС может способствовать инвазивности этих- 15023013 клеток. Одной молекулой внеклеточного матрикса, которая имеет чрезмерно повышенный уровень в глиомах, является BCAN, специфический для головного мозга хондроитин сульфат протеогликан. Уровень экспрессии BCAN увеличивается также во время периодов повышенной подвижности глиальных клеток во время развития и после травмы головного мозга. В глиоме может быть установлено приблизительно семикратное увеличение уровня экспрессии по отношению к нормальным тканям (Gary et al.,2000; Gary et al., 1998). В дополнение к увеличению уровня BCAN в глиоме протеолитический процессинг белка полной длины может также вносить свой вклад в инвазию (Gary et al., 1998; Nutt et al., 2001). Как могло быть показано, протеолитический процессинг BCAN металлопротеазами семейства ADAMTS является необходимым этапом в опосредовании его про-инвазивного эффекта в глиоме. Мутант, "не расщепляемая" форма BCAN не способна усиливать инвазивность глиомных клеток in vitro и прогрессию опухоли in vivo (Viapiano et al., 2008). мРНК не детектируется для BCAN в нормальной взрослой коре головного мозга или в какой-либо опухоли не глиомного ряда, так что BCAN считается единственным и селективным маркером в глиоме (Jaworski et al., 1996). Кроме того, анализ белка раскрыл не только повышенный уровень экспрессии BCAN полной длины, но и также присутствие дополнительных уникальных изоформ в глиоме. Таким образом, B/bg является продуктом BCAN мРНК полной длины, который возникает из неполного или сокращенного гликозилирования центрального белка. B/bg отсутствует в нормальном взрослом мозге, но обнаруживается в пробах глиом высокой степени злокачественности (Viapiano et al., 2005).BCAN описывали как селективно гиперэкспрессированный в типе образованных из глиобластомы опухолевых клеток, подобных стволовым (Gunther et al., 2008). Этот подтип клеток, подобных стволовым, проявлял наиболее высокую плюрипотентность и онкогенность. Хитиназа-3-подобный белок 1 (хрящевой гликопротеин-39) (CHI3L1).CHI3L1, член семейства "хитиназа-подобных белков млекопитающих", экспрессируется и секретируется несколькими видами солидных опухолей. Он вырабатывается раковыми клетками и опухолеассоциированными макрофагами, проявляет активность фактора роста для клеток, задействованных в процессах ремоделирования тканей, и может играть роль в пролиферации, дифференциации, инвазивности,метастазах раковых клеток, в ангиогенезе и воспалительных процессах и ремоделировании внеклеточного матрикса, окружающего опухоль (Johansen et al., 2006; Johansen et al., 2007; Ringsholt et al., 2007). Кроме того, CHI3L1 является аутологичным антигеном-кандидатом при ревматоидном артрите. ЛинииCD4 Т-клеток здоровых доноров, направленные против CHI3L1, экспрессировали CD25, глюкокортикоид-индуцированный фактор некроза, и молекулы Foxp3 были в состоянии подавлять антигенспецифические Т-клеточные ответы. Ответы 50% пациентов с ревматоидным артритом проявляют поляризацию по отношению к провоспалительному Т-хелперному 1 фенотипу и являются существенно менее действенными в подавлении антиген-специфических ответов (van Bilsen et al., 2004).CHI3L1 находится в повышенном количестве в онкостатине М, о котором известно, что он индуцируется в нервной системе в результате клеточного стресса, он экспрессирован в большинстве человеческих опухолей головного мозга и активирует сигнальный путь JAK/STAT (Krona et al., 2007). ЭкспрессияCHI3L1 была также ассоциирована с экспрессией р-МАРК, p-mTOR и p-p70S6K в глиобластоме (Pelloskiet al., 2006). В нескольких клинических исследованиях генной экспрессии было показано, что CHI3L1 был экспрессирован в более высоком количестве в глиобластоме в сравнении с нормальным головным мозгом с превышением уровня в нормальном головном мозге от 3 до 62 крат (Saidi et al., 2007; Kroes et al., 2007;Shostak et al., 2003; Tanwar et al., 2002) Во время иммуногистохимических исследований обнаружилось,что клетки с функционирующим ядром способны экспрессировать CHI3L1 в своей цитоплазме, но интенсивность экспрессии CHI3L1 зависела от клеточной активности. Таким образом, клетки, о которых известно, что они вызывают высокую метаболическую активность, имели тенденцию демонстрировать наиболее интенсивное цитоплазматическое окрашивание (Ringsholt et al., 2007). В дальнейшем с помощью иммуногистохимии могло быть продемонстрировано, что глиобластомы демонстрируют поразительно большую экспрессию CHI3L1, чем анапластические олигодендроглиомы (Nutt et al., 2005). Анализ методом иммунного блотинга (Western-blot) проб глиомы на уровень белка CHI3L1 показал значительное повышение в 65% глиобластом и не обнаруживаемые уровни в глиомах низкой степени злокачественности (II и III степени) или нормальной ткани головного мозга (Tanwar et al., 2002). В сравнении с пилоидной астроцитомой, которая не распространяется и может быть вылечена с помощью операции,только глиобластома экспрессирует CHI3L1 (Colin et al., 2006). Уровень сыворотки CHI3L1 повышен во множестве злокачественных образований и был ассоциирован с худшей выживаемостью. Наиболее высокие уровни сыворотки CHI3L1 были обнаружены у пациентов с метастатическим раком с наиболее коротким безрецидивным периодом и наиболее коротким сроком общей выживаемости. В особенности в сыворотке пациентов с глиобластомой наблюдалась повышенная экспрессия CHI3L1 (Kim et al., 2007; Johansen et al., 2007; Johansen et al., 2006; Junker et al.,2005; Tanwar et al., 2002). Пациенты с глиобластомой с активной опухолью имеют значительно повышенный уровень CHI3L1 по сравнению с пациентами, не имеющими радиографического подтверждения- 16023013 заболевания. Кроме того, существует значительное обратнопропорциональное соотношение междуCHI3L1 и выживаемостью при GBM (Hormigo et al., 2006; Pelloski et al., 2005). В дополнение, повышенная экспрессия CHI3L1 может наблюдаться при раке молочной железы, где она соотносится с увеличенным размером опухоли, худшей дифференциацией опухоли и более краткими сроками выживаемости без заболевания (Kim et al., 2007; Coskun et al., 2007). Более того, при сквамозной клеточной карциноме головы и шеи повышенные уровни сыворотки CHI3L1 были обнаружены в 53%. Пациенты с высоким уровнем сыворотки CHI3L1 имеют более краткий срок выживаемости, чем пациенты с нормальной сывороткой CHI3L1 (33 по отношению к 84 месяцам) (Roslind et al., 2008). Пациенты, страдающие от рака предстательной железы, демонстрировали значительно более высокие уровни сыворотки CHI3L1 в сравнении с пациентами с ДКПЖ или здоровыми индивидами (Kucur etCAP-GLY доменсодержащий линкерный белок 2 (CLIP2). Белок, кодируемый CLIP2, принадлежит к семейству цитоплазматических линкерных белков, которые были предложены для опосредования взаимодействия между специфическими мембранными органеллами и микротрубочками. Было обнаружено, что CLIP2 ассоциирован как с микротрубочками, так и с органеллами, называемыми дендритная ламеллярная структура (общая информация с веб-сайта NCBI).CLIP2 расположен на концах тирозинированных микротрубочек, но не на концах детирозинированных микротрубочек. Тубулин-тирозин-лигаза (TTL), фермент, который катализирует добавление Стерминального тирозинового остатка к альфа-тубулину в тубулин-тирозинационном цикле, задействован в опухолевой прогрессии и играет жизненно важную роль в нейрональной организации (Peris et al., 2006). В одном исследовании геномной ДНК методом GenoSensor Array 300 на замороженных срезах 30 случаев первичной глиобластомы характеризовали генные амплификации, генные делеции и хромосомную информацию всего домена. Гены, которые часто были амплифицированы, включали =CLIP2 (63,3%),EGFR (53,3%), IL6 (53,3%), АВСВ 1 (MDR1) (36,7%) и PDGFRA (26,7%) (Suzuki et al., 2004). Семейство транспортеров жидких переносчиков органического аниона, член 1 С 1 (SLCO1C1).SLCO1C1 селективно экспрессирован на гематоэнцефалическом барьере (Chu et al., 2008).SLCO1C1 характеризуется селективным предпочтением субстрата и может играть важную роль при распределении тироидных гормонов в головном мозге и семеннике (Pizzagalli et al., 2002). SLCO1C1 не детектировался специфически методом иммунофлуоресценции. SLCO1A2 и SLCO2B1 детектировались методом иммунофлуоресцентной микроскопии в люминальной мембране эндотелиальных клеток, формирующих гематоэнцефалический барьер и барьер между кровью и опухолью, но не в клетках глиомы(Bronger et al., 2005). Дистробревин, альфа (DTNA). Альфа-дистробревин первоначально был описан как компонент цитоплазмы дистрофингликопротеинового комплекса в клетках скелетных мышц. Изоформы DTNA демонстрируют различную локализацию в клетках и тканях; на базолатеральных мембранах в эпителиальных клетках дистробревины опосредуют контакт с внеклеточным матриксом, периферийными и трансмембранными белками и филаментным актиновым цитоскелетом. Помимо их структурообразующей роли, предполагается, чтоDTNA имеют важное значение в клеточных сигнальных реакциях и в клеточной дифференциации и они ассоциированы с плотными сочленениями во время их реорганизации (Sjo et al., 2005). DTNA может быть задействован в формировании и стабилизации синапсов, в равной степени как и в образовании кластеров никотиновых ацетилхолиновых рецепторов. Рецептор эпидермального фактора роста (эритробластический лейкемия вирусный (v-erb-b) онкогенный гомолог, птичий) (EGFR). Рецептор эпидермального фактора роста (EGFR) находится в последнее время в центре внимания,так как его нарушения являются одним из наиболее распространенных молекулярных отклонений от нормы при глиобластоме. В особенности EGFRvIII (рецептор эпидермального фактора роста, вариант III) является онкогенной, конститутивно активной мутантной формой белка EGFR, которая обычно экспрессируется в глиобластоме (Zawrocki and Biernat, 2005). EGFR задействован в активации спектра сигнальных путей, которые регулируют фенотип клеток-предшественников. Активированная EGFR тирозинокиназная активность улучшает миграционные процессы, пролиферацию нейтральных стволовых клеток и выживаемость. Так как, как известно, сигнальные реакции EGFR играют роль при глиобластоме, то можно сделать заключение, что глиобластома образуется из стволовых раковых клеток и что сигналы EGFR обычно изменены в этих клетках-предшественниках (Yuso-Sacido et al., 