Взаимодействие moraxella catarrhalis с эпителиальными клетками, внеклеточными матриксными белками и системой комплемента





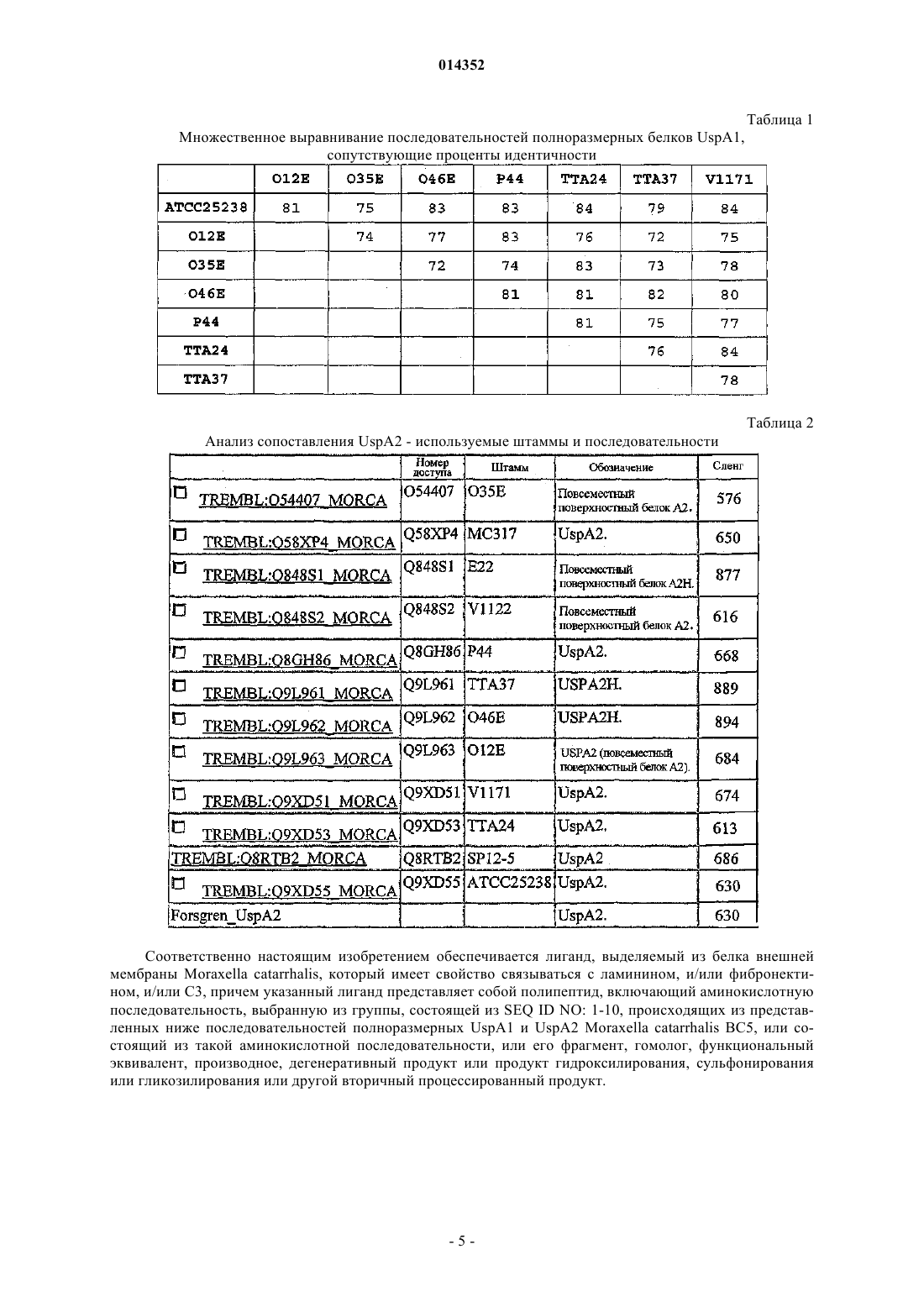


Формула / Реферат
1. Пептид, состоящий из последовательности ID NO. 2, или его фрагмент, гомолог, функциональный эквивалент, производное, дегенеративный продукт или продукт гидроксилирования, сульфонирования или гликозилирования или другой вторичный процессированный продукт, который сохраняет свойства связывать фибронектин.
2. Пептид, состоящий из последовательности ID NO. 3, или его фрагмент, гомолог, функциональный эквивалент, производное, дегенеративный продукт или продукт гидроксилирования, сульфонирования или гликозилирования или другой вторичный процессированный продукт, который сохраняет свойства связывать фибронектин.
3. Применение по крайней мере одного пептида по любому из пп.1, 2 для получения лекарственного средства для лечения или профилактики инфекции.
4. Применение по п.3, при котором инфекция вызвана Moraxella catarrhalis.
5. Применение по п.3 или 4 для профилактики или лечения воспаления среднего уха, синусита или инфекций нижних дыхательных путей.
6. Лиганд, включающий связывающий фибронектин домен и состоящий из аминокислотной последовательности, выбранной из группы, состоящей из последовательности ID NO. 2 или последовательности ID NO. 3, или его фрагмент, гомолог, функциональный эквивалент, производное, дегенеративный продукт или продукт гидроксилирования, сульфонирования или гликозилирования или другой вторичный процессированный продукт, который сохраняет свойства связывать фибронектин.
7. Слитый белок, включающий один или несколько лигандов по п.6.
8. Лекарственное средство, содержащее один или несколько лигандов по п.6 или 7 и один или несколько фармацевтически приемлемых адъювантов, наполнителей, вспомогательных веществ, связующих веществ, носителей или консервантов.
9. Вакцина, содержащая один или несколько лигандов по п.6 или 7 и один или несколько фармацевтически приемлемых адъювантов, наполнителей, вспомогательных веществ, связующих веществ, носителей или консервантов.
10. Способ лечения или профилактики у индивидуума инфекции, включающий введение фармацевтически эффективного количества лекарственного средства по п.8 или вакцины по п.9.
11. Способ по п.10, в котором инфекция вызвана Moraxella catarrhalis.
12. Последовательность нуклеиновой кислоты, кодирующая лиганд, определенный в п.6, или пептид, определенный в п.1 или 2.
13. Полипептид или усеченный полипептид, включающий по крайней мере одну из консервативных последовательностей последовательности ID NO. 2 или последовательности ID NO.3, или его фрагмент, гомолог, функциональный эквивалент, производное, дегенеративный продукт или продукт гидроксилирования, сульфонирования или гликозилирования или другой вторичный процессированный продукт, который обладает способностью связывать фибронектин.
14. Вакцинная композиция, содержащая эффективное количество UspA1 и/или UspA2 в комбинации с эффективным количеством белка MID и один или несколько фармацевтически приемлемых адъювантов, наполнителей, вспомогательных веществ, связующих веществ, носителей или консервантов.
15. Применение эффективного количества UspA1 и/или UspA2 в комбинации с эффективным количеством белка MID для получения лекарственного средства для лечения или профилактики инфекции.
16. Применение по п.15, при котором инфекция вызвана Moraxella catarrhalis.
Текст
ВЗАИМОДЕЙСТВИЕ MORAXELLA CATARRHALIS С ЭПИТЕЛИАЛЬНЫМИ КЛЕТКАМИ, ВНЕКЛЕТОЧНЫМИ МАТРИКСНЫМИ БЕЛКАМИ И СИСТЕМОЙ КОМПЛЕМЕНТА Настоящее изобретение относится к внеклеточным матриксным белкам Moraxella catarrhalis и их способности взаимодействовать с эпителиальными клетками через ассоциированные с клеткой фибронектин и ламинин, а также к их способности ингибировать систему комплемента. Эти внеклеточные матриксные белки пригодны для приготовления вакцин. Настоящим изобретением обеспечиваются пептиды, взаимодействующие с фибронектином, ламинином и системой комплемента.(71)(73) Заявитель и патентовладелец: АРНЕ ФОРСГРЕН АБ (SE) 014352 Область техники Настоящее изобретение относится к Moraxella catarrhalis и их способности взаимодействовать с эпителиальными клетками через внеклеточные матриксные белки, такие как фибронектин и ламинин, а также к их способности ингибировать систему комплемента. Взаимодействие с этими внеклеточными белками используется для приготовления вакцин. Уровень техники Способность связываться с эпителиальными клетками имеет огромное значение для нескольких видов бактерий. Например, Staphylococcus aureus и Streptococcus pyogenes обладают связывающими фибронектин белками (FnBP) с родственной организацией последовательностей. Эти FnBP известны как микробные поверхностные компоненты, узнающие адгезивные матриксные молекулы (MSCRAMM). При связывании с фибронектином они используют модульную структуру фибронектина, образующего обширные тандемные бета-молнии [27, 39, 47, 73]. Назначение фибронектина заключается в опосредовании адгезии бактерий и проникновения в клетки хозяина. Важный патоген слизистой оболочки Moraxella catarrhalis является третьей ведущей бактериальной причиной острого воспаления среднего уха у детей после Streptococcus pneumoniae и Haemophilus influenzae [14, 40, 55]. М. catarrhalis также является одним из наиболее частых обитателей глотки здоровых детей. Более того, М. catarrhalis также является частой причиной синусита и инфекций нижних дыхательных путей у взрослых с хроническим обструктивным заболеванием легких (COPD) [74]. Вероятно, частое появление этого вида у больных COPD отчасти связано с наличием у него большого набора адгезинов. В последние годы исследование фокусировалось на белках внешней мембраны и их взаимодействиях с хозяином, являющимся человеком [6, 48, 56]. Некоторые из этих белков внешней мембраны, в том числе среди прочих связывающий IgD M. catarrhalis (MID, также обозначаемый Hag), белок CD, белок адгезии М. catarrhalis (McaP) и повсеместные поверхностные белки (Usp), имеют, по-видимому, адгезивные функции [1, 22, 33, 48, 61, 81, 84]. Сущность изобретения Ввиду того факта, что М. catarrhalis является, как обнаружено, ведущей причиной инфицирования верхних и нижних дыхательных путей, существует текущая необходимость в разработке вакцин, которые можно использовать против М. catarrhalis. Поэтому цель настоящего изобретения состояла в обнаружении того, каким образом М. catarrhalis взаимодействует с эпителиальными клетками в организме и влияет на иммунную систему. Таким способом можно разработать вещества, которые могут действовать в качестве вакцин против M. catarrhalis. В этом исследовании авторы настоящего изобретения, используя происходящие из клинических изолятов мутанты М. catarrhalis, смогли показать, что и UspA1, и А 2 связывают фибронектин и ламинин. Кроме того, авторы настоящего изобретения смогли показать, что М. catarrhalis блокирует классический путь системы активации комплемента, а также объяснить, каким образом они осуществляют блокирование. Многие бактерии адгезируются к эпителиальным клеткам через связывающие фибронектинMSCRAMM [54, 77]. Pseudomonas aeruginosa имеет FnBP, который связывается с ассоциированным с клеткой фибронектином на эпителиальных клетках полости носа [69]. Блокирование взаимодействий бактерии-белок фибронектин может помочь ткани хозяина побороть инфекцию. Действительно было показано, что антитела против FnBP S. siureus приводят к быстрому обезвреживанию бактерий у инфицированных мышей [71]. Рекомбинантные усеченные белки UspA1/A2 вместе с более маленькими фрагментами, перекрывающими целую молекулу, проверили в соответствии с настоящим изобретением на связывание с фибронектином. И UspA1, и А 2 связывали фибронектин, и обнаружено, что связывающие фибронектин домены расположены в пределах UspA1299-452 и UspA2165-318. Эти два усеченных белка оба ингибировали связывание М. catarrhalis с эпителиальными клетками конъюнктивы Чанг в степени, сходной с ингибированием с использованием антител против фибронектина. Данные наблюдения показали, что и UspA1, и А 2 М. catarrhalis вовлечены в адгезию к эпителиальным клеткам через ассоциированный с клеткой фибронектин. Следовательно, в качестве возможных кандидатов, которые следует включать в вакцину против М. catarrhalis, предлагаются биологически активные сайты в пределах UspA1299-452 и UspA2165-318. Кроме того, авторы настоящего изобретения изучили и охарактеризовали связывание М. catarrhalis с ламинином. М. catarrhalis является частой причиной обострений инфекций у больных COPD. Вероятно,успех этого вида у больных COPD отчасти связан с наличием у него большого набора адгезинов. Кроме того, существуют патологические изменения, такие как потеря целостности эпителия с обнажением базальной мембраны, где у курильщиков утолщен сам слой ламинина [4]. Показано, что некоторые патогены способны связывать ламинин, и это может вносить вклад в их способность адгезироваться к таким поврежденным и оголенным поверхностям слизистой оболочки. Эти патогены включают патогены, которые, как известно, вызывают значимое заболевание дыхательных путей, такие как, среди прочих, S.aureus и P. aeruginosa [7, 63]. Авторы настоящего изобретения смогли показать, что повсеместный по-1 014352 верхностный белок (Usp) A1 и А 2 M. catarrhalis также связывается с ламинином. Связывающие ламинин домены UspA1 и А 2 были обнаружены, среди прочих, в пределах N-концевых половин UspA150-491 иUspA230-351. Эти домены также содержат связывающие фибронектин домены. Однако самые маленькие фрагменты, которые связывали фибронектин, UspA1299-452 и UspA2165-318, не связывали в какой-либо заметной степени ламинин. Фрагменты, меньшие чем N-концевая половина UspA1 (UspA150-491), теряют всю свою способность связывать ламинин, в то время как в случае UspA2 только UspA230-170 связывал ламинин, хотя на более низком уровне, чем целый рекомбинантный белок (UspA230-539). Эти открытия говорят о том, что разные части молекулы могут иметь разные функциональные роли. Также обнаружено, что UspA150-770 обладает свойствами связывать ламинин. При сравнении самых маленьких связывающих ламинин областей UspA1 и А 2 авторы настоящего изобретения обнаружили, что существует, однако, незначительное подобие в виде аминокислотной гомологии между UspA230-170 и UspA150-491 (данные не показаны). Это не удивительно, поскольку известен тот факт, что оба белка имеют структуру сферической головки в виде "леденца", несмотря на то, что ихN-концевые половины идентичны только на 22% [2, 32]. В качестве возможных кандидатов, которые следует включать в вакцину против М. catarrhalis,предлагаются биологически активные сайты в пределах UspA150-770 и UspA230-539. Наконец, авторы настоящего изобретения изучили взаимодействие между повсеместными поверхностными белками А 1 и А 2 М. catarrhalis и врожденной иммунной системой и обнаружили, что М. catarrhalis блокирует систему комплемента. Система комплемента является одной из первых линий врожденной защиты против патогенных микроорганизмов, и активация этой системы приводит к каскаду осаждения белков на поверхности бактерий, что приводит к образованию мембрано-атакующего комплекса или опсонизации патогена с последующим фагоцитозом [85, 86]. Одним из наиболее важных белков комплемента является С 3, который присутствует в кровотоке в концентрации, схожей с концентрацией некоторых иммуноглобулинов (1-1,2 мг/мл). С 3 не только играет решающую роль в качестве опсонина,но также является общим связующим звеном между классическим, лектиновым и альтернативным путями активации комплемента. Альтернативный путь функционирует в качестве петли усиления для классического и лектинового путей, и его можно также спонтанно активировать с помощью ковалентного присоединения С 3 к поверхности микробной клетки при отсутствии ингибиторов комплемента. Для осаждения С 3 требуется присутствие внутренней тиоэфирной связи, образованной в нативном белке при близости сульфгидрильной группы (Cys1010) и карбонильной группы глутамина (Gln1012) на -цепи С 3[76]. Протеолитическое отщепление с аминоконца -цепи С 3 пептида из 77 остатков приводит к образованию С 3 а (анафилатоксина) и С 3b. Присоединение С 3b затем осуществляется через ковалентную связь между карбонильной группой метастабильного тиоэфира и/или группами белков -NH2 или -ОН, или углеводными структурами на поверхности активатора [36, 37]. Обнаружено, что UspA1 и А 2 М. catarrhalis нековалентно и зависимым от дозы способом связываются с третьим компонентом комплемента (С 3) из обработанной EDTA сыворотки и обработанным метиламином С 3 (C3met). Обнаружено, что UspA150-770 и UspA230-539 связываются с С 3 и C3met. Обнаружено, что связывающая С 3 область UspA2 расположена главным образом в UspA2200-458. Однако UspA1, как обнаружено, играет минорную роль в этих взаимодействиях. В качестве возможных кандидатов, которые следует включать в вакцину против М. catarrhalis,предлагаются биологически активные сайты в пределах UspA150-770 и UspA230-539. Семейство UpsA состоит из UspA1 (молекулярная масса 88 кДа), UspA2 (62 кДа) и гибридного белка UspA2H (92 кДа) [2, 43]. Эти белки мигрируют в виде высокомолекулярных комплексов при электрофорезе в SDS-PAGE, являются относительно консервативными и, следовательно, важными вакцинными кандидатами. Аминокислотные последовательности UspA1 и А 2 идентичны на 43% и имеют 140 аминокислотных остатков, которые идентичны на 93% [2]. В ряду из 108 изолятов М. catarrhalis из носоглотки новорожденных с воспалением среднего уха гены uspA1 и uspA2 обнаружены в 107 (99%) и 108 (100%) изолятах соответственно. Двадцать один процент новорожденных были идентифицированы как имеющие гибридный вариант гена UspA2H [50]. Кроме того известно, что естественно приобретенные антитела против UspA1 и А 2 являются бактерицидными [15]. Семейству белков UpsA приписывается несколько функций. Экспрессия UpsA является существенной для прикрепления M. catarrhalis к эпителиальным клеткам конъюнктивы Чанг и к эпителиальным клеткам гортани Нер-2 [43, 49]. В более недавнем исследовании обнаружено, что UpsA связывается с относящимися к канцероэмбриональным антигенам клеточными адгезивными молекулами (CEACAM),экспрессируемыми линией эпителиальных клеток легкого А 549 [31]. Также показано, что очищенныйUspA1 связывает фибронектин в дот-блот анализах, в то время как очищенный UspA2 не связывает [49]. И UspA1, и А 2 могут играть важные роли в устойчивости сыворотки к М. catarrhalis [1, 5, 58, 60]. В настоящем изобретении продемонстрировано, что и UspA1, и А 2 являются детерминантами связывания М. catarrhalis с фибронектином и ламинином в клинических изолятах М. catarrhalis ВВН 18 иRH4. Интересно, что оба рекомбинантных UspA1 и А 2, происходящих из М. catarrhalis Bc5, связывали фибронектин в одинаковой степени. Связывающие фибронектин домены обнаружены в пределах аминокислотных остатков 229-452 UspA1 и 165-318 UspA2. Последовательности этих двух доменов идентичны-2 014352 по 31 аминокислотному остатку. Важно, что усеченные фрагменты белков, содержащие