Способ идентификации и получения (r)-специфической w-трансаминазы
Номер патента: 22764
Опубликовано: 29.02.2016
Авторы: Борншойер Уве, Робинс Карен, Хене Маттиас, Шетцле Себастьян
















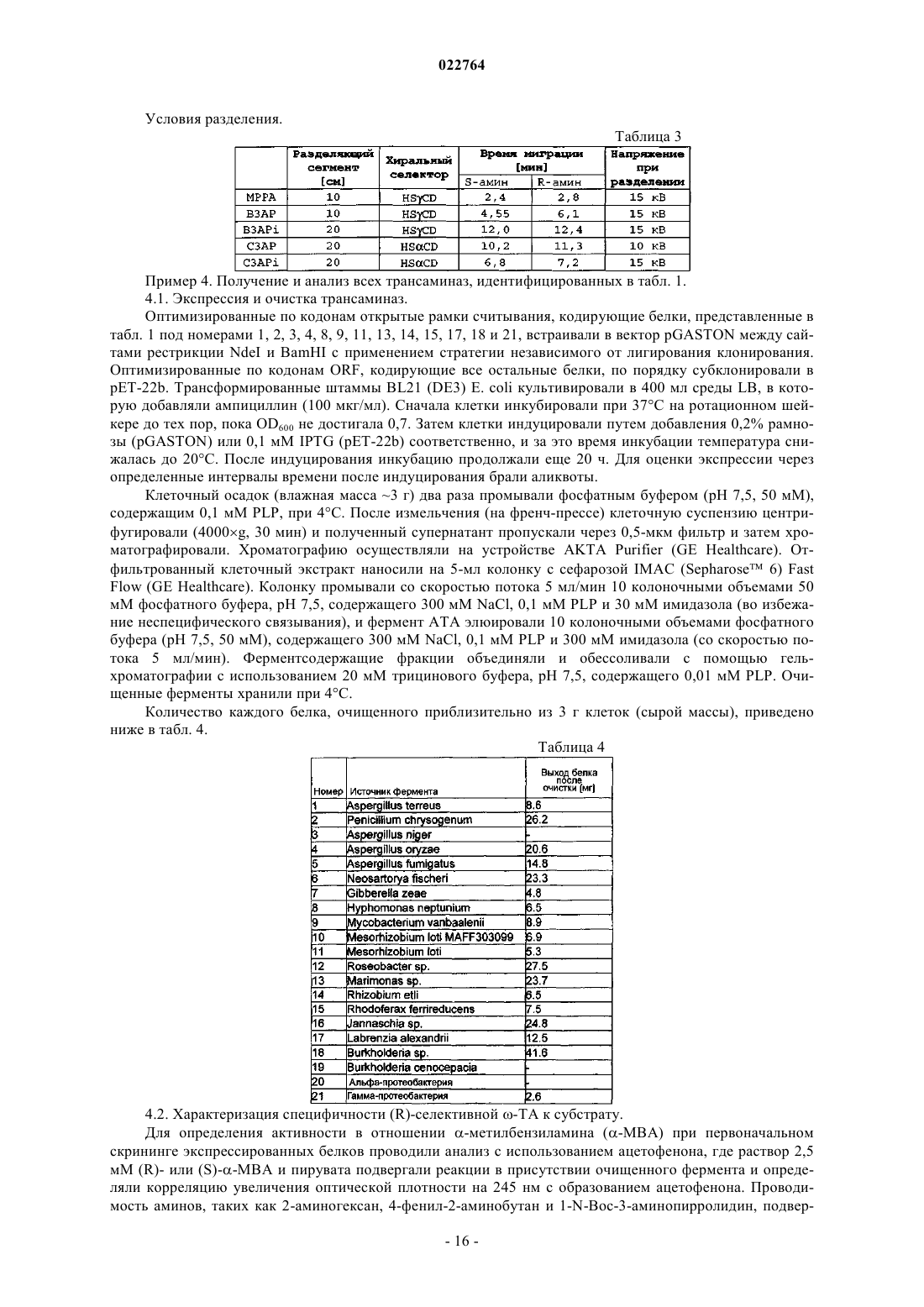

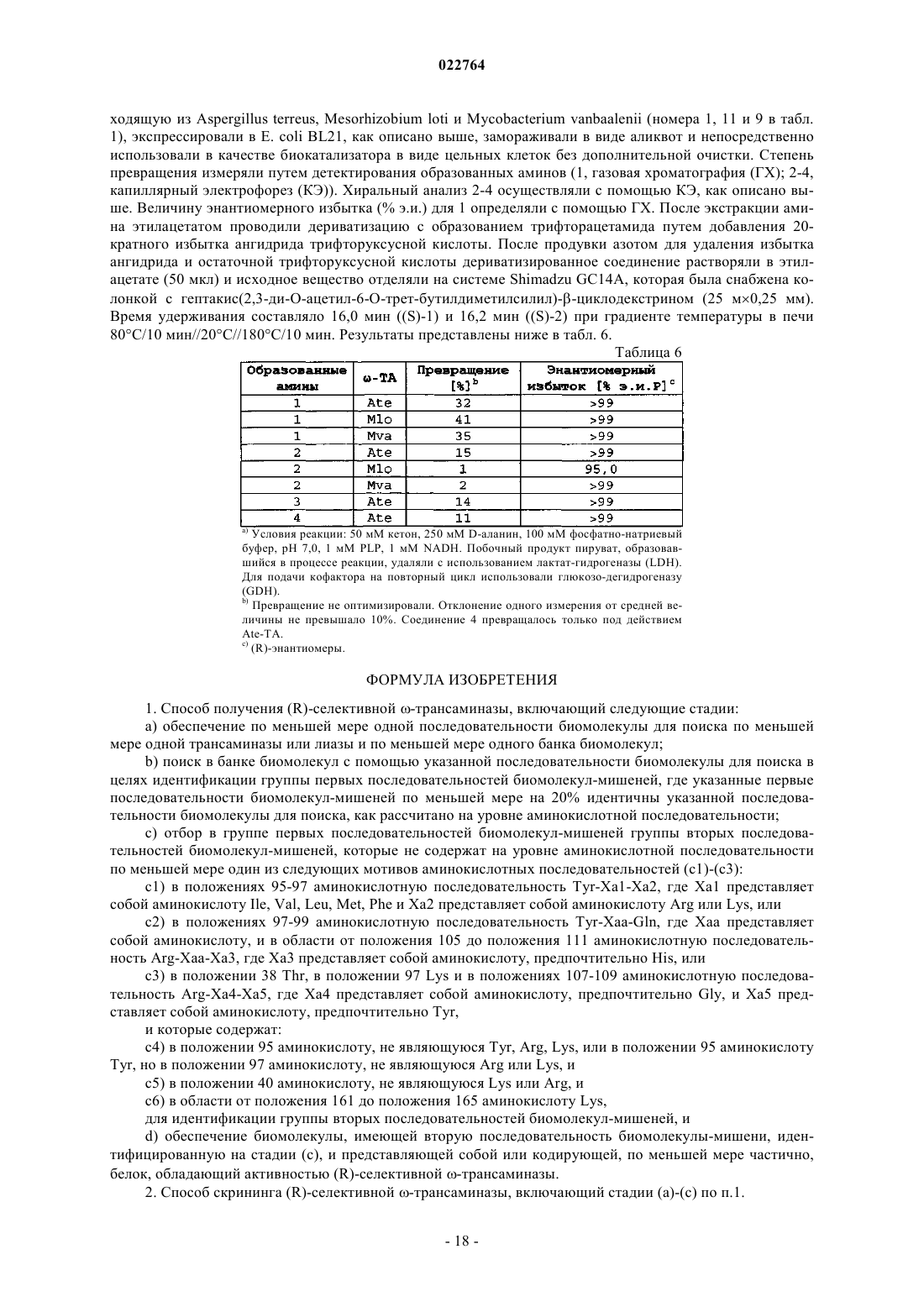

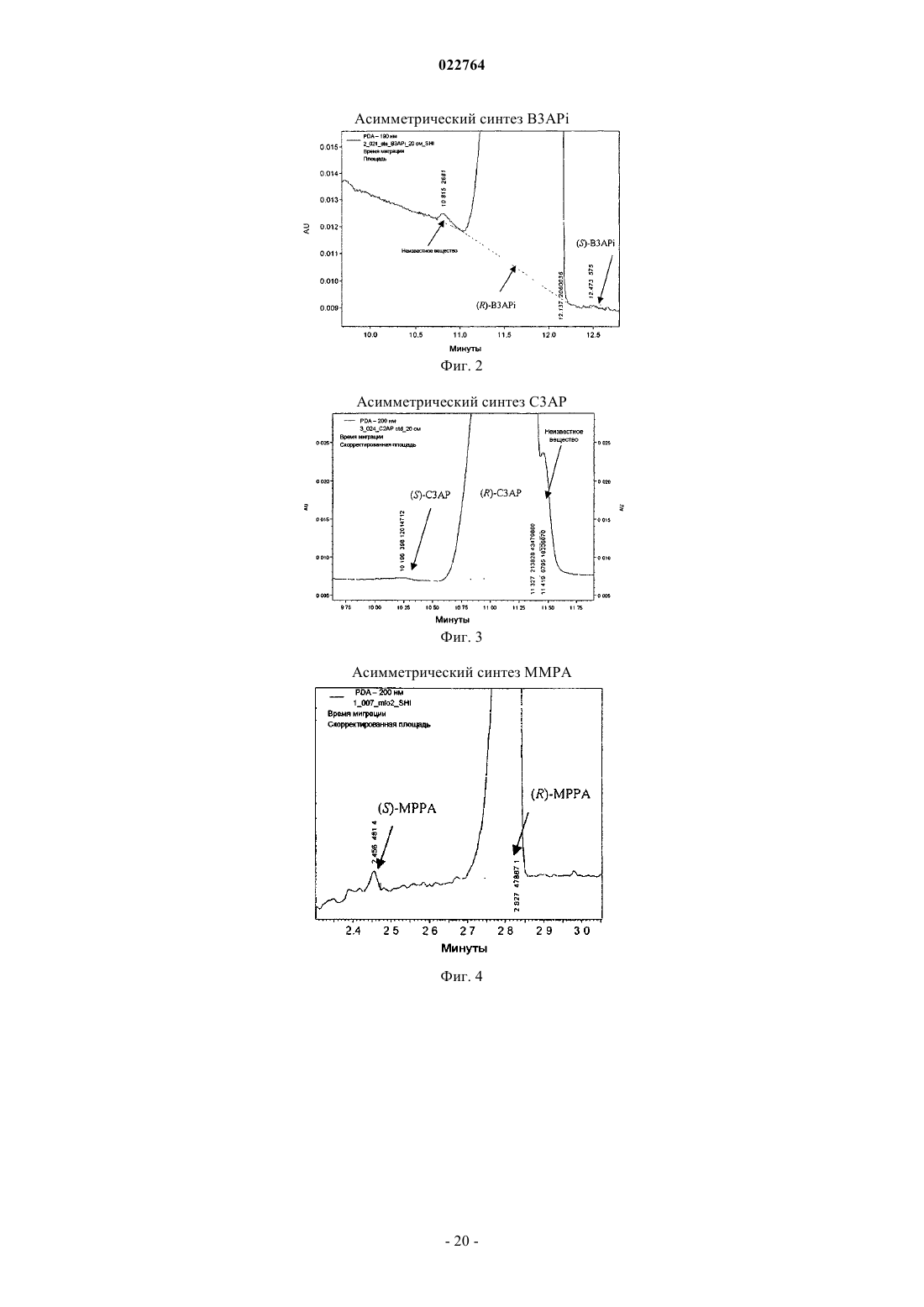
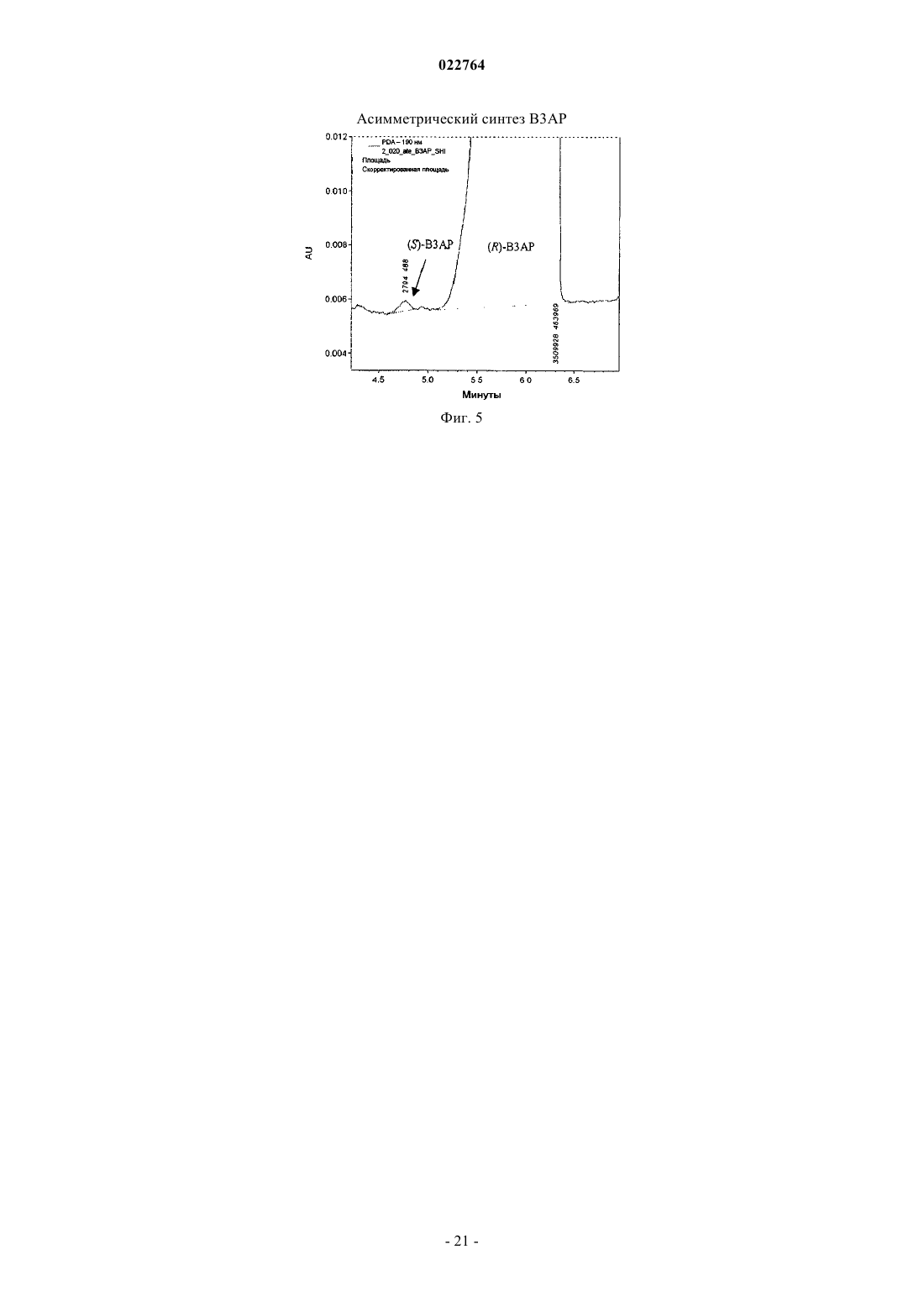








Формула / Реферат
1. Способ получения (R)-селективной w-трансаминазы, включающий следующие стадии:
a) обеспечение по меньшей мере одной последовательности биомолекулы для поиска по меньшей мере одной трансаминазы или лиазы и по меньшей мере одного банка биомолекул;
b) поиск в банке биомолекул с помощью указанной последовательности биомолекулы для поиска в целях идентификации группы первых последовательностей биомолекул-мишеней, где указанные первые последовательности биомолекул-мишеней по меньшей мере на 20% идентичны указанной последовательности биомолекулы для поиска, как рассчитано на уровне аминокислотной последовательности;
c) отбор в группе первых последовательностей биомолекул-мишеней группы вторых последовательностей биомолекул-мишеней, которые не содержат на уровне аминокислотной последовательности по меньшей мере один из следующих мотивов аминокислотных последовательностей (c1)-(c3):
c1) в положениях 95-97 аминокислотную последовательность Tyr-Xa1-Xa2, где Xa1 представляет собой аминокислоту Ile, Val, Leu, Met, Phe и Ха2 представляет собой аминокислоту Arg или Lys, или
с2) в положениях 97-99 аминокислотную последовательность Tyr-Xaa-Gln, где Хаа представляет собой аминокислоту, и в области от положения 105 до положения 111 аминокислотную последовательность Arg-Xaa-Ха3, где Ха3 представляет собой аминокислоту, предпочтительно His, или
с3) в положении 38 Thr, в положении 97 Lys и в положениях 107-109 аминокислотную последовательность Arg-Xa4-Xa5, где Ха4 представляет собой аминокислоту, предпочтительно Gly, и Ха5 представляет собой аминокислоту, предпочтительно Tyr,
и которые содержат:
с4) в положении 95 аминокислоту, не являющуюся Tyr, Arg, Lys, или в положении 95 аминокислоту Tyr, но в положении 97 аминокислоту, не являющуюся Arg или Lys, и
с5) в положении 40 аминокислоту, не являющуюся Lys или Arg, и
с6) в области от положения 161 до положения 165 аминокислоту Lys,
для идентификации группы вторых последовательностей биомолекул-мишеней, и
d) обеспечение биомолекулы, имеющей вторую последовательность биомолекулы-мишени, идентифицированную на стадии (с), и представляющей собой или кодирующей, по меньшей мере частично, белок, обладающий активностью (R)-селективной w-трансаминазы.
2. Способ скрининга (R)-селективной w-трансаминазы, включающий стадии (а)-(с) по п.1.
3. Способ по п.1 или 2, где указанная биомолекула представляет собой белок и последовательность биомолекулы представляет собой аминокислотную последовательность.
4. Способ по п.1 или 2, где биомолекула представляет собой молекулу ДНК и последовательность биомолекулы представляет собой последовательность ДНК.
5. Способ по любому из предшествующих пунктов, где банк биомолекул представляет собой базу данных биомолекул и поиск в указанной базе данных биомолекул на стадии (b) осуществляют с помощью программы выравнивания последовательностей биомолекул, в частности BLAST.
6. Способ по любому из пп.1, 3-5, где молекулу белка или ДНК, обеспеченную на стадии (d), получают синтезом de novo.
7. Способ по любому из пп.1, 2 и 4, где банк биомолекул представляет собой банк генов и поиск в указанном банке генов на стадии (b) осуществляют с использованием молекулы с последовательностью ДНК для поиска.
8. Способ по п.7, где на стадии (с) используют праймеры с ДНК-последовательностью для отбора группы вторых последовательностей биомолекул.
9. Способ по любому из предшествующих пунктов, где указанная последовательность биомолекулы для поиска представляет собой последовательность, представляющую всю функциональную (R)-селективную w-трансаминазу или ее часть.
10. (R)-селективная w-трансаминаза, которая может быть получена по любому из способов по пп.1 и 3-9, предпочтительно выбранная из группы последовательностей биомолекул, состоящей из SEQ ID NO: 1-4.
11. Способ определения характеристик трансаминазы, в частности для характеризации свойств трансаминазы, включающий следующие стадии:
i) обеспечение заряженного аминоакцептора, заряженного аминодонора и трансаминазы;
ii) осуществление реакции аминоакцептора и аминодонора с трансаминазой в реакционной среде и тем самым
iii) определение проводимости реакционной среды в первом наборе реакционных условий;
iv) после проведения стадии iii) определение проводимости реакционной среды во втором наборе реакционных условий с получением по меньшей мере двух величин проводимости, отражающих свойства трансаминазы.
12. Способ по п.11, где проводимость реакционной среды определяют по меньшей мере три раза, по меньшей мере, в третьем наборе реакционных условий.
13. Способ по п.11 или 12, где наборы реакционных условий являются одинаковыми.
14. Способ по любому из пп.11-13, где реакционная среда представляет собой буфер с низкой проводимостью.
15. Способ по любому из пп.11 или 14, где заряженный аминоакцептор представляет собой пируват.
16. Способ получения (R)-селективной w-трансаминазы по любому из пп.1 и 3-9, где после получения (R)-селективной w-трансаминазы осуществляют способ определения характеристик указанной (R)-селективной w-трансаминазы по любому из пп.11-15.
17. Способ скрининга (R)-селективной w-трансаминазы по любому из пп.2-9, где после скрининга (R)-селективной w-трансаминазы осуществляют способ определения характеристик указанной (R)-селективной w-трансаминазы по любому из пп.11-15.
18. Способ получения оптически активного хирального амина, включающий осуществление реакции по меньшей мере одного аминоакцептора и по меньшей мере одного аминодонора с (R)-селективной w-трансаминазой по п.10 и получение оптически активного хирального амина.
Текст
-трансаминаз, к полученным таким образом трансаминазам и к их применению в различных способах переаминирования. Настоящее изобретение относится к способам скрининга, характеризации и получения (R)специфических -трансаминаз, к полученным таким образом -трансаминазам и к их применению в различных способах переаминирования. Хиральные амины играют важную роль в фармацевтической, агрохимической и химической промышленности. Они часто используются в качестве промежуточных соединений или синтонов для получения различных физиологически, например, фармацевтически активных веществ, таких как производные цефалоспоринов или пирролидинов. Из большого числа хиральных аминов, применяемых в различных целях, физиологически активной является фактически только одна оптически активная форма, такая как (R)- или (S)-энантиомер. Поэтому существует необходимость в разработке способов получения хиральных аминов в оптически активной форме. Эти потребности могут быть частично удовлетворены благодаря получению хиральных аминов путем кристаллизации диастереомерных солей посредством добавления хиральных карбоновых кислот(Breuer et al., Angewandte Chemie (2004) 116, 806-843). В других химических методах используется энантиоселективный синтез, осуществляемый путем восстановления прохиральных предшественников с двойными C=N-связями. В последнее время из нескольких ферментативных методов, применяемых для синтеза оптически активных аминокислот и аминов, особое внимание привлекают методы с использованием сотрансаминаз (-ТА) благодаря их способности осуществлять асимметрический синтез оптически активных аминов, которые часто используются в качестве структурных блоков для получения множества фармацевтических средств.-Трансаминазы представляют собой PLP (перидоксалфосфат)-зависимые ферменты, которые катализируют реакции переноса аминогрупп. При использовании -трансаминаз, так называемые энантиообогащенные и/или чистые оптически активные амины могут быть, главным образом, продуцированы посредством реакции с применением двух стратегий: (i) асимметрического синтеза исходных из кетонов,и (ii) кинетического разделения исходных из рацемических аминов. Хотя -трансаминазы в основном обладают хорошей энантиоселективностью, однако обычно они не используются для асимметрического синтеза, хотя теоретически в данном случае можно ожидать 100% выхода. Одним из конкретных требований при осуществлении асимметрического синтеза с использованием -трансаминаз является сдвиг равновесия в сторону продукта, особенно при использовании аминокислоты, подобной аланину, в качестве аминодонора, поскольку в данном случае равновесие смещается в сторону субстратов (кетона, аланина), а не в сторону продуктов (амина, пирувата); другим требованием является достижение стереоселективности фермента, что не всегда происходит в случае использования -трансаминаз. Поэтому сотрансаминазы используются, главным образом, для кинетического разделения рацемических аминов, при котором достижение энантиоселективности не является необходимым условием. Таким образом, хотя были проведены исследования по кинетическому разделению рацемических аминов, ограничение выхода, который составляет максимум 50%, существенно мешает их применению. С другой стороны, для асимметрического синтеза требуются методы, позволяющие изменять нежелательный сдвиг равновесия в сторону синтеза отдельных энантиомеров оптически чистых аминов, для которого было разработано несколько методов, что и явилось одной из ключевых предпосылок создания эффективных способов, позволяющих использовать трансаминазы в промышленном масштабе. Такие методы описаны в заявке WO 2007/093372. В ЕР 0987332 А 1 описан способ продуцирования оптически активных аминосоединений, а именно(R)-аминосоединений, с использованием микробного фермента, в частности (R)-селективной -трансаминазы, выделенной из Arthrobacter sp. В ЕР 1038953 А 1 описана другая (R)-селективная -трансаминаза, которая, однако, была выделена из Mycobacterium aurum.Iwasaki at al. (Biotechnol. Let., 2003 (25), 1843-1846), Koszelewski at al. (Adv. Synth. Catal., 2008 (350),2761-2766) и Iwasaki at al. (Appl. Microbiol. Biotechnol., 2006 (69), 499-505) раскрывают R-специфическую трансаминазу из Arthrobacter sp. Идентификацию микроорганизмов, обеспечивающих пригодные (S)или (R)-селективные со-трансаминазы, обычно осуществляют путем отбора микроорганизмов, выделенных, например, из образцов почвы, и обогащения ими культуры (Jun et al., Арр. Microbiol. Biotechnol.,2004 (70), 2529-2534 и Shin et al. (Biosc. Biotechnol. Biochem. 