Способ переноса клеточных ядер, способ получения генетически модифицированного или трансгенного млекопитающего, способ клонирования млекопитающего, генетически модифицированное млекопитающее и генетически модифицированный эмбрион
Номер патента: 17100
Опубликовано: 28.09.2012
Авторы: Ду Юйтао, Крагх Петер Михаэль, Вайта Габор, Болун Ларс Аксель





















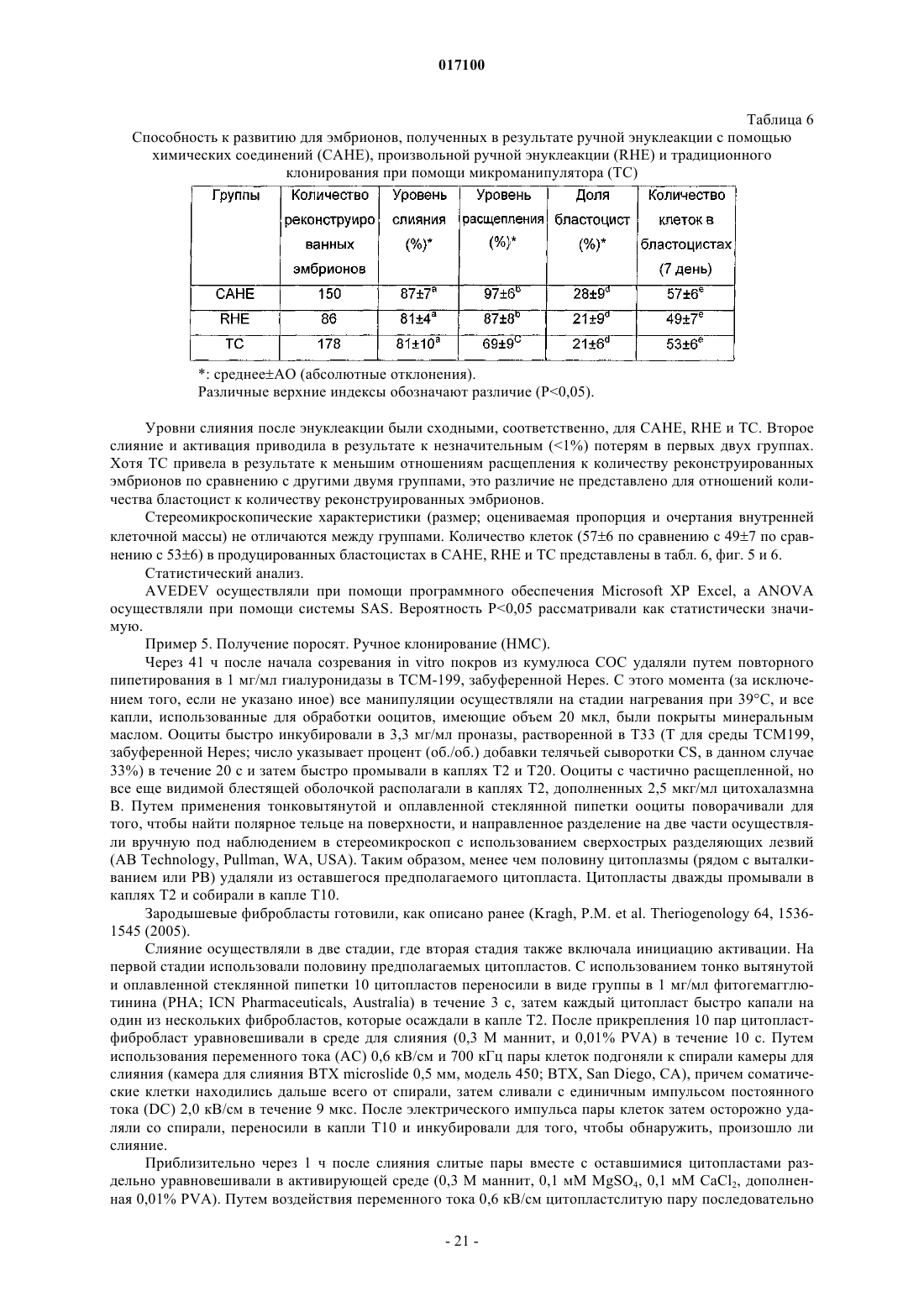

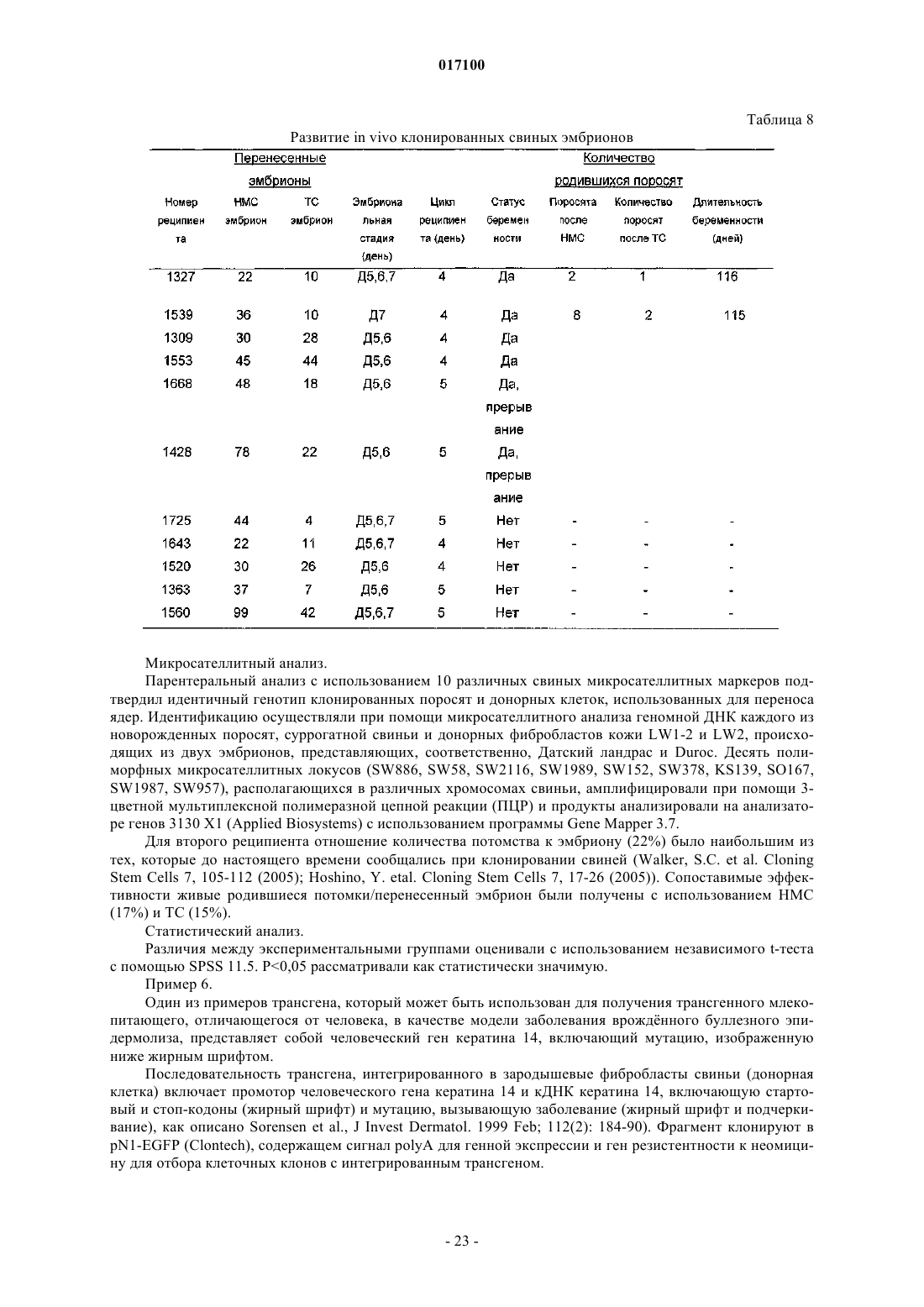





Формула / Реферат
1. Способ переноса клеточных ядер, включающий стадии:
а) получения по меньшей мере одного ооцита, где по меньшей мере часть блестящей оболочки удаляют частично,
б) разделения ооцита по меньшей мере на две части с получением ооцита, содержащего ядро, и по меньшей мере одного цитопласта,
в) получения донорной клетки или клеточного ядра, обладающего желаемыми генетическими свойствами,
г) слияния по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной,
д) получения реконструированного эмбриона.
2. Способ по п.1, где ооцит разделяют по меньшей мере на три части с получением по меньшей мере двух цитопластов.
3. Способ по любому из пп.1 и 2, где блестящую оболочку частично удаляют с помощью ферментов.
4. Способ по любому из пп.1-3, где ооцит разделяют по меньшей мере на три части с получением по меньшей мере двух цитопластов.
5. Способ по любому из пп.1-4, где желаемые генетические свойства донорной клетки или клеточного ядра получают путем модификации желаемого гена или генов с помощью мутации, делеции и/или инсерции.
6. Способ по любому из пп.1-5, где способ слияния выбирают из химического слияния, слияния с использованием электрического тока и биослияния.
7. Способ по любому из пп.1-6, где слияние осуществляют по меньшей мере в одну стадию.
8. Способ по любому из пп.1-7, где слияние осуществляют по меньшей мере в две стадии.
9. Способ по п.8, где первую стадию слияния осуществляют по меньшей мере между одним цитопластом и донорной клеткой или клеточным ядром, окруженным мембраной.
10. Способ по п.9, где вторую стадию слияния осуществляют по меньшей мере между одной слитой парой и по меньшей мере одним цитопластом.
11. Способ по любому из пп.1-10, где донорная клетка представляет собой соматическую клетку.
12. Способ по п.11, где соматическую клетку выбирают из группы, состоящей из эпителиальных клеток, нервных клеток, эпидермальных клеток, кератиноцитов, гематопоэтических клеток, меланоцитов, хондроцитов, лимфоцитов (B и T лимфоцитов), эритроцитов, макрофагов, моноцитов, мононуклеарных клеток, фибробластов, клеток сердечной мышцы и других мышечных клеток.
13. Способ по п.12, где соматические клетки получают из группы, состоящей из клеток кожи, клеток легких, клеток поджелудочной железы, клеток печени, клеток желудка, интестинальных клеток, сердечных клеток, клеток репродуктивных органов, клеток мочевого пузыря, клеток почки, клеток мочеиспускательного канала и клеток других органов мочевой системы.
14. Способ по любому из пп.1-13, где соматическая клетка представляет собой клетку фибробласта.
15. Способ по п.14, где соматическая клетка представляет собой клетку фибробласта, полученную от млекопитающего.
16. Способ по п.15, где млекопитающее представляет собой свинью.
17. Способ по пп.1 и/или 2, где донорная клетка имеет происхождение из клетки зародышевой линии.
18. Способ по любому из пп.1-17, где ооцит получают от свиньи.
19. Способ по п.1, дополнительно включающий стадии:
а) активации митоза в реконструированном эмбрионе с образованием эмбриона,
б) выращивания указанного эмбриона.
20. Способ по п.19, где эмбрион последовательно выращивают в различных средах.
21. Способ получения генетически модифицированного или трансгенного млекопитающего, отличного от человека, включающий осуществление способа по любому из пп.1-19 и последующие стадии:
а) активации митоза в реконструированном эмбрионе с образованием эмбриона;
б) выращивания указанного эмбриона и
в) переноса указанного выращенного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш.
22. Способ по п.21, где способ активации реконструированного эмбриона выбирают из группы, состоящей из электрического импульса, химически индуцированного шока, увеличения внутриклеточных концентраций двухвалентных катионов и уменьшения фосфорилирования.
23. Способ по п.21, дополнительно включающий вторую стадию слияния между по меньшей мере одной слитой парой цитопласта и донорной клетки и по меньшей мере одним цитопластом, где указанную вторую стадию слияния осуществляют последовательно или одновременно.
24. Способ по п.21, где эмбрион выращивают in vitro.
25. Способ по п.24, где эмбрион последовательно выращивают в различных средах.
26. Способ по п.21, где эмбрион подвергают криоконсервации перед переносом в млекопитающее-хозяин.
27. Способ по п.26, где эмбрион находится на стадии бластоцисты.
28. Способ по п.26, где по меньшей мере один ооцит, полученный на стадии (а) п.1, подвергают делипидизации путем мягкой обработки ферментативным агентом перед криоконсервацией эмбриона.
29. Способ по п.28, где делипидизированный ооцит разделяют по меньшей мере на две части с получением ооцита, содержащего ядро, и по меньшей мере одного цитопласта.
30. Способ по п.26, где указанный эмбрион выращивают до стадии бластоцисты перед криоконсервацией.
31. Способ клонирования млекопитающего, отличного от человека, включающий:
а) получение эмбриона в соответствии с любым из пп.1-18,
б) перенос указанного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш.
32. Генетически модифицированное млекопитающее, отличное от человека, получаемое при помощи способа по любому из пп.21-31, содержащее в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из двух различных материнских источников.
33. Генетически модифицированный эмбрион, отличный от человеческого, получаемый при помощи способа по любому из пп.21-31, содержащий в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из двух различных материнских источников.
34. Генетически модифицированное млекопитающее, отличное от человека, по п.32, содержащее в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из трех различных материнских источников.
35. Генетически модифицированный эмбрион, отличный от человеческого, по п.33, содержащий в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из трех различных материнских источников.
36. Генетически модифицированное млекопитающее, отличное от человека, по п.32, содержащее в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из четырех различных материнских источников.
37. Генетически модифицированный эмбрион, отличный от человеческого, по п.33, содержащий в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из четырех различных материнских источников.
38. Генетически модифицированное млекопитающее, отличное от человека, по п.32, где млекопитающее представляет собой свинью.
39. Генетически модифицированный эмбрион, отличный от человеческого, по п.33, где эмбрион имеет происхождение от свиньи.