эти остатки вUspA1 и UspA2, были способны ингибировать связывание М. catarrhalis с эпителиальными клетками Чанг, что говорит о том, что взаимодействия с этими клетками осуществлялись через ассоциированный с клеткой фибронектин. Связывающие ламинин домены обнаружили в пределах отмеченных выше аминокислотных остатков. С помощью анализов связывания с использованием рекомбинантных белков обнаружили, что главные связывающие области расположены в N-концевых частях, где оба белка образуют сферическую головку. Бактериальные факторы, опосредующие адгезию к ткани, и компоненты внеклеточного матрикса(ЕСМ) группируются вместе в одно семейство, называемое "микробные поверхностные компоненты,распознающие адгезивные матриксные молекулы" (MSCRAMM). Поскольку и UspA1, и UspA2 связывают фибронектин и ламинин, эти белки можно обозначитьMSCRAMM. В соответствии с одним аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 1, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии с другим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 2, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии со следующим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 3, и его фрагменты, гомологи, функциональные эквиваленты, производные, дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии с другим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 4, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии со следующим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 5, и его фрагменты, гомологи, функциональные эквиваленты, производные, дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии со следующим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 6, и его фрагменты, гомологи, функциональные эквиваленты, производные, дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии с другим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 7, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии с другим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 8, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процссированные продукты. В соответствии с другим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 9, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии с другим аспектом настоящим изобретением обеспечивается пептид, имеющий последовательность ID NO. 10, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. В соответствии с другим аспектом настоящим изобретением обеспечивается применение по крайней мере одного пептида в соответствии с настоящим изобретением для получения лекарственного средства для лечения или профилактики инфекции, предпочтительно инфекции, вызванной М. catarrhalis, в частности вызванной носительством М. catarrhalis на поверхностях слизистых оболочек. В соответствии с другим аспектом настоящим изобретением, кроме того, обеспечивается включающий связывающий фибронектин домен лиганд, который состоит из аминокислотной последовательности, выбранной из группы, состоящей из последовательности ID NO. 1, последовательности ID NO. 2 и последовательности ID NO. 3, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и-3 014352 другие вторичные процессированные продукты. Кроме того, настоящим изобретением обеспечивается включающий связывающий ламинин домен лиганд, который состоит из аминокислотной последовательности, выбранной из группы, состоящей из последовательности ID NO. 4 - последовательности ID NO. 8, и его фрагменты, гомологи, функциональные эквиваленты, производные, дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. Кроме того, настоящим изобретением обеспечивается лиганд, включающий связывающий С 3 илиC3met домен, который состоит из аминокислотной последовательности, выбранной из группы, состоящей из последовательности ID NO. 4, последовательности ID NO. 6, последовательности ID NO. 9 и последовательности ID NO. 10, и его фрагменты, гомологи, функциональные эквиваленты, производные,дегенеративные продукты или продукты гидроксилирования, сульфонирования или гликозилирования и другие вторичные процессированные продукты. Кроме того, настоящим изобретением обеспечивается лекарственное средство, содержащее один или несколько лигандов в соответствии с настоящим изобретением и один или несколько фармацевтически приемлемых адъювантов, наполнителей, вспомогательных веществ, связующих веществ, носителей или консервантов. Кроме того, настоящим изобретением обеспечивается вакцина, содержащая один или несколько лигандов в соответствии с настоящим изобретением и один или несколько фармацевтически приемлемых адъювантов, наполнителей, вспомогательных веществ, связующих веществ, носителей или консервантов. Настоящим изобретением также обеспечивается способ лечения или профилактики у индивидуума инфекции, предпочтительно инфекции, вызванной М. catarrhalis, в частности вызванной носительством М. catarrhalis на поверхностях слизистых оболочек, который включает введение фармацевтически эффективного количества лекарственного средства или вакцины в соответствии с настоящим изобретением. Наконец, настоящим изобретением также обеспечивается последовательность нуклеиновой кислоты, кодирующая лиганд, белок или пептид по настоящему изобретению, а также ее гомологи, полиморфные варианты, вырожденные последовательности и варианты сплайсинга. Дальнейшее раскрытие целей, задач, решений и особенностей настоящего изобретения будет очевидно из следующего детального описания настоящего изобретения со ссылкой на фигуры и прилагаемую формулу изобретения. Используемое здесь выражение "лиганд" предназначено для обозначения и целой молекулы, которая связывается с рецептором, и любой ее части, которая включает связывающий рецептор домен так,что он сохраняет свойство связываться с рецептором. Лиганды, включающие эквивалентные связывающие рецептор домены, также включены в настоящее изобретение. Выражения "фрагмент", "гомолог", "функциональный эквивалент" и "производное" относятся к вариантам, модификациям и/или частям пептидов и фрагментам белков в соответствии с настоящим изобретением, которые сохраняют желаемые свойства связываться с фибронектином, ламинином, С 3 илиC3met. Гомолог UspA1 в соответствии с настоящим изобретением определяется как последовательность,обладающая по крайней мере 72% идентичностью, как видно из табл. 1 ниже. Фрагмент в соответствии с настоящим изобретением определяется как любая гомологичная последовательность, которая является усеченной или удлиненной на 1, 2, 5, 10, 15, 20 аминокислот с N-конца и/или усеченной или удлиненной на 1, 2, 5, 10, 15, 20 аминокислот с С-конца. Выражения "дегенеративные продукты, продукты гидроксилирования, сульфонирования и гликозилирования или другие вторичные процессированные продукты" относятся к вариантам и/или модификациям пептидов и фрагментов белков в соответствии с настоящим изобретением, которые изменены по сравнению с первоначальным пептидом или фрагментом белка в результате дегенерации, гидроксилирования, сульфонирования или гликозилирования, но которые сохраняют желаемые свойства связываться с фибронектином, ламинином, С 3 или C3met. Настоящее изобретение особенно касается инфекций, вызванных Moraxella catarrhalis. Пептид в соответствии с настоящим изобретением можно использовать для лечения или профилактики воспаления среднего уха, синусита или инфекций нижних дыхательных путей.-4 014352 Таблица 1 Множественное выравнивание последовательностей полноразмерных белков UspA1,сопутствующие проценты идентичности Соответственно настоящим изобретением обеспечивается лиганд, выделяемый из белка внешней мембраны Moraxella catarrhalis, который имеет свойство связываться с ламинином, и/или фибронектином, и/или С 3, причем указанный лиганд представляет собой полипептид, включающий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, происходящих из представленных ниже последовательностей полноразмерных UspA1 и UspA2 Moraxella catarrhalis BC5, или состоящий из такой аминокислотной последовательности, или его фрагмент, гомолог, функциональный эквивалент, производное, дегенеративный продукт или продукт гидроксилирования, сульфонирования или гликозилирования или другой вторичный процессированный продукт. В предпочтительном варианте осуществления настоящего изобретения лиганд представляет собой полипептид [или усеченный полипептид по сравнению с полипептидом дикого типа], включающий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, или состоящий из такой аминокислотной последовательности, или его фрагмент, гомолог, функциональный эквивалент,производное, дегенеративный продукт или продукт гидроксилирования, сульфонирования или гликозилирования или другой вторичный процессированный продукт. Термин "лиганд" используется здесь для обозначения и целой молекулы, которая связывается с ламинином, и/или фибронектином, и/или С 3, и любой ее части, которая включает связывающий ламинин,и/или фибронектин, и/или С 3 домен так, что он сохраняет соответствующее связывающее свойство. Таким образом, "лиганд" включает молекулы, которые состоят только из связывающего ламинин, и/или фибронектин, и/или С 3 домена, т.е. области или областей пептида, требуемых для связывания. Для целей настоящего изобретения свойства полипептида связывать ламинин, фибронектин или С 3 можно установить следующим образом. Полипептиды можно пометить 125 йодом или другими радиоактивными соединениями и проверить на связывание в радиоиммуноанализах (РИА) в виде жидкой или твердой фазы (например, дот-блотов). Кроме того, полипептиды можно проанализировать на связывание с помощью твердофазных иммуноферментных анализов (ELISA) или проточной цитометрии с использованием подходящих антител и систем обнаружения. Взаимодействия между полипептидами и ламинином, фибронектином или С 3 можно, кроме того, исследовать с помощью резонанса поверхностных плазмонов (Biacore). Примеры методов подробно приведены в разделе "Материалы и методы". В другом предпочтительном варианте осуществления настоящего изобретения полипептид [или усеченный полипептид по сравнению с полипептидом дикого типа] включает по крайней мере одну из консервативных последовательностей из SEQ ID NO: 1-10, которые идентифицированы при продемонстрированном здесь выравнивании, или состоит по крайней мере из одной такой консервативной последо-6 014352 вательности. Следовательно, в этом варианте осуществления настоящего изобретения полипептид [или полипептид, усеченный по сравнению с полипептидом дикого типа] включает или состоит по крайней мере из одной из следующих последовательностей: Из UspA1 (консервативных фрагментов связывающего фибронектин домена - '/' отделяет альтернативные выборы аминокислоты в каком-либо положении) Из UspA2 (консервативных фрагментов связывающего фибронектин домена - '/' отделяет альтернативные выборы аминокислоты в каком-либо положении) Из UspA2 (консервативных фрагментов связывающего С 3 домена '/' отделяет альтернативные выборы аминокислоты в каком-либо положении) Понятно, что являющиеся полипептидами лиганды настоящего изобретения могут включать связывающий ламинин, и/или фибронектин, и/или С 3 домен упоминаемой здесь последовательности, модифицированной добавлением или делецией аминокислотных остатков к или из упоминаемой здесь последовательности с одного из двух или с обоих N- и C-концов, причем модифицированные пептиды сохраняют способность связывать ламинин, и/или фибронектин, и/или С 3 соответственно. Соответственно настоящим изобретением, кроме того, обеспечивается лиганд, включающий или состоящий из полипептида, к которому добавлены или из которого делетированы 50, 40, 30, 20, 10, 5, 3 или 1 аминокислотный остаток упоминаемой здесь последовательности с одного из двух или с обоих N- и С-концов, причем указанный модифицированный пептид сохраняет способность связывать ламинин, и/или фибронектин,и/или С 3; и/или индуцировать иммунный ответ против не модифицированного пептида. Под удлинением понимается удлинение последовательности с использованием пептида из полноразмерной аминокислотной последовательности, из которой он получен. Что касается фрагментов полипептидов настоящего изобретения, то в настоящем изобретении можно использовать фрагменты любого размера (основанные на гомологичных последовательностях/консервативных областях/функциональных доменах, обсуждаемых здесь) при условии, что эти фрагменты сохраняют способность связывать ламинин, и/или фибронектин, и/или С 3. Может быть желательным выделение минимального пептида, содержащего только такие области, которые требуются для связывания с рецептором. Являющиеся полипептидами лиганды в соответствии с настоящим изобретением можно получить из известных белков UspA1 или UspA2 Moraxella catarrhalis с помощью усечения одного из двух или обоих N- и С-концов. Усеченные полипептиды не являются полноразмерными нативными молекуламиUspA1 или UspA2. Соответственно настоящим изобретением, кроме того, обеспечивается последовательность UspA1 дикого типа, в которой отсутствует по крайней мере (или точно) 20, 30, 40, 50, 60, 70,80, 100, 120, 140, 160 и т.п. до 298 аминокислот с N-конца и/или в которой отсутствует по крайней мере(или точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140, 160, 180, 200 и т.п. до 450 аминокислот с С-конца. Предпочтительно усеченные полипептиды сохраняют функцию связывания с фибронектином (необязательно также связывания с ламинином и/или С 3).-7 014352 Таблица 3 Возможные комбинации усечений с N- и C-концов белка UspA1 дикого шипа Соответственно настоящим изобретением, кроме того, обеспечивается последовательность UspA2 дикого типа, в которой отсутствует по крайней мере (или точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140,160, 164 аминокислоты с N-конца и/или в которой отсутствует по крайней мере (или точно) 20, 30, 40, 50,60, 70, 80, 100, 120, 140, 180, 200 и т.п. до 312 аминокислот с С-конца. Предпочтительно усеченные полипептиды сохраняют функцию связывания с фибронектином (необязательно также связывания с ламинином и/или С 3). Возможные усеченные полипептиды можно выбрать из усеченных полипептидов,представленных в следующей таблице, все из которых находятся в пределах объема настоящего изобретения. Таблица 4 Возможные комбинации усечений с N- и C-концов белка UspA2 дикого типа Соответственно настоящим изобретением, кроме того, обеспечивается последовательность UspA2 дикого типа, в которой отсутствует по крайней мере (или точно) 5, 10, 15, 20, 25 или 29 аминокислот с Nконца и/или в которой отсутствует по крайней мере (или точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140,160, 180, 200 и т.п. до 453 аминокислот с С-конца. Предпочтительно усеченные полипептиды сохраняют функцию связывания с ламинином (необязательно также связывания с фибронектином и/или С 3). Возможные усеченные полипептиды можно выбрать из усеченных полипептидов, представленных в следующей таблице, все из которых находятся в пределах объема настоящего изобретения.-8 014352 Таблица 5 Возможные комбинации усечений с N- и C-концов белка UspA2 дикого типа Соответственно настоящим изобретением, кроме того, обеспечивается последовательность UspA2 дикого типа, в которой отсутствует по крайней мере (или точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140,160 и т.