Связывающий днк-сайт активатора транскрипции, пригодный для экспрессии генов
Номер патента: 14783
Опубликовано: 28.02.2011
Авторы: Пареникова Люси, Пейдж Ван Ноэль Николас Мария Элизабет



























Формула / Реферат
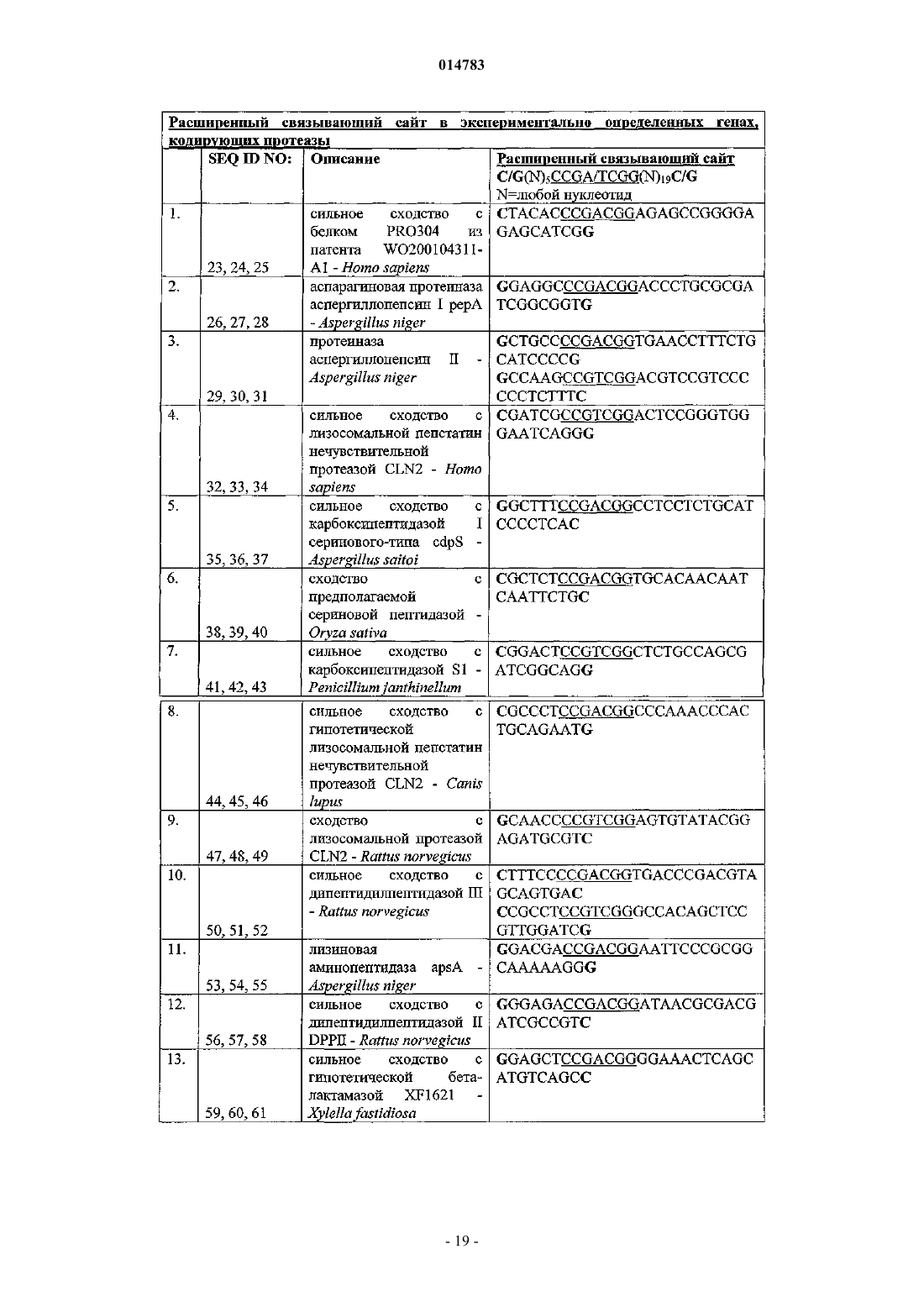
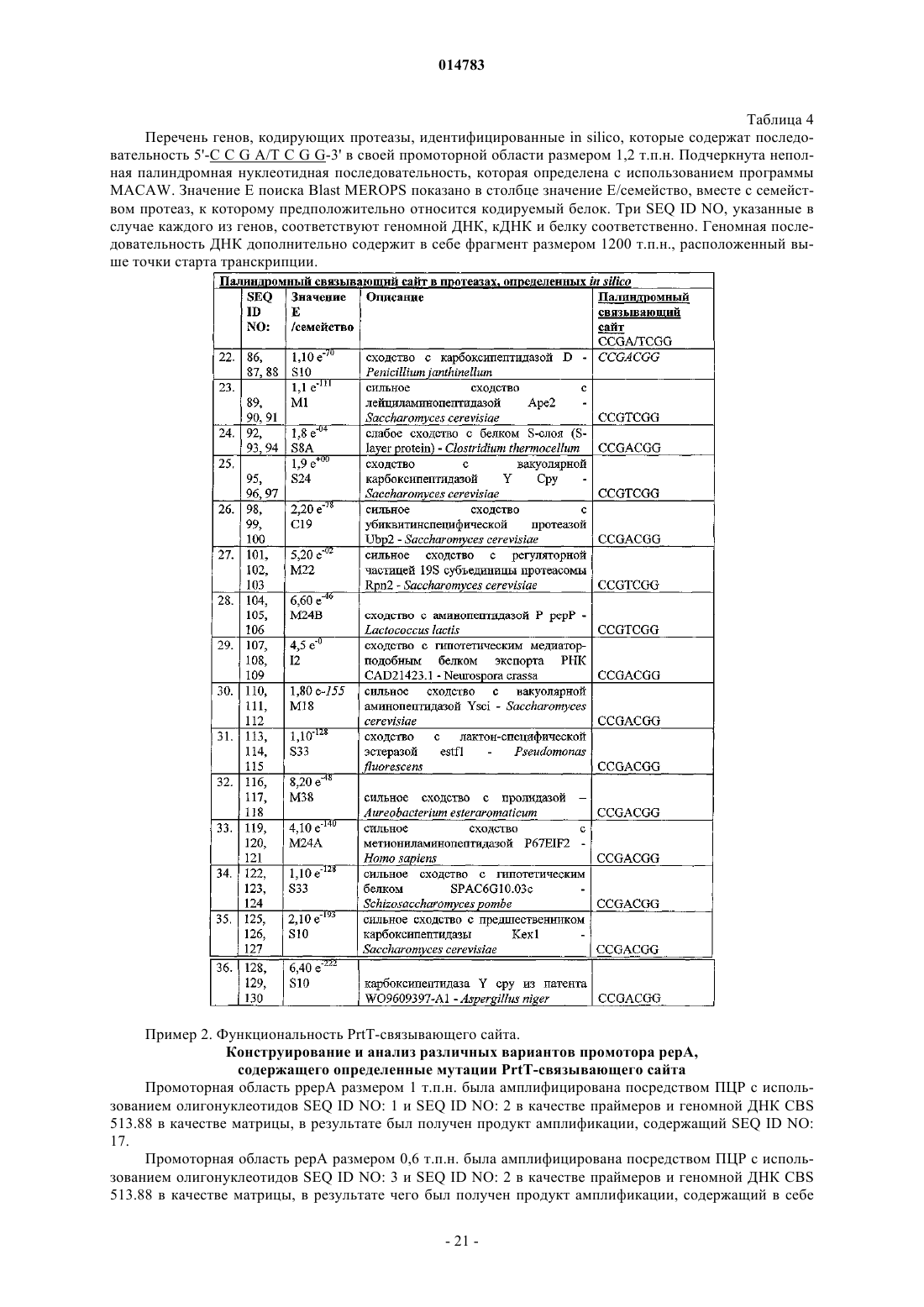
1. Изолированный полинуклеотид, включающий двухцепочечный связывающий ДНК-сайт для активатора транскрипции PrtT, где по меньшей мере 32 основания первой цепи сайта идентичны по последовательности с 5'-G/C(N)5ССGА/ТСGG(N)19G/C-3' (SEQ ID NO:22), вторая цепь сайта комплементарна первой цепи и связывание PrtT с сайтом активирует транскрипцию расположенной ниже нуклеотидной последовательности в клетке-хозяине.
2. Изолированный полинуклеотид, включающий двухцепочечный мутантный нефункциональный связывающий ДНК-сайт, где по меньшей мере 32 основания первой цепи немутантного сайта идентичны по последовательности с 5'-G/C(N)5ССGА/ТСGG(N)19G/C-3' (SEQ ID NO:22), вторая цепь мутантного нефункционального сайта комплементарна первой цепи, немутантный сайт связывается активатором транскрипции PrtT и связывание PrtT с немутантным сайтом активирует транскрипцию с расположенной ниже нуклеотидной последовательностью в клетке-хозяине, но по меньшей мере одно основание первой цепи мутантного нефункционального сайта изменено по сравнению с нуклеотидной последовательностью немутантного сайта таким образом, что PrtT больше не связывается с мутантным нефункциональным сайтом или что PrtT больше не активирует транскрипцию расположенной ниже нуклеотидной последовательности.
3. Изолированный полинуклеотид, включающий двухцепочечный мутантный улучшенный связывающий ДНК-сайт, где по меньшей мере 32 основания первой цепи немутантного сайта являются идентичными по последовательности с 5'-G/C(N)5ССGА/ТСGG(N)19G/C-3' (SEQ ID NO:22), вторая цепь мутантного улучшенного сайта комплементарна первой цепи, немутантный и мутантный улучшенные сайты связываются активатором транскрипции PrtT и связывание PrtT как с немутантным, так и с мутантным улучшенным сайтом активирует транскрипцию расположенной ниже нуклеотидной последовательности в клетке-хозяине, но по меньшей мере одно основание первой цепи мутантного улучшенного сайта изменено по сравнению с нуклеотидной последовательностью немутантного сайта таким образом, что транскрипция расположенной ниже нуклеотидной последовательности является улучшенной.
4. Рекомбинантный экспрессионный вектор, включающий связывающий ДНК-сайт по п.1 или 3 в составе промотора, стоп-сигнал транскрипции и стоп-сигнал трансляции.
5. Вектор по п.4, дополнительно включающий расположенную ниже нуклеотидную последовательность, которая кодирует полипептид, где транскрипция расположенной ниже нуклеотидной последовательности активируется PrtT.
6. Клетка-хозяин, включающая полинуклеотид по п.2 или 3 или экспрессионный вектор по п.4 или 5.
7. Клетка-хозяин, где по меньшей мере один связывающий ДНК-сайт в геноме клетки-хозяина мутирован в соответствии с п.2 таким образом, что PrtT не может связываться с мутантным сайтом или PrtT не может активировать транскрипцию расположенной ниже нуклеотидной последовательности.
8. Способ получения клетки-хозяина по п.7, где указанный способ включает в себя:
(a) введение по меньшей мере одного мутантного, нефункционального связывающего ДНК-сайта в промотор клетки-хозяина посредством мутагенеза или рекомбинации и
(b) необязательное подтверждение уменьшенного связывания PrtT с мутантным нефункциональным сайтом или уменьшенной PrtT-зависимой активации транскрипции в клетке-хозяине.
9. Клетка-хозяин, где связывающий ДНК-сайт в одном или более протеазном гене клетки-хозяина мутирован в соответствии с п.2 таким образом, что PrtT не может связываться с мутантным сайтом или что PrtT не может активировать транскрипцию протеазного гена, что приводит к получению клетки-хозяина с фенотипом сниженной протеазной активности.
10. Способ получения полипептида, где указанный способ включает в себя:
(a) культивирование клетки-хозяина по п.9 в питательной среде в условиях, приводящих к экспрессии полипептида,
(b) экспрессию полипептида в клетке-хозяине и
(c) необязательное извлечение полипептида из питательной среды или из клетки-хозяина.
11. Способ получения полипептида, где указанный способ включает в себя:
(a) трансформацию клетки-хозяина по п.9 экспрессионным вектором, где вектор экспрессирует полипептид,
(b) культивирование клетки-хозяина в питательной среде в условиях, приводящих к экспрессии полипептида,
(c) экспрессию полипептида в клетке-хозяине и
(d) необязательное извлечение полипептида из питательной среды или из клетки-хозяина.
12. Способ получения полипептида, где указанный способ включает в себя:
(a) культивирование в питательной среде клетки-хозяина, содержащей вектор по п.5, в условиях, приводящих к экспрессии полипептида, кодируемого расположенной ниже нуклеотидной последовательностью, содержащейся в данном векторе,
(b) экспрессию полипептида в клетке-хозяине и
(d) необязательное извлечение полипептида из питательной среды или из клетки-хозяина.
