Способ придания растению сахарной свеклы резистентности к вирусу некротического пожелтения жилок свеклы, используемый для этого днк-вектор и растение и его части, резистентные к данному вирусу
Номер патента: 6701
Опубликовано: 24.02.2006
Авторы: Жонар Жерар, Гиллей Юбер, Ван Дюн Корнелис Мария Петрус, Ричардз Кеннет










Формула / Реферат
1. Способ придания растению сахарной свеклы резистентности к вирусу некротического пожелтения жилок свеклы (BNYVV), включающий следующие стадии:
(a) получение ДНК-фрагмента, который состоит по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 вируса некротического пожелтения жилок свеклы (BNYVV), причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы;
(b) введение указанного ДНК-фрагмента, функционально связанного с промотором, который является активным в растениях сахарной свеклы, в клетку растения сахарной свеклы с целью получения трансформированной клетки растения сахарной свеклы и
(c) регенерацию трансгенного растения сахарной свеклы из трансформированной клетки растения сахарной свеклы.
2. Способ по п.1, отличающийся тем, что указанный фрагмент имеет последовательность, которая соответствует нуклеотидам 153-3258 РНК 1 BNYVV.
3. Способ по любому из пп.1 или 2, отличающийся тем, что указанный фрагмент вводят в клетку в составе ДНК-вектора, содержащего транскрипционную и трансляционную регуляторные последовательности, функционально присоединенные к указанному фрагменту.
4. ДНК-вектор, содержащий фрагмент, который состоит по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 (BNYVV), причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы и транскрипционную и трансляционную регуляторные последовательности, функционально присоединенные к указанному фрагменту.
5. ДНК-вектор по п.4, отличающийся тем, что указанный фрагмент имеет последовательность, которая соответствует нуклеотидам 153-3258 РНК 1 BNYVV.
6. Применение ДНК-вектора по любому из пп. 4 или 5 для трансформации клетки растения сахарной свеклы с целью получения растений, которые являются резистентными к BNYVV.
7. Клетка растения сахарной свеклы, обладающего резистентностью к BNYVV, которая содержит в своем геноме ДНК-фрагмент, состоящий по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 BNYVV, причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы.
8. Клетка растения по п.7, отличающаяся тем, что указанный фрагмент имеет последовательность, которая соответствует нуклеотидам 153-3258 РНК 1 BNYVV.
9. Применение клетки растения по любому из пп.7-8 для регенерации из нее растения сахарной свеклы, обладающего резистентностью к BNYVV.
10. Растение сахарной свеклы, регенерированное из клетки по любому из пп.7-8.
11. Потомство растения сахарной свеклы по п.10.
12. Семена растения сахарной свеклы по п.10 или потомства по п.11.
13. Вегетативно репродуцируемые структуры, такие как каллюс, почки, зародыши, происходящие от растения по п.10 или от потомства по п.11.
14. Применение семян по п.12 для регенерации из них растения сахарной свеклы, которое является резистентным к BNYVV.
15. Применение вегетативно репродуцируемых структур по п.13 для регенерации из них растения сахарной свеклы, которое является резистентным к BNYVV.
Текст
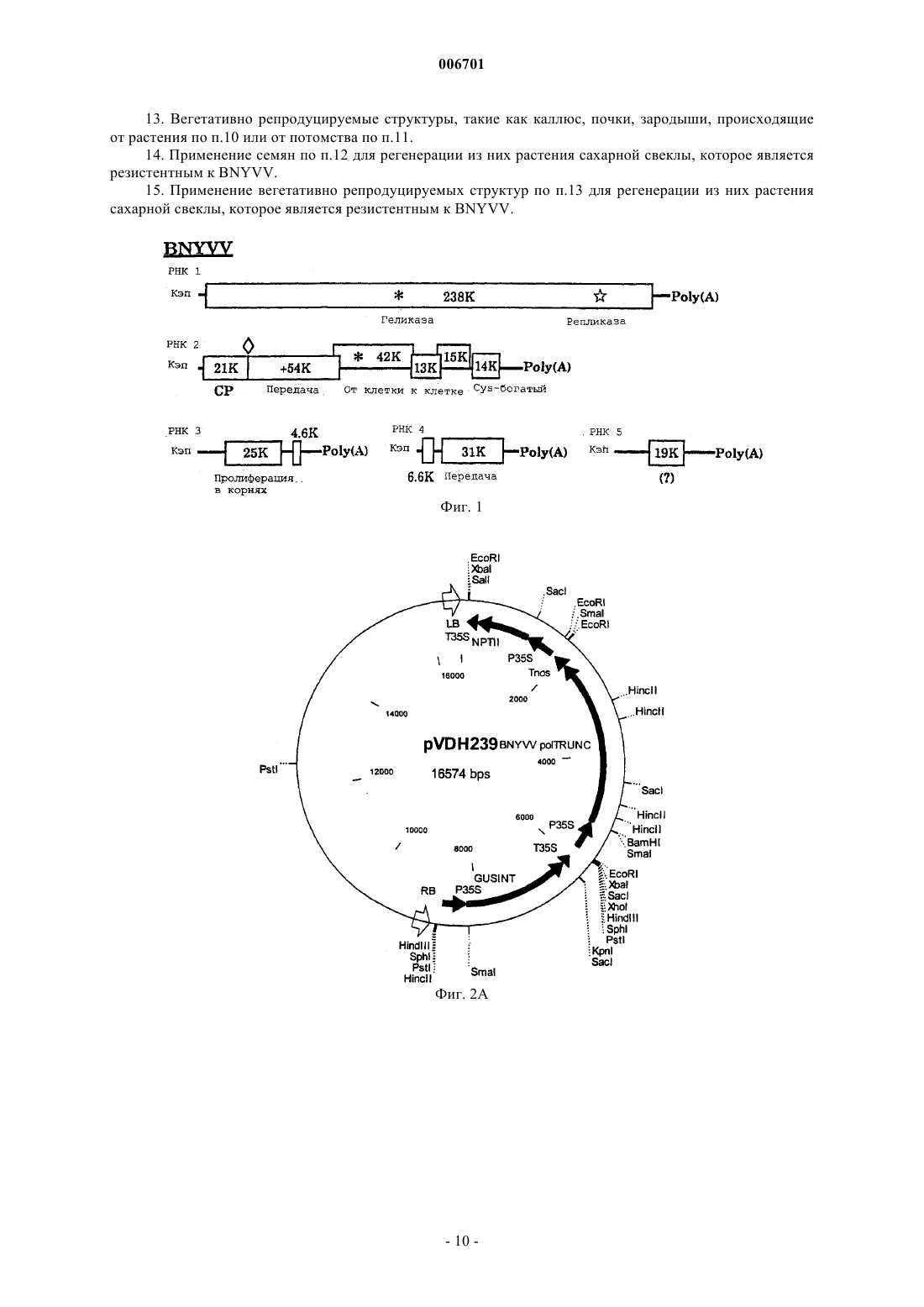
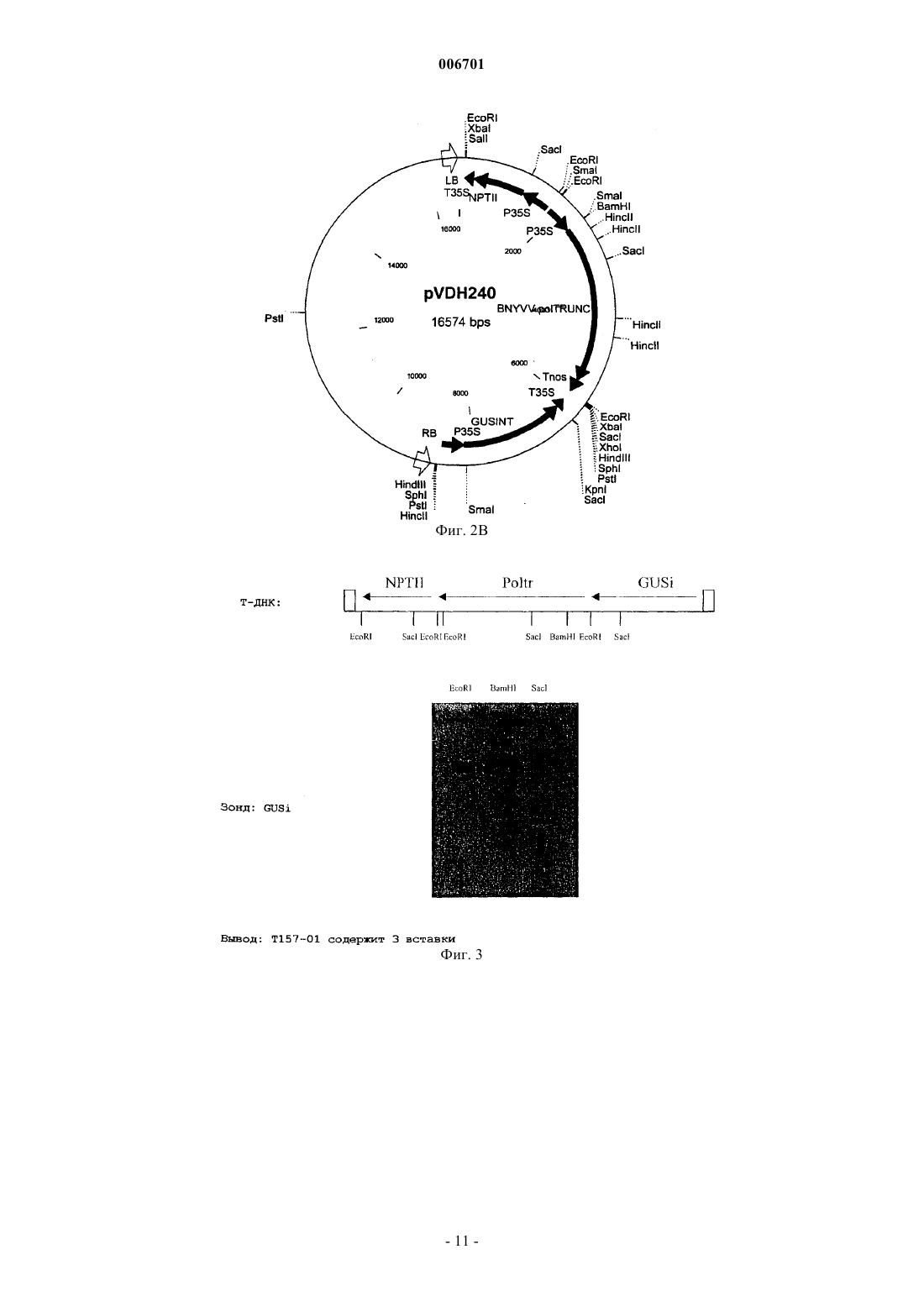
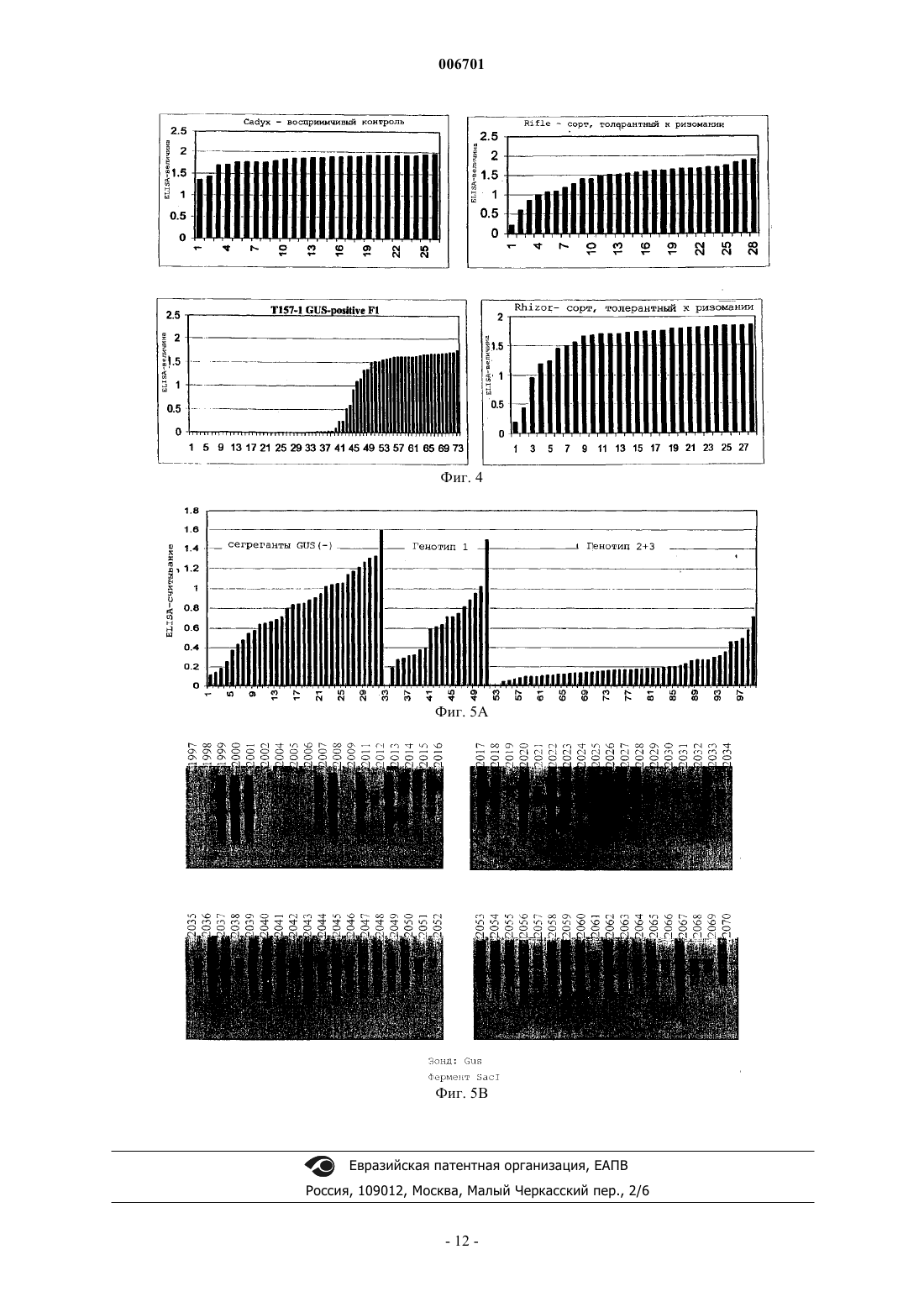
006701 Настоящее изобретение относится к способу придания растению сахарной свеклы резистентности к вирусу некротического пожелтения жилок свеклы (BNYVV - beet necrotic yellow vein virus). Также настоящее изобретение относится к ДНК-вектору, содержащему фрагмент, который состоит по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 (BNYVV), причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы и транскрипционную и трансляционную регуляторные последовательности, функционально присоединенные к указанному фрагменту, и к применению ДНК-вектора для трансформации клетки растения сахарной свеклы с целью получения растений, которые являются резистентными к BNYVV. Настоящее изобретение, таким образом, относится также к клетке растения сахарной свеклы, обладающей резистентностью к BNYVV, которая содержит в своем геноме такой ДНК-фрагмент, и к применению клетки растения для регенерации из нее растения сахарной свеклы, обладающего резистентностью к BNYVV. Настоящее изобретение относится к растению сахарной свеклы, регенерированному из такой клетки. Настоящее изобретение также относится к потомству растения сахарной свеклы и к семенам растения сахарной свеклы или потомства. Настоящее изобретение относится к вегетативно репродуцируемым структурам, таким как каллюс, почки, зародыши, происходящие от такого растения или потомства. Настоящее изобретение относится к применению вышеуказанных семян для регенерации из них растения сахарной свеклы, которое является резистентным к BNYVV, к применению вегетативно репродуцируемых структур для регенерации из них растения сахарной свеклы, которое является резистентным к BNYVV. Вирусы растений представляют собой серьезную угрозу для многих основных сельскохозяйственных культур. Так, например, вирус BNYVV, который передается почвенными грибками плазмодиофоромицетамми Polymuxa betae, является возбудителем болезни, известной, как ризомания, которая поражает экономически ценную культуру сахарной свеклы (Beta vulgaris). Это заболевание было впервые обнаружено в Италии в 1950-х годах, и с тех пор оно стало представлять серьезную угрозу для культур сахарной свеклы в большинстве регионов мира, где возделывается эта культура. В полевых условиях поражение вирусом BNYVV, обычно, ограничивается подземными частями растения. Инфицирование корней сахарной свеклы проявляется в повышенной пролиферации и некрозе боковых корней, в снижении общей массы главного корня и в последующем снижении выхода сахара. Гибель растения может быть также вызвана инфекцией, особенно, если эта культура инфицирована в начале сельскохозяйственного сезона. В некоторых случаях этот вирус поражает листья растения, где он может вызывать инвазивные жилкоассоциированные хлоротические и некротические поражения.BNYVV является типичным членом семейства бенивирусов, которые представляют собой передающиеся грибками палочковидные вирусы, обладающие составным геномом, состоящим из двух или более компонентов одноцепочечной (оц) РНК. Были идентифицированы четыре молекулы плюс-оцРНК вируса BNYVV, которые были обозначены в порядке уменьшения их размера (РНК 1 - 6,8 т.