п. до 301 аминокислоты с N-конца и/или в которой отсутствует по крайней мере (или точно) 20,30, 40, 50, 60, 70, 80, 100, 120, 140, 160 или 172 аминокислоты с С-конца. Предпочтительно усеченные полипептиды сохраняют функцию связывания с С 3 (необязательно также связывания с фибронектином и/или ламинином). Возможные усеченные полипептиды можно выбрать из усеченных полипептидов,представленных в следующей таблице, все из которых находятся в пределах объема настоящего изобретения.-9 014352 Таблица 6 Возможные комбинации усечений с N- и C-концов белка UspA2 дикого типа Известные последовательности UspA1 дикого типа, которые можно подвергнуть усечению таким образом, представляют собой последовательности штаммов АТСС 2538 (МХ 2; номер доступа в GenBank AAD43495), Р 44 (AAN84895), О 35 Е (ААВ 96359), ТТА 37 (AAF40122), O12 Е (AAF40118), О 46 Е(AAF36416), V1171 (AAD43469), ТТА 24 (AAD43467) (см. табл. 1/фиг. 19) или ВС 5 (см. выше). Известные последовательности UspA2 дикого типа, которые можно подвергнуть усечению таким образом,представляют собой последовательности штаммов О 35 Е (номер доступа в GenBank - O4407), MC317 (номер доступа в GenBank - Q58XP4), Е 22 (номер доступа в GenBank - Q848S1), V1122 (номер доступа вBC5 (ForsgrenUspA2) (см. выше). Идеально, если усеченный полипептид UspA1 или UspA2 этого варианта осуществления настоящего изобретения включает или состоит из аминокислотной последовательности, выбранной из группы,состоящей из SEQ ID NO: 1-10, или ее фрагмента, гомолога, функционального эквивалента, производного, дегенеративного продукта или продукта гидроксилирования, сульфонирования или гликозилирования или другого вторичного процессированного продукта; или включает или состоит по крайней мере из одной из консервативных последовательностей внутри этих областей, которые идентифицированы при продемонстрированном здесь выравнивании, например,из UspA1 (консервативных фрагментов связывающего фибронектин домена - '/' отделяет альтернативные выборы аминокислоты в каком-либо положении) Из UspA2 (консервативных фрагментов связывающего фибронектин домена - '/' отделяет альтернативные выборы аминокислоты в каком-либо положении) Из UspA2 (консервативных фрагментов связывающего С 3 домена - '/' отделяет альтернативные выборы аминокислоты в каком-либо положении) Может быть подходящим получение слитых белков, содержащих являющиеся полипептидами лиганды, описанные здесь. Соответственно в дополнительном варианте осуществления настоящим изобретением обеспечиваются слитые белки, включающие являющиеся полипептидами лиганды в соответствии с настоящим изобретением. Предпочтительно слитый белок в соответствии с этим вариантом осуществления настоящего изобретения идентичен менее чем на 50% любой известной полноразмерной последовательности на протяжении полной длины. Такие слитые белки могут составлять производное полипептидов настоящего изобретения. Кроме того, в производных могут использоваться полипептиды настоящего изобретения в качестве носителя для ковалентного присоединения пептидных или сахаридных составляющих. Их можно соединить, например, с пневмококковыми капсульными олигосахаридами, или полисахаридами, или липоолигосахаридами Moraxella catarrhalis, или липоолигосахаридами не типируемых Haemophilus influenzae. Гомологичные пептиды настоящего изобретения можно идентифицировать с помощью сравнения последовательностей. Предпочтительно гомологичные пептиды идентичны по крайней мере на 60%, более предпочтительно по крайней мере на 70, 80, 90, 95 или 99%, в порядке восхождения предпочтения,раскрытой здесь пептидной последовательности или ее фрагментам или усеченным полипептидам настоящего изобретения на протяжении их полной длины. Предпочтительно гомологичный пептид сохраняет способность связывать фибронектин, и/или ламинин, и/или С 3; и/или индуцировать иммунный ответ против раскрытых здесь пептидных последовательностей или их фрагментов. На фиг. 19 и 20 показано выравнивание пептидных последовательностей UspA1 и UspA2 различного происхождения, при котором указаны области последовательности, которые можно модифицировать с образованием гомологичных последовательностей при сохранении функции (т.е. способности связывать фибронектин, и/или ламинин, и/или С 3). Пептидами, гомологичными пептидам SEQ ID NO: 1-10 ВС 5,являются, например, такие последовательности, которые соответствуют последовательности ВС 5, из других штаммов, представленных на фиг. 19 и 20. Вакцины настоящего изобретения Полипептиды/пептиды/функциональные домены/гомологи/фрагменты/усеченные полипептиды/ производные настоящего изобретения в идеале готовят в виде вакцины, содержащей эффективное количество указанного(ых) компонента(ов) и фармацевтически приемлемое вспомогательное вещество. Вакцины настоящего изобретения можно использовать для назначения пациенту для профилактики или лечения инфекций, вызванных Moraxella catarrhalis, или воспаления среднего уха, или синусита, или инфекций нижних дыхательных путей. Их можно вводить любым известным способом, в том числе- 11014352 внутримышечно, парентерально, на слизистые оболочки и интраназально. Комбинированные вакцины настоящего изобретения Вакцины настоящего изобретения можно скомбинировать с другими антигенами Moraxella catarrhalis для профилактики или лечения отмеченных выше заболеваний. Авторы настоящего изобретения, в частности, обнаружили, что Moraxella catarrhalis имеет по крайней мере два способа торможения иммунной системы хозяина от наступления на организм. В дополнение к взаимодействию с С 3 (и С 4 ВР), упоминаемому в приведенных ниже примерах, М. catarrhalis имеет сильное сродство к растворимому и связанному с мембраной IgD человека благодаря белку MID (также известному как OMP106). Moraxella-зависимое связывание IgD с В-лимфоцитами приводит к синтезу поликлональных иммуноглобулинов, который может препятствовать продукции специфических моноклональных антител против Moraxella. Тот факт, что М. catarrhalis тормозит иммунную систему человека несколькими способами, может служить объяснением того, почему М. catarrhalis является таким частым обитателем дыхательных путей. Авторы настоящего изобретения полагают, что комбинация антигенов, вовлеченных в функцию связывания IgD (MID) и функцию связывания С 3 (UspA1 и/или UspA2), может обеспечить иммуногенную композицию, увеличивающую защитные способности хозяина против торможения бактериями родаMoraxella иммунной системы человека, обеспечивая таким образом усиленное снижение носительстваM. catarrhalis на поверхностях слизистых оболочек. Следовательно, дальнейшим аспектом настоящего изобретения является вакцинная композиция,содержащая эффективное количество UspA1 и/или UspA2 (особенно последнего) (например полноразмерных полипептидов или полипептидов/пептидов/функциональных доменов/гомологов/фрагментов/ усеченных полипептидов/производных настоящего изобретения, описанных здесь, которые предпочтительно сохраняют функцию связывания С 3) в комбинации с эффективным количеством белка MID (например, полноразмерных полипептидов или их полипептидов/пептидов/функциональных доменов/гомологов/фрагментов/усеченных полипептидов/производных, которые предпочтительно сохраняют функцию связывания IgD человека) и фармацевтически приемлемое вспомогательное вещество. Белок MID и связывающие IgD его гомологи/фрагменты/усеченные полипептиды описаны в заявкеWO 03/004651 (включенной сюда посредством ссылки). Особенно подходящими для этой цели фрагментами является полипептид, включающий (или состоящий из) F2-фрагмент, описанный в заявке WO 03/004651, или последовательности, идентичные с ним на 60, 70, 80, 90, 95, 99%, которые предпочтительно сохраняют активность связывания IgD человека. Компоненты MID и UspA этой комбинированной вакцины могут быть отделены друг от друга, или они могут быть подходящим образом слиты вместе с помощью известных методов молекулярной биологии. Краткое описание чертежей На фиг. 1 демонстрируются тринадцать штаммов М. catarrhalis, проверенных на связывание с фибронектином (А). Сильное связывание фибронектина коррелировало с экспрессией UspAl/A2, определенной с помощью pAb против UspA1/A2 (B-I). Профили проточной цитометрии М. catarrhalis BBH18 дикого типа и дефицитных в отношении UspA1/A2 мутантов демонстрируют зависимое от UspA1/A2 связывание с растворимым фибронектином. Представлены профили клинического изолята дикого типа (В и F) и соответствующих мутантов, лишенных UspA1 (С и G) или UspA2 (D и Н), и двойных мутантов (Е и I),у которых отсутствует и UspA1, и UspA2. Бактерии инкубировали с кроличьими антителами противFITC-конъюгированные pAb с последующим анализом проточной цитометрией. Продемонстрирован типичный эксперимент из трех со средней интенсивностью флуоресценции (MFI) для каждого профиля. На фиг. 2 демонстрируется, что дефицитные в отношении UspA2 мутанты М. catarrhalis RH4 не связывают 125I-меченный фибронектин. В качестве отрицательного контроля, не связывающего фибронектин, включили Е. coli BL21. Бактерии инкубировали с 125I-меченным фибронектином с последующими несколькими промывками и анализировали в гамма-счетчике. За 100% приняли связывание фибронектина с RH4 дикого типа, экспрессирующим и UspA1, и А 2. Показаны средние значения трех независимых экспериментов. Планки погрешностей представляют стандартные отклонения (SD). Схожие результаты были получены с М. catarrhalis BBH18. На фиг. 3 демонстрируются фотографии, подтверждающие, что мутанты М. catarrhalis, лишенныеUspA1 и UspA2, не связываются с иммобилизованным фибронектином. М. catarrhalis дикого типа мог адгезироваться к покрытым фибронектином предметным стеклам с высокой плотностью (А). Мутант М.catarrhalis AuspA1 также сохранял высокую плотность адгезии (В), в то время как М. catarrhalis AuspA2 и двойные мутанты AuspA1/A2 слабо адгезировались (С и D). Предметные стекла покрывали фибронектином и инкубировали с М. catarrhalis RH4 и его соответствующими мутантами UspAl/A2. После нескольких промывок бактерии окрашивали по Граму. Фиг. 4 представляет собой график, на котором демонстрируется, что рекомбинантные UspA1 и А 2 связываются с фибронектином зависимым от дозы способом. Для UspA150-770 и UspA230-539 показано специфическое связывание фибронектина. Обоими белками UpsA (40 нМ) покрывали микротитрационные- 12014352 планшеты и инкубировали с возрастающими концентрациями фибронетина с последующим обнаружением с помощью кроличьих pAb против фибронектина человека и HRP-конъюгированных антикроличьих pAb. Показаны средние значения трех отдельных экспериментов, а планки погрешностей указывают стандартные отклонения (SD). Фиг. 5. Активные связывающие фибронектин домены UspA1 и UspA2 находятся между аминокислотами 299-452 и 165-318 соответственно. Показаны усеченные белки, полученные из UspAl (А) и UspA2(В). Все фрагменты проверяли на связывание с фибронектином в ELISA. 40 нМ каждого усеченного фрагмента покрывали микротитрационные планшеты и инкубировали с 80 мкг/мл и 120 мкг/мл фибронектина для UspA1 и UspA2 соответственно. Связанный фибронектин обнаруживали с помощью кроличьих pAb против фибронектина, а затем HRP-конъюгированных антикроличьих pAb. Показаны результаты трех наборов экспериментов. Планки погрешностей представляют стандартные отклонения(SD). На фиг. 6 демонстрируется последовательность в соответствии с последовательностью ID NO: 1 и гомология последовательностей между UspA1299-452 и UspA2165-318. В скобках находятся 31 идентичных аминокислотных остатков. На фиг. 7 демонстрируется, что усеченные фрагменты UspAl50-491 и UspA1299-452 конкурентно ингибируют зависимое от UpsA связывание М. catarrhalis с фибронектином. В качестве отрицательных контролей включали двойные мутанты AuspA1/A2 M. catarrhalis, которые не связывают фибронектин. Рекомбинантные белки UspAl предварительно инкубировали с 2 мг/100 мл фибронектина до инкубации с М. catarrhalis. Представлены средние значения флуоресценции (MFI) M. catarrhalis со связанным фибронектином, обнаруживаемым с помощью FITC-конъюгированных pAb против фибронектина в анализе проточной цитометрией. UspA150-491 и UspA1299-452 приводили к ингибированию на 95 и 63% соответственно. Планки погрешностей представляют средние значениястандартные отклонения (SD) трех независимых экспериментов. На фиг. 8 демонстрируется, что UspA1299-452 и UspA2165-318 ингибируют адгезию М. catarrhalis к клеткам конъюнктивы Чанг через ассоциированный с клетками фибронектин. Эпителиальные клетки Чанг экспрессируют фибронектин на поверхности, что обнаружено с помощью pAb против фибронетина и проточной цитометрии (А). Предварительная инкубация со связывающими фибронектин белкамиUspA1299-452, UspA2165-318 или pAb против фибронектина приводила к значительному уменьшению связывания М. catarrhalis RH4 по сравнению с контрольными рекомбинантными белками (UspA1433-580 иUspA230-177) и контрольным антителом (мAb против ICAM1) (В). Р 0,05 при проверке по критерию Стьюдента на основе двойной выборки. Показаны средние значения трех отдельных экспериментов, а планки погрешностей указывают стандартные отклонения (SD). На фиг. 9 А демонстрируется связывание М. catarrhalis RH4 с ламинином через UspA1 и А 2. М. catarrhalis RH4 дикого типа (дикий тип) сильно связывался с иммобилизованным ламинином со среднейOD, составляющей 1,27. RH4AuspA1 продемонстрировал среднюю OD, составляющую 1,14 (89,9% дикого типа). RH4AuspA2 и двойной мутант RH4AuspA1/A2 имели среднюю OD, составляющую 0,19 и 0,23 соответственно (15,0 и 18,1% дикого типа). Это не значительно отличалось от остающейся необъясненной адгезии к покрытым бычьим сывороточным альбумином планшетам. Микротитрационные планшеты покрывали 30 мкг/мл ламинина или бычьего сывороточного альбумина. Их блокировали с последующей инкубацией с суспензией бактерий и, наконец, промывали. Связанные бактерии обнаруживали с помощью pAb против MID и HRP-конъюгированных антикроличьихpAb. Показаны усредненные результаты трех представляющих экспериментов. Планки погрешностей представляют стандартные отклонения (SD). На фиг. 9 В демонстрируется связывание рекомбинантных UspA1 и А 2 с ламинином зависимым от дозы способом. Специфическое связывание с ламинином показано для UspA150-770 и UspA230-539. Микротитрационные планшеты покрывали обоими белками UpsA (40 нМ) и инкубировали с возрастающими концентрациями ламинина с последующим обнаружением с помощью кроличьих pAb против ламинина иHRP-конъюгированных антикроличьих pAb. Показаны средние значения трех отдельных экспериментов,а планки погрешностей указывают стандартные отклонения (SD). На фиг. 10 А и В демонстрируется, что активные связывающие ламинин домены UspA150-770 (А) иUspA230-539 (В) находятся в N-концевых половинах. Микротитрационные планшеты покрывали 40 нМ рекомбинантных UspA150-770 и UspA230-539 вместе с усеченными белками и инкубировали с 20 мкг/мл ламинина с последующим обнаружением с помощью кроличьих pAb против ламинина и HRPконъюгированных антикроличьих pAb. Показаны средние