2006). Первоначально глиобластомы появляются de novo у пациентов постарше и часто гиперэкспрессируют EGFR. Гиперэкспрессия EGFR соотносится с повышенным ангиогенезом, эдемой (отечностью) и инвазией (Aghi et al., 2005). Кроме того, EGFR-амплифицированные глиобластомы имеют резистентность к лучевой терапии (Barker et al., 2001) и рецидивируют быстрее после лечения (Schlegel et al.,1994).GBM - единственная неэпителиальная опухоль человека, для которой эксцессивная активацияEGFR является протоонкогеном erbB. Гиперэкспрессия EGFR может усиливать рост клеток из-за увеличения формирования активных комплексов лиганд-рецептор. Генная амплификация - это механизм,усиливающий гиперэкспрессию рецепторов EGF в опухолях GBM (Thompson and Gill, 1985). О генеEGFR на хромосоме 7 известно, что он часто прибавляет число копий в глиомах высокой степени злокачественности (Okada et al., 2007). Элиминация EGFR при короткой РНК-интерференции ликвидирует онкогенез клеток глиобластомы (Huang et al., 2007). Гиперэкспрессия EGFR детектируется в 40-70% глиобластом, причем астроцитома: пилоидная,низкой степени злокачественности или анапластичная являются неизменно EGFR-негативными. (Agostiet al., 2006a). Высокие уровни сыворотки EGFR указывают на сокращенный срок выживаемости (Quaranta et al., 2007). Кроме того, как было показано, пациенты с долгим сроком выживаемости с астроцитомами высокой степени злокачественности являются EGFRvIII-негативными (Liang et al., 2008). Локус Notch-1 повышает уровень экспрессии EGFR и соотношения между уровнями EGFR, иNotch-1 мРНК может быть обнаружен на первичных глиомах человека высокой степени злокачественности (Purow et al., 2008). EGFR сам по себе задействован в конститутивной активации c-Jun NH2 терминальной киназы (JNK), которая способствует пролиферации, выживаемости и онкогенезу некоторых опухолей, включая глиомы (Li et al., 2008a). Хотя EGFRvIII экспрессирован лишь малой долей глиомных клеток, у большинства клеток имеется трансформированный фенотип. Как было продемонстрировано, экспрессия EGFRvIII в клетках индолентной глиомы стимулирует формирование относящихся к липидному рафту микрополостей, содержащих EGFRvIII, которые высвобождаются в клеточное окружение и могут сливаться с плазменными мембранами раковых клеток, в которых недостаточно EGFRvIII, приводя к перемещению онкогенной активности (А 1-Nedawi et al., 2008). Белок, связывающий жирную кислоту 7, головной мозг (FABP7). Белки, связывающие жирную кислоту (FABPs), являются цитозольными белками 14-15 кДа, которые, как предполагается, задействованы в поглощении, транспортировке и нацеливании жирной кислоты(ЖК). Они могут модулировать концентрацию ЖК и таким способом влиять на работу ферментов, мембран, ионных каналов и рецепторов, а также на генную экспрессию и клеточный рост и дифференциацию. Различают девять видов FABP со специфическим распространением в ткани. Вместо 30-70% идентичности аминокислотной последовательности они имеют похожую третичную бета-структуру "скользящей застежки", в которой связана ЖК. Нервная ткань содержит четыре вида FABP с различным пространственно-временным распространением (Veerkamp and Zimmerman, 2001). FABP7 высоко экспрессирован в развивающемся головном мозге и сетчатке глаза, и его экспрессия значительно снижается во взрослой ЦНС (Godbout et al., 1998). Основываясь на результатах, полученных in vitro, предполагается,что FABP7 требуется для образования радиальной глиальной системы развивающегося головного мозга(Mita et al., 2007). В нормальном головном мозге белок FABP7 практически не детектируется, но проявляет от умеренной до сильной ядерной и цитоплазмической экспрессии в нескольких мультиформных глиобластомах (GBM). Трансфицированные FABP7 клетки проявляют в 5 раз большую миграционную активность, чем контрольные клетки. Таким образом, более короткая общая выживаемость, ассоциированная с гиперэкспрессией FABP7, в особенности в глиобластоме, может быть обусловлена повышенной миграцией и инвазией опухолевых клеток в окружающую мозговую паренхиму (Liang et al., 2005). Ядерная транслокация FABP7 в особенности соотносится с амплификацией EGFR и более инвазивными опухолями (Kaloshi et al., 2007). Таким образом, FABP7 может быть индуцирован активацией EGFR, чтобы способствовать миграции опухолевых клеток GBM (Liang et al., 2006). Экспрессия FABP7, как было установлено, является также маркером для почечно-клеточной карциномы. Экспрессия FABP7 может быть детектирована только в раковых тканях, но не в не пораженных раком частях проб почек (Teratani et al., 2007). Экспрессия FABP7 в почечно-клеточной карциноме, как было показано, в 20 раз выше в опухоли в сравнении с нормальной почкой (Domoto et al., 2007; Buchneret al., 2007). Было показано, что FABP7 часто экспрессирован в меланоме, где он может быть задействован в клеточной пролиферации и инвазии (Goto et al., 2006). Глиофибриллярный кислый белок (GFAP).GFAP кодирует один из крупнейших белков промежуточных филаментов зрелых астроцитов. Он используется в качестве маркера для отличия астроцитов от других глиальных клеток во время развития. Мутации этого гена приводят к болезни Александера, редкое нарушение деятельности астроцитов в центральной нервной системе. Был описан дополнительный вариант транскрипта, однако его последовательность полной длины определена не была. Сообщалось о повышенном уровне в аутистическом головном мозге, в то время как в головном мозге людей с серьезной депрессией проявлялся пониженный уровень GFAP. Анализу подвергли головной мозг приматов, у которых заново возникли опухоли десять лет спустя после облучения головного мозга. Предшественники опухоли проявляли клеточный атипизм и митозы и были негативными для опухолеассоциированных маркеров GFAP, EGFR и р 53 (Lubensky et al., 2006).B новообразованиях с участием астроцитов количество GFAP-положительных клеток и интенсивность ок- 18023013 рашивания были прямо пропорциональны степени злокачественности. Во всех 3 случаях олигодендроглиомы происходила негативная реакция на GFAP (Reyaz et al., 2005). Чистые олигодендроглиомы являются иммуногистологически негативными для GFAP (Mokhtari et al., 2005). Уровни сыворотки GFAP у пациентов с глиомой высокой степени демонстрировали линейную корреляцию с объемом опухоли(Brommeland et al., 2007). Даже среди пациентов с глиобластомой может быть установлена значительная корреляция между объемом опухоли, объемом некроза опухоли, количеством некротических GFAPположительных клеток и уровнем GFAP в сыворотке (Jung et al., 2007). Последующее воздействие на клеточные линии глиобластомы ингибитором гистонной дезацетилазы 4-фенилбутиратом приводили к повышению концентрации нефосфорилированного GFAP, в то время как фосфорилированные изоформы оставались без изменений (Asklund et al., 2004). В клеточной линии глиобластомы, обработанной TGF-альфа, приблизительно на 50% снизились генная транскрипция GFAP, уровень мРНК и специфический синтез белка (Zhou and Skalli, 2000).C технической точки зрения промотор GFAP часто используется в качестве инструмента в моделях с мышами для индуцирования экспрессии желаемых белков, в особенности в нервной системе. Островковые клетки Лангерганса поджелудочной железы окутаны околоостровковыми швановскими клетками (pSC), экспрессирующими GFAP. Аутоиммунное нацеливание элементов ткани панкреатической нервной системы, вероятно, является неотъемлемой, рано формируемой, частью естественно развившегося диабета типа 1 (Winer et al., 2003). Эта панкреатическая экспрессия не отражена в данных о генной экспрессии в смешанных тканях фирмы "immatics" или других источниках. GFAP-001 был опубликован в качестве эпитопа, против которого пациенты с диабетом типа 1 в равной степени как и их не болеющие диабетом родственники с ответами антител против диабетических аутологичных антигенов(повышенный риск заболевания диабетом) проявляли усиленную реактивность секретирующих гранзим В ЦТЛ (ex vivo ELISPOT) по сравнению со здоровыми донорами (Standifer et al., 2006). Интересно, что, вероятно, существует обратное соотношение между обнаружением аутоиммунных заболеваний, в особенности диабета, и риском развития глиомы (Aronson и Aronson, 1965; Schlehofer etGPR56 - атипичный рецептор, связанный с G-белком (GPCR), с необычно крупным Nтерминальным внеклеточным участком, который содержит длинный Ser/Thr-обогащенный участок, формирующий муцин-подобный "стебель", и из-за этой черты считается, что GPR56 играет роль во взаимодействиях клетка-клетка или клетка-матрикс. Вместе с высоким уровнем экспрессии мРНК и его широким распространением было сделано предположение о возможной роли для этого рецептора в процессах взаимодействия клетка-клетка (Liu et al., 1999). GPR56 принадлежит к GPCR семейства секретинов, которое играет роль в развитии клеток-предшественников нервной системы и было соотнесено с пороками развития человеческого головного мозга. Более высокая экспрессия GPR56 соотносится с клеточными трансформационными фенотипами нескольких раковых тканей по сравнению с их нормальными эквивалентами, заставляя предположить потенциальную онкогенную функцию. Опосредованное РНКинтерференциями "выключение" GPR56 приводит к индукции апоптоза и снижению субстратнезависимого роста раковых клеток посредством увеличения аноикоза. "Выключение" GPR56 сокращает также клеточную адгезию к внеклеточному матриксу (Ke et al., 2007). Повышенный уровень GPR56 был продемонстрирован для мультиформной глиобластомы при использовании функциональной геномики. Иммуногистохимические исследования подтвердили экспрессию GPR56 в большинстве проб опухолевой ткани глиобластомы/астроцитомы с не устанавливаемыми уровнями экспрессии в нормальном взрослом головном мозге (Shashidhar et al., 2005). В панкреатических раковых клетках мРНК GPR56 экспрессирована на высоком уровне, причем белок GPR56 ничтожно мал и не детектируется в этих клетках, давая возможность предположить, что экспрессия белка GPR56 подавляется в человеческих панкреатических раковых клетках (Huang et al., 2008). Более ранние исследования метастазов показали, что количествоGPR56 заметно снижено в высоко метастатических вариантах клеточной линии человеческой меланомы,заставляя предположить, что гиперэкспрессия GPR56 подавляет рост опухоли и метастазы. Данное подавление роста, как