Процедура получения клеточных линий в безбелковых средах и клеточные линии, полученные этим способом
Номер патента: 7465
Опубликовано: 27.10.2006
Авторы: Кастильо Витльоч Адольфо, Боджано Ае Тамми, Виторес Сарасола Свьета, Рохас Дель Кальво Луис, Перес Родригес Роландо





Формула / Реферат
1. Способ получения линии клеток млекопитающих, адаптированных к росту в бессывороточных и безбелковых средах, который включает две стадии:
I) первую стадию, в ходе которой показатели жизнеспособности клеток в клеточной линии составляют от 80 до 100% и клетки выращивают в культуральных средах, в которых последовательно снижают содержание белка до критической концентрации белка, при которой жизнеспособность клеток падает до 0%;
II) вторую стадию, в ходе которой, на основе уже определенной критической концентрации белка, поддерживают предкритическую концентрацию как такую концентрацию белка, при которой клеточный рост возможен, фиксируя ее как исходную точку для дальнейшего медленного снижения концентрации белка до момента, когда клеточная культура достигнет исходных показателей жизнеспособности клеток и времени удвоения клеток в популяции.
2. Способ по п.1, отличающийся тем, что первая стадия состоит из следующих стадий:
i) высеивают в 3 ячейки в шестиячеечном планшете для культивирования клеток рекомбинантную клеточную линию с использованием стандартной культуральной среды для культивирования клеток (с исходной концентрацией белка); плотность клеток должна составлять 1-5 х 105 клеток/мл; через 48 ч половину супернатанта заменяют свежей безбелковой средой, что приводит к достижению конечной концентрации белка, составляющей 50% от ее значения в исходных условиях;
ii) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 50% от исходных условий;
iii) клетки растят до слияния при данной белковой концентрации;
iv) клетки со стадии iii высеивают по меньшей мере в 3 ячейки с плотностью в диапазоне 1-5 х 105 клеток/мл в культуральной среде, в которой содержание белка составляет 50% от его концентрации в исходных условиях; через 48 ч половину супернатанта заменяют свежей безбелковой средой, что приводит к достижению конечной концентрации белка, составляющей 50% от ее предыдущего значения;
v) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 50% от ее предыдущего значения;
vi) клетки растят до слияния при данной белковой концентрации;
vii) стадии (iv)-(vi) повторяют, при этом в ходе каждого цикла концентрацию белка снижают до 50% относительно концентрации в предыдущем цикле; процедуру повторяют до достижения концентрации белка, которая вызывает гибель клеток.
3. Способ по п.1, отличающийся тем, что вторая стадия состоит из следующих стадий:
viii) клетки из клеточной культуры с показателями жизнеспособности 80% или выше высеивают с использованием предкритической концентрации белка по меньшей мере в 3 ячейки с плотностью в диапазоне 2-6 х 105 клеток/мл; клетки растят при предкритической концентрации белка и через 48 ч 25% супернатанта заменяют свежей безбелковой средой, с достижением при этом конечной концентрации белка, которая составляет 75% от предкритической концентрации белка;
ix) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 75% от предкритической концентрации белка;
х) клетки растят до слияния при данной белковой концентрации;
xi) клетки со стадии (х) высеивают по меньшей мере в 3 ячейки с плотностью 2-6 х 105 клеток/мл в культуральной среде, в которой содержание белка составляет 75% от предкритической концентрации белка; через 48 ч 25% супернатанта заменяют свежей безбелковой средой, с достижением при этом конечной концентрации белка, которая составляет 75% от концентрации на предыдущей стадии;
xii) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 75% от концентрации на стадии (х);
xiii) клетки растят до слияния при данной белковой концентрации;
xiv) стадии (xi)-(xiii) повторяют, при этом в ходе каждого цикла концентрацию белка снижают до 75% относительно концентрации в предыдущем цикле и далее процедуру повторяют до достижения концентрации белка, которая не вызывает потери жизнеспособности клеток и снижения времени удвоения клеток в популяции; когда клетки переносят в среду со сниженной концентрацией белка и они способны расти без потери жизнеспособности клеток и без снижения времени удвоения клеток в популяции перед первым субкультивированием, авторы считают, что такие клетки достигли снова некритической стадии, и высеивают их непосредственно в безбелковой среде (концентрация белка 0 мг/мл).
4. Способ по пп.1-3, где бессывороточная и безбелковая среда, в которую клетки первоначально высеивают, включает от 5 до 10% фетальной сыворотки теленка.
5. Способ по пп.1-4, где линия клеток млекопитающих, адаптированных для роста в бессывороточных и безбелковых средах, представляет собой миеломную линию.
6. Способ по п.5, где миеломная линия представляет собой клеточную линию NSO.
7. Способ по п.6, где указанная клеточная линия NSO содержит последовательность, кодирующую рекомбинантный полипептид или рекомбинантный белок.
8. Способ по п.7, где последовательность, кодирующая рекомбинантный полипептид или рекомбинантный белок, кодирует рекомбинантное антитело или его фрагмент.
9. Линия клеток млекопитающих, полученная способом по пп.1-8, где указанная клеточная линия адаптирована для роста в бессывороточных и безбелковых средах.
10. Линия клеток млекопитающих по п.9, где указанная линия клеток млекопитающих представляет собой миеломную линию.
11. Линия клеток млекопитающих по п.10, где указанная клеточная линия представляет собой клеточную линию NSO.
12. Линия клеток млекопитающих по п.11, где клеточная линия NSO содержит последовательность, кодирующую рекомбинантный полипептид или рекомбинантный белок.
13. Линия клеток млекопитающих по п.12, где последовательность, кодирующая рекомбинантный полипептид или рекомбинантный белок, кодирует рекомбинантное антитело или его фрагмент.
14. Линия клеток млекопитающих по п.13, где последовательность кодирует гуманизированное рекомбинантное анти-EGF-R hR3 антитело или его фрагмент.
15. Линия клеток млекопитающих по п.13, где последовательность кодирует гуманизированное рекомбинантное анти-СD6 T1hT антитело или его фрагмент.