значения трех отдельных экспериментов, а планки погрешностей указывают стандартные отклонения (SD). Фиг. 11 является схематической иллюстрацией С 3, ковалентно связанного С 3b и C3met. (А) С 3 молекула в сыворотке состоит из одной -цепи и одной -цепи. (В) -Цепь содержит внутренний тиоэфирный сайт, который после активации может ковалентно присоединиться к поверхности микробов. (С) С 3 обрабатывали метиламином, который становится ковалентно связанным с тиоэфиром. На фиг. 12 иллюстрируется, что М. catarrhalis нейтрализует классический и альтернативные пути- 13014352 системы активации комплемента с помощью белков внешней мембраны UspA1 и А 2. (А) М. catarrhalisRH4 дикого типа (дикий тип), мутанты AuspA1, AuspA2 или AuspA1/A2 инкубировали в присутствии 10% NHS. (В) Мутант AuspA1/А 2 инкубировали с 10% NHS, дополненной или EDTA, или Mg-EGTA. Бактерии собирали в указанные моменты времени. После инкубации в течение ночи подсчитывали колониеобразующие единицы (cfu). За 100% принимали число бактерий в начале экспериментов. Показаны средние значения трех отдельных экспериментов, а планки погрешностей указывают стандартные отклонения (SD). (А) Средние значения после 5 мин для мутантов AuspA1, AuspA2 или AuspA1/A2 значительно отличались от дикого типа (Р 0,05). (В) Средние значения после 5 мин для мутанта AuspA1/A2 и после 10 мин для мутанта AuspA1/A2, проинкубированного с Mg-EGTA, значительно отличались от дикого типа (Р 0,05). На фиг. 13 иллюстрируется, что Moraxella catarrhalis связывает С 3 в сыворотке независимо от активации комплемента. Профили проточной цитометрии, демонстрирующие связывание С 3 с (А) М. catarrhalis или (В) Streptococcus pneumoniae. Бактерии инкубировали с NHS или NHS, предварительно обработанной EDTA. После этого добавляли кроличий pAb против C3d человека и в качестве вторичных антител козьи FITC-конъюгированные антикроличьи pAb с последующим анализом проточной цитометрией. За фоновую флуоресценцию принимали бактерии в отсутствие NHS, но в присутствии обоих pAb. Показан один представляющий эксперимент из трех. На фиг. 14 иллюстрируется, что М. catarrhalis нековалентно связывает очищенный обработанный метиламином С 3 зависимым от дозы способом, и что связывание основывается на ионных взаимодействиях. Профили проточной цитометрии, демонстрирующие (А) связывание с возрастающими концентрациями C3met. (В) Показана средняя интенсивность флуоресценции (MFI) каждого профиля в окне (А).C3met с или без NaCl, как указано. Связывание C3met измеряли с помощью проточной цитометрии, как описано в описании к фиг. 13. Планки погрешностей указывают стандартные отклонения (SD). Р 0,05,Р 0,01, Р 0,001. На фиг. 15 иллюстрируется, что профили проточной цитометрии М. catarrhalis RH4 дикого типа и дефицитных в отношении UspA1/A2 мутантов демонстрируют зависимое от UspA1/UspA2 связываниеC3met/C3. Показаны профили клинического изолята дикого типа (А, F, K) и соответствующих мутантов,лишенных белков MID (В, G, L), UspA1 (С, Н, М) , UspA2 (D, I, N) или обоих белков UspA1 и UspA2 (Е,J, О). Бактерии инкубировали с C3met (А-Е), NHS-EDTA (F-J) или NHS (K-O) и определяли, как указано в подписи к фиг. 3. Показан один типичный эксперимент из трех со средней интенсивностью флуоресценции (MFI) для каждого профиля. На фиг. 16 иллюстрируется, что C3met связывается с очищенным рекомбинантным UspA230-539, в то время как наблюдается только слабое связывание C3met с UspA150-770. Кроме того определено, что связывающая C3met область UspA2 находится между аминокислотными остатками 200-458. (А) Рекомбинантные UspAl50-770 и UspA230-539 иммобилизовали на нитроцеллюлозную мембрану. Мембрану инкубировали с [125I]-меченным C3met в течение ночи и связанный белок визуализировали с помощью PersonalFX (Bio-Rad) с использованием усиливающих экранов. В качестве отрицательного контроля включали рекомбинантный белок MID962-1200. (В) Микротитрационные планшеты покрывали UspA150-770, UspA230-539 и рядом усеченных белковUspA2 и инкубировали с C3met с последующей инкубацией с козьими pAb против С 3 человека и HRPконъюгированными антикозьими pAb. Показаны средние значения трех экспериментов. Из всех образцов вычитали фоновое связывание. Планки погрешностей соответствуют стандартным отклонениям (SD). Р 0,05, Р 0,01, Р 0,001. На фиг. 17 иллюстрируется, что добавление рекомбинантных UspA150-770 и UspA230-539 к сыворотке ингибирует осаждение С 3b и гибель М. catarrhalis через альтернативный путь. Профили проточной цитометрии демонстрируют осаждение С 3b на RH4AuspA1/A2 после инкубации с (A) NHS или NHS, предварительно проинкубированной с рекомбинантными UspA150-770 и UspA230-539, или (В) NHS-Mg-EGTA или NHS-Mg-EGTA, предварительно проинкубированной с UspA150-770 и UspA230-539. После добавления различных комбинаций NHS бактерии анализировали, как описано в подписи к фиг. 13. (С)RH4AuspA1/A2 инкубировали с 10% NHS или NHS-Mg-EGTA. Для ингибирования NHS-Mg-EGTA инкубировали со 100 нМ UspAl50-770 и/или UspA230-539 до добавления бактерий. Бактерии собирали в различные моменты времени. За 100% принимали число бактерий в начале экспериментов. Показаны средние значения трех отдельных экспериментов, а планки погрешностей указывают стандартные отклонения (SD). Моменты времени 10, 20 и 30 мин для мутанта AuspA1/A2, предварительно проинкубированного с рекомбинантными белками, значительно отличались от таких моментов времени для мутантаAuspA1/A2, проинкубированного только с Mg-EGTA (P 0,05). На фиг. 18 иллюстрируется, что рекомбинантные UspA150-770 и UspA230-539 уменьшают гемолиз эритроцитов кролика посредством ингибирования альтернативного пути. NHS инкубировали с или без 100 нМ UspA150-770 и/или UspA230-539 при 37 С в течение 30 мин. После этого NHS в указанных концентрациях добавляли к эритроцитам кролика. После инкубации в течение 30 мин суспензии центрифугиро- 14014352 вали и супернатанты измеряли с помощью спектрофотометрии. Максимальный гемолиз в каждом эксперименте принимали за 100%. Показаны средние значения трех отдельных экспериментов, а планки погрешностей соответствуют стандартным отклонениям (SD). Результаты, полученные с использованиемNHS + UspA230-539 и NHS + UspA150-770/UspA230-539 при концентрациях NHS, составляющих 2, 3 и 4%,значительно отличались от NHS-контролей (Р 0,05). На фиг. 19 иллюстрируется анализ сопоставления UspA1 восьми различных штаммов для того, чтобы показать гомологию различных частей UspA1. На фиг. 20 иллюстрируется анализ сопоставления UspA2 тринадцати различных штаммов для того,чтобы показать гомологию различных частей UspA1. На фиг. 21 иллюстрируется % идентичности в областях, идентифицированных в Forsgrenпоследовательности, подсчитанный как отношение между числом точных соответствий и длиной выровненной области, причем выровненная область - это та часть вышеуказанного общего выравнивания, которая содержит Forsgren-область. Материалы и методы Взаимодействие между М. catarrhalis и фибронектином. Бактериальные штаммы и условия культивирования. В табл. 7 перечислены источники клинических штаммов М. catarrhalis. Мутанты М. catarrhalisBBH18 и RH4 конструировали, как описано ранее [23, 58]. Штаммы М. catarrhalis в заведенном порядке культивировали в жидком бульоне с мозговым и сердечным экстрактами (BHI) или на чашках с BHIагаром при 37 С. Дефицитные в отношении UspA мутанты культивировали в BHI, дополненном 1,5 мкг/мл хлорамфеникола (Sigma, St. Louis, МО), а дефицитные в отношении UspA2 - мутанты инкубировали с 7 мкг/мл зеоцина (Invitrogen, Carlsbad, CA). Для выращивания двойных мутантов использовали и хлорамфеникол, и зеоцин. Таблица 7 Клинические штаммы М. catarrhalis, используемые в настоящем изобретении Примечание: Штаммы С 10, R4 не имеют гена UspA1, в то время как у штаммов F16, R14, Z14 отсутствует ген UspA2 [10]. Остальные штаммы содержали оба гена UspA1 и А 2 (данные не показаны). ДНК-метод. Для обнаружения присутствия генов uspA1, A2 и А 2 Н в тех штаммах, для которых это не известно,использовали праймеры и условия ПЦР, описанные Meier и др. [50]. Частичное секвенированиеUspA1299-452 и UspA2165-318 проводили с использованием 5'- и 3'-праймеров соответствующих генов uspA1 и UspA2 RH4 и ВВН 18. Подтверждение присутствия аминокислотных остатков "DQKADIDNNINNIYELAQQQDQHSSDIKTLK" также проводили с помощью ПЦР с использованием праймера (5'CGCTGACATCCAAGCACTTG-3'), сконструированного с 5'-конца этой последовательности, и 3'праймеров для UspA1 и А 2, как описано Meier и др. [50]. Конструирование и экспрессия рекомбинантных белков Недавно были описаны рекомбинантные UspA150-770 и UspA2230-539, лишенные своих гидрофобных С-концов [58]. Геномную ДНК экстрагировали из М. catarrhalis Вс 5, используя набор DNeasytissue (Qiagen, Hilden, Germany). Кроме того, с помощью того же самого метода были также сконструированы рекомбинантные белки, соответствующие множественным областям, охватывающим UspA150-770 и UspA230-539. В табл. 8 перечислены использованные праймеры. Все конструкции секвенировали в соответствии со стандартными методами. Экспрессию и очистку рекомбинантных белков осуществляли, как- 15014352 описано ранее [59]. Белки очищали, используя колонки, содержащие смолу с никелем (Novagen), в соответствии с инструкциями производителя для нативных условий. Рекомбинантные белки анализировали с помощью электрофореза в SDS-PAGE [21]. Таблица 8 Антитела. Недавно подробно описаны кроличьи поликлональные антитела (pAb) против UspA1/A2 [58]. Другими использованными антителами были кроличьи pAb против фибронектина человека, свиные конъюгированные с FITC (флуоресцинизотиоцианатом) антикроличьи pAb, свиные конъюгированные с пероксидазой хрена (HRP) антикроличьи pAb и, наконец, мышиные моноклональные антитела (mAb) противCD54 человека (ICAM1). Антитела получали от Dakopatts (Glostrup, Дания). Анализ проточной цитометрией. Экспрессию белков UspA1/A2 и способность М. catarrhalis связываться с фибронектином анализировали с помощью проточной цитометрии. Штаммы М. catarrhalis дикого типа и дефицитные в отношении UspA1/A2 мутанты выращивали в течение ночи и промывали дважды забуференным фосфатом солевым раствором, содержащим 3% рыбьего желатина (PBS-желатин). Затем бактерии (108) инкубировали с антисывороткой против UspA1/A2 или 5 мкг фибронектина (Sigma, St. Louis, МО). Затем их промывали и инкубировали в течение 30 мин при комнатной температуре с FITC-конъюгированными антикроличьимиpAb против фибронектина человека (если сначала добавляли фибронектин) в течение 30 мин при комнатной температуре перед инкубацией с FITC-конъюгированными антикроличьими pAb. После трех дополнительных промывок бактерии анализировали с помощью проточной цитометрии (EPICS, XL-MCL,Coulter, Hialeah, FL). Все инкубации проводили в конечном объеме 100 мкл PBS-желатин, а промывки осуществляли с использованием того же буфера. Для каждого анализируемого штамма в качестве отрицательного контроля использовали pAb против фибронектина и FITC-конъюгированные антикроличьиpAb. Исследования ингибирования фибронектина проводили с помощью предварительного инкубирования в течение 1 ч 0,25 мкмоль фрагментов UspA с 2 мкг фибронектина до инкубации с бактериями М.catarrhalis (108). Оставшееся свободным количество фибронектина, который связывался с М. catarrhalis,определяли с помощью проточной цитометрии, как отмечено выше. Связывание М. catarrhalis с иммобилизованным фибронектином. Предметные стекла покрывали 30 мкл-аликвотными пробами фибронектина (1 мг/мл) и высушивали на воздухе при комнатной температуре. После промывки один раз PBS предметные стекла инкубировали в чашках Петри с предварительно охлажденными бактериями в поздней экспоненциальной фазе(оптическая плотность (OD) при 600 нм = 0,9). После 2 ч при комнатной температуре предметные стекла промывали один раз PBS с последующим окрашиванием по Граму. Мечение белков и радиоиммуноанализ (РИА). Фибронектин метили 125 йодом (Amersham, Buckinghamshire, England) до высокой специфической активности (0,05 моль йода на моль белка) методом с использованием хлорамина Т [21]. Штаммы ВВН 18 и RH4 М. catarrhalis вместе с их соответствующими мутантами выращивали в течение ночи на твердой- 16014352 среде и промывали PBS с 2% бычьего сывороточного альбумина (BSA). Бактерии (108) инкубировали в течение 1 ч при 37 С с 125I-меченным фибронектином (1600103 имп/мин/образец) в PBS, содержащем 2% BSA. После трех промывок PBS с 2% BSA 125I-меченный фибронектин, связанный с бактериями, измеряли в гамма-счетчике (Wallac, Espoo, Финляндия). Твердофазный иммуноферментный анализ (ELISA) Микротитрационные планшеты (Nunc-Immuno Module; Roskilde, Дания) покрывали в течение ночи при 4 С 40 нМ очищенных рекомбинантных белков UspA150-770 и UspA230-539 в 75 мМ натрия карбоната,рН 9,6. Планшеты промывали четыре раза буфером для промывок (50 мМ Tris-HCl, 0,15 М NaCl и 0,1% твин-20, рН 7,5) и блокировали в течение 2 ч при комнатной температуре буфером для промывок, содержащим 3% рыбьего желатина. После четырех дополнительных промывок лунки инкубировали в течение 1 ч при комнатной температуре с фибронектином (120 мкг/мл), последовательно разведенным трехквадратично каждый раз в 1,5% рыбьем желатине (в буфере для промывок). После этого планшеты промывали и инкубировали с кроличьими pAb против фибронектина человека в течение 1 ч. После дополнительных промывок добавляли HRP-конъюгированные антикроличьи pAb и инкубировали в течение 1 ч при комнатной температуре. И pAb против фибронектина человека, и HRP-конъюгированные антикроличьиpAb разводили 1:1000 в буфере для промывок, содержащем 1,5% рыбьего желатина. Лунки промывали четыре раза и планшеты проявляли и измеряли OD450. ELISA с усеченными белками, охватывающимиUspA150-770 и UspA230-539, выполняли с использованием установленных доз фибронектина при 80 мкг/мл и 120 мкг/мл соответственно. Анализ ингибирования адгезии клеточной линии Клетки конъюнктивы Чанг (АТСС CCL 20.2) культивировали в среде RPMI 1640 (Gibco BRL, LifeTechnologies, Paisley, Шотландия), дополненной 10% фетальной телячьей сыворотки, 2 мМ L-глутамина и 12 мкг гентамицина/мл. За день до экспериментов по ингибированию адгезии клетки собирали, дважды промывали свободной от гентамицина средой RPMI 1640 и добавляли в 96-луночные планшеты для культуры тканей (Nunc) в конечной концентрации 104 клеток/лунку в 200 мкл свободной от гентамицина культуральной среды. После этого клетки инкубировали в течение ночи при 37 С в увлажненной атмосфере 5% CO2 и 95% воздуха. В день эксперимента ингибирование адгезии М. catarrhalis проводили с помощью предварительной инкубации возрастающих концентраций рекомбинантных усеченных белковUspA1/A2, содержащих связывающие фибронектин домены (UspA1229-452 и UspA2165-318), или кроличьихpAb против фибронектина человека (разведенных 1:50) в течение 1 ч. В качестве контролей использовали не связывающие фибронектин рекомбинантные белки (UspA1433-580 и UspA230-177). Известно, что эпителиальные клетки Чанг экспрессируют ICAM1 [18]. Следовательно, антитела против ICAM1 использовали для определения, является ли ингибиторный эффект антител