Текст
СВЯЗЫВАЮЩИЙ ДНК-САЙТ АКТИВАТОРА ТРАНСКРИПЦИИ, ПРИГОДНЫЙ ДЛЯ ЭКСПРЕССИИ ГЕНОВ(71)(73) Заявитель и патентовладелец: ДСМ АйПи АССЕТС Б.В. (NL) Пареникова Люси, Пейдж Ван Ноэль Николас Мария Элизабет (NL) Представитель: Описаны связывающие ДНК-сайты, которые специфически узнаются активатором транскрипции протеазных генов PrtT. Связывающий ДНК-сайт может быть определен структурно по консенсусной нуклеотидной последовательности, а также функционально, исходя из способности PrtT регулировать активацию транскрипции через данную последовательность. Как PrtT, так и соответствующий ему связывающий ДНК-сайт, (то есть нуклеотидная последовательность в каждом промоторе, которая узнается PrtT) могут применяться в системе экспрессии генов. Наличие только одного активатора транскрипции PrtT является недостаточным, соответствующий ему связывающий ДНК-сайт является необходимым для узнавания PrtT (то есть связывания с сайтом и активации транскрипции в подходящих условиях). Функциональный сайт, такой как сайт, полученный из гена гриба дикого типа, будет обеспечивать PrtT-зависимую активацию транскрипции с 3'-нижерасположенных последовательностей. Мутация промотора дикого типа, которая приводит к получению нефункционального сайта, будет нарушать PrtT-зависимую активацию транскрипции 3'-нижерасположенных последовательностей. Мутация промотора дикого типа, которая приводит к получению более функционального сайта, будет усиливать PrtT-зависимую активацию транскрипции с 3'-нижерасположенных последовательностей. 014783 Область техники, к которой относится изобретение Настоящее изобретение касается полинуклеотидов, имеющих функциональный, мутантный нефункциональный и/или мутантный усиленный участок, связывающий ДНК-сайт, который специфически узнается активатором транскрипции протеазных генов PrtT, и применения данных полинуклеотидов в регуляции экспрессии генов. Сведения о предшествующем уровне техники Активаторы транскрипции у грибов, называемые PrtT, были недавно описаны в WO 00/20596, WO 01/68864 и WO 06/040312. Данные активаторы транскрипции были выделены из Aspergillus niger (A. niger) и Aspergillus oryzae (A. oryzae). Они универсально активируют транскрипцию с промоторов, расположенных выше 5'-концов протеазных генов у грибов. До последнего времени модуляция PrtTактивируемой транскрипции была возможна только на общем уровне посредством изменения активатора транскрипции самого по себе. Здесь мы представляем консервативные связывающие ДНК-сайты в промоторах протеазных генов,которые являются необходимыми и достаточными для обеспечения PrtT-зависимой активации транскрипции, расположенной ниже нуклеотидной последовательности. Настоящее изобретение предоставляет новый связывающий ДНК-сайт, расположенный в промоторной области и узнаваемый активаторами транскрипции PrtT, который имеет улучшенные свойства для регуляции транскрипции генов у грибов по сравнению с ранее описанными последовательностями. Другие преимущества и улучшения описаны ниже или будут очевидны из нижеследующего раскрытия изобретения. Раскрытие изобретение Целью настоящего изобретения является контроль (регулирование) при помощи генной инженерии реактивности PrtT, улучшенной реактивности или нереактивности природного гена грибов или другой гетерологичной нуклеотидной последовательности по способности к узнаванию или неузнаванию функционального или мутантного связывающего ДНК-сайта промотора в клетках грибов. В первом аспекте настоящего изобретения предоставляются полинуклеотиды, которые содержат в себе один или более связывающий ДНК-сайт, где по меньшей мере один сайт узнается PrtT и обеспечивает PrtT-зависимую активацию транскрипции. Во втором аспекте настоящего изобретения предоставляются полинуклеотиды, которые содержат в себе один или более мутантный, нефункциональный ДНК-связывающий сайт, где по меньшей мере один сайт не узнается PrtT и/или не обеспечивает PrtT-зависимой активации транскрипции. В третьем аспекте настоящего изобретения предоставляются полинуклеотиды, которые содержат в себе один или более мутантный, улучшенный связывающий ДНК-сайт, где по меньшей мере один сайт узнается PrtT и обеспечивает усиленную PrtT-зависимую активацию транскрипции. В четвертом аспекте настоящего изобретения предоставляются экспрессионные векторы, содержащие в себе по меньшей мере один из вышеупомянутых немутантньгх и/или мутантных, улучшенных связывающих ДНК-сайтов. Вектор может дополнительно содержать в себе расположенную ниже нуклеотидную последовательность, которая транскрибируется с промотора, который содержит в себе функциональный или функциональный и улучшенный связывающий ДНК-сайт и активируется посредством PrtT. Кроме того, предоставляются клетки, содержащие в себе вышеупомянутый связывающий ДНК-сайт (например, немутантный, мутантный, нефункциональный и мутантный, усиленный), и/или вышеупомянутый вектор. В пятом аспекте настоящего изобретения предоставляются клетки, в которых по меньшей мере один эндогенный ген мутирован таким образом, что его промотор больше не связывается с PrtT или транскрипционно не активируется PrtT, или клетки, содержащие в себе мутантные, нефункциональные вышеупомянутые полинуклеотиды, или соответствующий экспрессионный вектор. Также предоставляются такие клетки, которые обладают, по меньшей мере, сниженной протеазной активностью. В шестом аспекте настоящего изобретения предоставляются способы идентификации протеазных генов. Один или более дифференциально экспрессируемых генов детектируются в клетках грибов, как содержащих, так и не содержащих активатор транскрипции PrtT (например, в случае удаления prtT или инактивации PrtT иным способом). Те дифференциально экспрессируемые гены, которые кодируют протеазы (например, определенные на основании сходства последовательности с известными протеазами, по протеолитической активности продукта гена и по наличию связывающего ДНК-сайта для PrtT), идентифицируются как протеазные гены, контролируемые PrtT. Также предоставляются идентифицированные таким способом новые протеазы и кодирующие их полинуклеотиды. В седьмом аспекте настоящего изобретения предоставляются способы получения клетки, содержащей в промоторе по меньшей мере один мутантный, нефункциональный связывающий ДНК-сайт. Мутация может быть введена в хромосому хозяина (то есть в эндогенный ген, предпочтительно в протеазный ген) посредством мутагенеза или с помощью рекомбинантной техники. Введение мутации в геном хозяина может быть подтверждено или уменьшением связывания PrtT с мутантным связывающим ДНКсайтом, или уменьшением PrtT-зависимой активации транскрипции с данного промотора. Является предпочтительной инактивация эндогенных промоторов по меньшей мере 13 идентифицированных экс-1 014783 периментально, более предпочтительно всех нативных протеазных генов посредством мутирования соответствующего количества связывающих ДНК-сайтов. В восьмом аспекте настоящего изобретения предоставляются способы получения полипептида. Один или более полипептидов экспрессируются в клетке с экспрессионного вектора или с эндогенного промотора нативного гена грибов. В случае чувствительных к протеазам полипептидов, клетка предпочтительно является клеткой с уменьшенной активностью протеиназ. В случае полипептидов (таких как многие протеазы), которые секретируются из клетки, они могут быть выделены из питательной среды. В ином случае полипептиды выделяются из клеток (предпочтительно из клеточной массы или осадка): (i) растворимые полипептиды выделяются из клеточного лизата и (ii) нерастворимые полипептиды или полипептиды, встраиваемые в мембраны или ассоциированные с клеточными мембранами, из клеточной фракции. Также предоставляются другие способы применения и получения вышеупомянутых полинуклеотидов, экспрессионных векторов, новых протеаз и кодирующих их полинуклеотидов, а также клеток. Дополнительные аспекты настоящего изобретения будут ясны из нижеследующего описания и формулы изобретения, в которых они обобщены. Краткий перечень фигур Фиг. 1 показывает измерение протеазной активности в супернатантах культур клеток. Результаты измерения по методу Anson (J. Gen. Physiol. 22:79-89, 1938) были получены с использованием супернатанта, полученного после 10 дней ферментации во встряхиваемой колбе. Измерения протеазной активности в трансформантах CBS 513.88 дельта рерА, несущих в себе конструкты с различным образом модифицированными промоторами pepA (см. табл. 5), показаны в столбиках с 4 по 9. Столбики 2 и 3 соответствуют трансформантам, несущим в себе конструкты с 1,0 т.п.н. промотором pepA дикого типа (содержащим в себе PrtT-связывающий сайт) и с 0,6 т.п.н. промотором pepA дикого типа (не содержащим в себе PrtT-связывающего сайта) соответственно. Фон был определен как протеазная активность нетрансформированного Aspergillus niger CBS 513.88 дельта pepA (столбик 1). Каждый столбик представляет собой среднее значение протеазной активности двух независимых трансформантов. Фиг. 2 показывает физическую плазмидную карту вектора 1, на которой показаны сайты узнавания ферментами рестрикции, расположение генов и их ориентация. Эта информация используется для переклонирования конструктов AscI/XhoI ррерА-рерА. Фиг. 3 показывает физическую плазмидную карту экспрессионного вектора pGBFIN-23, на которой показаны сайты узнавания ферментами рестрикции, расположение генов и их ориентация. Показаны фланкирующие glaA области по отношению к промотору glaA и сайту клонирования HindIII-XhoI. ДНК Е. coli может быть удалена посредством расщепления вектора с использованием фермента рестрикцииNoiI перед трансформацией штамма A. niger. Фиг. 4 показывает физическую плазмидную карту замещающего вектора pGBDEL, на которой показаны сайты узнавания ферментами рестрикции, расположение генов и их ориентация. Показаны множественные сайты клонирования для клонирования фланкирующих областей по отношению к маркеруamdS. Фиг. 5 показывает физическую плазмидную карту замещающего вектора pGBDEL-PRT2, на которой показаны сайты узнавания ферментами рестрикции, расположение генов и их ориентация. Показаны 5' prtT фланкирующая область и 3' prtT фланкирующая область по отношению к маркеру amdS. Последовательности 3' prtT фланкирующих областей перекрываются, по меньшей мере, участками в несколько сотен пар оснований. ДНК Е. coli была удалена посредством расщепления вектора при использовании ферментов рестрикции BstBI и XmaI с последующим замыканием вектора в кольцо перед трансформацией штамма A. niger. Фиг. 6 иллюстрирует процедуру удаления хромосомного prtT гена. Линейный ДНК-конструктpGBDEL-PRT2, содержащий в себе селективный маркер amdS, фланкированный гомологичными областями (5' и 3') гена prtT (1), интегрируется в геном посредством гомологичной рекомбинации с двойным перекрестом (double-crossover) (X) в генном локусе prtT (2) и замещает хромосомный ген prtT (3). После этого в результате рекомбинации между прямыми повторами (3', 3' область) происходит удаление маркера amdS, что приводит к точному вырезанию гена prtT (4). Сведения, подтверждающие возможность осуществления изобретения Связывающий ДНК-сайт может быть определен структурно по консенсусной нуклеотидной последовательности и функционально по способности PrtT регулировать активацию транскрипции через данную последовательность. Связывающий ДНК-сайт был идентифицирован как консервативная нуклеотидная последовательность (SEQ ID NO: 22 представляет собой удлиненную последовательность) в эндогенных промоторах природных протеазных генов грибов. Как PrtT, так и соответствующий ему связывающий ДНК-сайт (то есть нуклеотидная последовательность в каждом из промоторов, которая узнаетсяPrtT и обеспечивает способность PrtT контролировать транскрипцию гена), могут применяться в системе экспрессии генов как (i) для усовершенствования способа получения полипептидов в PrtT-содержащей клетке, так и (ii) для усовершенствования способа получения чувствительного к протеазам полипептида в PrtT-содержащей клетке. Наличие только одного активатора транскрипции PrtT является недостаточ-2 014783 ным, поскольку локализация соответствующего ему связывающего ДНК-сайта в промоторе необходима для узнавания активатором PrtT (то есть для связывания с сайтом и активации транскрипции с промотора в подходящих условиях). Функциональный сайт, такой как полученный из протеазного гена грибов дикого типа, будет обеспечивать PrtT-зависимую активацию транскрипции с расположенных ниже 3'-конца последовательностей. Напротив, мутация промотора дикого типа, которая приводит к появлению нефункционального сайта, будет нарушать PrtT-зависимую активацию транскрипции 3'-расположенных ниже последовательностей. Представленная в перечне последовательностей, являющихся предметом настоящего изобретения, последовательность SEQ ID NO: 22 описывает ДНК-сайт, связывающий PrtT,как [snnnnnccgw cggnnnnnnn nnnnnnnnnn nns]. Для внесения ясности, в описании настоящего изобретенияSEQ ID NO: 22 будет описываться как 5'-G/C (N)5 С С G А/Т С G G (N)19 G/C-3', то есть G/C в случае S и А/Т в случае W. В одном воплощении настоящего изобретения предоставляется полинуклеотид, содержащий в себе один или более двухцепочечный связывающий ДНК-сайт. По меньшей мере 32 основания первой цепи сайта являются идентичными по последовательности с 5'-G/C (N)5 С С G А/Т С G G (N)19 G/C-3' (SEQ IDNO:22). Поскольку 24 основания в удлиненном связывающем ДНК-сайте могут быть вырождены, то для изменения аффинности при связывании PrtT и/или для изменения PrtT-активируемой транскрипции расположенной ниже нуклеотидной последовательности замены в последовательности могут производиться только в девяти позициях. Вторая цепь сайта комплементарна первой цепи. Данный сайт специфически связывается с активатором транскрипции PrtT в условиях, позволяющих связывание, и такое связывание будет активировать транскрипцию расположенной ниже нуклеотидной последовательности. Асимметрия приведенной выше удлиненной последовательности или более короткой последовательности 5'-С С G А/Т С G G-3' полезна при определении направления транскрипции, а именно направления 5'3'. Предпочтительно, чтобы функциональный связывающий ДНК-сайт сохранял неполную палиндромную последовательность из семи последовательных оснований. Двухцепочечный связывающий ДНК-сайт может быть получен из эукариот (например, из грибов). В другом воплощении настоящего изобретения предоставляется полинуклеотид, содержащий в себе один или более двухцепочечных мутантных, нефункциональных связывающих ДНК-сайтов. Двухцепочечный связывающий ДНК-сайт (например, по меньшей мере 32 основания первой цепи немутантного сайта являются идентичными по последовательности с последовательностью 5'-G/C (N)5 С С G А/Т С GG (N)19 G/C-3', SEQ ID NO: 22) получен из эукариот (например, из грибов), где немутантный, функциональный сайт специфически связывается активатором транскрипции PrtT в условиях, разрешающих связывание, и такое связывание будет активировать транскрипцию расположенной ниже нуклеотидной последовательности. Но по меньшей мере одно основание (например, одно, два, три, четыре, пять, шесть,семь, восемь или девять оснований) первой цепи мутантного сайта изменено по сравнению с последовательностью немутантного сайта (например, посредством добавления, удаления, перемещения, трансверсии оснований или любой комбинации действий в удлиненной последовательности) таким образом, чтоPrtT больше специфически не распознает мутантный, нефункциональный сайт, или PrtT больше не активирует транскрипцию. Поскольку 24 основания в удлиненном связывающем ДНК-сайте могут быть вырожденны, изменения в последовательности с целью изменения связывающей аффинности PrtT и/или для изменения активируемой PrtT транскрипции расположенной ниже нуклеотидной последовательности могут производиться максимум в девяти позициях. Вторая цепь мутантного нефункционального сайта является комплементарной к первой цепи. Немутантный сайт специфически связывался с активатором транскрипции PrtT в условиях, разрешающих связывание, и такое связывание будет обеспечивать