2001 (65), 1782-1788. В частности, все описанные R-селективные -трансаминазы были получены путем обогащения культуры. Такие методы требуют много времени, поскольку для их эффективного осуществления часто предпочтительна сверхэкспрессия фермента в гетерологичном организме, подобного Escherichia coli. Для этого требуется выделение генной последовательности из организмов дикого типа, но клонирование ДНК не всегда является успешным и представляет собой очень длительный процесс. В частности, в ходе процесса разработки или идентификации нового фермента -ТА очистка фермента и характеризация его ферментативных свойств представляет особый интерес. Однако, поскольку активность -ТА обычно определяют лабораторными методами, такими как ВЭЖХ или капиллярный электрофорез (СЕ), то определение свойств фермента чаще носит ограничительный характер. Как правило, по сравнению с (S)-селективными -трансаминазами число доступных (R)-1 022764 селективных -трансаминаз еще более ограничено. Это находится в резком противоречии с высокой потребностью в (R)-селективных -трансаминазах, которые являются наиболее пригодными для асимметрического синтеза (R)-энантиомеров различных хиральных аминов. Таким образом, существует потребность в обеспечении дополнительными (R)-селективными -трансаминазами и средствами и способами их получения в целях продуцирования R-энантиомеров, в частности оптически активных аминов, например, для фармацевтического или агрохимического применений, что в настоящее время является либо недоступным, либо считается экономически невыгодным процессом, особенно в промышленном масштабе. По этой причине настоящее изобретение, основанное на данной технической проблеме, обеспечивает простые, быстрые и надежные средства и способы идентификации, характеризации и получения (R)селективных -трансаминаз, в частности продуцирования желаемых (R)-энантиомеров, предпочтительно в оптически чистой форме, которые являются предпочтительно пригодными для идентификации, характеризации и получения в промышленном масштабе. Настоящее изобретение решает эти технические проблемы путем предоставления идеи, сформулированной в независимых пунктах формулы изобретения, в частности в пунктах формулы изобретения,относящихся к способам, продуктам и их применению. Таким образом, настоящее изобретение предоставляет конкретный вариант осуществления способа получения (R)-селективных со-трансаминаз (также называемых -ТА), включающего следующие стадии:a) обеспечение по меньшей мере одной запрашиваемой последовательности биомолекулы, по меньшей мере одной трансаминазы или лиазы, предпочтительно трансаминазы или лиазы, которые принадлежат к PLP-зависимым ферментам укладки типа IV, и по меньшей мере одного банка биомолекул;b) поиск в банке биомолекул с помощью запрашиваемой последовательности биомолекулы в целях идентификации группы первых последовательностей биомолекул-мишеней, где указанные первые последовательности биомолекул-мишеней по меньшей мере на 20%, предпочтительно на 32% идентичны запрашиваемой последовательности биомолекул, как вычислено на уровне аминокислотной последовательности;c) отбор в группе первых последовательностей биомолекул-мишеней группы вторых последовательностей биомолекул-мишеней, которые не содержат на уровне аминокислотной последовательности по меньшей мере один из следующих мотивов аминокислотных последовательностей (c1)-(с 3):c1) в положениях 95-97 аминокислотную последовательность Tyr-Xa1-Xa2, где Xa1 представляет собой аминокислоту Ile, Val, Leu, Met, Phe и Ха 2 представляет собой аминокислоту Arg или Lys, или с 2) в положениях 97-99 аминокислотную последовательность Tyr-Xaa-Gin, где Хаа представляет собой предпочтительно обычную аминокислоту, и в области от положения 105 до положения 111, предпочтительно в положениях 107-109 аминокислотную последовательность Arg-Xaa-Ха 3, где Ха 3 представляет собой аминокислоту, предпочтительно His, или с 3) в положении 38 Thr, в положении 97 Lys и в положениях 107-109 аминокислотную последовательность Arg-Xa4-Xa5, где Ха 4 представляет собой аминокислоту, предпочтительно Gly, и Ха 5 представляет собой аминокислоту, предпочтительно Tyr,и которые содержат: с 4) в положении 95 аминокислоту, не являющуюся Tyr, Arg, Lys, или в положении 95 аминокислотуTyr, но в положении 97 аминокислоту, не являющуюся Arg или Lys, и с 5) в положении 40 аминокислоту, не являющуюся Lys или Arg, и с 6) в области от положения 161 до положения 165, предпочтительно в положении 163, аминокислоту Lys, для идентификации группы вторых последовательностей биомолекул-мишеней, иd) обеспечение, предпочтительно получение, биомолекулы, имеющей вторую последовательность биомолекул-мишеней, идентифицированную на стадии (с), и представляющей собой или кодирующей,по меньшей мере частично, белок, обладающий активностью (R)-селективной -трансаминазы. Термин "биомолекула, представляющая собой или кодирующая, по меньшей мере частично, белок",предпочтительно означает, что указанная биомолекула может или представлять собой белок, обладающий активностью (R)-селективной -трансаминазы, или, в случае, когда данная биомолекула представляет собой нуклеотидную последовательность, по меньшей мере часть указанной нуклеотидной последовательности кодирует указанный белок, предпочтительно полноразмерный белок. Соответственно в случае, когда обеспеченная биомолекула представляет собой молекулу нуклеотидной последовательности,по меньшей мере часть указанной молекулы нуклеотидной последовательности кодирует белок, обладающий активностью (R)-селективной -трансаминазы, благодаря чему другая часть указанной нуклеотидной молекулы может иметь некоторые другие функции, например регуляторную или репликативную функцию. Таким образом, термин "обеспечение биомолекулы, имеющей вторую последовательность биомолекулы-мишени, идентифицированную на стадии (с) и представляющую собой или кодирующую,по меньшей мере частично, белок, обладающий активностью (R)-селективной -трансаминазы", предпочтительно эквивалентен термину "получение биомолекулы, имеющей вторую последовательность биомолекулы-мишени, идентифицированную на стадии (с), которая может представлять собой белок, обла-2 022764 дающий активностью (R)-селективной со-трансаминазы, или которая может представлять собой молекулу нуклеотидной последовательности, которая включает последовательность, кодирующую указанный белок, при условии, что такой белок обладает активностью (R)-селективной -трансаминазы". В одном из вариантов осуществления настоящего изобретения на стадии (с 2) положение мотиваArg-Xaa-Ха 3 может смещаться на 1-2 аминокислоты от положений 107-109, то есть эти аминокислоты могут присутствовать в положениях 105-111. В одном из вариантов осуществления настоящего изобретения на стадии (с 6) Lys в положении 163 может смещаться на 1-2 аминокислоты, то есть эта аминокислота может присутствовать в положениях 161-165. В контексте настоящего описания каждая аминокислотная последовательность, в частности мотив аминокислотной последовательности или последовательность ДНК, или данный мотив последовательности ДНК всегда записываются в направлении от аминоконца до карбоксиконца или в направлении 5'3',если это не оговорено особо. Предпочтительно последовательности, представленные в данном направлении, всегда указываются как непрерывные последовательности нуклеотидов, без интронов, или непрерывные последовательности аминокислот, в зависимости от обстоятельств. В предпочтительном варианте осуществления настоящего изобретения по меньшей мере одной запрашиваемой последовательностью биомолекул, по меньшей мере одной трансаминазы, используемой на стадии (а), предпочтительно является (R)-селективная -трансаминаза. В другом предпочтительном варианте осуществления изобретения по меньшей мере одну запрашиваемую последовательность биомолекулы по меньшей мере одной трансаминазы, используемой на стадии (а), выбирают из группы, состоящей из (R)-селективной -трансаминазы, аминотрансферазы с разветвленной аминокислотной цепью (ВСАТ) и D-аминокислотной трансаминазы (DATA). Предпочтительной аминотрансферазой с разветвленной аминокислотной цепью является аминотрансфераза, полученная из Е. coli. В предпочтительном варианте осуществления изобретения по меньшей мере одной запрашиваемой последовательностью биомолекулы по меньшей мере одной лиазы, используемой на стадии (а), является аминодезоксихоризмат-лиаза (ADCL). Настоящее изобретение относится способу получения (R)-селективной -трансаминазы, являющемуся быстрым, эффективным, простым, надежным и нетрудоемким. Следовательно, настоящее изобретение дает возможность идентифицировать и получить (R)-селективную -трансаминазу, что до настоящего времени было неизвестно или недоступно, открывая множество путей обеспечения хиральных аминов, предпочтительно (R)-энантиомеров, в частности очень эффективных путей асимметрического синтеза. Об этом явно свидетельствует тот факт, что даже белки с низкой степенью идентичности последовательности, например только 35%, известной (R)-селективной -трансаминазе, в противоположность ожиданиям, фактически представляют собой (R)-селективные -трансаминазы. Особенно преимущественным и неожиданным в способе настоящего изобретения является то, что он дает возможность обеспечить предпочтительно (R)-селективные -трансаминазы из Aspergillus terreus и Mesorhizobium loti, благодаря чему трансаминаза из Aspergillus представляет собой первую эукариотическую -трансаминазу, и она превращает в субстраты с (R)-селективностью. В основу настоящего изобретения, главным образом, была положена техническая идея, заключающаяся в том, что белки или кодирующие их нуклеотидные последовательности даже с низкой степенью идентичности последовательностей известным последовательностям трансаминаз или лиаз, в частности(R)-селективных -трансаминаз, могут быть использованы в качестве потенциального источника для идентификации и получения (R)-селективных -трансаминаз, где предполагаемые (R)-селективные трансаминазы скринируют и получают путем выявления нежелательных белков и тем самым позитивного отбора белков, обнаруживающих конкретные мотивы последовательностей в аминокислотной последовательности, которой может быть приписана желаемая ферментативная активность. Таким образом, в настоящем изобретении для идентификации и получения (R)-селективных -трансаминаз специально используются конкретные структурные признаки, в частности отсутствие и присутствие конкретных аминокислот в предполагаемой (R)-селективной -ТА. В соответствии с этим на первой стадии (а) настоящего способа запрашиваемую последовательность биомолекулы по меньшей мере одной трансаминазы или лиазы, например известной (R)селективной со-трансаминазы, или по меньшей мере одной характерной части последовательности такой лиазы или трансаминазы, предпочтительно (R)-специфической со-трансаминазы, которая предпочтительно может быть использована для идентификации PLP-зависимого фермента укладки типа IV, обеспечивают вместе по меньшей мере с одним банком биомолекул. Предпочтительно запрашиваемая последовательность биомолекулы представляет собой ORF (открытая рамка считывания) трансаминазы или лиазы, в частности (R)-селективной -ТА, кодирующую нуклеотидную последовательность или ее характерную часть. В другом варианте осуществления изобретения указанная запрашиваемая последовательность биомолекулы представляет собой саму ORF-аминокислотную последовательность или ее характерную часть. В контексте настоящего изобретения характерной частью запрашиваемой последовательности био-3 022764 молекулы по меньшей мере одной трансаминазы или лиазы, в частности известной (R)-селективной трансаминазы, является последовательность биомолекулы, которая в форме ее молекулы последовательности ДНК гибридизуется в соответствующих условиях с полноразмерной запрашиваемой последовательностью биомолекулы, в частности с ДНК-последовательностью ORF. Методы гибридизации нуклеиновых кислот, таких как ДНК, хорошо известны и описаны, например, в руководстве Molecular Cloning, Third Edition (2001); Methods for General and Molecular Bacteriology,ASM Press (1994); Immunology methods manual, Academic Press (Molecular) и во многих других известных руководствах. Ниже описаны примеры гибридизации в жестких условиях. Фильтр с иммобилизованной на нем нуклеиновой кислотой и нуклеиновую кислоту, используемую в качестве зонда, инкубируют в растворе,содержащем 50% формамид, 5XSSC (750 мМ хлорид натрия и 75 мМ цитрат натрия), 50 мМ фосфат натрия (рН 7,6), 5 Х раствор Денхардта, 10% сульфат декстрана