против фибронектина вторичным по отношению к стерическому препятствию. Впоследствии на конфлюэнтные монослои наносили М. catarrhalis RH4 (106) в PBS-желатине. Во всех экспериментах планшеты для культуры тканей центрифугировали при 3000g в течение 5 мин и инкубировали при 37 С в 5% СО 2. Через 30 мин инфицированные монослои ополаскивали несколько раз PBSжелатином для удаления не прикрепившихся бактерий и затем обрабатывали трипсином-EDTA (0,05% трипсина и 0,5 мМ EDTA) для отсоединения клеток Чанг от пластиковой подложки. После этого полученные в результате клетки/бактериальную суспензию помещали при разведении на чашки, содержащиеBHI-агар, и инкубировали в течение ночи при 37 С в 5% СО 2. Определение экспрессии фибронектина в эпителиальных клетках конъюнктивы Чанг Эпителиальные клетки конъюнктивы Чанг собирали соскобом с последующим ресуспендированием в PBS-желатине. Клетки (1106/мл) метили с использованием кроличьих pAb против фибронектина человека с последующими промывкой и инкубацией с FITC-конъюгированными антикроличьими pAb. После трех дополнительных промывок клетки анализировали с помощью проточной цитометрии, как отмечено выше. Взаимодействие между М. catarrhalis и ламинином Бактериальные штаммы и условия культивирования. Клинические штаммы ВВН 18 и RH4 М. catarrhalis и их соответствующие мутанты описаны ранее[58]. Оба штамма экспрессируют относительно больше UspA2, чем UspA1 [58]. Мутанты экспрессировали равное количество связывающего IgD белка М. catarrhalis (MID) по сравнению со штаммами дикого типа. Бактерии в заведенном порядке культивировали в жидком бульоне с мозговым и сердечным экстрактами (BHI) или на чашках с BHI-агаром при 37 С. Дефицитные в отношении UspA1 мутанты, дефицитные в отношении UspA2 мутанты и двойные мутанты культивировали в BHI, дополненном антибиотиками, как описано [58]. Конструирование и экспрессия рекомбинантных белков. Были получены рекомбинантные UspA150-770 и UspA230-539, лишенные своих гидрофобных С-концов- 17014352 Антитела. Использовали кроличьи поликлональные антитела (pAb) против UspA1/A2 и MID [22, 58]. Кроличьи pAb против ламинина получали от Sigma (St Louis, МО, США). Свиные конъюгированные с пероксидазой хрена (HRP) антикроличьи pAb получали от Dakopatts (Glostrup, Дания). Связывание М. catarrhalis с иммобилизованным ламинином. Микротитрационные планшеты (Nunc-Immuno Module; Roskilde, Дания) покрывали в течение ночи при 4 С ламинином саркомы мыши Engelbreth-Holm-Swarm (Sigma, Saint Louis, США) или бычьим сывороточным альбумином (BSA) (30 мкг/мл) в Tris-HCl, pH 9. Планшеты промывали забуференным фосфатом солевым раствором и 0,05% твин-20, рН 7,2 (PBS-твин) и впоследствии блокировали 2% BSA вPBS + 0,1% твин-20, рН 7,2. Затем добавляли М. catarrhalis RH4 и ВВН 18 (108) в 100 мкл с последующей инкубацией в течение 1 ч. Несвязанные бактерии удаляли промывкой 3 раза PBS-твином. Оставшиеся связанными бактерии определяли посредством pAb против MID с последующим обнаружением с помощью HRP-конъюгированных антикроличьи pAb. Планшеты проявляли и измеряли OD450 в соответствии со стандартным протоколом. Твердофазный иммуноферментный анализ (ELISA) Микротитрационные планшеты (Nunc-Immuno Module) покрывали при 4 С 40 нМ очищенных рекомбинантных белков UspA150-770 и UspA230-539 в 75 мМ натрия карбоната, рН 9,6. Планшеты промывали четыре раза буфером для промывок (50 мМ Tris-HCl, 0,15 М NaCl и 0,1% твин-20, рН 7,5) и блокировали при комнатной температуре буфером для промывок, содержащим 3% рыбьего желатина. После дополнительных промывок лунки инкубировали в течение 1 ч при комнатной температуре с ламинином в различных разведениях, как указано, в 1,5% рыбьем желатине (в буфере для промывок). После этого планшеты промывали и инкубировали с кроличьими pAb против ламинина. После дополнительных промывок добавляли HRP-конъюгированные антикроличьи pAb и инкубировали при комнатной температуре. ИOD450. В качестве являющихся фоном контролей использовали непокрытые лунки, проинкубированные с идентичными разведениями ламинина. ELISA с усеченными белками, охватывающими UspA150-770 иUspA230-539, выполняли с использованием установленных доз ламинина (20 мкг/мл). Взаимодействие между М. catarrhalis и С 3 и C3met Бактериальные штаммы и условия культивирования. Недавно были подробно описаны клинические изоляты М. catarrhalis и родственные подвиды [21,53]. Типовые штаммы получали из коллекции культур, University of Gothenburg (CCUG; Department ofClinical Bacteriology, Sahlgrenska Hospital, Gothenburg, Швеция) или американской коллекции типовых культур (АТСС; Manassas, Va); Neisseria gonorrheae CCUG 15821, Streptococcus pyogenes CCUG 25570 и 25571, Streptococcus agalactiae CCUG 4208, Streptococcus pneumoniae ATCC 49619, Legionella pneumophilaATCC 33152, Pseudomonas aeruginosa ATCC 10145, Staphylococcus aureus ATCC 29213 и, наконец, Staphylococcus aureus ATCC 25923. Остальные штаммы, представленные в табл. 9, были клиническим изолятами из Medical Microbiology, Department of Laboratory Medicine, Malmo University Hospital, Lund University, Швеция. Таблица 9M. catarrhalis является уникальной связывающей C3/C3met бактерией. Родственные подвидыMoraxella и другие частые патогены человека не связывают C3/C3met (средняя интенсивность флуоресценции (MFI) 2,0). После инкубации с EDTA-обработанной NHS или C3met бактерии анализировали с помощью проточной цитометрии с использованием кроличьих pAb против C3d и FITCконъюгированных козьих антикроличьих pAb. Различные не являющиеся Moraxella виды выращивали в соответствующих стандартных средах для культивирования. Штаммы M. catarrhalis в заведенном порядке культивировали в жидком бульоне с мозговым и сердечным экстрактами (BHI) или на чашках с BHI-агаром при 37 С. Мутанты М. catarrhalisBBH18 и RH4 получали, как описано ранее [22, 23, 58]. Дефицитные в отношении MID мутанты выращивали в BHI, содержащем 50 мкг/мл канамицина. Дефицитные в отношении UspA1 мутанты культивировали в BHI, дополненном 1,5 мкг/мл хлорамфеникола (Sigma, St. Louis, МО), а дефицитные в отношении UspA2 мутанты инкубировали с 7 мкг/мл зеоцина (Invitrogen, Carlsbad, CA). Для выращивания двойных мутантов UspA1/A2 использовали и хлорамфеникол, и зеоцин. Антитела Кроликов иммунизировали внутримышечно 200 мкг рекомбинантного полноразмерного UspAl,эмульгированного в полном адъюванте Фрейнда (Difco, Becton Dickinson, Heidelberg, Германия), и на 18 и 36 день вторично инъецировали такую же дозу белка в неполном адъюванте Фрейнда [22]. Кровь брали спустя 3 недели. Для увеличения специфичности антисыворотку против UspA1 подвергали аффинной очистке с использованием коъюгированного с Сефарозой рекомбинантного UspA150-770 [58]. Антисыворотка в одинаковой степени связалась с UspA1 и UspA2, и поэтому ее обозначили pAb против UspA1/A2. Кроличьи pAb против C3d человека и свиные конъюгированные с FITC антикроличьи pAb покупали уDakopatts (Glostrup, Дания), а козьи антитела против С 3 человека получали от Advanced Research Technologies (San Diego, CA). Ослиные конъюгированные с пероксидазой хрена (HRP) антикозьи pAb получали от Serotec (Oxford, Соединенное Королевство). Белки и мечение йодом Недавно было описано получение рекомбинантных UspA150-770 и UspA230-539, лишенных своих гидрофобных C-концов [23]. Усеченные белки UspA1 и UspA2 получали, как подробно описано Tan и др.[78]. С 3b покупали у Advanced Research Technologies. С 3 (H2O) получали с помощью замораживания и оттаивания очищенного С 3. Подобную С 3b молекулу (C3met) получали с помощью инкубации очищенного С 3 со 100 мМ метиламина (рН 8,0) в течение 2 ч при 37 С и последующим диализом против 100 мМTtis-HCl (рН 7,5), 150 мМ NaCl. Для исследований связывания C3met метили 0,05 моль 125I (Amersham,Buckinghamshire, England) на моль белка методом с использованием хлорамина Т [25]. Анализ проточной цитометрией Связывание С 3 с М. catarrhalis и другими видами анализировали с помощью проточной цитомет- 19014352 рии. Бактерии выращивали на твердой среде в течение ночи и промывали дважды PBS, содержащим 2%BSA (Sigma) (PBS-BSA). После этого бактерии (108 колониеобразующих единиц; cfu) инкубировали сPBS-BSA в течение 30 мин при 37 С. После промывок бактерии инкубировали с pAb против C3d человека в течение 30 мин на льду с последующими отмывками и инкубацией в течение дополнительных 30 мин на льду с козьими конъюгированными с FITC антикроличьими pAb. После трех дополнительных промывок бактерии анализировали с помощью проточной цитометрии (EPICS, XL-MCL, Coulter, Hialeah,FL). Все инкубации проводили в конечном объеме 100 мкл PBS-BSA, а промывки осуществляли с использованием того же буфера. Для каждого анализируемого штамма в качестве отрицательного контроля отдельно добавляли pAb против C3d человека и FITC-конъюгированные антикроличьи pAb. В исследованиях ингибирования сыворотку предварительно инкубировали со 100 мМ рекомбинантных белковUspA150-770 и UspA230-539 в течение 30 мин при 37 С. Для анализа свойств взаимодействия С 3 с М. catarrhalis к бактериям и C3met добавляли возрастающие концентрации NaCl (0-1,0 М). Для анализа экспрессии UspA1/A2 бактерии (108 cfu) инкубировали с pAb против UspA1/A2 и промывали, как описано выше. Для обнаружения использовали козьи FITC-конъюгированные антикроличьи pAb, разведенные в соответствии с инструкциями производителя. Для гарантии того, что EDTA не разрушало белки внешней мембраны UspA1 и UspA2, M. catarrhalis инкубировали с или без EDTA с последующим определением экспрессии UspA1/A2. При концентрациях, используемых в экспериментах с NHS-EDTA, EDTA не изменял плотность UspA1/A2. Сыворотка и бактерицидный анализ с использованием сыворотки Нормальную сыворотку человека (NHS) получали от пяти здоровых добровольцев. Крови предоставляли возможность коагулировать в течение 30 мин при комнатной температуре и после этого инкубировали на льду в течение 60 мин. После центрифугирования сыворотки объединяли, разливали по аликвотным пробам и хранили при -70 С. Для инактивации и классического, и альтернативного путей добавляли 10 мМ EDTA. Напротив, для инактивации классического пути включали Mg-EGTA. Сыворотку человека, дефицитную в отношении С 4 ВР, готовили пропусканием свежей сыворотки через колонку HiTrap (Amersham Biosciences) с присоединенными mAb 104, мышиными mAb, направленными против ССР 1 -цепи С 4 ВР [41]. Не связавшийся материал собирали и истощенную сыворотку хранили в аликвотных пробах при -70 С. Истощенную в отношении Clq сыворотку получали через первый шаг очисткиC1q [79] с использованием ионообменной хроматографии на Biorex 70 (Bio-Rad, Hercules, CA). Получаемая в результате сыворотка проявляла нормальную гемолитическую активность. Сыворотка, дефицитная в отношении фактора D и пропердина, была любезно предоставлена Dr. Anders Sjholm (Department of(DGVB). Бактерии (108 cfu) инкубировали вместе с 10% NHS и EDTA или Mg-EGTA в конечном объеме 100 мкл. Бактерии/NHS инкубировали при 37 С, и в различные моменты времени брали 10 мклаликвотные пробы и распределяли по чашкам с BHI-агаром. При исследовании ингибирования до добавления бактерий 10% сыворотку инкубировали со 100 нМ рекомбинантных белков UspA150-770 иUspA230-539 в течение 30 мин при 37 С. Дот-блот анализы Очищенные рекомбинантные белки UspA150-770 и UspA230-539, последовательно разведенные трехквадратично каждый раз (1,9-150 нМ), в 100 мкл 0,1 М Tris-HCl, рН 9,0 наносили на нитроцеллюлозные мембраны (SchleicherSchll, Германия), используя дот-блот устройство. После пропитки мембраны инкубировали в течение 2 ч с PBS-твин, содержащим 5% молочного порошка, при комнатной температуре и промывали четыре раза PBS-твин. После этого добавляли 5103 имп/мин [125I]-меченного C3met вPBS-твин с 2% молочного порошка и инкубировали в течение ночи при 4 С. Связанный белок визуализировали с помощью Personal FX (Bio-Rad) с использованием усиливающих экранов. Резонанс поверхностных плазмонов (Biacore) Взаимодействие между UspA150-770 или UspA230-539 и С 3 далее анализировали с помощью резонанса поверхностных плазмонов (Biacore 2000; Biacore, Uppsala, Швеция), как недавно описано для взаимодействия UspA1/2-C4BP [58]. KD (константу диссоциации при равновесии) рассчитывали из кривой связывания, представленной на графике зависимости ответа при равновесии от концентрации, используя модель аффинности в стационарном состоянии, предоставляемую в программном обеспечении Biaevaluation(Biacore). Твердофазный иммуноферментный анализ (ELISA) Микротитрационные планшеты (Nunc-Immuno Module; Roskilde, Дания) покрывали в течение ночи при 4 С в трех повторах очищенными рекомбинантными белками UspA150-770, UspA230-539 или усеченными фрагментами UspA1 и UspA2 (40 нМ в 75 мМ натрия карбоната, рН 9,6). Планшеты промывали четыре раза буфером для промывок (PBS с 0,1% твин-20, рН 7,2) и блокировали в течение 2 ч при комнатной температуре буфером для промывок, дополненным 1,5% овальбумина (блокирующим буфером). После промывок лунки инкубировали в течение ночи при 4 С с 0,25 мкг C3met в блокирующем буфере. После- 20014352 этого планшеты промывали и инкубировали с козьими антителами против С 3 человека в блокирующем буфере в течение 1 ч при комнатной температуре. После дополнительных промывок добавляли ослиныеHRP-конъюгированные антикозьи pAb и инкубировали в течение еще 1 ч при комнатной температуре. Лунки промывали четыре раза и планшеты проявляли и измеряли OD450. Гемолитическое исследование Эритроциты кролика промывали трижды охлажденным на льду 2,5 мМ Veronal-буфером, рН 7,3,содержащим 0,1% (вес./об.) желатина, 7 мМ MgCl2, 10 мМ EGTA и 2,5% декстрозы (MgEGTA), и ресуспендировали при концентрации 0,5109 клеток/мл. Эритроциты инкубировали с различными концентрациями (от 0 до 4%) сыворотки, разведенной в MgEGTA. Спустя 1 ч при 37 С эритроциты центрифугировали и количество лизированных эритроцитов определяли с помощью спектрофотометрического измерения высвободившегося гемоглобина при 405 нм. Для ингибирования белками UspA1 и UspA2 10% сыворотку предварительно инкубировали со 100 нМ рекомбинантных белков UspA150-770 и/илиUspA230-539 в течение 30 мин при 37 С и после этого добавляли к эритроцитам при 0-4%. Выделение нейтрофильных лейкоцитов и фагоцитоз Нейтрофильные лейкоциты (PMN) человека выделяли из свежей крови здоровых добровольцев, используя Macrodex (Pharmalink AB, Upplands Vsby, Швеция). PMN центрифугировали в течение 10 мин при 300g, отмывали в PBS и ресуспендировали в среде RPMI 1640 (Life Technologies, Paisley, Шотландия). Суспензию бактерий (0,5108) подвергали опсоническому влиянию 3% или нормальной сыворотки человека (NHS), или NHS-EDTA, или 20 мкг очищенного C3met в течение 15 мин при 37 С. После промывок бактерии смешивали с PMN (1107 клеток/мл) в соотношении бактерии/PMN = 1:10 с последующей инкубацией при 37 С при непрерывным вращении. Выжившие после 0, 30, 60 и 120 мин инкубации бактерии определяли с помощью счета жизнеспособных бактерий. Число поглощенных обработанныхNHS бактерий сравнивали с бактериями, подвергнутыми фагоцитозу в отсутствие NHS. В качестве положительного контроля