активацию транскрипции расположенной ниже нуклеотидной последовательности. Предпочтительно, чтобы мутантный нефункциональный связывающий ДНК-сайт имел одно или более изменений в неполной палиндромной последовательности из семи последовательных оснований. В еще одном воплощении настоящего изобретения предоставляется полинуклеотид, который содержит в себе один или более двухцепочечных мутантных, улучшенных связывающих ДНК-сайтов, которые демонстрируют усиленную транскрипцию расположенной ниже нуклеотидной последовательности по сравнению с немутантным связывающим ДНК-сайтом. Двухцепочечный связывающий ДНК-сайт(например, по меньшей мере 32 основания первой цепи немутантного сайта являются идентичными по последовательности с 5'-G/C (N)5 С С G А/Т С G G (N)19 G/C-3', SEQ ID NO: 22) получен из эукариот (например, из грибов), где немутантный и мутантный улучшенные сайты специфически связываются активатором транскрипции PrtT в условиях, разрешающих связывание, и такое связывание будет активировать транскрипцию расположенной ниже нуклеотидной последовательности. Но по меньшей мере одно основание (например, одно, два, три, четыре, пять, шесть, семь, восемь, или девять оснований) первой цепи мутантного сайта изменены по сравнению с последовательностью немутантного сайта (например,посредством добавления, удаления, перемещения, трансверсии, или любого их сочетания, оснований в удлиненной последовательности) таким образом, что PrtT специфически распознает сайт и активирует и усиливает транскрипцию расположенной ниже нуклеотидной последовательности. Поскольку 24 основания в удлиненном связывающем ДНК-сайте могут быть вырожденными, изменения в последовательности для изменения сродства связывания PrtT и/или для изменения активируемой PrtT транскрипции рас-3 014783 положенной ниже нуклеотидной последовательности могут производиться максимум в девяти позициях. Вторая цепь мутантного сайта является комплементарной к первой цепи. Немутантный и мутантный улучшенные сайты специфически связываются активатором транскрипции PrtT в условиях, разрешающих связывание, и такое связывание будет обеспечивать активацию транскрипции расположенной ниже нуклеотидной последовательности. Предпочтительно, чтобы мутантный улучшенный связывающий ДНК-сайт имел одно или более изменение как в неполной палиндромной последовательности из семи последовательных оснований, так и в удлиненной последовательности. Термин "усиленная транскрипция нуклеотидной последовательности, расположенной ниже мутантного связывающего ДНК-сайта" в контексте настоящего изобретения означает, по меньшей мере, увеличение на 10%, по меньшей мере, увеличение на 20%, по меньшей мере, увеличение на 30%, по меньшей мере, увеличение на 40%, по меньшей мере, увеличение на 80%, по меньшей мере, увеличение на 100%,по меньшей мере, увеличение на 200% или, по меньшей мере, увеличение на 300% количества продукта транскрипции по сравнению с соответствующей нуклеотидной последовательностью, расположенной ниже немутантного связывающего ДНК-сайта при экспрессии в одних и тех же условиях с применением одного и того же способа определения. Методы оценки количества продукта экспрессии известны специалисту в данной области техники. Примерами таких методов оценки количества продукта экспрессии являются Нозерн-блоттинг, количественная полимеразная цепная реакция (ПЦР), полимеразная цепная реакция в реальном времени, анализ на матрицах генов (Gene Array Analysis). Полинуклеотид может быть выделен из эукариотического источника (например, из грибов). Например, он может быть клонирован посредством скрининга библиотеки рекомбинантных последовательностей источника или посредством амплификации фрагментарной последовательности ДНК источника. Таким образом, он может быть очищен до любой желаемой степени из эукариот (например, по меньшей мере 90% растворенных веществ в композиции представляют собой молекулы желаемой нуклеиновой кислоты). Последовательность промотора из источника, не являющегося грибом, который не имеет активатора транскрипции PrtT, может случайно быть идентична SEQ ID NO: 22 или достаточно похожа на не для того, чтобы быть исправленной до SEQ ID NO: 22 и, таким образом, регулироваться PrtT в клетке. Она может быть рекомбинантной, если ДНК, которые не являются коллинеарными в геноме, соединяются вместе для конструирования рекомбинантного полинуклеотида. По меньшей мере, некоторые части полинуклеотида могут быть получены из источников, отличных от клетки, в которой реплицируется данный полинуклеотид. В экспрессионном векторе контакты оснований, ориентация и расположение одного или более связывающих ДНК-сайтов является типичным для содержащих цинковые пальцы транскрипционных факторов, таких как PrtT. Связывающий ДНК-сайт может быть встроен в экспрессионный вектор для оптимальной или субоптимальной активации транскрипции расположенной ниже нуклеотидной последовательности (например, протеазного гена или другого гена как природного, так и гетерологичного), расположенной в пределах 1200 оснований, 800 оснований или в пределах от 100 до 1200 оснований от сайта инициации транскрипции. Расположенная ниже нуклеотидная последовательность (например, кодирующая последовательность) может представлять собой ген не протеазы, ген не грибов, или и то, и другое. Другие регуляторные последовательности промотора могут быть, а могут и не быть, происходящими из гена грибов. Два различных типа PrtT-экспрессирующих клеток грибов могут быть сконструированы с применением вышеупомянутых воплощений настоящего изобретения. Клетки грибов сконструированы посредством встраивания функционального связывающего ДНК-сайта в хромосому или в эписому для обеспечения PrtT-зависимой активации транскрипции нуклеотидной последовательности, расположенной ниже данного сайта. Такой тип грибов будет совместно транскрибировать такие последовательности с генами протеаз. Альтернативно этому клетки грибов сконструированы посредством замещения хромосомных или эписомных связывающих ДНК-сайтов мутантными нефункциональными сайтами с целью ликвидации зависимости от PrtT, поскольку промотор, содержащий в себе мутантный нефункциональный сайт,больше не узнается PrtT. Природные гены протеаз могут быть инактивированы таким способом; PrtTзависимая активация транскрипции нуклеотидной последовательности, расположенной ниже функционального сайта, может быть также инактивирована. Данный тип грибов является пригодным для экспрессии природных или гетерологичных полипептидов, которые являются чувствительными к расщеплению протеазами. Таким образом, полинуклеотид, являющийся предметом настоящего изобретения,может представлять собой рекомбинационный вектор, содержащий в себе функциональный или нефункциональный связывающий ДНК-сайт, фланкированный гомологичными последовательностями локусамишени, и селективный маркер для детектирования рекомбинации, при этом не требуется присутствия расположенной ниже кодирующей последовательности для экспрессии с промотора. Предпочтительно,чтобы гомологичные фланкирующие последовательности содержали в себе по меньшей мере 30 оснований, более предпочтительно по меньшей мере 50 оснований, более предпочтительно по меньшей мере 100 оснований, по меньшей мере 200 оснований, более предпочтительно по меньшей мере 500 оснований, еще более предпочтительно по меньшей мере 1 т.п.н., наиболее предпочтительно по меньшей мере 2 т.п.н. локуса-мишени клетки-хозяина. Посредством предоставления нефункционального PrtT-4 014783 связывающего сайта специфические протеазы могут быть инактивированы в зависимости от интересующего белка, который должен продуцироваться (например, инактивация пролин-специфических протеаз в том случае, когда интересующий белок богат пролином, или инактивация карбоксипептидаз в том случае, когда интересующий белок является чувствительным к деградации карбоксипептидазами). Напротив, полная инактивация протеаз посредством удаления гена PrtT имеет недостаток, заключающийся в том, что эти протеазы необходимы для внутриклеточных процессов, и белковая регуляция в этом случае будет также инактивирована, что приведет к ухудшению экспрессии клеткой-хозяином. Кроме того, другие гены, отличные от протеаз (например, гены, кодирующие белки-переносчики), контролируемые PrtT,могут быть существенны для данной клетки, и инактивация PrtT может являться летальной для таких клеток. Полипептиды PrtT представляют собой активаторы транскрипции генов протеаз, действующие через связывающие PrtT ДНК-сайты в их промоторах. Термин "активатор транскрипции" в используемом в настоящем изобретении значении относится к полипептиду, который обладает способностью активировать транскрипцию с одного из следующих: специфического протеазного промотора или группы протеазных промоторов. PrtT является необходимым для инициации транскрипции гена протеазы (или иной гетерологичной нуклеотидной последовательности), с которым функционально связан промотор. Биологическая активность связывающего ДНК-сайта, который содержится в протеазном промоторе,может быть определена посредством измерения протеазной активности (например, протеолиза), как это описано в настоящем изобретении для определения активности кислых эндопротеаз с использованием бычьего сывороточного альбумина (BSA) в качестве субстрата. Данный способ также описан van denHombergh et al. (Curr. Tent. 28:299-308, 1995). Другие способы измерения протеазной активности могут быть найдены в WO 02/068623. Альтернативно этому биологическая активность связывающего ДНКсайта может быть определена посредством измерения уровня мРНК транскриптов протеаз. Уровни мРНК могут, например, быть количественно оценены посредством гибридизации (например, с помощью Нозерн- или слот-блотинга) или полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР). Анализ связывания на фильтре, сшивание белков и иммунопреципитация хроматина могут использоваться для идентификации связывания полипептида со связывающим ДНК-сайтом. Репортерный ген, находящийся под контролем промотора, содержащего в себе связывающий ДНКсайт, может быть использован как суррогатный заменитель протеазы или ее активности. Было описано измерение биологической активности -галактозидазы (lacZ) или зеленого флуоресцентного белка (GFP),использующихся в качестве репортерных генов (Luo, Gene 163:127-131, 1995; Henriksen et al., Microbiol. 145:729-734, 1999). Альтернативно этому один или более репортерный ген специфической протеазы, такой как рерА ген, кодирующий пепстатин-чувствительную внеклеточную аспарагиновую протеазу, может быть использован для измерения активности связывающего ДНК-сайта. Один или более сайт регулирует транскрипцию с промотора репортерного гена. По меньшей мере одна мутация может быть введена в связывающий ДНК-сайт при помощи стандартных приемов, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Примерами процедур мутагенеза являются процедуры, используемые при применении наборов QuickChangesite-directed mutagenesis kit (Stratagene), Altered Sites II in vitro mutagenesis system (Promega), достраивания перекрывающихся последовательностей (SOE-PCR, как это описано Но et al., Gene 77:51-59, 1989),или иные способы ПЦР (GriffinGriffin, eds., Molecular Biology: Current Innovations and Future Trends,Horizon Scientific Press, Norfolk, UK). Случайный мутагенез (например, химическое или радиационное повреждение) или подверженная ошибкам репликация ДНК (например, неправильное встраивание нуклеотидов) могут быть также использованы для внесения мутаций в связывающий ДНК-сайт. Связывающий ДНК-сайт, являющийся предметом настоящего изобретения, может быть получен из любых мицелиальных грибов (гифомицетов). "Мицелиальные грибы (гифомицеты)" включают в себя все мицелиальные формы из подотделов Eumycota и Oomycota. Мицелиальные грибы характеризуются наличием клеточной стенки, состоящей из хитина, целлюлозы, глюкана, хитозана, маннана и других сложных полисахаридов. Вегетативный рост происходит за счет удлинения гифов, и катаболизм углерода является облигатно аэробным. Виды мицелиальных грибов включают в себя, но ими не ограничиваются,представителей таких родов, как Acremonium, Aspergillus, Aureobasidium, Cryptococcus, Filibasidium,Fusarium, Humicola, Magnaporthe, Mucor, Myceliophthora, Neocallimastix, Neurospora, Paecilomyces, Penicillium, Piromyces, Schizophyllum, Talaromyces, Thermoascus, Thielavia, Tolypocladium и Trichoderma. Связывающий ДНК-сайт может быть получен из видов Aspergillus, таких как А. awamori, A. nidulans, A. niger, A. oryzae, A. sojae или A. fumigatus. Предпочтительно, когда он получен из штамма A. niger, A. oryzae, или A. fumigatus. В ином случае он получен из видов Penicillium, таких как P. chrysogenum,или из видов Fusarium, таких как F. oxysporum или F. venenatum. Следует понимать, что для вышеупомянутых видов настоящее изобретение включает в себя как совершенные, так и несовершенные стадии, а также другие таксономические эквиваленты, например анаморфы, независимо от названия вида, под которым они известны. Специалист в данной области техники с легкостью распознает идентичность подходящих эквивалентов. Например, полипептиды могут быть-5 014783 получены из микроорганизмов, которые являются таксономическими эквивалентами Aspergillus, как это определено RaperFennel (The Genus Aspergillus, Wilkins Company, Baltimore MD, 1965) независимо от видовых названий, под которыми они известны.Aspergilli представляют собой митоспоровые грибы, характеризующиеся как аспергилл, имеющие конидиоспорную ножку, для которых неизвестны телеоморфные состояния, завершающиеся везикулой,которая, в свою очередь, несет один или два слоя синхронно формирующихся специализированных клеток, именуемых по разному или стеригмами, или фиалидами, и споры бесполого размножения, называемые конидиями. Известные телеоморфы Aspergillus включают в себя Eurotium, Neosartorya и Emericella. Штаммы Aspergillus и их телеоморфы легко доступны пользователям в большом числе коллекций культур клеток. Связывающие ДНК-сайты могут быть получены из штаммов мицелиальных грибов, таких как, например, Aspergillus niger CBS 513.88, Aspergillus oryzae ATCC 20423, IFO 4177, ATCC 1011, ATCC 9576,ATCC 14488-14491, ATCC 11601, ATCC12892, Aspergillus fumigatus AF293 (CBS101355), P. chrysogenumATCC 36225, или ATCC 48272, Trichoderma reesei ATCC 26921, или ATCC 56765, или ATCC 26921, Aspergillus sojae ATCC 11906, Chrysosporium lucknowense ATCC44006 и их производных. Кроме того, связывающие ДНК-сайты могут быть идентифицированы и получены из других источников, включая микроорганизмы, выделенные из природной среды (например, почвы, компостов, воды,и т.