Текст
СПОСОБ ПЕРЕНОСА КЛЕТОЧНЫХ ЯДЕР, СПОСОБ ПОЛУЧЕНИЯ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННОГО ИЛИ ТРАНСГЕННОГО МЛЕКОПИТАЮЩЕГО, СПОСОБ КЛОНИРОВАНИЯ МЛЕКОПИТАЮЩЕГО, ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННОЕ МЛЕКОПИТАЮЩЕЕ И ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЙ ЭМБРИОН Ду Юйтао, Вайта Габор, Болун Ларс Аксель, Крагх Петер Михаэль (DK) Ильмер Е.Г., Пантелеев А.С.,Кузнецова Ю.В. (RU) В настоящем изобретении раскрыты способы переноса клеточного ядра, включающие, например,модификацию блестящей оболочки ооцита и/или разделение ооцита на несколько частей. В настоящем изобретении также раскрыты способы получения генетически модифицированного млекопитающего, отличного от человека. Генетически модифицированные млекопитающие и генетически модифицированные эмбрионы, отличные от человека, получаемые при помощи раскрытых способов, также находятся в объеме настоящего изобретения. Также раскрыт способ клонирования млекопитающего, отличного от человека. 017100 Область изобретения Настоящее изобретение относится к способу переноса клеточных ядер у млекопитающих и к полученным генетически модифицированным млекопитающим или генетически модифицированным животным, которые могут быть получены при помощи этого способа. Предпосылки изобретения Способность генетически модифицировать донорные клетки и использовать их для переноса ядер обеспечивает средство для получения генетически модифицированных животных, которые могут быть использованы, например, в качестве моделей заболеваний для исследования серьезных заболеваний людей и тестирования лекарств. Традиционные способы переноса клеточных ядер включают две стадии микроманипуляции. Первая стадия включает энуклеацию зрелого ооцита, а вторая стадия охватывает перенос донорного ядра. Тем не менее, оказалось, что микроманипуляция обладает несколькими недостатками, например потребность в дорогостоящем оборудовании, потребность в высококвалифицированном персонале и требует много времени. Недавно был разработан улучшенный способ переноса ядер, в котором в качестве донорных клеток используют соматические клетки, известный как ручное клонирование (НМС), который включает применение блестящей оболочки (zona pellucida) свободных ооцитов. Способ является упрощенным по сравнению с традиционным переносом ядер, поскольку микроманипуляция больше не является необходимой. Способ использован в отношении коров (Vajta et al., 2001, Cloning 3, 89-95; Vajta et al., 2003, Bioll.(Booth et al., 2001, Cloning Stem Cells 3, 191-197) описано применение переноса ядер без блестящей оболочки с одной стадией микроманипуляции. Фактически то, что этот способ обладает меньшими техническими требованиями и требует меньших затрат времени, подталкивает исследователей предложить применение способа НМС в отношении других видов. Тем не менее, множество технических проблем привело к тому, что применение НМС в отношении свиней потребовало больших затрат по сравнению с теми, которые исходно предполагались. Одна из проблем, с которой столкнулись, связана с низкими плавучими плотностями ооцитов свиней, как ооцитов свиней с интактной блестящей оболочкой (ZI), так и в особенности лишенных блестящей оболочки (ZF). Следовательно, ооциты свиней не осаждаются на дно пробирки. Кроме того, поверхность ооцитов является липкой и сложно избежать их прикрепления друг к другу при удалении оболочки. Более того, свиные ооциты ZF являются очень хрупкими и сложно разделять их при помощи такого способа, который описан для ооцитов коров. Недавно способ НМС, однако, с низкой эффективностью был применен к переносу ядер в отношении свиней с использованием генетически модифицированных соматических клеток, фибробластов, в качестве донорных клеток, что привело в результате к получению генетически модифицированных клонированных бластоцистов (Kragh et al., 2004, Reproduction, Fertility and Development 16, 315-318). Настоящее изобретение улучшает способ соматического переноса клеточных ядер при помощи НМС, что приводит в результате к увеличенной скорости эмбриональной реконструкции и, следовательно, к значительному увеличению вероятности получить генетически модифицированных животных. Препятствие для получения генетически модифицированных животных при помощи способов переноса ядер в большом масштабе представляет собой невозможность криоконсервации ооцитов и эмбрионов свиней с использованием способов, применяемых в отношении других видов. Это является следствием высокого содержания липидов в ооцитах и эмбрионах свиней. Криоконсервация клонированных эмбрионов свиней может значительно улучшать выход клонирования соматических клеток путем того, что избегают логистических проблем. Тем не менее, недавно была опубликован неинвазивный способ делипидизации эмбрионов свиней путем центрифугирования, но без последующей микроманипуляции (Esaki et al., 2004, Biol Reprod. 71, 432-6). Краткое описание изобретения Настоящее изобретение в одном из аспектов относится к способу переноса клеточных ядер, включающему стадии а) получения по меньшей мере одного ооцита, имеющего по меньшей мере часть модифицированной блестящей оболочки, б) разделения ооцит по меньшей мере на две части с получением по меньшей мере одного цитопласта, в) получения донорной клетки или клеточного ядра, обладающего желаемыми генетическими свойствами, г) слияния по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной, д) получения реконструированного эмбриона. В одном из воплощений данного способа осуществляют разделение ооцита по меньшей мере на три части с получением по меньшей мере двух цитопластов. Другой аспект изобретения относится к способу получения генетически модифицированного или трансгенного млекопитающего, отличного от человека, включающему стадии а) получения по меньшей мере одного ооцита, имеющего по меньшей мере часть модифицированной блестящей оболочки, б) разделения ооцита по меньшей мере на две части с получением ооцита, имеющего ядро и по меньшей мере один цитопласт, в) получения донорной клетки или клеточного ядра с желаемыми генетическими свойствами, г) слияния по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окру-1 017100 женным мембраной, д) получения реконструированного эмбриона, е) активации реконструированного эмбриона с образованием эмбриона, ж) выращивания указанного эмбриона и з) переноса указанного выращенного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш. В еще одном аспекте изобретение относится к способу клонирования млекопитающего, отличного от человека, включающему стадии а) получения эмбриона при помощи способов по настоящему изобретению, б) переноса указанного выращенного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш. В другом аспекте изобретение относится к генетически модифицированному млекопитающему, отличному от человека, получаемому при помощи определенного здесь способа. В еще одном аспекте изобретение относится к генетически модифицированному эмбриону, отличному от человека, получаемому при помощи определенного здесь способа. В еще одном аспекте изобретение относится к генетически модифицированному эмбриону, отличному от человека, получаемому при помощи определенного здесь способа, имеющего в клетках его тканей митохондрии по меньшей мере из трех различных материнских источников. В заключительном аспекте изобретение относится к способу, дополнительно к указанным стадиям способа переноса клеточных ядер включающему стадии а) активации реконструированного эмбриона с образованием эмбриона и б) выращивания указанного эмбриона. Описание графических материалов Фиг. 1 - (а) деление ооцитов на три части; (б) комплексы фрагмента фибробласт-ооцит для первого слияния; (в) эмбрионы реорганизовывали триплетами (при переменном токе); (г) слияние триплетов. Измерительная линейка = 50 м; фиг. 2 - (а) зрелые in vitro ооциты после частичного расщепления блестящей оболочки, (б) делипидизированные ооциты после центрифугирования, (в) разделение на две части делипидизированных ооцитов, (г) комплексы фрагмента фибробласт-ооцит для первого слияния, (д) четырехклеточная стадия реконструированных эмбрионов, развившихся из делипидизированных ооцитов, (е) четырехклеточная стадия реконструированных эмбрионов, развившихся из интактных ооцитов, (ж) повторное вытягивание бластоцистов из делипидизированных эмбрионов после нагревания, (з) окрашивание по Хехст и УФ визуализация повторного вытягивания бластоцистов из делипидизированных эмбрионов после нагревания. Линейка представляет 100 мкм; фиг. 3 - разделение на две части при энуклеакции с помощью химических соединений. Отмечен конус экструзии или полярное тело, связанное с меньшей частью (предполагаемый кариопласт). Стереомикроскопическое изображение. Линейка представляет 50 мкм; фиг. 4 - окрашивание по Хехст и УФ визуализация в отсутствие и присутствии хроматина. УФ свет,инвертированное флуоресцентное микроскопическое изображение. Линейка представляет 50 мкм; (а) отсутствие хроматина в предполагаемых цитопластах, (б) присутствие хроматина в предполагаемых кариопластах; фиг. 5 - стереомикроскопическое изображение бластоцист на 7 день, полученных путем ручной энуклеации при помощи химических соединений (САНЕ). Линейка представляет 50 мкм; фиг. 6 - окрашивание по Хехст и УФ визуализация бластоциты, проявляющейся после ручной энуклеакции при помощи химических соединений (САНЕ). Линейка представляет 50 мкм. Подробное описание изобретения В настоящем изобретении предложены улучшенные способы клонирования млекопитающего путем переноса ядер, которые связаны с введением полного комплемента ядерной ДНК из одной клетки в энуклеированную клетку. Перенос ядер соматических клеток. При клонировании перенос ядра соматической (телесной) клетки или соматической клетки в яйцеклетку (ооцит), у которой собственное ядро удалено (денуклеированную или энуклеированную), назван перенос ядер соматических клеток. Новый индивид, который развивается из этого реконструированного эмбриона, является генетически идентичным донору соматической клетки. В настоящем изобретении способ переноса ядер соматических клеток представляет собой способ переноса клеточных ядер, включающий стадии а) получения по меньшей мере одного ооцита, имеющего по меньшей мере часть модифицированной блестящей оболочки, б) разделения ооцита по меньшей мере на две части с получением по меньшей мере одного цитопласта, в) получения донорной клетки или клеточного ядра, обладающего желаемыми генетическими свойствами, г) слияния по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной, д) получения реконструированного эмбриона. Тем не менее, настоящее изобретение также относится к способу переноса клеточных ядер, включающему стадии а) получения по меньшей мере одного ооцита, б) разделения ооцита по меньшей мере на три части с получением по меньшей мере двух цитопласт, в) получения донорной клетки или клеточного ядра,обладающего желаемыми генетическими свойствами, г) слияния по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной, д) получения реконструированного эмбриона.-2 017100 Параметры перечисленных стадий могут варьировать для обеспечения наиболее эффективного ядерного переноса для данных видов животных. Различные параметры подробно описаны ниже. Ооцит. Термин ооцит по настоящему изобретению обозначает незрелую репродуктивную клетку самки,которая не завершила процесс созревания с образованием яйцеклетки (гаметы). В настоящем изобретении энуклеированный ооцит представляет собой клетку-реципиента в процессе переноса ядер. Ооциты по настоящему изобретению выделяют из яйцеводов и/или яичников млекопитающего. Обычно ооциты выделяют из умерщвленных животных, хотя они могут быть выделены также из яйцеводов и/или яичников живых животных. В одном из воплощений ооциты выделяют при помощи способов выделения маточной трубы или способов трансвагинального выделения. В предпочтительном воплощении ооциты выделяют путем аспирации. Ооциты, как правило, перед энуклеацией созревают в различных средах, известных специалистам в данной области техники. Ооциты также могут быть выделены из яичников недавно умерщвленного животного или когда яичник замораживали и/или отмораживали. Предпочтительно ооциты выделяют свежими из яйцеводов. Ооциты или цитопласты также могут быть криоконсервированы перед применением. Хотя специалисту в данной области техники понятно, что предпочтительны свежевыделенные и зрелые ооциты, также понятно, что можно криоконсервировать ооциты после сбора или после созревания. Если используют криоконсервированные оциты, то они должны быть исходно оттаяны перед помещением ооцитов в среду для созревания. Способы оттаивания криоконсервированного материала, осуществляемые таким образом, что они активны после способа оттаивания, хорошо известны специалистам в данной области техники. Тем не менее, как правило, криоконсервация ооцитов и цитопласт требует больших затрат, и ее особенно сложно осуществлять у свиней ввиду вышеупомянутой общей хрупкости свиных ооцитов и цитопласт, и ввиду высокого содержания липидов, что делает их очень чувствительными к повреждению при охлаждении (т.е. повреждению, которое возникает между +15 и +5C во время процедуры охлаждения и нагревания). В еще одном воплощении зрелые (метафаза II) ооциты, которые созрели in vivo, могут быть собраны и использованы в раскрытых здесь способах переноса ядер. По существу, зрелые (метафаза II) ооциты выделяют хирургическим путем у млекопитающих, не подвергнутых или подвергнутых суперовуляции, через 35-48 ч после начала течки или после инъекции человеческого хорионического гонадотропина (hCG) или похожего гормона. Когда ооциты выращены in vitro, клетки кумулюса, окружающие ооциты in vivo, могут образовывать скопление и могут быть удалены с получением ооцитов, которые находятся на более подходящей стадии созревания для энуклеакции. Клетки кумулюса могут быть удалены путем пипетирования или перемешивания, например, в присутствии в диапазоне от 0,1 до 5% гиалуронидазы, таком как в диапазоне от 0,2 до 5% гиалуронидазы, например в диапазоне от 0,5 до 5% гиалуронидазы, таком как в диапазоне от 0,2 до 3% гиалуронидазы, например в диапазоне от 0,5 до 3% гиалуронидазы, таком как в диапазоне от 0,5 до 2% гиалуронидазы, например в диапазоне от 0,5 до 1% гиалуронидазы, таком как 0,5% гиалуронидазы. Первая стадия в предпочтительных способах включает выделение реципиентного ооцита у соответствующего животного. В этой связи ооцит может быть получен из любого животного источника и на любой стадии созревания. Сообщали о том, что стадия созревания ооцита при энуклеакции и ядерном переносе имеет значение для успеха способов ядерного переноса. Незрелые (профаза I) ооциты из яичников млекопитающих часто собирают путем аспирации. При использовании способов, таких как генетическая инженерия, перенос ядер и клонирование, такие собранные ооциты предпочтительно подвергают созреванию in vitro перед тем как ооциты могут быть использованы в качестве клеток-реципиентов для ядерного переноса. Предпочтительно для успешного клонирования эмбриона млекопитающего в качестве реципиентного ооцита используют ооцит на стадии метафаза II, поскольку полагают, что на этой стадии созревания ооцит является достаточно активированным для того, чтобы вводить ядро, как если бы оно представляло собой оплодотворяющий сперматозоид. Тем не менее, настоящее изобретение относится к любой стадии созревания ооцита, подходящей для осуществления переноса ядер соматических клеток, эмбрионов, бластоцист и/или животных, которые получены при помощи способа переноса ядер соматических клеток по настоящему изобретению. Созревание ооцитов in vitro обычно происходит в среде для созревания до тех пор, пока ооцит не достигнет стадии метафазы II или не произойдет вытеснение первого полярного тела. Время, которое потребуется незрелому ооциту для достижения созревания, названо периодом созревания. В предпочтительном воплощении настоящего изобретения ооцит представляет собой ооцит свиньи или молодой свиньи, предпочтительно молодой свиньи. Животные. Донор (соматическая клетка или ядро соматической клетки) и реципиент (цитопласт), вовлеченные в способ переноса клеточных ядер по настоящему изобретению, представляют собой млекопитающее,-3 017100 отличающееся от человека. Аналогично, животное, которому могут быть имплантированы реконструированные эмбрионы по настоящему изобретению, представляет собой млекопитающее, отличающееся от человека. Млекопитающее может представлять собой копытное животное, выбранное из группы, состоящей из домашних или диких представителей коров, овец, оленей, свиней, лошадей и верблюдов. В конкретном воплощении млекопитающее представляет собой корову или быка, бизона, буйвола, овцу,снежного барана, лошадь, пони, осла, мула, оленя, лося, карибу, козу, азиатского буйвола, верблюда, ламу, альпака или свинью. В конкретном воплощении настоящего изобретения млекопитающее представляет собой свинью. В одном из воплощений свинья представляет собой дикую свинью. В еще одном воплощении свинья представляет собой домашнюю свинью Sus scrofa или S. domesticus. В еще одном воплощении изобретение относится к минисвиньям, а также к инбредным свиньям. В конкретном воплощении свинья может быть выбрана из группы, состоящей из ландраса, йоркшира, хэмпшира, дюрка, китайского мейшана, беркшира и пьетрена. В еще одном воплощении настоящее изобретение относится к группе, состоящей из ландраса, йоркшира, хэмпшира и дюрка. Тем не менее, настоящее изобретение также относится к группе, состоящей из ландраса, дюрка и китайского мейшана. Аналогично, группа, состоящая из беркшира, пьетрена, ландраса и китайского мейшана, может представлять собой задачи по настоящему изобретению. Также группа, состоящая из ландраса и китайского мейшана, представляет собой задачи настоящего изобретения. В конкретном воплощении свинья представляет собой ландрас или йоркшир. В конкретном воплощении изобретение относится к свиньям породы хэмпшир, а также дюрок. В еще одном предпочтительном воплощении свинья представляет собой породу китайский мейшан. Тем не менее, также беркшир охвачен изобретением, и в конкретном воплощении пьетрен охвачен настоящим изобретением. В еще одном воплощении настоящее изобретение относится к минисвиньям, выбранным из группы,состоящей из геттингена, юкатана, Bama Xiang Zhu, Wuzhishan, Xi Shuang Banna. В других воплощениях изобретение относится к группе, состоящей из геттингена, юкатана. Альтернативно, изобретение относится к группе, состоящей из Bama Xiang Zhu, Wuzhishan, Xi Shuang Banna. В частности, изобретение относится к геттинген. Но также для изобретения подходит юкатан. Аналогично,изобретением охвачены Bama Xiang Zhu, также Wuzhishan и, в частности Xi Shuang Banna. Донорные млекопитающие по настоящему изобретению могут представлять собой самок или самцов. Млекопитающее может иметь любой возраст, такой как взрослое млекопитающее, или, например,зародыш. Эмбрион. В соответствии с настоящим изобретением реконструированный эмбрион (т.