п.н.; РНК 2 4,7 т.п.н.; РНК 3 - 1,8 т.п.н.; РНК 4 - 1,5 т.п.н.). Некоторые изоляты, полученные в Японии, также содержат пятый РНК-компонент (РНК 5 - 1,45 т.п.н.). Все РНК были клонированы и секвенированы. Генетическая карта вирусных РНК представлена на фиг. 1. Все пять РНК имеют последовательность из полиадениловой кислоты, а РНК 1-4 на 3'-конце имеют кэп-структуры на своих 5'-концах. РНК 1 и 2 кодируют основные независимые от хозяина функции "домашнего хозяйства", а более мелкие РНК конкретно участвуют в природных процессах инфицирования,включая опосредованное вектором инфицирование корней сахарной свеклы, пролиферацию в корневой системе и индуцирование симптомов ризомании. Так, например, было показано, что РНК 1 кодирует вирусную РНК-полимеразную активность, а РНК 2 кодирует 21 кДа-белок вирусного покрова. 3'Проксимальная половина РНК 2 несет группу из трех следующих друг за другом незначительно перекрывающихся вирусных генов, известных как тройной блок генов (TGB3), который имеет близкое сходство с кластером из трех генов в других палочковидных вирусах растений и который участвует в переносе вируса от клетки к клетке. РНК 3 ассоциируется с массивной пролиферацией мелких корешков сахарной свеклы и облегчает распространение вируса в ткани корней, а РНК 4 повышает эффективность передачи вируса грибами. Передаваемые грибами вирусы, такие как BNYVV, могут сохраняться в находящихся в состоянии покоя спорах, присутствующих в почве, в течение нескольких лет после заражения земель. Так как в настоящее время не существует эффективных химических или физических методов элиминации вируса как в растениях, так и в почве, то единственной возможностью для каждого фермера, культивирующего сахарную свеклу, является использование генетически резистентных сортов. Некоторые компании получают ряд резистентных, и даже частично резистентных сортов благодаря переносу генов устойчивости дикого типа в коммерчески ценные сорта посредством селекции. Однако этот процесс является очень длительным и трудоемким, и для того чтобы получить полезные резистентные растения, обычно, требуется очень много времени. Кроме того, у толерантных или частично резистентных растений при высоком давлении болезни, будут развиваться симптомы болезни, поскольку уровень резистентности является слишком низким. Другая проблема, связанная с толерантностью или лишь с частичной резистентностью растений, заключается в том, что популяции вируса продолжают расти, что приводит к возможному возникновению штамма BNYVV, который может разрушать гены резистентности/толерантности.-1 006701 Быстрый прогресс в области генной инженерии растений привел к разработке новых стратегий придания растениям генетической резистентности к вирусам. Резистентность к вирусным болезням, сообщаемая посредством введения фрагментов последовательностей вирусного генома, является новой отправной точкой для достижения резистентности. Вирусную последовательность (конструкцию) вводят в растение путем использования комбинированных методов с применением соответствующих клеточных или тканевых культур и системы доставки ДНК, такой как хорошо известная система трансформацииAgrobacterium tumefaciens или путем прямого переноса ДНК, опосредованного химическими соединениями, такими как полиэтиленгликоль (ПЭГ). Известные методы индуцирования механизмов защиты растений от патогенов описаны, например, в ЕР 9687106.0, который относится к способу индуцирования резистентности к вирусу, содержащему последовательность TGB3. В этой публикации описано, что резистентность к BNYVV может быть индуцирована методом, предусматривающим трансформацию клеток растения ДНК-конструкцией, соответствующей фрагменту последовательности геномной или субгеномной РНК 2 BNYVV. Однако недостаток этого метода заключается в том, что в этом случае достигается лишь толерантность, а не резистентность. У толерантных растений-хозяев еще может происходить репликация вируса растений на нормальном уровне, и эти растения обнаруживают небольшие видимые признаки инфицирования или эти признаки отсутствуют, тогда как у резистентных хозяев, репликация происходит на низком уровне или вообще отсутствует. В заявке WO 93/25068 резистентность к вирусу у растений индуцируют путем трансформации растения репликазной частью (РНК 1) генома растительного вируса. Эта публикация не относится ни к сахарной свекле, ни к BNYVV. Известно, что сахарная свекла является трудным объектом для генной инженерии, что осложняет успешное индуцирование у него резистентности к BNYVV. Однако если учесть степень и последствия вирусной болезни, вызываемой BNYVV, то особый интерес представляет улучшение источников генетической резистентности растений сахарной свеклы. Поэтому целью настоящего изобретения является обеспечение средств для получения растений сахарной свеклы, которые обладают резистентностью к BNYVV. Предпочтительно, если они обладают полной резистентностью или иммунитетом, сообщаемым