д.). Методы выделения микроорганизмов из окружающей среды хорошо известны в данной области техники. Нуклеотидная последовательность может быть получена с использованием меченого зонда посредством скрининга геномной библиотеки другого микроорганизма. После того как нуклеотидная последовательность по меньшей мере одного связывающего ДНК-сайта была выявлена с использованием зонда, последовательность может быть выделена или клонирована с применением известных методовSpring Harbor, NY, 2001; и Ausubel et al., Current Protocols in Molecular Biology, Wiley InterScience, NY,1995). Условия низкой, средней, высокой строгости означают предгибридизацию и гибридизацию при температуре 42 С в 5SSPE, 0,3% SDS, 200 пг/мл деградированной и денатурированной ДНК спермы лосося (salmon sperm DNA) и одной из концентраций формамида 25, 35 или 50% для условий низкой,средней, высокой строгости соответственно. После этого реакция гибридизации промывается три раза в течение 30 мин с использованием 2SSC, 0,2% SDS при одной из температур 55, 65 или 75 С для условий низкой, средней, высокой строгости соответственно. Могут использоваться олигонуклеотидные зонды. Для детектирования соответствующих связывающих ДНК-сайтов они обычно являются мечеными(например, с использованием 32 Р, 33 Р, 3 Н, 35S, биотина, авидина или флуоресцентного/люминесцентного маркера). Например, молекулы, с которыми гибридизуется 32 Р-, 33 Р-, 3 Н- или 35S-меченый олигонуклеотидный зонд, могут быть детектированы с применением рентгеновской пленки или Phospho-Image анализа. Одним из вариантов нуклеотидной последовательности может также являться гомологичный или паралогичный связывающий ДНК-сайт в протеазном промоторе. В контексте настоящего изобретения определения гомологичная или паралогичная означают нуклеотидную последовательность, идентичную или близкую последовательности 5'-С С G А/Т С G G-3' и полученную из A. niger, A. oryzae, A. fumigatus, P. chrysogenum, F. oxysporum или из F. venenatum. Например, штаммы Aspergillus могут быть подвергнуты скринингу на предмет наличия гомологичного или паралогичного связывающего ДНКсайта в протеазном промоторе посредством гибридизации. Для детектирования гомологичной или паралогичной нуклеотидной последовательности, в соответствии с настоящим изобретением, геномная ДНКбиблиотека может быть подвергнута скринингу с использованием зонда, который гибридизуется с последовательностью 5'-С С G А/Т С G G-3', или более удлиненным ее вариантом, например с удлиненными вариантами, показанными в табл. 2 и 3. Методы, используемые для выделения или клонирования нуклеотидной последовательности, хорошо известны в данной области техники и включают в себя прямое выделение из геномной ДНК. Клонирование нуклеотидной последовательности, являющейся предметом настоящего изобретения, из такой геномной ДНК может быть выполнено с использованием известных гибридизационных методов."Полинуклеотид" определяется в рамках настоящего изобретения как двухцепочечная молекула нуклеиновой кислоты, которая изолирована из существующего в природе гена или которая была модифицирована таким образом, что она содержит сегменты нуклеиновой кислоты, которые комбинированы и расположены в таком порядке, который в природе не встречается. "Экспрессионный вектор" определяется в рамках настоящего изобретения как двухцепочечная молекула нуклеиновой кислоты, которая содержит в себе регуляторные последовательности, необходимые для экспрессии кодирующей последовательности и сохранения вектора (по меньшей мере, временного) в клетке. Определение "кодирующая последовательность", как это определяется в рамках настоящего изобретения, соответствует последовательности, которая транскрибируется в мРНК и транслируется в полипептид. Границы кодирующей по-6 014783 следовательности, как правило, определены стартовым кодоном ATG, расположенным на 5'-конце мРНК,и последовательностью стоп-кодона трансляции, ограничивающей открытую рамку считывания на 3'конце мРНК. Кодирующая последовательность может включать в себя, но ими не ограничивается, ДНК,кДНК и рекомбинантные нуклеотидные последовательности. Подразумевается, что термин экспрессия включает в себя любую стадию, вовлеченную в процесс образования полипептида, включая, но ими не ограничиваясь, транскрипцию, сплайсинг, посттранскрипционную модификацию, трансляцию, посттрансляционную модификацию, секрецию и протеолитический процессинг. Определение "регуляторная последовательность" в рамках настоящего изобретения соответствует последовательности, включающей в себя все компоненты, необходимые или предпочтительные для экспрессии полипептида. Каждая регуляторная последовательность может быть нативной или чужеродной по отношению к нуклеотидной последовательности, кодирующей полипептид. Такие регуляторные последовательности включают в себя, но не ограничиваются, лидерную последовательность, оптимальные последовательности инициации трансляции (как это описано в Kozak, J. Biol. Chem. 266:19867-19870,1991), последовательность полиаденилирования, пропептидную последовательность, препропептидную последовательность, промотор, сигнальную последовательность, а также терминатор транскрипции. Как минимум, регуляторные последовательности включают в себя промотор и стоп-сигналы для транскрипции и трансляции. Изменение нуклеотидной последовательности, кодирующей полипептид, перед ее встраиванием в экспрессионный вектор в зависимости от вектора может быть желательным или необходимым. Методы модификации нуклеотидных последовательностей с применением методов клонирования являются известными в данной области техники. Регуляторные последовательности могут предоставляться с линкерами для введения специфических сайтов рестрикции, что облегчает легирование регуляторных последовательностей с кодирующей областью нуклеотидной последовательности, кодирующей полипептид. Определение "функционально связанный" в рамках настоящего изобретения соответствует такой конфигурации, при которой регуляторная последовательность подходящим образом расположена в определенной позиции по отношению к кодирующей последовательности так, что регуляторная последовательность направляет экспрессию полипептида или транскрипцию с других расположенных ниже последовательностей. Регуляторная последовательность может представлять собой промотор, который узнается клетками и обеспечивает экспрессию расположенной ниже нуклеотидной последовательности. Промотор содержит в себе последовательности, регулирующие транскрипцию (например, один или более связывающий ДНК-сайт, являющийся предметом настоящего изобретения), которые регулируют экспрессию полипептида или транскрипцию других расположенных ниже последовательностей. Промотор может представлять собой любую нуклеотидную последовательность, которая демонстрирует транскрипционную активность в клетке, включая мутантные, измененные (укороченные) и гибридные промоторы, и может быть получен из генов, кодирующих внеклеточные или внутриклеточные полипептиды как нативные, так и гетерологичные для клетки. Регуляторная последовательность может также являться пригодной последовательностью терминатора транскрипции, где нуклеотидная последовательность узнается клетками и терминирует транскрипцию. Терминирующая последовательность функционально связана с 3'-концом нуклеотидной последовательности, кодирующей полипептид. Любой терминатор, который в клетке является функциональным,может быть применен в рамках настоящего изобретения. Предпочтительно, когда терминаторы для клеток мицелиальных грибов получают из генов, кодирующих амилазу TAKA A. oryzae, глюкоамилазу(glaA) A. niger, антранилатсинтазу A. nidulans, -глюкозидазу A. niger, ген trpC, а также трипсинподобную протеазу Fusarium oxysporum. Регуляторная последовательность может также представлять собой пригодную лидерную последовательность, нетранслируемый участок мРНК, которая важна для трансляции в клетке. Лидерная последовательность функционально связана с 5'-концом нуклеотидной последовательности, кодирующей полипептид. Любая лидерная последовательность, которая является функциональной в клетке, может быть использована в рамках настоящего изобретения. Предпочтительно, когда лидерные последовательности для клеток мицелиальных грибов получают из генов, кодирующих амилазу TAKA A. oryzae и триозофосфатизомеразу A. nidulans, а также glaA A. niger. Другие регуляторные последовательности могут быть получены из гена IPNS Penicillium или гена pcbC, гена -тубулина. Все регуляторные последовательности, приведенные в WO 01/21779, включаются в настоящее изобретение путем отсылки. Регуляторная последовательность может также представлять собой последовательность полиаденилирования, последовательность, которая функционально связана с 3'-концом нуклеотидной последовательности и которая при транскрипции узнается клеткой мицелиальных грибов как сигнал для добавления полиаденозиновых остатков к транскрибируемой мРНК. Любая последовательность полиаденилирования, которая является функциональной в клетке, может быть использована в рамках настоящего изобретения. Предпочтительно, когда последовательности полиаденилирования для клеток мицелиальных грибов получают из генов, кодирующих амилазу TAKA A. oryzae, глюкоамилазу A. niger, антранилат-7 014783 синтазу A. nidulans, трипсинподобную протеазу Fusarium oxysporum, а также -глюкозидазу A. niger. Экспрессионные векторы Еще одним воплощением настоящего изобретения являются экспрессионные векторы, содержащие в себе связывающий ДНК-сайт внутри промотора, расположенную ниже промотора транскрибируемую последовательность, стоп-сигнал транскрипции, стоп-сигнал трансляции, а также другие регуляторные последовательности, необходимые для поддержания вектора в клетке, полученные с использованием рекомбинантной технологии. Различные нуклеотидные и регуляторные последовательности, описанные выше, могут быть объединены для получения рекомбинантного экспрессионного вектора, который может включать в себя один или более подходящий сайт рестрикции для осуществления встраивания или замещения нуклеотидной последовательности, кодирующей полипептид, по данному сайту. Нуклеотидная последовательность, кодирующая желаемый полипептид, может быть экспрессирована посредством встраивания нуклеотидной последовательности или полинуклеотида, содержащего в себе нуклеотидную последовательность, в подходящий вектор для экспрессии. При конструировании экспрессионного вектора кодирующая последовательность располагается в векторе таким образом, чтобы кодирующая последовательность была функционально связана с соответствующими регуляторными последовательностями (например, с одним или более связывающим ДНК-сайтом), обеспечивающими экспрессию и, возможно, секрецию. Экспрессионный вектор может представлять собой любой вектор (например, плазмиду или вирус),который может быть легко сконструирован с использованием рекомбинантной техники и может обеспечить экспрессию нуклеотидной последовательности, кодирующей желаемый полипептид. Выбор вектора будет, как правило, зависеть от совместимости вектора с клеткой (например, мицелиальных грибов), в которую вектор должен быть введен. Векторы могут быть линейными или кольцевыми (то есть эписомами). Вектор может представлять собой автономно реплицирующийся вектор, то есть вектор, который существует как внехромосомная единица, репликация которой является независимой от хромосомной репликации, например плазмиду, внехромосомный элемент, минихромосому или искусственную хромосому. Автономно реплицирующийся вектор для клонирования может содержать в себе последовательность АМА 1 (см., например, AleksenkoClutterbuck, Fungal Genet. Biol. 21:373-397, 1997). Позитивные и негативные селективные маркеры в составе вектора могут быть использованы для целей генной инженерии. Функции поддержания (сохранения) и ориджин репликации являются существенными для эписом. Для сконструированных хромосом могут быть необходимы наличие центромеры, теломер и ориджинов репликации для их сохранения и сегрегации. Альтернативно этому вектор может быть одним из тех векторов, которые при введении в клетку(например, мицелиальных грибов) интегрируются в геном и реплицируются вместе с хромосомой, в которую они интегрировались. Интегративный вектор для клонирования может встраиваться в случайное место или в предопределенный локус-мишень в хромосоме клетки. В воплощении настоящего изобретения клонируемый интегративный вектор содержит в себе фрагментарную область, которая гомологична нуклеотидной последовательности в предопределенном локусе-мишени в геноме клетки, обеспечивающую направленное встраивание клонируемого вектора в данный локус. С целью облегчения направленного встраивания клонируемый вектор предпочтительно линеаризуется перед проведением трансформации клеток. Линеаризация предпочтительно осуществляется таким образом, чтобы по меньшей мере один, но предпочтительно оба конца клонируемого вектора были фланкированы последовательностями,гомологичными локусу-мишени для обеспечения встраивания посредством гомологичной рекомбинации. Специалист в данной области техники определит оптимальную длину фланкирующей последовательности для специфической клетки-хозяина, которая необходима для обеспечения встраивания посредством гомологичной рекомбинации. Длина гомологичных последовательностей, фланкирующих локус-мишень,составляет предпочтительно по меньшей мере 30 п.н., предпочтительно по меньшей мере 50 п.н., предпочтительно по меньшей мере 0,1 т.п.н., еще более предпочтительно по меньшей мере 0,2 т.п.н., более предпочтительно по меньшей мере 0,5 т.п.н., еще более предпочтительно по меньшей мере 1 т.п.н., наиболее предпочтительно по меньшей мере 2 т.п.н. Предпочтительно, когда эффективность направленной интеграции в геном клетки-хозяина, то есть встраивания в предопределенный локус-мишень, увеличивается за счет усиленной способности клетки-хозяина к гомологичной рекомбинации. Такой фенотип клетки предпочтительно включает дефектный ген ku70, как это описано в WO 2005/095624. WO 2005/095624,который включен в настоящее изобретение путем отсылки, раскрывает предпочтительный способ получения клетки мицелиальных грибов, обладающей повышенной эффективностью направленного встраивания. Предпочтительно, когда последовательность ДНК в клонируемом векторе, которая гомологична локусу-мишени, происходит из высокоэкспрессируемого локуса, то есть когда она происходит из гена,который демонстрирует высокий уровень экспрессии в клетке-хозяине мицелиальных грибов. Ген, демонстрирующий высокий уровень экспрессии, то есть высокоэкспрессируемый ген, определяется в рамках настоящего изобретения как ген, мРНК которого может составлять по меньшей мере 0,5% (вес./вес.) тотальной клеточной мРНК, например, в условиях индукции, или в ином случае, ген, продукт которого может составлять по меньшей мере 1% (вес./вес.) от тотального клеточного белка, или в случае, когда продукт гена является секретируемым, ген, продукт которого может секретироваться до уровня по-8 014783 меньшей мере 0,1 г/л (как это описано в ЕР 357127). Большое число предпочтительных высокоэкспрессируемых генов грибов приведено в качестве примера: гены амилазы, глюкоамилазы, алкогольдегидрогеназы, ксиланазы, глицеральдегидфосфатдегидрогеназы или целлобиогидролазы (cbh) из Aspergilli илиTrichoderma. Наиболее предпочтительными высокоэкспрессируемыми генами для данных целей являются ген глюкоамилазы, предпочтительно ген глюкоамилазы A. niger, ген TAKA-амилазы A. oryzae, генgpdA A. nidulans, ген cbh Trichoderma reesei, предпочтительно cbh1. Они могут представлять собой гены грибов, регулируемые связывающими ДНК-сайтами, являющимися предметом настоящего изобретения. Более чем одна копия нуклеотидной последовательности, кодирующей полипептид, может быть введена в клетку для увеличения экспрессии продукта гена. Это может быть осуществлено предпочтительно посредством встраивания в хромосому клетки нуклеотидной последовательности, более предпочтительно посредством направленного встраивания нуклеотидной последовательности в один из высокоэкспрессируемых локусов, только что перечисленных выше. Эффективность встраивания может быть увеличена при использовании рекомбиназы. В ином случае, это может быть осуществлено посредством включения амплифицируемого селективного маркерного гена с нуклеотидной последовательностью, когда клетки,содержащие амплифицированные копии селективного маркерного гена и, следовательно, дополнительные копии нуклеотидной последовательности, могут быть отобраны посредством культивирования клеток в присутствии подходящего селективного агента. Для еще большего увеличения числа копий последовательности ДНК для достижения суперэкспрессии может быть применена техника конверсии гена,описанная в WO 98/46772. Векторы предпочтительно содержат один или более селективных маркеров, наличие которых позволяет легко проводить селекцию трансформированных клеток. Селективный маркер представляет собой ген, продукт которого обеспечивает устойчивость к биоциду или противовирусную устойчивость,устойчивость к тяжелым металлам, прототрофию у ауксотрофов, и тому подобное. Селективный маркер для применения в клетках мицелиальных грибов может выбираться из группы, включающей, но ими не ограничивающейся, amdS (ацетамидазу), argB (орнитинкарбамоилтрансферазу), bar (фосфинотрицинацетилтрансферазу), bleA (связывание блеомицина), hygB (гигромицинфосфотрансферазу), niaD (нитратредуктазу), pyrG (оротидин-5'-фосфатдекарбоксилазу), sC (сульфатаденилтрансферазу) и trpC (антранилатсинтазу), а также их эквиваленты из других видов. Предпочтительными для применения в клетке Aspergillus и Penicillium являются гены amdS (ЕР 635574, WO 97/06261) и pyrG A. nidulans или A. oryzae, а также ген bar Streptomyces hygroscopicus. Более предпочтительно применение гена amdS, еще более предпочтительно гена amdS из A. nidulans или A. niger. Наиболее предпочтительно селективный маркерный ген представляет собой кодирующую последовательность amdS A.nidulans, слитую с промоторомgpdA A.nidulans, как раскрыто в ЕР 635574, который включен в настоящее изобретение путем отсылки. Также могут применяться гены AmdS, например те, которые раскрыты в WO 97/06261. Способы объединения описанных выше элементов при конструировании экспрессионного вектора,являющегося предметом настоящего изобретения, хорошо известны в данной области техники (например, SambrookRussell, Molecular Cloning: A Laboratory Manual, 3rd Ed., CSHL Press, 2001; Ausubel etal., Current Protocols in Molecular Biology, Wiley InterScience, 1995). Клетки-хозяева и другие сконструированные клетки Связывающие ДНК-сайты, являющиеся предметом настоящего изобретения, предпочтительно применяются для создания одного из двух типов клеток:(i) первого типа клеток, которые будут хорошо пригодны для продуцирования желаемого полипептида, где желаемый полипептид находится под контролем одного или более связывающего ДНК-сайта, и(ii) второго типа клеток, которые будут хорошо пригодны для продуцирования желаемого полипептида, где желаемый полипептид является чувствительным к деградации протеиназами. В данном типе клеток по меньшей мере один связывающий ДНК-сайт в геноме клетки-хозяина является мутантным в соответствии со вторым аспектом настоящего изобретения таким образом, что PrtT больше специфически не распознает мутантный нефункциональный сайт или PrtT больше не активирует транскрипцию. Оба типа клеток необязательно дополнительно содержат в себе полинуклеотид или экспрессивный вектор, содержащий в себе по меньшей мере один функциональный или мутантный (то есть нефункциональный или улучшенный) связывающий ДНК-сайт, который расположен выше и функционально связан с транскрибируемой нуклеотидной последовательностью (например, кодирующей полипептид, чувствительный к протеазной деградации, полипептид, находящийся под контролем активатора транскрипцииPrtT, активатор транскрипции PrtT сам по себе или другой полипептид, который необходимо продуцировать). Клетка-хозяин необязательно обладает повышенным ответом на несвернутые белки (unfolded protein response, UPR), что увеличивает возможность продуцирования интересующего полипептида. UPR может быть повышен с применением способов, описанных в US 2004/0186070 и/или в US 2001/0034045 и/или в WO 01/72783. Более конкретно, уровень белка НАС 1 и/или IRE1, и/или РТС 2 был изменен с целью получения клетки-хозяина, обладающей повышенным UPR. Альтернативно или в сочетании с повышенным UPR и/или фенотипом, демонстрирующим сниженную экспрессию протеиназ и/или секрецию протеиназ, клетка-хозяин имеет оксалат-дефицитный фенотип для увеличения уровня продукции-9 014783 интересующего полипептида. Оксалат-дефицитный фенотип может быть получен с использованием способов, описанных в WO 2004/070022 A2. Альтернативно или в сочетании с повышенным UPR и/или дефицитностью по оксалату клеткахозяин демонстрирует набор фенотипических различий по сравнению с клеткой дикого типа, свидетельствующих о возможности увеличения уровня продукции интересующего полипептида. Такие различия могут включать в себя, но ими не ограничиваются, пониженную экспрессию глюкоамилазы и/или нейтральной -амилазы А, и/или нейтральной -амилаза В, протеазы, а также оксалатгидролазы. Указанные фенотипические различия, демонстрируемые клеткой-хозяином, могут быть получены посредством генетических модификаций в соответствии со способами, описанными в US 2004/0191864 А 1. Выбор клетки-хозяина будет в значительной степени зависеть от источника нуклеотидной последовательности, кодирующей желаемый полипептид, который необходимо продуцировать. Предпочтительно клетка-хозяин представляет собой клетку мицелиальных грибов, как они определены ранее в настоящем изобретении в качестве источника, из которого может быть получен связывающий ДНК-сайт. Клетка-хозяин может также представлять собой клетку-хозяина, как это раскрыто в WO 01/68864 или WO 00/20596. Введение экспрессионного вектора или иного полинуклеотида в клетку мицелиальных грибов может включать процесс, состоящий из получения протопластов, трансформации протопластов и регенерации клеточной стенки известным способом. Подходящие способы трансформации клеток Aspergillus описаны в ЕР 238023 и в работе Yelton et al. (Proc. Natl. Acad. Sci. USA 81:1470-1474, 1984). Пригодный способ трансформации видов Fusarium описан Malardier et al. (ген 78:147-156, 1989) или в WO 96/00787. Экспрессионный вектор или конструкт нуклеиновой кислоты, которые могут применяться, были ранее описаны в соответствующих разделах. Продукция полипептида, чувствительного к протеазной деградации В соответствии с другим своим воплощением, настоящее изобретение касается клетки-хозяина с фенотипом, соответствующим сниженной протеазной активности, где клетка представляет собой мутантную родительскую клетку, пригодную для продуцирования полипептида, чувствительного к деградации протеиназами, где родительская клетка содержит в себе одну или более нуклеотидную последовательность, кодирующую протеазы, транскрипция которых активируется активатором транскрипции PrtT,и мутантная клетка транскрибирует меньшее количество протеазных генов, по сравнению с родительской клеткой при культивировании их в одних и тех же условиях, поскольку протеазные промоторы содержат в себе один или более мутантный нефункциональный PrtT-связывающий сайт. Предпочтительный способ измерения протеазной активности в клетке-хозяине описан в приведенном в настоящем изобретении разделе Примеры для определения активности кислой эндо-протеазы с использованием бычьего сывороточного альбумина (BSA) в качестве субстрата. Детальное описание данного способа также приводится в работе van den Hombergh et al. (Curr. Genet. 28:299-308, 1995). Измерение активности протеаз(ы) также может быть выполнено с использованием других известных способов. В соответствии с одним из таких способов, аликвота культуральной среды 48-часовой культуры клеток инкубируется в присутствии миоглобина кашалота, меченного 3 Н, при рН 4,0, затем измеряется радиоактивность фракции, растворимой в трихлоруксусной кислоте (ТСА) (van Noort et al., Anal. Biochem. 198:385-390, 1991). Другие способы были описаны для идентификации, например, аспарагиновой протеиназы A A. niger (Takahashi, Meth. Enzymoh 248:146-155, 1991), эндопептидаз (Morihara, Meth.al., Meth. Enzymoh 45:495-503, 1976). Альтернативно этому могут быть использованы другие способы оценки протеазной активности, такие как, например, описанный в WO 02/068623. Альтернативно этому используемым способом может являться Нозерн-блоттинг, использование репортерного гена, находящегося под контролем протеазного промотора, или Вестерн-блоттинг, или анализ с применением ДНКмикрочипов (EisenBrown, Meth. Enzymoh 303:179-205, 1999), как это также описано в настоящем изобретении. Мутантная клетка может продуцировать меньшее количество протеаз и проявлять меньшую протеазную активность по сравнению с родительской клеткой, которая используется в качестве контроля при проведении измерения с применением любого из данных способов. Мутант A. niger может продуцировать меньшее количество протеаз и проявлять меньшую протеазную активность по сравнению с представленным в коллекции штаммом A. niger CBS 513.88. Мутант A. oryzae может продуцировать меньшее количество протеаз и проявлять меньшую протеазную активность по сравнению с представленным в коллекции штаммом A. oryzae. Мутант P. chrysogenum может продуцировать меньшее количество протеаз и проявлять меньшую протеазную активность по сравнению с представленным в коллекции штаммомP. chrysogenum CBS 455.95. Мутант A. fumigatus может продуцировать меньшее количество протеаз и проявлять меньшую протеазную активность по сравнению с представленным в коллекции штаммом A.fumigatus AF293 (CBS 101355). Мутантная клетка может быть получена посредством генной инженерии с использованием техник рекомбинантных генетических манипуляций, путем проведения мутагенеза мицелиальных грибов или и того и другого. При использовании техник генетических манипуляций предпочтительным является по- 10014783 лучение рекомбинантных грибов предпочтительно посредством удаления связывающего ДНК-сайта, узнаваемого активатором транскрипции PrtT, более предпочтительно, когда удаленный связывающий ДНК-сайт заменяется на его нефункциональный вариант, и наиболее предпочтительно, когда удаление и замещение проводятся так, как это описано в ЕР 357127. Мутантная клетка может быть получена посредством модификации или инактивации связывающего ДНК-сайта, узнаваемого активатором транскрипции PrtT, присутствующего в клетке и необходимого для экспрессии расположенной ниже последовательности. Экспрессия протеаз в мутантной клетке может быть, таким образом, уменьшена или полностью устранена. Модификация или инактивация связывающего ДНК-сайта, являющегося предметом настоящего изобретения, может быть достигнута в результате проведения мутагенеза родительской клетки и отбора мутантных клеток, в которых способность экспрессировать протеазы снижена по сравнению с родительской клеткой. Мутагенез, который может быть специфическим или случайным, может быть осуществлен,например, с применением подходящего физического или химического мутагена, с применением подходящего олигонуклеотида или когда последовательность ДНК подвергается ПЦР-опосредованному мутагенезу. Кроме того, мутагенез может быть осуществлен при использовании любой комбинации этих мутагенных агентов. Примеры физического или химического мутагенного агента, пригодного для данной цели, включают гамма- или ультрафиолетовое (UV) излучение, гидроксиламин, N-метил-N'-нитро-N-нитрозогуанидин(MNNG), О-метилгидроксиламин, азотистую кислоту, этилметансульфонат (EMS), бисульфит натрия,муравьиную кислоту, а также аналоги нуклеотидов. При использовании таких агентов мутагенез, как правило, осуществляется посредством инкубации родительской клетки, которую необходимо мутировать, в присутствии выбранного мутагенного агента в подходящих условиях, далее следует отбор мутантных клеток, демонстрирующих сниженный уровень экспрессии гена. Впоследствии может быть проведена селекция полученных мицелиальных грибов посредством измерения уровня экспрессии желаемого полипептида и/или любой протеазы, для которой известно, что она находится под контролем активатора транскрипции PrtT. Мицелиальные грибы необязательно впоследствии подвергаются селекции посредством измерения уровня экспрессии данного интересующего нас гена, который должен экспрессироваться в клетке-хозяине. Мутантная клетка, которая была модифицирована или инактивирована посредством любого из описанных выше способов и продуцирует меньшее количество протеаз, а также демонстрирует меньшую протеазную активность по сравнению с родительской клеткой при культивировании клеток в идентичных условиях, которая измеряется с применением одинаковых способов, как это определено ранее, может нести другую нуклеотидную последовательность. Мутантная клетка демонстрирует протеазную активность, предпочтительно по меньшей мере на 25% меньшую, более предпочтительно по меньшей мере на 50% меньшую, еще более предпочтительно по меньшей мере на 75% меньшую и наиболее предпочтительно по меньшей мере на 95% меньшую по сравнению с родительской клеткой при культивировании клеток в идентичных условиях и применении одинаковых способов определения активности, как это определено ранее. Мутантная клетка мицелиальных грибов Aspergillus niger, Aspergillus oryzae, Aspergillusfumigatus, Penicillium chrysogenum, F. oxysporum или F. venenatum может продуцировать меньшее количество протеаз и демонстрировать меньшую протеазную активность по сравнению с соответствующими представленными в коллекциях мицелиальными грибами, упоминавшимися ранее, при культивировании клеток в идентичных условиях и применении одинаковых способов измерения, как это определено ранее. В соответствии с воплощением настоящего изобретения полипептиды далее продуцируются в клетке-хозяине, являющейся предметом настоящего изобретения, характеризующейся фенотипом, соответствующим сниженной протеазной активности, где клетка представляет собой мутантную родительскую клетку, пригодную для продукции полипептида, чувствительного к деградации протеиназами, где родительская клетка содержит в себе одну или более нуклеотидную последовательность, кодирующую протеазы, транскрипция которых активируется активатором транскрипции PrtT, и мутантная клетка транскрибирует меньшее количество протеазных генов по сравнению с родительской клеткой при культивировании клеток в одинаковых условиях, поскольку протеазные промоторы содержат в себе один или более мутантный нефункциональный связывающий ДНК-сайт. В соответствии с предпочтительным воплощением настоящего изобретения полипептид продуцируется согласно способу, включающему:(a) культивирование клетки-хозяина с фенотипом, соответствующим сниженной протеазной активности, в питательной среде в условиях, подходящих для экспрессии полипептида,(b) экспрессию полипептида в данной клетке-хозяине и(c) необязательное выделение полипептида из питательной среды или из клетки-хозяина. В соответствии с другим предпочтительным воплощением настоящего изобретения полипептид продуцируется согласно способу, включающему:(а) трансформацию клетки-хозяина с фенотипом, соответствующим сниженной протеазной активности, экспрессионным вектором, где вектор экспрессирует полипептид,(b) культивирование клетки-хозяина в питательной среде в условиях, подходящих для экспрессии(d) необязательное выделение полипептида из питательной среды или из клетки-хозяина. Продукция других нативных или гетерологичных полипептидов и других последовательностей В соответствии с еще одним воплощением настоящее изобретение касается способов транскрибирования расположенной ниже нуклеотидной последовательности с использованием активатора транскрипции PrtT в клетке-хозяине, где транскрибируемая последовательность кодирует желаемый полипептид или представляет собой функциональную молекулу нуклеиновой кислоты, включающих:(iv) немутантный и/или мутантный, улучшенный связывающий ДНК-сайт, являющийся предметом настоящего изобретения, а также содержащей в себе расположенную ниже нуклеотидную последовательность, которая кодирует полипептид, где транскрипция расположенной ниже нуклеотидной последовательности активируется PrtT,(b) экспрессию полипептида в клетке-хозяине и(d) необязательное выделение полипептида из питательной среды или из клетки-хозяина. Продуцируемый полипептид может быть чувствителен к деградации протеазами. В таком случае будет применяться мутантная клетка-хозяин, которая является дефицитной по протеазам. Клетка-хозяин,дефицитная по протеазам, предпочтительно получена в соответствии со способом настоящего изобретения. Грибы могут расти или сохраняться в питательной среде, являющейся пригодной для продуцирования желаемого полипептида, при использовании способов, известных в данной области техники. Например, клетки могут быть помещены на твердый субстрат, встряхиваться в колбе, культивироваться в условиях ферментации в малых или в больших объемах (включая непрерывную и периодическую ферментацию, ферментацию с доливом среды или твердофазную ферментацию) в лабораторных или промышленных ферментерах в пригодной среде и в условиях, обеспечивающих экспрессию и/или выделение полипептида. Культивирование проводят в пригодной питательной среде, содержащей в себе источники углерода и азота, а также неорганические соли, с использованием способов, известных в данной области техники (см., например, BennettLaSure, eds., More Gene Manipulations in Fungi, Academic Press, CA, 1991). Пригодные культуральные среды доступны от коммерческих поставщиков или могут быть приготовлены с использованием опубликованных составов (например, см. в каталогах American Type CultureCollection). Если полипептид секретируется в питательную среду, то полипептид может быть получен непосредственно из среды. Если полипептид не секретируется, то он может быть получен из клеточных лизатов. Полученный в результате полипептид может быть выделен с применением способов, известных в данной области техники. Например, полипептид может быть выделен из питательной среды с применением обычных процедур, включающих, но ими не ограничивающихся, центрифугирование, фильтрацию,экстрагирование, распылительную сушку, выпаривание или осаждение. Выделенный полипептид может быть дополнительно очищен с помощью разнообразных способов, известных в данной области техники,включающих, но ими не ограничивающихся, хроматографию (например, ионообменную, аффинную,гидрофобную, хроматофокусирование или гель-фильтрацию), электрофорез (например, препаративное изоэлектрическое фокусирование), дифференциальное растворение (например, осаждение ацетоном или сульфатом аммония) или экстракцию (например, хаотропным агентом, солью или рН). См., например,JansonRyden, eds., Protein Purification, VCH Publishers, New York, 1989. Полипептид может быть детектирован с применением способов, известных в данной области техники, которые являются специфическими для полипептида. Такие способы детектирования могут включать в себя применение специфических антител, образование продукта фермента, исчезновение субстрата фермента или электрофорез в полиакриламидном геле в денатурирующих условиях (SDS-PAGE). Например, для определения активности полипептида может быть использован ферментативный метод. Способы определения ферментативной активности являются известными в данной области техники для большого числа ферментов. Клетки могут продуцировать по меньшей мере на 20% больше, по меньшей мере на 50% больше, по меньшей мере на 100% больше, по меньшей мере на 200% больше либо по меньшей мере на 300% больше полипептида по сравнению с соответствующей родительской клеткой при культивировании в одинаковых условиях с применением одного из данных способов. Предпочтительно, когда родительская клетка представляет собой один из представленных в коллекциях штаммов, упоминавшихся выше, и является клеткой-хозяином или источником последовательности связывающего ДНК-сайта. Полипептид может представлять собой любой полипептид как нативный, так и гетерологичный для клетки мицелиальных грибов. Определение "гетерологичный полипептид" в рамках настоящего изобретения означает полипептид, который не продуцируется клеткой мицелиальных грибов дикого типа. Определение "полипептид" не означает соответствия какой-либо определенной длине кодируемого продук- 12014783 та и, таким образом, включает в себя пептиды, олигопептиды и белки. Нуклеотидная последовательность, кодирующая гетерологичный полипептид, может быть получена из любого прокариотического,эукариотического или иного источника, а также может представлять собой синтетический ген. Определение "полученный из" так, как оно используется в настоящем изобретении, в связи с данным источником должно означать, что полипептид продуцируется источником или клеткой, в которую был введен ген из данного источника. Желаемый полипептид может представлять собой антитело или его антигенсвязьшающую часть,антиген, фактор свертывания крови, фермент, пептидный гормон или его вариант, рецептор или его лиганд-связывающую часть, регуляторный белок, структурный белок, репортерный белок, транспортный белок, внутриклеточный белок, белок, вовлеченный в процесс секреции, белок, вовлеченный в процесс укладки белков, шаперон, пептидный переносчик аминокислот, фактор гликозилирования или транскрипционный фактор. Полипептид может секретироваться во внеклеточную культуральную среду. Ферменты включают в себя оксидоредуктазы, трансферазы, гидролазы, лиазы, изомеразы, лигазы,каталазы, целлюлазы, хитиназы, кутиназы, дезоксирибонуклеазы, декстраназы, а также эстеразы. Полипептид может представлять собой карбогидразу, например целлюлазы, такие как эндоглюканазы, глюканазы, целлобиогидролазы, а также -глюкозидазы, гемицеллюлазы или пектинолитические ферменты, такие как ксиланазы, ксилозидазы, маннаназы, галактоназы, галактозидазы, пектинметилэстеразы, пектинлиазы, пектатлиазы, эндополигалактуроназы, экзополигалактуроназы, рамногалактуроназы,арабаназы, арабинофуранозидазы, арабиноксилангидролазы, галактуроназы, лиазы, а также амилолитические ферменты; гидролазу, изомеразу или лигазу, фосфатазы, такие как фитазы, эстеразы, такие как липазы, фосфолипазы, галактолипазы, протеолитические ферменты, карбоксипептидазу, эндопротеазу,металлопротеазу, сериновую протеазу, аминопептидазу, оксидоредуктазы, такие как оксидазы, трансферазы, а также изомеразы. Более предпочтительно, когда желаемый полипептид представляет собой фосфолипазу свиньи. Полипептид может представлять собой амилазу, карбогидразу, каталазу, хитиназу,кутиназу, циклодекстрингликозилтрансферазу, дезоксирибонуклеазу, эстеразу, -галактозидазу, галактозидазу, глюкоамилазу, -глюкозидазу, -глюкозидазу, галопероксидазу, протеолитический фермент, инвертазу, лакказу, липазу, маннозидазу, мутаназу, оксидазу, пектинолитический фермент, пероксидазу, фосфолипазу, полифенолоксидазу, рибонуклеазу, трансглутаминазу, глюкооксидазу, гексозооксидазу или монооксигеназу. Полипептид может представлять собой инсулин человека или его аналог, гормон роста человека,эритропоэтин человека, тканевой активатор плазминогена (tPA) человека или инсулинотропин человека. Альтернативно этому полипептид может представлять собой внутриклеточный белок или фермент,такой как, например, шаперон, протеаза или фактор транскрипции. Пример такого случая описан Punt etal. (Appl. Microbiol. Biotechnol. 50:447-454, 1998). Такой подход может быть применен, например, для увеличения эффективности клетки-хозяина как продуцента белка в том случае, если известно, что данный полипептид, такой как шаперон, протеаза или фактор транскрипции, является лимитирующим фактором для продуцирования белка. В способах настоящего изобретения клетки мицелиальных грибов могут также применяться для продуцирования рекомбинантных полипептидов, которые являются нативными для клетки. Нативные полипептиды могут рекомбинантно продуцироваться посредством, например, помещения гена, кодирующего полипептид, под контроль другого промотора с целью увеличения экспрессии полипептида для быстрого экспорта интересующего природного полипептида из клетки посредством использования сигнальной последовательности, а также для увеличения числа копий гена, кодирующего полипептид, продуцируемый клеткой в норме. Настоящее изобретение также включает в рамки определения "гетерологичный полипептид" рекомбинантную продукцию полипептидов, нативных для клетки, в тех случаях,где такая экспрессия вовлекает применение генетических элементов, не являющихся эндогенными для клетки, или использование последовательностей эндогенных элементов, которые были изменены для функционирования так, как они в норме не функционируют в клетке мицелиальных грибов. Способы,используемые для выделения или клонирования нуклеотидных последовательностей, кодирующих гетерологичный полипептид, известны в данной области техники и включают в себя выделение из геномной ДНК, приготовление из кДНК или их сочетание. В способах настоящего изобретения гетерологичные полипептиды могут также включать в себя слитый или гибридный полипептид, в котором другой полипептид слит с N-концом или с С-концом полипептида или его фрагмента. Слитый полипептид получается посредством слияния нуклеотидной последовательности (или ее части), кодирующей один полипептид, с нуклеотидной последовательностью(или ее частью), кодирующей другой полипептид. Способы получения слитых полипептидов являются известными в данной области техники и включают в себя лигирование кодирующих последовательностей, кодирующих полипептиды, таким образом,что при этом не нарушается рамка считывания и экспрессия слитого полипептида осуществляется под контролем того же промотора(ов) и терминатора. Гибридные полипептиды могут содержать в себе сочетание частей или полных полипептидных последовательностей, полученных по меньшей мере из двух- 13014783 различных полипептидов, где один или более из них может являться гетерологичным для мутантной клетки гриба. Выделенная нуклеотидная последовательность, кодирующая интересующий гетерологичный полипептид, может быть изменена различными способами с целью обеспечения экспрессии полипептида. Подразумевается, что экспрессия включает в себя любую стадию, вовлеченную в процесс образования полипептида, включая, но ими не ограничиваясь, транскрипцию, посттранскрипционную модификацию, трансляцию, посттрансляционную модификацию, а также секрецию. Изменение нуклеотидной последовательности, кодирующей полипептид, перед ее встраиванием в вектор, может быть желательно или необходимо в зависимости от экспрессионного вектора. Способы модификации нуклеотидных последовательностей с использованием способов клонирования хорошо известны в данной области техники. Альтернативно этому расположенная ниже транскрибируемая последовательность не кодирует полипептид, но вместо этого представляет собой функциональную молекулу нуклеиновой кислоты. Может быть необходим посттранскрипционный сплайсинг или эндонуклеолитический/экзонуклеолитический процессинг новообразующихся транскриптов. Молекула нуклеиновой кислоты может представлять собой антисмысловую или малую интерферирующую молекулу РНК. Модификация или инактивация гена хозяина может быть осуществлена посредством принятых антисмысловых способов с использованием нуклеотидной последовательности, комплементарной нуклеотидной последовательности гена. Более конкретно, экспрессия гена клеткой мицелиальных грибов может быть снижена или полностью подавлена посредством введения нуклеотидной последовательности, комплементарной нуклеотидной последовательности гена гриба, которая может транскрибироваться в клетке и способна гибридизоваться с мРНК, продуцирующейся в клетке. В условиях, позволяющих комплементарной антисмысловой нуклеотидной последовательности гибридизоваться с мРНК, количество транслированного белка соответственно уменьшается или полностью подавляется. Примеры экспрессии антисмысловой РНК предоставляются в работах Ngiam et al. (Appl. Environ. Microbiol. 66:775-782, 2000) и Zrenner et al. (Planta 190:247-252, 1993). Модификация, снижение активности или инактивация гена хозяина может быть достигнута посредством способов РНК-интерференции (РНКи) (FEMS Microb. Lett. 237:317-324, 2004). Более конкретно,экспрессия гена клеткой мицелиальных грибов может быть снижена или полностью подавлена посредством клонирования идентичных смысловой и антисмысловой частей нуклеотидной последовательности,экспрессия которой должна быть изменена, во взаимообратной ориентации с нуклеотидным спейсером между ними, вставки в экспрессионный вектор и введения экспрессионного вектора в клетку, где двухцепочечная РНК (дцРНК) может быть транскрибирована и затем процессирована до более короткой малой интерферирующей РНК, которая способна гибридизоваться с мРНК-мишенью. После того как дцРНК транскрибирована, образование малых (21-23) нуклеотидных фрагментов миРНК приводит к направленной деградации мРНК, на которую должно быть оказано действие. Элиминирование специфической мРНК может осуществляться до разной степени. Способы РНК-интерференции, описанные в WO 2005/05672 и WO 2005/026356, могут применяться для модификации, снижения активности или инактивации гена хозяина. Способы идентификации протеазных генов В дополнительном аспекте настоящего изобретения предоставляются способы идентификации протеазных генов. Один или более дифференциально экспрессируемых генов детектируются в соответствующих клетках грибов как при наличии, так и в отсутствие активатора транскрипции PrtT (например, в случае удаления prtT или иного способа инактивации PrtT). Наличие экспрессии гена может быть определено с применением способов, известных специалисту в данной области техники. Примеры таких анализов экспрессии приведены выше в настоящем изобретении и включают в себя Нозерн-блотинг, ПЦР в реальном времени и ПЦР с обратной транскрипцией (ОТ-ПЦР). Те дифференциально экспрессируемые гены, которые кодируют протеазы (например, как определено по подобию последовательности с последовательностями известных протеаз, протеолитической активности продукта гена и наличию связывающего ДНК-сайта для PrtT), идентифицируются как протеазные гены, контролируемые PrtT. Идентифицированные таким способом новые протеазы и полинуклеотиды, кодирующие данные новые протеазы,входят в рамки настоящего изобретения. Сведения, подтверждающие возможность осуществления изобретения Далее настоящее изобретение описывается посредством нижеследующих примеров, которые не должны рассматриваться как ограничивающие рамки настоящего изобретения. Примеры В примерах, приведенных в настоящем изобретении, стандартные способы молекулярного клонирования, такие как выделение и очистка нуклеиновых кислот, электрофорез нуклеиновых кислот, ферментативная модификация, расщепление и/или амплификация нуклеиновых кислот, трансформацияE.coli, и т.д., были проведены так, как это описано в литературе (Sambrook et al. (2000) "Molecular Cloning: a laboratory manual", third edition, Cold Spring Harbor Laboratories, Cold Spring Harbor, New York; InnisCBS 513.88 дельта prtT. Данный штамм A. niger несет в себе делецию гена, кодирующего протеазный регулятор PrtT. CBS 513.88 дельта prtT был создан с применением подхода "MARKER-GENE FREE", как это описано в ЕР 635574. В данном патенте подробно описано, как провести делецию специфических последовательностей ДНК glaA в геноме CBS 513.88. Данный подход обеспечил получение MARKER-GENE FREE рекомбинантного A. niger CBS 513.88 дельта prtT, который не содержит никаких чужеродных последовательностей ДНК. Вектор для замещения гена в случае гена prtT, кодирующего протеазный регулятор, был сконструирован в соответствии с известными принципами и создан с использованием рутинных способов клонирования. В сущности, данные векторы включают фланкирующие области из примерно 1000-3000 пар оснований для открытой рамки считывания (ORF) prtT для обеспечения гомологичной рекомбинации в заранее определенный геномный локус. Кроме того, они содержат в себе двунаправленный селективный маркер amdS, расположенный между прямыми повторами. Характерная конструкция таких делеционных векторов раскрыта в ЕР 635574 и WO 98/46772, которые включены в настоящее изобретение путем отсылки. С применением олигонуклеотидов SEQ ID NO:15 и SEQ ID NO: 16 в качестве праймеров и геномной ДНК CBS 513.88 в качестве матрицы была проведена ПЦР для амплификации 1,5 т.п.н., расположенных ниже prtT фланкирующей области, и для введения сайтов рестрикции KpnI и XmaI на концы последовательности для обеспечения клонирования в pGBDEL (фиг. 4). Данный фланкирующий фрагмент размером 1,5 т.п.н., расположенный ниже prtT, был расщеплен KpnI и XmaI и встроен в вектор pGBDEL,расщепленный KpnI и XmaI, что привело к получению вектора pGBDEL-PRT1. С применением олигонуклеотидов SEQ ID NO: 11 и SEQ ID NO: 12 в качестве праймеров и геномной ДНК CBS 513.88 в качестве матрицы, фланкирующая область размером 3 т.п.н., расположенная выше prtT, обозначенная как фрагмент А, была амплифицирована с применением ПЦР. Кроме того, к 5'концу был добавлен сайт рестрикции BstBI с перекрыванием последовательности, расположенной нижеprtT области на 3'-конце фрагмента А. С применением олигонуклеотидов SEQ ID NO: 13 и SEQ ID NO: 14 в качестве праймеров и геномной ДНК CBS 513.88 в качестве матрицы фланкирующая область размером 500 п.