предполагается, опосредуется взаимодействием GPR56 с тканевой трансглютаминазой (TG2), широко распространенным компонентом ткани и стромы, который задействован в подавлении прогрессии опухоли как таковой (Xu et al., 2006; Xu and Hynes, 2007). О другом ингибирующем воздействии GPR56 сообщалось в связи с миграцией клеток-предшественников нервной системы (NPCs).GPR56 высоко экспрессирован в NPCs и, вероятно, принимает участие в регуляции движения NPC через сигнальные пути Galpha( 12/13) и Rho, заставляя предположить, что GPR56 играет важную роль в развитии центральной нервной системы (Iguchi et al., 2008). Рецептор глютамата, ионотрофический, АМРА 4 (GRIA4). Глютаматный рецептор типа -Амин-3-гидрокси-5-метил-4-изоксазолпропионат (АМРА)(AMPARs) опосредует быструю нейротрансмиссию в возбудительных синапсах ЦНС и состоят из субъединиц, взятых из комплекта из четырех белков, GluR1 по GluR4 (GRIA4). Субъединицы GRIA4 повсеместно экспрессированы в клетках человеческой глиобластомы, действуя как Са 2+-пропускающие AMPARs. Конверсия в не пропускающие Са 2+ рецепторы ингибирует дви- 19023013 жение клеток и индуцирует апоптоз, причем гиперэкспрессия Са 2+-пропускающих рецепторов АМРА упрощает миграцию и пролиферацию опухолевых клеток. Поэтому Са 2+-пропускающие рецепторы АМРА, как предполагается, играют центральную роль в росте глиобластомы (Ishiuchi et al., 2002). Инсулиноподобный фактор роста, 2 мРНК-связывающий белок 3 (IGF2BP3).IGF2BP3 является членом белкового семейства инсулиноподобных факторов роста II, связывающих мРНК, вовлеченным в локализацию, оборот и трансляционный контроль мРНК. Закодированный белок содержит несколько КН-доменов, которые важны в связывании РНК, и о которых известно, что они вовлечены в синтез и метаболизм РНК. Он экспрессируется, в основном, во время эмбрионального развития и в некоторых опухолях. Таким образом, IGF2BP3 рассматривается как онкофетальный белок (Liao etal., 2005). Специальной информации об экспрессии IGF2BP3 в глиобластоме найдено не было, но, как описывается, IGF2BP3 гиперэкспрессирован в нескольких других злокачественных заболеваниях. Таким образом, IGF2BP3 экспрессирован в 30% из 716 проанализированных образцов почечно-клеточной карциномы, светлоклеточного типа. В этом исследовании его экспрессия была ассоциирована с высокой стадией и степенью злокачественности первичных опухолей в равной степени как и с другими побочными чертами, включая коагуляционный некроз опухоли и саркомоподобную дифференциацию. Кроме того, положительная экспрессия IGF2BP3 была ассоциирована с 5-10-кратно возросшим риском отдаленных метастазов и 42-50% ростом риска летального исхода от RCC (почечно-клеточная карцинома), позволяя предположить, что экспрессия IGF2BP3 представляет собой независимый прогностический фактор поведения агрессивной почечно-клеточной карциномы, светлоклеточного типа. (Hoffmann et al.,2008; Jiang et al., 2006; Jiang et al., 2008). Экспрессия IGF2BP3 обнаруживалась также в злокачественной меланоме в сравнении с доброкачественными невусами, где экспрессии установлено не было, даже если присутствовали диспластические черты (Pryor et al., 2008). При эндометриальном раке экспрессияIGF2BP3 тесно связана с эндометриальным раком типа II и агрессивным гистологическим фенотипом среди эндометриальных неопластических образований (Zheng et al., 2008). У 20 пациентов, страдающих плоскоклеточной карциномой пищевода, индукция специфических Т-клеточных ответов TIL (лимфоцитов, инфильтрирующих опухоль), лимфоцитов региональных лимфатических узлов и лимфоцитов периферической крови против рестриктированного по HLA-A2402 пептид-эпитопа из IGF2BP3 могла наблюдаться в 40% всех случаев (Mizukami et al., 2008). IGF2BP3 также высоко экспрессирован в карциномах поджелудочной железы. В 2 исследованиях 90% проб тканей опухолей поджелудочной железы проявляли экспрессию IGF2BP3 после иммуноокрашивания, в то время как не-неопластические ткани поджелудочной железы были негативными для IGF2BP3. Кроме того, IGF2BP3 мРНК был гиперэкспрессирован в карциномах поджелудочной железы в сравнении с не-неопластическими пробами тканей, и экспрессия прогрессивно возрастала вместе со стадией опухоли (Yantiss et al., 2005; Yantiss et al., 2008). Экспрессия IGF2BP3 была также обнаружена в существенно повышенном состоянии в уротелиальных опухолях высокой степени, в то время как он обычно не экспрессирован в доброкачественном уротелии или уротелиальных опухолях низкой степени. Более того, пациенты с IGF2BP3-положительными опухолями имеют намного более низкий процент выживаемости без прогрессирования и выживаемости без болезни, чем те, что имеют IGF2 ВР 3-негативные опухоли. У IGF2 ВР 3-положительных пациентов с поверхностно инвазивной уротелиальной карциномой при первоначальном диагнозе также продолжали развиваться метастазы, в то время как у IGF2BP3-негативных пациентов метастазов обнаружено не было. Помимо этого, данные этих исследований позволяют сделать предположение, что IGF2BP3 может быть задействован в прогрессии уротелиальных опухолей с низкой до высокой стадии как при папиллярных,так и при поражениях с ровной поверхностью (Li et al., 2008b; Sitnikova et al., 2008). Мегало-энцефалитная лейкодистрофия с подкорковой кистой 1 (MLC1). Мегало-энцефалитная лейкодистрофия с подкорковыми кистами (MLC) является аутосомальным рецессивным заболеванием белого вещества головного мозга у детей. MLC вызывается мутациями генаMLC1 (Ilja Boor et al., 2006). В соответствии со знаниями изобретателей никаких сообщений о взаимосвязи MLC1 с опухолями головного мозга в имеющейся литературе найдено не было. В одной работе исследовалось клеточное и региональное распространение MLC1 в головном мозге мышей (Schmitt et al., 2003). Наивысшая экспрессия MLC1 была обнаружена во время пре- и перинатального периода в многопотенциальных клетках-предшественниках нервной системы. В головном мозге взрослых мышей MLC1 мРНК был обнаружен исключительно в глиальных клетках. В отличие от развивающихся и зрелых астроцитов, олигодендроциты были лишены экспрессии MLC1. Нестин (NES). Во время развития происходит экстенсивная экспрессия промежуточного филамента нестина в нейроэпителиальных клетках брюшного слоя в возрасте 11 недель пост-гравидарно во всех частях ЦНС, в то время как иммунореактивность нестина сокращается во время второго и третьего триместра (Takano andBecker, 1997; Lendahl et al., 1990; Zimmerman et al., 1994; Tohyama et al., 1992). Во время и после миграции из пролиферативного брюшного слоя экспрессия нестина резко снижается в пост-митотических клетках-нейронах (Arnold и Trojanowski, 1996). Окрашивание нестина не-неопластических тканей взрослого головного мозга показало лишь слабое окрашивание очень малого количества эндотелиальных клеток (Dahlstrand et al., 1992). Нестин может экспрессироваться повторно во время неопластической транс- 20023013 формации (Veselska et al., 2006). В глиомных тканях иммунореактивность нестина появляется лишь в опухолевых клетках, и количество вырабатываемого нестина увеличивается, когда степень глиомы становится более злокачественной, приближаясь к глиобластоме. Глиобластомы (IV-я степень злокачественности) экспрессируют наиболее высокое число возникающих нестин-положительных клеток и вообще наиболее высокие уровни окрашивания нестина. Экспрессия нестина может быть детектирована в опухолевых клетках различных видов первичных опухолей ЦНС, которые имеют нейроэктодермальное происхождение, но не в метастазирующих клетках карцином (Dahlstrand et al., 1992; Tohyama et al.,1992). Нестин практически не экспрессирован в диффузных астроцитомах, переменно экспрессирован в анапластических астроцитомах и сильно и нерегулярно экспрессирован в глиобластомах, где его распространение варьируется в комплементарном пути GFAP и виментина (Schiffer et al., 2006). С клинической точки зрения нестин-негативные опухоли зародышевых клеток ЦНС не демонстрировали распространения, в то время как все опухоли, которые демонстрировали распространение, также сильно экспрессировали белок нестин (Sakurada et al., 2007). Опухолевые клетки, сильно экспрессирующие нестин, часто расположены рядом с кровеносными сосудами (Dahlstrand et al., 1992), (Kurihara et al., 2000; Sugawara et al., 2002; Florenes et al., 1994; Strojniket al., 2007), и, как предполагалось, экспрессия нестина активированным эндотелием является маркером ангиогенеза (Teranishi et al., 2007; Maderna et al., 2007; Amoh et al., 2005; Mokry et al., 2004).GBM включает трансформированных предшественников, которые несут полный комплемент функциональных характеристик, ожидаемых от стволовых клеток, включая способность генерирования опухоли. Эти клетки могут сформировать GBM даже при серийной трансплантации и могут поэтому быть идентифицированы как опухолевые стволовые клетки нервной системы (Galli et al., 2004). Эти клетки принадлежат к клеточной субпопуляции CD133+ из опухолей человеческого головного мозга и коэкспрессируют NSC (стволовые клетки нервной системы)-маркер нестин, но не дифференцированные маркеры линии дифференцировки нервной системы (Singh et al., 2004b; Singh et al., 2003; Singh et al.,2004a; Mao et al., 2007). Присутствие популяции CD133+/нестин+ в опухолях головного мозга предполагает, что нормальная стволовая клетка нервной системы могут быть клеткой происхождения глиом (Shiras et al., 2007). Так как сигнальные пути Notch активны в опухолях головного мозга и стволовых клетках, было продемонстрировано, что промотор нестина активируется в культуре посредством активностиNotch (Shih and Holland, 2006). Трансфекция клеточной линии С 6 астроцитомы у крыс с дуплексом siPHK нестина раскрыл эффективное супрессионное влияние siPHK нестина на клеточный рост культивированных клеток астроцитомы с зависимостью от дозы (Wei et al., 2008). Экспрессии нестина, как сообщается, происходит также в стволовых раковых клетках предстательной железы (Gu et al., 2007; Gipp et al., 2007) и рака поджелудочной железы (Carriere et al., 2007) в равной степени как и меланомы (Klein et al., 2007). Кроме того, нестин экспрессируется также в следующих опухолях: GIST (гастроинтестинальная стромальная опухоль) (Tsujimura et al., 2001; Sarlomo-Rikala et al.,2002), меланомы (Florenes et al., 1994; Brychtova et al., 2007), колоректальный рак (Teranishi et al., 2007) и опухоли поджелудочной железы (Ohike et al., 2007; Kleeberger et al., 2007). Экспрессия нестина может быть также обнаружена в различных нормальных тканях: об экспрессии нестина сообщалось для подоцитов нормальных зрелых клубочков почки человека. В нормальных условиях нестин экспрессируется в нескольких видах клубочковых клеток на ранних стадиях развития и ограничивается подоцитами в зрелых клубочках (Ishizaki et al., 2006), указывая на то, что нестин чрезвычайно важен для некоторых функциональных аспектов подоцитов. Взрослые клубочки проявляют иммунореактивность нестина в виментин-экспрессирующих клетках с подоцитной морфологией (Bertelli et al.,2007). Возможно, посредством взаимодействия с виментином нестин служит для улучшения механической прочности подоцитов, которые подвержены сильному растяжению во время клубочковой фильтрации (Perry et al., 2007). Таким образом, в человеческой почке нестин экспрессируется, начиная с первых этапов гломерулогенеза в подоцитах, мезангиальных и эндотелиальных клеток. Эта экспрессия ограничивается, затем, подоцитами в зрелых клубочках и не может быть обнаружена в других структурах почки взрослого человека (Su et al., 2007). Иммуногистохимический анализ раскрыл существование постоянной экспрессии нестина в корковом веществе нормальных взрослых надпочечников. Клетки, экспрессирующие нестин, преобладают в ретикулярной зоне, причем карциномы надпочечников выявляют вариабельное число нестин-иммунореактивных клеток (Toti et al., 2005). Также сообщалось об экспрессии нестина в интерстициальных клетках Кахала (ICC) в нормальном желудочно-кишечном тракте. Таким образом, большинство внутримышечных ICC антрума и все мышечно-кишечные ICC тонкого кишечника являются нестин-иммунореактивными, в равной степени как и некоторые мышечно-кишечные ICC и большинство ICC кольцевой мускулатуры толстого кишечника(Vanderwinden et al., 2002). На поджелудочной железе нестин-иммунореактивные клетки могут быть обнаружены на островковых клетках и на экзокринном участке. В зоне главного протока поджелудочной железы нестин-положительные клетки представляют малые капилляры, рассеянные на соединительной ткани, окружающей эпителий протока. Таким образом, нестин экспрессируется васкулярными эндотелиальными клетками поджелудочной железы человека (Klein et al., 2003). В самих островковых клетках- 21023013 могут быть обнаружены клетки-предшественники, экспрессирующие нестин. Выдвигается гипотеза, что эти нестин-положительные, образованные из островковых клеток, клетки-предшественники являются другой популяцией клеток, которая постоянно находится внутри островковых клеток поджелудочной железы и могут участвовать в неогенезе островковых эндокринных клеток (Zulewski et al., 2001). Из нормальной печени взрослого может быть изолирована популяция человеческих стволовых клеток печени,положительных для виментина и нестина (Herrera et al., 2006). При анализах клеточной культуры, анализе цитоскелета и матричной композиции при иммуноокрашивании обнаружилось, что клетки, происходящие из фетальных легких и взрослого костного мозга, экспрессируют белки виментин и нестин в промежуточных филаментах (Sabatini et al., 2005). Нестин был обнаружен в функциональных одонтобластах молодых постоянных зубов. Его экспрессия снижается, и нестин отсутствует в более взрослых постоянных зубах, в то время как его уровень снова повышается в кариозных и поврежденных зубах (About et al.,2000). Нестин-экспрессирующие взрослые стволовые клетки могут быть также обнаружены в перилюминальном регионе зрелой передней доли гипофиза и, при использовании генетически индуцируемой карты зачатков, было продемонстрировано, что они служат для генерирования подклассов всех шести терминально дифференцированных эндокринных видов клеток мозгового придатка. Данные стволовые клетки,так как они не играют значимой роли при органогенезе, подвергаются послеродовой экспансии и начинают вырабатывать дифференцированное потомство, которое колонизирует орган, который первоначально полностью состоял из дифференцированных клеток, образованных из эмбриональных предшественников (Gleiberman et al., 2008). Подсемейство ядерных рецепторов 2, группа Е, член 1 (NR2E1).NR2E1 (TLX) - это фактор транскрипции, который необходим для пролиферации стволовых клеток нервной системы и самообновления посредством рекрутинга пистонных дезацетилаз (HDACs) в его гены-мишени по ходу транскрипции, чтобы репрессировать их транскрипцию, что в свою очередь регулирует пролиферацию стволовых клеток нервной системы. Рекрутирование HDACs приводит к транскрипционной репрессии TLX генов-мишений, циклинзависимого ингибитора киназы, p21(CIPl/WAF1)(p21) и гена-супрессора опухоли, PTEN (Sun et al., 2007). Подсемейство TLX/HOX11 дивергентных содержащих гомеобокс генов задействовано в различных аспектах эмбриогенеза и, в случае TLX1/HOX11 иTLX3/HOX11L2, выступает главным образом в качестве онкогена при человеческой Т-клеточной острой лимфобластической лейкемии (Dixon et al., 2007). NR2E1 лежит в основе фундаментальной программы развития ретинальной организации и контролирует образование необходимого количества потомства ретинальных клеток и развитие глиальных клеток во время длительного периода ретиногенеза (Miyawakiet al., 2004). Для глиобластомы специфической информации обнаружено не было. Нейрональная молекула клеточной адгезии (NRCAM). Человеческая NRCAM (молекула клеточной адгезии, связанная с нейроглией) гиперэкспрессирована в ткани мультиформной глиобластомы (GMT) в сравнении с нормальной тканью головного мозга(NBT). NRCAM описывается как однопроходной трансмембранный белок типа I, взаимодействующий с анкирином. Антисмысловой hNRCAM был причиной снижения нативной экспрессии hNRCAM, изменений в клеточной морфологии, пониженной клеточной пролиферации и удлинения клеточного цикла. Кроме того, гиперэкспрессия антисмыслового hNRCAM в этих клетках вызывала обширную редукцию числа колоний мягкого агара и инвазию через гель внеклеточного матрикса (ЕСМ) in vitro. Подкожная инъекция антисмыслового hNRCAM, гиперэкспрессирующего клетки глиобластомы в "голых" мышей,вызывала полное ингибирование опухолевого образования по сравнению с вектором только трансфецированных клеток. Внутриопухолевая инокуляция плазмидом, экспрессирующим антисмысловойhNRCAM, также была причиной медленного роста опухоли у "голых" мышей in vivo (Sehgal et al., 1999). Ген-специфический анализ RT-PCR указал на то, что hNRCAM гиперэкспрессирован в астроцитомах высокой степени злокачественности, глиомах и опухолевых тканях глиобластомы по сравнению с нормальной тканью головного мозга (Sehgal et al., 1998). NRCAM, молекула межклеточной адгезии семейства иммуноглобулин-подобных молекул клеточной адгезии, известная своей функцией в разрастании и управлении нейронами, недавно была идентифицирована как ген-мишень сигнального пути бетакатенина в меланоме человека, а также клетках и ткани карциномы толстой кишки. Опосредованная ретровирусами трансдукция NRCAM в фибробласты индуцирует клеточную подвижность и онкогенез(Conacci-Sorrell et al., 2005). Индукция транскрипции NRCAM бета-катенином или плакоглобином играет роль при онкогенезе в меланоме и раке толстого кишечника, вероятно, способствуя росту и подвижности клеток (Conacci-Sorrell et al., 2002). Также и другие мишени в сигнальных путях бета-катенина находятся в повышенном количестве, такие как MYC (Liu et al., 2008). NrCAM гиперэкспрессирован в папиллярных карциномах щитовидной железы человека на уровне мРНК и белка на любой стадии опухоли (Gorka etal., 2007). Гиперэкспрессия NRCAM мРНК в опухолях ассоциирована с индикаторами высокой пролиферации и была ассоциирована с неутешительным исходом при эпендимомах (Zangen et al., 2007). Подопланин (PDPN).PDPN является муцинподобным интегральным мембранным гликопротеином типа I с различным- 22023013 распространением в человеческих тканях. Он задействован в миграции раковых клеток, инвазии, метастазах и злокачественной прогрессии, а также он участвует в накоплении тромбоцитов. CLEC-2 является первым идентифицированным патофизиологическим рецептором подопланина (Kato et al., 2008). Было исследовано 115 случаев глиобластомы с помощью иммуногистохимического метода с использованием анти-PDPN-антитела. 47% глиобластом экспрессировали PDPN на поверхностных клетках в особенности вокруг некротических участков и пролиферирующих эндотелиальных клеток. Кроме того, PDPN мРНК и белковая экспрессия были значительно выше в глиобластоме, чем в анапластичных астроцитомах, позволяя предположить, что экспрессия PDPN может быть ассоциирована с злокачественностью астроцитов (Mishima et al., 2006). Было также продемонстрировано, что PDPN экспрессируется в 82,9% глиобластом (29/35) в других анализах (Shibahara et al., 2006). В исследовании, изучающем экспрессию PDPN и действия по накоплению тромбоцитов клеточных линий глиобластомы, LN319 экспрессировал в высокой степени PDPN и индуцировал накопление тромбоцитов. NZ-1, высоко реактивное анти-PDPN-антитело,нейтрализовало накопление тромбоцитов LN319, позволяя предположить, что PDPN является основной причиной для накопления тромбоцитов, индуцированного (Kato et al., 2006). Уровни экспрессии геновPDPN были значительно выше в глиобластомах, чем в не-неопластическом белом веществе, что было подтверждено иммуногистохимией (Scrideli et al., 2008). PDPN специфически экспрессирован лимфатическими, но не эндотелиальными клетками кровеносных сосудов в культуре и при опухольассоциированном лимфангиогенезе. Хотя PDPN первоначально отсутствовал в нормальном человеческом эпидермисе, его экспрессия была сильно индуцирована в 22 из 28 сквамозных карцином, позволяя предположить, что PDPN играет роль при прогрессии опухоли (Schacht et al., 2005). PDPN находится в повышенном количестве на инвазивном фронте многих раковых заболеваний человека. Исследование экспрессии PDPN на культивированных клетках человеческого рака молочной железы, на модели с мышами с панкреатическим бета-клеточным канцерогенезом и биоптатах рака человека указывают на то,что PDPN способствует инвазии опухолевых клеток in vitro и in vivo. PDPN индуцирует коллективную клеточную миграцию при образовании филоподий посредством снижения активности малых GTP-аз семейства Rho. В заключение, PDPN