путем комбинирования различных методов и путем использования методов, которые не позволяют вирусам реплицироваться. Эта цель достигается благодаря настоящему изобретению путем разработки способа придания растению сахарной свеклы резистентности к вирусу некротического пожелтения жилок свеклы (BNYVV),включающего следующие стадии:(a) получение ДНК-фрагмента, который состоит по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 вируса некротического пожелтения жилок свеклы (BNYVV), причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы;(b) введение указанного ДНК-фрагмента, функционально связанного с промотором, который является активным в растениях сахарной свеклы, в клетку растения сахарной свеклы с целью получения трансформированной клетки растения сахарной свеклы; и(c) регенерацию трансгенного растения сахарной свеклы из трансформированной клетки растения сахарной свеклы. Таким образом, получают растения сахарной свеклы, которые обладают стабильной резистентностью и в которых указанный вирус не реплицируется. Такой трансформант является уникальным аспектом настоящего изобретения и нигде ранее не был описан. В соответствии с настоящим изобретением, существенная гомология последовательности данного фрагмента представляет собой гомологию по крайней мере на 70%, предпочтительно по крайней мере на 80%, более предпочтительно по крайней мере на 90%, а наиболее предпочтительно по крайней мере на 95%. Термин "существенная гомология последовательности" данного фрагмента используется для обозначения нуклеотидных последовательностей, которые при их сопоставлении имеют аналогичные (идентичные или консервативно замененные) нуклеотиды в аналогичных положениях или областях. Так, например, две нуклеотидные последовательности по крайней мере с 85%-ной гомологией имеют по крайней мере на 85% гомологичные (идентичные или консервативно замененные) нуклеотиды в аналогичных положениях, что выявляется при их сопоставлении с введением оптимальной поправки до 3 "брешей",при условии, что указанные "бреши" затрагивают не более чем 15 аминокислотных остатков. Степень сходства может быть определена методами, хорошо известными специалистам (см., например, WilburSpace". Comput. Appl. Biosci. 4:11-17 (1988. Одной из программ, которая может быть использована для определения степени сходства, является метод одной пары MegAlign Lipman-Pearson (с использованием параметров по умолчанию), который может быть получен из DNAstar Inc, 1228, Selfpark Street, Madison,Wisconsin, 53715, USA как часть системы Lasergene. Тест на гомологию последовательности основан на определении процента идентичности, который вычисляется методом Fast DB, исходя из следующих па-2 006701 раметров: "штраф" на расхождение - 1,0, штраф на "брешь" (1,00), штраф на размер "бреши" - 0,33 и штраф на присоединение - 30,0. В своем предпочтительном варианте настоящее изобретение относится к использованию различных фрагментов, имеющих нуклеиновокислотные последовательности, соответствующие указанной гомологии или полностью гомологичные нуклеотидам 153-3258, 169-539, 1226-1683, 2754-3192 или всем из 6746 нуклеотидов РНК 1. Настоящее изобретение также относится к ДНК, которая гибридизируется с ДНК настоящего изобретения и которая кодирует РНК 1. Такая гибридизация предпочтительно происходит в условиях низкой или высокой жесткости или в промежуточных условиях. В общих чертах, условия низкой жесткости могут быть определены как: 3 х SSC в интервале примерно от комнатной температуры до температуры примерно 65 С, а условия высокой жесткости могут быть определены как: 0,1 х SSC при температуре примерно 65 С. SSC обозначает буфер, состоящий из 0,15 М NaCl, 0,15 цитрата натрия. "3 х SSC" означает в три раза более высокую концентрацию, чем SSC, и т.п. Данный фрагмент может быть введен в клетку регенерируемого растения с помощью ДНК-вектора,содержащего данный фрагмент и транскрипционную и трансляционную регуляторную последовательности, функционально присоединенные к этому фрагменту, стандартными методами трансформации растений, такими как Agrobacterium-опосредованная трансформация клеток, введенных в ткани растения, такие как семядоли (Krens et al., Plant Science 116: 97-106; 1996), или опосредованное полиэтиленгликолем включение ДНК в одиночные клетки, такие как замыкающие клетки-протопласты (Hall et al., Nature Biotechnology 14: 1133-1138; 1996). ДНК-вектор, содержащий данный фрагмент, также является частью настоящего изобретения. Цели использования конструкций, полученных так, чтобы данная последовательность гена ингибировала или стимулировала экспрессию гена, абсолютно очевидны. Полная последовательность гена, находящегося под контролем промотора, который эффективно функционирует в данном растении, будет, в основном, экспрессировать повышенное количество генного продукта, что будет приводить к усилению действия продуцируемого таким образом белка. Иногда количество генного продукта снижается, и этот феномен называют "косуппрессией". Негативная регуляция этого гена может быть осуществлена несколькими методами. Это может быть достигнуто с использованием "доминантно-негативных" конструкций. Эти конструкции содержат специфический ДНК-связывающий домен, а также возможные домены димеризации, которые, однако, являются транскрипционно неактивными. Они "сидят" на промоторах целевых генов, и тем самым препятствуют связыванию эндогенного белка. Кроме того, снижение количества генного продукта может быть также достигнуто с использованием доминантно-негативной мутации, либо путем изменения на обратную ориентации генной последовательности по отношению к промотору так, чтобы он продуцировал тип генного продукта, называемого "антисмысловой" матричной РНК. ДНК-конструкция настоящего изобретения может быть "антисмысловой" конструкцией, генерирующей "антисмысловую" РНК, или "смысловой" конструкцией (кодирующей по крайней мере часть функционального белка), генерирующей "смысловую" РНК."