е. единичная клетка эмбриона) содержит генетический материал донорной клетки. Затем реконструированный эмбрион последовательно делится до мультиклеточного эмбриона после начала митоза. In vitro начало митоза типично осуществляют путем активации как здесь описано. В настоящем изобретении термин эмбрион также относится к реконструированным эмбрионам,которые представляют собой эмбрионы, полученные после процесса ядерного переноса после начала митоза путем активации. Реконструированные эмбрионы выращивают in vitro. Когда эмбрион содержит приблизительно 12-16 клеток, он назван морулой. Затем эмбрион дополнительно делится и образуется множество клеток и заполненная жидкостью полость цисты в своем центре, представляющая собой полость бластоцеля. На этой стадии эмбрион назван бластоцитом. Стадия развития оплодотворенного ооцита готова для имплантации; образована из морулы и состоит из внутренней клеточной массы, внутренней полости и внешнего слоя клеток, названного трофоэктодермальными клетками. Бластоцист по настоящему изобретению может быть имплантирован в матку млекопитающегохозяина и продолжает расти в зародыш и затем в животное. В предложенных здесь способах получения генетически модифицированного или трансгенного млекопитающего, отличающегося от человека, клонирования млекопитающего, отличающегося от человека, выращивания реконструированного эмбриона и/или криоконсервации свиного эмбриона эмбрион может быть выращен in vitro. Эмбрион, например,может быть выращен в последовательной культуре. Понятно, что эмбрион может представлять собой нормальный эмбрион или реконструированный эмбрион, как здесь определено. Цитопласт. Ооцит или часть ооцита, из которого удалено ядро. Донорная клетка. Под термином донорная клетка по настоящему изобретению подразумевают соматическую клетку и/или клетки, полученные из зародышевой линии. Под термином соматическая клетка по настоящему изобретению подразумевают любую клетку(организма) животного на любой стадии развития. Например, соматические клетки могут происходить из зародышевой или взрослой ткани. Особенно предпочтительные соматические клетки представляют собой клетки, имеющие зародышевое происхождение. Тем не менее, также могут быть использованы клетки из зародышевой линии. В соответствии с настоящим изобретением донорная клетка представляет со-4 017100 бой соматическую клетку. В еще одном воплощении по настоящему изобретению донорная клетка представляет собой клетку, происходящую из зародышевой линии клеток. В предпочтительном воплощении настоящего изобретения донорная клетка обладает желаемыми генетическими свойствами. Тем не менее, донорная клетка может обладать желаемыми генетическими свойствами, которые получают путем генетической манипуляции как здесь описано. Соматические клетки выбраны из группы, состоящей из эпителиальных клеток, нервных клеток,эпидермальных клеток, кератиноцитов, гематопоэтических клеток, меланоцитов, хондроцитов, лимфоцитов (B и T лимфоцитов), эритроцитов, макрофагов, моноцитов, мононуклеарных клеток, фибробластов, клеток сердечной мышцы и других мышечных клеток. Они могут быть получены из различных органов, например кожи, легкого, поджелудочной железы,печени, желудка, тонкой кишки, сердца, репродуктивных органов, мочевого пузыря, почки, мочеиспускательного канала и других органов мочевой системы. Животные, из которых могут быть получены соматические клетки, здесь описаны. Предпочтительное воплощение изобретения представляет собой применение соматических клеток, происходящих из тех же самых видов как реципиентный ооцит (цитопласт). Предпочтительно соматические клетки представляют собой клетки фибробласта, которые могут быть получены из развивающихся плодов и взрослых животных в больших количествах. Кроме того,фибробласты легко могут быть размножены путем деления in vitro. Наиболее предпочтительно соматические клетки представляют собой фибробласты плодного происхождения, выращенные in vitro. В предпочтительном воплощении соматические клетки являются генетически модифицированными. В еще одном предпочтительном воплощении по настоящему изобретению соматические клетки представляют собой клетки свиньи предпочтительно зародышевого происхождения или, например, выделенные у взрослых особей. Энуклеакция. Способ энуклеакции ооцита может быть выбран из группы способов, состоящей из аспирации, физического удаления, применения ДНК-специфических флюорохромов, воздействия ультрафиолетовым светом и/или энуклеакции путем воздействия химическими соединениями. В одном из воплощений настоящее изобретение относится к применению ДНК-специфических флюорохромов. Тем не менее, энуклеакция может быть осуществлена путем воздействия ультрафиолетовым светом. В конкретном воплощении энуклеакцию осуществляют путем воздействия химическими соединениями перед физическим удалением ядра. Энуклеакция путем воздействия химическими соединениями с использованием, например, противоопухолевых агентов, таких как демеколцин (N-деацетил-N-метилколхицин), и/или, например, этопозид или родственные агенты, может быть осущствлена перед ферментативной модификацией блестящей оболочки. Энуклеация путем химического воздействия включает выращивание зрелых СОС(cumulus-oocyte complexes, комплексы кумулюс-ооцит) в среде для созревания, как здесь описано, дополненной демеколцином, в течение определенного периода времени. Среда для созревания может быть дополнена демеколцином в диапазоне от 0,1 до 10 мкг/мл, таком как от 0,2 до 10 мкг/мл, например от 0,3 до 10 мкг/мл, таком как от 0,25 до 5 мкг/мл, например от 0,3 до 1 мкг/мл, таких как от 0,25 до 0,5 мкг/мл,например 0,4 мкг/мл. Аналогично, среда для созревания может быть дополнена этопозидом, например к среде для созревания может быть добавлен этопозид в диапазоне от 0,1 до 10 мкг/мл этопозида, таким как от 0,2 до 10 мкг/мл, например от 0,3 до 10 мкг/мл, таким как от 0,25 до 5 мкг/мл, например от 0,3 до 1 мкг/мл, таким как от 0,25 до 0,5 мкг/мл, например 0,4 мкг/мл. Время выращивания СОС в присутствии противоопухолевых агентов составляет от 10 мин до 5 ч, такое как от 30 мин до 5 ч, например от 10 мин до 2 ч, такое как от 30 мин до 2 ч, например от 10 мин до 1,5 ч, такое как от 20 мин до 3 ч, например от 10 мин до 3 ч, такое как от 30 мин до 1,5 ч, например 45 мин. В конкретном воплощении энуклеакцию путем воздействия химическими соединениями осуществляют с использованием 0,45 мкг/мл демеколцина и/или этопозида, добавленного к среде для созревания в течение 45 мин. В конкретном воплощении предпочтительно, чтобы энуклеакцию осуществляли путем физического удаления ядра. Физическое удаление может быть осуществлено путем разделения, например путем разделения ооцита на две половины (две части), одна из которых содержит ядро, а другая энуклеированный ооцит, известный как цитопласт, удаления половины ооцита с ядром и отбора получающегося в результате цитопласта для дальнейшего способа по изобретению. Альтернативно, разделение осуществляют путем деления на три, приводящее в результате к трем частям, из которых две части представляют собой цитопласты. В еще одном воплощении ооцит может быть разделен на четыре части, что приводит в результате к получению трех цитопласт. Ооцит даже может быть разделен на пять частей путем физического удаления, что приводит в результате к четырем цитопластам. Аналогично, ооцит может быть разделен на шесть частей, например семь частей, таких как восемь частей, например девять частей, таких как десять или более частей. Физическое разделение ооцита и последующее удаление части, несущей ядро ооцита, может быть осуществлено путем применения микрохирургического скальпеля. Ооциты могут быть отобраны для идентификации тех ооцитов, которые были успешно энуклеированы. Части ооцитов, которые несут ядерную ДНК, могут быть идентифицированы путем окрашивания флюорохромом по Хехст, причем-5 017100 указанная процедура окрашивания известна специалистам в данной области техники. Части ооцитов,несущие ядерную ДНК, отбрасывают, а энуклеированные ооциты (цитопласты) отбирают для дальнейшего способа. Блестящая оболочка. Блестящая оболочка представляет собой толстый, прозрачный, неклеточный слой или оболочку,имеющую однородную толщину, окружающую ооцит. Как правило, интактную блестящую оболочку считают важной для переноса клеточных ядер ввиду множества параметров. Один из параметров заключается в удерживании полярного тельца рядом с метафазной пластиной ооцита, что указывает на соответствующую точку для энуклеакции. Другой параметр относится к удерживанию донорной клетки близко к цитопласту ооцита перед и во время слияния. Также полагают, что блестящая оболочка обеспечивает защиту донорной клетки и цитопласта во время слияния. Наконец полагают, что развитие эмбриона после восстановления и активации поддерживается блестящей оболочкой. Модификация по меньшей мере части блестящей оболочки может быть осуществлена при помощи множества способов. Например, для модификации блестящей оболочки может быть использована физическая манипуляция. Но также может быть использована химическаая обработка агентами, такими как кислые растворы (кислый Тироде). Один из примеров химических агентов, которые могут быть использованы в настоящем изобретении, представляют собой кислые растворы, например Тироде. В конкретном воплощении изобретения блестящая оболочка модифицирована путем ферментативного расщепления. Такое ферментативное расщепление может быть осуществлено при помощи ферментов, включающих, например, трипсин. Альтернативно, может быть использована специфическая протеаза, такая как проназа. В предпочтительном воплощении ферментативное расщепление приводит в результате по меньшей мере к частичному расщеплению части блестящей оболочки, что в предпочтительном воплощении по настоящему изобретению обозначает, что по меньшей мере часть блестящей оболочки удалена или что блестящая оболочка частично удалена. В контексте по настоящему изобретению блестящая оболочка полностью не удалена. В соответствии с особенно предпочтительным воплощением по настоящему изобретению частично расщепленная часть блестящей оболочки характеризуется тем, что блестящая оболочка остается видимой, и тем, что ооцит не деформирован. Частичное расщепление может быть достигнуто путем воздействия протеазой. В еще одном воплощении по настоящему изобретению частичное расщепление может сопровождаться путем применения проназы. В еще одном воплощении частичное расщепление может быть достигнуто путем комбинации протеазы и проназы. В предпочтительном воплощении концентрация проназы находится в диапазоне от 0,1 до 10 мг/мл,таком как от 0,5 до 10 мг/мл, например от 1 до 10 мг/мл, таком как от 1,5 до 10 мг/мл, например от 2 до 10 мг/мл, таком как от 2,5 до 10 мг/мл, например от 2,75 до 10 мг/мл, таком как от 3 до 10 мг/мл, например от 3,25 до 10 мг/мл, таком как от 3,3 до 10 мг/мл, например от 3,5 до 10 мг/мл. Предпочтительное воплощение представляет собой концентрация проназы в диапазоне от 2 до 5 мг/мл, таком как от 2,25 до 5 мг/мл, например от 2,5 до 5 мг/мл, таком как от 2,75 до 5 мг/мл, например от 2,8 до 5 мг/мл, таком как от 2,9 до 5 мг/мл, например от 3 до 5 мг/мл, таком как от 3,1 до 5 мг/мл, например от 3,2 до 5 мг/мл, таком как от 3,3 до 5 мг/мл. Конкретное воплощение по настоящему изобретению представляет собой концентрация проназы в диапазоне от 1 до 4 мг/мл, например от 1 до 3,9 мг/мл, таком как от 1 до 3,8 мг/мл, например от 1 до 3,7 мг/мл, таком как от 1 до 3,6 мг/мл, например от 1 до 3,5 мг/мл таком как от 1 до 3,4 мг/мл, например от 1 до 3,3 мг/мл. В предпочтительном воплощении концентрация проназы находится в диапазоне от 2,5 до 3,5 мг/мл,таком как 2,75 до 3,5 мг/мл, например от 3 до 3,5 мг/мл. В конкретном воплощение концентрация проназы составляет 3,3 мг/мл. Специалисту в данной области техники ясно, что проназа должна быть растворена в подходящей среде, одна из предпочтительных сред по настоящему изобретению представляет собой T33 (забуференная Hepes среда TCM 199, содержащая 33% сыворотки крупного рогатого скота (как описано ранее - Vajta, et al., 2003). Время инкубации ооцита в растворе проназы находится в диапазоне от 1 до 30 с, таком как от 2 до 30 с, например от 3 до 30 с, таком как от 4 до 30 с, таком как от 5 до 30 с. В еще одном воплощении по настоящему изобретению время инкубации находится в диапазоне от 2 до 15 с, таком как от 2 до 14 с, например от 2 до 13 с, таком как от 2 до 12 с, например от 2 до 11 с, таком как от 2 до 10 с, например от 2 до 9 с, таком как от 2 до 8 с, например от 2 до 7 с, таком как от 2 до 6 с, например от 2 до 5 с. В конкретном воплощении по настоящему изобретению время инкубации находится в диапазоне от 3 до 10 с, таком как от 3 до 9 с, например от 4 до 10 с, таком как от 3 до 8 с, например от 4 до 9 с, таком как от 3 до 7 с, например от 4 до 8 с, таком как от 3 до 6 с, например от 4 до 7 с, таком как от 3 до 5 с,-6 017100 например от 4 до 6 с, таком как от 4 до 5 с. Особенно предпочтительное время инкубации составляет 5 с. В предпочтительном воплощении по настоящему изобретению ооцит обрабатывают в течение 5 с в растворе проназы концентрации 3,3 мг/мл при 39C. Реконструированный эмбрион. Под термином реконструированный эмбрион подразумевают клетку, которая образуется путем встраивания донорной клетки или ядра донорной клетки в энуклеированный ооцит, которые соответствует зиготе (во время нормального оплодотворения). Тем не менее, термин реконструированный эмбрион также относится к восстановленной клетке. В настоящем изобретении донорная клетка представляет собой соматическую клетку. Тем не менее, донорная клетка также может быть получена из клетки зародышевой линии. Слияние. Перенос донорной клетки или клеточного ядра, окруженного мембраной, из донорной клетки, по меньшей мере, в цитопласт по настоящему изобретению осуществляют путем слияния. В описанных ниже сценариях термин донорная клетка также относится к клеточному ядру, окруженному мембраной,из донорной клетки. Слияние может быть достигнуто при помощи множества способов. Слияние могут быть осуществлено между донорной клеткой и по меньшей мере одним цитопластом, такое как между донорной клеткой и по меньшей мере двумя цитопластами, например между донорной клетктой и по меньшей мере двумя цитопластами, таким как между донорной клеткой и по меньшей мере тремя цитопластами, таким как между донорной клеткой и по меньшей мере четырьмя цитопластами, например между донорной клеткой и по меньшей мере пятью цитопластами, таким как между донорной клеткой и по меньшей мере шестью цитопластами, например между донорной клеткой и по меньшей мере семью цитопластами, таким как между донорной клеткой и по меньшей мере восемью цитопластами. Слияние может быть осуществлено в соответствии с перечисленными выше комбинациями одновременно или последовательно. В одном из воплощений по настоящему изобретению слияние осуществляют одновременно. В еще одном воплощении слияние по меньшей мере одного цитопласта и донорной клетки осуществляют последовательно. Например, слияние может быть осуществлено путем химического слияния, при котором донорная клетка и по меньшей мере один цитопласт подвергают агентам, способствующим слиянию, таким как,например, белки, гликопротеины или углеводороды или их комбинациям. Известно множество агентов,способствующих слиянию, например полиэтиленгликоль (ПЭГ), трипсин, диметилсульфоксид (ДМСО),лектины, агглютинин, вирусы и вирус Сендай. Предпочтительно использован фитогемаглютинин (РНА). Тем не менее, может быть использован маннит и/или поливиниловый спирт. Альтернативно, слияние может быть осуществлено путем индукции постоянным током (DC) через плоскость слияния. Часто используют переменный ток (АС) для совмещения донорной и реципиентной клетки. Слияние с использованием электрического тока продуцирует достаточно высокий импульс электричества, который временно способен разрушить мембраны цитопласта и донорной клетки и затем восстановить мембраны. В результате открываются небольшие каналы между