использовали S. aureus, подвергнутый опсоническому влиянию нормальной сыворотки человека. Примеры и результаты Взаимодействие между М. catarrhalis и фибронектином М. catarrhalis, лишенным UspA1 и А 2, не связывается с растворимым или иммобилизованным фибронектином. Был отобран произвольный ряд клинических штаммов М. catarrhalis (n=13) (табл. 7) и проведена их проверка на связывание фибронектина относительно экспрессии ими UspA1/A2 с помощью анализа проточной цитометрией. Высокая экспрессия UspA1/A2, определенная по высокой средней интенсивности флуоресценции (MFI), коррелировала с экспрессией UspA1/A2 (коэффициент персональной корреляции 0,77, Р 0,05) (фиг. 1 А). Однако используя pAb против UspA1/A2, было невозможно провести дифференциацию между экспрессией UspA1 и А 2. Более того, являлось невероятным присутствие белка UspA2H,вносящего вклад в связывание, поскольку в штаммах, используемых в этом исследовании, ген UspA2H не был обнаружен (данные не показаны). Два изолята М. catarrhalis (BBH18 и RH4) и их специфические мутанты, у которых отсутствовалBBH18 сильно связывал фибронектин со средней интенсивностью флуоресценции (MFI), составляющей 96,1 (фиг. 1F). Напротив BBH18AuspA1 продемонстрировал уменьшенное связывание с фибронектином с MFI, составляющей 68,6 (фиг. 1G). При связывании фибронектина с BBH18AuspA2 и двойным мутантом BBH18AuspA1/A2 обнаружилась MFI, составляющая только 10,7 и 11,5 соответственно (фиг. 1 Н, 1I). Похожие результаты были получены с мутантами по UspA1/А 2 клинического штамма RH4 М. catarrhalis. Вместе взятые эти результаты говорят о том, что UspA1 и А 2 связывают фибронектин, и что способность бактерий связывать фибронектин сильно зависит от экспрессии UspA1/A2. Для дальнейшего анализа взаимодействия между фибронектином и М. catarrhalis 125I-меченный фибронектин инкубировали с двумя клиническими изолятами М. catarrhalis (BBH18 и RH4) и их соответствующими мутантами. М. catarrhalis RH4 дикого типа сильно связывал 125I-фибронектин, в то время как соответствующий мутант AuspA1 продемонстрировал 80% связывания дикого типа. Напротив, AuspA2 и двойной мутант связывали 125I-фибронектин при 14 и 12% соответственно, что было как раз выше уровней фона (5-10%) (фиг. 2). Похожие результаты были получены с М. catarrhalis BBH18 и соответствующими мутантами по UspA1/A2. Следовательно, результаты говорят о том, что для максимального связывания растворимого фибронектина бактериями М. catarrhalis требуется и UspA1, и А 2. Для исследования присоединения бактерий к иммобилизованному фибронектину М. catarrhalis RH4 и его соответствующие мутанты AuspA1/A2 наносили на покрытые фибронектином предметные стекла. После инкубации в течение 2 ч предметные стекла промывали и впоследствии окрашивали по Граму. Обнаружено, что М. catarrhalis дикого типа и мутант AuspA1 сильно адгезировались к покрытым фибронектином предметным стеклам (фиг. 3 А и 3 В). Напротив, М. catarrhalis AuspA2 и двойные мутантыAuspA1/A2 слабо адгезировались к покрытому фибронектином предметному стеклу, при этом только малое число бактерий оставалось после промывки (фиг. 3 С и 3D соответственно). Эксперименты с другим клиническим изолятом М. catarrhalis (BBH18) и полученными из него мутантами продемонстрирова- 21014352 ли схожую картину, что указывает на то, что UspA2 имеет главное значение при связывании М. catarrhalis с иммобилизованным фибронектином. Связывающие фибронектин домены включают аминокислотные остатки, находящиеся между 299 и 452 UspA1 и между 165 и 318 UspA2. Для дальнейшего анализа взаимодействий UspA1 и А 2 с фибронектином в Е. coli рекомбинантно получили усеченные UspA150-770 и UspA230-539, ими покрывали микротитрационные планшеты и инкубировали с возрастающими концентрациями фибронетина. Связанный фибронектин обнаруживали с помощью pAb против фибронектина человека с последующей инкубацией с конъюгированными с пероксидазой хрена антикроличьими pAb. Оба рекомбинантных UspA150-770 и UspA230-539 связывали растворимый фибронектин, и взаимодействия зависели от дозы (фиг. 4). Для определения связывающего фибронектин домена UspA1 получили рекомбинантные белки, охватывающие целую молекулу UspA150-770. Фибронектин инкубировали с иммобилизованными фрагментами белков UspA1 и взаимодействия количественно определяли с помощью ELISA. UspA150-491 связывал фибронектин почти также эффективно, как и UspA150-770, что говорит о том, что связывающий домен находится в пределах этой части белка. Среди других усеченных фрагментов UspA1299-452 эффективно связывал фибронектин (фиг. 5 А). Параллельно анализировали взаимодействия между фибронектином и несколькими рекомбинантными фрагментами UspA2, включающими аминокислоты UspA230-539. Два фрагмента UspA2101-318 и UspA2165-318 сильно связывали фибронектин (фиг. 5 В). Открытия обеспечивают значимое доказательство того, что связывающие домены включают остатки, обнаруживаемые в пределахUspA1299-452 и UspA2165-318. Сравнение последовательностей этих двух связывающих фрагментов позволило обнаружить, что для UspA1 и А 2 идентичными являются 31 аминокислотных остатковDQKADIDNNINNIYELAQQQDQHSSDIKTLK (фиг. 6). Более того, эта повторяющаяся последовательность также была обнаружена в гене UspA1 и А 2 М. catarrhalis BBH18 и RH4 (данные не показаны). Фрагменты UspA150-491 и UspA1299-452 конкурентно ингибируют связывание М. catarrhalis с фибронектином. Для дальнейшего подтверждения открытий в отношении связывающих фибронектин доменов рекомбинантные усеченные белки UspA1 проверяли в отношении их способности блокировать связывание фибронектина с М. catarrhalis. Фибронектин (2 мкг) предварительно инкубировали с 0,25 мкмоль рекомбинантных фрагментов UspA1 и впоследствии инкубировали с М. catarrhalis. Наконец, UpsA-зависимое связывание М. catarrhalis с фибронектином измеряли с помощью проточной цитометрии. Предварительная инкубация с UspA150-431 и UspA1299-452 приводила к уменьшению связывания фибронектина с уменьшением на 95% для UspA150-491 и уменьшением на 63% для UspA1299-452 (фиг. 7). Когда фибронектин предварительно инкубировали с усеченным UspA2101-318, получали ингибирование на 50%. Таким образом, связывающие фибронектин домены UspA1 и А 2 блокируют взаимодействия между фибронектином и М. catarrhalis.UspA1299-452 и UspA2165-318 ингибируют адгезию М. catarrhalis к эпителиальным клеткам Чанг. Известно, что эпителиальные клетки экспрессируют фибронектин, и многие бактерии прикрепляются к эпителиальным клеткам через ассоциированный с клетками фибронектин [46, 54, 69, 77]. Предварительные исследования показали, что М. catarrhalis адгезируется к эпителиальным клеткам [46, 49]. Были проанализированы клетки конъюнктивы Чанг, которые часто использовали в экспериментах по адгезии с патогенами дыхательных путей. Клетки Чанг сильно экспрессировали фибронектин, что обнаружено с помощью анализа проточной цитометрией (фиг. 8 А). Для анализа важности для адгезии бактерий UpsA-зависимого связывания с фибронектином эпителиальные клетки Чанг предварительно инкубировали с pAb против фибронектина человека или рекомбинантными белками UspA1299-452 и UspA2165-318. После этого добавляли М. catarrhalis RH4 и анализировали адгезию бактерий. Относительная адгезия (измеренная по числу колониеобразующих единиц) после предварительной инкубации с 0,4 мкмоль на 200 мкл UspA1299-452, UspA2165-318 или pAb против фибронектина человека составляла 36, 35 и 32% соответственно. Более высокие концентрации рекомбинантных пептидов не приводили к дальнейшему ингибированию. Напротив, не связывающие фибронектин фрагменты UspA1433-580 и UspA230-177 не ингибировали взаимодействия между М. catarrhalis и эпителиальными клетками Чанг(фиг. 8 В). Таким образом, фибронектин на эпителиальных клетках Чанг может функционировать в качестве рецептора для М. catarrhalis, a аминокислотные остатки 299-452 UspA1 и 165-318 UspA2 содержат лиганд, ответственный за взаимодействия. Взаимодействие между М. catarrhalis и ламинином. М. catarrhalis связывает ламинин через UspA1 и А 2. Два клинических изолята М. catarrhalis (BBH18 и RH4) и их специфические мутанты, у которых отсутствовал UspA1, UspA2 или оба белка, анализировали с помощью ELISA с использованием целых клеток. М. catarrhalis RH4 сильно связывался с иммобилизованным ламинином (фиг. 9 А). Напротив, мутант по uspA1 M. catarrhalis RH4 (RH4AuspA1) продемонстрировал связывание с ламинином, составляющее 89,9% дикого типа. Мутант по uspA2 М. catarrhalisRH4 (RH4AuspA2) и двойной мутант RH4AuspA1/A2 обладали 15,2 и 18,1% связывающей способности дикого типа соответственно. Это незначительно отличалось от остающейся необъясненной адгезии к- 22014352 покрытым BSA планшетам. Похожие результаты были получены с мутантами по UspA1/A2, происходящими из клинического штамма ВВР 18 М. catarrhalis. В этих двух штаммах (ВВН 18 и RH4) UspA2 является преимущественно экспрессируемым белком по сравнению с UspA1, что является объяснением минимальной разницы в связывании между диким типом и RH4AuspA1. Вместе взятые эти результаты показывают, что UspA1 и А 2 связывают ламинин. Для дальнейшего анализа связывания UspA1/A2 с ламинином в Е. coli получили усеченныеUspA150-770 и UspA230-539. Рекомбинантными белками покрывали микротитрационные планшеты и инкубировали с возрастающими концентрациями ламинина. Связанный ламинин обнаруживали с помощью кроличьих pAb против ламинина с последующей инкубацией с HRP-конъюгированными антикроличьими pAb. Оба рекомбинантных UspA150-770 и UspA230-539 сильно связывали растворимый ламинин, и связывание было зависимым от дозы и насыщаемым (фиг. 9 В). Для определения связывающих ламинин доменов получали рекомбинантные белки UspA1 и А 2, охватывающие целые молекулы. Ламинин инкубировали с иммобилизованными усеченными фрагментами белков UspA1 и А 2 и затем количественно определяли с помощью ELISA. UspA150-431 связывал ламинин почти также эффективно, как и UspA150-770, что говорит о том, что связывающий домен находится в пределах этой части белка. Однако среди других усеченных фрагментов, охватывающих эту область, ни один из них, по-видимому, не связывал ламинин. N-концевая часть, UspA230-351, была способна сохранять 44,7% связывающей способности полноразмерного белка. Более короткий белок UspA230-177 продемонстрировал 43,7% связывающей способности (фиг. 10B). Эти результаты показывают, что связывающие домены включают остатки, обнаруживаемые в N-концах и UspA1, и UspA2. Взаимодействие между М. catarrhalis и С 3 и C3met Белки внешней мембраны UspA1 и UpsA2 M. catarrhalis ингибируют и классический, и альтернативный путь активации комплемента. Поверхностная экспрессия UpsA2 является решающей для выживания М. catarrhalis в нормальной сыворотке человека (NHS) [1, 58], т.е. дефицитные в отношении UpsA2 мутанты Moraxella быстро погибают под воздействием NHS. Было недавно продемонстрировано, что и UspA1, и А 2 связываются с C4 ВР и, следовательно, могут ингибировать классический путь активации комплемента [58]. Чтобы далее пролить свет на взаимодействия М. catarrhalis с системой комплемента, исследовали выживание двойных мутантов по UspA1/A2 в сыворотке, обработанной или EGTA с добавлением MgCl2 (Mg-EGTA), илиEDTA. Mg-EGTA ингибирует классический и лектиновый пути и, следовательно, делает возможным отдельный анализ альтернативного пути. Напротив, EDTA ингибирует все пути активации комплемента в результате абсорбции двухвалентных катионов (Mg2+ и Са 2+). М. catarrhalis RH4 дикого типа выживал после 30 мин инкубации, в то время как двойной мутант RH4AuspA1/A2 погибал под влиянием интактной NHS после 10 мин (фиг. 12). При ингибировании классического пути (NHS + Mg-EGTA) мутантRH4AuspA1/A2 жил в течение значительного большего периода времени по сравнению с NHS без какихлибо хелатообразующих агентов, но не так долго, как бактерия дикого типа. Более того, при блокировании EDTA и классического, и альтернативного путей М. catarrhalis RH4AuspA1/A2 выживал. Схожую картину получали с изолятом М. catarrhalis BBH18 и соответствующими мутантами BBH18AuspA1/A2(не показано). Параллельно эксперименты с C1q и дефицитной в отношении фактора D/пропертина сывороткой продемонстрировали, что и классический, и альтернативный путь ингибировались М. catarrhalis(не показано). Таким образом, М. catarrhalis, патоген, который часто заселяет дыхательные пути человека, не только нейтрализует классический путь, но также и альтернативный путь системы активации комплемента с помощью белков внешней мембраны UspA1 и А 2. М. catarrhalis абсорбирует С 3 из EDTA-инактивированной сыворотки. С 3 ковалентно присоединяется к поверхности микроба и, отсюда, индуцируют альтернативный путь (фиг. 11 В). Для анализа того, может ли М. catarrhalis взаимодействовать с С 3, штамм RH4 дикого типа инкубировали с NHS или NHS, обработанной EDTA. Присоединение или осаждение (через ковалентную связь) С 3/С 3b на поверхности бактерий М. catarrhalis RH4 определяли с помощью анализа проточной цитометрией с использованием поликлонального антитела (pAb), направленного против C3d, узнающего и С 3, и С 3b. Инкубация бактерий с NHS, содержащей интактный комплемент, приводила к осаждению С 3 (фиг. 13). Интересно, что, когда путь активации комплемента был инактивирован в присутствии EDTA, M. catarrhalis RH4 все еще связывал С 3 (фиг. 13 А). Streptococcus pneumoniae, включенный для сравнения, не абсорбировал С 3 из обработанной EDTA сыворотки (фиг. 13 В). В противоположность пневмококки М. catarrhalis, таким образом, связывал С 3 безотносительно активации комплемента. Внутренний тиоэфир С 3 спонтанно гидролизуется в жидкой фазе до С 3(H2O). Следовательно, интактный С 3 или С 3(H2O) были наиболее вероятными формами С 3, взаимодействующими с М. catarrhalis. Поскольку M. catarrhalis также связывает С 4 ВР [58], хотели исключить вовлечение С 4 ВР в связывание С 3 и с этой целью использовали истощенную в отношении С 4 ВР сыворотку. М. catarrhalis абсорбировал С 3 из истощенной в отношении С 4 ВР сыворотки в той же степи, что и из NHS (не показано). Связывание C3met с М. catarrhalis является зависимым от дозы и нековалентным. Из экспериментов можно заключить, что С 3 связывается с поверхностью М. catarrhalis безотносительно активации комплемента. Поэтому проанализировали, может ли преобразованный С 3, не являю- 23014352 щийся функциональным, связываться с бактериями. Нативный С 3 очищали из сыворотки человека и обрабатывали метиламином, который преобразует С 3 в молекулу C3met, эквивалентную С 3b, не обладающую способностью ковалентно связываться с микробами (фиг. 11 С). С помощью анализа проточной цитометрией обнаружено, что штамм RH4 М. catarrhalis дикого типа эффективно связывал C3met зависимым от дозы и насыщаемым способом (фиг. 14 А и В). Это взаимодействие не опосредовалось С 3, а частью молекулы С 3, поскольку С 3b и С 3(H2O) также связывались с М. catarrhalis (не показано). Связывание М. catarrhalis RH4 с C3met основывалось в большой степени на ионных взаимодействиях, поскольку возрастающие концентрации NaCl ингибировали взаимодействие (фиг. 14 С). Похожие результаты были получены со