и 20 мкг/л денатурированной ДНК спермы лосося, при 42 С в течение ночи. После инкубации фильтр промывают в 0,2 Х растворе SSC (примерно при 65 С). Такие жесткие условия гибридизации могут быть изменены путем коррекции концентрации формамида (условия становятся менее жесткими по мере снижения концентрации формамида) и путем изменения концентрации солей и температуры. Гибридизацию в условиях низкой жесткости осуществляют, например, путем инкубации фильтра с иммобилизованной на нем нуклеиновой кислотой и нуклеиновой кислотой, используемой в качестве зонда, в растворе, содержащем 6XSSCE (20XSSCE: 3 моль/л хлорида натрия, 0,2 моль/л дигидрофосфата натрия и 0,02 моль/л EDTA, рН 7,4), 0,5% ДСН, 30% формамида и 100 мкг/л денатурированной ДНК спермы лосося при 37 С в течение ночи и промывки фильтра IX раствором SSC, содержащим 0,1% ДСН(50 С). В особенно предпочтительном варианте осуществления настоящего изобретения термин "по меньшей мере одна запрашиваемая последовательность биомолекулы по меньшей мере одной трансаминазы или лиазы" следует понимать как последовательность биомолекулы, которая пригодна для отбора последовательностей биомолекул в соответствии с присутствием или отсутствием мотивов последовательностей (с 1)-(с 6), как идентифицировано в данном описании. Соответственно такая запрашиваемая последовательность представляет собой последовательность для скрининга и идентификации таких последовательностей биомолекул, которые на уровне аминокислотной последовательности не содержат по меньшей мере один из мотивов аминокислотных последовательностей (c1)-(c3) и которые содержат на аминокислотном уровне мотивы последовательностей (с 4)-(с 6). Такая запрашиваемая последовательность биомолекулы может быть включена в качестве информации об аминокислотной последовательности или в качестве информации о молекуле ДНК-последовательности или о последовательности ДНК. В предпочтительном варианте осуществления изобретения характерная часть запрашиваемой последовательности биомолекулы, используемой на стадии (а), включает, предпочтительно состоит из области от положения 30 до положения 170, более предпочтительно от положения 30 до положения 120ORF трансаминазы, предпочтительно (R)-селективной -трансаминазы или лиазы. На следующей стадии (b) в банке биомолекул осуществляют поиск запрашиваемой последовательности биомолекулы для идентификации группы первых последовательностей биомолекул-мишеней,имеющей, по меньшей мере, минимальную степень идентичности последовательностей, которые по меньшей мере на 20%, предпочтительно на 25%, предпочтительно на 32%, предпочтительно по меньшей мере на 33%, более предпочтительно на 34%, по меньшей мере на 35%, по меньшей мере на 36%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90% или по меньшей мере на 95% идентичны запрашиваемой последовательности биомолекулы, основанной на уровне аминокислотной последовательности, и где указанная группа первых биомолекул-мишеней представляет собой первый отбор из банка биомолекул,используемого на стадии (а). Указанная степень идентичности последовательностей представляет собой идентичность последовательностей, оцененную предпочтительно между последовательностями, по меньшей мере, характерной части последовательности, предпочтительно, по существу, полноразмерной последовательности, в частности полноразмерной ORF запрашиваемой последовательности биомолекулы и, по меньшей мере, характерной частью последовательности, предпочтительно, по существу, полноразмерной последовательности, в частности полноразмерной ORF скринируемых последовательностей биомолекул. После проведения указанной стадии (b) в этой группе первых последовательностей биомолекулы-мишени на стадии (с) проводят отбор таких последовательностей, которые не содержат в качестве мотива последовательности (c1) в положениях 95-97 аминокислотную последовательность Tyr-Xa1Xa2, где Xa1 представляет собой аминокислоту Ile, Val, Leu, Met, Phe и Ха 2 представляет собой аминокислоту Arg или Lys, или которые не содержат в качестве мотива последовательности (с 2) в положениях 97-99 аминокислотную последовательность Tyr-Xaa-Gln, где Хаа предпочтительно представляет собой обычную аминокислоту, и в области 105-111, предпочтительно в положениях 107-109 аминокислотную последовательность Arg-Xaa-Ха 3, где Ха 3 представляет собой обычную аминокислоту, предпочтительноHis, или которые не содержат в качестве мотива последовательности (с 3) в положении 38 аминокислотуArg-Xa4-Xa5, где Ха 4 предпочтительно представляет собой обычную аминокислоту, предпочтительноGly, и Ха 5 предпочтительно представляет собой обычную аминокислоту, предпочтительно Tyr; что дает возможность дифференцировать и отделять эти последовательности от последовательностей, которые имеют по меньшей мере один из мотивов последовательностей (c1), (с 2) или (с 3), идентифицированных выше. На другой последующей или одновременной стадии отбора осуществляют отбор таких последовательностей биомолекул, которые соответствуют мотиву последовательности (с 4), где в положении 95 присутствует другая аминокислота, не являющаяся Tyr, Arg, Lys, или в положении 95 присутствует аминокислота Tyr, и в положении 97 присутствует аминокислота, не являющаяся Arg и Lys; и которые соответствуют мотиву последовательности (с 5), где в положении 40 присутствует аминокислота, не являющаяся Lys и Arg; и мотиву последовательности (с 6), в области от положения 161 до положения 165, в которой предпочтительно в положении 163 присутствует по меньшей мере одна аминокислота Lys и предпочтительно одна аминокислота Lys, где указанная стадия дает возможность проводить отбор и идентификацию группы последовательностей второй биомолекулы-мишени. Группа последовательностей второй биомолекулы-мишени, идентифицированных и скринированных для идентифицированных выше мотивов последовательностей, представляет собой последовательности биомолекул, представляющих собой или кодирующих белок, обладающий активностью (R)-селективной -трансаминазы. В контексте настоящего изобретения трансаминаза представляет собой пиридоксалфосфатзависимый фермент, катализирующий перенос аминогрупп, предпочтительно классифицированный в укладку типа IV. Трансаминазы классифицированы в соответствии с Системой классификации ферментов Е.С. 2.6.1.Х. В особенно предпочтительном варианте осуществления настоящего изобретения трансаминаза представляет собой (R)-селективную трансаминазу, в частности в предпочтительном варианте осуществления -трансаминазу. В контексте настоящего изобретения белком, обладающим активностью(R)-селективной -трансаминазы, является белок, который в соответствующих реакционных условиях катализирует перенос азотных групп, таких как аминогруппы, передаваемые от донора к акцептору, например, как это в состоянии делать (R)-селективная (омега)-трансаминаза (бета-аланин-пируваттрансаминазы). В контексте настоящего изобретения (R)-селективная -трансаминаза является предпочтительным ферментом, классификационным кодом Е.С.2.6.1.18. В контексте настоящего изобретения термин "оптически активный хиральный амин" относится к такому же понятию, как термин "энантиомерно активный хиральный амин". Эти термины, в частности,означают препарат, который, по существу, не содержит даже в более предпочтительном варианте осуществления нежелательного энантиомера. Соответственно оптически активный хиральный амин, по существу, содержит избыток одного энантиомера или даже состоит только из одного энантиомера. В частности, в контексте настоящего изобретения оптически активный хиральный амин имеет оптическую чистоту по меньшей мере 70, 80, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99,5, 99,6, 99,7, 99,8%, в частности по меньшей мере 99,9%. В настоящем изобретении оптическая чистота выражена в % избытка одного энантиомера по отношению к другому энантиомеру. Таким образом, оптическая чистота в % означает отношение разности концентраций (R)- и (S)-энантиомеров к сумме концентраций указанных знантиомеров (оптическая чистота А в %=([А]-[В]):([А]+[В])100, где А и В представляют собой концентрации (R)- и (S)энантиомеров или наоборот). В контексте настоящего изобретения банк биомолекул представляет собой источник самих биомолекул или набор данных о последовательностях биомолекул, в частности данных о последовательностях полинуклеотидов или полипептидов. В контексте настоящего изобретения биомолекулой предпочтительно является полинуклеотидная молекула, несущая генетическую информацию, в частности молекула ДНК. В другом предпочтительном варианте осуществления настоящего изобретения биомолекулой является полипептид, в частности белок, содержащий ряд аминокислот. Таким образом, банк биомолекул может представлять собой или физический источник биомолекул, в частности банк генов, например библиотеку кДНК или геномную библиотеку, или совокупность данных об указанных биомолекулах, в частности совокупность данных о последовательностях, например об аминокислотных последовательностях или полинуклеотидных последовательностях, в частности о последовательностях ДНК. Настоящее изобретение также относится к последовательностям биомолекул, аминокислотным последовательностям, полинуклеотидным последовательностям или нуклеотидным последовательностям, в частности к последовательностям ДНК, где указанный термин, с одной стороны, означает физическое вещество per se, что означает белок, полипептид или молекулу ДНК, и, с другой стороны, несет в себе информацию об указанном химическом веществе, которая в случае полинуклеотида указывает на тип,порядок и число нуклеотидов, присутствующих в данном полинуклеотиде, и в случае полипептида на тип, порядок и число отдельных аминокислот, образующих данный полипептид. В контексте настоящего изобретения термины "биомолекула", "молекула ДНК", "полинуклеотидная молекула", "полипептид" или "белок" означают химические вещества per se. В данном случае настоящее изобретение конкретно относится к информации об аминокислотной последовательности, о последовательности ДНК, о полинуклеотидной последовательности или о последовательности биомолекулы, при этом она не относится к физической форме биомолекулы, но скорее к содержащейся в ней информации,то есть к типу, порядку и числу ее составляющих, а именно аминокислот или нуклеотидов. В одном из предпочтительных вариантов осуществления (А) на стадиях (а)-(с) настоящее изобретение относится к банку биомолекул, представляющему собой совокупность данных о последовательностях, в одном варианте осуществления о полинуклеотидных последовательностях, в частности о последовательностях ДНК, или в другом варианте осуществления об аминокислотных последовательностях,где поиск указанных полинуклеотидных и аминокислотных последовательностей осуществляют на стадии (b) с применением соответствующих инструментов выравнивания последовательностей, предпочтительно программы BLAST, которая представляет собой основной инструмент поиска путем локального выравнивания, проводимого в целях идентификации группы последовательностей первых биомолекулмишеней (Altschul, S.F. et al., 1990. J. Mol. Biol., 215:403; Altschul, S.F. et al., 1997, Nucleic Acid Res.,25:3389-3402). Другими подходящими программами являются программы GAP, BESTFIT и FASTA, входящие в пакет программ Wisconsin Genetics Software Package (Genetics Computer Group (GCG), Madison,Wl, USA). Ha дополнительной стадии (с) группу идентифицированных последовательностей первых биомолекул подвергают дополнительным стадиям отбора, в процессе которых мотивы идентифицированных последовательностей используют для идентификации и негативного отбора мотивов последовательностей (c1)-(c3) и позитивного отбора мотивов последовательностей (с 4)-(с 6) желательной последовательности биомолекул. После идентификации второй группы последовательностей биомолекул авторами настоящего изобретения была использована информация об указанных последовательностях в целях получения соответствующих молекул олиго- или полинуклеотидов, например праймеров для гибридизации, а также в целях скрининга и физического отбора молекул последовательностей ДНК из банка биомолекул, где указанные молекулы ДНК кодируют ферменты, идентифицированные во второй группе. Таким образом, в предпочтительном варианте осуществления информация о последовательностях, полученная на стадии (с), может быть использована для клонирования соответствующего гена из библиотеки геномных ДНК или кДНК и экспрессии белка, например в Е. coli, Sacischaromyces cerevisiae, Pichia pastoris или т.