16. Линия клеток млекопитающих по п.13, где последовательность кодирует химерное рекомбинантное анти-СD3 T3Q антитело или его фрагмент.
17. Применение способа по пп.1-8 для получения линии клеток млекопитающих, адаптированных для роста в бессывороточной и безбелковой среде.
18. Гуманизированное моноклональное антитело к EGF-R hR3 или его фрагменты, секретируемое клетками, полученными способом по пп.1-8.
19. Гуманизированное моноклональное антитело против антигена CD6 T1hT или его фрагменты, секретируемое клетками, полученными способом по пп.1-8.
20. Химерное моноклональное антитело против антигена CD3 T3Q или его фрагменты, секретируемое клетками, полученными способом по пп.1-8.
Текст
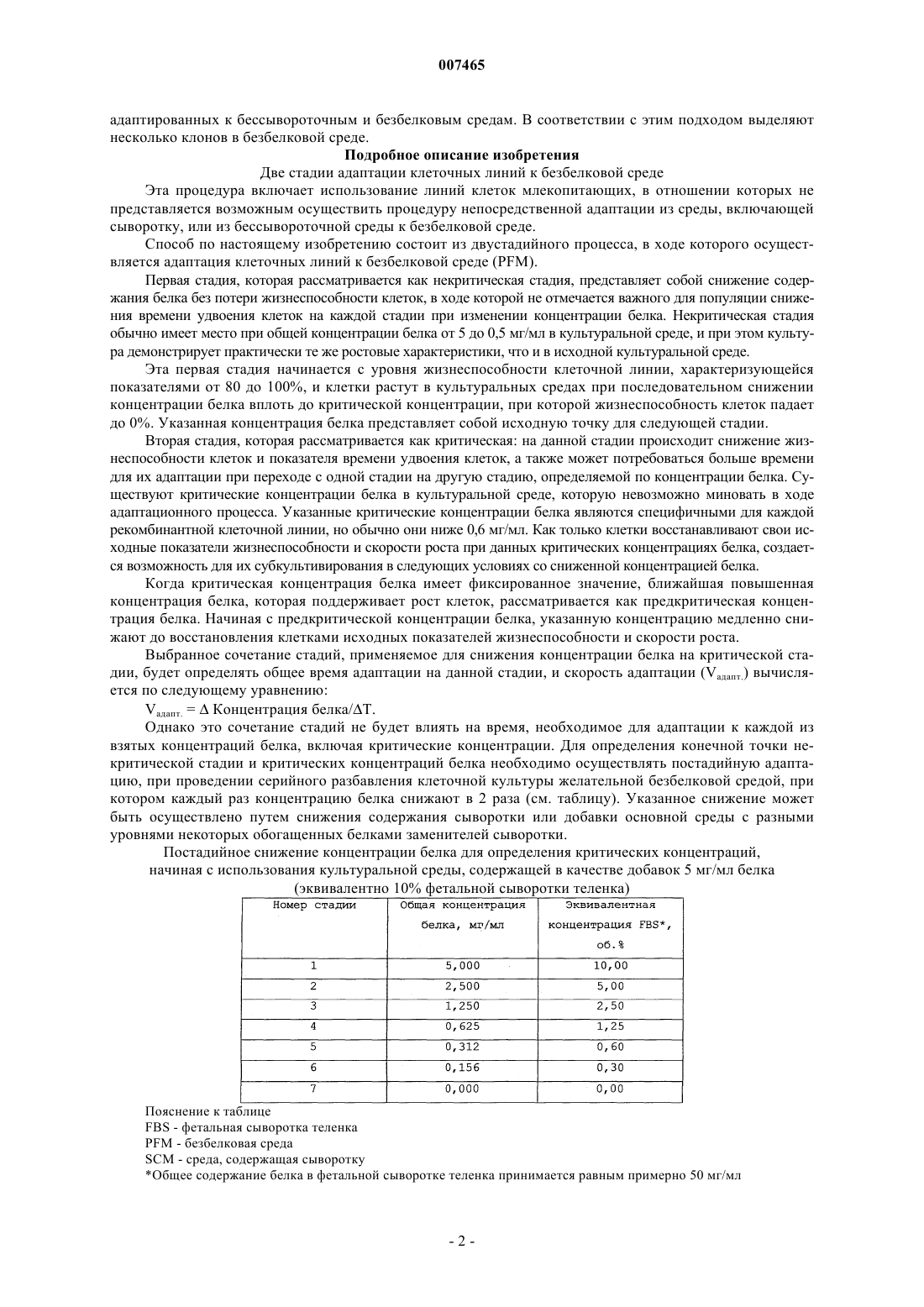
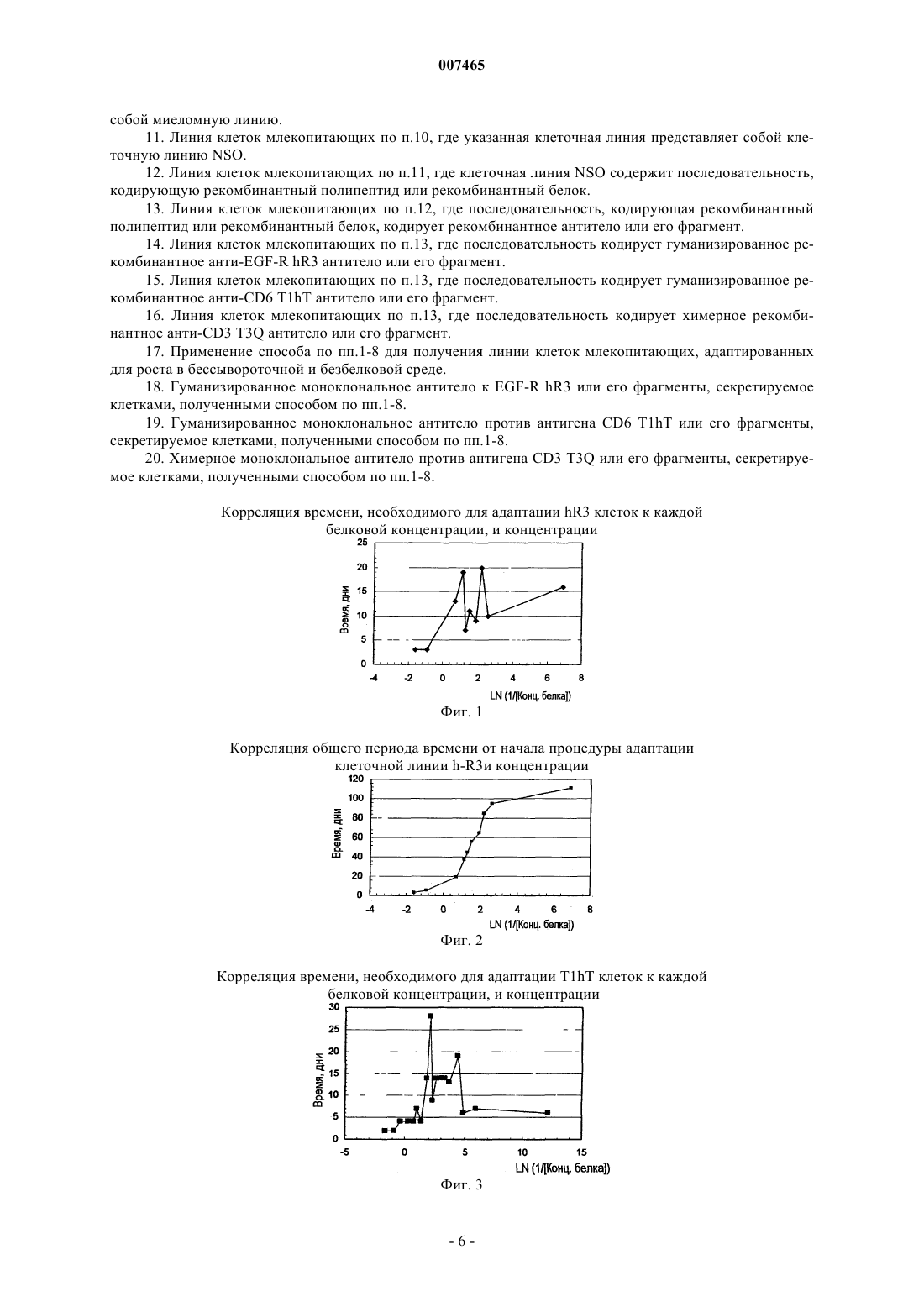
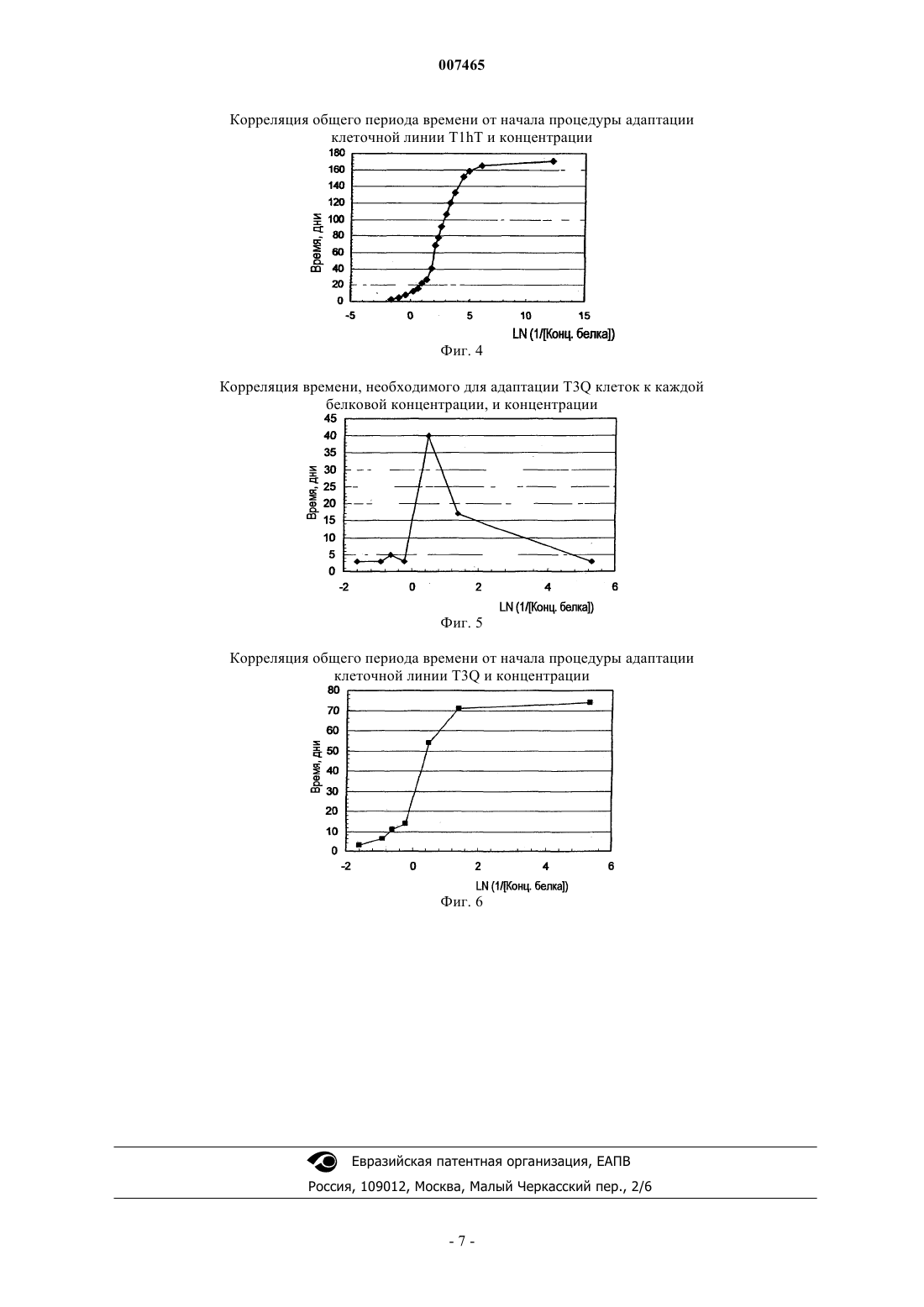
007465 Область техники, к которой относится изобретение Настоящее изобретение относится к биотехнологии, конкретно к способу получения стабильных клеточных клонов, адаптированных к бессывороточной и безбелковой среде в ходе двустадийного процесса адаптации. Предпосылки создания изобретения С момента начала внедрения методов культивирования клеток млекопитающих in vitro выросла потребность в крупномасштабном производстве таких клеток в связи с тем, что многие получаемые при этом продукты имеют диагностическое и терапевтическое применение. Указанные полезные продукты включают моноклональные антитела, гормон роста человека, лимфокины, эритропоэтин, факторы свертывания крови и тканевые активаторы плазминогена. Использование рекомбинантных моноклональных антител (rMab) при лечении и диагностике in vivo разных заболеваний включает во многих случаях применение для лечения высоких доз. Этот факт подводит к необходимости производства больших количеств необходимых rMab с очень высокой степенью чистоты. Некоторые рекомбинантные моноклональные антитела, которые имеют потенциал, позволяющий использовать их при лечении и диагностике рака и аутоиммунных заболеваний, экспрессируются в клетках миеломы NSO, получаемых в Центре молекулярной иммунологии (Center of Molecular Immunology). В патенте США 5891996 описывается получение химерных и гуманизированных антител против рецептора эпидермального фактора роста (EGF-R), применяемых при диагностике и лечении опухолей, экспрессирующих указанный рецептор. В WO 97/19111 описываются анти-СD6 моноклональные антитела,применяемые для диагностики и лечения псориаза у пациентов. Gavilondo et al. in Hybridoma 9, No. 5,1999 описали анти-СD3 моноклональные антитела, названные IOR-Т 3 а. Для культивирования в безбелковой среде были разработаны различные методики. Так, в одном конкретном случае, были разработаны полностью безбелковые среды, которые позволяют осуществлять выращивание клеток в безбелковых условиях. В WO 97/05240 описывается экспрессия рекомбинантных белков в безбелковых условиях. В JP 2696001 описывается использование безбелковой среды для получения фактора VIII в СНО клетках при добавлении неионного поверхностно-активного вещества или циклодекстрина для повышения продуктивной способности хозяйских клеток. Для повышения эффективности указанных добавок рекомендуется внесение, например, бутирата и лития. В WO 96/26266 описывается культивирование клеток в среде, которая включает глутаминсодержащий белковый гидролизат, в котором содержание свободных аминокислот составляет менее 15% от всего веса белка и в котором пептиды имеют вес менее 44 кДа. В качестве культуральной среды для культивирования клеточных культур используют синтетическую минимальную среду в качестве базовой среды,к которой, в дополнение к белковому гидролизату, добавляют, в том числе, фетальную сыворотку теленка, гентамицин и меркаптоэтанол. Использование среды, включающей сыворотку, для получения рекомбинантными методами факторов крови в указанном документе не рассматривается. В патенте США 5393668 А описываются специальные синтетические поверхности, которые позволяют растить прикрепляющиеся клетки в условиях безбелковой среды. Для стимуляции пролиферации клеток СНО клетки, которые осуществляют суперэкспрессию человеческого инсулина, размножают на искусственном субстрате, с которым ковалентно связывается инсулин (Ito et al., 1996 PNAS U.S.A 93:3598-3601).Reiter et al. (1992, Cytotechnology 9:247-253) описывают иммобилизацию r-CHO клеток, выращенных в бессывороточной среде с высокой плотностью, на носителях с последующей перфузией через иммобилизованные клетки безбелковой среды в ходе фазы продукции, при которой происходит непрерывное высвобождение белка