донорной клеткой и реципиентной клеткой. Мембраны донорной клетки и реципиентной клетки постепенно связываются и в итоге две клетки сливаются в одну клетку. Совмещение по меньшей мере одного цитопласта и донорной клетки может быть осуществлено путем использования переменного тока в диапазоне от 0,06 до 0,5 кВ/см, такого как от 0,1 до 0,4 кВ/см,например от 0,15 до 0,3 кВ/см. В предпочтительном воплощении совмещение по меньшей мере одного цитопласта и донорной клетки может быть осуществлено путем использования переменного тока при 0,2 кВ/см. Слияние может быть вызвано путем применения постоянного тока поперек плоскости слияния по меньшей мере одного цитопласта и донорной клетки. Постоянный ток находится в диапазоне от 0,5 до 5 кВ/см, таком как от 0,75 до 5 кВ/см, например от 1 до 5 кВ/см, таком как от 1,5 до 5 кВ/см, например от 2 до 5 кВ/см. Другое предпочтительные воплощение по настоящему изобретению представляет собой применение постоянного тока в диапазоне от 0,5 до 2 кВ/см. В еще одном предпочтительном воплощении постоянный ток может составлять 2 кВ/см. Постоянный ток предпочтительно может быть применен в диапазоне 1-15 мкс, таком как от 5 до 15 мкс, например от 5 до 10 мкс. Конкретное воплощение может составлять 9 мкс. В особенно предпочтительном воплощении слияние с постоянным током могут быть осуществлено путем использования постоянного тока 2 кВ/см в течение 9 мкс. Тем не менее, слияние с использованием электрического тока и химическое слияние также могут быть комбинированы. Типично слияние с использованием электрического тока осуществляют в камерах для слияния, известных специалистам в данной области техники. Слияние может быть осуществлено по меньшей мере в одну стадию, такое как в две стадии, например три стадии, такое как в четыре стадии, например в пять стадий, такое как шесть стадий, например-7 017100 семь стадий, такое как в восемь стадий. Слияние может быть осуществлено, например, на первой стадии, на которой по меньшей мере один цитопласт сливают с донорной клеткой. Вторая стадия слияния может включать слияние слитой пары(цитопласт-донорная клетка, реконструированный эмбрион) по меньшей мере с одним цитопластом, таким как по меньшей мере два цитопласта, например три цитопласта, такие как четыре цитопласта, например пять цитопластов, такиие как шесть цитопластов, например семь цитопластов, такие как восемь цитопластов. Вторая стадия слияния с слиянием по меньшей мере одного цитопласта и слитой пары может быть осуществлена последовательно или одновременно. В одном из воплощений по меньшей мере два цитопласта слиты со слитой парой одновременно. В еще одном воплощении по меньшей мере два цитопласта слиты со слитой парой последовательно. В одном из воплощений изобретения вторая стадия слияния также может предлставлять собой стадию активации, на которой реконструированный эмбрион активируют для вхождения в митоз, как здесь описано. Активация. В предпочтительном воплощении реконструированный эмбрион могут оставить перед активацией в течение некоторого времени для того, чтобы дать возможность ядру донорной клетки реинициировать свой геном и обеспечить тотипотентность в новом окружении энуклеированного цитопласта. Реконструированный эмбрион, например, могут оставлять в течение 1 ч перед активацией. Предпочтительно реконструированный эмбрион может быть активирован для индукции митоза. Способы активации предпочтительно могут быть выбраны из группы, состоящей из электрического импульса, вызванного химическими соединениями шока, увеличения внутриклеточных уровней двухвалентных катионов или уменьшения фосфорилирования. Для активации может быть предпочтительна комбинация способов. В одном из конкретных воплощений изобретения активация и вторая стадия слияния могут быть осуществлены одновременно. Тем не менее, активация реконструированного эмбриона и по меньшей мере одна дополнительная стадия слияния между реконструированным эмбрионом и по меньшей мере одним цитопластом могут быть осуществлены последовательно. Уменьшение фосфорилирования клеточных белков в реконструированном эмбрионе при помощи известных способов, таких как, например, путем добавления ингибиторов киназы, может активировать реконструированный эмбрион. Предпочтительное воплощение может включать применение агентов,которые ингибируют белковый синтез, например циклогексимид. В еще одном предпочтительном воплощении могут быть использованы агенты, ингибирующие образование веретена, например цитохалазин В. В одном из воплощений изобретения внутриклеточные уровни двухвалентных катионов могут быть увеличены. Двухвалентные катионы, такие как, например, кальций, могут быть включены в активирующую среду. Предпочтительно катионы могут проникать в реконструированный эмбрион, в частности, при воздействии на реконструированный эмбрион электрического импульса. В предпочтительном воплощении электрический импульс может вызвать проникновение кальция в реконструированный эмбрион. Применение электрического импульса с использованием постоянного тока может составлять стадию активации. Тем не менее, в предпочтительном воплощении электрический импульс, применяемый для активации, также может служить в качестве дополнительной стадии слияния. Перед применением электрического импульса с использованием постоянного тока по меньшей мере один цитопласт и по меньшей мере один реконструированный эмбрион могут быть совмещены путем применения переменного тока. Переменный ток может находиться в диапазоне от 0,06 до 0,5 кВ/см, таком как от 0,1 до 0,4 кВ/см, например от 0,15 до 0,3 кВ/см. В предпочтительном воплощении совмещение по меньшей мере одного цитопласта и донорной клетки может быть осуществлено с использованием переменного тока при 0,2 кВ/см. Активация может быть вызвана путем применения постоянного тока через плоскость слияния по меньшей мере одного цитопласта и донорной клетки. Постоянный тока находится в диапазоне от 0,2 до 5 кВ/см, такой как от 0,4 до 5 кВ/см, например от 0,5 до 5 кВ/см. Еще одно предпочтительное воплощение по настоящему изобретению представляет собой применение постоянного тока в диапазоне от 0,5 до 2 кВ/см. В еще одном предпочтительном воплощении постоянный ток могут составлять 0,7 кВ/см. Постоянный ток предпочтительно может быть применен в течение от 10 до 200 мкс, такое как от 25 до 150 мкс, например от 50 до 100 мкс. Конкретное воплощение изобретения может составлять 80 мкс. В особенно предпочтительном воплощении слияния с постоянным током может быть использован постоянный тока 0,7 кВ/см в течение 80 мкс. Особенно предпочтительное воплощение активации по настоящему изобретению может представлять собой применение электрического импульса в комбинации с воздействием на реконструированный эмбрион агентов, ингибирующих белковый синтез, образование веретена и двухвалентных катионов. Активация может быть осуществлена путем любой комбинации вышеописанных способов. Тип генетической модификации. Донорные клетки могут быть генетически модифицированы при помощи любого из стандартных-8 017100 способов, известных в области техники. Генетическая модификация может представлять собой модификацию геномной ДНК путем делеции, инсерции, дупликации и/или других форм мутаций, включающих точечную мутацию. Модификация может быть проведена в кодирующих последовательностях и/или некодирующих последовательностях. Конструкции ДНК для встраивания могут нести ген интересующих и/или регуляторных последовательностей, таких как промоторы, изоляторы, энхансеры, репрессоры или сайты связи с рибосомами. В некоторых воплощениях в геном вводят только одну генетическую модификацию. В других воплощениях, тем не менее, геном может быть модифицирован в более чем одном сайте. Подходящие способы генетической модификации клеток млекопитающих, таких как фибробласты, включают способы, такие как добавление генов путем негомологичной рекомбинации, замену генов путем гомологичной рекомбинации, и редактирование генов. Эти способы могут включать применение встраивания ретровируса, транспозонное перемещение и/или способы с искусственной хромосомой. Например, негомологичная рекомбинация ДНК может быть осуществлена, как описано в Kragh et al. (2004)Reprod. Fert. Dev. 16:290 или Kragh et al. (2004) Reprod. Fert. Dev. 16:315. Основанное на транспозонах перемещение генов может быть осуществлено, как описано в Izsvak et al. (1997) Cell. 91:501. Замена генов путем гомологичной рекомбинации может, например, включать способы, описанные Urnow et al.(2005) Nature 435:646. Способы редактирования генов описаны в Andersen et al. (2002) J. Mol. Med. 80:770, Liu et al. (2002) Gene Ther. 9:118 и Sorensen et al. (2005) J. Mol. Med. 83:39. Выращивание эмбрионов in vitro. Один из аспектов изобретения относится к способу выращивания эмбрионов in vitro, при котором доля бластоциста увеличивается до 25,3%. Таким образом, способ выращивания реконструированного эмбриона, находящийся в объеме настоящего изобретения, включает стадии а) получения по меньшей мере одного ооцита, имеющего по меньшей мере часть блестящей оболочки, б) разделения ооцита по меньшей мере на две части с получением ооцита, имеющего ядро, и по меньшей мере одного цитопласта,в) получения донорной клетки или клеточного ядра, обладающего желаемыми генетическими свойствами, г) слияния по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной, д) получения реконструированного эмбриона, е) активации реконструированного эмбриона с образованием эмбриона и ж) выращивания указанного эмбриона. Еще один аспект изобретения относится к способу переноса клеточных ядер, в который включена стадия выращивания эмбриона. В предпочтительном воплощении в отношении описанных здесь способов эмбрионы выращивают в наборе сред последовательно. Предпочтительно бластоцисты выращивают в традиционной среде, такой как, например NCSU37 или эквивалентная среда, известная специалисту в данной области техники, из которой глюкоза удалена и заменена другими агентами. Один из агентов может представлять собой пируват. Другой агент может представлять собой лактат. Агенты также могут быть комбинированы и заменять глюкозу в традиционной среде. Эмбрионы могут быть выращены в замещенных средах, как описано выше, на 0-3 день, такой как 02 день. Концентрация пирувата может находиться в диапазоне от 0,05 до 1 мМ, таком как от 0,1 до 1 мМ,например от 0,125 до 1 мМ, таком как от 0,15 до 1 мМ. Тем не менее, концентрация пирувата натрия также может находиться в диапазоне от 0,05 до 0,9 мМ, таком как от 0,05 до 0,8 мМ, например от 0,05 до 0,7 мМ, таком как от 0,05 до 0,6 мМ, например от 0,05 до 0,5 мМ, таком как от 0,05 до 0,4 мМ, например от 0,05 до 0,3 мМ, таком как от 0,05 до 0,2 мМ. Предпочтительно концентрация находится в диапазоне от 0,05 до 0,17 мМ. Предпочтительная концентрация пирувата натрия составляет 0,17 мМ. Концентрация лактата может находиться в диапазоне от 0,5 до 10 мМ, таком как от 0,75 до 10 мМ,например от 1 до 10 мМ, таком как от 1,5 до 10 мМ, таком как от 1,75 до 10 мМ, например от 2 до 10 мМ,таком как от 2,5 до 10 мМ. Тем не менее, концентрация лактата натрия также может находиться в диапазоне от 0,5 до 9 мМ, таком как от 0,5 до 8 мМ, например от 0,5 до 7 мМ, таком как от 0,5 до 6 мМ, например от 0,5 до 5 мМ, таком как от 0,5 до 4 мМ, например от 0,5 до 3 мМ. Предпочтительно концентрация находится в диапазоне от 1 до 5 мМ, таком как от 2 до 4 мМ, например от 2 до 3 мМ. Предпочтительная концентрация лактата натрия составляет 2,73 мМ. После исходной инкубации в среде без глюкозы последнюю вновь заменяют пируватом и лактатом. Эмбрионы могут быть выращены в среде, содержащей глюкозу, на 3-4 день, предпочтительно на 3-7 день. Концентрация глюкозы может находиться в диапазоне от 1 до 10 мМ, таком как от 2 до 10 мМ, например от 3 до 10 мМ, таком как от 4 до 10 мМ, например от 5 до 10 мМ. Тем не менее, концентрация глюкозы также может находиться в диапазоне от 1 до 9 мМ, таком как от 2 до 8 мМ, например от 3 до 7 мМ, таком как 4-6 мМ. Предпочтительная концентрация глюкозы по настоящему изобретению составляет 5,5 мМ глюкозы. В еще одном предпочтительном воплощении эмбрион представляет собой эмбрион свиньи. Генетически модифицированные животные. В соответствии с одним из воплощений по настоящему изобретению предложены генетически модифицированные или трансгенные животные, обладающие желаемыми генотипами. Понятно, что изобретение не включает способы модификации генетической идентичности живот-9 017100 ных, которые вероятно приводили бы к их страданию без какой-либо значительной медицинской пользы для человека или животного, или животные, возникающие в результате таких способов. Настоящее изобретение относится к способам получения генетически модифицированного или трансгенного млекопитающего, отличающегося от человека, включающим а) получение по меньшей мере одного ооцита, имеющего по меньшей мере часть модифицированной блестящей оболочки, б) разделение ооцита по меньшей мере на две части с получением ооцита, имеющего ядро, и по меньшей мере одного цитопласта, в) получения донорной клетки или клеточного ядра, обладающего желаемыми генетическими свойствами, г) слияние по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной, д) получения реконструированного эмбриона, е) активации реконструированного эмбриона с образованием эмбриона, ж) выращивания указанного эмбриона; и з) переноса указанного выращенного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш. Тем не менее, генетически сконструированное или трансгенное млекопитающее, отличающееся от человека, также может быть получено при помощи способа,включающего а) получение по меньшей мере одного ооцита, б) разделение ооцита по меньшей мере на три части с получением ооцита, имеющего ядро, и по меньшей мере одного цитопласта, в) получения донорной клетки или клеточного ядра, обладающего желаемыми генетическими свойствами, г) слияние по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной,д) получения реконструированного эмбриона, е) активации реконструированного эмбриона с образованием эмбриона, ж) выращивания указанного эмбриона и з) переноса указанного выращенного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш. Донорство органа или ткани. В одном из воплощений животные по изобретению могут быть использованы в качестве источника органа или ткани для людей или других животных, причем животным тех же самых видов или животных других видов. Перенос между видами обычно назван ксенотрансплантацией. Целые органы, которые могут быть трансплантированы, включают сердце, почку, печень, поджелудочную железу или легкое. Альтернативно, части органов, такие как специфические ткани органов, могут быть трансплантированы или перенесены людям или другим животным. В еще одном воплощении индивидуальная клетка или популяция индивидуальных клеток из животного по изобретению может быть перенесена человеку или другому животному для терапевтических задач. Модели заболевания. Настоящее изобретение также относится к способу клонирования млекопитающего, отличающегося от человека, в соответствии со способами по настоящему изобретению. Таким образом, один из аспектов изобретения относится к способу клонирования млекопитающего, отличающегося от человека, включающему а) получение бластоцисты как здесь описано, возможно с оттаиванием эмбриона, б) переноса указанного выращенного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш. Генетически модифицированный зародыш может развиваться в млекопитающее, отличающееся от человека. Настоящее изобретение также охватывает генетически модифицированное животное в качестве моделей заболеваний, которое получают при помощи описанных здесь способов. Таким образом, второй аспект изобретения представляет собой генетически модифицированное млекопитающее, отличающееся от человека, которое получено при помощи описанных здесь способов. Еще один аспект относится к генетически модифицированному эмбриону, отличающемуся от человека, который получают при помощи описанных здесь способов. Описанные здесь способы не включают хирургическую стадию на организме, отличающемся от организма человека. Способ переноса клеточных ядер по настоящему изобретению представляет собой средство получения модельных животных для любого соответствующего заболевания, которые желательно получать для исследования развития заболевания, возможных схем лечения, тестирования лекарств и профилактики. Выбор заболевания не ограничивается какой-либо конкретной группой заболеваний. Примеры применения настоящего изобретения для разработки моделей заболеваний на основе генетически модифицированного животного представлены ниже. Тем не менее, изобретение не ограничено перечисленными ниже примерами. Генетические модификации вводят в соматическую клетку перед переносом ядер соматических клеток (SCNT) при помощи способа ручного клонирования (НМС). Тем не менее, генетическая модификация в еще одном воплощении может быть введена во время ручного клонирования (НМС), например путем добавления на различных стадиях способа НМС трансгенов, которые затем вводят в геном эмбриона. Генетические модификации включают случайную интеграцию гена, вызывающего заболевание, мутированного гена, в геном соматической клетки. Также может быть осуществлена случайная интеграция нормального не подвергшегося мутации гена, которая вызывает заболевание при экспрессии в специфической ткани или при специфическом уровне экспрессии.