п. В другом предпочтительном варианте осуществления настоящего изобретения информацию о последовательностях, полученную на стадии (с), используют для синтеза de novo желаемой (R)селективной -трансаминазы. В таком предпочтительном варианте осуществления информацию о последовательностях, полученную на стадии (с), возможно использовать для синтеза гена, например, с оптимизированной встречаемостью кодонов и стабильной мРНК и для клонирования и экспрессии такого гена. Согласно такому предпочтительному варианту осуществления в качестве по меньшей мере одного банка биомолекул на стадии (а) возможно использовать любой банк биомолекул, содержащий информацию о последовательностях ДНК или информацию об аминокислотных последовательностях. В данном случае банк биомолекул содержит информацию о последовательностях ДНК, где любая указанная информация транслируется в информацию об аминокислотных последовательностях для последующей обработки на стадиях (b)-(с), в которых аминокислотные последовательности используют в качестве запрашиваемых последовательностей биомолекул и в качестве мотивов аминокислотной последовательности; или на стадиях (b)-(с), которые проводят в банке последовательностей ДНК с использованием запрашиваемых последовательностей биомолекул и мотивов последовательностей (с 1)-(с 6), подвергнутых обратной трансляции в последовательность ДНК. В другом предпочтительном варианте осуществления настоящего изобретения идея настоящего изобретения применяется в варианте осуществления (В) на стадии (а), проводимой с использованием банка биомолекул в физическом формате банка генов, в частности генной библиотеки, такой как библиотека кДНК, метагеномная или геномная библиотека, где используются молекулы ДНК, кодирующие всю или, по меньшей мере, характерную часть по меньшей мере одной (R)-селективной -трансаминазы для поиска, идентификации и отбора группы последовательностей первых биомолекул, в частности молекул последовательности ДНК, которые, как рассчитано на уровне аминокислотной последовательности, по меньшей мере на 20%, предпочтительно на 32% идентичны запрашиваемой последовательности молекулы ДНК. Последующую стадию (с) предпочтительно осуществляют в указанной группе первых молекул ДНК с использованием нуклеотидных молекул-праймеров, позволяющих идентифицировать и проводить позитивный и негативный отбор желаемых мотивов последовательностей (c1)-(с 6). В контексте настоящего изобретения термин "мотив последовательности" означает специфические селективные признаки аминокислотной последовательности предполагаемой (R)-селективной трансаминазы, как конкретно идентифицировано на стадии (с) заявленного способа. В частности, первым мотивом последовательности (c1) является мотив последовательности, который был идентифицирован в положениях 95-97, имеющих аминокислотную последовательность Tyr-Xa1-Xa2 в указанном порядке,где Xa1 представляет собой аминокислоту Ile, Val, Leu, Met, Phe и Ха 2 представляет собой аминокислотуArg или Lys; и который согласно настоящему изобретению не должен присутствовать в конечной отобранной последовательности биомолекулы. Вторым мотивом последовательности (с 2) является мотив последовательности, который был идентифицирован в положениях 97-99, имеющих аминокислотную последовательность Tyr-Xaa-Gln в указанном порядке, где Хаа представляет собой любую аминокислоту; и в положениях 105-111, предпочтительно в положениях 107-109, имеющих аминокислотную последовательность Arg-Xaa-Ха 3, в указанном порядке, где Хаа представляет собой любую аминокислоту и Ха 3 предпочтительно представляет собой аминокислоту His, причем указанный мотив также не должен присутствовать в конечной отобранной последовательности биомолекулы. Третьим мотивом последовательности (с 3) является мотив последовательности, характеризующийся присутствием Thr в положении 38,Lys в положении 97, и присутствием аминокислотной последовательности Arg-Xa4-Xa5 в положениях 107-109, где Ха 4 предпочтительно представляет собой аминокислоту Gly, и Ха 5 предпочтительно представляет собой аминокислоту Tyr, причем указанный мотив (с 3) не должен присутствовать в конечной отобранной последовательности биомолекул. Четвертый мотив последовательности (с 4) должен иметь в положении 95 другую аминокислоту, не являющуюся Tyr, Arg, Lys, или он должен иметь Tyr в положении 95, но в положении 97 он не должен иметь Arg или Lys, что позволяет идентифицировать предполагаемую (R)-селективную -трансаминазу,полученную согласно настоящему изобретению. Пятый мотив последовательности (с 5), присутствующий в полученной и идентифицированной предполагаемой (R)-селективной -трансаминазе, должен иметь в положении 40 аминокислоту, не являющуюся Lys и Arg. Шестой мотив последовательности (с 6),присутствующий в полученной и идентифицированной (R)-селективной -ТА, должен иметь в области от положения 161 до положения 165 по меньшей мере одну аминокислоту Lys, предпочтительно одну аминокислоту Lys, и в положении 163 предпочтительно аминокислоту Lys. В контексте настоящего изобретения идентификация и определение локализации положений аминокислот описаны ниже. Первые последовательности биомолекул-мишеней подвергают выравниванию,предпочтительно множественному выравниванию друг с другом и с запрашиваемой последовательностью биомолекулы. Выравнивание может быть осуществлено с использованием стандартной компьютерной программы для выравнивания, такой как STRAP, в частности ClustalW, предпочтительноClustalW3D. В другом варианте осуществления выравнивание этих последовательностей можно также осуществлять попарно, то есть каждую первую последовательность биомолекулы-мишени можно сопоставить с запрашиваемой последовательностью биомолекулы. В предпочтительном варианте осуществления первые последовательности биомолекулы-мишени можно сопоставить с ВСАТ Е. coli. В другом предпочтительном варианте осуществления первую последовательность биомолекулы-мишени можно сопоставить с другой запрашиваемой последовательностью биомолекулы, используемой в настоящем изобретении, например (R)-селективной -трансаминазы. Нумерация положений аминокислот, представленных в настоящем изобретении, определяется положением соответствующего мотива последовательности в запрашиваемой последовательности биомолекулы, используемой в качестве позиционного стандарта. Описанное в данном описании сопоставление соответствующих положений аминокислот осуществляют путем выравнивания последовательностей первой биомолекулы-мишени с последовательностями мотивов, присутствующих в запрашиваемой последовательности биомолекулы. В качестве примера, в целях идентификации положений 95-99 в предполагаемой -трансаминазе,которые были проанализированы на отсутствие мотивов (c1)-(с 3) и на присутствие мотива (с 4), в качестве стандарта для указания положений использовали известную аминокислотную последовательность ВСАТ Е. coli от положения 92 до положения 100, а именно последовательность TSAYIRPLI (SEQ ID NO: 9). Известная аминокислотная последовательность от положения 35 до положения 42, а именно последовательность VFEGIRCY (SEQ ID NO: 10) ВСАТ Е. coli имеет G в положении 38 для мотива последовательностей (с 3) и (с 5). Аминокислотная последовательность DVGMGVNP в аминокислотной последовательности ВСАТ Е. coli (SEQ ID NO: 11) от положения 104 до положения 111 имеет мотив (с 3) в положениях 105-111. Аминокислотная последовательность PTAAKAGGN в положениях 159-167 известной последовательности ВСАТ Е. coli (SEQ ID NO: 12) имеет в положении 163 аминокислоту K в мотиве последовательности (с 6). Таким образом, в предпочтительном варианте осуществления настоящего изобретения указанная биомолекула представляет собой белок и последовательность биомолекулы представляет собой аминокислотную последовательность. Соответственно в одном из предпочтительных вариантов изобретения банком биомолекул является банк, в частности база данных, где указанным банком является банк, содержащий собранную информацию о различных белках, в частности информацию о конкретных аминокислотных последовательностях. Предпочтительными банками являются база данных белков NCBI, база данных UniProtKB/SwissProt и база данных UniProtKB/TrEMBL. В предпочтительном варианте осуществления изобретения биомолекула также может представлять собой молекулу ДНК и последовательность биомолекулы может представлять собой последовательность ДНК. Соответственно в одном из вариантов осуществления банком биомолекул является банк, в частности база данных, в которой собрана информация о различных полинуклеотидных последовательностях, в частности о последовательностях ДНК. В предпочтительном варианте осуществления изобретения применяется способ, описанный выше,где банком биомолекул является база данных биомолекул, и поиск в этой базе данных биомолекул осуществляют, как описано на стадии (b) с помощью программы выравнивания последовательностей биомолекул, в частности BLAST. В предпочтительном варианте осуществления предусматривается, что если банком биомолекул является база данных, то эту базу данных скринируют на стадии (b) либо на любую аминокислотную последовательность, если такой базой данных биомолекул является база данных аминокислотных последовательностей, либо на последовательность ДНК, если такой базой данных биомолекул является база данных последовательностей ДНК. В настоящим изобретением предусмотрена конечная стадия (d) для обеспечения биомолекулы,предпочтительно молекулы ДНК или молекулы аминокислотной последовательности, то есть белка, либо путем синтеза трансаминазы de novo, либо путем выделения из существующего банка генов полинуклеотидов, кодирующих желаемую трансаминазу, с помощью праймеров, где полинуклеотиды определены на основе вторых последовательностей биомолекул. Полученные полинуклеотиды, в частности молекулы последовательности ДНК, экспрессируют в соответствующих условиях в культуральной среде,подходящей для экспрессии желаемой трансаминазы. Настоящее изобретение также относится к способу скрининга и идентификации (R)-селективной трансаминазы, включающему вышеописанные стадии (а)-(с), в частности обеспечение по меньшей мере одной запрашиваемой последовательности биомолекулы, по меньшей мере одной (R)-селективной трансаминазы и по меньшей мере одного банка биомолекул; поиск в банке биомолекул запрашиваемой последовательности биомолекулы для идентификации группы первых последовательностей биомолекулмишеней, где первые последовательности биомолекул по меньшей мере на 20%, предпочтительно на 25% и более предпочтительно на 32% идентичны запрашиваемой