в клеточный супернатант. Таким образом, клетки могут поддерживаться в течение менее 10 генераций в безбелковой среде. Способы, успешно применявшиеся ранее для крупномасштабного получения клеточных культур в безбелковых условиях, описаны применительно к клеточным линиям; в частности для VERO клеток (WO 96/15231). Таким образом, клетки могут расти в бессывороточных и безбелковых условиях, начиная с исходной ампулы до крупного технического процесса масштабом до 1200 л. Процесс адаптации клеток, которые вначале выращивались в условиях, включающих сыворотку, к безбелковой среде представляет собой трудную задачу, требующую обычно много времени; кроме того, неоднократно было показано, что выход экспрессированного белка и производительность рекомбинантных СНО клеток существенно снижаются при адаптации к безбелковым условиям в сравнении с условиями, включающими сыворотку (Paterson et al., 1994, Appl. Microbiol. Biotechnol. 40:691-658). Указанное явление представляет собой следствие нестабильности или сниженного роста рекомбинантных клонов в измененных условиях культивирования. Несмотря на использование стабильных исходных клонов, большая часть клеток снова становится клетками со сниженной экспрессией или даже непродуцентами, которые обгоняют в росте клеткипродуценты в ходе процесса продуцирования, за счет чего культура в ферментере в итоге полностью становится культурой непродуцирующих клеток или таких клеток, которые характеризуются низким уровнем экспрессии. В настоящем изобретении авторы предлагают подход для разработки стабильных клеточных линий,-1 007465 адаптированных к бессывороточным и безбелковым средам. В соответствии с этим подходом выделяют несколько клонов в безбелковой среде. Подробное описание изобретения Две стадии адаптации клеточных линий к безбелковой среде Эта процедура включает использование линий клеток млекопитающих, в отношении которых не представляется возможным осуществить процедуру непосредственной адаптации из среды, включающей сыворотку, или из бессывороточной среды к безбелковой среде. Способ по настоящему изобретению состоит из двустадийного процесса, в ходе которого осуществляется адаптация клеточных линий к безбелковой среде (PFM). Первая стадия, которая рассматривается как некритическая стадия, представляет собой снижение содержания белка без потери жизнеспособности клеток, в ходе которой не отмечается важного для популяции снижения времени удвоения клеток на каждой стадии при изменении концентрации белка. Некритическая стадия обычно имеет место при общей концентрации белка от 5 до 0,5 мг/мл в культуральной среде, и при этом культура демонстрирует практически те же ростовые характеристики, что и в исходной культуральной среде. Эта первая стадия начинается с уровня жизнеспособности клеточной линии, характеризующейся показателями от 80 до 100%, и клетки растут в культуральных средах при последовательном снижении концентрации белка вплоть до критической концентрации, при которой жизнеспособность клеток падает до 0%. Указанная концентрация белка представляет собой исходную точку для следующей стадии. Вторая стадия, которая рассматривается как критическая: на данной стадии происходит снижение жизнеспособности клеток и показателя времени удвоения клеток, а также может потребоваться больше времени для их адаптации при переходе с одной стадии на другую стадию, определяемой по концентрации белка. Существуют критические концентрации белка в культуральной среде, которую невозможно миновать в ходе адаптационного процесса. Указанные критические концентрации белка являются специфичными для каждой рекомбинантной клеточной линии, но обычно они ниже 0,6 мг/мл. Как только клетки восстанавливают свои исходные показатели жизнеспособности и скорости роста при данных критических концентрациях белка, создается возможность для их субкультивирования в следующих условиях со сниженной концентрацией белка. Когда критическая концентрация белка имеет фиксированное значение, ближайшая повышенная концентрация белка, которая поддерживает рост клеток, рассматривается как предкритическая концентрация белка. Начиная с предкритической концентрации белка, указанную концентрацию медленно снижают до восстановления клетками исходных показателей жизнеспособности и скорости роста. Выбранное сочетание стадий, применяемое для снижения концентрации белка на критической стадии, будет определять общее время адаптации на данной стадии, и скорость адаптации (Vадапт.) вычисляется по следующему уравнению:Vадапт. =Концентрация белка/T. Однако это сочетание стадий не