штаммом BBH18 М. catarrhalis дикого типа (не показано). Для определения того, является ли связывание С 3 общей особенностью всех штаммов M. catarrhalis,отобрали произвольный ряд клинических изолятов (n=13) и проанализировали их способность связыватьC3met. Все штаммы М. catarrhalis связывали C3met, как обнаружено с помощью анализа проточной цитометрией с использованием pAb против C3d. Значения MFI варьировали от 4 до 39. Однако S. pneumoniae и Е. coli, включенные для сравнения, не связывали C3met. М. catarrhalis является уникальной связывающей С 3 и C3met бактерией. Чтобы продолжить анализы в отношении абсорбции С 3 из NHS к бактериям, родственные подвидыMoraxella (n=13), а также частые патогены человека (n=13) инкубировали в присутствии NHS-EDTA. Интересно, что среди всех проверенных бактериальных видов М. catarrhalis был единственным видом бактерий, связывающим С 3 в комплемент-инактивированной сыворотке (табл. 9). Все родственные штаммы Moraxella, а также другие патогены человека также проанализировали на связывание C3met. Параллельно со связыванием С 3 М. catarrhalis был единственным видом, связывающим C3met. Обобщая,М. catarrhalis имеет уникальную особенность сильно связывать С 3 и C3met нековалентным способом. М. catarrhalis связывает C3met через белки внешней мембраны UspA1 и UspA2. Для определения белка М. catarrhalis, ответственного за связывание С 3, проверили ряд бактериальных мутантов, лишенных белков внешней мембраны MID, UspA1 и/или UspA2 [22, 58]. Интересно, что связывание C3met значительно коррелировало с экспрессией Ups (фиг. 15). М. catarrhalis RH4Amid связывал C3met в той же степени, что и соответствующий штамм дикого типа (фиг. 15 А-В). МутантRH4AuspA1 продемонстрировал только слегка сниженное связывание, в то время как RH4AuspA2 был слабым связующим по сравнению с соответствующим штаммом дикого типа (фиг. 15 С-D). Параллельно полностью отсутствовало связывание C3met с двойным мутантом RH4AuspA1/A2 (фиг. 15 Е). Более того,когда те же эксперименты проводили с использованием NHS-EDTA, наблюдали ту же картину (фиг. 15FJ). При использовании нормальной сыворотки человека все мутанты продемонстрировали на своей поверхности схожее количество С 3, поскольку это была смесь ковалентного осаждения и связывания С 3(фиг. 15K-O). Похожие результаты были получены с изолятом М. catarrhalis BBP18 и соответствующими мутантами ВВР 18. Для дальнейшего анализа взаимодействия между С 3 и UspA1/A2 в Е. coll получили UspA150-770 иUspA230-533 и их очистили. Рекомбинантные белки наносили на нитроцеллюлозную мембрану с использованием устройства для дот-блота с последующей инкубацией с меченным йодом C3met. В качестве отрицательного контроля включили рекомбинантный MID362-1200, который получен из белка MID внешней мембраны [58]. Обнаружили слабое связывание с UspA150-770, в то время как [125I]-C4met сильно связывался с UspA230-533 (фиг. 16 А). Эти открытия в дальнейшем подкреплены с использованием резонанса поверхностных плазмонов (т.е. Biacore). UspA150-770 и UspA230-533 иммобилизовали на поверхности СМ 5 чипа, используя связывание через аминогруппу, и C3met вносили до достижения насыщения. KD для взаимодействия между C3met и UspA150-770 или UspA230-533 составляла 3 и 14 мкМ соответственно. В заключение, было обнаружено, что UspA2 является главным связывающим C3met белком М. catarrhalis, в то время как UspAl вносит вклад в меньшей степени. Связывающий С 3 домен находится между аминокислотными остатками 200 и 458 UspA2. Для определения связывающего С 3 домена UspA2 получали рекомбинантные белки, охватывающие целую молекулу UspA230-539. C3met инкубировали с иммобилизованными полноразмерными белкамиUspA150-770, UspA230-539 и рядом усеченных белков UspA2. После этого взаимодействия количественно определяли с помощью ELISA. В соответствии с дот-блот анализом (фиг. 16 А) UspA150-770 связывалC3met в гораздо меньшей степени, чем UspA230-533 в ELISA (фиг. 16 В). Среди усеченных фрагментов белка UspA2165-318, UspA2200-539 и UspA2302-458 эффективно связывали C3met, что говорит о том, что связывающий домен находится в пределах аминокислотных остатков 200 и 458. Рекомбинантный UspA1/A2 нейтрализует активность С 3. Для того чтобы детально исследовать роль зависимого от UspA1/A2 ингибирования альтернативного пути, провели ряд анализов проточной цитометрией с использованием бактерий, проинкубированных с 10% NHS или сывороткой, которую предварительно проинкубировали со 100 нМ рекомбинантногоUspA150-770 и UspA230-533. Интересно, что значительное снижение осаждения/связывания С 3 на поверхности М. catarrhalis RH4AuspA1/A2 наблюдали, когда NHS предварительно обрабатывали UspA150-770 иUspA230-539 (фиг. 17 А). При выключении классического пути с помощью Mg-EGTA получали похожие результаты (фиг. 17 В). Таким образом, рекомбинантные белки UspA1350-770 и UspA230-539 абсорбировали- 24014352 С 3 из NHS и ингибировали осаждение/связывание С 3. Для определения того, увеличивает ли абсорбция С 3 рекомбинантными UspA150-770 и UspA230-539 выживаемость бактерий, двойной мутант М. catarrhalis RH4AuspA1/A2 инкубировали с сывороткой, дополненной UspA150-770 и UspA230-539, с последующим определением числа выживших бактерий. MgEGTA включали в реакции для того, что ингибировать классический путь. Интересно, что добавление рекомбинантных UspA150-770 и UspA230-539 к NHS предотвращало гибель дефицитных в отношенииUspA1/A2 М. catarrhalis (фиг. 17 С). UspA230-539 был наиболее эффективен в ингибировании гибели бактерий по сравнению с UspA150-770. Когда оба рекомбинантных белка добавляли вместе, дополнительного ингибирования альтернативного пути не определялось. 10% NHS соответствует приблизительно 600 нМ С 3. Для исследования того, может ли большее число молекул UspA1 нейтрализовать активность С 3, добавляли UspA150-770 и/или UspA230-531 до 600 нМ. Однако более высокие концентрации рекомбинантных белков не увеличивали дополнительно ингибирование (не показано). Был также включен гемолитический анализ альтернативного пути, состоящий из эритроцитов кролика и NHS, для установления роли UspAl и А 2 в качестве ингибиторов альтернативного пути. NHS предварительно инкубировали с рекомбинантными UspA150-770, UspA230-539 или обоими белками вместе с последующим добавлением эритроцитов. После инкубации в течение 1 ч определяли величину лизиса эритроцитов. Интересно, что значительно сниженный гемолиз наблюдали, когда NHS предварительно инкубировали с UspA150-770 или UspA230-539 по сравнению с необработанной NHS (фиг. 18). Параллельно с увеличением выживаемости бактерий в присутствии UspA150-770 или UspA230-539 (фиг. 17 С) предварительная инкубация только с UspA230-539 приводила к более эффективному ингибированию альтернативного пути по сравнению с ингибированием в случае, когда NHS предварительно инкубировали с UspA150-77. В заключение, рекомбинантные UspA150-770 или UspA230-539 блокировали активность альтернативного пути благодаря их способности захватывать С 3. Помимо того, что С 3 является ключевой молекулой в пути активации комплемента, осажденные С 3b и iC3b (инактивированный С 3b) направляет микробы для удаления в процессе опсонофагоцитоза. Для исследования того, может ли С 3 или C3met, который нековалентно связан с поверхностью М. catarrhalis, все еще функционировать в качестве опсонина, провели ряд исследований фагоцитоза. М. catarrhalis предварительно инкубировали с C3met, NHS или NHS, обработанной EDTA, с последующим добавлением нейтрофильных лейкоцитов. Интересно, что М. catarrhalis не поглощался в присутствииC3met, в то время как NHS сильно индуцировала фагоцитоз (данные не представлены). Однако, когда NHS предварительно обрабатывали EDTA, М. catarrhalis не фагоцитировался нейтрофильными лейкоцитами. Таким образом, C3/C3met был не активен на поверхности клеток М. catarrhalis и не функционировал в качестве опсонина. Осуждение. Взаимодействие между М. catarrhalis и фибронектином.UspA1299-452 и UspA2165-318 из клинического штамма Вс 5 М. catarrhalis были самыми короткими фрагментами, которые все еще связывали фибронектин. Интересно, что более длинные фрагменты,включающие аминокислотную последовательность, обнаруживаемую в пределах UspA1299-452 и UspA2165318, проявляли более эффективное связывание с фибронектином (фиг. 5 А и В). Это может означать, что эти две области представляют частичные связывающие домены, или что сайт связывания в высокой степени зависит от специфической молекулярной структуры. UspA1299-452 и UspA2165-318 имеют последовательность из 31 идентичных аминокислотных остатков, включающую 23 остатка NNINNIYELAQQQDQHSSDIKTL (последовательность NNINNIY). Эта последовательность содержит эпитоп для протективного моноклонального антитела (mAb) 17C7, которое обладает общей реактивностью [2, 50,30]. В мышиной модели пассивная иммунизация mAb 17C7 обеспечивала защиту и увеличивала обезвреживание М. catarrhalis в легких [30]. Отсюда самое интересное то, что связывающие фибронектин домены UspA1/A2 содержат эти остатки, и это доказывает важность этой области в патогенезе инфицирования М. catarrhalis дыхательных путей. Связывающие фибронектин М. catarrhalis BBH18 и RP4, использованные в экспериментах, также несут указанный 31 аминокислотный остаток в их белке UspA1/A2. Большинство М. catarrhalis имеют часть этой последовательности (т.е. последовательность NNINNIY). Однако штаммы, подобные штамму О 35 Е, имеющему последовательность NNINNIY в гене UspA2, не экспрессируют связывающий фибронектин белок UspA2 [49]. Вероятным объяснением могло бы быть то, что вариации во фланкирующих областях могут влиять на взаимодействие с фибронектином. Также сама консервативная последовательность NNINNIY может иметь минорные единичные изменения в аминокислотных основаниях [28]. Таким образом, вероятно, что связывание с фибронектином зависит не просто от экспрессии UspA1/A2, но также от индивидуальной организации каждого белка UpsA. Интересно, что почти идентичную аминокислотную последовательность можно обнаружить в гибридном белке UspA2H с адгезивными свойствами (М. catarrhalis TTA37 и 046 Е) [43]. Это служит поддержкой открытию того, что последовательность из 31 аминокислоты важна для адгезии. В последнем наборе экспериментов было проверено, можно ли ингибировать адгезию М. catarrhalis к клеткам конъюнктивы Чанг с помощью связывающих фибронектин фрагментов (UspA1299-452 иUspA2165-318) (фиг. 8 В). Предварительная инкубация с UspA1299-452, UspA2165-318 или pAb против фибронектина приводила к снижению связывания с эпителиальными клетками Чанг. Эти результаты подтверждают важность этих связывающих доменов для взаимодействий UspA1/A2 с эпителиальными клетками Чанг и, кроме того, говорят о том, что фибронектин является важным рецептором для UpsA. Кроме того,известно, что FnBP облегчают адгезию бактерий к недифференцированным и поврежденным дыхательным путям [54, 69]. Экспрессию фибронектина фибробластами легких также увеличивает дым от сигарет[87]. Связывание UspA1/A2 М. catarrhalis с фибронектином ЕСМ или ассоциированным с эпителиальными клетками фибронектином имеет, следовательно, важное значение у больных COPD и может служить объяснением частых случаев инфекции М. catarrhalis в этой группе больных [40]. В заключение, было показано, что UspA1/A2 M. catarrhalis ВВН 18, RH4 и Вс 5 являются решающими FnBP. Оба рекомбинантных UspA1 и А 2, происходящих из Вс 5, связывают фибронектин с помощью связывающего домена, имеющего идентичные аминокислотные остатки, в том числе консервативную последовательность NNINNIY. Более того, взаимодействие UspA1/A2 М. catarrhalis с эпителиальными клетками происходит через ассоциированный с клетками фибронектин. Следовательно, определение этих связывающих фибронектин доменов представляет собой важный шаг к разработке вакцины против М. catarrhalis. Взаимодействие между М. catarrhalis и ламинином. М. catarrhalis является частой причиной обострений инфекций у больных COPD. Вероятно, успех этого вида у больных COPD отчасти связан с наличием у него большого набора адгезивных молекул. Кроме того, существуют патологические изменения, такие как потеря целостности эпителия с обнаружением базальной мембраны, где у курильщиков утолщен сам слой ламинина [4]. Показано, что некоторые патогены способны связывать ламинин, и это может вносить вклад в их способность адгезироваться к таким поврежденным и денудированным поверхностям слизистой оболочки. Эти патогены включают патогены, которые, как известно, вызывают значимое заболевание дыхательных путей, такие как среди прочих S. aureus и P. aeruginosa [7, 63]. Авторы настоящего изобретения недавно показали, что и UspAl, и А 2 связывают фибронектин [78]. Связывающие фибронектин домены находятся в пределах UspA1299-452 и UspA2165-318. В этом исследовании N-концевые половины UspA150-491 и UspA230-351 (содержащие связывающие фибронектин домены) также связывали ламинин. Однако самые маленькие фрагменты, которые связывали фибронектин,UspA1299-452 и UspA2165-318, не связывали ламинин в какой-либо заметной степени. В действительности фрагменты, меньшие чем N-концевая половина UspA1 (UspA150-491), теряют всю свою способность связывать ламинин, в то время как в случае UspA2 только UspA230-170 связывал ламинин, хотя на более низком уровне, чем целый рекомбинантный белок (UspA230-539). Эти открытия говорят о том, что различные части молекул, возможно, могут иметь разные функциональные роли. При сравнении самых маленьких связывающих ламинин областей UspA1 и А 2 было обнаружено,что существует, однако, незначительная схожесть, между прочим аминокислотная гомология, междуUspA230-170 и UspA150-491 (данные не показаны). Это не удивительно, поскольку известен тот факт, что оба белка имеют структуру сферической головки в виде "леденца", несмотря на то, что их N-концевые половины идентичны только на 22% [2, 32]. Авторы настоящего изобретения постулируют, что за in vivo взаимодействия с ламинином в области головки, вероятно, ответственна третичная структура. Нахождение связывающих доменов в N-конце было бы логичным, поскольку они были бы наиболее выставленными и контактировали с базальной мембраной человека in vivo. Бактериальные факторы, опосредующие адгезию к компонентам ткани и внеклеточного матрикса(ЕСМ), группируются вместе в одно семейство, названное "микробные поверхностные компоненты, узнающие адгезивные матриксные молекулы (MSCRAMM). Поскольку UspA1/A2 связывают и фибронектин, и ламинин, эти белки можно обозначить MSCRAMM. Результаты говорят о том, что UspA1 и А 2 являются многофункциональными адгезинами с различными доменами, взаимодействующими с различными лигандами в дыхательных путях. О похожих профилях связывания широкого спектра сообщалось для других бактериальных белков, таких как YadA Yersinia enterocolitica, с которым UspA1 и UspA2 структурно связаны [45, 70]. YadA также связывает и фибронектин, и ламинин [32]. В итоге было показано, что UspA1/A2 являются решающими во взаимодействии М. catarrhalis с являющимся гликопротеином ламинином базальной мембраны, и это играет важную роль в патогенезе инфицирования