Антисмысловая РНК" представляет собой РНК-последовательность, которая комплементарна последовательности оснований в соответствующей мРНК, то есть комплементарна в том смысле, что каждое основание (или множество оснований) в данной антисмысловой последовательности (при чтении в направлении 3'5') может спариваться с соответствующим основанием (G с С, А с U) в последовательности мРНК, читаемой в направлении 5'3'. Такая антисмысловая РНК может быть продуцирована в клетке при ее трансформации соответствующей ДНК-конструкцией, расположенной так, чтобы в результате генерировался транскрипт, у которого по крайней мере часть последовательности комплементарна по крайней мере части кодирующей цепи релевантного гена (или ДНК-последовательности, в основном,гомологичной этому гену)."Смысловая РНК" представляет собой РНК-последовательность, которая, в основном, гомологична по крайней мере части соответствующей мРНК-последовательности. Такая смысловая РНК может быть продуцирована в клетке при ее трансформации соответствующей ДНК-конструкцией, расположенной в нормальной ориентации, так, чтобы генерировался транскрипт, имеющий последовательность, идентичную по крайней мере части кодирующей цепи релевантного гена (или ДНК-последовательности, в основном, гомологичной этому гену). Подходящие смысловые конструкции могут быть использованы для ингибирования экспрессии гена (как описано в международной патентной заявке WO 91/08299). ДНК-конструкции настоящего изобретения могут содержать нуклеотидную последовательность длиной по крайней мере в 10 оснований (предпочтительно по крайней мере 35 оснований) для транскрипции в РНК. Данная нуклеотидная последовательность не имеет теоретического верхнего предела она может иметь и длину, равную длине продуцированной клеткой, релевантной мРНК - но, в основном,предпочтительно использовать последовательности длиной от 100 до 1000 оснований. Получение таких конструкций более подробно описано ниже. В качестве источника нуклеотидной ДНК-последовательности для транскрипции могут быть использованы подходящие кДНК или геномная ДНК, РНК или синтетический полинуклеотид. Областью-3 006701 инициации транскрипции (или промотором), функционирующей в растениях, может быть конститутивный промотор (такой как промотор 35S вируса мозаики цветной капусты), либо индуцибельный или стадиеспецифический регулируемый промотор, если это необходимо. Подходящие ДНК-последовательности, регулирующие экспрессию генов, экспрессируемых в растениях (включая маркерные гены),такие как области инициации транскрипции, энхансеры, лидерные последовательности, нетранслируемые лидерные последовательности и т.п., могут быть получены от любого гена, который экспрессируется в растительной клетке. Могут быть также использованы и гибридные промоторы, объединяющие функциональные части различных промоторов, или их синтетические эквиваленты. Помимо конститутивных промоторов, регулирующих экспрессию экспрессируемых генов настоящего изобретения, могут быть использованы индуцибельные промоторы или промоторы, осуществляющие регуляцию соответствующего типа экспрессии, например, стадиеспецифическую или клеткоспецифическую экспрессию. Так,например, может оказаться желательным модифицировать активность белка на определенной стадии развития растения. Использование конститутивного промотора имеет своей целью воздействовать на уровни и функции белка во всех органах данного растения, тогда как использование тканеспецифического промотора позволяет осуществлять более эффективную регуляцию экспрессии гена и подвергаемых воздействию функций. В другом своем варианте настоящее изобретение относится к применению индуцибельных промоторов. Известны промоторы, которые индуцируются патогенами, стрессом, химическими соединениями и воздействиями окружающей среды. Индуцирование активности гена внутренними или внешними факторами входит в объем настоящего изобретения. Промоторы этого типа позволяют индуцировать активность гена регулируемым образом и тем самым позволяют растению нормально развиваться без чрезмерного воздействия на них трансгенного гена. Индуцибельными промоторами являются промоторы,описанные в DE 4446342 (промотор PRP-1, индуцируемый грибами и ауксином), в WO 96/28561 (промотор PRP-1, индуцируемый грибами), в ЕР 0712273 (промотор, индуцируемый нематодами), в ЕР 0330479 и патенте США 5510474 (промотор, индуцируемый стрессом), в WO/96/12814 (индуцируемый холодом) и промотор, индуцируемый спиртом Zeneca. Другие индуцибельные промоторы описаны в ЕР 0494724,ЕР 0619844, WO 92/19724. Таким образом, этот генный продукт, независимо от того, является ли он антисмысловой или смысловой РНК или пептидом, продуцируется в ткани лишь в то время, когда необходимо его действие. Как упоминалось выше, термин "индуцибельный промотор" включает промоторы, которые могут быть индуцированы химически. Использование промоторной последовательности, которая регулируется с применением внешних химических стимуляторов, является наиболее предпочтительным. Внешними химическими стимуляторами являются предпочтительно агрономически приемлемые химические соединения, использование которых совместимо с сельскохозяйственной практикой и не оказывает негативного воздействия на растения или млекопитающих. Наиболее предпочтительная индуцибельная промоторная область включает систему индуцибельных промоторов-переключателей, такую как, например, двухкомпонентная система, такая как система промоторов-переключателей генов