будет влиять на время, необходимое для адаптации к каждой из взятых концентраций белка, включая критические концентрации. Для определения конечной точки некритической стадии и критических концентраций белка необходимо осуществлять постадийную адаптацию, при проведении серийного разбавления клеточной культуры желательной безбелковой средой, при котором каждый раз концентрацию белка снижают в 2 раза (см. таблицу). Указанное снижение может быть осуществлено путем снижения содержания сыворотки или добавки основной среды с разными уровнями некоторых обогащенных белками заменителей сыворотки. Постадийное снижение концентрации белка для определения критических концентраций,начиная с использования культуральной среды, содержащей в качестве добавок 5 мг/мл белкаSCM - среда, содержащая сыворотку Общее содержание белка в фетальной сыворотке теленка принимается равным примерно 50 мг/мл-2 007465 Перед началом процедуры адаптации клетки поддерживают в Т-бутылях при показателях жизнеспособности не ниже 80% с использованием стандартной среды, обычно используемой для культивирования клеток. Процесс адаптации осуществляют постадийно, выполняя описанные ниже стадии.i. Высеивают в 3 ячейки в шестиячеечном планшете для культивирования клеток рекомбинантную клеточную линию с использованием стандартной культуральной среды для культивирования клеток (с исходной концентрацией белка). Плотность клеток должна составлять 1-5 х 105 клеток/мл. Через 48 ч половину супернатанта заменяют свежей безбелковой средой, что приводит к достижению конечной концентрации белка, составляющей 50% от ее значения в исходных условиях.ii. Каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 50% от исходных условий.iii. Клетки растят до слияния при данной белковой концентрации.iv. Клетки со стадии iii высеивают по меньшей мере в 3 ячейки с плотностью в диапазоне 1-5 х 105 клеток/мл в культуральной среде, в которой содержание белка составляет 50% от его концентрации в исходных условиях. Через 48 ч половину супернатанта заменяют свежей безбелковой средой, что приводит к достижению конечной концентрации белка, составляющей 50% от ее значения в предыдущих условиях.v. Каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 50% от исходных условий.vi. Клетки растят до слияния при данной белковой концентрации.vii. Стадии (iv)-(vi) повторяют, при этом в ходе каждого цикла концентрацию белка снижают до 50% относительно концентрации в предыдущем цикле. Процедуру повторяют до достижения концентрации белка, которая вызывает гибель клеток.viii. Клетки из клеточной культуры с показателями жизнеспособности 80% или выше высеивают с использованием предкритической концентрации белка по меньшей мере в 3 ячейки с плотностью в диапазоне 2-6 х 105 клеток/мл. Клетки растят при предкритической концентрации белка и через 48 ч 25% супернатанта заменяют свежей безбелковой средой, с достижением при этом конечной концентрации белка, которая составляет 75% от предкритической концентрации белка.ix. Каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 75% от предкритической концентрации белка. х. Клетки растят до слияния при данной белковой концентрации.xi. Клетки со стадии (х) высеивают по меньшей мере в 3 ячейки с плотностью 2-6 х 105 клеток/мл в культуральной среде, в которой содержание белка составляет 75% от предкритической концентрации белка. Через 48 ч 25% супернатанта заменяют свежей безбелковой средой, с достижением при этом конечной концентрации белка, которая составляет 75% от концентрации на предыдущей стадии.xii. Каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 75% от концентрации на стадии (х).xiii. Клетки растят до слияния при данной белковой концентрации.xiv. Стадии (xi)-(xiii) повторяют, при этом в ходе каждого цикла концентрацию белка снижают до 75% относительно концентрации в предыдущем цикле и далее процедуру повторяют до достижения концентрации белка, которая не вызывает потери жизнеспособности клеток и снижения времени удвоения клеток в популяции. Когда клетки переносят в среду со сниженной концентрацией белка и они способны расти без потери жизнеспособности клеток и снижения времени удвоения клеток в популяции перед первым субкультивированием, авторы считают, что такие клетки достигли снова некритической стадии, и высеивают их непосредственно в безбелковой среде (концентрация белка 0 мг/мл). В процедуре осуществления настоящего изобретения исходная культуральная среда содержит фетальную сыворотку теленка в количестве от 5 до 10%. Линия клеток млекопитающих, адаптированная для роста в безбелковой