индуцирует альтернативный путь инвазии опухолевых клеток при отсутствии эпителиально-мезенхимального перехода (Wicki et al., 2006); уровень экспрессии PDPN был повышен у большинства пациентов с колоректальным раком (Kato et al., 2003); предполагается, что TGFбета является физиологическим регулятором PDPN в опухолевых клетках (Suzuki et al., 2008); PDPN экспрессируется раковыми клетками, происходящими из пищевода, легких, печени, толстого кишечника и молочной железы, а также лимфатических эндотелиальных клеток (Kono et al., 2007). Тенасцин С (гексабрахион) (TNC). Экспрессия внеклеточного матричного гликопротеина TNC в глиобластоме, но не в нормальном головном мозге, и его ассоциация с пролиферативными эндотелиальными мембранами основания при глиобластоме позволили предположить уже в 1983 г., что TNC может быть полезным маркером для глиом(Bourdon et al., 1983). Во время прогрессии опухоли ЕСМ опухолевых тканей переделывается, обеспечивая теперь окружение, которое более благоприятно для прогрессии опухоли, для которой ангиогенез является ключевым этапом (Carnemolla et al., 1999). TNC гиперэкспрессируется в сосудах опухоли, которые имеют высокий пролиферативный индекс, указывающий на то, что TNC задействован в неопластическом ангиогенезе (Kim et al., 2000). Экспрессия TNC в опухолях может быть вызвана гипоксией (Lal et al.,2001). Индукция TNC опосредована TGF-бета 1, обеспечивая глиомам высокой степени злокачественности механизм для инвазии в здоровую паренхиму (Hau et al., 2006). Также гиперэкспрессия TNC является последствием специфической активации промотора гена тенасцина-С гастрином, который, как известно,значительно модулирует миграцию клеток человеческой глиобластомы (Kucharczak et al., 2001). TNC снижает количество тропомиозина-1 и, таким образом, дестабилизирует стрессовые волокна актина. Это вызывает снижение количества ингибитора сигнального Wnt-каскада, "Dickkopfl". Так как снижение экспрессии тропомиозина-1 и возрастание Wnt-сигналинга тесно связаны с трансформацией и онкогенезом,то TNC специфически модулирует данные сигнальные пути, усиливая пролиферацию клеток глиомы(Ruiz et al., 2004). Периваскулярное окрашивание TNC вокруг кровеносных сосудов, снабжающих опухоль, наблюдалось в тканях глиобластомы, причем периваскулярное окрашивание TNC встречается менее часто в глиомах II и III степени по ВОЗ, указывая на то, что интенсивность окрашивания TNC соотносится со степенью опухоли, а наиболее сильное окрашивание указывает на плохой прогноз (Herold-Mende et al.,2002; Zukiel et al., 2006). Наиболее высокая экспрессия TNC наблюдается в соединительной ткани, окружающей опухоли (Chiquet-Ehrismann and Tucker, 2004). TNC также вносит свой вклад в генерацию ниши стволовых клеток внутри субвентрикулярной зоны (SVZ), принимая участие в управлении сигналами фактора роста для ускорения развития нервных стволовых клеток. TNC необходим для временной экспрессии EGFR в стволовые клетки нервной системы, и он усиливает сигналинг FGF2. Доминирующим эффектом TNC на клетках SVZ-зоны является регуляция процессов, связанных с прогрессией (Garcion etal., 2004). TNC является сильнейшим инициатором направленной миграции NSC человека (нервных стволовых клеток) (гаптотаксис). Вырабатываемый опухолью ЕСМ, таким образом, обеспечивает терпимое окружение для тропизма NSC к рассеянным опухолевым клеткам (Ziu et al., 2006).- 23023013 Путь TNC играет также важную роль в росте опухоли молочной железы и метастазах. Таким образом, блокада сигналинга или резкое снижение TNC в клетках MDA-MB-435 привела к значительному ухудшению клеточной миграции и субстрат-независимой клеточной пролиферации. Мыши, инъецированные клональными клетками MDA-MB-435 со сниженной экспрессией TNC, демонстрировали значительное снижение роста первичной опухоли, в равной степени как и снижение рецидивов после хирургического удаления первичной опухоли и снижение частоты метастазов в легких (Calvo et al., 2008). Сурвивин (BIRC5). Экспрессия BIRC5 (сурвивин), члена семейства белков-ингибиторов апоптоза (IAP), содержится в повышенном количестве в фетальных тканях и различных видах рака человека и со значительно сниженной экспрессией в нормальных взрослых дифференцированных тканях, в особенности, если у них низкий индекс пролиферации. Предполагается, что сурвивин способен к регуляции как клеточной пролиферации, так и отмирания апоптических клеток. Хотя сурвивин обычно находится на цитоплазматическом участке клетки и ассоциируется с плохим прогнозом при раке, также сообщалось о ядерной локализации,указывающей на благоприятный прогноз (O'Driscoll et al., 2003). Регуляцию сурвивина и с его помощью описывают несколькими механизмами. Предполагается, что сурвивин ассоциирован с молекулярным шапероном Hsp60. In vivo Hsp60 с избытком экспрессирован в первичных опухолях человека по сравнению с соответствующими нормальными тканями. Экстренная абляция Hsp60 небольшой интерферирующей РНК дестабилизирует митохондриальный пул сурвивина, вызывает митохондриальную дисфункцию и активирует каспаза-зависимый апоптоз (Ghosh et al., 2008). Кроме того, ингибирование Ras приводит к снятию сурвивинового "тормоза" на апоптозе и активации митохондриального апоптотического каскада реакций. В особенности в глиобластоме устойчивость к апоптозу может быть ликвидирована посредством Ras-ингибитора, мишенью которого является сурвивин (Blum et al., 2006). Вероятно,существует взаимосвязь между NF-kappaB гиперактивностью в глиомах и гиперэкспрессией сурвивина,одного из генов-мишеней NF-kappaB. Таким образом, активированные NF-kappaB анти-апоптотические гены гиперэкспрессированы в пробах опухолей. В особенности, в глиобластоме обнаруживаются очень высокие уровни экспрессии сурвивина (Angileri et al., 2008). Предполагается, что гиперэкспрессия сурвивина в глиомах головного мозга может играть важную роль в злокачественной пролиферации, антиапоптозе и ангиогенезе (Zhen et al., 2005; Liu et al., 2006b). Было произведено несколько анализов для изучения экспрессии сурвивина и его влияния на выживаемость при глиобластоме. В качестве обобщения, экспрессия сурвивина, в особенности одновременная экспрессия в ядре и цитоплазме в астроцитных опухолях была значительно ассоциирована со степенью злокачественности (с наибольшей экспрессией сурвивина в глиобластоме) и более короткими общими сроками выживаемости по сравнению с пациентами, у которых были сурвивин-негативные опухоли (Kajiwara et al., 2003; Saito et al., 2007; Uematsu etal., 2005; Mellai et al., 2008; Grunda et al., 2006; Xie et al., 2006; Sasaki et al., 2002; Chakravarti et al., 2002). Гиперэкспрессия сурвивина описывалась также для других опухолевых форм. При раке молочной железы гиперэкспрессия сурвивина ассоциирована с более высокой степенью злокачественности и более коротким сроком выживаемости без заболевания (Yamashita et al., 2007; Al-Joudi et al., 2007; Span et al.,2004). Как было зафиксировано, в клеточных линиях рака пищевода промоторная активность сурвивина была в 28,5 раз выше, чем в нормальных тканях (Sato et al., 2006). При колоректальном раке экспрессия сурвивина также ассоциирована с патологической степенью и метастазами лимфатических узлов (Tan etal., 2005). Как было показано, агрессивность светлоклеточной почечной карциномы ассоциирована с экспрессией сурвивина. Кроме того, экспрессия сурвивина обратно пропорционально ассоциирована со специфичной для рака выживаемостью (Kosari et al., 2005). Экспрессия сурвивина может быть обнаружена в наборе неоплазмов с участием кератиноцитов и гиперпролиферативных поражениях кожи, но не в нормальной коже (Bowen et al., 2004). В клеточных линиях рака поджелудочной железы сурвивин был амплифицирован в 58% проанализированных клеточных линий (Mahlamaki et al., 2002). При плоскоклеточной карциноме экспрессия сурвивина может помочь идентифицировать случаи с более агрессивным и инвазивным клиническим фенотипом (Lo et al., 2001). Так как сурвивин является такой многообещающей мишенью при лечении рака, то исследования с использованием образованных из сурвивина пептидов показали, что сурвивин является иммуногенным для пациентов с опухолями, вызывая ответы, опосредованные CD8+ Т-клетками. В дополнение к этому,сурвивин специфически стимулировал CD4+ Т-клеточную реактивность лимфоцитов в периферической крови у тех же пациентов (Casati et al., 2003; Piesche et al., 2007). Сурвивин (SVN, BIRC) гиперэкспрессирован во множестве раковых форм. Итак, гиперэкспрессия сурвивина в целом, как считается, ассоциирована с более коротким общим сроком выживаемости и более высокими степенями злокачественности. Настоящее изобретение в дальнейшем относится к конкретным белкам-маркерам, которые могут быть использованы для прогнозирования глиобластомы. Кроме того, настоящее изобретение относится к применению этих новых мишеней для лечения рака. В соответствии с приводимой здесь информацией белки GFAP, FABP7, DTNA, NR2E1, SLCO1C1, CHI3L1, ACSBG1, IGF2BP3, NLGN4X, MLC1, NRCAM,BCAN, EGFR, PDPN, NES и CLIP2 сильно гиперэкспрессированы в глиобластомах по сравнению с нормальным головным мозгом и другими жизненно важными тканями (к примеру, печень, почка, сердце).- 24023013 Белки GRP56, CSPG4, NRCAM, TNC, BIRC5, CLIP2, NES, PDPN, EGFR, BCAN, GRIA4, как продемонстрировано, играют важную роль в онкогенезе, так как они задействованы в злокачественной трансформации, клеточном росте, пролиферации, ангиогенезе или инвазии нормальной ткани. Белки NES, TNC, BIRC5, EGFR ассоциированы с раковыми стволовыми клетками или нишами стволовых клеток при глиобластоме. Раковые стволовые клетки являются субпопуляцией раковых клеток с потенциалом для самообновления, необходимым для постоянного роста опухоли. Эти клетки находятся в специализированных и высоко организованных структурах, так называемых нишах раковых стволовых клеток, которые требуются для сохранения потенциала самообновления раковых стволовых клеток. Гиперэкспрессия белков BIRC5, NRCAM, IGF2BP3 в опухолях, как было продемонстрировано,ассоциирована с поздними стадиями заболевания и неутешительным прогнозом для пациентов. ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN, как было продемонстрировано, играют важную роль в ремоделировании ткани, требуемом для роста опухоли в нервной системе. Поэтому экспрессия ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN может быть использована в качестве маркера, чтобы отличить глиобластому от других форм рака. Таким образом, настоящее изобретение обеспечивает способы идентификации животного, предпочтительно человека, у которого, вероятно, имеется глиобластома. В одном воплощении установленная вероятность составляет от 80 до 100%. Один таковой способ включает определение уровня по крайней мере одного из белков ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN в опухолевой пробе животного субъекта. В одном воплощении проба изымается методом радикальной хирургии. В другом воплощении проба изымается методом игольной биопсии. Если уровень ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN, как установлено, выше на 20% или более в клетках относительно уровня, установленного в доброкачественных эпителиальных клетках того же самого образца, то у животного субъекта, вероятно, имеется глиобластома. Чем выше уровень различным белков группы, включающей ВСА, CLIP2, DTNA, NLGNAX, NR2E1,NRCAM и PDPN, чем выше вероятность того, что будет установлено, что у животного субъекта имеется глиобластома. В одном воплощении определение уровня ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM илиPDPN производится in situ. В другом воплощении определение уровня ВСА, CLIP2, DTNA, NLGNAX,NR2E1, NRCAM или PDPN производится in vitro. В другом отличном воплощении определение уровня ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN производится in vivo. В предпочтительном воплощении определение уровня ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN производится методом лазерной захватывающей микроскопии вместе с анализом Western blot. В одном конкретном воплощении определение уровня ВСА, CLIP2, DTNA, NLGNAX, NR2E1,NRCAM или PDPN производится с помощью антитела, специфического для ВСА, CLIP2, DTNA,NLGNAX, NR2E1, NRCAM или PDPN. В другом воплощении определение уровня ВСА, CLIP2, DTNA,NLGNAX, NR2E1, NRCAM или PDPN производится методом ПЦР с праймером, специфическим для мРНК, кодирующей ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN. В другом отличном воплощении определение уровня ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN производится методом с нуклеотидным зондом, специфическим для мРНК, кодирующей ВСА, CLIP2, DTNA,NLGNAX, NR2E1, NRCAM или PDPN. В одном воплощении определение уровня ВСА, CLIP2, DTNA,NLGNAX, NR2E1, NRCAM или PDPN производится с использованием анализа Northern blot. В другом воплощении определение уровня ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN выполняется методом анализа с помощью защиты от рибонуклеазы. В других воплощениях могут быть использованы иммунологические тесты, такие как методы иммуноферментных зон (ELISA), радиоиммунные методы (RIA) и методы иммунного блоттинга (Western blot) для определения полипептидов ВСА, CLIP2,DTNA, NLGNAX, NR2E1, NRCAM и PDPN в жидкой пробе организма (такой как кровь, сыворотка, мокрота, моча или перитонеальная жидкость). Биоптаты, тканевые пробы и клеточные пробы (такие как из яичника, лимфатических узлов, соскобы эпителиальных клеток с поверхности яичника, биоптаты легкого, биоптаты печени и любые жидкостные пробы, содержащие клетки (такие как перитонеальная жидкость, мокрота и плейральный секрет), могут быть проанализированы посредством дезагрегации и/или растворения ткани или клеточного образца и проведения иммуноанализа на обнаружение полипептида,такого как ELISA, RIA или иммунного блоттинга (Western blotting). Такие клеточные или тканевые пробы могут также быть проанализированы методами, основанными на нуклеиновых кислотах, к примеру,методом полимеразной цепной реакции с обратной транскрипцией (RT-PCR) с амплификацией, гибридизации Нозерн-блоттинг или слот- или дот-блоттинг. Для визуализации распределения опухолевых клеток в тканевых образцах в целях обнаружения полипептидных маркеров глиобластомы или мРНК, соответственно, могут быть применены диагностические тесты, которые сохраняют структуру ткани, например иммуногистологическое окрашивание, in situ гибридизация РНК или in situ RT-PCR. Для локализации опухолевой массы in vivo могут применяться методы визуализации, такие как магнитно-резонансная томография (МРТ) посредством введения в субъект антитела, которое специфически связывается с полипептидом ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN (в особенности полипептидом, находящимся на клеточной поверхности), где антитело конъюгировано или по-другому связано с парамаг- 25023013 нитным индикатором (или другой подходящей детектируемой единицей в зависимости от используемого метода визуализации); альтернативно локализацию не меченого опухолевого маркер-специфического антитела можно определить с помощью вторичного антитела, связанного с детектируемой единицей. В дополнение настоящее изобретение в дальнейшем обеспечивает химерные/белки слияния/пептиды, включающие полипептиды ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и/или PDPN и их фрагменты, включительно функциональные, протеолитические и атигенные фрагменты. Партнеры по слиянию или части гибридной молекулы адекватно обеспечивают эпитопы, которые стимулируют CD4+ Т-клетки. Стимулирующие эпитопы CD4+ хорошо известны из уровня техники и включают те, что были выявлены в столбнячном токсине. В дальнейшем предпочтительном воплощении пептид является рекомбинантным белком, в частности включающим N-терминальные аминокислоты антиген-ассоциированной инвариантной цепи (Ii) HLA-DR. В одном воплощении пептид по изобретению является укороченным белком человека или гибридным белком белкового фрагмента и другого сегмента полипептида, при условии, что человеческий сегмент включает одну или более аминокислотную последовательность изобретения. Антитела к полипептидам ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN, к химерным/белкам слияния, включающим полипептиды ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM илиPDPN, в равной степени как и фрагменты полипептидов ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN, включая протеолитические и антигенные фрагменты, и к химерным/белкам слияния/пептидам, включая эти фрагменты, являются также частью настоящего изобретения. Помимо этого,методы использования таких антител для прогнозирования рака и глиобластомы в частности также являются частью настоящего изобретения. Антитела по настоящему изобретению могут быть поликлональными антителами, моноклональными антителами и/или химерными антителами. Бессмертные линии клеток, которые вырабатывают моноклональное антитело по настоящему изобретению, являются также частью настоящего изобретения. Обычному специалисту данной области будет понятно, что в некоторых частных случаях более высокая экспрессия ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN в качестве опухолевого гена-маркера будет указывать на худший прогноз для субъекта, имеющего глиобластому. Например, относительно более высокие уровни экспрессии ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM илиPDPN могут указывать на относительно крупную первичную опухоль, более высокую тяжесть заболевания (к примеру, больше метастазов) или относительно более злокачественный фенотип опухоли. Чем более различна гиперэкспрессия различных белков группы, включающей ВСА, CLIP2, DTNA,NLGNAX, NR2E1, NRCAM и PDPN, тем хуже прогноз для пациента. Диагностические и прогностические методы по изобретению охватывают использование известных методов, к примеру, основанных на антителах методов детекции полипептидов ВСА, CLIP2, DTNA,NLGNAX, NR2E1, NRCAM и PDPN, а также методов, основанных на гибридизации и/или амплификации нуклеиновых кислот для детекции ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и/или PDPN мРНК. Помимо этого, в связи с тем, что быстрое уничтожение опухолевых клеток часто приводит к образованию авто-антитела, опухолевые маркеры глиобластомы по изобретению могут использоваться в серологических анализах (например, в тесте ELISA для сыворотки субъекта) для обнаружения автоантител против ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN у субъекта. Уровни полипептид-специфического авто-антитела для ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN, которые по крайней мере приблизительно в 3 раза выше (и предпочтительно по крайней мере в 5 раз или 7 раз выше, наиболее предпочтительно по крайней мере в 10 или 20 раз выше), чем в контрольной пробе,являются признаком глиобластомы. Находящиеся на поверхности клетки внутриклеточные и секретируемые полипептиды BCA, CLIP2,DTNA, NLGNAX, NR2E1, NRCAM и PDPN могут быть также использованы в анализе биоптатов, например, ткани или клеточных проб (включая клетки, полученные из жидкостных проб, таких как полосная перитонеальная жидкость), для определения, содержит ли тканевый или клеточный биоптат клетки глиобластомы. Биоптат может быть проанализирован как интактная ткань или проба с целыми клетки,или же ткань или клеточная проба могут быть дисагрегированы и/или растворены, как то необходимо для конкретного вида используемого диагностического анализа. Например, биоптаты или пробы могут подвергаться цельнотканевому или цельноклеточному анализу для определения уровня полипептида или мРНК ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN in situ, например, используя иммуногистохимический метод, гибридизацию мРНК in situ или ПЦР с обратной транскрипцией in situ. Опытному специалисту известно, как приготавливать ткани или клетки к анализу для определения уровня полипептида или мРНК с использованием иммуногистохимических методов, таких как ELISA, иммунный блоттинг или равноценные методы, или же методов анализа уровня мРНК посредством аналитических методов, основанных на нуклеиновых кислотах, таких как ПЦР с обратной транскрипцией, нозернгибридизация или слот- или дот-блоттинг.