alcA/alcR, описанная в международной заявке WО 93/21334, система промоторов-переключателей гена экдизона, описанная в международной заявке WO 96/37609, или промтор GST, описанный в международных заявках WO 90/08826 и WO 93/031294, содержание которых вводится в настоящее описание посредством ссылки. Такие промоторные системы называются "промоторами-переключателями". Химическими соединениямипереключателями, используемыми в сочетании с промоторами-переключателями, являются агрономически приемлемые химические соединения, делающие эту систему особенно эффективной в способе настоящего изобретения. Каждый специалист может самостоятельно выбрать ДНК-вектор для использования в данных способах. Примером вектора, подходящего для Agrobacterium-опосредованной трансформации, являетсяpBluescript или pIGPD7 (Hall et al., Nature Biotechnology 14: 1133-1138, 1996). Введение данного фрагмента в эти векторы может быть осуществлено стандартными методами молекулярной биологии, описанными, например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor LaboratoryPress; 1989. Трансформированные растения, полученные данным способом настоящего изобретения, обнаруживают абсолютную резистентность, или иммунитет, к BNYVV. В противоположность этому предшествующие попытки сообщения растениям резистентности к BNYVV (Kallerhof et al., Plant Cell Reports 9:224-228, 1990; и Mannenrlof et al., Euphytica 90:293-299, 1996), или к другим вирусам, таким как вирус табачной мозаики (TMV) (Donson et al., Mol. Plant-Microbe Interact. 6:635-642; 1993) путем трансформации растений фрагментами вирусного гена, оказались менее успешными. Инокулированные листья еще обнаруживали симптомы инфекции, что свидетельствует о том, что данная резистентность не является абсолютной. А поэтому тот факт, что способ настоящего изобретения позволяет сообщать растениям сахарной свеклы абсолютную резистентность к BNYVV, оказался неожиданностью.-4 006701 Кроме того, настоящее изобретение относится к трансформированной клетке растения и к трансгенному растению, резистентному к BNYVV, а также к репродуцируемым структурам, таким как семена,каллюсы, почки, зародыши, полученные от трансгенных растений, и к их потомству. В предпочтительном варианте осуществления изобретения описанная здесь резистентность может быть объединена с резистентностью других типов или с толерантностью к BNYVV. Настоящее изобретение может быть, кроме того, проиллюстрировано нижеследующими примерами или графическим материалом, но они не должны рассматриваться как ограничение изобретения. На фиг. 1 схематически представлена характерная геномная организация вируса некротического пожелтения жилок свеклы (на основе работы Jupin et al., Seminars in Virology vol. 2.2: 112-129; 1991). На фиг. 2 показаны физические карты pVDH239 и pVDH240. LB = левый край, RB = правый край,P35S = промотор 35S CaMV, NPTII = неомицин-фосфотрансфераза II, T35S = сигнал полиаденилирования 35S CaMV, GUSINT = ген бета-глюкуронидазы, BNYVVpolTRUNC = фрагмент кДНК 1 BNYVV,Tnos = происходящий от гена нопалин-синтазы сигнал полиаденилирования. Указаны положения сайтов распознавания главных рестриктирующих ферментов. На фиг. 3 проиллюстрирован блот-анализ по Саузерну, посредством которого было определено число Т-ДНК-вставок, интегрированных в геном первичного трансформанта Т 157-01 сахарной свеклы. В верхней части схематически показана Т-ДНК-структура бинарного вектора PVDH239. На фиг. 4 показаны диаграммы отдельных ELISA-величин для экстрактов из корней растений сахарной свеклы от популяций Cadyx (восприимчивый контроль), Rifle (сорт, толерантный к ризомании),Rhizor (сорт, толерантный к ризомании) и Т 157-01 (GUS-позитивные F1-растения) после инокуляцииBNYVV-инфицированной почвой. Каждое число на горизонтальной оси представляет отдельное растение. На фиг. 5 В проиллюстрирован блот-анализ по Саузерну, посредством которого было определено число Т-ДНК-вставок, интегрированных в геном F1-потомства растений Т 157-01, а также представлена диаграмма отдельных ELISA-величин для экстрактов из корней F1-потомства растений Т 157-01 после инокуляции BNYVV-инфицированной почвой (фиг. 5 А). Числа в верхней части Саузерн-блотов представляют лабораторные коды отдельных растений потомства F1. ELISA-величины, обозначенные "Генотипом 1" в нижней панели, соответствуют отдельным растениям, обнаруживающим одну полосу в Саузерн-блоте (2012, 2019, 2021, 2029, 2030, 2031, 2034, 2035, 2038, 2042, 2044, 2046, 2051, 2052, 2061, 2066,2068, 2069), тогда как ELISA-величины, обозначенные "Генотипом 2+3" в нижней панели, соответствуют отдельным растениям, обнаруживающим 2 или 3 полосы в Саузерн-блоте (1999, 2000, 2001, 2007, 2008,2011, 2013, 2014, 2015, 2016, 2017, 2018, 2020, 2022, 2023, 2024, 2025, 2026, 2027, 2028, 2032, 2033, 2036,2037, 2039, 2040, 2041, 2043, 2045, 2047, 2048, 2049, 2050, 2053, 2054, 2055, 2056, 2057, 2058, 2059, 2060,2062, 2063, 2064, 2065, 2067, 2070). ELISA-величины, обозначаемые "сегрегантами GUS(-)" в нижней панели, соответствуют отдельным растениям потомства F1, которые являются GUS-негативными (1997,1998, 2002, 2004, 2005, 2006, 2009, остальные не показаны). Примеры Пример 1. Создание конструкции с усеченной последовательностью репликазы BNYVV. Для получения кДНК-клонов BNYVV использовали комбинации из двух праймеров для клонирования в трансформирующем векторе (Bouzoubaa et al., J. Gen. Virol. 