- 10017100 Введенный ген или трансген может происходить из любых видов, включающих бактерии, свинью,человека, мышь, крысу, дрожжи, беспозвоночные или растения. Регуляторные последовательности трансгена могут управлять повсеместной или индуцируемой, или ткане- и/или времяспецифической экспрессией и также могут происходить из любых видов, включающих свинью, человека, мышь, крысу,дрожжи, беспозвоночные или растения. Важно, что генетическая модификация в соматической клетке может быть направлена на специфическую область в геноме свиньи путем гомологичной рекомбинации целевой конструкции или при помощи способов редактирования гена. Она может представлять собой инактивацию (например, нокаут) специфических генов, что вызывает заболевание или фенотип, или может представлять собой интеграцию (нокин) специфических мутаций в специфические гены, что затем вызывает заболевание. Кроме того, трансгены, вызывающие заболевание, могут быть встроены в специфические регуляторные области генома свиньи при помощи способов гомологичной рекомбинации. Генетические модификации, введенные в геном свиньи перед или во время способа НМС, также могут представлять собой эпигенетические модификации (например, метилирование ДНК или метилирование или ацетилирование/деацетилирование гистонов) путем инкубации соматических клеток, ооцитов или реконструированных путем НМС эмбрионов с химическими компонентами, такими как трикостатин или соединения, обладающие похожим действием. Изобретение относится к генетически модифицированным животным в качестве моделей заболеваний, например моделей дегенеративных заболеваний, нарушений укладки митохондриальных белков,болезни Альцгеймера, болезни Паркинсона, хореи Гентингтона или склероза. Тем не менее, также модель наследственной болезни Альцгеймера представляет собой воплощение настоящего изобретения. В еще одном воплощении модели заболевания могут включать все виды раковых заболеваний, например рак молочной железы. Но могут быть исследованы все раковые заболевания, такие как рак толстой кишки или рак легкого. Другие воплощения относятся к моделям генетических сенсорных систем для анализа проникновения через кожу терапевтически активных молекул или пластичных липосом. Еще одно воплощение относится к моделям заболеваний для лечения ран или лечения язвы. Кроме того, модели заболеваний для лечения пороков развития, например путем реконструктивной хирургии, находятся в объеме настоящего изобретения. Также в объеме настоящего изобретения находятся модели заболеваний, связанные с конструированием ткани, такие как трансплантация клеток, трансплантация ткани, трансплантация органов. Другие модели заболеваний представляют собой модели псориаза и/или модели эпидермолитических расстройств, такие как врожднный буллезный эпидермолиз. Кроме того, воплощения настоящего изобретения представляют собой модели лечения и профилактики заболеваний, вызванных атеросклерозом, ишемической болезнью сердца. Модели также включают модели метаболических расстройств, которые ведут к спектру обычных заболеваний, таких как, например, диабет или ожирение. Кроме того, атеросклероз и сердечнососудистое заболевание исходно могут быть вызваны метаболическими расстройствами. Повреждение почки представляет собой еще один пример заболевания, которое может быть вызвано метаболической дисфункцией. Аналогично, высокое кровяное давление (гипертензия) также может являться следствием исходно метаболической дисфункции и может быть исследовано в моделях метаболических расстройств у генетически модифицированных животных. Кроме того, заболевание вызвано мутацией митохондриальных белков, например дефицит ацил-КоА-дегидрогеназы с короткой цепью, нейромышечная слабость, дегенерация путем экспрессии варианта орнитинтранскарбамилазы с делецией. Витрификация. Термин криоконсервация использован для различных способов замораживания клетки, включенных в процесс замораживания, хранения и оттаивания живых клеток. Витрификация представляет собой форму криоконсервации, при котором живые клетки быстро охлаждают таким образом, что жидкость в клетке не образует лед. Таким образом, витрификация относится к способу охлаждения, при котором клетки или целые ткани консервируют путем охлаждения до низких температур меньше нуля градусов,таких как (типично) -80C или -196C (температура кипения жидкого азота). При этих небольших температурах любая биологическая активность, включающая биохимические реакции, которые могут приводить к клеточной гибели, эффективно прекращаются. Тем не менее, витрификация относится к специальному подходу, при котором в среде и консервированных клетках или тканях не допускают образования льда. Это охлаждение без льда может быть достигнуто путем применения высоких концентраций растворов криоконсерванта и чрезвычайно высоких скоростей охлаждения. Нагревание также должно быть осуществлено с быстрым увеличением температуры. Один из аспектов настоящего изобретения относится к возможности витрификации (криоконсервации) ооцита, цитопласта, клеток, эмбрионов или бластоцист. Таким образом, в настоящем изобретении раскрыты способы криоконсервации свиного эмбриона, включающие а) получение по меньшей мере одного свиного ооцита, б) делипидизацию ооцитов, в) активацию реконструированного эмбриона с образованием эмбриона, г) выращивание указанного эмбриона, д) витрификацию эмбриона. Кроме того, дели- 11017100 пидизированный ооцит может быть разделен по меньшей мере на две части, как здесь описано, с получением ооцита, имеющего ядро, и по меньшей мере одного цитопласта. В частности, изобретение относится к витрификации ооцита, тем не менее, изобретение также относится к витрификации эмбрионов, предпочтительно эмбрионов на стадии бластоцисты. В одном из воплощений эмбрион выращивают до стадии бластоцисты перед витрифиацией. В особенности настоящее изобретение охватывает эмбрионы. Кроме того, подвергнутые витрификации цитопласты охвачены настоящим изобретением, так же как и клетки. Еще один аспект изобретения относится к криоконсервации свиного эмбриона, полученного при помощи способа переноса клеточных ядер, как здесь описано, включающего стадию витрификации свинного эмбриона. Еще один аспект изобретения относится к свинному эмбриону, полученному или получаемому при помощи предложенных здесь способов. Использованный здесь термин криоконсервация может относится к витрификации ооцита, цитопласта, клетки, эмбриона или животного по изобретению. Температуры, использованные для криоконсервации, предпочтительно составляют меньше -80C и более предпочтительно меньше чем -196C. Ооциты, клетки и эмбрионы по изобретению могут быть подвергнуты криоконсервации в течение неопределенного периода времени. Известно, что биологические материалы могут быть подвергнуты криоконсервации на срок более чем пятьдесят лет. В объеме настоящего изобретения эмбрионы могут быть подвергнуты криоконсервации перед переносом в млекопитающее-хозяин при использовании способов получения генетически сконструированного или трансгенного млекопитающего, отличающегося от человека. Такая криоконсервация перед переносом может быть осуществлена на стадии бластоцисты во время развития эмбриона. Один из аспектов изобретения относится к неинвазивной делипидизации ооцитов путем мягкой обработки ферментативным агентом, например проназой. В предпочтительном воплощении концентрация проназы предпочтительно находится в диапазоне от 0,5 до 5 мг/мл, таком как от 0,5 до 3 мг/мл, например от 0,5 до 2 мг/мл. Предпочтительно концентрация проназы составляет 1 мг/мл. В еще одном воплощении настоящего изобретения неинвазивную делипидизацию ооцитов осуществляют путем обработки проназой в концентрации 3,3 мг/мл. Делипидизацию ооцитов осуществляют в присутствии подходящей среды, например среды, включающей 50% сыворотку крупного рогатого скота. Процессу делипидизации дают возможность протекать в течение периода предпочтительно в диапазоне от 1 до 5 мин, в частности в течение 3 мин. Тем не менее, в случаях, когда концентрация проназы находится в диапазоне от 2,5 до 5 мг/мл, период, в течение которого дают возможность протекать процессу делипидизации, находится в диапазоне от 5 до 15 с, например от 5 до 10 с, таком как 10-15 с. Одно из воплощений настоящего изобретения представляет собой делипидизацию ооцитов с использованием проназы в концентрации 3,3 мг/мл в течение 10 с. Предпочтительно ооциты затем промывают в подходящей среде, например среде ТСМ-199, забуференной Hepes, дополненной телячьей сывороткой, например 20% телячьей сывороткой. Расщепленные проназой и отмытые ооциты предпочтительно подвергают центрифугированию в диапазоне от 8000 до 15000g, например от 9000 до 14000g. В особенно предпочтительном воплощении ооциты центрифугируют при 12000g. Центрифугирование может быть осуществлено в диапазоне от 10 до 30 мин, таком как в течение 20 мин. В особенно предпочтительном воплощении настоящего изобретения ооциты подвергают делипидизации при помощи проназы в концентрации 1 мг/мл в течение 3 мин, после чего ооциты могут быть отмыты и затем подвергнуты центрифугированию при 12000g в течение 20 мин. В предпочтительном воплощении изобретения делипидизированные ооциты могут быть подвергнуты витрификации. В соответствии с одним из воплощений изобретения делипидизированные ооциты могут быть подвернуты витрификации и затем нагреты для использования в способах по настоящему изобретению. В альтернативном воплощении делипидизированные ооциты могут быть использованы в описанных здесь способах с получением, например, эмбрионов, в частности эмбрионов на стадии бластоцисты, которые предпочтительно могут быть подвергнуты витрификации. Таким образом, витрификации могут быть подвергнуты ооциты, цитопласты, клетки, эмбрионы или эмбрионы на стадии бластоцисты. Подвергнутые витрификации бластоцисты, полученные при помощи способа витрификации по настоящему изобретению, могут храниться и при нагревании могут быть имплантированы в подходящее млекопитающее, отличающееся от человека, с получением генетически модифицированного млекопитающего в соответствии со способами переноса клеточных ядер по настоящему изобретению. Митохондрии. Клетки ткани генетически модифицированного млекопитающего, отличающегося от человека,и/или эмбрионов, отличающихся от человеческого, полученные при помощи настоящего изобретения,могут нести митохондрии различных материнских источников. В предпочтительном воплощении млекопитающее, отличающееся от человека, и/или эмбрион, отличающийся от человеческого, могут нести митохондрии только из одного материнского источника. Тем не менее, в другом предпочтительном вопло- 12017100 щении млекопитающее, отличающееся от человека, и/или эмбрион, отличающийся от человеческого,могут нести митохондрии по меньшей мере из двух материнских источников, таких как три материнских источника, например четыре материнских источника, таких как пять материнских источников, например шесть материнских источников, таких как семь материнских источников, например восемь материнских источников, таких как девять материнских источников, например десять материнских источников. Вероятность определенного количества материнских источников может быть рассчитана, основываясь на обнаруженных типах митохондрий. Примеры Если не указано иное, все химические соединения получены из Sigma Chemical Co. (St Louis, MO,USA). Сбор ооцитов и созревание in vitro (IVM). Комплексы кумулюс-ооцит (СОС) аспирировали из фолликул размером 2-6 мм свиней со скотобойни или яичников молодых свиней. СОС подвергали созреванию в группах по 50 в 400 мкл забуференного бикарбонатом ТСМ-199 (GIBCO BRL), дополненного 10% (об./об.) сыворотки крупного рогатого скота(CS), 10% (об./об.) фолликулярной жидкости свиней, 10 Е/мл хорионического гонадотропина жеребой кобылы (eCG), 5 Е/мл человеческого хорионического гонадотропина (hCG) (Suigonan Vet; Skovlunde,Denmark) при 38,5C в "Системе инкубации подводного судна" (SIS; Vajta, et al., 1997) в 5% CO2 во влажной атмосфере в течение 41-44 ч. Оплодотворение in vitro (IVF). Эксперименты IVF осуществляли с использованием ооцитов, подвергнутых созреванию in vitro, в 3 идентичных повторах. После созревания СОС дважды промывали mTBM, содержащим 2 мМ кафеин(mTBMfert) и переносили в группах по 50 в 400 мкл mTBMfert. Свежеэякулированную сперму обрабатывали, как описано ранее (Booth, et al., в печати). После капацитации в течение 2 ч при 38,5C и в 5% CO2 в увлажненной атмосфере сперму добавляли к ооцитам с получением конечной концентрации 1105 сперматозоидов/мл. Оплодотворение осуществляли при 38,5C и в 5% CO2 в увлажненной атмосфере в SIS в течение 3 ч. После оплодотворения предполагаемые зиготы подвергали перемешиванию в mTBMfert для удаления клеток кумулюса перед промыванием в среде IVC и помещали в культуральные чашки (см. Выращивание и анализ эмбриона). Ручное клонирование (НМС). Применение способа НМС основано на предшествующей работе авторов изобретения на крупном рогатом скоте и свиньях (Kragh, et al., 2004; Peura and Vajta, 2003; Vajta, et al., 2003), но со значительными модификациями. Кратко, через 41 ч после начала созревания покров из кумулюса СОС удаляли путем повторного пипетирования в 1 мг/мл гиалуронидазы в ТСМ-199, забуференной Hepes. С этого момента (за исключением того, если не указано иное) все манипуляции осуществляли на стадии нагревания при 39C, и все капли, использованные для обработки ооцитов, имеющие объем 20 мкл, были покрыты минеральным маслом. Ооциты недолго инкубировали в 3,3 мг/мл проназы, растворенной в T33 (Т для среды ТСМ 199,забуференной Hepes; число указывает процент (об./об.) добавки телячьей сыворотки CS, в данном случае 33%) в течение 5 с. Перед тем как ооциты теряли форму в растворе проназы, их извлекали и быстро промывали в каплях Т 2 и Т 20. Ооциты с частично расщепленной, но все еще видимой блестящей оболочкой располагали в каплях T2, дополненных 3 мг/мл поливинилового спирта (TPVA) и 2,5 мкг/мл цитохалазина В. Разделение на три части вместо разделения на две части осуществляли вручную под наблюдением в стереомикроскоп с использованием сверхострых разделяющих лезвий (АВ Technology, Pullman, WA,USA; фиг. 1a). Фрагменты разделенных на три части ооцитов собирали и окрашивали каплями 5 мкг/мл флюорохрома Хехст 33342 в TPVA в течение 5 мин, затем помещали в 1 мкл капли среды TPVA на дно чашки Петри Falcon диаметром 60 мм с маслом (3-4 фрагмента на каплю). С использованием инвертированного микроскопа и УФ освещения регистрировали позиции фрагментов без окрашивания хроматина(цитопласты) и позже собирали под стереомикроскопом в каплях Т 10 до начала слияния. Фетальные клетки фибробласта готовили, как описано ранее (Kragh, et al., в печати). Слияние осуществляли в две стадии, где вторая стадия также включала начало активации. Для первой стадии использовали одну треть отобранных цитопластов (предпочтительно меньшие части). При помощи тонко вытянутой и оплавленной стеклянной пипетки 10 цитопластов переносили в виде группы в 1 мг/мл фитогемагглютинина (PHA; ICN Pharmaceuticals, Australia) на 3 с, затем быстро капали на одну из нескольких клеток фибробласта индивидуально, которые осаждали в капле Т 2. После прикрепления 10 пар цитопласт-клетка фибробласта уравновешивали в среде для слияния (0,3 М маннит и 0,01% поливинолового спирта PVA) в течение 10 с. С использованием переменного тока (АС) 0,6 кВ/см и 700 кГц, пары клеток подгоняли к спирали камеры для слияния (камера для слияния ВТХ microslide 0,5 мм, модель 450; ВТХ,SanDiego, СА, USA), причем донорные клетки располагались дальше всего от спирали (фиг. 16), затем использовали постоянный ток (DC) 2,0 кВ/см в течение 9 мкс. После электрического импульса пары клеток осторожно удаляли со спирали, переносили в капли Т 10 и инкубировали для обнаружения того, произошло ли слияние.