среде, представляет собой миеломную линию, в частности NSO клетки. Настоящее изобретение также может быть полезно в том случае, когда используют линию клетокNSO, трансфицированных полипептидом или рекомбинантным белком, в частности, когда осуществляется трансфекция последовательностью, кодирующей рекомбинантное антитело или его фрагменты. Описываются также линии клеток млекопитающих, модифицированные в соответствии с процедурой по настоящему изобретению, которые растут в безбелковой среде. Другой вариант настоящего изобретения относится к любой линии клеток млекопитающих, экспрессирующих гуманизированные или химерные антитела, выбранные из группы, состоящей из антител против рецептора EGF hR3, анти-СD6 T1hT, анти-СD3 T3Q или их фрагментов, растущих в безбелковой среде, так что эти антитела могут секретироваться этими клеточными линиями. Клеточные линии, получаемые способом согласно изобретению, поддерживают стабильный рост в безбелковой среде в течение по меньшей мере 40 генераций. Примеры Пример 1. Адаптация рекомбинантной клеточной линии hR3 к безбелковой среде. Рекомбинантную клеточную линию hR3 получают путем трансфекции миеломной клеточной линииNSO векторными конструкциями для достижения экспрессии легкой и тяжелой цепей гуманизированного моноклонального антитела против рецептора человеческого EGF hR3. Адаптацию этой клеточной линии к безбелковой среде осуществляют по описанной ранее процедуре, в процессе двустадийного снижения содержания белка в среде. Указанные клетки вначале культивируют в среде RPMI-1640 с добавкой 10% FBS. Замещение FBS осуществляют за счет внесения обогащенной белком добавки Nutridoma NS (Boehringer Manheim) к безбелковой среде RPMI-1640, когда концентрация белка достигнет 0,15 мг/мл. Снижение содержания белка в исходной среде осуществляют путем последовательных разбавлений с использованием безбелковой среды PFHM-II (Gibco). На фиг. 1 и 2, соответственно, показаны результаты вычисления критических концентраций и скоростей адаптации Vадапт Пример 2. Адаптация рекомбинантной клеточной линии T1hT к безбелковой среде. Рекомбинантную клеточную линию T1hT получают путем трансфекции миеломной клеточной линии NSO векторными конструкциями для достижения экспрессии легкой и тяжелой цепей гуманизированного по методу Т-супрессии эпитопа моноклональными антителами против человеческого CD6. Адаптацию этой клеточной линии к безбелковой среде осуществляют по процедуре, описанной в п.2, в ходе двустадийного процесса снижения содержания белка в среде. Указанные клетки культивируют в среде RPMI-1640 с добавкой 10% FBS. Снижение содержания белка в исходной среде осуществляют путем последовательных разбавлений с использованием безбелковой среды PFHM-II (Gibco). Результаты вычисления критических концентраций и скоростей адаптации Vадапт. показаны на фиг. 3 и 4, соответственно. Пример 3. Адаптация рекомбинантной клеточной линии T3Q к безбелковой среде. Рекомбинантную клеточную линию T3Q получают путем трансфекции миеломной клеточной линии NSO векторными конструкциями для достижения экспрессии легкой и тяжелой цепей гуманизированного моноклонального антитела T3Q, который распознает рецептор CD3 на человеческих лимфоцитах. Адаптацию этой клеточной линии к безбелковой среде проводят по процедуре, описанной в п.2, в ходе двустадийного процесса снижения содержания белка в среде. Указанные клетки культивируют в среде RPMI-1640 с добавкой 10% FBS. Снижение содержания белка в исходной среде осуществляют путем последовательных разбавлений с использованием безбелковой среды PFHM-II (Gibco). Результаты вычисления критических концентраций и скоростей адаптации Vадапт. показаны на фиг. 5 и 6, соответственно. Краткое описание чертежей Фиг. 1. Корреляция времени, необходимого для адаптации h-R3 клеток к каждой из исследуемых концентраций (до восстановления жизнеспособности и показателя времени удвоения клеток), и натурального логарифма величины, обратной концентрации белка. Значения критических концентраций для линии клеток h-R3 составляют 0,32 и 0,11 мг/мл применительно к общей концентрации белка. Фиг. 2. Корреляция суммарного периода времени от момента начала процесса адаптации по используемой процедуре и натурального логарифма величины, обратной концентрации белка, для линии клетокh-R3. Показатели скоростей адаптации для линии клеток h-R3 составляют 0,0053 мг/день. Фиг. 3. Корреляция времени, необходимого для адаптации T1hT клеток к каждой из исследуемых концентраций (до восстановления жизнеспособности и показателя времени удвоения клеток), и