н., расположенная ниже prtT, обозначенная как фрагмент В, была амплифицирована с применением ПЦР. Оба фрагмента, А и В, получившиеся в результате, были слиты с использованием ПЦР с достраиванием перекрывающихся последовательностей (sequence overlap extension, SOE-PCR, как это описано Но et al., Gene 77:51-59, 1989), олигонуклеотидов SEQ ID NO: 11 и SEQ ID NO: 14 в качестве праймеров и фрагментов А и В; в результате был получен фрагмент С размером 3,5 т.п.н. Данный фрагмент С был расщеплен с использованием BstBI и AscI и встроен в вектор pGBDEL-PRT1, расщепленныйBstBI и AscI, с образованием pGBDEL-PRT2 (фиг. 5). Последовательность введенных ПЦР фрагментов,содержащих в себе вышестоящую и расположенную ниже области гена prtT, была подтверждена при проведении сиквенсного анализа. Линейная ДНК из вектора, расщепленного BstBI/XmaI для удаления pGBDEL-PRT2, была выделена и использована для трансформации CBS 513.88. Данная линейная ДНК может интегрироваться в геном в локусе prtT, замещая, таким образом, кодирующую последовательность prtT конструктом, содержащим в себе amdS (см. фиг. 6). Трансформанты были отобраны на ацетамидной культуральной среде и колонии были очищены в соответствии со стандартными способами. Растущие колонии были диагностированы посредством ПЦР на встраивание в локус prtT. Удаление гена prtT было определено по наличию амплификации фрагмента с размером, специфическим для вставки pGBDEL-PRT2, и отсутствием фрагмента,специфического для локуса prtT дикого типа. Споры были посеяны на фторацетамидную культуральную среду для отбора штаммов, которые не содержат маркера amdS. Выбранные штаммы были протестированы с использованием Саузерн-анализа для подтверждения делеции гена prtT. Штаммы дельта prtT были отобраны как репрезентативные штаммы с инактивированным геном prtT (см. фиг. 6). Конструирование pepA нокаута Aspergillus niger CBS 513.88CBS 513.88 дельта рерА. Дефицитный по pepA штамм Aspergillus niger CBS 513.88 был создан, как это описано van den Hombergh et al. (Eur. J. Biochem. 247:605-613, 1997). Создание prtT нокаута CBS 513.88 дельта pepACBS 513.88 дельта prtT дельта рерА. Способ, описанный для конструирования CBS 513.88 дельта prtT, был использован для аналогичного конструирования prtT нокаута CBS 513.88 дельта pepA для получения CBS 513.8 8 дельта prtT дельтаpepA. Ферментация A. niger в колбе со встряхиванием Штаммы A. niger предварительно культивировали в 20 мл предкультуральной среды, как это описано в WO 99/32617. После выращивания в течение ночи 10 мл данной культуры клеток перенесли в ферментационную среду 1 (FM1), содержащую 7% глюкозы, как это описано в WO 99/32617. Данная средаFM1 содержит в себе из расчета на 1 л 25 г гидролизата казеина, 12,5 г дрожжевого экстракта, 1 гKH2PO4, 2 г K2SO4, 0,5 г MgSO47H2O, 0,03 г ZnCl2, 0,02 г CaCl2, 0,01 г MnSO44H2O, 0,3 г FeSO47H2O, 10 мл раствора пеницилин-стрептомицин (5000 ед./мл пенициллина и 5 мг/мл стрептомицина) и доводится до рН 5,6 с использованием 4 Н H2SO4. Ферментация осуществляется в 500 мл колбах с перегородкой на дне, содержащих 100 мл ферментационного бульона при 34 С и встряхивании при 170 об./мин в течение указанного количества дней. Для индукции протеаз мицелий был собран после культивирования в течение 16-24 ч в FM1, промыт при комнатной температуре средой для индукции (IM) и перенесен в IM с источником углерода, как это указано. Среда для индукция (IM) содержит из расчета на 1 л 6 г NaNO3, 0,5 г KCl, 1,5 г KH2PO4, 1,13 мл 4MKOH, 0,5 г MgSO47H2O, 0,01% (вес./об.) казаминовых кислот, 0,1% (вес./об.) дрожжевого экстракта, 1 мл концентрированного раствора микроэлементов (концентрированный раствор микроэлементов содержит из расчета на 1 л 22 г ZnSO47H2O, 11 г Н 3 ВО 3, 5 г FeSO47H2O, 1,7 г CoCl26H2O, 1,6 г Cu2SO45H2O,5 г MnCl24H2O, 1,5 г Na2MoO42H2O, 50 г ЭДТА, рН доводится до 6,5 с использованием 4 М KOH, раствор стерилизуют фильтрованием и хранят в темноте при температуре 4 С), 10 мл концентрированного раствора витаминов (концентрированный раствор витаминов содержит из расчета на 1 л 200 мг рибофлавина, 200 мг тиамин-HCl, 200 мг никотинамида, 100 мг пиридоксин-HCl, 20 мг пантотеновой кислоты,0,4 мг биотина, рН доводится до 6 с использованием 4 М NaOH, стерилизуется фильтрованием и хранится в темноте при температуре 4 С), рН доводится до 5,6, содержит также 1% (вес./об.) коллагена или 2%(вес./об.) обезжиренной соевой муки. Конструирование индуцированной коллагеном кДНК-библиотеки Штамм CBS 513.88 A. niger культивировался в 100 мл среды, как описано выше, при 34 С и встряхивании при 170 об./мин в вибрационном инкубаторе с использованием 500 мл колб для встряхивания с перегородкой на дне. A. niger CBS 513.88 был предварительно прокультивирован в течение ночи, после чего мицелий был перенесен в ферментационную среду 1 (FM1 содержит из расчета на 1 л 25 г гидролизата казеина, 12,5 г дрожжевого экстракта, 1 г KH2PO4, 2 г K2SO4, 0,5 г MgSO47H2O, 0,03 г ZnCl2, 0,02 гCaCl2, 0,01 г MnSO44H2O, 0,3 г FeSO47H2O, 10 мл раствора пенициллин-стрептомицин (5000 ед./мл пенициллина и 5 мг/мл стрептомицина), рН доведен до 5,6 с использованием 4 Н H2SO4). После культивирования в течение 20 ч мицелий был перенесен в среду для индукции (IM). Мицелий, собранный спустя 18, 28 или 48 ч после переноса в IM, содержащую 1% (вес./об.) коллагена или 2% (вес./об.) обезжиренной соевой муки, был использован для экстракции РНК. Экстракция РНК и выделение мРНК были осуществлены, как это детально описано в WO 99/32617. Создание экспрессионной библиотеки кДНК, включающее в себя синтез кДНК, лигирование линкеров и трансформацию Е. coli, также описано в WO 99/32617. Линкеры, применяемые для реакций кДНК, состояли из сайтов рестрикции HindIII и XhoI. Полученные в результате пулы кДНК были легированы в вектор pGBFIN23, расщепленный HindIII-XhoI, создание и использование которого описано в WO 99/32617. Физическая карта pGBFIN-23 может быть найдена на фиг. 3. Для трансформации электрокомцетентных клетокDH10B (Invitrogen) применялись лигазные смеси, что привело к образованию более 105 колоний на одну библиотеку кДНК, полученную из мицелия, индуцированного как соевой мукой, так и коллагеном. Секвенирование 96 случайно выбранных клонов каждой из двух библиотек выявило низкий процент векторов, не несущих вставку. Размеры вставки для секвенированных клонов составили от 0,5 до 4,7 т.п.н. со средним размером 1,7 т.п.н. Для обеспечения эффективного формата для скрининга библиотека была сформирована в пулы по 103 клонов. Для каждого из этих пулов была приготовлена суспензия в глицерине, полученные пробы хранили для дальнейшего использования. Способы определения протеазной активности Общие активности кислых эндопротеаз в супернатантах культур клеток были определены как количество деградированного бычьего сывороточного альбумина (BSA). Для этого 450 мкл 1% (вес./об.) BSA в 0,1 М NaOAc pH 4,0 инкубировали с 50 мкл супернатанта культуры клеток при температуре 37 С в течение различных отрезков времени. В конце периода инкубации оставшийся BSA был осажден 500 мкл 10% (вес./об.) трихлоруксусной кислоты (ТСА) с последующей инкубацией во льду в течение 10 мин. Преципитат был отцентрифугирован в течение 10 мин при 13000 об./мин в центрифуге Eppendorf. Оптическая плотность супернатанта измерялась при 280 нм. Одна единица протеазной активности была определена по методу Anson (J. Gen. Physiol. 22:79-89, 1938) как разница в единицах оптической плотности при 280 нм за 1 ч. Более подробное описание и ссылки на данный способ также приводятся в работе vanden Hombergh et al. (Curr. Genet. 28:299-308, 1995). Пример 1. Детекция PrtT-связывающего сайта. Детекция дифференциально экспрессируемых генов в CBS 513.88 дельта prtTPrtT является регулятором транскрипции протеаз, активатором некоторых протеаз WO 00/20596,WO 01/68864 и WO 06/040312. Посредством сравнения экспрессии генов в штаммах CBS 513.88 и CBS 513.88 дельта prtT, культивируемых в идентичных условиях, могут быть определены гены, экспрессия которых изменяется при удалении регулятора транскрипции PrtT. Для определения дифференциально- 16014783 экспрессирующихся генов может быть использовано несколько подходов, которые известны в данной области техники:PrtT-1, культивируемых в идентичных условиях, с использованием специфических зондов для известных генов, кодирующих протеазы (информацию о специфических зондах см. в работах van den Hombergh, An(ii) создание вычитающей библиотеки кДНК (способы, доступные в таких компаниях, какInvitrogen, Genomax и др.) и определение дифференциально экспрессируемых генов посредством секвенирования кДНК клонов; а также(iii) анализа на микрочипах полного транскриптора с применением такого микрочипа, как микрочипAfrymetrix GeneChip. Для определения дифференциально экспрессируемых генов в штаммах CBS 513.88 и CBS 513.88 дельта prtT, выращиваемых в условиях FM1, мы использовали изготовленный на заказ микрочип A. nigerAffymetrix GeneChip. Гены, кодирующие протеазы, которые показали значительно более низкие уровни экспрессии в штамме CBS 513.88 дельта prtT по сравнению с CBS 513.88, перечислены в табл. 1. Идентификация предполагаемых PrtT-связывающих сайтов Промоторный участок генов, кодирующих представленные в табл. 1 протеазы, размером 1,0 т.п.н. был получен из нуклеотидной последовательности штамма CBS 513.88 и содержит в себе последовательность, расположенную непосредственно выше стартовой точки инициации трансляции соответствующего гена. Промоторные участки размером 1,0 т.п.н. были проанализированы с использованием компьютерной программы Multiple Alignment Construction and Analysis Workbench (MACAW) (версия 2.0.5 для Macintosh). Данное программное обеспечение использует алгоритм множественного выравнивания(KarlinAltschul, Proc. Natl. Acad. Sci. USA 87:2264-2268, 1990; Schuler et al., Proteins: Structure,Function, and Genetics 9:180-190, 1991; Lawrence et al., Science 262:208-214, 1993). Были заданы следующие параметры: минимальная ширина рамки (pattern): 6, максимальная ширина рамки: 25, случайная затравка (seed): 12345, количество испытаний: 3, количество тераций в одном испытании: 50. Данный поиск привел к выявлению 7-нуклеотидной неполной палиндромной последовательности 5'-С С G А/Т С G G-3' во всех экспериментально выявленных генах, кодирующих протеазы (табл. 1). Дальнейший анализ расположенной выше последовательности или последовательности, расположенной ниже палиндромной последовательности в промоторах, привел к определению более длинной последовательности из 33 нуклеотидов 5'-G/C (N)5 С С G А/Т С G G (N)19 G/C-3' (SEQ ID NO: 22), где N = С, илиG, или А или, Т (см. табл. 2). PrtT содержит в себе ДНК-связывающий домен с цинковым двухядерным кластером Zn(II)2-Cys6. Для большого числа факторов транскрипции дрожжей и грибов типа Zn(II)2-Cys6 были обнаружены сайты связывания и большинство из этих сайтов содержат два полусайта CGG в виде инвертированных, перевернутых (последовательность, ориентированная в обратном направлении, например, прямая последовательность представляет собой CGG, перевернутая - CCG) или прямых повторов, разделенных фиксированным числом нуклеотидов, характерным для фактора транскрипции (Kim etBiol 22:2642-2649, 2002. Предполагаемый сайт связывания, определенный выше, содержит в себе перевернутый повтор, CCGA/TCGG, что свидетельствует о том, что данный сайт является частью последовательности для связывания PrtT. Анализ полного генома A. niger на наличие PrtT-связывающих сайтов Определив вероятный PrtT-связывающий сайт в экспериментально определенном наборе протеаз,мы проанализировали область размером 1,0 т.п.н. во всех предсказанных открытых рамках считывания(ORF's) в геноме CBS 513.88 на предмет других генов, кодирующих протеазы, которые могли бы регулироваться посредством 33-нуклеотидной последовательности 5'-G/C (N)5 C C G A/TC G G (N)19 G/C-3'(SEQ ID NO: 22). Данный анализ позволил выявить более чем 400 ORF's. Для того чтобы выбрать из данного набора генов гены, которые возможно кодируют протеазы, мы применили следующий подход:(i) мы создали архив всех доменов Pfam, которые содержатся в базе данных пептидаз MEROPS peptidase database (выпуск 7.20, Rawlings et al., Nucl. Acids Res. 32:D160-D164, 2004), поскольку домены(ii) мы использовали перечень доменов Pfam для поиска в геноме CBS 513.88 всех ORF's, которые кодируют белок, содержащий домен Pfam, идентифицированный в (i). При наложении списка, включающего более чем 400 ORF's, которые содержат в промоторной области размером 1,0 т.п.н. 33-нуклеотидную последовательность 5'-G/C (N)5 С С G А/Т С G G (N)19 G/C-3'in silico идентифицировали еще 16 протеаз, которые могли бы быть потенциальной мишенью регулятораPrtT. Для того чтобы подтвердить соответствие гена пептидазе/протеазе, мы осуществили поискMEROPS Blast (http://merops.sanger.ac.uk/) с идентифицированной последовательностью. Из 16 последо- 17014783 вательностей (табл. 3) восемь имели значение Е в MEROPS Blast-поиске ниже, чем е-25, которое было установлено как граничное значение для того, чтобы ген был идентифицирован как протеаза/пептидаза. Сходным образом, как это описано выше в случае поиска с использованием удлиненного PrtTсвязывающего сайта, мы осуществили поиск с использованием только неполной палиндромной последовательности, 5'-С С G Т/А С G G-3'. Результаты приведены в табл. 4. Было идентифицировано еще 15 генов, предположительно кодирующих пептидазы/протеазы. При применении поиска MEROPS Blast с использованием указанного выше отсекающего значения Е 11 генов могли быть определены как гены,кодирующие пептидазу/протеазу. Поиск in silico позволил дополнительно идентифицировать 8 генов, кодирующих пептидазу/протеазу, которые содержат в своей промоторной области размером 1,0 т.п.н. удлиненный предполагаемый PrtT-связывающий сайт, 5'-G/C (N)5 С С G A/TC G G (N)19-G/C-3' (SEQ ID NO: 22), и 11 генов,кодирующих пептидазу/протеазу, которые содержат в своей промоторной области размером 1,0 т.п.н. предполагаемый PrtT-связывающий сайт, неполную палиндромную последовательность, 5'-С С G Т/А СG G-3'. Таблица 1 Гены, кодирующие протеазы, в значительной степени негативно регулируемые в штамме Aspergillus niger CBS 513.88 дельта prtT по сравнению с родительским штаммом Aspergillus niger CBS513.88. ТриSEQ ID NO, указанные для каждого гена, соответствуют геномной ДНК, кДНК и белку соответственно. Геномная последовательность ДНК дополнительно содержит в себе фрагмент размером 1200 т.п.н., расположенный выше точки начала транскрипции. Таблица 2 Перечень расширенных нуклеотидных последовательностей 5'-C/G (N)5 С С G А/Т С G G (N)19 C/G3' (SEQ ID NO:22) вероятных PrtT-связывающий сайтов для генов, кодирующих протеазы, из табл. 1. Подчеркнута неполная палиндромная нуклеотидная последовательность, определенная с использованием программы MACAW. Три SEQ ID NO, указанные в случае каждого из генов, соответствуют геномной ДНК, кДНК и белку соответственно. Геномная последовательность ДНК дополнительно содержит в себе фрагмент размером 1200 т.п.н., расположенный выше точки начала транскрипции.- 19014783 Таблица 3 Перечень расширенных нуклеотидных последовательностей 5'-C/G (N)5 С С G А/Т С G G (N)19 C/G3' (SEQ ID NO:22) вероятных PrtT-связывающий сайтов, детектированных в генах, кодирующих протеазы, путем анализа in silico нуклеотидной последовательности CBS513.88. Неполная палиндромная нуклеотидная последовательность, определенная с использованием программы MACAW, подчеркнута. ТриSEQ ID NO, указанные в случае каждого из генов, соответствуют геномной ДНК, кДНК и белку соответственно. Геномная последовательность ДНК дополнительно содержит в себе фрагмент размером 1200 т.п.н., расположенный выше точки старта транскрипции.- 20014783 Таблица 4 Перечень генов, кодирующих протеазы, идентифицированные in silico, которые содержат последовательность 5'-С С G А/Т С G G-3' в своей промоторной области размером 1,2 т.п.