- 13017100 Приблизительно через 1 ч после первого слияния слитые пары вместе с оставшимися двумя третями цитопластов уравновешивали в отдельных каплях активирующей среды (0,3 М маннит, 0,1 мМMgSO4, 0,1 мМ CaCl2 и 0,01% поливиниловый спирт (PVA. При переменном токе 0,6 кВ/см триплеты цитопласт-слитая пара-цитопласт последовательно подгоняли к спирали в группах по 10, причем слитые пары располагались посередине (фиг. 1 в). Для второго слияния и инициации активации использовали единичный импульс постоянного тока 0,7 кВ/см в течение 80 мкс. Затем триплеты удаляли со спирали и осторожно переносили в капли Т 10 для контроля слияния (фиг. 1 г). Реконструированные эмбрионы инкубировали в культуральной среде (см. Выращивание и оценка эмбрионов), дополненной 5 мкг/мл цитохалазином В и 10 мкг/мл циклогексимидом в течение 4 ч при 38,5C в 5% CO2, 5% O2 и 90% N2 с максимальной влажностью, затем тщательно трижды промывали в среде IVC перед выращиванием культуры. Партеногенетическая активация (РА). Партеногенетически активированные ооциты готовили отдельно или параллельно с НМС. Ооциты обнажали также, как описано выше, за исключением того, что использовали более длительную инкубацию в проназе для полного удаления блестящей оболочки. Ооциты без блестящей оболочки (ZF) затем уравновешивали в течение 10 с в активирующей среде (0,3 М маннит, 0,1 мМ MgSO4, 0,1 мМ CaCl2 и 0,01% PVA) и переносили в камеру для слияния (камера для слияния ВТХ microslide 0,5 мм, модель 450; ВТХ, SanDiego, CA, USA). Единичный импульс постоянного тока 0,85 кВ/см в течение 80 мкс генерировали при помощи аппарата для слияния клеток BLS CF-150/B (BLS, Budapest, Hungary) и применяли в отношении ооцитов ZF. Для ооцитов с интактной блестящей оболочкой (ZI) использовали единичный импульс постоянного тока 1,25 кВ/см в течение 80 мкс (в соответствии с неопубликованными предшествующими экспериментами авторов изобретения, эти параметры привели в результате к более высокой активации и последующему развитию in vitro соответственно ооцитов ZI и ZF). Процедура после электрического импульса была такой же, как для реконструированных эмбрионов НМС. Выращивание и анализ эмбрионов. Все свиные эмбрионы, полученные при помощи вышеприведенных обработок, выращивали в модифицированной среде NCSU37 (Kikuchi, et al., 2002), содержащей 4 мг/мл бычьего сывороточного альбумина (BSA) при 38,5C в 5% О 2, 5% CO2 и 90% N2 с максимальной влажностью. Культуральную среду дополняли 0,17 мм пируватом натрия и 2,73 мм лактатом натрия с 0 дня (день оплодотворения и активации) по 2 день, затем лактат натрия и пируват натрия заменяли 5,5 мм глюкозы с 2 по 7 день. Все эмбрионы ZF выращивали в системе WOW (Vajta, et al., 2000) в той же самой культуральной среде и использованной выше смеси газов, с осторожной заменой среды на 2 день без удаления эмбрионов изWOW. Уровень бластоцисты регистрировали на 7 день. Для определения общего количества клеток бластоцисты фиксировали и наносили на предметные стекла для микроскопии в глицероле с 20 мкг/мкл флюорохрома Хехст 33342. После окрашивания в течение 24 ч эмбрионы визуализировали под инвертированным микроскопом Diaphot 200 с эпифлюоресцентной насадкой и фильтром UV-2A (Nikon, Tokyo,Japan). Пример 1. Различия в способности к развитию между ооцитами свиньи (2,5 года, масса 170 кг) и молодой свиньи (5,5-6 месяцев, масса 75 кг) исследовали путем ZF и ZI РА после 44 ч созревания in vitro. Исследовали четыре объединенные группы в 3 идентичных повторах: (1) ооциты ZF свиней (2) ооциты ZI свиней(3) ооциты ZF молодых свиней (4) ооциты ZI молодых свиней. Для активации ZF применяли единичный импульс постоянного тока 0,85 кВ/см в течение 80 мкс, тогда как для активации ооцитов ZI использовали единичный импульс 1,25 кВ/см. После выращивания в течение 7 дней, как описано выше, определяли процентную долю бластоцист на активированный эмбрион. Исследовали способность к развитию in vitro для партеногенетически активированных ооцитов, полученных у свиней или молодых свиней. Как представлено в табл. 1, уровень бластоцист после ZF или ZI РА для партеногенетически активированных ооцитов из свиней был значительно выше по сравнению с уровнем из молодых свиней.- 14017100 Таблица 1 Развитие бластоцисты на 7 день для партеногенетически активированных ооцитов свиньи и молодой свиньи Различные верхние индексы обозначают значимые различия (p 0,05). Различные верхние индексы обозначают значимые различия (p 0,05).Процент (среднееCO) эмбрионов, развившихся до бластоцисты. Различие в способности к развитию ооцитов у свиней и молодых свиней раздельно исследовали при продукции in vitro (IVP) и переносе ядер соматических клеток (SCNT) эмбрионов, что привело в результате к заключению, похожему на заключение в более ранней публикации других исследовательских групп (Sherrer, et al., 2004; Hyun, et al., 2003), то есть что эмбрионы, развившиеся из ооцитов, полученных у свиней, превосходят эмбрионы, развившиеся из ооцитов, полученных у молодых свиней, в отношении поддержания развития бластоцисты. Хотя молодые свиньи, использованные в исследовании авторов настоящего изобретения, имели пограничный возраст зрелости, различие между бластоцистами на 7 день после РА было значимым, что приводит к лучшей способности к развитию ооцитов свиней. Пример 2. Реализуемость модифицированного НМС на ооцитах свиньи исследовали в 6 идентичных повторах с IVF и параллельно с ZF РА в качестве контроля. Более компетентные ооциты свиньи (по примеру 1) использовали в примере 2. Через семь дней после реконструкции и/или активации определяли количество бластоцист на реконструированный эмбрион и общее количество клеток в случайно выбранной бластоцисте. Более чем 90% фрагментов ооцитов, полученных из морфологически интактных ооцитов, могут быть получены для НМС после разделения на три части. В среднем 37 эмбрионов могут быть реконструированы из 100 зрелых ооцитов. Способность к развитию для всех источников свиных эмбрионов представлена в табл. 2. На 7 день развитие реконструированных эмбрионов до стадии бластоцисты составляло 174% при среднем количестве клеток 465, тогда как доля бластоцист для IVF и ZF РА составляла соответственно 306% и 474% (n=243, 170, 97). Таблица 2 Развитие in vitro эмбрионов, продуцированных путем НМС, IVF и ZF РА Различные верхние индексы обозначают значимые различия (p 0,05). Различные верхние индексы обозначают значимые различия (p 0,05). Хотя теоретическая максимальная эффективность не была достигнута, интеграция частично расщепленной блестящей оболочки и расщепление на три части ооцита почти удваивало количество реконструированных эмбрионов по сравнению с более ранней системой (Kragh, et al., 2004 Reprod. Fertil. Dev 16,315-318). Это увеличение эффективности реконструкции может обладать специальными преимуществами при клонировании у свиней, поскольку выделение ооцитов после аспирации требует больше затрат ресурсов и времени по сравнению с крупным рогатым скотом. Еще более важный момент представляет собой большое количество эмбрионов, требующееся для обеспечения беременности после переноса свиных ядер. IVC у свиней также рассматривают как ресурсоемкую и неэффективную процедуру (Reed, et- 15017100 достигать по меньшей мере компактной морулы или стадии ранней бластоцисты in vitro для того, чтобы избежать разрушения в яйцеводе без защитного слоя блестящей оболочки. С другой стороны, на стадии бластоцисты эмбрионы без блестящей оболочки могут быть перенесены успешно, о чем свидетельствует рождение телят после переноса ядер эмбриональных или соматических клеток (Peura et al., 1998; Tecirlioglu et al., 2004; Oback et al., 2003; Vajta, et al., 2004), а также рождение поросят после IVP ооцитов без блестящей оболочки (Wu, et al., 2004). Среда NCSU37 представляла собой наиболее широко и успешно применяемую среду для выращивания свиных эмбрионов. Тем не менее, несмотря на улучшенное развитие эмбрионов по сравнению с другими средами, жизнеспособность IVP свиных эмбрионов все же нарушена после IVC. В некоторых обзорах указано, что глюкоза не метаболизируется легко эмбрионами свиньи на ранней стадии перед восьмиклеточной стадией, но используется в более высоких количествах у эмбрионов между стадиями компактной морулы и бластоцисты (Flood, et al., 1988). Замена глюкозы на пируват и лактат в NCSU37 в течение первых 2 дней выращивания культуры приводит в результате к доле бластоцист 25,3% для IVP свиных эмбрионов в исследовании Кикуши (Kukuchi, et al., 2002), что дополнительно подтверждено в настоящем исследовании авторов изобретения с долей IVP бластоцист в среднем 30%. Кроме того, первая оценка этой последовательной системы выращивания на свиных эмбрионах НМС и ZF РА привела в результате к доле бластоцист, соответственно, 17 и 47%. Иногда доля бластоцист для ZI РА может даже достичь уровня 90% (Du, неопубликованное). Статистический анализ. Дисперсионный анализ ANOVA осуществляли с использованием SPSS 11.0, вероятность P0,05 рассматривали как статистически значимую. Пример 3. Витрификация подвергнутых ручному клонированию бластоцист свиньи, продуцируемых из делипидизированных созревших in vitro ооцитов. Недавно был опубликован неинвазивный способ делипидизации эмбрионов свиньи путем центрифугирования, но без последующей микроманипуляции (Esaki et al., 2004, Biol Reprod. 71, 432-6). Криоконсервацию эмбрионов/бластоцист осуществляли путем витрификации с использованиемCryotop (Kitazato Supply Co, Fujinomiya Japan), как описано ранее (Kuwayama et al. 2005a; 2005b). В момент витрификации эмбрионы/бластоцисты переносили в уравновешивающий раствор (ES), состоящий из 7,5% (об./об.) эталенгликоля (EG) и 7,5% диметилсульфоксида (DMSO) в ТСМ 199, дополненный 20% синтетическим заместителем сыворотки крови (SSS) при 39C в течение 5-15 мин. После исходного уменьшения размеров эмбрионы восстанавливали свой исходный объем. 4-6 эмбрионов/бластоцист переносили в 20 мкл каплю раствора для витрификации (VS), состоящего из 15% (об./об.) EG и 15%(DMSO) и 0,5 М сахарозы, растворенной в ТСМ 199, дополненной 20% SSS. После инкубации в течение 20 с эмбрионы загружали в Cryotop и погружали в жидкий азот. Процесс от воздействия VS до погружения завершали в течение 1 мин. Эмбрионы/бластоцисты отмораживали путем погружения Cryotop непосредственно в раствор для оттаивания (TS), состоящий из 1,0 М сахарозы в ТСМ 199 плюс 20% SSS в течение 1 мин, затем переносили в разбавляющий раствор (DS), состоящий из 0,5 М сахарозы в ТСМ 199 плюс 20% SSS в течение 3 мин. Для удаления криоконсерванта эмбрионы/бластоцисты дважды промывали в промывающем растворе (WS; TCM199 плюс 20% SSS), 5 мин для каждого момента времени. Жизнеспособность подвергнутых стеклованию бластоцист определяли в соответствии с уровнем обратного расширения после восстановления в течение 24 ч в культуральной среде, дополненной 10% телячьей сывороткой (CS). Способ неинвазивной делипидизации применяли в отношении созревших in vitro свиных ооцитов и дальнейшее развитие делипидизированных ооцитов после партеногенетической активации исследовали в 4 идентичных параллелях. Ооциты произвольным образом разделяли на группы ооцитов, предназначенных для делипидизации, и контрольные ооциты. Для делипидизации ооциты расщепляли 1 мг/мл проназы в присутствии 50% сыворотки крупного рогатого скота (CS) в течение 3 мин и промывали в среде ТСМ-199, забуференной Hepes, дополненной 20% CS, что приводило в результате к частичному расщеплению блестящей оболочки (фиг. 2 а). Затем 4050 ооцитов центрфугировали (12000g, 20 мин) при комнатной температуре в среде ТСМ-199, забуференной Hepes, дополненной 2% CS, 3 мг/мл PVA и 7,5 мкг/мл цитохалазином В (СВ) (фиг. 2 б). Блестящие оболочки центрифугированных и интактных ооцитов полностью удаляли с дополнительным расщеплением в 2 мг/мл растворе проназы. Для активации в отношении обеих групп применяли единичный импульс постоянного тока 85 кВ/см в течение 80 мкс с последующей обработкой 5 мкг/мл СВ и 10 мкг/мл циклогексимида (СНХ) в течение 4 ч. Все эмбрионы затем выращивали в модифицированной среде NCSU37. Семидневные бластоцисты подвергали витрификации и нагревали с использованием способа Cryotop (Kuwayama et al.,RBM Online, в печати) при 38,5C. Жизнеспособность подвергнутых витрификации бластоцистов определяли в соответствии с уровнями повторного расширения после восстановления в течение 24 ч в культуральной среде, дополненной 10% CS. Количество клеток в подвергнутых повторному расширению бластоцистах в обеих группах определяли после окрашивания по Хехст. Результаты сравнивали путем анализа ANOVA. Частичное расщепление блестящей оболочки и центрифугирование приводило в ре- 16017100 зультате к успешной делипидизации у 173/192 (90%) ооцитов. Развитие до бластоцист не различалось между делипидизированными и интактными ооцитами (287% по сравнению с 285% соответственно;P0,05). Тем не менее, уровни жизнеспособности бластоцист, происходящих из делипидизированных ооцитов, были значительно выше, чем уровни жизнеспособности бластоцист, происходящих из интактных ооцитов (85+6% по сравнению с 327% соответственно; P0,01). Отсутствует различие между средним количеством клеток в повторно расширившихся бластоцистах, происходящих из делипидизированных или интактных ооцитов (367 по сравнению с 389 соответственно; P0,05). Результаты демонстрируют, что способ простой делипидизации не препятствует способности к развитию in vitro активированных свиных ооцитов и улучшает криовыживание продуцируемых бластоцист без существенной потери в количестве клеток. После делипидизации блестящую оболочку ооцитов из обеих групп полностью удаляли. Те же самые параметры электрической активации, как описанные выше, применяли для обеих групп. Через семь дней после активации определяли долю бластоцист и количество клеток в бластоцистах. Исследовали реализуемость применения неинвазвного способа делипидизации в отношении ооцитов свиньи, подвергнутых созреванию in vitro. 90% (173/192) ооцитов успешно могут быть подвергнуты делипидизации. Как представлено в табл. 3, развитие в бластоцисты не различалось между делипидизированными и интактными ооцитами (287% по сравнению с 285% соответственно; P0,05). Тем не менее, уровни жизнеспособности бластоцист, происходящих из делипидизированных ооцитов, были значительно выше, чем уровни жизнеспособности бластоцист, развившихся из интактных ооцитов (856% по сравнению с 327% соответственно; P0,01). Отсутствует различие в среднем количестве клеток для повторно расширившихся бластоцист, происходящих из делипидизированных или интактных ооцитов(367 по сравнению с 389 соответственно; P0,05). Таблица 3 Способность к развитию и криовыживание подвергнутых витрификации-оттаиванию эмбрионов из делипидизированных и интактных активированных ооцитов Ручное клонирование делипидизированных ооцитов. Делипидизированные ооциты использовали для НМС в 5 повторах. В качестве контрольной системы для НМС использовали четыре идентичных повтора с неделипидизированными ооцитами. Через семь дней после реконструкции бластоцисты, продуцированные из обеих групп, подвергали витрификации сCryotop. Уровни выживания и количество клеток в повторно расширившихся бластоцистах определяли,как описано для бластоцист, продуцированных РА. Если не указано иное, все манипуляции осуществляли на стадии нагревания при 39C, и все капли,использованные для обработки ооцитов, имели объем 20 мкл с покрытием из минерального масла. Для переноса ядер соматических клеток применяли ручное клонирование (НМС), описанное в предшествующей работе авторов изобретения (Du, et al., 2005) с единственной модификацией: для энуклеакции как делипидизированных, так и контрольных ооцитов применяли разделение на две части вместо разделения на три части. Кратко, после удаления кумулюса контрольные ооциты инкубировали в 3,3 мг/мл проназы, растворенной в T33 в течение 10 с. Перед тем как ооциты теряли форму в растворе проназы, их извлекали и быстро промывали в каплях Т 2 и Т 20. Делипидизированные ооциты после центрифугирования расщепляли в 3,3 мг/мл растворе проназы в течение еще 5 с. Как контрольные, так и делипидизированные ооциты с частично расщепленной, разрыхленной и смягченной блестящей оболочкой выстраивали в каплях Т 2, дополненной 2,5 мкг/мл цитохалазином В. Разделение на две части. Разделение на три части вместо разделения на две части осуществляли вручную под наблюдением в стереомикроскоп (фиг. 2 в) с использованием сверхострых разделяющих лезвий (АВ Technology, Pullman,WA, USA). Половинки собирали и окрашивали каплями 5 мкг/мл флюорохрома Хехст 33342 в каплях Т 2 в течение 5 мин и затем помещали в 1 мкл капли среды Т 2 на дно чашки Петри Falcon диаметром 60 мм с маслом (3-4 фрагмента на каплю). С использованием инвертированного микроскопа и УФ освещения регистрировали позиции половинок без окрашивания хроматина (цитопласты). Цитопласты позже соби- 17017100 рали под стереомикроскопом и хранили в каплях Т 10. Свиные зародышевые фибробластов готовили путем расщепления трипсином из монослоев, как описано ранее (Kragh, et al., 2005). Слияние осуществляли в две стадии, где вторая стадия также включала начало активации. Для первой стадии использовали 50% имеющихся цитопластов переносили в 1 мг/мл фитогемагглютанина (PHA; ICN Pharmaceuticals, Australia), растворенного в T0, на 3 с, затем быстро капали на единичную клетку фибробласта. После прикрепления пары цитопласт-фибробласт уравновешивали в среде для слияния (0,3 М маннит и 0,01% PVA) в течение 10 с и переносили в камеру для слияния. С использованием переменного тока (АС) 0,6 кВ/см и 700 кГц, пары клеток подгоняли к спирали камеры для слияния, причем соматические клетки располагались дальше всего от спирали (фиг. 2 г),затем использовали постоянный ток (DC) 2,0 кВ/см в течение 9 мкс. После электрического импульса пары клеток осторожно удаляли со спирали, переносили в капли Т 10 и инкубировали для обнаружения того, произошло ли слияние. Приблизительно через 1 ч после первого слияния каждую пару сливали с другим цитопластом в активирующей среде. Переменный ток и единичный импульс постоянного тока 0,7 кВ/см в течение 80 мкс применяли, как описано выше. Слияние детектировали в каплях Т 10, затем реконструированные эмбрионы переносили в среду IVC0-2 (см. Выращивание и анализ эмбрионов), дополненную 5 мкг/мл цитохалазином В и 10 мкг/мл циклогексимидом. После инкубации в течение 4 ч при 38,5C в 5% CO2, 5% O2 и 90% N2 с максимальной влажностью эмбрионы 3 раза промывали в среде IVC0-2 перед выращиванием культуры. Таблица 4 Способность к развитию и криовыживание подвергнутых витрификации-оттаиванию свиных эмбрионов SCNT, происходящих из делипидизированных и интактных ооцитов Различные верхние индексы обозначают значимые различия (p 0,05).: среднееСО. Способность к развитию in vitro регистрировали при НМС с делипидизированными ооцитами при сравнении доли бластоцист на 7 день с контрольной группой НМС (216% по сравнению с 236% соответственно; P0,05; табл. 4). Уровень криовыживания после витрификации клонированных бластоцист,продуцированных из делипидизированных ооцитов, был значительно выше по сравнению с уровнем криовыживания бластоцист, развившихся из интактных ооцитов (796% по сравнению с 328 соответственно; P0,01). Пример 4. Ручная энуклеакция (САНЕ) при помощи химических соединений и сравнение с существующими способами. После созревания in vitro в течение 41-42 ч СОС дополнительно выращивали в течение 45 мин в том же самом растворе, который дополнен 0,4 мкг/мл демеколцином. Клетки кумулюса затем удаляли путем пипетирования в 1 мг/мл гиалуронидазы, растворенной в ТСМ-199, забуференной Hepes. С этого момента (если не указано иное) все манипуляции осуществляли на стадии нагревания при 39C. Все капли, использованные для обработки ооцитов, имели объем 20 мкл и были покрыты минеральным маслом. Основные стадии способа НМС описаны ранее. Кратко, ооциты без клеток кумулюса инкубировали в 3,3 мг/мл проназы, растворенной в T33 (Т для среды ТСМ 199, забуференной Hepes; число указывает на процент (об./об.) добавки телячьей сыворотки CS, в данном случае 33%) в течение 20 с. Когда происходил частичный лизис блестящих оболочек и легкая деформация ооцитов, их извлекали и быстро промывали в каплях Т 2 и Т 20. Девять ооцитов выстаивали в одной капле Т 2, дополненной 2,5 мкг/мл цитохалазином В. Путем применения тонко вытянутой и оплавленной стеклянной пипетки ооциты поворачивали для того, чтобы найти светлый конус выталкивания и/или прочно прикрепленное полярное тельце на поверхности, и направленное разделение на две части осуществляли вручную под наблюдением в стереомикроскоп с использованием сверхострых разделяющих лезвий (АВ Technology, Pullman, WA, USA). Менее чем половину цитоплазмы (рядом с местом выталкивания или РВ) выделяли из оставшейся части(фиг. 3). После разделения на две часта всех 9 ооцитов в капле более крупные части и более мелкие части(с выталкиванием или присоединенным РВ) собирали и помещали в отдельные капли Т 2 соответственно.