натурального логарифма величины, обратной концентрации белка. Значения критических концентраций для линии клеток T1hT составляют 0,12 и 0,01 мг/мл применительно к общей концентрации белка. Фиг. 4. Корреляция суммарного периода времени от момента начала процесса адаптации по используемой процедуре и натурального логарифма величины, обратной концентрации белка, для линии клетокT1ht. Показатели скоростей адаптации для линии клеток T1ht составляют 0,0014 мг/день. Фиг. 5. Корреляция времени, необходимого для адаптации T3Q клеток к каждой из исследуемых концентраций (до восстановления жизнеспособности и показателя времени удвоения клеток), и натурального логарифма величины, обратной концентрации белка. Значения критических концентраций для линии клеток T3Q составляют 0,63 мг/мл применительно к общей концентрации белка. Фиг. 6. Корреляция суммарного периода времени от момента начала процесса адаптации по используемой процедуре и натурального логарифма величины, обратной концентрации белка, для линии клетокT3Q. Показатели скоростей адаптации для линии клеток T3Q составляют 0,0172 мг/день. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ получения линии клеток млекопитающих, адаптированных к росту в бессывороточных и безбелковых средах, который включает две стадии:I) первую стадию, в ходе которой показатели жизнеспособности клеток в клеточной линии состав-4 007465 ляют от 80 до 100% и клетки выращивают в культуральных средах, в которых последовательно снижают содержание белка до критической концентрации белка, при которой жизнеспособность клеток падает до 0%;II) вторую стадию, в ходе которой, на основе уже определенной критической концентрации белка,поддерживают предкритическую концентрацию как такую концентрацию белка, при которой клеточный рост возможен, фиксируя ее как исходную точку для дальнейшего медленного снижения концентрации белка до момента, когда клеточная культура достигнет исходных показателей жизнеспособности клеток и времени удвоения клеток в популяции. 2. Способ по п.1, отличающийся тем, что первая стадия состоит из следующих стадий:i) высеивают в 3 ячейки в шестиячеечном планшете для культивирования клеток рекомбинантную клеточную линию с использованием стандартной культуральной среды для культивирования клеток (с исходной концентрацией белка); плотность клеток должна составлять 1-5 х 105 клеток/мл; через 48 ч половину супернатанта заменяют свежей безбелковой средой, что приводит к достижению конечной концентрации белка, составляющей 50% от ее значения в исходных условиях;ii) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 50% от исходных условий;iii) клетки растят до слияния при данной белковой концентрации;iv) клетки со стадии iii высеивают по меньшей мере в 3 ячейки с плотностью в диапазоне 1-5 х 105 клеток/мл в культуральной среде, в которой содержание белка составляет 50% от его концентрации в исходных условиях; через 48 ч половину супернатанта заменяют свежей безбелковой средой, что приводит к достижению конечной концентрации белка, составляющей 50% от ее предыдущего значения;v) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 50% от ее предыдущего значения;vi) клетки растят до слияния при данной белковой концентрации;vii) стадии (iv)-(vi) повторяют, при этом в ходе каждого цикла концентрацию белка снижают до 50% относительно концентрации в предыдущем цикле; процедуру повторяют до достижения концентрации белка, которая вызывает гибель клеток. 3. Способ по п.1, отличающийся тем, что вторая стадия состоит из следующих стадий:viii) клетки из клеточной культуры с показателями жизнеспособности 80% или выше высеивают с использованием предкритической концентрации белка по меньшей мере в 3 ячейки с плотностью в диапазоне 2-6 х 105 клеток/мл; клетки растят при предкритической концентрации белка и через 48 ч 25% супернатанта заменяют свежей безбелковой средой, с достижением при этом конечной концентрации белка, которая составляет 75% от предкритической концентрации белка;ix) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 75% от предкритической концентрации белка; х) клетки растят до слияния при данной белковой концентрации;xi) клетки со стадии (х) высеивают по меньшей мере в 3 ячейки с плотностью 2-6 х 105 клеток/мл в культуральной среде, в которой содержание белка составляет 75% от предкритической концентрации белка; через 48 ч 25% супернатанта заменяют свежей безбелковой средой, с достижением при этом конечной концентрации белка, которая составляет 75% от концентрации на предыдущей стадии;xii) каждые 48 ч супернатант