- 26023013 Оборудование для измерения уровней экспрессии ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM или PDPN. Настоящим изобретением обеспечивается оборудование для обнаружения повышенного уровня экспрессии ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN в качестве гена-маркера глиобластомы у субъекта. Оборудование для обнаружения полипептидного маркера глиобластомы предпочтительно содержит антитело, которое специфически связывается с выбранным полипептидным маркером глиобластомы. Оборудование для обнаружения мРНК-маркера глиобластомы предпочтительно содержит одну или более нуклеиновых кислот (к примеру, один или более олигонуклеотидный праймер или зонд,зонды ДНК, зонды РНК или шаблоны для получения зондов РНК), которая специфически гибридизируется с мРНК белков ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN. Оборудование, основанное на антителах, в особенности может использоваться для обнаружения присутствия и/или измерения уровня полипептида ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и/или PDPN, который специфически связан с антителом или его иммунореактивным фрагментом. Оборудование может включать антитело,реактивное по отношению к антигену, и реагент для обнаружения реакции антитела с антигеном. Такое оборудование может быть оборудованием ELISA и может содержать контроль (например, специфицированное количество конкретного полипептидного маркера глиобластомы), первичные и вторичные антитела, когда это целесообразно, и любые другие необходимые реагенты, такие как детектируемые составляющие, субстраты ферментов и цветные реагенты, как описывалось выше. Диагностическое оборудование альтернативно может быть оборудованием для иммуноблоттинга, обычно включающим компоненты и реагенты, описываемые здесь. Оборудование, основанное на нуклеиновой кислоте, может использоваться для обнаружения и/или измерения уровня экспрессии ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN посредством обнаружения и/или измерения количества мРНК белков ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM иPDPN в пробе, такой как тканевый или клеточный биоптат. Например, оборудование для ПЦР с обратной транскрипцией для обнаружения повышенной экспрессии ВСА, CLIP2, DTNA, NLGNAX, NR2E1,NRCAM и PDPN предпочтительно содержит олигонуклеотидные праймеры, которых достаточно для проведения обратной транскрипции мРНК-маркера глиобластомы до кДНК и амплификацию кДНКмаркера глиобластомы посредством ПЦР, и будет также предпочтительно содержать контрольные шаблонные молекулы ПЦР и праймеры для проведения адекватного негативного и положительного контроля, а также внутреннего контроля для квантизации. Обычный специалист данной области в состоянии отобрать подходящие праймеры для проведения реакций обратной транскрипции и ПЦР, а также адекватные контрольные реакции, которые должны быть выполнены. Такое руководство приводится, например, в работе F. Ausubel et al., Current Protocols in Molecular Biology, John WileySons, New York, N.Y.,1997. Многочисленные разновидности ПЦР с обратной транскрипцией известны из уровня техники. Нацеленная доставка иммунотоксинов к ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN может осуществляться в качестве терапевтических мишеней для лечения или предупреждения глиобластомы. Например, молекула антитела, которая специфически связывается с находящимся на поверхности клетки полипептидом ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN, может быть конъюгирована с радиоизотопом или другим токсическим соединением. Конъюгаты антител вводятся в субъекта, так что связывание антитела с родственным ему полипептидом глиобластомы приводит к нацеленной доставке терапевтического соединения в клетки глиобластомы, тем самым производя лечение рака яичника. Терапевтическим составляющим может быть токсин, радиоизотоп, лекарственное, химическое средство или белок (см., например, Bera et al. "Pharmacokinetics and antitumor activity of a bivalent disulfide-stabilized Fv immunotoxin with improved antigen binding to erbB2" Cancer Res. 59:4018-4022 (1999. Например, антитело может быть связано или конъюгировано с бактериальным токсином (например, токсин дифтерии, экзотоксин псевдомонас А, холерный токсин) или растительным токсином (например,рициновый токсин) для нацеленной доставки токсина к клетке, экспрессирующей ВСА, CLIP2, DTNA,NLGNAX, NR2E1, NRCAM и PDPN. Этот иммунотоксин может быть доставлен к клетке, и при соединении с находящимся на поверхности клетки полипептидным маркером глиобластомы токсин, конъюгированный с антителом, специфическим для маркера глиобластомы, будет доставлен к клетке. Еще один другой аспект настоящего изобретения относится к антителу, которое специфически связывается с главным комплексом гистосовместимости человека (МНС) I или II класса в комплексе с HLAрестриктированным антигеном (именуемое в дальнейшем также "комплекс-специфическое антитело"). Еще один другой аспект настоящего изобретения относится, затем, к методу получения указанного антитела, специфически связывающегося с главным комплексом гистосовместимости человека (МНС) I илиII класса в комплексе с HLA-рестриктированным антигеном, метод, включающий иммунизацию полученного с помощью генной инженерии нечеловеческого млекопитающего, включающего клетки, экспрессирующие указанный главный комплекс гистосовместимости человека (МНС) I или II класса с растворимой формой молекулы МНС I или II класса в комплексе с указанным HLA-рестриктированным антигеном; изолирование молекул мРНК из антитела, вырабатывающего клетки указанного нечеловеческого млекопитающего; получение библиотеки фагового отображения, отображающей белковые молекулы,закодированные указанными молекулами мРНК; и изолирование по крайней мере одного фага из указан- 27023013 ной библиотеки фагового отображения, указанный по крайней мере один фаг, отображающий указанное антитело, специфически связываемое с указанным главным комплексом гистосовместимости человека(МНС) I или II класса, в комплексе с указанным HLA-рестриктированным антигеном. Соответствующие методы для получения таких антител и одноцепочных главных комплексов гистосовместимости I класса,в равной степени как и другие инструменты для получения данных антител раскрыты в WO 03/068201,WO 2004/084798, WO 01/72768, WO 03/070752 и у Cohen C.J., Denkberg G., Lev A., Epel M., Reiter Y. Recombinant antibodies with MHC-restricted, peptide-specific, T-cell receptor-like specificity: new tools to studyApr 15; 170(8):4349-61, которые в целях настоящего изобретения в открытой форме включены сюда в своей целостности путем ссылки. Предпочтительно, если антитело связывается с аффинностью связывания ниже 20 нмоль, предпочтительно ниже 10 нмоль, с комплексом, который называется "специфическим" в контексте настоящего изобретения. Термин "антитело" используется здесь в широком смысле и включает, как поликлональные, так и моноклональные антитела. В дополнение к интактным молекулам иммуноглобулина в термин "антитела" включены также фрагменты или полимеры таких молекул иммуноглобулина и гуманизированные версии молекул иммуноглобулина, при условии что они проявляют одно из желаемых свойств (например, являются комплекс-специфическим антителом, как упомянуто выше, доставляют токсин к раковой клетке,экспрессирующей раковый, предпочтительно ген-маркер глиобластомы, на повышенном уровне и/или ингибируют активность ракового полипептида-маркера, такого как сурвивин), описанных здесь. Помимо этого, для любого полипептида ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM и PDPN,для которого существует специфический лиганд (например, лиганд, который связывается с находящимся на поверхности клетки белком), лиганд может быть использован вместо антитела для нацеливания токсичного соединения на клетку глиобластомы, как описывалось выше. Если возможно, антитела по изобретению могут быть куплены в коммерческих источниках. Антитела по изобретению могут быть также генерированы при использовании хорошо известных методов. Специалисту будет понятно, что для генерирования антител по изобретению могут использоваться либо полипептиды-маркеры глиобластомы полной длины, либо их фрагменты. Полипептид, необходимый для генерирования антитела по изобретению, может быть частично или полностью очищенным из естественного источника или же может быть получен с помощью техники рекомбинантной ДНК. Например,кДНК, кодирующая полипептид ВСА, CLIP2, DTNA, NLGNAX, NR2E1, NRCAM or PDPN или его фрагмент, может быть экспрессирована в прокариотические клетки (например, бактерии) или эукариотические клетки (например, клетки дрожжей, насекомых или млекопитающих), после чего рекомбинантный белок может быть очищен и использован для получения препарата из моноклонального или поликлонального антитела, которое специфически связывается с полипептидом-маркером глиобластомы, использованным для получения антитела. Специалисту данной области будет известно, что получение двух или более различных панелей моноклональных или поликлональных антител увеличивает вероятность получения антитела со специфичностью и аффинностью, необходимыми для предназначенного для него использования (например,ELISA, иммуногистохимия, визуализация in vivo, терапия на основе иммунотоксина). Антитела протестированы на желаемую для них активность с помощью известных методов в соответствии с целью использования антител (например, ELISA, иммуногистохимия, иммунотерапия и т.д.; для получения дальнейшей информации по генерированию и испытанию антител см., например, Harlow and Lane,Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1988). Например, антитела могут быть протестированы с помощью анализов ELISA, метода иммунного блоттинга(Western-blot), иммуногистохимического окрашивания законсервированных формальдегидом образцов глиобластомы или замороженных тканевых срезов. После их первоначальной характеристики in vitro антитела, предназначаемые для терапевтического или диагностического использования in vivo тестируются в соответствии с известными клиническими методами анализа. Термин "моноклональное антитело", используемый здесь, обозначает антитело, полученное из, по существу, гомогенной популяции антител, т.