- 18017100 Направленная ручная энуклеакция без обработки демеколцином (ОНЕ). Все стадии напоминали описанный ранее способ, но без применения преинкубации с демеколцином. Произвольное ручное разделение на две части для энуклеакции (RHE). Преинкубацию с демеколцином также устраняли из предварительной обработки этой группы. После удаления клеток кумулюса блестящие оболочки частично расщепляли при помощи проназы, как описано выше. Произвольное ручное разделение на две равные части применяли в каплях Т 2, дополненных 2,5 мкг/мл СВ. Все половины ооцитов отбирали и окрашивали 10 мкг/мл Хехст 33342 в каплях Т 2 в течение 10 мин, затем помещали в 1 мкл капли среды Т 2, покрытые минеральным маслом (по три половины ооцитов в каждой капле). С использованием инвертированного микроскопа и УФ света регистрировали позиции не содержащих хроматин половин ооцитов, т.е. цитопласты. Эти цитопласты позже собирали под стереомикроскопом и хранили в каплях Т 2 перед дальнейшими манипуляциями. Слияние и инициация активации. Свиные зародышевые фибробласты готовили, как описано ранее (Kragh, et al., 2005, Du, et al., 2005). Слияние осуществляли в две стадии, где вторая стадия также включала инициацию активации. На первой стадии с использованием тонко вытянутой и оплавленной стеклянной пипетки приблизительно 100 соматических клеток помещали в каплю Т 2 и 20-30 цитопластов помещали в каплю Т 10. После уравновешивания в течение короткого периода времени группы из 3 цитопластов переносили в 1 мг/мл фитогемагглютинина (РНА) в течение 2-3 с, затем каждый цитопласт быстро капали на единичную соматическую клетку. После прикрепления пары цитопласт-соматическая клетка вновь извлекали и переносили в среду для слияния (0,3 М маннит, дополненный 0,01% [мас./об.] PVA). Путем использования переменного тока(АС) 0,6 кВ/см и 700 кГц, уравновешенные пары подгоняли к одной спирали камеры для слияния (камера для слияния ВТХ microslide 0,5 мм, модель 450; ВТХ, San Diego, CA), причем соматические клетки находились дальше всего от спирали, затем сливали с единичным импульсом постоянного тока (DC) 2,0 кВ/см в течение 9 мкс. Пары затем осторожно удаляли со спирали в каплю Т 10 и дополнительно наблюдали, произошло ли слияние. Приблизительно 1 ч после слияния слитые пары и оставшиеся цитопласты раздельно уравновешивали в активирующей среде (0,3 М маннит, 0,1 мМ MgSO4, 0,1 мМ CaCl2, дополненная 0,01% [мас./об.]PVA). Путем применения постоянного тока 0,6 кВ/см одну пару и один цитопласт подгоняли к одной спирали камеры для слияния, причем слитые пары контактировали со спиралью. Единичный импульс постоянного тока 0,86 кВ/см в течение 80 мкс использовали для второго слияния и инициации активации. Слияние контролировали после инкубации в каплях Т 10. Традиционное клонирование (ТС). Микроманипуляцию осуществляли с использованием инвертированного микроскопа Diaphot 200(Nikon, Tokyo, Japan), как описано ранее (Chen et al., 1999; Zhang et al., 2005). Кратко, после 42-44 ч созревания in vitro клетки кумулюса удаляли, как описано выше. Все манипуляции осуществляли на стадии нагревания при 39C. Единичную каплю раствора для микроманипуляции объемом 50 мкл наносили в центральную область на крышку культуральной чашки диаметром 60 мм и покрывали минеральным маслом. Группы из 20-30 ооцитов и фетальных клеток фибробласта помещали в ту же самую каплю. После инкубации в течение 15-30 мин ооцит удерживали при помощи пипетки (внутренний диаметр 25-35 мкм, а внешний диаметр 80-100 мкм). После помещения в позицию 5-6 ч первое полярное тельце и прилегающую цитоплазму (приблизит. 10% от общего объема ооцита), предположительно содержащую метафазную пластину, аспирировали и удаляли при помощи срезанной на конус инъекционной пипетки(внутренний диаметр 20 мкм). Фетальную клетку фибробласта затем инъецировали в пространство через ту же самую щель. После ядерного переноса (NT) реконструированные комплексы переносили в капли среды, покрытые минеральным маслом, для восстановления в течение 1-1,5 ч до проведения слияния и активации. Среда для восстановления представляла собой NCSU-23, дополненная 4 мг/мл BSA и 7,5 мкг/мл СВ. Реконструированные комплексы инкубировали в среде для слияния в течение 4 мин. Комплексы вручную совмещали с использованием тонко вытянутого и оплавленного стеклянного капилляра с получением плоскости контакта параллельно электродам. Затем применяли единичный импульс постоянного тока 2,0 кВ/см длительностью 30 мкс. После выращивания культуры в каплях IVC0-2 (определенной в "Выращивание и анализ эмбрионов"), дополненной 7,5 мкг/мл СВ, в течение 30-60 мин результаты слияния проверяли под стереомикроскопом. Слитые комплексы подвергали второму импульсу в активирующем растворе. После инкубации в течение 30 мин в Т 10 их переносили в IVC0-2 для оценки развития in vitro. Дальнейшие стадии активации. После активирующего все реконструированные эмбрионы инкубировали в IVC0-2, дополненной 5 мкг/мл СВ и 10 мкг/мл циклогексимида при 38,5C в 5% CO2, 5% O2, и 90% N2, с максимальной влажностью. Выращивание и анализ эмбрионов. Через 4 ч все реконструированные и активированные эмбрионы промывали и выращивали в четырехлуночных планшетах Nunc в 400 мкл IVC0-2, покрытых минеральным маслом, при 38,5C в 5% CO2,- 19017100 5% O2, и 90% N2 с максимальной влажностью. IVC0-2 представлял собой модифицированную средуNCSU37 (Kikuchi, et al., 1999), содержащую 4 мг/мл BSA, 0,17 мМ пируват натрия и 2,73 мМ лактат натрия на день с 0 (день активации) по 2 день. Пируват натрия и лактат натрия заменяли 5,5 мМ глюкозой на 2-7 день (IVC2-7). Все эмбрионы без блестящей оболочки выращивали в системе лунка лунки(WOW) (Vajta et al., 2000) в той же самой культуральной среде и смеси газов, которые были использованы выше, с осторожной заменой среды на 2 день без удаления эмбрионов из WOW. ТС эмбрионы выращивали в группах по 15-30 в лунках четырехлуночных планшетов с использованием того же самого количества и состава среды. Расщепление и доли бластоцист регистрировали соответственно на 2 и 7 день. Для определения общего количества клеток бластоцисты фиксировали и наносили на предметное стекло для микроскопии в небольшом количестве (2 мкл) глицерина, содержащего 10 мкг/мл Хехст 33342. После окрашивания в течение нескольких часов при комнатной температуре эмбрионы визуализировали под инвертированным микроскопом Diaphot 200 с эпифлюоресцентной насадкой и фильтром UV-2A(Nikon, Tokyo, Japan). Сравнение эффективности CAHE с OHE. Эффективность и достоверность CAHE тестировали в 12 идентичных повторах с использованием в общей сложности 620 ооцитов. После созревания в течение 41-42 ч ооциты подвергали инкубации с демеколцином. Направленное разделение на две части осуществляли в отношении ооцитов, в которых после частичного расщепления проназой обнаружен конус выталкивания и/или прочно присоединенное РВ. Регистрировали процент подвергнутых разделению на две части ооцитов относительно общего количества ооцитов и выживание относительно подвергнутых расщеплению на две части ооцитов. Затем предполагаемые цитопласты и кариопласты собирали отдельно и окрашивали при помощи Хехст 33342 (10 мкг/мл в Т 2 в течение 10 мин). Наличие или отсутствие хроматина обнаруживали под инвертированным флуоресцентным микроскопом (фиг. 4). Эффективность и достоверность OHE исследовали в 9 идентичных повторах с использованием в общей сложности 414 ооцитов. После созревания in vitro в течение 42-43 ч осуществляли направленное разделение на две части в зрелых ооцитах, в которых после частичного расщепления проназой обнаруживали конус выталкивания и/или РВ. Результаты оценивали, как описано в предыдущем параграфе. Результаты представлены в табл. 5. Таблица 5 Эффективность ручной энуклеакции с помощью химических соединений (CAHE) и направленной ручной энуклеакции (ОНЕ): среднееАО (абсолютные отклонения). Различные верхние индексы обозначают различие (P0,05). Отсутствие различий между группами в отношении конусов выталкивания и/или прикрепленных полярных тел, что дает возможность для направленного разделения на две части, или в уровне лизиса, и показатель успешной энуклеакции на количество разделенных на две части ооцитов также было сходным. Тем не менее, общая эффективность способа, измеренная в соответствии с количеством цитопластов на общее количество ооцитов, была выше для группы CAHE по сравнению с группой ОНЕ. Сравнение развития in vitro эмбрионов, продуцированных путем CAHE, RHE и ТС. В 8 повторах в общей сложности 468 созревших in vitro ооцитов случайным образом распределяли и подвергали трем из описанных выше способов энуклеакции. Регистрировали уровни слияния между цитопластом и донорными фибробластами. Реконструированные эмбрионы активировали и выращивали,как описано ранее. Расщепление и доли бластоцист определяли, соответственно, на 2 и 7 день. Стереомикроскопические характеристики развившихся бластоцист сравнивали между группами.- 20017100 Таблица 6 Способность к развитию для эмбрионов, полученных в результате ручной энуклеакции с помощью химических соединений (CAHE), произвольной ручной энуклеакции (RHE) и традиционного клонирования при помощи микроманипулятора (ТС): среднееАО (абсолютные отклонения). Различные верхние индексы обозначают различие (P0,05). Уровни слияния после энуклеакции были сходными, соответственно, для CAHE, RHE и ТС. Второе слияние и активация приводила в результате к незначительным (1%) потерям в первых двух группах. Хотя ТС привела в результате к меньшим отношениям расщепления к количеству реконструированных эмбрионов по сравнению с другими двумя группами, это различие не представлено для отношений количества бластоцист к количеству реконструированных эмбрионов. Стереомикроскопические характеристики (размер; оцениваемая пропорция и очертания внутренней клеточной массы) не отличаются между группами. Количество клеток (576 по сравнению с 497 по сравнению с 536) в продуцированных бластоцистах в CAHE, RHE и ТС представлены в табл. 6, фиг. 5 и 6. Статистический анализ.AVEDEV осуществляли при помощи программного обеспечения Microsoft ХР Excel, a ANOVA осуществляли при помощи системы SAS. Вероятность P0,05 рассматривали как статистически значимую. Пример 5. Получение поросят. Ручное клонирование (НМС). Через 41 ч после начала созревания in vitro покров из кумулюса СОС удаляли путем повторного пипетирования в 1 мг/мл гиалуронидазы в ТСМ-199, забуференной Hepes. С этого момента (за исключением того, если не указано иное) все манипуляции осуществляли на стадии нагревания при 39C, и все капли, использованные для обработки ооцитов, имеющие объем 20 мкл, были покрыты минеральным маслом. Ооциты быстро инкубировали в 3,3 мг/мл проназы, растворенной в T33 (Т для среды ТСМ 199,забуференной Hepes; число указывает процент (об./об.) добавки телячьей сыворотки CS, в данном случае 33%) в течение 20 с и затем быстро промывали в каплях Т 2 и Т 20. Ооциты с частично расщепленной, но все еще видимой блестящей оболочкой располагали в каплях Т 2, дополненных 2,5 мкг/мл цитохалазмнаB. Путем применения тонковытянутой и оплавленной стеклянной пипетки ооциты поворачивали для того, чтобы найти полярное тельце на поверхности, и направленное разделение на две части осуществляли вручную под наблюдением в стереомикроскоп с использованием сверхострых разделяющих лезвий(АВ Technology, Pullman, WA, USA). Таким образом, менее чем половину цитоплазмы (рядом с выталкиванием или РВ) удаляли из оставшегося предполагаемого цитопласта. Цитопласты дважды промывали в каплях Т 2 и собирали в капле Т 10. Зародышевые фибробласты готовили, как описано ранее (Kragh, P.M. et al. Theriogenology 64, 15361545 (2005). Слияние осуществляли в две стадии, где вторая стадия также включала инициацию активации. На первой стадии использовали половину предполагаемых цитопластов. С использованием тонко вытянутой и оплавленной стеклянной пипетки 10 цитопластов переносили в виде группы в 1 мг/мл фитогемагглютинина (PHA; ICN Pharmaceuticals, Australia) в течение 3 с, затем каждый цитопласт быстро капали на один из нескольких фибробластов, которые осаждали в капле Т 2. После прикрепления 10 пар цитопластфибробласт уравновешивали в среде для слияния (0,3 М маннит, и 0,01% PVA) в течение 10 с. Путем использования переменного тока (АС) 0,6 кВ/см и 700 кГц пары клеток подгоняли к спирали камеры для слияния (камера для слияния ВТХ microslide 0,5 мм, модель 450; ВТХ, San Diego, CA), причем соматические клетки находились дальше всего от спирали, затем сливали с единичным импульсом постоянного тока (DC) 2,0 кВ/см в течение 9 мкс. После электрического импульса пары клеток затем осторожно удаляли со спирали, переносили в капли Т 10 и инкубировали для того, чтобы обнаружить, произошло ли слияние. Приблизительно через 1 ч после слияния слитые пары вместе с оставшимися цитопластами раздельно уравновешивали в активирующей среде (0,3 М маннит, 0,1 мМ MgSO4, 0,1 мМ CaCl2, дополненная 0,01% PVA). Путем воздействия переменного тока 0,6 кВ/см цитопластслитую пару последовательно- 21017100 подгоняли к спирали в группах по 10, причем слитые пары располагались дальше всего от спирали. Для второго слияния и инициации активации использовали единичный импульс постоянного тока 0,7 кВ/см в течение 80 мкс. Пары затем удаляли со спирали и осторожно переносили в капли Т 10 для проверки слияния. Реконструированные эмбрионы инкубировали в среде PZM-3, дополненной 5 мкг/мл СВ и 10 мкг/мл циклогексимида, в течение 4 ч при 38,5C в 5% CO2, 5% O2 и 90% N2 с максимальной влажностью, затем тщательно промывали перед выращиванием культуры. Традиционное клонирование (ТС). Микроманипуляцию осуществляли с использованием инвертированного микроскопа Diaphot 200(Nikon, Tokyo, Japan). После 42-44 ч созревания клетки кумулюса удаляли, как описано выше. Все манипуляции осуществляли на стадии нагревания при 39C. Единичную каплю раствора для микроманипуляции (NCSU-23, дополненная 4 мг/мл BSA и 7,5 мкг/мл СВ) объемом 50 мкл наносили в центральную область на крышку культуральной чашки диаметром 60 мм и покрывали минеральным маслом. Группы из 20-30 ооцитов и зародышевых фибробластов помещали в ту же самую каплю. После инкубации в течение 15-30 мин ооцит удерживали при помощи пипетки (внутренний диаметр 25-35 мкм, а внешний диаметр 80-100 мкм). После помещения в позицию 5-6 ч первое полярное тельце и прилегающую цитоплазму (приблизит. 