н. Подчеркнута неполная палиндромная нуклеотидная последовательность, которая определена с использованием программыMACAW. Значение Е поиска Blast MEROPS показано в столбце значение Е/семейство, вместе с семейством протеаз, к которому предположительно относится кодируемый белок. Три SEQ ID NO, указанные в случае каждого из генов, соответствуют геномной ДНК, кДНК и белку соответственно. Геномная последовательность ДНК дополнительно содержит в себе фрагмент размером 1200 т.п.н., расположенный выше точки старта транскрипции. Пример 2. Функциональность PrtT-связывающего сайта. Конструирование и анализ различных вариантов промотора рерА,содержащего определенные мутации PrtT-связывающего сайта Промоторная область ррерА размером 1 т.п.н. была амплифицирована посредством ПЦР с использованием олигонуклеотидов SEQ ID NO: 1 и SEQ ID NO: 2 в качестве праймеров и геномной ДНК CBS 513.88 в качестве матрицы, в результате был получен продукт амплификации, содержащий SEQ ID NO: 17. Промоторная область рерА размером 0,6 т.п.н. была амплифицирована посредством ПЦР с использованием олигонуклеотидов SEQ ID NO: 3 и SEQ ID NO: 2 в качестве праймеров и геномной ДНК CBS 513.88 в качестве матрицы, в результате чего был получен продукт амплификации, содержащий в себеSEQ ID NO: 18. Кодирующая последовательность рерА была амплифицирован посредством ПЦР с использованием олигонуклеотидов SEQ ID NO: 5 и SEQ ID NO: 6 в качестве праймеров и кДНК библиотеки (коллаген) в качестве матрицы, в результате чего был получен продукт амплификации, содержащий в себе SEQ ID NO: 21. ПЦР-фрагменты были очищены на колонке для очистки продуктов ПЦР (QIAGEN) и использованы в ПЦР со слиянием (fusion PCR) для получения фрагмента кДНК ррерА-рерА размером 1 т.п.н. и фрагмента кДНК ррерА-рерА размером 0,6 т.п.н. Они, в свою очередь, были клонированы и секвенированы. Плазмида pCR-Blunt-Topo, содержащая ррерА-рерА кДНК размером 1 т.п.н., была использована в качестве матрицы для осуществления сайт-направленного мутагенеза с использованием набора QuickChange site-directed mutagenesis kit (Stratagene). Табл. 5 содержит перечень мутаций, которые были внесены в расширенную 33-нуклеотидную PrtT-связывающую последовательность 5'-G/C (N)5 С С G А/Т СG G (N)19 G/C-3' промотора рерА. Мутации покрывают неполную палиндромную последовательностьCCGACGG и/или консервативные концевые положения C/G. Олигонуклеотиды, применяемые для внесения данных мутаций, перечислены в табл. 5. После мутагенеза полученная вставка ррерА-рерА кДНК размером 1 т.п.н. в плазмиде pCR-Blunt-Topo была повторно секвенирована для подтверждения наличия желаемой мутации и исключения возможности наличия других мутаций, возможно, внесенных при проведении ПЦР. Один положительный клон в случае каждого из мутантов или вариантов был отобран и использован для конструирования конечного экспрессионного вектора с использованием XhoI/AscI в качестве сайтов клонирования (фиг. 2). Полученные конструкты были трансформированы в штаммы CBS 513.88 дельтаpepA (см. фиг. 1) и в CBS 513.88 дельта prtT дельта pepA. Полученные трансформанты были проверены методом проведения ПЦР с колоний с использованием SEQ ID NO: 1 и SEQ ID NO: 2 (в случае всех конструктов, несущих в себе промотор ррерА размером 1 т.п.н.) или с использованием SEQ ID NO: 2 и SEQID NO: 3 в качестве праймеров (в случае конструктов, несущих в себе промотор ррерА размером 0,6 т.п.н.). Встраивание конструктов было направлено в глюкоамилазный локус (glaA, см. фиг. 2) с использованием фланкирующих последовательностей, присутствующих в векторе. Правильные трансформанты выращивали в течение 10 дней в 100 мл среды CSM/MES (150 г/кг мальтозаH2O, 60 г/кг Bacto soyton, 1 г/кг NaH2PO4, 15 г/кг (NH4)2SO4, 1 г/кг MgSO47H2O, 0,08 г/кг Tween-80, 0,02 г/кг Basildon, 20 г/кг MES, 1 г/кг L-аргинин, рН 6,2) в 500 мл колбах с отбойниками при температуре 30 С и встряхивании при 250 об./мин. Спустя 10 дней супернатант был собран, и протеазная активность была измерена при использовании метода Anson (J. Gen. Physiol. 22:79-89, 1938). Результаты представлены на фиг. 1. Результаты, представленные на фиг. 1, четко показывают, что внеклеточная протеазная активность сильно снижена, когда в ррерА-рерА конструкте в CBS 513.88 дельта pepA палиндром CCGACGG мутирован или отсутствует, что свидетельствует об уменьшенной транскрипционной активности с мутантного промотора ррерА. Мутации в двух других консервативных положениях (концевых G/C остатках) в 33 нуклеотидном расширенном PrtT-связывающем сайте нарушают эффективность транскрипции с промотора pepA примерно на 25%. Удивительно, что сочетание мутации на 5'-конце связывающего сайта (замена G на Т) и вставки одного дополнительного нуклеотида в неполный палиндром (замена CCGACGG на CCGATCGG) привело к получению мутантного промотора рерА с повышенной примерно на 40% транскрипционной эффективностью. Последовательность более сильного промотора рерА приведена под номером SEQ ID NO:139. Также ниже продемонстрирована зависимость транскрипции pepA от PrtT. Когда немодифицированный конструкт ррерА-рерА кДНК размером 1,0 т.п.н. был трансформирован в CBS 513.88 дельтаpepA, наблюдалось сильное увеличение внеклеточной протеазной активности. Однако когда данный конструкт был трансформирован в CBS 513.88 дельта prtT дельта pepA (данные не показаны), никакой внеклеточной активности не наблюдалось, что свидетельствует о том, что активность рерА в высокой степени зависит от PrtT-опосредованной транскрипции. Более детально, палиндром CCGACGG крайне важен для PrtT-опосредованной транскрипции pepA. Это позволяет заключить, что CCGACGG представляет собой PrtT-связывающий сайт в промоторе рерА. Роль двух концевых консервативных остатков расширенного PrtT-связывающего сайта может заключаться в стабилизации комплекса белок-ДНК.- 22014783 Таблица 5 Перечень мутаций, внесенных в промотор pepA. Мутантные нуклеотиды выделены жирным шрифтом, а PrtT-связьшающий сайт подчеркнут. Последовательности олигонуклеотидных праймеров, которые применялись для внесения данных мутаций, приведены в двух правых столбцах. Описанное и заявленное здесь настоящее изобретение не должно ограничиваться рамками специфических воплощений, входящих в настоящее описание, поскольку данные воплощения предназначены для иллюстрации некоторых аспектов настоящего изобретения. Любые эквивалентные воплощения должны входить в рамки настоящего изобретения. На самом деле различные модификации настоящего изобретения в дополнение к тем, которые показаны и описаны в данной заявке, будут понятны специалисту в данной области техники из предшествующего описания. Такие модификации также должны быть включены в рамки приведенной ниже формулы изобретения. В случае конфликта настоящее раскрытие изобретения, включая определения, будет использовано для контроля. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Изолированный полинуклеотид, включающий двухцепочечный связывающий ДНК-сайт для активатора транскрипции PrtT, где по меньшей мере 32 основания первой цепи сайта идентичны по последовательности с 5'-G/C(N)5CCGA/TCGG(N)19G/C-3' (SEQ ID NO:22), вторая цепь сайта комплементарна первой цепи и связывание PrtT с сайтом активирует транскрипцию расположенной ниже нуклеотидной последовательности в клетке-хозяине. 2. Изолированный полинуклеотид, включающий двухцепочечный мутантный нефункциональный связывающий ДНК-сайт, где по меньшей мере 32 основания первой цепи немутантного сайта идентичны по последовательности с 5'-G/C(N)5CCGA/TCGG(N)19G/C-3' (SEQ ID NO:22), вторая цепь мутантного нефункционального сайта комплементарна первой цепи, немутантный сайт связывается активатором транскрипции PrtT и связывание PrtT с немутантным сайтом активирует транскрипцию с расположенной ниже нуклеотидной последовательностью в клетке-хозяине, но по меньшей мере одно основание первой цепи мутантного нефункционального сайта изменено по сравнению с нуклеотидной последовательностью немутантного сайта таким образом, что PrtT больше не связывается с мутантным нефункциональным сайтом или что PrtT больше не активирует транскрипцию расположенной ниже нуклеотидной последовательности. 3. Изолированный полинуклеотид, включающий двухцепочечный мутантный улучшенный связывающий ДНК-сайт, где по меньшей мере 32 основания первой цепи немутантного сайта являются идентичными по последовательности с 5'-G/C(N)5CCGA/TCGG(N)19G/C-3' (SEQ ID NO:22), вторая цепь мутантного улучшенного сайта комплементарна первой цепи, немутантный и мутантный улучшенные сайты связываются активатором транскрипции PrtT и связывание PrtT как с немутантным, так и с мутантным улучшенным сайтом активирует транскрипцию расположенной ниже нуклеотидной последовательности в клетке-хозяине, но по меньшей мере одно основание первой цепи мутантного улучшенного сайта изменено по сравнению с нуклеотидной последовательностью немутантного сайта таким образом, что транскрипция расположенной ниже нуклеотидной последовательности является улучшенной. 4. Рекомбинантный экспрессионный вектор, включающий связывающий ДНК-сайт по п.1 или 3 в составе промотора, стоп-сигнал транскрипции и стоп-сигнал трансляции. 5. Вектор по п.4, дополнительно включающий расположенную ниже нуклеотидную последовательность, которая кодирует полипептид, где транскрипция расположенной ниже нуклеотидной последовательности активируется PrtT. 6. Клетка-хозяин, включающая полинуклеотид по п.2 или 3 или экспрессионный вектор по п.4 или 5. 7. Клетка-хозяин, где по меньшей мере один связывающий ДНК-сайт в геноме клетки-хозяина мутирован в соответствии с п.2 таким образом, что PrtT не может связываться с мутантным сайтом или PrtT- 23014783 не может активировать транскрипцию расположенной ниже нуклеотидной последовательности. 8. Способ получения клетки-хозяина по п.7, где указанный способ включает в себя:(a) введение по меньшей мере одного мутантного, нефункционального связывающего ДНК-сайта в промотор клетки-хозяина посредством мутагенеза или рекомбинации и(b) необязательное подтверждение уменьшенного связывания PrtT с мутантным нефункциональным сайтом или уменьшенной PrtT-зависимой активации транскрипции в клетке-хозяине. 9. Клетка-хозяин, где связывающий ДНК-сайт в одном или более протеазном гене клетки-хозяина мутирован в соответствии с п.2 таким образом, что PrtT не может связываться с мутантным сайтом или что PrtT не может активировать транскрипцию протеазного гена, что приводит к получению клеткихозяина с фенотипом сниженной протеазной активности. 10. Способ получения полипептида, где указанный способ включает в себя:(c) необязательное извлечение полипептида из питательной среды или из клетки-хозяина. 11. Способ получения полипептида, где указанный способ включает в себя:(a) трансформацию клетки-хозяина по п.9 экспрессионным вектором, где вектор экспрессирует полипептид,(b) культивирование клетки-хозяина в питательной среде в условиях, приводящих к экспрессии полипептида,(c) экспрессию полипептида в клетке-хозяине и(d) необязательное извлечение полипептида из питательной среды или из клетки-хозяина. 12. Способ получения полипептида, где указанный способ включает в себя:(a) культивирование в питательной среде клетки-хозяина, содержащей вектор по п.5, в условиях,приводящих к экспрессии полипептида, кодируемого расположенной ниже нуклеотидной последовательностью, содержащейся в данном векторе,(b) экспрессию полипептида в клетке-хозяине и(d) необязательное извлечение полипептида из питательной среды или из клетки-хозяина.
МПК / Метки
МПК: C12N 15/80
Метки: пригодный, днк-сайт, связывающий, транскрипции, активатора, генов, экспрессии
Код ссылки
<a href="https://eas.patents.su/30-14783-svyazyvayushhijj-dnk-sajjt-aktivatora-transkripcii-prigodnyjj-dlya-ekspressii-genov.html" rel="bookmark" title="База патентов Евразийского Союза">Связывающий днк-сайт активатора транскрипции, пригодный для экспрессии генов</a>
Конструкции нуклеиновых кислот для экспрессии генов

Номер патента: 8247
Опубликовано: 27.04.2007
Автор: Браун Ральф Патрик
МПК: C12N 15/86, C12N 15/869
Метки: нуклеиновых, конструкции, экспрессии, кислот, генов
Формула / Реферат:
1. Конструкция нуклеиновой кислоты, содержащая нуклеиновую кислоту вирусного генома, которая представляет собой нуклеиновую кислоту генома вируса герпеса или последовательность, по меньшей мере на 80% ей гомологичную, где указанная нуклеиновая кислота вирусного генома содержит по меньшей мере два эндогенных регуляторных элемента экспрессии гена, каждый из которых содержит эндогенный промотор, способный экспрессироваться в клетках млекопитающего,...
Промотор для экспрессии генов в вирусах осповакцины и/или в клетках, инфицированных вирусом осповакцины, и способы его применения

Номер патента: 12846
Опубликовано: 30.12.2009
Автор: Лейрер Соня
МПК: A61K 39/275, C12N 15/86
Метки: осповакцины, способы, инфицированных, экспрессии, промотор, клетках, вирусом, вирусах, генов, применения
Формула / Реферат:
1. Промотор, состоящий из нуклеотидной последовательности или содержащий нуклеотидную последовательность, выбранную из группы, включающей: (i) любую нуклеотидную последовательность SEQ ID NO: 2-4, 6 и 8-12, (ii) нуклеотидную последовательность длиной по меньшей мере 15 нуклеотидов любой последовательности SEQ ID NO: 2-4 и 6 и (iii) производные любой последовательности SEQ ID NO: 8-12, причем в производном не более 10 нуклеотидов являются...
Способы измерения экспрессии генов из фиксированного и залитого в парафин образца ткани

Номер патента: 8832
Опубликовано: 31.08.2007
Авторы: Даненберг Питер, Даненберг Кэтлин, Свенсон Стивен
МПК: C12N 15/10, C12Q 1/68, G01N 33/48...
Метки: фиксированного, генов, способы, измерения, залитого, экспрессии, парафин, образца, ткани
Формула / Реферат:
1. Способ количественного измерения экспрессии целевого гена из фиксированного и залитого в парафин образца ткани, предусматривающий: (a) депарафинирование образца ткани для получения депарафинированного образца; (b) выделение мРНК из депарафинированного образца путем первого нагревания образца ткани в хаотропном растворе, содержащем эффективную концентрацию хаотропного агента, до температуры в диапазоне от примерно 50 до примерно 100шС в...
Полипептиды, содержащие домены протеина gax, участвующие в подавлении транскрипции и/или взаимодействующие с другими протеинами, соответствующие нуклеиновые кислоты и их применение

Номер патента: 3694
Опубликовано: 28.08.2003
Авторы: Марсиро Кристоф, Мафуди Абдерраим, Бранеллек Дидье, Фурнье Алан
МПК: C07K 14/47, C12N 15/12, A61K 38/17...
Метки: протеинами, домены, протеина, полипептиды, участвующие, другими, транскрипции, применение, нуклеиновые, содержащие, соответствующие, кислоты, взаимодействующие, подавлении
Формула / Реферат:
1. Полипептид, отличающийся тем, что он представляет собой фрагмент протеина GAX (Growth-Arrest-Specific Homeobox) человека, содержащий, по меньшей мере, остатки 1-32 протеина GAX и обладающий репрессорной активностью в отношении транскрипции и/или оказывающий положительное или отрицательное влияние на репликацию ДНК. 2. Полипептид по п.1, отличающийся тем, что он дополнительно содержит остатки 140-230 протеина GAX человека. 3. Полипептид,...
Выявление индуцибельных генов в микроорганизмах

Номер патента: 6579
Опубликовано: 24.02.2006
Автор: Карльсен Франк
МПК: C12Q 1/68
Метки: микроорганизмах, генов, индуцибельных, выявление
Формула / Реферат:
1. Способ выявления наличия микроорганизма в тестируемом образце, предусматривающий воздействие на тестируемый образец индуктором, который способен индуцировать экспрессию по меньшей мере одного гена в микроорганизме, и тестирование наличия мРНК, транскрибированной с гена, который индуцирован посредством воздействия указанным индуктором. 2. Способ по п.1, где микроорганизмом является медленно растущая бактерия. 3. Способ по п.2, где медленно...
Предыдущий патент: Сложные эфиры молочной кислоты в качестве агента для улучшения воздействия пестицидов на растения, способ улучшения воздействия пестицидов
Следующий патент: Способ производства монопентаэритритола высокой чистоты и монопентаэритритол, произведенный посредством процесса
Случайный патент: Магнитная головка для построения изображений по удельной проводимости при ее использовании в скважине