68: 615-626; 1987). Для 5'-конца были использованы праймеры:P1: 5'-CGCGGATCCACCATGGCAGATTCGTTC-3' (содержащий рестрикционные BamHI- и NcoIсайты и нуклеотиды, идентичные нуклеотидам 153-168) и Р 2: 5'-GACGAATTCAAGTCGTCTTTC-3' (рестрикционный EcoRI-сайт и нуклеотиды, комплементарные нуклеотидам 288-301). Для 3'-конца были использованы праймеры: Р 3: 5'-GACGAATTCGAAGATGAGTCTA-3' (EcoRI-сайт и нуклеотиды, идентичные нуклеотидам 2799-2812) и Р 4: 5'-CGCAGATCTTTAACTGCTCATCACCAAC-3' (BglII-сайт и нуклеотиды, комплементарные нуклеотидам 3244-3258 и стоп-кодон). Перед клонированием фрагментов в вектор pBluescript (Stratagene, La Jolla, CA, USA),Sall/HincIII/Accl-сайт заменяли на BgIII-сайт. После амплификации кДНК BNYVV с использованием Р 1 и Р 2 получали ДНК-фрагмент, который гидролизовали ферментами BamHI и EcoRI и встраивали в модифицированный BamHI/EcoRI-расщепленный вектор pBluescript, в результате чего получалиpRSNBPol1. Затем кДНК 1 BNYVV амплифицировали с использованием Р 3 и Р 4. Полученный ДНКфрагмент расщепляли EcoRI и BgIII и встраивали в плазмиду pRSNBPol1, расщепленную EcoRI и BgIII, в результате чего получали плазмиду pRSNBPol2. Затем Acc1-фрагмент кДНК 1 BNYVV, идентичный нуклеотидам 250-2815, клонировали в Асс 1 расщепленный pRSNBPol2. Таким образом получали pRSNBPoltrunc, который включал фрагмент кДНК 1BgIII-фрагментом у 3'-конца. Фрагмент poltrunc клонировали в виде BamHI-BgIII-фрагмента в BamHIсайте pVDH4 так, что функциональный смысловой фрагмент poltrunc находился за промотором 35SCaMV и перед терминаторной последовательностью гена нопалина. Полную конструкцию, несущую промотор 35S CaMV, фрагмент poltrunc и терминаторную последовательность нопалина, вырезали из плазмиды ферментом ClaI и клонировали в бинарный трансформирующий вектор pVDH212, в которомBamHI-сайт был преобразован в ClaI-сайт путем встраивания молекулярного линкера.pVDH212 представляет собой происходящий от pBIN19 бинарный трансформирующий вектор, который содержит 35S-NPTII (ген неомицинфосфотрансферазы, находящийся под контролем промотора 35S CaMV и используемый в качестве селективного маркерного гена, сообщающего резистентность к канамицину), а также 35S-GUSi (ген, кодирующий бета-глюкуронидазу (Vancanneyt et al., Mol.Gen.Genet. 220:245-250; 1990). Таким образом, были получены два различных трансформирующих вектора, содержащих указанную конструкцию poltrunc: один из которых, pVDH239, содержал конструкцию poltrunc в том же самом направлении транскрипции, что и гены NPTII и GUS, а другой, pVDH240, содержал конструкцию poltrunc, имеющую противоположное направление транскрипции. Бинарные трансформирующие векторы pVDH239 и 240 (фиг. 2) конъюгировали и переносили в Agrobacterium tumefaciens LBA44 03(Phabagen Collection, Utrecht, the Netherlands), в результате чего получали штаммы НАТ 1239 (дляpVDH239) и НАТ 1240 (для pVDH240). HAT1239 и НАТ 1240 были использованы для трансформации сахарной свеклы. Пример 2. Трансформация сахарной свеклы. Методы Agrobacterium-опосредованной трансформации с использованием бинарных векторов хорошо разработаны и известны каждому специалисту. Для получения трансформированных растений сахарной свеклы, были трансформированы растения В 8 М.5.9 О-типа методом Klens et al. (Plant Science 116:97-106; 1996). В результате трансформации Agrobacterium HAT1239 получали 8 трансформантов: Т 156-03, Т 156-06, Т 156-09, Т 156-10, Т 156-13, Т 156-20,Т 150-30 и Т 157-01, а в результате трансформации Agrobacterium HAT1240 получали 12 трансформантов: Т 183-01, Т 183-02, Т 183-06, Т 183-10, Т 183-16, Т 183-19, Т 183-21, Т 183-23, Т 183-26, Т 184-26, Т 184-02,Т 184-03 и Т 184-04. У этих трансформантов индуцировали цветение путем яровизации, и они были использованы для получения семян либо посредством самоопыления (семена, полученные самоопылением,или семена S1) или путем перекрестного опыления с растениями с мужской стерильностью (гибридные семена или семена F1). Для предупреждения опыления пыльцой сахарной свеклы от другого источника,опыление осуществляли в закрытых пыльцесодержащих камерах. Семена F1 использовали в качестве исходного материала для проведения биологического анализа на резистентность к BNYVV. Проростки предварительно скринировали на GUS-активность, что указывало на присутствие Т-ДНК. GUS-негативные сегреганты были использованы в качестве негативного контроля в данном биологическом анализе. В зависимости от полного числа вставок Т-ДНК, резистентность, если она существует, может подвергаться сегрегации в GUS-позитивной популяции. И лишь потомство, происходящее от Т 157-01, обнаруживало резистентность к BNYVV. Затем первичный трансформант Т 157-01 анализировали более тщательно на блотах по Саузерну. Геномную ДНК выделяли из листьев, расщепляли отдельно ферментами EcoRI, BamHI и SacI и использовали для получения блота по Саузерну, который затем зондировали GUS. Исходя из этого эксперимента был сделан вывод, что первичный трансформант содержал три вставки Т-ДНК (фиг. 3). Пример 3. Биологический анализ. Биологический анализ боковых корней был разработан как тест одного растения для отбора резистентных к ризомании растений сахарной свеклы. Однонедельные проростки пересаживали в стандартную смесь из 10% инфицированной ризоманией почвы, а затем инкубировали в течение 4 недель. Условия инфицирования оптимизировали и стандартизировали по уровню давления инфекции и роста корешков (только образование волосистых корней). Использовали следующие условия инкубирования: 18,919 С день/ночь, относительная влажность 70%, освещенность дневным светом 10,000 люкс, вегетационные сосуды находились на расстоянии 32,5 см от ламп. Количество вируса в проростках, которое непосредственно коррелировало с механизмом резистентности у растений, измеряли ELISA-методом. Величину отсечки для резистентных растений устанавливали при значении ОП 0,2 (Clark et al., J.Virol.Meth. 15:213-222; 1987). Через четыре недели после пересадки нижние части корней использовали для ELISA. Кусочки корней сушили на