е. отдельные антитела внутри популяции идентичны за исключением возможных естественных мутаций, которые могут быть представлены в небольших количествах. Моноклональные антитела здесь специфически включают "химерные" антитела, в которых участок тяжелой и/или легкой цепи идентичен или гомологичен с соответствующими последовательностями антител, образованных из конкретных видов или относящихся к конкретному классу или подклассу антител, в то время как остальная(ые) цепь(и) идентична с или гомологична с соответствующими последовательностями антител, образованных из других видов или относящихся к другому классу или подклассу- 28023013 антител, в равной степени как и фрагменты таких антител, пока они проявляют желаемую антагонистическую активность (патент США 4816567). Моноклональные антитела по изобретению могут быть приготовлены при использовании гибридомного метода. В рамках гибридомного метода мышь или другое подходящее животное-хозяин обычно иммунизируется иммунизирующим веществом, чтобы инициировать лимфоциты, которые вырабатывают или способны вырабатывать антитела, которые будут специфически связываться с иммунизирующим веществом. Альтернативно лимфоциты могут быть иммунизированы in vitro. Моноклональные антитела могут быть также получены с помощью методов с рекомбинантной ДНК, таких как описываемые в патенте США 4816567. ДНК, кодирующая моноклональные антитела по изобретению, может быть без труда изолирована и разделена на последовательности с помощью стандартной методики (например, при использовании олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител).In vitro-методы также подходят для приготовления моновалентных антител. Расщепление антител для получения их фрагментов, в особенности Fab-фрагментов, может быть произведено при использовании стандартных техник, известных из уровня техники. К примеру, расщепление может производиться при использовании папаина. Примеры расщепления под воздействием папаина описываются в заявкеWO 94/29348, опубликованной 22.12.1994 г., и в патенте США 4342566. Расщепление антител под воздействием папаина обычно приводит к двум идентичным фрагментам, связывающимся с антигеном,называемым Fab-фрагментами, каждый из которых имеет отдельный антиген-связывающий сайт и остаточный Fe-фрагмент. В результате воздействия пепсином получается фрагмент, который имеет два антиген-комбинирующих сайта и все еще способен к кросс-линкингу антигена. Фрагменты антитела как связанные с другими последовательностями, так и не связанные, могут также включать вставки, делеции, замещения или другие избранные модификации конкретных участков или специфических аминокислотных остатков при условии, что активность фрагмента незначительно изменена или повреждена по сравнению с немодифицированным антителом или фрагментом антитела. Данные модификации могут обеспечивать некоторые дополнительные свойства, такие как добавление/удаление аминокислот, способных к дисульфидному связыванию, увеличение их биостойкости, изменение их секреторных характеристик и т.д. В любом случае фрагмент антитела должен обладать свойством биоактивности, таким как активностью связывания, регуляцией связывания на связывающем домене и т.д. Функциональные или активные участки антитела могут быть идентифицированы при мутагенезе специфического участка белка с последующей экспрессией и контролем экспрессированного полипептида. Такие методы полностью очевидны для опытного специалиста данной области техники и могут включать сайт-специфический мутагенез нуклеиновой кислоты, кодирующей фрагмент антитела. Антитела по изобретению могут в дальнейшем включать гуманизированные антитела или человеческие антитела. Гуманизированные формы нечеловеческих (например, мышиных) антител - это химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab' или другие антиген-связывающие субпоследовательности антител), которые содержат минимальную последовательность, полученную из нечеловеческого иммуноглобулина. Гуманизированные антитела включают человеческие иммуноглобулины (антитело-реципиент), в которых остатки из определяющего комплементарного участка (CDR) реципиента замещены остатками из CDR нечеловеческих видов (донорское антитело), таких как мыши, крысы или кролики, имеющими желаемую специфичность, аффинность и емкость. В некоторых случаях остатки Fv-каркаса (FR) человеческого иммуноглобулина замещены соответствующими нечеловеческими остатками. Гуманизированные антитела могут также включать остатки, которые не встречаются ни в антителе-реципиенте, ни в импортированном CDR или каркасных последовательностях. В целом, гуманизированное антитело будет включать, по существу, все из по крайней мере одного и, типично, двух вариабельных доменов, в которых все или по существу все участки CDR соответствуют таковым из нечеловеческого иммуноглобулина, и все или по существу все из участков FR являются таковыми консенсусной последовательности человеческого иммуноглобулина. Гуманизированное антитело оптимально также будет содержать по крайней мере один сегмент константного участка иммуноглобулина (Fc), типично - человеческого иммуноглобулина. Методы гуманизации нечеловеческих антител хорошо известны из уровня техники. В целом, гуманизированное антитело имеет один или более аминокислотный остаток, введенный в него из источника,являющегося нечеловеческим. Такие нечеловеческие аминокислотные остатки называются часто "импортированными" остатками, которые обычно берутся из "импортированного" вариабельного домена. Гуманизация может быть произведена в обязательном порядке посредством замены участков CDR или последовательностей CDR грызунов на соответствующие последовательности человеческого антитела. Соответственно, такие "гуманизированные" антитела являются химерными антителами (патент США 4816567), где в обязательном порядке менее чем один интактный человеческий вариабельный домен был заменен соответствующей последовательностью нечеловеческого вида. На практике гуманизированные антитела являются типично человеческими антителами, в которых некоторые остатки CDR и, возможно,остатки FR заменены на остатки аналогичных сайтов антител грызунов. Использоваться могут трансгенные животные (например, мыши), которые способны при иммуниза- 29
МПК / Метки
МПК: A61K 39/00, A61P 35/00, A61K 38/16, A61K 38/08
Метки: применение, индуцирующий, иммунный, ответ, противоопухолевый, пептид
Код ссылки
<a href="https://eas.patents.su/30-23013-peptid-induciruyushhijj-protivoopuholevyjj-immunnyjj-otvet-i-ego-primenenie.html" rel="bookmark" title="База патентов Евразийского Союза">Пептид, индуцирующий противоопухолевый иммунный ответ, и его применение</a>
Глюкагоноподобный пептид-1, улучшающий ответ &beta-клеток на глюкозу у субъектов с нарушением толерантности к глюкозе

Номер патента: 4538
Опубликовано: 24.06.2004
Авторы: Бирне Мария, Куллидж Томас Р., Гоке Буркхард
МПК: A61K 38/26
Метки: ответ, субъектов, глюкозе, глюкозу, глюкагоноподобный, пептид-1, нарушением, beta-клеток, толерантности, улучшающий
Формула / Реферат:
1. Способ лечения человека с нарушенной толерантностью к глюкозе до наступления инсулиннезависимого сахарного диабета, предусматривающий введение указанному человеку композиции, содержащей соединение, которое связывается с рецептором глюкагоноподобного пептида-1, и фармацевтический носитель, причем указанная композиция содержит указанный компонент в количестве, эффективном для увеличения чувствительности и ответа b-клеток поджелудочной железы на...
Препарат, потенцирующий противоопухолевый эффект, противоопухолевый препарат и способ лечения рака

Номер патента: 11573
Опубликовано: 28.04.2009
Авторы: Утида Дзундзи, Такети Теидзи, Коизуми Кацухиса, Нукацука Мамору
МПК: A61K 31/506, A61K 31/44, A61K 31/132...
Метки: способ, эффект, препарат, лечения, противоопухолевый, рака, потенцирующий
Формула / Реферат:
1. Препарат, потенцирующий противоопухолевый эффект, для потенцирования противоопухолевой активности противоопухолевого препарата, включающего в себя тегафур в терапевтически эффективном количестве, гимерацил в количестве, эффективном для потенцирования противоопухолевого эффекта, и отерацил калия в количестве, эффективном для ингибирования побочного эффекта, где препарат, потенцирующий противоопухолевый эффект, включает...
Агент, индуцирующий апоптоз, способ его получения и пищевой продукт, содержащий его

Номер патента: 2024
Опубликовано: 24.12.2001
Авторы: Като Икуносин, Икаи Катсусиге, Ю Фу-Гонг, Татсуми Йоко, Сагава Хироаки, Сакаи Такеси, Кояма Нобуто
МПК: A61P 43/00, A61K 35/78
Метки: продукт, пищевой, способ, агент, индуцирующий, получения, апоптоз, содержащий
Формула / Реферат:
1. Агент, индуцирующий апоптоз, который содержит в качестве эффективного компонента глицеролипид и/или глицерогликолипид, свободный в своей молекулярной структуре от фосфатного эфира и фосфонатного эфира. 2. Агент по п.1, в котором вышеупомянутый глицеролипид и/или глицерогликолипид происходит из природных источников, например растений, микроорганизмов или животных. 3. Агент по п.2, в котором вышеупомянутыми природными источниками являются...
Способ, вызывающий т-клеточный ответ

Номер патента: 12066
Опубликовано: 28.08.2009
Авторы: Браун Ральф Патрик, Донг Личун
МПК: A61K 48/00, A61K 39/00, A61K 31/70...
Метки: ответ, т-клеточный, вызывающий, способ
Формула / Реферат:
1. Способ, вызывающий T-клеточный ответ против T-клеточного эпитопа у субъекта млекопитающего-хозяина, при этом указанный способ включает: (i) первую иммунизацию, которая состоит по меньшей мере из двух введений, которые осуществляют субъекту с интервалом от 1 до 14 дней, при этом каждое введение заключается во введении представляющей интерес нуклеотидной последовательности (NOI), кодирующей T-клеточный эпитоп, и необязательно (ii) вторую...
Способ прогнозирования резерва яичника в ответ на индукцию овуляции и набор для его осуществления

Номер патента: 12444
Опубликовано: 30.10.2009
Авторы: Сейфер Дэвид Б., Маклафлин Дэвид Т.
МПК: C07K 16/00, A61K 39/395, C12N 5/06...
Метки: яичника, прогнозирования, резерва, ответ, способ, индукцию, овуляции, набор, осуществления
Формула / Реферат:
1. Способ прогнозирования резерва яичника в ответ на индукцию овуляции у субъекта, который нуждается в индукции овуляции, включающий: (a) определение уровня Мюллеровой ингибирующей субстанции (MIS) в сыворотке у субъекта; (b) сравнение определенного сывороточного уровня MIS со стандартом, в котором коррелируют уровень MIS в сыворотке с числом ооцитов, полученных после индукции овуляции; и (c) прогнозирование ожидаемого ответа яичника. 2. Способ...
Предыдущий патент: Неорганические наночастицы высокой плотности, используемые для разрушения клеток in vivo
Следующий патент: Синтез стирилпиридинов, меченных радиоактивным изотопом 18f, из предшественников тозилата и их стабильные фармацевтические композиции
Случайный патент: Устройство и способ обработки для несмешивающихся жидкостей