последовательности биомолекулы, как рассчитано на уровне аминокислотной последовательности; отбор в группе первых последовательностей биомолекул-мишеней, которые не содержат на уровне аминокислотной последовательности каких-либо мотивов последовательностей (c1)-(с 3), а именно мотива (c1), имеющего в положениях 95-97 аминокислотную последовательность Tyr-Xa1-Xa2, где Xa1 представляет собой аминокислоту Ile, Val, Leu, Met,Phe и Ха 2 представляет собой аминокислоту Arg или Lys, или мотива (с 2), имеющего в положениях 9799 аминокислотную последовательность Tyr-Xaa-Gln, где Хаа представляет собой любую аминокислоту,и в области от положения 105 до положения 111, предпочтительно в положениях 107-109, аминокислотную последовательность Arg-Xaa-Ха 3, где Ха 3 представляет собой аминокислоту, предпочтительно His,или мотива (с 3), имеющего Thr в положении 38, Lys в положении 97, и в положении 107-109 аминокислотную последовательность Arg-Xa4-Ха 5, где Ха 4 представляет собой аминокислоту, предпочтительноGly, и Ха 5 представляет собой аминокислоту, предпочтительно Tyr, и которые содержат мотивы (с 4),(с 5) и (с 6), а именно мотив (с 4), который имеет на аминокислотном уровне в положении 95 другую аминокислоту, не являющуюся Tyr, Arg, Lys, либо в положении 95 аминокислоту Tyr, и в положении 97 аминокислоту, не являющуюся Arg или Lys; мотив (с 5), который в положении 40 не имеет Lys или Arg, и мотив (с 6), который в области от положения 161 до положения 165, предпочтительно в положении 163 имеет Lys, чтобы идентифицировать группу вторых последовательностей биомолекул, которые представляют собой последовательности биомолекул (R)-селективной -ТА. В наиболее предпочтительном варианте осуществления настоящее изобретение относится к (R)селективным -трансаминазам, которые могут быть получены с применением способов согласно изобретению. В частности, настоящее изобретение относится к белкам и к последовательностям ДНК, в частности к молекулам ДНК, кодирующим указанный белок, происходящий из Mesorhizobium loti и Aspergillusterreus, последовательности которых представлены в SEQ ID NO: 1 и 2 для Mesorhizobium loti и SEQ IDNO: 3 и 4 для Aspergillus terreus. В конкретном и в наиболее предпочтительном варианте осуществления изобретение предусматривает, что (R)-селективные -трансаминазы, которые могут быть получены или получены согласно настоящему изобретению, например трансаминазы, которые были идентифицированы в SEQ ID NO: 1 и 3, могут быть использованы в реакции переаминирования, в частности они могут быть использованы в способе получения оптически активного хирального амина, предпочтительно (R)энантиомера хирального амина, включающем осуществление реакции по меньшей мере одного аминоакцептора и по меньшей мере одного аминодонора с (R)-селективной -трансаминазой согласно изобретению, в частности SEQ ID NO: 1 или 3, и получение оптически активного хирального амина. В предпочтительном варианте осуществления способ получения оптически активного хирального амина представляет собой асимметрический синтез, осуществляемый с использованием соединения, содержащего кетогруппу, предпочтительно кетон, и аминодонора. В другом предпочтительном варианте осуществления способ синтеза осуществляют посредством кинетической реакции разделения. Аминодонором предпочтительно является смесь рацемических аминов, и аминоакцептором предпочтительно являются кетоны,используемые в качестве продуктов извлечения. В предпочтительном варианте осуществления оптически активный хиральный амин синтезируют из кетонов с использованием (R)-селективной -трансаминазы согласно изобретению методом асимметрического синтеза, где степень превращения в требуемый оптически активный хиральный амин, то есть(R)-энантиомер, составляет по меньшей мере 70, 80, 90, 95, 98, 99, 99,5% и наиболее предпочтительно 100%. В другом предпочтительном варианте осуществления (R)-селективную -трансаминазу согласно изобретению используют в кинетической реакции разделения рацемических аминов, где степень превращения в требуемый оптически активный хиральный амин, то есть (S)-энантиомер, предпочтительно составляет по меньшей мере 30, 40, 45, 46, 47, 48, 49%, в частности 50%. Концентрации для анализа оптической чистоты и степени превращения могут быть определены,например, с помощью ВЭЖХ, капиллярного электрофореза (КЭ), газовой хроматографии (ГХ) или оптическими или флуориметрическими методами. Таким образом, в предпочтительном варианте осуществления настоящее изобретение относится к способу получения оптически активного хирального амина, включающему осуществление реакции аминоакцепторного соединения, содержащего кетогруппу, и рацемической смеси амина в присутствии трансаминазы, в частности (R)-селективной -трансаминазы, предпочтительно трансаминазы согласно изобретению, с получением (S)-энантиомера хирального амина. В другом предпочтительном варианте осуществления настоящее изобретение относится к способу получения оптически активного хирального амина, в частности (R)-энантиомера указанного амина,включающему осуществление реакции аминоакцепторного соединения, содержащего кетогруппу, и аминодонора в присутствии (R)-селективной -трансаминазы, в частности трансаминазы, которая может быть синтезирована согласно настоящему изобретению, с получением (R)-энантиомера указанного амина. В контексте настоящего изобретения аминоакцептором является молекула, способная принимать аминогруппу, переносимую от аминодонора посредством трансаминазы, в частности -трансаминазы. В особенно предпочтительном варианте осуществления настоящего изобретения аминоакцептор содержит функциональную кетоновую группу. В особенно предпочтительном варианте осуществления настоящего изобретения аминоакцептор выбран из группы, состоящей из фенилпировиноградной кислоты и ее соли, пировиноградной кислоты и ее соли, ацетофенона, 2-кетоглутарата, 3-оксобутирата, 2-бутанона, 3-оксопирролидина (3-ОР), 3 пиридилметилкетона (3-PMK), этилового эфира 3-оксомасляной кислоты (3-ОВЕЕ), метилового эфира 3 оксопентановой кислоты (3-ОРМЕ), N-1-boc-оксопиперидинона, N-1-boc-3-оксопирролидина (В 3 ОР), 3 оксопиперидина, N-1-boc-3-оксопиперидина (B3OPi), 1-Cbz-3-оксопиперидина (C3OPi), 1-Cbz-3 оксопирролидина (С 3 ОР), алкил-3-оксобутаноатов, метоксиацетона и 1-оксотетралона. В контексте настоящего изобретения аминодонором является молекула, способная переносить аминогруппу на аминоакцептор под действием трансаминазы, в частности -трансаминазы. В особенно предпочтительном варианте осуществления указанным аминодонором является амин или аминокислота. В особенно предпочтительном варианте осуществления аминодонор выбран из группы, состоящей из -аланина, аланина, -метилбензиламина (-МВА), глутамата, фенилаланина, глицина, 3 аминобутирата, изопропиламина, 2-аминобутирата и -аминобутирата или их солей, например хлорида любого из указанных соединений. В особенно предпочтительном варианте осуществления полученным кетоновым продуктом может быть фенилпировиноградная кислота и ее соль, пировиноградная кислота и ее соль, глиоксиловая кислота и ее соль, ацетофенон, 2-кетоглутарат, ацетон, 3-оксобутират, 2-бутанон,3-оксопирролидин (3-ОР), 3-пиридилметилкетон (3-PMK), этиловый эфир 3-оксомасляной кислоты (3 ОВЕЕ), метиловый эфир 3-оксопентановой кислоты (3-ОРМЕ), N-1-boc-оксопиперидинон и N-1-boc-3 оксопирролидин (В 3 ОР), или их соли, например хлорид любого из этих соединений. В другом предпочтительном варианте настоящее изобретение относится к способу получения оптически активного хирального амина, выбранного из группы аминов, имеющих оптически активную аминогруппу, в частности аминов с алкильными группами, разветвленными алкильными группами или с арилалкильными группами. Более конкретно, указанные амины, в частности моно- или бициклические амины, представляют собой аминопроизводные 5-6-членных циклических углеводородов или S-, О- илиN-замещенных гетероциклических углеводородов, или ароматические амины, в частности алкил- или алкоксизамещенные ароматические амины. В предпочтительном варианте осуществления полученные хиральные амины выбраны из группы, состоящей из фенилаланина, аланина, 3-аминопиперидина, алкил 3-аминобутаноатов, 3-аминопирролидина (3-АР), 3-пиридил-1-этиламина (3-РЕА), N-1-boc-3 аминопирролидина (В 3 АР), N-1-boc-3-аминопиперидина (B3APi), 1-Cbz-3-аминопиперидина (C3APi), 1Cbz-3-аминопирролидина (С 3 АР), этилового эфира 3-аминомасляной кислоты (3-АВМЕ), метилового эфира 3-аминопентановой кислоты (3-АРМЕ), -метилбензиламина (-МВА), 1-аминотетралина, 3,4 диметоксифенилацетона, -метил-4-(3-пиридил)бутанамина, -аминобутирата, глутамата, изопропиламина, -аминобутирата, втор-бутиламина, метоксиизопропиламина, производных 3-аминопирролидина,1-N-Boc-3-аминопиперидина, цефалоспорина и производных цефалоспорина. В особенно предпочтительном варианте настоящее изобретение включает проведение реакции 3 ОР с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-3 АР. В другом предпочтительном варианте настоящее изобретение включает проведение реакции 3-РМК с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-3-РЕА. В другом предпочтительном варианте настоящее изобретение включает проведение реакции 3 ОВЕЕ с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-3 АВЕЕ. В другом предпочтительном варианте настоящее изобретение включает проведение реакции 3 ОРМЕ с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-3 АРМЕ. В другом предпочтительном варианте настоящее изобретение включает проведение реакции В 3 ОР с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-В 3 АР. В другом предпочтительном варианте настоящее изобретение включает проведение реакции B3OPi с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-B3APi. В другом предпочтительном варианте настоящее изобретение включает проведение реакции C3OPi с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-C3APi. В другом предпочтительном варианте настоящее изобретение включает проведение реакции С 3 ОР с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-С 3 АР. В другом предпочтительном варианте настоящее изобретение включает проведение реакции ацетофенона с (R)-селективной -трансаминазой и аминодонором, с получением оптически активного (R)-MBA. В другом предпочтительном варианте настоящее изобретение включает проведение реакции аминоакцептора, в частности моно- или бициклических 5-6-членных циклических углеводородов, содержащих оксогруппу, или из S-, О- или N-замещенных гетероциклических углеводородов, или ароматических аминов, в частности алкил- или алкоксизамещенных ароматических аминов, с аминодонором и (R)селективной -трансаминазой, с получением аминов, в частности моно- или бициклических аминов, например аминопроизводных 5-6-членных циклических углеводородов или S-, О- или N-замещенных гетероциклических углеводородов, или ароматических аминов, в частности алкил- или алкокси-замещенных ароматических аминов, например, в (R)-форме. В особенно предпочтительном варианте осуществления аминоакцептор и аминодонор подвергают реакции с трансаминазой в водной среде, например в физиологическом буфере. В особенно предпочтительном варианте осуществления реакцию переаминирования осуществляют при рН от 5 до 9, в частности от 7 до 8,5. В особенно предпочтительном варианте осуществления реакцию осуществляют при температуре 10-65 С, предпочтительно 20-50 С, например при 18-25 С, и предпочтительно при комнатной температуре или при 34-39 С, в частности 37 С. В другом предпочтительном варианте осуществления настоящего изобретения молярное отношение