фильтровальной бумаге и переносили в ступку. Затем добавляли буфер для экстракции при отношении 10 объемных эквивалентов массы корней (разведение 1/10). Сок из корней экстрагировали посредством ручной выжимки. Этот сок переносили в 1,5 мл-пробирки и центрифугировали (300 об./мин, 10 мин) и супернатанты выдерживали на льду и анализировали. Были протестированы следующие популяции:-6 006701 В группе Т 157-01 могут наблюдаться две категории GUS(+)-растений (фиг. 4). Одна категория обнаруживает иммунитет против BNYVV (ELISA-отсечка 0,2) со средней ELISA-величиной 0,006, которое представляет собой фоновый уровень экспериментальной системы. Другая категория обнаруживает нормальную восприимчивость со средней ELISA-величиной в пределах восприимчивого контроля. Давление инфекции в данном биоанализе было высоким вследствие повышения температуры в течение определенного отрезка времени инфекционного периода. Поэтому явного отличия между Cadyx иRhizor/Rifle не наблюдалось (фиг. 4). С другой стороны, резистентные растения, выбранные в F1 потомстве Т 157-01, могут рассматриваться как абсолютно резистентные независимо от давления инфекционной ризомании (фиг. 4). Исходя из этого, можно сделать вывод, что введенная конструкция, содержащая фрагмент кДНК 1 BNYVV, приводила к сильному негативному воздействию на размножениеBNYVV в боковых корнях инокулированных трансгенных растений сахарной свеклы, что позволяло эффективно обеспечивать полную резистентность этих растений к ризомании. Для объяснения сегрегации GUS(+)-популяции на категории резистентных и восприимчивых растений, отдельные растения были проанализированы в независимом эксперименте на резистентность кBNYW и одновременно на присутствие Т-ДНК с помощью блот-анализа по Саузерну с использованиемSacI в качестве рестриктирующего фермента и GUS в качестве зонда. Результаты, представленные на фиг. 5 А и 5 В, свидетельствовали о том, что все GUS(+)-растения в той популяции, которая содержала одну полосу на блоте по Саузерну, были восприимчивыми (средняя ELISA-величина - 0,63), тогда как все GUS(+)-растения, которые содержали 2 или 3 полосы на блоте по Саузерну, были резистентными(средняя ELISA-величина - 0,21). Все GUS(-)-сегреганты оказались восприимчивыми (средняя ELISAвеличина - 0,80). Очевидно, что резистентный фенотип ассоциирован с присутствием верхней и нижней полосы этой конкретной картины полос, полученной на блотах по Саузерну, тогда как присутствие средней полосы не ассоциировано с резистентным фенотипом. Исходя из этого, можно сделать вывод, что расщепление GUS(+)-популяции на восприимчивую и резистентную категорию может быть объяснено тем фактом, что одна из Т-ДНК, которая приводит к GUS(+)-фенотипу, не сообщает резистености к ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ придания растению сахарной свеклы резистентности к вирусу некротического пожелтения жилок свеклы (BNYVV), включающий следующие стадии:(a) получение ДНК-фрагмента, который состоит по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 вируса некротического пожелтения жилок свеклы (BNYVV), причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы;(b) введение указанного ДНК-фрагмента, функционально связанного с промотором, который является активным в растениях сахарной свеклы, в клетку растения сахарной свеклы с целью получения трансформированной клетки растения сахарной свеклы и(c) регенерацию трансгенного растения сахарной свеклы из трансформированной клетки растения сахарной свеклы. 2. Способ по п.1, отличающийся тем, что указанный фрагмент имеет последовательность, которая соответствует нуклеотидам 153-3258 РНК 1 BNYVV. 3. Способ по любому из пп.1 или 2, отличающийся тем, что указанный фрагмент вводят в клетку в составе ДНК-вектора, содержащего транскрипционную и трансляционную регуляторные последовательности, функционально присоединенные к указанному фрагменту. 4. ДНК-вектор, содержащий фрагмент, который состоит по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 (BNYVV), причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы и транскрипционную и трансляционную регуляторные последовательности, функционально присоединенные к указанному фрагменту. 5. ДНК-вектор по п.4, отличающийся тем, что указанный фрагмент имеет последовательность, которая соответствует нуклеотидам 153-3258 РНК 1 BNYVV. 6. Применение ДНК-вектора по любому из пп.4 или 5 для трансформации клетки растения сахарной свеклы с целью получения растений, которые являются резистентными к BNYVV. 7. Клетка растения сахарной свеклы, обладающего резистентностью к BNYVV, которая содержит в своем геноме ДНК-фрагмент, состоящий по крайней мере из 15 последовательно расположенных нуклеотидов геномной РНК 1 BNYVV, причем указанный фрагмент включает по крайней мере часть открытой рамки считывания геликазы. 8. Клетка растения по п.7, отличающаяся тем, что указанный фрагмент имеет последовательность,которая соответствует нуклеотидам 153-3258 РНК 1 BNYVV. 9. Применение клетки растения по любому из пп.7-8 для регенерации из нее растения сахарной свеклы, обладающего резистентностью к BNYVV. 10. Растение сахарной свеклы, регенерированное из клетки по любому из пп.7-8. 11. Потомство растения сахарной свеклы по п.10. 12. Семена растения сахарной свеклы по п.10 или потомства по п.11.-9 006701 13. Вегетативно репродуцируемые структуры, такие как каллюс, почки, зародыши, происходящие от растения по п.10 или от потомства по п.11. 14. Применение семян по п.12 для регенерации из них растения сахарной свеклы, которое является резистентным к BNYVV. 15. Применение вегетативно репродуцируемых структур по п.13 для регенерации из них растения сахарной свеклы, которое является резистентным к BNYVV.