полностью заменяют свежей культуральной средой, в которой концентрация белка составляет 75% от концентрации на стадии (х);xiii) клетки растят до слияния при данной белковой концентрации;xiv) стадии (xi)-(xiii) повторяют, при этом в ходе каждого цикла концентрацию белка снижают до 75% относительно концентрации в предыдущем цикле и далее процедуру повторяют до достижения концентрации белка, которая не вызывает потери жизнеспособности клеток и снижения времени удвоения клеток в популяции; когда клетки переносят в среду со сниженной концентрацией белка и они способны расти без потери жизнеспособности клеток и без снижения времени удвоения клеток в популяции перед первым субкультивированием, авторы считают, что такие клетки достигли снова некритической стадии,и высеивают их непосредственно в безбелковой среде (концентрация белка 0 мг/мл). 4. Способ по пп.1-3, где бессывороточная и безбелковая среда, в которую клетки первоначально высеивают, включает от 5 до 10% фетальной сыворотки теленка. 5. Способ по пп.1-4, где линия клеток млекопитающих, адаптированных для роста в бессывороточных и безбелковых средах, представляет собой миеломную линию. 6. Способ по п.5, где миеломная линия представляет собой клеточную линию NSO. 7. Способ по п.6, где указанная клеточная линия NSO содержит последовательность, кодирующую рекомбинантный полипептид или рекомбинантный белок. 8. Способ по п.7, где последовательность, кодирующая рекомбинантный полипептид или рекомбинантный белок, кодирует рекомбинантное антитело или его фрагмент. 9. Линия клеток млекопитающих, полученная способом по пп.1-8, где указанная клеточная линия адаптирована для роста в бессывороточных и безбелковых средах. 10. Линия клеток млекопитающих по п.9, где указанная линия клеток млекопитающих представляет-5 007465 собой миеломную линию. 11. Линия клеток млекопитающих по п.10, где указанная клеточная линия представляет собой клеточную линию NSO. 12. Линия клеток млекопитающих по п.11, где клеточная линия NSO содержит последовательность,кодирующую рекомбинантный полипептид или рекомбинантный белок. 13. Линия клеток млекопитающих по п.12, где последовательность, кодирующая рекомбинантный полипептид или рекомбинантный белок, кодирует рекомбинантное антитело или его фрагмент. 14. Линия клеток млекопитающих по п.13, где последовательность кодирует гуманизированное рекомбинантное анти-EGF-R hR3 антитело или его фрагмент. 15. Линия клеток млекопитающих по п.13, где последовательность кодирует гуманизированное рекомбинантное анти-СD6 T1hT антитело или его фрагмент. 16. Линия клеток млекопитающих по п.13, где последовательность кодирует химерное рекомбинантное анти-СD3 T3Q антитело или его фрагмент. 17. Применение способа по пп.1-8 для получения линии клеток млекопитающих, адаптированных для роста в бессывороточной и безбелковой среде. 18. Гуманизированное моноклональное антитело к EGF-R hR3 или его фрагменты, секретируемое клетками, полученными способом по пп.1-8. 19. Гуманизированное моноклональное антитело против антигена CD6 T1hT или его фрагменты,секретируемое клетками, полученными способом по пп.1-8. 20. Химерное моноклональное антитело против антигена CD3 T3Q или его фрагменты, секретируемое клетками, полученными способом по пп.1-8. Корреляция времени, необходимого для адаптации hR3 клеток к каждой белковой концентрации, и концентрации Фиг. 1 Корреляция общего периода времени от начала процедуры адаптации клеточной линии h-R3 и концентрации Фиг. 2 Корреляция времени, необходимого для адаптации Т 1hТ клеток к каждой белковой концентрации, и концентрации-6 007465 Корреляция общего периода времени от начала процедуры адаптации клеточной линии Т 1hT и концентрации Фиг. 4 Корреляция времени, необходимого для адаптации T3Q клеток к каждой белковой концентрации, и концентрации Фиг. 5 Корреляция общего периода времени от начала процедуры адаптации клеточной линии T3Q и концентрации
МПК / Метки
МПК: C12N 5/00, C12P 21/08
Метки: процедура, получения, безбелковых, клеточные, способом, полученные, средах, линии, этим, клеточных, линий
Код ссылки
<a href="https://eas.patents.su/8-7465-procedura-polucheniya-kletochnyh-linijj-v-bezbelkovyh-sredah-i-kletochnye-linii-poluchennye-etim-sposobom.html" rel="bookmark" title="База патентов Евразийского Союза">Процедура получения клеточных линий в безбелковых средах и клеточные линии, полученные этим способом</a>
Способ обработки табака для снижения содержания нитрозоаминов и продукты, полученные этим способом

Номер патента: 2448
Опубликовано: 25.04.2002
Автор: Вилльямс Джонни Р.