больных COPD [74]. Взаимодействие между M. catarrhalis и С 3 и C3met. Устойчивость к комплементу является один из самых важных факторов вирулентности бактерий[66]. Большинство (89%) изолятов М. catarrhalis от больных с инфекциями нижних дыхательных путей устойчивы к опосредуемому комплементом уничтожению [34]. UspA1 и А 2 М. catarrhalis являются решающими для выживания бактерий в сыворотке человека in vivo [1, 15], и было показано, что эти два белка внешней мембраны связываются с жидкофазным регулятором классического пути активации комплемента, С 4 ВР [58]. В настоящем исследовании продемонстрировано, что М. catarrhalis может ингибировать альтернативный путь с помощью нековалентного связывания С 3 (фиг. 17 и 18). Связывание С 3,наиболее вероятно, также ингибирует классический путь. Однако это невозможно было детально про- 26014352 анализировать, поскольку М. catarrhalis также связывает С 4 ВР. Интересно, что зависимое от М. catarrhalis связывание С 3 является уникальным, так как несколько родственных подвидов Moraxella, а также частые патогенные для человека бактерии не связывают С 3 (табл. 9). Взаимодействия с С 3 и обработанным метиламином С 3 опосредуются главным образом UspA2, в то время как UspA1 имеет минорную роль (фиг. 15 и 16). Связывающая С 3 область UspA2 находится между аминокислотными остатками 200458. Эта область содержит участок из 140 аминокислотных остатков, которые на 93% идентичны области в UspA1 [2]. Однако несмотря на эту схожесть последовательностей, UspA1 связывает С 3 в гораздо меньшей степени. Это может быть следствием специфических различий в конформации белков. Различие в связывании С 3 белками UspA1 и UspA2 контрастирует со взаимодействием UspA1/A2 с С 4 ВР [58].M. catarrhalis одинаково устойчив и к классическому, и к альтернативному пути (фиг. 12 В). Бактерии связывают С 4 ВР, что ингибирует классический путь [58], и в этом документе продемонстрировано взаимодействие с альтернативным путем благодаря связыванию С 3. Трудно определить, какой из этих механизмов имеет наибольшее значение для устойчивости М. catarrhalis в сыворотке при различных ситуациях in vivo. Например, важность классического пути сильно зависит от анамнеза инфицирования М.catarrhalis и способности вырабатывать комплемент-активирующие антитела. Однако всякий обеспечивающий защиту от комплемента механизм, конечно, полезен для патогена. Поскольку С 3 является ключевой молекулой системы комплемента, наиболее вероятно, что связывание С 3 приводит к регулированию всех трех путей активации комплемента и может вносить наибольший вклад в устойчивость в сыворотке. Важность системы комплемента в качестве первичного защитного механизма отражает тот факт,что микробы выработали различные стратегии блокирования и/или нейтрализации компонентов системы комплемента [42, 35, 88]. В дополнение к М. catarrhalis специфические поверхностные молекулы, которые связывают С 4 ВР и, как следствие, защищают бактерии от классического пути активации комплемента, экспрессируют S. pyogenes, Bordetella pertussis, E. coli K1, Candida albicans и N. gonorrhoeae [8, 9, 52,58, 64, 65, 80]. В дополнение к ингибированию классического пути несколько бактерий (например, С.albicans, N. meningitides, S. pyogenes и S. pneumoniae; для обзора см. [68, 89]) связывают фактор Н и схожую с фактором Н молекулу и, следовательно, частично защищаются от альтернативного пути активации комплемента.UspA1 и А 2 абсорбируют С 3 из сыворотки и, следовательно, наиболее вероятно, ингибируют активацию комплементы. Схожим образом поверхностный белок А пневмококков (PspA), по-видимому, ингибирует альтернативный путь и in vitro, и in vivo. PspA является важным фактором вирулентности S.pneumoniae. Дефицитные в отношении PspA штаммы пневмококков легко обезвреживаются в крови, в то время как экспрессирующие PspA штаммы выживают [82]. Кроме того, в крысиной модели бактериемии дефицитные в отношении PspA пневмококки имеют значительно сниженную вирулентность по сравнению с пневмококками, экспрессирующими PspA [11]. Продемонстрировано, что на PspA-отрицательных пневмококках осаждается больше С 3b, чем на PspA-положительных пневмококках [67, 82]. Таким образом, экспрессия PspA уменьшает опосредуемое комплементом обезвреживание и фагоцитоз S. pneumoniae посредством предельной опсонизации с помощью С 3b [12, 67]. Дефицитные в отношении PspA пневмококки, не являющиеся вирулентными для нормальных мышей, становятся вирулентными для мышей, дефицитных в отношении С 3 и фактора В [82]. Насколько известно, существует только два примера бактериальных белков, которые нековалентно связывают С 3 и отсюда блокируют функцию комплемента. Первым белком является внеклеточный связывающий фибронектин белок Staphylococcus aureus (Efb), который, как обнаружено, связывает С 3b [44].Efb ингибирует и классический, и альтернативный путь независимо от тиоэфирной конформации, т.е. связывание с С 3b не является ковалентным. Вторым примером является связывающий холин белок пневмококков (CbpA), который, как показано, связывает обработанный метиламином С 3, что говорит о нековалентном взаимодействии, которое не зависит от активации комплемента [16]. CbpA является компонентом клеточной стенки пневмококков, но может только связывать С 3 при секреции CbpA. Чтобы проверить эту гипотезу, для которой в литературе нет твердого доказательства, было проанализировано одиннадцать различных изолятов пневмококков на связывание С 3 (обработанного метиламином С 3 илиNHS-EDTA) с помощью проточной цитометрии (фиг. 12 В и табл. 9). Связанный С 3 невозможно было обнаружить на поверхности S. pneumoniae. С помощью анализа лизатов S. pneumoniae и супернатантов культур вестерн-блоттингом с использованием обработанного метиламином С 3 с последующим использованием pAb против С 3 человека авторы настоящего изобретения подтвердили результаты Cheng и соавторов [16] (не показано). В свете Efb и CbpA, оба из которых являются связывающими С 3 белками,секретируемыми грамположительными бактериями, грамотрицательный М. catarrhalis является уникальным видом с закрепленными на мембране белками, которые связывают С 3 и ингибируют альтернативный путь активации комплемента на поверхности бактерии. Показано, что дрожжи Candida albicans связывают С 3b, iC3b и C3d. Однако С 3b связывается со значительно меньшим сродством, чем iC3b и C3d [29]. Была обнаружена большая разница между связыванием С 3 с М. catarrhalis и С. albicans (не показано); несмотря на то, что Candida связывали C3met (56% положительных клеток), средняя интенсивность флуоресценции (MFI) составляла только 2,0 по сравне- 27014352 нию с MFI, составляющей 36,9 для М. catarrhalis. Кроме того, связывание не определялось, когда С. albicans инкубировали с обработанной EDTA сывороткой. Из С. albicans были выделены два связывающих С 3 белка, и наиболее охарактеризованным белком является маннобелок с молекулярной массой 60 кДа,который первоначально определили с помощью антитела, направленного против рецептора 2 комплемента человека (CD21) [13]. Однако UspA1 и А 2 М. catarrhalis не узнавались поликлональным антителом,направленным против CD21 (не показано). Параллельно со стафилококками и пневмококками существует также связывающий C3d белок, секретируемый С. albicans [72]. Наконец, выделили рецептор для iC3b С. albicans, и он структурно схож с CR3 человека (CD11b) [3]. Механизмы, с помощью которых эти рецепторы могут участвовать в патогенезе, полностью не известны. Вышеуказанные примеры связывающих С 3 патогенов весьма отличаются от М. catarrhalis тем, что эти виды часто выделяют из кровотока. М. catarrhalis является патогеном слизистых оболочек с редкими случаями бактериемических инфекций. Следовательно, связывание и инактивация С 3, наиболее вероятно, происходят на поверхности слизистых оболочек. Это подтверждается тем фактом, что происходит сильная активация комплемента и являющееся следствием этого воспаление при таком болезненном состоянии, как острое воспаление среднего уха [57]. Полагают, что белки комплемента транспортируются на поверхность слизистых оболочек благодаря экссудации плазмы [26, 62]. В выпотах в полость среднего уха (МЕЕ), например, у детей также обнаруживаются сильно увеличенные концентрации продуктов С 3[51]. Кроме того показано, что факторы комплемента в жидкости МЕЕ важны для бактерицидной активности против других агентов слизистых оболочек, таких как не типируемые H. influenzae [75]. М. catarrhalis является исключительно патогеном человека. Он не вызывает заболеваний, таких как воспаление среднего уха или пневмония, у животных. В нескольких случаях использовали мышиную модель очищения легких и модель воспаления среднего уха с использованием шиншиллы. Однако ни воспаления среднего уха, ни пневмонии не развивается, и бактерии быстро обезвреживаются [19, 83]. Таким образом,трудно проверить биологическое значение связывания С 3 с бактериями in vivo. Поскольку UspA1 и А 2 являются многофункциональными белками [1, 15, 31, 43, 58, 78], невозможно соотнести какое-либо различие в обезвреживании М. catarrhalis со связыванием С 3. В частности, тот факт, что UspA1 является важным адгезином М. catarrhalis и связывает и СЕАСАМ 1, и фибронектин [31, 78], будет, наиболее вероятно, оказывать влияние на обезвреживание. Тем не менее, благодаря сильной активации комплемента при таких болезненных состояниях, как воспаление среднего уха, зависимое от Moraxella связывание С 3,может представлять важный способ снятия защиты слизистых оболочек. Тот факт, что М. catarrhalis тормозит иммунную систему человека несколькими путями, может служить объяснением того, почему М. catarrhalis является таким частым обитателем дыхательных путей[73]. В заключение, М. catarrhalis выработал усложненные способы противостояния гуморальной и врожденной иммунным системам. Представленные данные показывают, что М. catarrhalis обладает уникальной способностью связывать С 3 на поверхности бактериальной клетки, что невозможно обнаружить у других бактериальных видов.
МПК / Метки
МПК: C07K 14/21, A61P 31/04, A61K 39/104
Метки: эпителиальными, взаимодействие, матриксными, moraxella, внеклеточными, белками, клетками, catarrhalis, системой, комплемента
Код ссылки
<a href="https://eas.patents.su/30-14352-vzaimodejjstvie-moraxella-catarrhalis-s-epitelialnymi-kletkami-vnekletochnymi-matriksnymi-belkami-i-sistemojj-komplementa.html" rel="bookmark" title="База патентов Евразийского Союза">Взаимодействие moraxella catarrhalis с эпителиальными клетками, внеклеточными матриксными белками и системой комплемента</a>
Полипептид белка-106 наружной мембраны moraxella catarrhalis, днк, его кодирующие, и их применение

Номер патента: 2743
Опубликовано: 29.08.2002
Авторы: Такер Кеннет, Плосила Лаура
МПК: A01N 63/00, C07H 21/02, A61K 39/00...
Метки: наружной, moraxella, белка-106, днк, полипептид, кодирующие, catarrhalis, применение, мембраны
Формула / Реферат:
1. Выделенный полипептид ОМP 106, который является полипептидом наружной мембраны Moraxella catarrhalis и имеет молекулярную массу приблизительно от 180 кДа до приблизительно 230кДа, как определено электрофорезом в SDS-полиакриламидном геле с использованием миозина скелетных мышц кролика и 3-галактозидазы E.coli в качестве стандартов молекулярных масс 200 кДа и 116,25 кДа соответственно, и включает либо последовательность SEQ ID NО:1, либо...
Способ получения яичного желтка, обогащенного натуральными антисекреторными белками, яичный желток, полученный данным способом, и продукты, содержащие указанный желток

Номер патента: 4644
Опубликовано: 24.06.2004
Авторы: Ланге Стефан, Йеранссон Лейф, Леннрот Ивар
МПК: A23L 1/305, A61K 35/54, A23K 1/16...
Метки: яичный, натуральными, содержащие, данным, желтка, белками, антисекреторными, яичного, обогащенного, указанный, способ, получения, способом, полученный, желток, продукты
Формула / Реферат:
1. Способ получения от яйценесущей птицы яичного желтка, обогащенного натуральными антисекреторными белками (NASP), предусматривающий, что яйценесущей птице дают корм, индуцирующий образование NASP. 2. Способ по п.1, отличающийся тем, что птица представляет собой птицу отряда куриных. 3. Способ по п.1 или 2, отличающийся тем, что корм, индуцирующий образование NASP, содержит аминокислоты, сахара и амиды в количестве, стимулирующем образование...
Препарат выделенных кератиноцитов, носитель, покрытый этими клетками, способ криоконсервации препарата и способ лечения ран

Номер патента: 9073
Опубликовано: 26.10.2007
Авторы: Райф Катарина, Эберхардт Петра, Нё Вольфганг
МПК: C12N 5/06, A61L 27/60, A61P 17/02...
Метки: способ, криоконсервации, носитель, препарата, этими, клетками, препарат, кератиноцитов, ран, выделенных, покрытый, лечения
Формула / Реферат:
1. Препарат выделенных кератиноцитов человека, которые представляют собой клетки культуры KC-BI-1 (DSM АСС 2514) или производные данных клеток, являются не иммортализованными и могут реплицироваться по меньшей мере 150 раз при выращивании in vitro. 2. Препарат по п.1, где репликация кератиноцитов не возможна: а) в отсутствие фетальной телячьей сыворотки, и/или б) в отсутствие подкармливающих клеток, и/или в) в отсутствие эпидермального фактора...
Жевательная резинка, содержащая в покрытии натуральные растительные компоненты с неповрежденными клетками в качестве ароматического агента, способ получения жевательной резинки и применение указанныхнатуральных растительных компонентов в покрытии жевательной резинки

Номер патента: 4371
Опубликовано: 29.04.2004
Автор: Сталь Бронислав-Ян
МПК: A23G 3/30
Метки: ароматического, применение, качестве, клетками, покрытии, резинки, получения, указанныхнатуральных, натуральные, жевательная, компоненты, растительных, компонентов, растительные, способ, содержащая, агента, резинка, жевательной, неповрежденными
Формула / Реферат:
1. Жевательная резинка, содержащая a) нерастворимую жевательную основу, b) водорастворимую часть, c) покрытие, содержащее ароматический агент, в котором по меньшей мере 10 вес.% ароматического агента является натуральным растительным ароматическим веществом, содержащим, по существу, неповрежденные клетки растения. 2. Жевательная резинка по п.1, в которой натуральное растительное ароматическое вещество нанесено на покрытие в форме порошка. 3....
Биореактор со сдерживающей системой

Номер патента: 12226
Опубликовано: 28.08.2009
Автор: Лутц Петер
МПК: C12M 1/107
Метки: биореактор, системой, сдерживающей
Формула / Реферат:
1. Биореактор для метанирования биомассы, имеющий перегниватель (2), выполненный с возможностью его газонепроницаемого закрытия створкой (14) и предназначенный для размещения биомассы; систему дренажа просачивающегося вещества в полу и/или в стенках биореактора (2), при этом сдерживающее средство (100), которое взаимодействует, по меньшей мере, со сторонами перегневателя (2) и расположенное, если смотреть в направлении заполнения, за створкой, в...
Предыдущий патент: Сельскохозяйственная композиция
Следующий патент: Вакцинные композиции, содержащие сапониновый адъювант
Случайный патент: Установка погружная электрогидроприводная