МПК / Метки
МПК: C12N 15/40, C12N 15/82, C12N 5/10, A01H 5/00
Метки: придания, пожелтения, резистентности, части, данному, резистентные, некротического, растению, днк-вектор, этого, используемый, жилок, способ, сахарной, свеклы, вирусу, растение
Код ссылки
<a href="https://eas.patents.su/13-6701-sposob-pridaniya-rasteniyu-saharnojj-svekly-rezistentnosti-k-virusu-nekroticheskogo-pozhelteniya-zhilok-svekly-ispolzuemyjj-dlya-etogo-dnk-vektor-i-rastenie-i-ego-chasti-rezistentn.html" rel="bookmark" title="База патентов Евразийского Союза">Способ придания растению сахарной свеклы резистентности к вирусу некротического пожелтения жилок свеклы, используемый для этого днк-вектор и растение и его части, резистентные к данному вирусу</a>
Линия клеток растения сахарной свеклы и растение сахарной свеклы, устойчивые к имидазолиноновому гербициду, семена указанного растения, способы получения указанной клеточной линии и гибридных растенийсвеклы, устойчивых к данному гербициду, и применение указанного гербицида для борьбы с сорняками, растущими на полях с указанными растениями сахарной свеклы

Номер патента: 3296
Опубликовано: 24.04.2003
Авторы: Пеннер Доналд, Райт Терри Р.
Метки: свеклы, растущими, гибридных, полях, указанного, получения, линии, устойчивые, способы, устойчивых, имидазолиноновому, сорняками, растенийсвеклы, растения, указанными, растение, клеточной, гербицида, борьбы, гербициду, применение, семена, указанной, растениями, данному, сахарной, линия, клеток
Формула / Реферат:
1. Линия клеток растения сахарной свеклы с мутированным геном ацетолактатсинтазы, в котором в положении 337 гуанин замещен на аденин, устойчивая к имидазолиноновому гербициду, способная регенерировать в растение сахарной свеклы, причем указанная устойчивость передается при общепринятом кроссбридинге регенерированных растений с растениями, чувствительными к данному гербициду. 2. Линия клеток по п.1, которая получена из чувствительных клеток...
Способ придания растительной клетке или растению устойчивости к вирусу, содержащему последовательность 3 тройного блока генов (tgb3), и трансгенное растение, устойчивое к указанному вирусу

Номер патента: 4328
Опубликовано: 29.04.2004
Авторы: Вейен Ги, Лефебвр Марк, Бузубаа Салах, Блейкастен-Гроссханс Клодин, Жонар Жерар, Гюйие Убер, Ричардс Кен
МПК: C07K 14/08, A01H 5/00, C12N 15/82...
Метки: устойчивое, вирусу, последовательность, устойчивости, растение, тройного, растению, клетке, указанному, придания, блока, трансгенное, растительной, tgb3, способ, генов, содержащему
Формула / Реферат:
1. Способ придания растительной клетке или растению устойчивости к вирусу, содержащему последовательность 3 из тройного блока генов (TGB3), с условием, что это не вирус X картофеля, состоящий из следующих этапов: получения конструкции нуклеиновой кислоты, содержащей нуклеотидную последовательность TGB3 указанного вируса или соответствующую ей кДНК или их варианты, основанные на вырожденности генетического кода, связанные с одной или более...
Линия клеток растения сахарной свеклы и растение сахарной свеклы, устойчивые к имидазолиноновым и сульфонилмочевинным гербицидам, семена указанного растения, способы получения указанной клеточной линии и гибридных растений свеклы, устойчивых к данным гербицидам, и применение имидазолинонового гербицида для борьбы с сорняками, растущими на полях с указанными растениями сахарной свеклы

Номер патента: 3297
Опубликовано: 24.04.2003
Авторы: Райт Терри Р., Пеннер Доналд
Метки: указанного, гибридных, растущими, сорняками, указанной, клеточной, указанными, имидазолиноновым, получения, растение, данным, растений, сахарной, растения, гербицидам, устойчивые, сульфонилмочевинным, свеклы, семена, применение, борьбы, способы, растениями, имидазолинонового, линии, полях, устойчивых, гербицида, линия, клеток
Формула / Реферат:
1. Линия клеток растения сахарной свеклы с мутированным геном ацетолактатсинтазы, в котором в положении 562 цитозин замещен на тимин и в положении 337 гуанин замещен на аденин, устойчивая к имидазолиноновым и сульфонилмочевинным гербицидам, способная регенерировать в растение сахарной свеклы, причем указанная устойчивость передается при общепринятом кроссбридинге регенерированных растений с растениями, чувствительными к данным гербицидам. 2....
Способ индуцирования у растительной клетки или растения устойчивости к вирусу, растение и растительная клетка, обладающие такой устойчивостью

Номер патента: 5168
Опубликовано: 30.12.2004
Авторы: Лобер Эммануэль, Гийе Убер, Ричард Кеннет, Жонар Жерар
МПК: A01H 5/00, C12N 15/40
Метки: устойчивости, способ, клетки, клетка, растения, растительной, обладающие, индуцирования, вирусу, растение, устойчивостью, растительная
Формула / Реферат:
1. Способ индуцирования у растительной клетки или растения устойчивости к вирусу, содержащему последовательность 2 из тройного блока генов (TGB2), включающий следующие этапы: получение конструкции нуклеиновой кислоты, содержащей нуклеотидную последовательность TGB2 указанного вируса или соответствующую ей кДНК или их варианты, основанные на вырожденности генетического кода, связанные с одной или более регуляторными последовательностями,...
Материал, используемый в приготовлении рассадочной грядки, способ регулирования прорастания сорняков в рассадочных грядках и применение этого материала и этого способа для регулирования прорастания сорняков

Номер патента: 2553
Опубликовано: 27.06.2002
Автор: Нильсен Карстен Хольмгор
МПК: A01G 13/02, A01C 1/04
Метки: прорастания, регулирования, приготовлении, рассадочных, материала, способа, материал, сорняков, рассадочной, используемый, способ, этого, грядки, применение, грядках
Формула / Реферат:
1. Материал, используемый в приготовлении рассадочной грядки для выращивания сельскохозяйственных культур, содержащий два слоя (2, 5) материала, сквозь который не могут прорастать растения, для закладки на уровне ниже уровня поверхности рассадочной грядки так, что они проходят от указанного уровня ниже уровня поверхности до уровня выше уровня поверхности рассадочной грядки, отличающийся тем, что с указанными двумя слоями материала на указанном...
Предыдущий патент: Носитель с твердым фибриногеном и твердым тромбином
Следующий патент: Способ определения последовательности полинуклеотида
Случайный патент: Способ получения смазочного масла и газойля