аминоакцептора к аминодонору составляет 1:1-1:5, в частности 1:1-1:2. В предпочтительном варианте осуществления настоящего изобретения ферментативная активность может составлять от 1 до 20000 мкмоль/мин. Настоящее изобретение также относится к способу анализа трансаминазы, в частности к способу характеризации свойств трансаминазы, включающему следующие стадии:i) обеспечение заряженного аминоакцептора, заряженного аминодонора и трансаминазы, предпочтительно -трансаминазы, и наиболее предпочтительно (R)-селективной -трансаминазы, которые могут быть синтезированы способом согласно изобретению;ii) осуществление реакции аминоакцептора и аминодонора с трансаминазой в реакционной среде и тем самымiii) определение проводимости реакционной среды в первом наборе реакционных условий;iv) после проведения стадии iii) определение проводимости реакционной среды во втором наборе реакционных условий с последующим вычислением по меньшей мере двух величин проводимости, отражающих свойства трансаминазы. В процессе проведения катализируемой трансаминазой и предпочтительно -ТА-катализируемой реакции заряженных субстратов, например аминодонора, предпочтительно амина, и аминоакцептора, и предпочтительно кетокислоты, например пирувата, проводимость снижается, поскольку в данном случае образуются незаряженный кетон и цвиттерионный аминокомпонент, предпочтительно аминокислота,например аланин. Способ анализа согласно изобретению позволяет осуществлять простой мониторинг прохождения реакции. Во избежание получения слишком высоких исходных значений проводимости наиболее предпочтительно использовать буфер с низкой проводимостью, в частности буфер на основе CHES (N- 10022764 циклогексил-2-аминоэтансульфоновой кислоты) с низкой проводимостью. Калибровка проводимости может быть предпочтительно осуществлена путем имитации различных превращений. В качестве примера для пары стандартных субстратов, таких как -метилбензиламин и пируват, 1 мМ превращение соответствует изменению 44 мкС. Сравнение измеренных скоростей реакции с помощью капиллярного электрофореза давало превосходное подтверждение надежности способа согласно изобретению. Клеточные экстракты не оказывали какого-либо значительного влияния на эффективность способа согласно изобретению. Поскольку пируват является наиболее распространенным аминоакцептором фактически для всех -ТА, то этот способ может быть применен в исследованиях трансаминазной активности в отношении различных аминодоноров. Кроме того, может быть также получена информация об энантиоселективности данного фермента. В данном варианте осуществления настоящее изобретение относится к способу анализа трансаминазы, в частности к способу характеризации свойств трансаминазы, которые позволяют проводить анализы на активность трансаминазы в зависимости, например, от величины рН или температуры реакционной среды, а также анализы на стабильность реакции, на влияние добавок или состава буферов. Согласно настоящему способу анализа первое измерение проводимости реакционной среды осуществляют в первом наборе реакционных условий и затем осуществляют по меньшей мере одно второе измерение проводимости реакционной среды для сравнения двух полученных величин проводимости и для анализа величин активности и свойств трансаминазы. Снижение проводимости указывает на активность трансаминазы согласно изобретению в трансаминазной реакции. Медленное снижение, ускоренное снижения или отсутствие снижения проводимости позволяет сделать выводы о свойствах трансаминазы. В контексте настоящего изобретения набором реакционных условий является предпочтительно набор реакционных условий, выбранный из группы, состоящей из температуры, величины рН и состава реакционной среды, предпочтительно реакционных условий, идентифицированных выше, за исключением концентрации выделенных веществ и продуктов. В одном из вариантов осуществления настоящего изобретения набор реакционных условий сохраняют постоянным в течение всего периода прохождения реакции. В другом варианте осуществления настоящего изобретения набор реакционных условий может быть изменен в течение всего периода прохождения реакции. В предпочтительном варианте осуществления настоящее изобретение также относится к такому способу анализа трансаминазы, в котором реакционной средой является буфер с низкой проводимостью. В особенно предпочтительном варианте осуществления указанного способа заряженным аминоакцептором является пируват. В предпочтительном варианте осуществления настоящего изобретения способ анализа трансаминазы осуществляют после получения (R)-селективной -трансаминазы согласно изобретению, и такой способ согласно изобретению может быть применен не только для получения новых и ценных (R)селективных -трансаминаз, но также для определения их свойств. Другими предпочтительными вариантами осуществления настоящего изобретения являются объекты изобретения, представленные в зависимых пунктах формулы изобретения. Настоящее изобретение более подробно описано в нижеследующих примерах и в прилагаемом списке последовательностей.SEQ ID NO: 1 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 2 представляет собой последовательность ДНК, кодирующую SEQ ID NO: 1;SEQ ID NO: 3 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 4 представляет собой последовательность ДНК, кодирующую SEQ ID NO: 3;SEQ ID NO: 5 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 6 представляет собой последовательность ДНК, кодирующую SEQ ID NO: 5;SEQ ID NO: 7 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 8 представляет собой последовательность ДНК, кодирующую SEQ ID NO: 7;SEQ ID NO: 9 представляет собой мотив последовательности ВСАТ Е. coli, используемый для определения аминокислот в положениях 95-99;SEQ ID NO: 10 представляет собой мотив последовательности ВСАТ Е. coli, используемый для определения аминокислоты в положении 38;SEQ ID NO: 11 представляет собой мотив последовательности ВСАТ Е. coli, используемый для определения аминокислоты в положении 107;SEQ ID NO: 12 представляет собой мотив последовательности ВСАТ Е. coli, используемый для определения аминокислоты в положении 163;SEQ ID NO: 13 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Penicillium chrysogenum;SEQ ID NO: 14 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 15 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Aspergillus niger;SEQ ID NO: 16 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 17 представляет собой последовательность ДНК, кодирующую (R)-селективную -TA,происходящую из Aspergillus oryzae;SEQ ID NO: 18 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 19 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Aspergillus fumigatus;SEQ ID NO: 20 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 21 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Neosartorya fischeri;SEQ ID NO: 22 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 23 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Gibberella zeae,SEQ ID NO: 24 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 25 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Hyphomonas neptunium;SEQ ID NO: 26 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 27 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Mesorhizobium loti MAFF303099;SEQ ID NO: 28 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 29 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Roseobacter sp.;SEQ ID NO: 30 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 31 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Marinomonas sp.;SEQ ID NO: 32 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 33 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Rhizobium etli;SEQ ID NO: 34 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 35 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Rhodoferax ferrireducens;SEQ ID NO: 36 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 37 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Jannaschia sp.;SEQ ID NO: 38 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 39 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Labrenzia alexandrii;SEQ ID NO: 40 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 41 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Burkholderia sp.;SEQ ID NO: 42 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 43 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Burkholderia cenocepacia;SEQ ID NO: 44 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 45 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из альфа-протеобактерии;SEQ ID NO: 46 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 47 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из гамма-протеобактерии;SEQ ID NO: 48 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 49 представляет собой последовательность ДНК, кодирующую (R)-селективную -ТА,происходящую из Mycobacterium vanbaalenii иSEQ ID NO: 50 представляет собой полноразмерную аминокислотную последовательность (ORF)SEQ ID NO: 49. На фиг. 1 представлена хроматограмма B3APi, полученного методом стандартного синтеза; на фиг. 2 представлена хроматограмма B3APi, полученного методом асимметрического синтеза согласно изобретению; на фиг. 3 представлена хроматограмма С 3 АР, полученного методом асимметрического синтеза согласно изобретению; на фиг. 4 представлена хроматограмма МРРА, полученного методом асимметрического синтеза согласно изобретению; на фиг. 5 представлена хроматограмма В 3 АР, полученного методом асимметрического синтеза согласно изобретению. Примеры Пример 1. Идентификация (R)-селективных -трансаминаз. Аминокислотную последовательность (R)-селективной -ТА Mycobacterium aurum, представленную в ЕР 1038953 A1 (SEQ ID NO: 5) и аминокислотную последовательность (R)-селективной -ТА Arthrobacter sp., представленную в ЕР 0987332 Al (SEQ ID NO: 7), использовали в качестве запрашиваемой последовательности биомолекулы. В данном примере используемым банком биомолекул является опубликованная база данных белков NCBI (http://.ncbi.nlm.nih.gov/pubmed (13 июля 2009 г.). Используя программу поиска BLAST с аминокислотной последовательностью -ТА из М. aurum или Arthrobacter sp. (SEQ ID NO: 5 или 7) в качестве запрашиваемой и используя стандартные параметры(оценочная матрица BLOSUM62, длина "слова": 3, "штрафы" на пробел: пробел на наличие - 11, пробел на удлинение - 1), идентифицировали группу из 100 различных аминокислотных последовательностей,из различных организмов, которые все были минимум на 30% идентичны запрашиваемой последовательности. Для BLAST "неизбыточные белковые последовательности (nr)"ROGRAMS=blastpPAGETYPE=BlastSearchSHOWDEFAULTS =onLINKLOC=blasthome) использовали 13 июля 2009 г. В этой первой группе из 100 различных аминокислотных последовательностей, представляющих собой первые последовательности биомолекул-мишеней, путем поиска мотивов последовательностей были выявлены и идентифицированы последовательности, которые не содержат мотивов последовательностей c1), с 2) или с 3) и которые содержат мотивы последовательностей с 4)-с 6). Была идентифицирована 21 ORF, которые перечислены ниже в табл. 1.vanbaalenii и Mesorhizobium loti. 2.1. -TA из Mycobacterium vanbaalenii, номер 9 в табл. 1 (SEQ ID NO: 49 и 50), из Aspergillus terreus, номер 1 в табл. 1 (SEQ ID NO: 3 и 4) и Mesorhizobium loti, номер 11 в табл. 1 (SEQ ID NO: 1 и 2) были получены и использованы в форме, адаптированной с точки зрения встречаемости кодонов (для Е.coli), для экспрессии указанных трансаминаз в Escherichia coli. Трансаминаза из Mycobacterium vanbaalenii далее обозначается Mva-TA, трансаминаза из Aspergillus terreus обозначается Ate-TA и трансаминаза из Mesorhizobium loti обозначается Mlo-TA. 2.2. Анализ на ацетофенон. В анализе на ацетофенон, проводимом с использованием 2,5 мМ амина и 2,5 мМ пирувата при рН 7,5 и 30 С, Mva-TA и Ate-TA превращались в (R)МВА по меньшей мере в 100 раз быстрее, чем в (S)энантиомер. Анализ: мониторинг увеличения абсорбции ацетофенона, образующегося в процессе реакции, проводили на длине волны 245 нм.Mlo-TA не превращался ни в (R)МВА, ни в (S)МВА. Затем тестировали другие амины в присутствии пирувата или -кетоглутарата, используемых в качестве аминоакцепторов (10 мМ амин, 10 мМ пируват, 0,1 мМ PLP, фосфатный буфер рН 7,5, инкубацию в течение 24 ч при 30 С, анализ методом тонкослойной хроматографии). Превращение наблюдалось для 2-аминогептана, 2-аминопентана, 1,3-диметилбутиламина и 4 фенил-2-аминобутана. Также детектировалось минимальное превращение изопропиламина. Какого-либо превращения с использованием других аминодоноров, таких как D-аланин, L-валин, аминобутират, этиламин, бензиламин, путресцин, 2-амино-3-метилбутан и 3,3-диметил-2-аминобутан, не наблюдалось. Таким образом, было обнаружено, что все три белка представляют собой -ТА. При использовании (R)- и (S)-2-аминогексана в качестве субстрата заметное превращение наблюдалось только для (R)-энантиомера. Таким образом, Mlo-ТА также представляет собой (R)-селективную ТА. Какой-либо DATA-активности (активность D-аминокислота-трансаминазы или ВСАТ-активности(активности аминотрансферазы аминокислот с разветвленной цепью) для всех трех белков не наблюдалось. 