10% от общего объема ооцита), предположительно содержащую метафазную пластину,аспирировали и удаляли при помощи конической инъекционной пипетки (внутренний диаметр 20 мкм). Зародышевый фибробласт затем инъецировали в пространство через ту же самую щель. После переноса ядер (NT) реконструированные комплексы переносили в капли среды, покрытые минеральным маслом,для восстановления в течение 1-1,5 ч до проведения слияния и активации. Реконструированные комплексы инкубировали в среде для слияния в течение 4 мин. Комплексы вручную совмещали с использованием тонковытянутого и оплавленного стеклянного капилляра с получением плоскости контакта параллельно электродам. Затем применяли единичный импульс постоянного тока 2,0 кВ/см длительностью 30 мкс. После выращивания культуры в каплях среды PZM-3, дополненной 7,5 мкг/мл СВ в течение 30-60 мин, результаты слияния проверяли под стереомикроскопом. Слитые комплексы подвергали действию второго импульса в активирующем растворе. После инкубации в течение 30 мин в Т 10 комплексы переносили в среду PZM-3 для оценки развития in vitro. Выращивание и перенос эмбрионов. Реконструированные эмбрионы выращивали в среде PZM-3 (Dobrinsky, J.T. et al. Biol Reprod 55,1069-1074 (1996), дополненной 4 мг/мл BSA. Эмбрионы без блестящей оболочки, продуцированные из НМС, выращивали в модифицированной системе WOW (Feltrin, C. Et al. Reprod Fertil Dev 18, 126 (2006). Две различные линии клеток (LW1-2 для НМС, LW2 для ТС) использовали в качестве клеток-доноров ядер для НМС и ТС для идентификации потомства, полученного двумя способами. LW1-2 и LW2 происходят соответственно из эмбрионов из гибридов (с Duroc) и чистых Датских ландрасов. Среднее отношение количества бластоцист на реконструированный эмбрион после выращивания invitro в течение 7 дней составляло 50,12,8% (среднееCO), что было значительно выше (p0,01) для НМС по сравнению с ТС, осуществленным параллельно в лаборатории авторов изобретения (табл. 7) и также было наибольшим, чем отношение, о котором сообщали для клонирования свиней. Таблица 7 Развитие in vitro эмбрионов, продуцированных в результате ручного и традиционного клонирования Значения различных верхних индексов в колонках значимо отличаются (p 0,05). Смешанные бластоцисты, продуцированные из НМС и ТС, хирургически переносили в 11 естественным образом синхронизированных свиней на 4 или 5 день цикла эструса. У шести (55%) реципиентов поставили диагноз беременность при помощи ультрасонографии, у 2 произошел аборт и ко времени написания 2 родили, соответственно, 3 и 10 поросят. Размер выводка 10 клонированных поросят в соответствии с нашими знаниями представляет собой наибольший размер выводка, достигнутый до настоящего времени при клонировании свиней. Все из них являются здоровыми и демонстрируют нормальное поведение за исключением одного, который демонстрирует жесткий изгиб дистального сустава одной из передних ног. Предварительные результаты свидетельствуют о том, что, когда переносили эмбрионы на аналогичных стадиях, реципиенты на 4 день цикла эструса поддерживали беременность лучше, чем реципиенты на 5 день (табл. 8). Микросателлитный анализ. Парентеральный анализ с использованием 10 различных свиных микросателлитных маркеров подтвердил идентичный генотип клонированных поросят и донорных клеток, использованных для переноса ядер. Идентификацию осуществляли при помощи микросателлитного анализа геномной ДНК каждого из новорожденных поросят, суррогатной свиньи и донорных фибробластов кожи LW1-2 и LW2, происходящих из двух эмбрионов, представляющих, соответственно, Датский ландрас и Duroc. Десять полиморфных микросателлитных локусов (SW886, SW58, SW2116, SW1989, SW152, SW378, KS139, SO167,SW1987, SW957), располагающихся в различных хромосомах свиньи, амплифицировали при помощи 3 цветной мультиплексной полимеразной цепной реакции (ПЦР) и продукты анализировали на анализаторе генов 3130 Х 1 (Applied Biosystems) с использованием программы Gene Mapper 3.7. Для второго реципиента отношение количества потомства к эмбриону (22%) было наибольшим из тех, которые до настоящего времени сообщались при клонировании свиней (Walker, S.C. et al. CloningStem Cells 7, 105-112 (2005); Hoshino, Y. etal. Cloning Stem Cells 7, 17-26 (2005. Сопоставимые эффективности живые родившиеся потомки/перенесенный эмбрион были получены с использованием НМС(17%) и ТС (15%). Статистический анализ. Различия между экспериментальными группами оценивали с использованием независимого t-теста с помощью SPSS 11.5. P0,05 рассматривали как статистически значимую. Пример 6. Один из примеров трансгена, который может быть использован для получения трансгенного млекопитающего, отличающегося от человека, в качестве модели заболевания врожднного буллезного эпидермолиза, представляет собой человеческий ген кератина 14, включающий мутацию, изображенную ниже жирным шрифтом. Последовательность трансгена, интегрированного в зародышевые фибробласты свиньи (донорная клетка) включает промотор человеческого гена кератина 14 и кДНК кератина 14, включающую стартовый и стоп-кодоны (жирный шрифт) и мутацию, вызывающую заболевание (жирный шрифт и подчеркивание), как описано Sorensen et al., J Invest Dermatol. 1999 Feb; 112(2): 184-90). Фрагмент клонируют вpN1-EGFP (Clontech), содержащем сигнал polyA для генной экспрессии и ген резистентности к неомицину для отбора клеточных клонов с интегрированным трансгеном. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ переноса клеточных ядер, включающий стадии: а) получения по меньшей мере одного ооцита, где по меньшей мере часть блестящей оболочки удаляют частично,б) разделения ооцита по меньшей мере на две части с получением ооцита, содержащего ядро, и по меньшей мере одного цитопласта,в) получения донорной клетки или клеточного ядра, обладающего желаемыми генетическими свойствами,г) слияния по меньшей мере одного цитопласта с донорной клеткой или клеточным ядром, окруженным мембраной,д) получения реконструированного эмбриона. 2. Способ по п.1, где ооцит разделяют по меньшей мере на три части с получением по меньшей мере двух цитопластов. 3. Способ по любому из пп.1, 2, где блестящую оболочку частично удаляют с помощью ферментов. 4. Способ по любому из пп.1-3, где ооцит разделяют по меньшей мере на три части с получением по меньшей мере двух цитопластов. 5. Способ по любому из пп.1-4, где желаемые генетические свойства донорной клетки или клеточного ядра получают путем модификации желаемого гена или генов с помощью мутации, делеции и/или инсерции. 6. Способ по любому из пп.1-5, где способ слияния выбирают из химического слияния, слияния с использованием электрического тока и биослияния. 7. Способ по любому из пп.1-6, где слияние осуществляют по меньшей мере в одну стадию. 8. Способ по любому из пп.1-7, где слияние осуществляют по меньшей мере в две стадии. 9. Способ по п.8, где первую стадию слияния осуществляют по меньшей мере между одним цитопластом и донорной клеткой или клеточным ядром, окруженным мембраной. 10. Способ по п.9, где вторую стадию слияния осуществляют по меньшей мере между одной слитой парой и по меньшей мере одним цитопластом. 11. Способ по любому из пп.1-10, где донорная клетка представляет собой соматическую клетку. 12. Способ по п.11, где соматическую клетку выбирают из группы, состоящей из эпителиальных клеток, нервных клеток, эпидермальных клеток, кератиноцитов, гематопоэтических клеток, меланоцитов, хондроцитов, лимфоцитов (B и T лимфоцитов), эритроцитов, макрофагов, моноцитов, мононуклеарных клеток, фибробластов, клеток сердечной мышцы и других мышечных клеток. 13. Способ по п.12, где соматические клетки получают из группы, состоящей из клеток кожи, клеток легких, клеток поджелудочной железы, клеток печени, клеток желудка, интестинальных клеток, сердечных клеток, клеток репродуктивных органов, клеток мочевого пузыря, клеток почки, клеток мочеиспускательного канала и клеток других органов мочевой системы. 14. Способ по любому из пп.1-13, где соматическая клетка представляет собой клетку фибробласта.- 25017100 15. Способ по п.14, где соматическая клетка представляет собой клетку фибробласта, полученную от млекопитающего. 16. Способ по п.15, где млекопитающее представляет собой свинью. 17. Способ по пп.1 и/или 2, где донорная клетка имеет происхождение из клетки зародышевой линии. 18. Способ по любому из пп.1-17, где ооцит получают от свиньи. 19. Способ по п.1, дополнительно включающий стадии: а) активации митоза в реконструированном эмбрионе с образованием эмбриона,б) выращивания указанного эмбриона. 20. Способ по п.19, где эмбрион последовательно выращивают в различных средах. 21. Способ получения генетически модифицированного или трансгенного млекопитающего, отличного от человека, включающий осуществление способа по любому из пп.1-19 и последующие стадии: а) активации митоза в реконструированном эмбрионе с образованием эмбриона; б) выращивания указанного эмбриона и в) переноса указанного выращенного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш. 22. Способ по п.21, где способ активации реконструированного эмбриона выбирают из группы, состоящей из электрического импульса, химически индуцированного шока, увеличения внутриклеточных концентраций двухвалентных катионов и уменьшения фосфорилирования. 23. Способ по п.21, дополнительно включающий вторую стадию слияния между по меньшей мере одной слитой парой цитопласта и донорной клетки и по меньшей мере одним цитопластом, где указанную вторую стадию слияния осуществляют последовательно или одновременно. 24. Способ по п.21, где эмбрион выращивают in vitro. 25. Способ по п.24, где эмбрион последовательно выращивают в различных средах. 26. Способ по п.21, где эмбрион подвергают криоконсервации перед переносом в млекопитающеехозяин. 27. Способ по п.26, где эмбрион находится на стадии бластоцисты. 28. Способ по п.26, где по меньшей мере один ооцит, полученный на стадии (а) п.1, подвергают делипидизации путем мягкой обработки ферментативным агентом перед криоконсервацией эмбриона. 29. Способ по п.28, где делипидизированный ооцит разделяют по меньшей мере на две части с получением ооцита, содержащего ядро, и по меньшей мере одного цитопласта. 30. Способ по п.26, где указанный эмбрион выращивают до стадии бластоцисты перед криоконсервацией. 31. Способ клонирования млекопитающего, отличного от человека, включающий: а) получение эмбриона в соответствии с любым из пп.1-18,б) перенос указанного эмбриона в млекопитающее-хозяин таким образом, что эмбрион развивается в генетически модифицированный зародыш. 32. Генетически модифицированное млекопитающее, отличное от человека, получаемое при помощи способа по любому из пп.21-31, содержащее в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из двух различных материнских источников. 33. Генетически модифицированный эмбрион, отличный от человеческого, получаемый при помощи способа по любому из пп.21-31, содержащий в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из двух различных материнских источников. 34. Генетически модифицированное млекопитающее, отличное от человека, по п.32, содержащее в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из трех различных материнских источников. 35. Генетически модифицированный эмбрион, отличный от человеческого, по п.33, содержащий в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из трех различных материнских источников. 36. Генетически модифицированное млекопитающее, отличное от человека, по п.32, содержащее в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из четырех различных материнских источников. 37. Генетически модифицированный эмбрион, отличный от человеческого, по п.33, содержащий в клетках своих тканей митохондрии, имеющие происхождение по меньшей мере из четырех различных материнских источников. 38. Генетически модифицированное млекопитающее, отличное от человека, по п.32, где млекопитающее представляет собой свинью. 39. Генетически модифицированный эмбрион, отличный от человеческого, по п.33, где эмбрион имеет происхождение от свиньи.
МПК / Метки
МПК: C12N 15/85, C12N 15/87
Метки: модифицированный, ядер, получения, млекопитающего, клонирования, переноса, модифицированное, способ, эмбрион, клеточных, генетически, модифицированного, трансгенного, млекопитающее
Код ссылки
<a href="https://eas.patents.su/30-17100-sposob-perenosa-kletochnyh-yader-sposob-polucheniya-geneticheski-modificirovannogo-ili-transgennogo-mlekopitayushhego-sposob-klonirovaniya-mlekopitayushhego-geneticheski-modificiro.html" rel="bookmark" title="База патентов Евразийского Союза">Способ переноса клеточных ядер, способ получения генетически модифицированного или трансгенного млекопитающего, способ клонирования млекопитающего, генетически модифицированное млекопитающее и генетически модифицированный эмбрион</a>
Способ получения модифицированного морденита аммония

Номер патента: 247
Опубликовано: 25.02.1999
Авторы: Пасселек Жан, Ван Жизель Огюст
МПК: C07C 209/16, B01J 29/18
Метки: модифицированного, аммония, морденита, получения, способ
Формула / Реферат:
1. Способ получения модифицированного морденита аммония, обладающего каталитической активностью, характеризующийся тем, что включает (1) сушку морденита аммония при условиях, обеспечивающих сохранение аммонийной формы морденита, и (2) обработку высушенного таким образом морденита аммония, тетрахлорсиланом в газовой фазе при температуре между 300 и 600°С.2. Способ по п.1, отличающийся тем, что стадию сушки проводят при температуре ниже 400°С,...
Производные аминогуанидина и алкоксигуанидина (варианты) и способ их получения (варианты), фармацевтическая композиция и способ ингибирования протеолиза у млекопитающего, способ лечения различных заболеваний млекопитающего, способы ингибирования индуцированной тромбином агрегации тромбоцитов и свертывания фибриногена в плазме, тромбина, агрегации тромбоцитов и образования тромбов в крови, устройство для сбора крови, искусственного кровообращения и хранения крови

Номер патента: 3461
Опубликовано: 26.06.2003
Авторы: Маркоутэн Томас П., Солл Ричард М., Томкзук Брюс Э., Лу Тианбао, Сайедем Коллин
МПК: A61K 31/445, A61P 7/02, C07C 279/00...
Метки: фармацевтическая, способы, различных, тромбов, тромбином, лечения, индуцированной, композиция, кровообращения, алкоксигуанидина, искусственного, плазме, протеолиза, тромбина, заболеваний, образования, сбора, ингибирования, производные, варианты, млекопитающего, способ, хранения, крови, получения, аминогуанидина, тромбоцитов, свертывания, фибриногена, агрегации, устройство
Формула / Реферат:
1. Соединение формулы I или его сольват, гидрат или фармацевтически приемлемая соль, где L - означает -C(O)- или -SO2-; R1 означает группу R2 означает группу или R1 и R2 могут быть объединены с атомом азота, к которому они присоединены, с образованием цикла, содержащего от трех до семи атомов, который необязательно содержит дополнительный атом азота или кислорода, и который по выбору является бензо- или пиридоконденсированным циклом, причем...
Синергетические комбинации биологически активных веществ, их применение, способ подавления вредных фитопатогенных грибов, способ получения фунгицидных средств, способы протравливания (трансгенного) посевного материала

Номер патента: 14410
Опубликовано: 30.12.2010
Авторы: Хадано Хироюки, Вахендорф-Нойманн Ульрике, Доллингер Маркус, Лэроу Бернар Марк, Савада Харуко, Штенцель Клаус, Шерб Кристиян, Дамен Петер, Гросжан Мари-Клэр, Гуо Жан-Мари, Хойзер-Хан Изольдэ
МПК: A01P 3/00, A01N 57/20, A01N 43/653...
Метки: трансгенного, применение, синергетические, подавления, активных, комбинации, грибов, способ, способы, фитопатогенных, средств, материала, протравливания, биологически, веществ, фунгицидных, получения, посевного, вредных
Формула / Реферат:
1. Синергетические комбинации активных веществ, содержащихгруппа (1) гербицид, выбранный из(1-2) глуфосината,(1-3) глуфосината аммония,и по крайней мере одно активное вещество, выбранное из группы (2):группа (2) стробилурины, выбранные из(2-1) азоксистробина,(2-2) флуоксастробина,(2-3) (2Е)-2-(2-{[6-(3-хлор-2-метилфенокси)-5-фтор-4-пиримидинил]окси}фенил)-2-(метоксиимино)-N-метилэтанамида,2-4) трифлоксистробина,(2-5)...
Производные гуанидина (варианты), способ их получения (варианты), фармацевтическая композиция для ингибирования протеолиза у млекопитающего, способ лечения, способ ингибирования: протеолиза у млекопитающего, тромбин-индуцированной агрегации тромбоцитов и образования сгустков фибриногена в плазме, тромбина в крови, образования аггрегатов тромбоцитов в крови, образования тромбов в крови, устройстводля сбора, циркуляции и хранения крови

Номер патента: 2740
Опубликовано: 29.08.2002
Авторы: Федде Синтия Л., Стэгнейро Томас П., Томкзук Брюс Э., Солл Ричард М., Маркоутэн Томас П., Иллиг Карл Р., Лу Тианбао
МПК: A61K 31/44, A61K 31/255, A61K 31/155...
Метки: ингибирования, способ, лечения, фибриногена, циркуляции, сбора, млекопитающего, протеолиза, сгустков, тромбов, устройстводля, аггрегатов, тромбина, фармацевтическая, крови, гуанидина, хранения, получения, композиция, производные, варианты, тромбин-индуцированной, образования, агрегации, тромбоцитов, плазме
Формула / Реферат:
1. Соединение формулы I или его сольват, гидрат, фармацевтически приемлемая соль или пролекарство, где R1 является фенильной, нафтильной, дигидродиоксобензо[b]тиофенильной или гетероарильной группой, каждая из которых может быть необязательно замещена, по крайней мере, одним заместителем, независимо выбранным из группы, состоящей из С1-6алкила, гидрокси, нитро, трифторметила, трифторметокси, галогена, С1-6алкокси, циано, амино, гидроксиамино,...
Рекомбинантная двухцепочная последовательность днк для экспрессии сахарозофосфорилазы в растении, способ получения трансгенного растения с повышенной экспрессией сахарозофосфорилазы в различных частяхрастения и линия трансформированных клеток растения

Номер патента: 634
Опубликовано: 29.12.1999
Авторы: Девирд Ян Виллем, Велдон Марсиа Ли, Бэрри Джерард Фрэнсис, Кишор Джениш Мерти
МПК: C12N 15/82
Метки: растении, клеток, двухцепочная, получения, линия, трансформированных, последовательность, способ, экспрессии, частяхрастения, растения, различных, повышенной, экспрессией, сахарозофосфорилазы, днк, трансгенного, рекомбинантная
Формула / Реферат:
1. Рекомбинантная двухцепочечная последовательность ДНК для экспрессии сахарозофосфорилазы в растении, содержащая следующие конструктивные элементы: - промотор, активный в определенных клетках растений; - ген, кодирующий фермент сахарозофосфорилазу Streptococcus mutans, имеющий следующую последовательность нуклеотидов: (SEQ ID NO:5); и 3` - нетранслируемую область, обеспечивающую терминацию транскрипции гена сахарозофосфорилазы и...
Предыдущий патент: Узел клинового коуша
Следующий патент: Фильтрующая секция и фильтрующее устройство с такой секцией
Случайный патент: Огнестойкая полимерная композиция, содержащая нанонаполнители