МПК: A24F 47/00, A24B 3/10
Метки: обработки, снижения, табака, способом, нитрозоаминов, этим, способ, полученные, продукты, содержания
Формула / Реферат:
1. Способ снижения количества или предотвращения образования нитрозоаминов в собранном табачном растении, предусматривающий (i) стадию (а) отделения стеблей от табачных листьев, или (b) прессования табачных листьев для удаления избыточной влаги, или (с) обработки табачных листьев паром, и (ii) стадию подвергания, по меньшей мере, части растения микроволновому облучению, при том, что упомянутая часть является невысушенной и находится в...
Способ производства гранул вспученной глины и гранулы, полученные этим способом

Номер патента: 3730
Опубликовано: 28.08.2003
Авторы: Ванденбюсш Фредерик, Бессирон Натали
МПК: B28B 1/50, C04B 20/06
Метки: производства, способом, способ, вспученной, глины, гранулы, гранул, этим, полученные
Формула / Реферат:
1. Способ производства гранул вспученной глины, отличающийся тем, что вспучивание глиняных гранул осуществляют путем нагрева последних посредством микроволнового излучения. 2. Способ производства гранул вспученной глины по п.1, отличающийся тем, что глиняные гранулы подвергают сушке до обработки микроволновым излучением, обеспечивающим их вспучивание. 3. Способ производства гранул вспученной глины по п.1, отличающийся тем, что глиняные гранулы...
Способ отделения гемицеллюлозы от биомассы, содержащей гемицеллюлозу, а также биомасса и гемицеллюлоза, полученные этим способом

Номер патента: 5492
Опубликовано: 24.02.2005
Авторы: Штайн Армин, Кеттенбах Герхард
МПК: C08B 37/14, D21C 3/22
Метки: способ, содержащей, гемицеллюлоза, способом, этим, гемицеллюлозы, полученные, отделения, биомассы, биомасса, также, гемицеллюлозу
Формула / Реферат:
1. Способ отделения гемицеллюлозы от биомассы, содержащей гемицеллюлозу, отличающийся посредством следующих стадий: a) экстрагирование гемицеллюлозы из биомассы, содержащей гемицеллюлозу, путем обработки комплексным соединением в водном растворе с образованием растворимого комплекса гемицеллюлозы, причем в качестве комплексного соединения в стадии a) используют координационное соединение переходного металла и моно- или полидентатного...
Способ введения этиленовоненасыщенных функциональных групп в полимер алкилстирола и полимерные производные, полученные этим способом

Номер патента: 1883
Опубликовано: 22.10.2001
Авторы: Фречет Джин М.Дж., Ван Сяньчан, Хэк Шах А.
МПК: C08F 8/42
Метки: этиленовоненасыщенных, алкилстирола, этим, полученные, функциональных, групп, введения, производные, полимер, способ, полимерные, способом
Формула / Реферат:
1. Способ введения этиленовоненасыщенной функциональной группы в полимер алкилстирола по месту первичного бензильного углеродного атома, включающий следующие стадии: приготовление раствора этого полимера в углеводородном растворителе, добавление в такой полимерный раствор алкоксида щелочного металла и литийалкильного соединения с получением раствора металлированного промежуточного продукта, добавление в раствор промежуточного продукта...
Cпособ получения фактора ix из биологических источников и фактор ix, полученный этим способом

Номер патента: 222
Опубликовано: 24.12.1998
Авторы: Хоффер Лутц, Йосич Дьюро, Морфельд Франк
Метки: cпособ, фактора, полученный, биологических, фактор, этим, источников, получения, способом
Формула / Реферат:
1. Способ получения фактора IX из биологических источников с помощью хроматографии, отличающийся тем, что перед разделением с помощью аффинной хроматографии указанный источник обрабатывается сульфатом аммония в концентрации от 1,5-2,3 моль/л. 2. Способ по п.1, отличающийся тем, что указанным биологическим источником является плазма крови или плазма, истощенная по фактору VIII и фибриногену путем отделения криопреципитата (криообедненная...
Предыдущий патент: Антагонисты метаботропных рецепторов глутамата
Следующий патент: Фармацевтическая композиция для лечения повреждения тканей, вызванного недостаточностью артериального кровотока
Случайный патент: Штанговый глубинный насос