2.3. Анализ проводимости 1-N-boc-3-аминопирролидин (В 3 АР), 1-N-boc-3-аминопиперидин.(B3APi) и 1-Cbz-3-аминопиперидин (C3APi) также использовали в качестве субстратов для определения относительной активности этих веществ в отношении субстрата-модели -МВА. Ход реакции амина и пирувата (при рН 7,5 оба субстрата заряжены) в аланин и кетон (кетон не заряжен, аланин представляет собой цвиттерионное соединение и не влияет на проводимость) подвергали мониторингу кинетики проводимости, что позволяет вычислить степень превращения. Перед началом реакции проводили вычисление различных уровней превращения в зависимости от различных концентраций аланина, пирувата, кетона и амина. Снижение проводимости при превращении 1 мМ -МВА, В 3 АР, B3APi и C3APi составляло 44, 50,48,5 и 49,3 мкСм. А дополнение к трем рекомбинантно экспрессированным трансаминазам тестировали коммерчески доступную (R)-селективную АТА-117, поставляемую Codexis, в следующих реакционных условиях: 50 мМ буфер CHES, рН 7,5, рН корректировали с использованием BIS-TRIS (=бис-(2-гидроксиэтил)аминотрис-(гидроксиметил)метан), 0,1 мМ PLP, 5-6 мМ амина и пирувата, при температуре реакции 25 С. Превращение субстратов было показано для Ate-TA. Относительная активность в отношении (R)-МВА составляла 2% для (R)-В 3 АР и (R,S)-C3Api и 1 % для (R)-B3APi. 2.4. Определение энантиоселективности при асимметрическом синтезе аминов (R)-селективных ТА, происходящих из Aspergillus terreus и Mesorhizobium loti. Независимый метод показал, что Mlo и Ate-TA являются (R)-селективными и обладают способностью превращать субстраты в требуемые продукты с превосходной энантиоселективностью. Окончательное подтверждение (R)-энантиоселективности обоих трансаминаз было проведено при асимметрическом синтезе различных аминов с последующим определением их оптической чистоты. В качестве аминодонора использовали 100-кратный избыток аланина. Превращение оценивали не точно, а путем приблизительной оценки. В других экспериментах могут быть проведены различные методы с увеличением степени превращения (PDC). Для обеих трансаминаз была получена высокая или очень высокая энантиоселективность, за исключением C3APi, для которой была получена только очень низкая энантиоселективность. Таблица 2 Методы. Трансаминазы экспрессировали в Е. coli BL21. Культуральную среду (50 мл среды LD-Amp) индуцировали 0,2% рамнозой, и когда оптическая плотность (OD) составила 0,5, среду культивировали в течение 12 ч при 25 С. Затем клетки промывали фосфатно-натриевым буфером, центрифугировали, суспендировали в 1 мл фосфатно-натриевого буфера(рН 7,5) и замораживали в виде 200 мкл аликвот. Для биокатализа ко всем аликвотам добавляли 5 мкл 500 мМ раствора кетона в ДМСО, 22 мг Dаланина и 10 мкл 10 мМ PLP. Реакционную смесь доводили до общего объема 500 мкл добавлением 50 мМ фосфатно-натриевого буфера (рН 7,5). Реакционные смеси инкубировали в течение ночи при 25 С и 500 об/мин. Для определения энантиомерного избытка посредством СЕ к 400 мкл образца добавляли 200 мкл 1 н. NaOH, экстрагировали 400 мкл дихлорметана и затем органическую фазу отделяли. После этого органическую фазу экстрагировали 100 мкл 5 мМ триэтиламмоний-фосфатного буфера (рН 3). Затем полученную водную фазу вводили в CE. Протокол разделения. Оборудование: капиллярный электрофорез: колонка СЕ-Capillare: высота 30 см, внутренний диаметр 50 мкм, температура 15 С. Промывка: триэтиламмоний-фосфатным буфером рН 3, 1 мин, 30 фунт/кв.дюйм. Промывка: 5% в высокой степени сульфированным циклодекстрином (HSCD или HSCD), 1 мин,10 фунт/кв.дюйм. Впрыск: 5-10 с, 1 фунт/кв.дюйм. Пропитка водой. Разделение: напряжение 10 или 15 кВ. Детекция: МРРА, С 3 АР и C3APi на 200 нм; В 3 АР, B3APi на 190 нм. Пример 4. Получение и анализ всех трансаминаз, идентифицированных в табл. 1. 4.1. Экспрессия и очистка трансаминаз. Оптимизированные по кодонам открытые рамки считывания, кодирующие белки, представленные в табл. 1 под номерами 1, 2, 3, 4, 8, 9, 11, 13, 14, 15, 17, 18 и 21, встраивали в вектор pGASTON между сайтами рестрикции NdeI и BamHI с применением стратегии независимого от лигирования клонирования. Оптимизированные по кодонам ORF, кодирующие все остальные белки, по порядку субклонировали в рЕТ-22b. Трансформированные штаммы BL21 (DE3) Е. coli культивировали в 400 мл среды LB, в которую добавляли ампициллин (100 мкг/мл). Сначала клетки инкубировали при 37 С на ротационном шейкере до тех пор, пока OD600 не достигала 0,7. Затем клетки индуцировали путем добавления 0,2% рамнозы (pGASTON) или 0,1 мМ IPTG (рЕТ-22b) соответственно, и за это время инкубации температура снижалась до 20 С. После индуцирования инкубацию продолжали еще 20 ч. Для оценки экспрессии через определенные интервалы времени после индуцирования брали аликвоты. Клеточный осадок (влажная масса 3 г) два раза промывали фосфатным буфером (рН 7,5, 50 мМ),содержащим 0,1 мМ PLP, при 4 С. После измельчения (на френч-прессе) клеточную суспензию центрифугировали (4000g, 30 мин) и полученный супернатант пропускали через 0,5-мкм фильтр и затем хроматографировали. Хроматографию осуществляли на устройстве AKTA Purifier (GE Healthcare). Отфильтрованный клеточный экстракт наносили на 5-мл колонку с сефарозой IMAC (Sepharose 6) FastFlow (GE Healthcare). Колонку промывали со скоростью потока 5 мл/мин 10 колоночными объемами 50 мМ фосфатного буфера, рН 7,5, содержащего 300 мМ NaCl, 0,1 мМ PLP и 30 мМ имидазола (во избежание неспецифического связывания), и фермент АТА элюировали 10 колоночными объемами фосфатного буфера (рН 7,5, 50 мМ), содержащего 300 мМ NaCl, 0,1 мМ PLP и 300 мМ имидазола (со скоростью потока 5 мл/мин). Ферментсодержащие фракции объединяли и обессоливали с помощью гельхроматографии с использованием 20 мМ трицинового буфера, рН 7,5, содержащего 0,01 мМ PLP. Очищенные ферменты хранили при 4 С. Количество каждого белка, очищенного приблизительно из 3 г клеток (сырой массы), приведено ниже в табл. 4. Таблица 4 4.2. Характеризация специфичности (R)-селективной -ТА к субстрату. Для определения активности в отношении -метилбензиламина (-МВА) при первоначальном скрининге экспрессированных белков проводили анализ с использованием ацетофенона, где раствор 2,5 мМ (R)- или (S)МВА и пирувата подвергали реакции в присутствии очищенного фермента и определяли корреляцию увеличения оптической плотности на 245 нм с образованием ацетофенона. Проводимость аминов, таких как 2-аминогексан, 4-фенил-2-аминобутан и 1-N-Boc-3-аминопирролидин, подвер- 16022764 гали мониторингу, используя следующий анализ на проводимость: раствор, содержащий 10 мМ амина и пирувата, подвергали реакции в присутствии очищенной аминтрансаминазы и определяли снижение проводимости в зависимости от степени превращения субстрата. Для исследования DATA- и ВСАТ-активности измеряли снижение уровня NADH на спектрофотометре при длине волны 340 нм, используя следующий анализ на связывание дегидрогеназы: раствор 5 мМ -кетоглутаровой кислоты и D-аланина подвергали реакции в присутствии очищенной трансаминазы, 1 ед./мл лактат-дегидрогеназы и 0,5 мМ NADH для измерения DATA-активности. Для измерения ВСАТ-активности использовали раствор, содержащий 5 мМ 3-метил-2-оксомасляной кислоты и Lглутамата, 10 мМ хлорида аммония, 1 ед/мл глутамат-дегидрогеназы и 0,5 мМ NADH. Все реакции проводили в 20 мМ трициновом буфере, рН 7,5, содержащем 0,01 мМ PLP. рН буфера корректировали добавлением 1,8-диазабицикло[5.4.0]ундек-7-ена. Результаты представлены ниже в табл. 5. Таблица 5 Специфические активности для различных субстратов Номера в первом столбце соответствуют номерам в табл. 1. Все измерения проводили, по меньшей мере, в дубликатах. Отклонение одного измерения от средней величины составляет 10%. а) Измерение не могло быть проведено, поскольку в процессе экспрессии выход белка был очень низким/белок был нестабильным в процессе очистки. 4.3. Асимметрический синтез (R)-аминов 1-4 (см. выше примечания к табл. 5) с -ТА из Aspergillusterreus, Mesorhizobium loti и Mycobacterium vanbaalenii. Асимметрический синтез осуществляли при 30 С в течение 24 ч в фосфатно-натриевом буфере (100 мМ, рН 7), содержащем моногидрат пиридоксил-5'-фосфата (PLP) (1 мМ) и NAD+(1 мМ) в 1,5-мл пробирках Эппендорфа. Реакционная смесь содержала 50 мМ кетон, L-аланин (5 экв., 250 мМ), лактат-дегидрогеназу, выделенную из коровьего сердца (90 ед.), глюкозу (150 мМ) и глюкозо-дегидрогеназу (15 ед.). -ТА, проис- 17022764 ходящую из Aspergillus terreus, Mesorhizobium loti и Mycobacterium vanbaalenii (номера 1, 11 и 9 в табл. 1), экспрессировали в Е. coli BL21, как описано выше, замораживали в виде аликвот и непосредственно использовали в качестве биокатализатора в виде цельных клеток без дополнительной очистки. Степень превращения измеряли путем детектирования образованных аминов (1, газовая хроматография (ГХ); 2-4,капиллярный электрофорез (КЭ. Хиральный анализ 2-4 осуществляли с помощью КЭ, как описано выше. Величину энантиомерного избытка (% э.и.) для 1 определяли с помощью ГХ. После экстракции амина этилацетатом проводили дериватизацию с образованием трифторацетамида путем добавления 20 кратного избытка ангидрида трифторуксусной кислоты. После продувки азотом для удаления избытка ангидрида и остаточной трифторуксусной кислоты дериватизированное соединение растворяли в этилацетате (50 мкл) и исходное вещество отделяли на системе Shimadzu GC14A, которая была снабжена колонкой с гептакис(2,3-ди-O-ацетил-6-O-трет-бутилдиметилсилил)циклодекстрином (25 м 0,25 мм). Время удерживания составляло 16,0 мин S)-1) и 16,2 мин S)-2) при градиенте температуры в печи 80 С/10 мин//20 С//180 С/10 мин. Результаты представлены ниже в табл. 6. Таблица 6a) Условия реакции: 50 мМ кетон, 250 мМ D-аланин, 100 мМ фосфатно-натриевый буфер, рН 7,0, 1 мМ PLP, 1 мМ NADH. Побочный продукт пируват, образовавшийся в процессе реакции, удаляли с использованием лактат-гидрогеназы (LDH). Для подачи кофактора на повторный цикл использовали глюкозо-дегидрогеназуb) Превращение не оптимизировали. Отклонение одного измерения от средней величины не превышало 10%. Соединение 4 превращалось только под действиемa) обеспечение по меньшей мере одной последовательности биомолекулы для поиска по меньшей мере одной трансаминазы или лиазы и по меньшей мере одного банка биомолекул;b) поиск в банке биомолекул с помощью указанной последовательности биомолекулы для поиска в целях идентификации группы первых последовательностей биомолекул-мишеней, где указанные первые последовательности биомолекул-мишеней по меньшей мере на 20% идентичны указанной последовательности биомолекулы для поиска, как рассчитано на уровне аминокислотной последовательности;c) отбор в группе первых последовательностей биомолекул-мишеней группы вторых последовательностей биомолекул-мишеней, которые не содержат на уровне аминокислотной последовательности по меньшей мере один из следующих мотивов аминокислотных последовательностей (c1)-(c3):c1) в положениях 95-97 аминокислотную последовательность Tyr-Xa1-Xa2, где Xa1 представляет собой аминокислоту Ile, Val, Leu, Met, Phe и Ха 2 представляет собой аминокислоту Arg или Lys, или с 2) в положениях 97-99 аминокислотную последовательность Tyr-Xaa-Gln, где Хаа представляет собой аминокислоту, и в области от положения 105 до положения 111 аминокислотную последовательность Arg-Xaa-Ха 3, где Ха 3 представляет собой аминокислоту, предпочтительно His, или с 3) в положении 38 Thr, в положении 97 Lys и в положениях 107-109 аминокислотную последовательность Arg-Xa4-Xa5, где Ха 4 представляет собой аминокислоту, предпочтительно Gly, и Ха 5 представляет собой аминокислоту, предпочтительно Tyr,и которые содержат: с 4) в положении 95 аминокислоту, не являющуюся Tyr, Arg, Lys, или в положении 95 аминокислотуTyr, но в положении 97 аминокислоту, не являющуюся Arg или Lys, и с 5) в положении 40 аминокислоту, не являющуюся Lys или Arg, и с 6) в области от положения 161 до положения 165 аминокислоту Lys,для идентификации группы вторых последовательностей биомолекул-мишеней, иd) обеспечение биомолекулы, имеющей вторую последовательность биомолекулы-мишени, идентифицированную на стадии (с), и представляющей собой или кодирующей, по меньшей мере частично,белок, обладающий активностью (R)-селективной -трансаминазы. 2. Способ скрининга (R)-селективной -трансаминазы, включающий стадии (а)-(с) по п.1. 3. Способ по п.1 или 2, где указанная биомолекула представляет собой белок и последовательность биомолекулы представляет собой аминокислотную последовательность. 4. Способ по п.1 или 2, где биомолекула представляет собой молекулу ДНК и последовательность биомолекулы представляет собой последовательность ДНК. 5. Способ по любому из предшествующих пунктов, где банк биомолекул представляет собой базу данных биомолекул и поиск в указанной базе данных биомолекул на стадии (b) осуществляют с помощью программы выравнивания последовательностей биомолекул, в частности BLAST. 6. Способ по любому из пп.1, 3-5, где молекулу белка или ДНК, обеспеченную на стадии (d), получают синтезом de novo. 7. Способ по любому из пп.1, 2 и 4, где банк биомолекул представляет собой банк генов и поиск в указанном банке генов на стадии (b) осуществляют с использованием молекулы с последовательностью ДНК для поиска. 8. Способ по п.7, где на стадии (с) используют праймеры с ДНК-последовательностью для отбора группы вторых последовательностей биомолекул. 9. Способ по любому из предшествующих пунктов, где указанная последовательность биомолекулы для поиска представляет собой последовательность, представляющую всю функциональную (R)-селективную -трансаминазу или ее часть. 10. (R)-селективная -трансаминаза, которая может быть получена по любому из способов по пп.1 и 3-9, предпочтительно выбранная из группы последовательностей биомолекул, состоящей из SEQ IDNO: 1-4. 11. Способ определения характеристик трансаминазы, в частности для характеризации свойств трансаминазы, включающий следующие стадии:ii) осуществление реакции аминоакцептора и аминодонора с трансаминазой в реакционной среде и тем самымiii) определение проводимости реакционной среды в первом наборе реакционных условий;iv) после проведения стадии iii) определение проводимости реакционной среды во втором наборе реакционных условий с получением по меньшей мере двух величин проводимости, отражающих свойства трансаминазы. 12. Способ по п.11, где проводимость реакционной среды определяют по меньшей мере три раза, по меньшей мере, в третьем наборе реакционных условий. 13. Способ по п.11 или 12, где наборы реакционных условий являются одинаковыми. 14. Способ по любому из пп.11-13, где реакционная среда представляет собой буфер с низкой проводимостью. 15. Способ по любому из пп.11 или 14, где заряженный аминоакцептор представляет собой пируват. 16. Способ получения (R)-селективной -трансаминазы по любому из пп.1 и 3-9, где после получения (R)-селективной -трансаминазы осуществляют способ определения характеристик указанной (R)селективной -трансаминазы по любому из пп.11-15. 17. Способ скрининга (R)-селективной -трансаминазы по любому из пп.2-9, где после скрининга(R)-селективной -трансаминазы осуществляют способ определения характеристик указанной (R)селективной -трансаминазы по любому из пп.11-15. 18. Способ получения оптически активного хирального амина, включающий осуществление реакции по меньшей мере одного аминоакцептора и по меньшей мере одного аминодонора с (R)-селективной-трансаминазой по п.10 и получение оптически активного хирального амина.
МПК / Метки
МПК: C12N 9/88, G06F 19/00, C12N 9/10
Метки: способ, получения, r)-специфической, w-трансаминазы, идентификации
Код ссылки
<a href="https://eas.patents.su/30-22764-sposob-identifikacii-i-polucheniya-r-specificheskojj-w-transaminazy.html" rel="bookmark" title="База патентов Евразийского Союза">Способ идентификации и получения (r)-специфической w-трансаминазы</a>
Способ получения оптически активного n-защищенного 3-аминопирролидина или оптически активного n-защищенного 3-аминопиперидина и соответствующих кетонов путем оптического разделения рацемических смесей аминов с использованием бактериальной омега-трансаминазы

Номер патента: 16471
Опубликовано: 30.05.2012
Авторы: Хене Маттиас, Робинс Карен, Борншойер Уве
МПК: C12P 41/00, C12P 17/12, C12P 17/10...
Метки: омега-трансаминазы, n-защищенного, активного, кетонов, способ, аминов, рацемических, разделения, смесей, путем, бактериальной, оптического, использованием, соответствующих, получения, 3-аминопиперидина, 3-аминопирролидина, оптически
Формула / Реферат:
1. Способ получения оптически активного амина, включающий:(a) обеспечение акцептора аминогруппы и рацемической смеси 3-аминопирролидина (3АР) или 3-аминопиперидина (3APi), в каждом из которых у атома азота в кольце имеется защитная группа;(b) осуществление взаимодействия акцептора аминогруппы и рацемической смеси соответствующего амина с (R)- или (S)-селективной трансаминазой;(c) получение оптически активного амина, аминного продукта и...
Рекомбинантный или мутантный полинуклеотид для придания растениям толерантности к гербицидам, ингибирующим ацетогидроксикислотную синтазу (ahas) (варианты), растение, не встречающееся в природе, и его семена, способ борьбы с сорняками, способ получения трансгенного растения, толерантного к соединениям, ингибирующим ahas, способ идентификации или отбора клетки растения, ткани растения или растения или части растения, способ предотвращения нежелательного вегетативного роста и способ идентификации растения

Номер патента: 21541
Опубликовано: 30.07.2015
Авторы: Мэкэлвер Джон А., Зинг Биджай
МПК: A01H 5/10, C12N 15/82, A01H 5/00...
Метки: нежелательного, мутантный, ткани, предотвращения, способ, растениям, ahas, рекомбинантный, трансгенного, соединениям, идентификации, борьбы, получения, природе, ингибирующим, отбора, варианты, семена, синтазу, встречающееся, толерантного, сорняками, растения, толерантности, придания, гербицидам, полинуклеотид, ацетогидроксикислотную, части, клетки, роста, вегетативного, растение
Формула / Реферат:
1. Рекомбинантный или мутантный полинуклеотид для придания растениям толерантности к гербицидам, ингибирующим ацетогидроксикислотную синтазу (AHAS), кодирующий полипептид большой субъединицы ацетогидроксикислотной синтазы (AHASL) с аминокислотными замещениями, выбранный из:a) полипептида, имеющего валин-, треонин-, глутамин-, цистеин- или метионин-замещение в положении, соответствующем положению 122 SEQ ID NO: 1 или положению 90 SEQ ID NO: 2, и...
Композиция для окрашивания in vivo, способ ее получения и способы ее применения для идентификации диспластической ткани

Номер патента: 2127
Опубликовано: 24.12.2001
Автор: Беркетт Дуглас Д.
МПК: C07D 279/20, A61K 49/00, A61K 31/54...
Метки: получения, окрашивания, способы, способ, идентификации, применения, диспластической, vivo, композиция, ткани
Формула / Реферат:
1. Композиция продукта толуидинового синего 0 (ТВО), отличающаяся тем, что соотношение суммарных площадей пиков ВЭЖХ при 254 нм, представляющих конформационные изомеры толуидинового синего 0, и суммарных площадей пиков, представляющих N-деметилированные производные указанных изомеров, составляет, по крайней мере, приблизительно 6:1. 2. Композиция продукта толуидинового синего 0 (ТВО), отличающаяся тем, что соотношение суммарных площадей пиков...
Способ идентификации низкомолекулярных веществ, имитирующих эпитопы млекопитающих, и способ получения специфических анти-идиотипических моноклональных антител

Номер патента: 7182
Опубликовано: 25.08.2006
Авторы: Кауфманн Петер, Франк Ханс-Георг
МПК: C07K 16/28, C07K 16/42
Метки: имитирующих, моноклональных, низкомолекулярных, эпитопы, млекопитающих, идентификации, получения, веществ, специфических, антител, анти-идиотипических, способ
Формула / Реферат:
1. Способ идентификации низкомолекулярных веществ, имитирующих эпитопы млекопитающих, включающий следующие стадии: a) выделение структур, содержащих эпитопы, которые участвуют в синцитиальном слиянии клетка-клетка трофобласта для получения эпитопного препарата; b) иммунизация немлекопитающих с помощью эпитопного препарата для получения иммунного ответа; c) иммортализация иммунного ответа для получения библиотеки антител; d) селекция антител в...
Способ идентификации пользователя, электронная идентификационная система и устройство для биометрической идентификации личности по характерным параметрам кисти руки пользователя

Номер патента: 8534
Опубликовано: 29.06.2007
Авторы: Тахири Азар Мамед Оглы, Зиновьев Виктор Валентинович
Метки: пользователя, устройство, идентификации, кисти, руки, личности, идентификационная, характерным, система, биометрической, способ, электронная, параметрам
Формула / Реферат:
1. Способ идентификации пользователя, использующий электронную систему идентификации, включающую по крайней мере первое и второе электронное устройство идентификации, связанные по крайней мере одной телекоммуникационной линией, при этом способ включает следующие операции: формирование базы данных пользователя в каждом электронном устройстве идентификации и идентификация пользователя, в котором операция формирования базы данных пользователя...
Предыдущий патент: Переработка углеводородного газа
Следующий патент: Фармацевтическая композиция
Случайный патент: Материал покрытия для излучающих поверхностей, вырабатывающих электромагнитные волны, и способ его получения